

ABSTRACT
Immunomodulation is an important part of lenalidomide's mode of action. We analyzed the impact of lenalidomide on T cells from patients with multiple myeloma during lenalidomide therapy in vivo and in patients with lenalidomide-refractory disease in vitro Patients enrolled in the German Speaking Myeloma Multicenter Group (GMMG) MM5 trial received a consolidation therapy with two cycles of lenalidomide after autologous stem cell transplantation (ASCT). Half of the study population continued treatment with lenalidomide maintenance therapy for 2 y, while the other patients received lenalidomide maintenance therapy until complete remission. We analyzed 58 patients with (n = 30) or without (n = 28) lenalidomide therapy and 12 patients refractory to lenalidomide with regards to their anti-myeloma-specific T-cell responses displayed by IFNγ, Granzyme B, and Perforin secretion. The immunophenotype of T-cells was investigated by flow cytometry. Significantly, more myeloma-specific T-cell responses were observed in patients during lenalidomide therapy, compared to patients without treatment. Furthermore, we found on T-cells from patients treated with lenalidomide a decreased CD45RA expression, indicating a maturated immunophenotype and a decreased expression of CD57, indicating functional T cells. An improved myeloma-specific T-cell response was observed in 6 out of 12 heavily pretreated patients (refractory to lenalidomide) after in vitro incubation with lenalidomide. Complementary to the results in vivo, lenalidomide decreased CD45RA expression on T cells in vitro.
KEYWORDS: Antigen-specific T-cells, CD45RA, CD57, immunotherapy, lenalidomide, multiple myeloma
Abbreviations
- ASCT
autologous stem cell transplantation
- CD45RA
cluster of designation 45 RA
- CCR7
chemokine receptor 7
- DC
dendritic celly
- ELISA
enzyme linked immunosorbent assay
- EliSpot
enzyme linked immunospot assay
- FACS
fluorescence-activated cell sorting
- Foxp3
forkhead-box-protein p3
- GMMG MM5
German Speaking Myeloma Multicenter Group
- GM-CSF
granulocyte macrophage colony-stimulating factor
- HLA-A2+
human leukocyte antigen serotype-A2+
- HLA-DR
human leukocyte antigen - antigen D related
- IFNγ
interferon-gamma
- IL
interleukin
- MM
multiple myeloma
- PD-1
programmed death-1
- PBMC
peripheral blood mononuclear cell
- TNF
tumor necrosis factor
Introduction
Lenalidomide plus dexamethasone is an established therapy for relapsed and newly diagnosed multiple myeloma (MM). Aside from the known classical tumoricidal effects of lenalidomide, recent work has focused on the immunomodulatory capacity of lenalidomide.
In vitro immunomodulation of lenalidomide
The immunostimulatory impact of lenalidomide on natural killer cells and T cells in patients with MM has been extensively described.1-7 In 2004, LeBlanc et al.6 found that lenalidomide increased T-cell co-stimulation via the B7-CD28 pathway, while Galustian showed that function and proliferation of regulatory T cells was inhibited by lenalidomide.5
Recently, Luptakova et al.7 demonstrated that upon unspecific activation of T cells with CD3/CD28 beads, the presence of lenalidomide resulted in Th1 polarization and increased interferon (IFN)-γ secretion by T cells. In addition, they showed that the proliferative response to allogenic dendritic cells (DC) was augmented by lenalidomide. De Keersmaecker et al.8 recently described the impact of lenalidomide on improving the quality of DC from MM patients by upregulation of costimulatory ligands. Of importance, regarding the expression of inhibitory molecules on T cells, the authors demonstrated that programmed death-1 (PD-1), an antigen that is upregulated in MM in terms of an immune escape mechanism, could be downregulated by lenalidomide in vitro.
Concerning the mode of action of lenalidomide, emerging data has suggested that Cereblon, a member of the cullin 4 ring ligase complex, is the molecular target of lenalidomide and pomalidomide on T cells and may be responsible for the observed immunomodulatory effects.9-12
In vivo immunomodulation of lenalidomide
Only a few reports have highlighted the immunomodulatory effect of lenalidomide in vivo. In relapsed MM patients, Harada et al. showed that lenalidomide has the capacity to enhance the production of inflammatory cytokines 8,13 while Busch et al. 14 identified activating, as well as inhibitory components of lenalidomide-induced immunomodulation. In the allogenic setting, lenalidomide causes a higher frequency of graft-versus-host disease, likely due to the observed activation of T- and natural killer cells.15-17
Recent clinical trials have analyzed the impact of lenalidomide as an induction therapy for newly diagnosed MM patients and as maintenance therapy after high-dose chemotherapy and ASCT. While the cytotoxic effects of lenalidomide in induction chemotherapy might be initially important for achieving rapid remission, the immunomodulatory effects of lenalidomide maintenance therapy might be more important in improving remission status disease after ASCT.
Recently, Govindaraj et al.18 demonstrated that lenalidomide maintenance therapy in patients with acute myeloid leukemia reduced TNF Receptor 2 on CD4+ T cells and enhanced the effector cytokine expression of T cells. In patients with MM, Clave et al. 19 showed a change of the T-cell immunophenotype during lenalidomide maintenance therapy, with a decreased CD8+CD45RA+CCR7− T-cell compartment.
We recently described that in an in vitro model, lenalidomide enhances antigen-specific T-cell response to a peptide from the MART-1 tumor-antigen (Melan-Aaa26–35*A27L) that cross reacts with the HM1.24aa22-30 myeloma antigen, which is expressed overall on malignant and non-malignant plasma cells.20-23 Aim of this study was to verify our in vitro results in vivo, because Chan et al.24 previously demonstrated that in a setting with natural killer cells, the impact of lenalidomide observed in vitro was not evident in vivo. Therefore, we analyzed the antigen-specific T-cell response against HM1.24 as a model for myeloma-specific T-cell responses in patients treated with lenalidomide.
A total of 51 patients enrolled in the GMMG MM5 clinical trial was assessed with regards to their anti-myeloma-specific T-cell responses and their T-cell immunophenotype. Antigen-specific T-cell activation using a DC-based protocol was analyzed by IFNγ, Granzyme B, and Perforin secretion using ELISA and EliSpot assay in HLA-A2+ MM patients. Furthermore, the expression of T-cell differentiation markers (CD45RA, CCR7, CD28 and CD279) was measured by flow cytometry. Eleven patients enrolled in the GMMG MM5 trial were also analyzed before and during lenalidomide therapy, including consolidation and maintenance therapy with lenalidomide. In addition, we included seven patients after ASCT who were not part of the GMMG MM5 trial, and who had not received lenalidomide therapy as control group.
Additionally, we analyzed 12 heavily pretreated HLA-A2+ patients refractory to lenalidomide, with regards to their myeloma-specific T-cell response and T-cell immunophenotype after in vitro T-cell stimulation with lenalidomide.
Results
Impact of lenalidomide therapy on the activation of T cells
The myeloma-specific T-cell response against the HM1.24 myeloma antigen was analyzed in MM patients with and without lenalidomide maintenance therapy in an antigen-specific model. Depending on the availability and amount of the peripheral blood samples, we performed IFNγ ELISpot-assays, IFNγ-, Granzyme B and Perforin-ELISAs. A patient with a positive antigen-specific T-cell response was defined by a significant T-cell response in at least one of the performed tests. We found an increased frequency of HM1.24aa22-30 specific T-cell responses in patients treated with lenalidomide (n = 13) compared to patients without lenalidomide treatment (n = 21) (p = 0.029, Fig. 1, supplemental data 1).
Figure 1.

Lenalidomide therapy enhances the antigen-specific immune response of CD8+ T cells from patients with multiple myeloma. The number of patients with specific or no specific immune response treated with or without lenalidomide is shown.
Impact of lenalidomide therapy on the expression of T-cell markers
We analyzed the T-cell compartment in patients with and without lenalidomide therapy and found no significant differences in the relative amount of CD3+ cells, CD8+ or CD4+ T cells (supplemental data 1). Regarding the modulation of T-cell immunophenotype and maturation, we recently showed that lenalidomide decreases the expression of CD45RA in vitro in patients with MM.23 Therefore, in this study we analyzed the expression of CD45RA, CD28 and CCR7, a set of maturation markers, on CD8+ T cells by flow cytometry. To estimate the activation status, we investigated the expression of CD38, CD154, HLA-DR, and CD57 on CD8+ T cells. In addition, we determined the expression of PD-1 (CD279) on the surface of CD8+ T cells, as recent publications demonstrated that PD-1 expressed on MM cells might deliver an inhibitory signal to T cells via PD-1 expressed on T cells 25 and that the expression of PD-1 can be downregulated by lenalidomide in vitro.7
We found in our in vitro studies that lenalidomide significantly downregulated the expression of CD45RA in vivo in patients with MM (n = 34, Fig. 2A). However, we found no correlation between the expression of CD28 (n = 37), CCR7 (n = 37), PD-1 (n = 39) CD38 (n = 19), CD154 (n = 19) and HLA-DR (n = 19) on the surface of CD8+ T cells and lenalidomide therapy (Fig. 2B, C, D, E, F and G). Of special interest, we found a significant downregulation of CD57 on CD8+ T-cells in the lenalidomide group (n = 19, Fig. 2H). For CD45RA, CD28, CCR7 and PD-1, we also analyzed the CD4+ T-cell compartment but found no significant differences between the two groups (data not shown).
Figure 2.
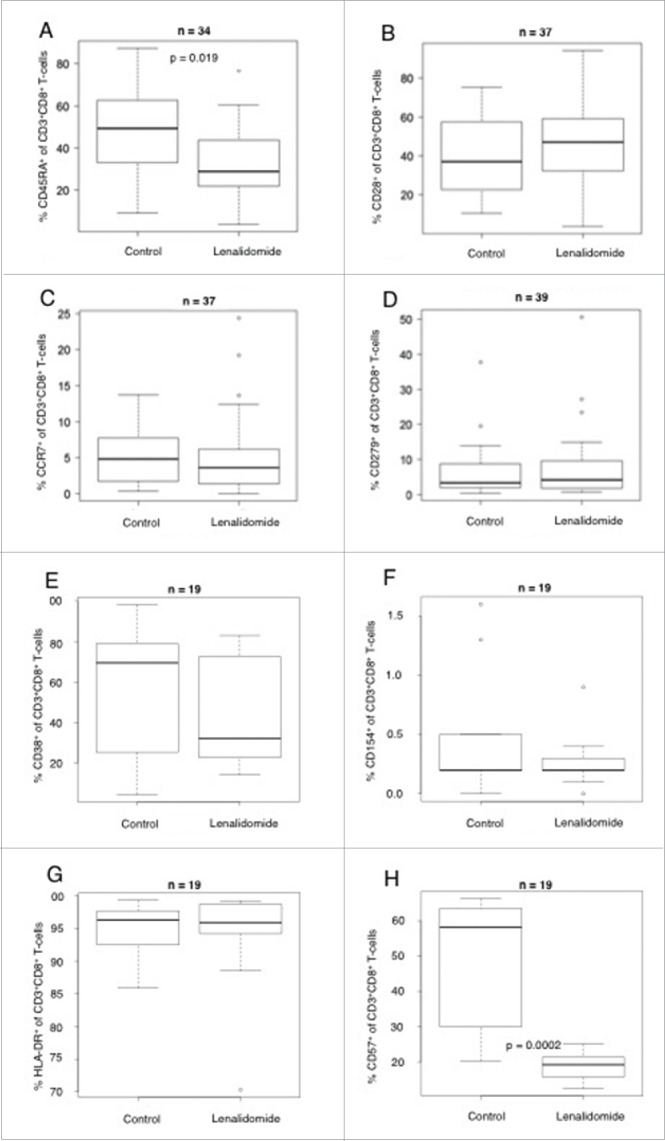
Impact of lenalidomide therapy on the expression of T-cell markers. Shown is the expression of (A) CD45RA, (B) CD28, (C) CCR7 and (D) CD279 (E) CD38, (F) CD154, (G) HLA-DR, (H) CD57 on CD8+ T cells (in % of all CD8+ T cells) from patients with MM treated with or without lenalidomide, analyzed by flow cytometry.
Impact of lenalidomide therapy on CD4+CD127dimCD25highFoxp3+ regulatory T cells
We assessed the percentage of CD4+CD127dimCD25highFoxp3+regulatory T-cells of CD4+ T cells in patients with or without lenalidomide treatment by flow cytometry. As shown in Fig. 3, we found an increase of these regulatory T cells in patients during lenalidomide therapy, likely due to the low patient number (n = 18) the difference between the two groups was not statistically significant (p = 0.115).
Figure 3.
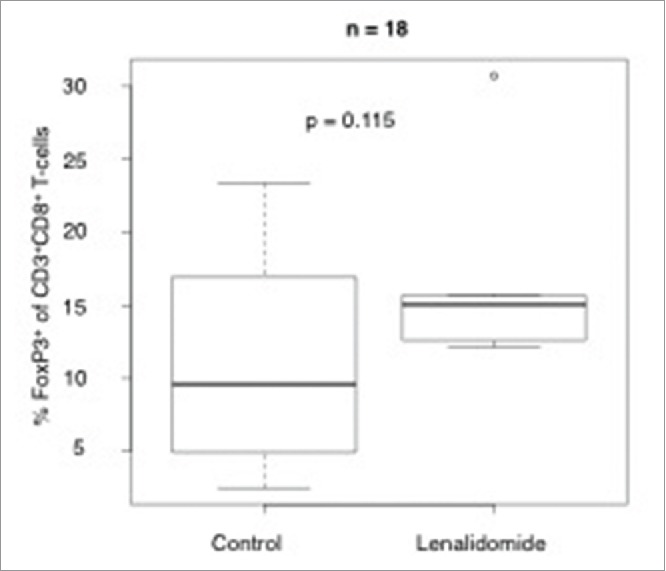
Impact of lenalidomide therapy on the CD4+CD127dimCD25high Foxp3+regulatory T-cell compartment. Shown is the percentage of CD4+CD127dimCD25highFoxp3+regulatory T cell of all CD4+ T cells in patients with MM treated with or without lenalidomide, analyzed by flow cytometry.
Impact of lenalidomide on the activation of T cells from lenalidomide-refractory patients in vitro
To determine if the immune system of patients who relapsed during or after lenalidomide treatment was still sensitive to lenalidomide in vitro, we investigated the T-cell responses against the HM1.24 antigen in 12 HLA-A2+ patients with refractory and advanced MM. Those heavily pretreated patients already had received a median of 6.4 (range 4–9) chemotherapeutic schedules. All investigated patients had relapsed after lenalidomide treatment, and most of the patients were analyzed in progressive disease (Table 2). We found that in 6 out of 12 patients, lenalidomide triggered a boosted immune response, defined as an enhanced immune response in at least one of the following assays: IFNγ EliSpot-assay or IFNγ, Granzyme B or Perforin ELISA. A representative EliSpot is shown in Fig. 4.
Table 2.
Patient characteristics (patients with lenalidomide-refractory multiple myeloma).
| Characteristics | |
|---|---|
| Total patients, n (%) | Number of patients, n (%) |
| Gender, n (%) | 12 (100%) |
| Female | 6 (50%) |
| Male | 6 (50%) |
| Median age at time of sampling, years (range) | 67.6 (47–85) |
| Median number of pretreatment regimens, n (range) | 8 (7–11) |
| Type, n (%) | |
| IgG Kappa | 3 (24.9%) |
| IgG Lambda | 1 (8.3%) |
| IgA Kappa | 2 (16.6%) |
| IgA Lambda | 2 (16.6%) |
| IgD Kappa | 1 (8.3%) |
| BJP Lambda | 1 (8.3%) |
| BJP Kappa | 1 (8.3%) |
| Hyposec. | 1 (8.3%) |
| ISS at time of sampling, n (%) | |
| I | 0 (0%) |
| II | 1 (8.3%) |
| III | 11 (91.7%) |
| Previous chemotherapy regiments (per patient), range | 6.4 (4–9) |
| Status at time of sampling, n (%) | |
| SD/MR | 1 (8.3%) |
| PR/VGPR | 1 (8.3%) |
| nCR/CR | 1 (8.3%) |
| PD | 9 (75.1%) |
| Immune response, n (%) | |
| Positive | 6 (50%) |
| Negative | 6 (50%) |
BJP, Bence–Jones protein; ISS, International Staging System; SD, stable disease; MR, minor response; PR, partial response; VGPR, very good partial response; nCR, near complete remission; CR, complete remission; PD, progressive disease.
Figure 4.
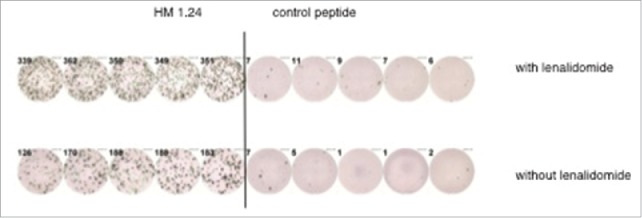
Representative EliSpot-analysis. The figure shows a representative EliSpot analysis from a lenalidomide refractory patient with T cells demonstrating an enhanced antigen-specific T-cell response against the myeloma antigen HM1.24 after in vitro incubation with lenalidomide.
Impact of lenalidomide on the expression of T-cell markers from lenalidomide-refractory patients in vitro
The expression of CD45RA on CD8+ T-cells from refractory patients was also downregulated after lenalidomide incubation, which was analogous to the in vivo data from patients in the MM5 trial (n = 5, p = 0.19, Fig. 5). No correlation between lenalidomide incubation and the expression of CD28 and CCR7 on CD8+ T cells from refractory patients was observed.
Figure 5.

Impact of lenalidomide in vitro in patients with refractory multiple myeloma on the expression of T-cell markers. The plot shows the expression of CD45RA on CD8+ T-cells (in % of all CD8+ T cells) from patients with refractory MM analyzed by flow cytometry.
Discussion
While lenalidomide is well established in the treatment of MM, its immunomodulatory mode of action in vivo is still elusive. Recent in vitro data has demonstrated the stimulatory impact of lenalidomide on T cells in MM patients, while in vivo data are rare. Our aim was to confirm recent in vitro results using data from lenalidomide treated MM patients enrolled in the MM5 clinical trial. Patient samples were obtained for immunological functional assays and for flow cytometry analysis of T-cell markers after high-dose chemotherapies and ASCT before and during consolidation or maintenance treatment with lenalidomide. Six patients, who were not part of the MM5 clinical trial and who had not undergone any lenalidomide therapy were also analyzed.
In the present study, we were able to confirm the experimental observation that lenalidomide enhances antigen-specific T-cell activation. We showed that patients with lenalidomide treatment had a higher frequency of antigen-specific T-cell responses compared to patients treated without lenalidomide. T-cell responses were analyzed by IFNγ-EliSpot-assay and IFNγ, Granzyme B and Perforin ELISA. Because of the limited volume of the peripheral blood samples, it was not possible to do all four assays in each patient. Furthermore, not all patients with a significant T-cell response showed a positive result in all performed assays. As such, we defined a patient as having a specific T-cell response if they had one significant positive assay. It is noteworthy, that we observed these enhanced T-cell responses although the number of CD4+CD127dimCD25highFoxp3+regulatory T-cells in our study was increased in patients with lenalidomide therapy, confirming recent data26,27 Next, we wanted to determine whether the immune system of patients who relapsed after or during lenalidomide therapy was still sensitive to lenalidomide. Therefore, we analyzed the impact of lenalidomide on T-cells from these patients in vitro, using the previously described antigen-specific model in 12 HLA-A2+ patients. Unexpectedly, we found that in six patients, lenalidomide enhanced the immune response, although most of the patients were in progressive disease at the time of analysis. These results show that although MM cells might be refractory to lenalidomide with regards to its cytotoxic mode of action, lenalidomide is still able to activate T cells in these patients. Nevertheless, these in vitro data do not necessarily reflect the in vivo situation, where MM cells from lenalidomide-refractory patients might escape the antigen-specific T-cell response by immunosuppressive mechanisms, even though the MM-specific T-cell response can be enhanced by lenalidomide.
We also assessed the impact of lenalidomide on the T-cell maturation markers, CD45RA, CD28, CCR7 and PD-1, and we confirmed previous in vitro findings 23 that the expression of CD45RA on CD8+ T cells was downregulated upon lenalidomide treatment. It is unclear whether this phenomenon contributes to the immunostimulatory effect of lenalidomide, especially because we recently showed that thalidomide can also reduce the expression of CD45RA on the surface of T cells, but did not observe any immunostimulatory effects of thalidomide in vivo.28
A component of our study that should be considered is the difference in the period from ASCT to peripheral blood sample analysis between patients with and without lenalidomide treatment. While much data exist on T-cell number and T-cell subset recovery after high dose chemotherapy and ASCT,29,30 only a few reports have analyzed T-cell function in this setting. Van der Velden et al.31 showed that T-cell-mediated immunity against the tetanus toxin in MM patients was impaired after ASCT, without recovery until 15 mo later. No time point of immune response reconstitution was determined in this report. Furthermore, they showed that the expression of CD45RA on T cells was constantly decreased in this period without recovery. As such, one could speculate that the diminished CD45RA expression we found during lenalidomide therapy is not due to a post-transplant phenomenon. Another important finding was the decrease of the CD8+ CD57+ T-cell compartment in patients during lenalidomide treatment. A few reports exist, that a CD8+ CD57+ T-cell compartment is expanded after ASCT in patients with MM.32 Based on the observation that this expansion was detectable for years after ASCT, it is unlikely that this result is due a longer follow up in the lenalidomde group in our study. While the expression of CD57 on T-cells is considered as a marker of senescent and functional deficient T cells,33 these data might indicate that lenalidomide is able to restore those T cells.
Summarizing, we found that lenalidomide enhances T-cell responses in vivo and unexpectedly has also immunostimulatory capacity on T cells of patients who are refractory to lenalidomide in vitro.
Materials and methods
Patients
In vivo analysis
To analyze T cells from patients with MM before and during lenalidomide consolidation and maintenance therapy, 50 mL of peripheral blood was drawn after obtaining written informed consent. Approval for the use of patients' peripheral blood mononuclear cells (PBMCs) was obtained from the ethics committee of the University of Heidelberg (approval No. 13—0023).
A total of 58 patients participated in the study, of which 51 were patients in the MM5 trial of the GMMG. Patients enrolled in the GMMG MM5 trial were randomized to receive induction therapy with bortezomib, cyclophosphamide and dexamethasone or bortezomib, doxorubicin and dexamethasone. After dose intensification with single or double high-dose melphalan and ASCT according to remission after first transplant, the patients received a consolidation therapy with two cycles of lenalidomide (25 mg on day 1–21, cycle duration 28 d). Half of the study population continued treatment with lenalidomide maintenance therapy for 2 y (10 mg daily for 3 mo, if tolerable dose escalation to 15 mg/d from the 4th mo), while the other patients received lenalidomide maintenance therapy until complete remission. In addition, in this analysis, we included seven patients who were not in the GMMG MM5 trial and did not receive lenalidomide therapy. The patients' characteristics are listed in Table 1.
Table 1.
Patient characteristics (newly diagnosed patients).
| Characteristics | |
|---|---|
| Total patients, n (%) | Number of patients, n (%) |
| Gender, n (%) | 58 (100%) |
| Female | 24 (41.4%) |
| Male | 34 (58.6%) |
| Median age at begin of induction, years (range) | 56.4 (35.0–71.8) |
| Type, n (%) | |
| IgG Kappa | 21 (36.2%) |
| IgG Lambda | 13 (22.4%) |
| IgA Kappa | 9 (15.5%) |
| IgA Lambda | 4 (6.9%) |
| IgD Lambda | 1 (1.7%) |
| BJP Kappa | 7 (12.1%) |
| BJP Lambda | 3 (5.2%) |
| Median initial β-2 microglobulin, mg/L (range) | 3.0 (1.6–226.0) |
| ISS at first diagnosis, n (%) | |
| I | 34 (58.6%) |
| II | 14 (24.1%) |
| III | 9 (15.5%) |
| n.a. | 1 (1.7%) |
| CRAB symptoms at first diagnosis, n (%) | |
| Hypercalcemia alone | 0 (0%) |
| Renal insufficiency | 2 (3.4%) |
| Anemia | 7 (12.1%) |
| Bone lesions | 39 (67.2%) |
| Hypercalcemia and bone lesions | 1 (1.7%) |
| Hypercalcemia, renal insufficiency and bone lesions | 1 (1.7%) |
| Renal insufficiency, anemia and bone lesions | 2 (3.4%) |
| Anemia and lytic bone lesions | |
| High-risk cytogenetics, n (%) | |
| del17p13 | 6 (10.3%) |
| t(4;14) | 4 (6.9%) |
| dupl 1q21 | 8 (13.8%) |
| del17p13 and t(4;14) | 3 (5.2%) |
| Induction therapy, n (%) | 1 (1.7%) |
| 3xPAd | 25 (43.1%) |
| 3xVCD | 33 (56.9%) |
| Number of autologous stem cell transplantation, n (%) | |
| 0 | 1 (1.7%) |
| 1 | 35 (60.3%) |
| 2 | 22 (37.9%) |
| Remission status before consolidation therapy, n (%) | |
| SD/MR | 1 (1.7%) |
| PR/VGPR | 24 (41.4%) |
| nCR/CR | 33 (56.9%) |
| Immune response, n (%) | |
| Positive | 19 (32.8%) |
| Negative | 15 (25.9%) |
| N.A. | 24 (41.4%) |
| Treatment with lenalidomide, n (%) | |
| no | 28 (48.3%) |
| yes | 30 (51.7%) |
| Median cumulative dose of lenalidomide, mg (range) | 2310 (525–8610) |
BJP, Bence–Jones protein; ISS, International Staging System; n.a., not available; del, deletion; t, translocation; dupl, duplication; SD, stable disease; MR, minor response; PR, partial response; VGPR, very good partial response; nCR, near complete remission; CR, complete remission; mg, milligram.
In vitro analysis
To determine whether the anti-myeloma-specific T-cell response could be enhanced by lenalidomide, even in lenalidomide-refractory patients, we analyzed the T cells from 12 MM patients with advanced and heavily pretreated disease. The patient's characteristics are listed in Table 2.
PBMCs for the in vitro expansion of peptide-specific T cells
PBMCs from patients with MM were purified using density centrifugation (Biochrom, Berlin, Germany). HLA-A typing was performed by flow cytometry, as previously described.34 To generate antigen-specific T cells, PBMCs were incubated for 7 d together with autologous DC (ratio: 1:15) in a T-cell medium containing IL-2 at a concentration of 50 IE/mL.
Synthesis of peptides
The Melan-Aaa26–35*A27L peptide (amino-acid sequence: ELAGIGILTV) was synthesized by the peptide-synthesis-department of the German Cancer Research Center Heidelberg (DKFZ, Heidelberg, Germany) using standard procedures.
In vitro generation of DC
PBMCs from HLA-A2+ MM patients were used. Immature DCs were obtained by culturing plastic adherent PBMCs for 5 d with GM-CSF (800 U/mL, Molgramostim, Essex Pharma, München, Germany) and IL-4 (500 U/mL, R&D). Afterwards, differentiation into mature DC was induced by stimulation with TNF-α (10 ng/mL, Sigma-Aldrich, Deisenhofen, Germany), IL-6 (1000 U/mL, R&D systems, Abingdon, Oxon, United Kingdom) and prostaglandin E2 (1 µg/mL, Sigma-Aldrich, Deisenhofen, Germany) for 2 d, as previously published.35
IFNγ EliSpot-assay
The generation of Melan-Aaa26–35*A27L peptide-specific CD8+ cells was investigated using an IFNγ EliSpot-assay. Expanded CD8+ cells (2 × 104 / well) were purified with immunomagnetic beads (MACS-system, Miltenyi Biotec, Bergisch Gladbach, Germany) and incubated with peptide-loaded T2 cells as targets (effector/target-ratio 1:5) for 48 h in anti-IFNγ-antibody (Mabtech AB, Nacka, Sweden) coated nitrocellulose-plates (Millipore, Eschborn, Germany). After detection with biotinylated anti-cytokine-antibodies (Mabtech AB, Nacka, Sweden) and conjugation with Avidin ALP (Sigma, Deisenhofen, Germany), the BCIP/NBT substrate was added (Sigma, Deisenhofen, Germany). EliSpots were counted with a CTL ImmunoSpot Analyzer. T2 cells were loaded with Melan-Aaa26–35*A27L or a HLA-A2 restricted control peptide (amino-acid sequence: LLIIVILGV) during two-hour incubation with 10 µg/mL of peptide according to LeBlanc et al.6
ELISA
Cytokine secretion of activated T cells was analyzed in the EliSpot supernatants by ELISA. Commercial ELISA kits were used to measure the secreted cytokines, IFNγ (BD Biosciences, Heidelberg, Germany), Granzyme B- and Perforin (both from Mabtech AB, Nacka, Sweden), according to the manufacturer's instructions. Briefly, a monoclonal antibody specific for IFNγ, Granzyme B- or Perforin respectively was pre-coated onto a microplate (Greiner Bio-One, Frickenhausen, Germany). Standards and samples were added into the wells, and any cytokine present was bound by the immobilized antibody. After washing to remove the unbound substances, an enzyme-linked monoclonal antibody specific for IFNγ, Granzyme B- or Perforin was added to the wells. Following washing to remove any unbound antibody-enzyme reagent, a substrate solution (TMB substrate set, BD Biosciences) was added to the wells, leading to differential coloring, in proportion to the amount of IFNγ, Granzyme B- and Perforin. The intensity of the color was analyzed with an automated plate reader.
Flow cytometry
The expression of surface T-cell markers was analyzed by flow cytometry. Cells were resuspended in saponin buffer and incubated with fluorochrome-labeled antibodies against CD3, CD8+, CD28, CD45-RA, CD279, CCR7, CD38, CD154, HLA-DR, CD25, CD127, CD4, CD57 and Foxp3 (all from BD Biosciences, Heidelberg, Germany, except CD25 was purchased from Dako, Glostrup, Denmark and FoxP3 from ebioscience, San Diego, USA), according to the manufacturer's instructions. Control cells were stained with the corresponding isotype antibodies. After a final wash with saponin buffer, the cells were resuspended in PBS. Flow cytometry analysis was performed with an FACS Calibur flow cytometer (BD Biosciences). The CD4+CD127dimCD25highFoxp3+regulatory T-cell compartment was analyzed by flow cytometry as described elsewhere.28
Statistics
The statistical significance of the difference between T cells stimulated by MART-1-loaded T2 cells with and without lenalidomide was calculated for EliSpot and ELISA by Student's t-test. Each EliSpot and ELISA assay was performed in five wells. An example for a representative Elispot analysis is shown in Fig. 4. The Student's t-test was significant if p values were ≤ 0.05. Non-significant p values > 0.05 and ≤ 0.5 are reported for descriptive reasons only. Four square analysis and fisher test was used to calculate differences regarding the statistical significance of the difference in antigen-specific T-cell response between patients with or without lenalidomide maintenance therapy.
To analyze the statistical significance of the difference of expression of T-cell markers between the two groups, we used Wilcoxon test.
Supplementary Material
Disclosure of potential conflict of interest
M.H. has received research funding from Celgene. The remaining authors declare no competing financial interests. H.G. has received research funding from Celgene, Janssen and Chugai.
Funding
This work was supported by Celgene GmbH.
References
- 1.Chang DH, Liu N, Klimek V, Hassoun H, Mazumder A, Nimer SD, Jagannath S, Dhodapkar MV. Enhancement of ligand-dependent activation of human natural killer T cells by lenalidomide: therapeutic implications. Blood 2006; 108:618–21; PMID:16569772; http://dx.doi.org/ 10.1182/blood-2005-10-4184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Davies FE, Raje N, Hideshima T, Lentzsch S, Young G, Tai YT, Lin B, Podar K, Gupta D, Chauhan D et al.. Thalidomide and immunomodulatory derivatives augment natural killer cell cytotoxicity in multiple myeloma. Blood 2001; 98:210–6; PMID:11418482; http://dx.doi.org/ 10.1182/blood.V98.1.210 [DOI] [PubMed] [Google Scholar]
- 3.Hsu AK, Quach H, Tai T, Prince HM, Harrison SJ, Trapani JA, Smyth MJ, Neeson P, Ritchie DS. The immunostimulatory effect of lenalidomide on NK-cell function is profoundly inhibited by concurrent dexamethasone therapy. Blood 2011; 117:1605–13; PMID:20978269; http://dx.doi.org/ 10.1182/blood-2010-04-278432 [DOI] [PubMed] [Google Scholar]
- 4.Song W, van der Vliet HJ, Tai YT, Prabhala R, Wang R, Podar K, Catley L, Shammas MA, Anderson KC, Balk SP et al.. Generation of antitumor invariant natural killer T cell lines in multiple myeloma and promotion of their functions via lenalidomide: a strategy for immunotherapy. Clin Cancer Res 2008; 14:6955–62; PMID:18980990; http://dx.doi.org/ 10.1158/1078-0432.CCR-07-5290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Galustian C, Meyer B, Labarthe MC, Dredge K, Klaschka D, Henry J, Todryk S, Chen R, Muller G, Stirling D et al.. The anti-cancer agents lenalidomide and pomalidomide inhibit the proliferation and function of T regulatory cells. Cancer Immunol Immunother 2009; 58:1033–45; PMID:19009291; http://dx.doi.org/ 10.1007/s00262-008-0620-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.LeBlanc R, Hideshima T, Catley LP, Shringarpure R, Burger R, Mitsiades N, Mitsiades C, Cheema P, Chauhan D, Richardson PG et al.. Immunomodulatory drug costimulates T cells via the B7-CD28 pathway. Blood 2004; 103:1787–90; PMID:14512311; http://dx.doi.org/ 10.1182/blood-2003-02-0361 [DOI] [PubMed] [Google Scholar]
- 7.Luptakova K, Rosenblatt J, Glotzbecker B, Mills H, Stroopinsky D, Kufe T, Vasir B, Arnason J, Tzachanis D, Zwicker JI et al.. Lenalidomide enhances anti-myeloma cellular immunity. Cancer Immunol Immunother 2013; 62:39–49; PMID:22733396; http://dx.doi.org/ 10.1007/s00262-012-1308-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De Keersmaecker B, Fostier K, Corthals J, Wilgenhof S, Heirman C, Aerts JL, Thielemans K, Schots R. Immunomodulatory drugs improve the immune environment for dendritic cell-based immunotherapy in multiple myeloma patients after autologous stem cell transplantation. Cancer Immunol Immunother 2014; 63:1023–36; PMID:24947180; http://dx.doi.org/ 10.1007/s00262-014-1571-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gandhi AK, Kang J, Havens CG, Conklin T, Ning Y, Wu L, Ito T, Ando H, Waldman MF, Thakurta A et al.. Immunomodulatory agents lenalidomide and pomalidomide co-stimulate T cells by inducing degradation of T cell repressors Ikaros and Aiolos via modulation of the E3 ubiquitin ligase complex CRL4(CRBN). Br J Haematol 2014 Mar; 164(6):811-21; PMID:24328678; http://dx.doi.org/ 10.1111/bjh.12708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lopez-Girona A, Mendy D, Ito T, Miller K, Gandhi AK, Kang J, Karasawa S, Carmel G, Jackson P, Abbasian M et al.. Cereblon is a direct protein target for immunomodulatory and antiproliferative activities of lenalidomide and pomalidomide. Leukemia 2012; 26:2326–35; PMID:22552008; http://dx.doi.org/ 10.1038/leu.2012.119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhu YX, Braggio E, Shi CX, Bruins LA, Schmidt JE, Van Wier S, Chang XB, Bjorklund CC, Fonseca R, Bergsagel PL et al.. Cereblon expression is required for the antimyeloma activity of lenalidomide and pomalidomide. Blood 2011; 118:4771–9; PMID:21860026; http://dx.doi.org/ 10.1182/blood-2011-05-356063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sehgal K, Das R, Zhang L, Verma R, Deng Y, Kocoglu M, Vasquez J, Koduru S, Ren Y, Wang M et al.. Clinical and pharmacodynamic analysis of pomalidomide dosing strategies in myeloma: impact of immune activation and cereblon targets. Blood 2015; 125:4042–51; PMID:25869284; http://dx.doi.org/ 10.1182/blood-2014-11-611426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ozaki S, Harada T, Fujii S, Nakamura S, Miki H, Nakano A, Kagawa K, Takeuchi K, Abe M, Matsumoto T. Transient inflammatory reaction during lenalidomide plus reduced-dose dexamethasone therapy in two patients with relapsed multiple myeloma. Int J Hematol 2011; 93:257–9; PMID:21279817; http://dx.doi.org/ 10.1007/s12185-011-0775-1 [DOI] [PubMed] [Google Scholar]
- 14.Busch A, Zeh D, Janzen V, Mugge LO, Wolf D, Fingerhut L, Hahn-Ast C, Maurer O, Brossart P, von Lilienfeld-Toal M. Treatment with lenalidomide induces immunoactivating and counter-regulatory immunosuppressive changes in myeloma patients. Clin Exp Immunol 2014; 177:439–53; PMID:24712857; http://dx.doi.org/ 10.1111/cei.12343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kneppers E, van der Holt B, Kersten MJ, Zweegman S, Meijer E, Huls G, Cornelissen JJ, Janssen JJ, Huisman C, Cornelisse PB et al.. Lenalidomide maintenance after nonmyeloablative allogeneic stem cell transplantation in multiple myeloma is not feasible: results of the HOVON 76 Trial. Blood 2011; 118:2413–9; PMID:21690556; http://dx.doi.org/ 10.1182/blood-2011-04-348292 [DOI] [PubMed] [Google Scholar]
- 16.Wolschke C, Stubig T, Hegenbart U, Schonland S, Heinzelmann M, Hildebrandt Y, Ayuk F, Atanackovic D, Dreger P, Zander A et al.. Postallograft lenalidomide induces strong NK cell-mediated antimyeloma activity and risk for T cell-mediated GvHD: Results from a phase I/II dose-finding study. Experimental Hemato 2013; 41:134-42 e3; PMID:23085463; http://dx.doi.org/ 10.1016/j.exphem.2012.10.004 [DOI] [PubMed] [Google Scholar]
- 17.Alsina M, Becker PS, Zhong X, Adams A, Hari P, Rowley S, Stadtmauer EA, Vesole DH, Logan B, Weisdorf D et al.. Lenalidomide maintenance for high-risk multiple myeloma after allogeneic hematopoietic cell transplantation. Biol Blood Marrow Transplant 2014; 20:1183–9; PMID:24769014; http://dx.doi.org/ 10.1016/j.bbmt.2014.04.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Govindaraj C, Madondo M, Kong YY, Tan P, Wei A, Plebanski M. Lenalidomide-based maintenance therapy reduces TNF receptor 2 on CD4 T cells and enhances immune effector function in acute myeloid leukemia patients. Am J Hematol 2014 Aug; 89(8):795-802; PMID:24757092; http://dx.doi.org/ 10.1002/ajh.23746 [DOI] [PubMed] [Google Scholar]
- 19.Clave E, Douay C, Coman T, Busson M, Bompoint C, Moins-Teisserenc H, Glauzy S, Carmagnat M, Gorin NC, Toubert A et al.. Lenalidomide consolidation and maintenance therapy after autologous stem cell transplant for multiple myeloma induces persistent changes in T-cell homeostasis. Leuk Lymphoma 2013 Aug; 55(8):1788-95; PMID:24237448; http://dx.doi.org/ 10.3109/10428194.2013.865182 [DOI] [PubMed] [Google Scholar]
- 20.Harada T, Ozaki S, Oda A, Tsuji D, Ikegame A, Iwasa M, Udaka K, Fujii S, Nakamura S, Miki H et al.. Combination with a defucosylated anti-HM1.24 monoclonal antibody plus lenalidomide induces marked ADCC against myeloma cells and their progenitors. PLoS One 2013 Dec 26; 8(12):e83905; PMID:24386306; http://dx.doi.org/ 10.1371/journal.pone.0083905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ozaki S, Kosaka M, Harada M, Nishitani H, Odomi M, Matsumoto T. Radioimmunodetection of human myeloma xenografts with a monoclonal antibody directed against a plasma cell specific antigen, HM1.24. Cancer 1998; 82:2184–90; PMID:9610698; http://dx.doi.org/ 10.1002/(SICI)1097-0142(19980601)82:11%3c2184::AID-CNCR13%3e3.0.CO;2-Q [DOI] [PubMed] [Google Scholar]
- 22.Ozaki S, Kosaka M, Wakatsuki S, Abe M, Koishihara Y, Matsumoto T. Immunotherapy of multiple myeloma with a monoclonal antibody directed against a plasma cell-specific antigen, HM1.24. Blood 1997; 90:3179–86; PMID:9376601 [PubMed] [Google Scholar]
- 23.Neuber B, Herth I, Tolliver C, Schoenland S, Hegenbart U, Hose D, Witzens-Harig M, Ho AD, Goldschmidt H, Klein B et al.. Lenalidomide enhances antigen-specific activity and decreases CD45RA expression of T cells from patients with multiple myeloma. J Immunol 2011; 187:1047–56; PMID:21677134; http://dx.doi.org/ 10.4049/jimmunol.1002460 [DOI] [PubMed] [Google Scholar]
- 24.Chan AC, Neeson P, Leeansyah E, Tainton K, Quach H, Prince HM, Harrison SJ, Godfrey DI, Ritchie D, Berzins SP. Natural killer T cell defects in multiple myeloma and the impact of lenalidomide therapy. Clin Exp Immunol 2014; 175:49–58; PMID:24032527; http://dx.doi.org/ 10.1111/cei.12196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hallett WH, Jing W, Drobyski WR, Johnson BD. Immunosuppressive effects of multiple myeloma are overcome by PD-L1 blockade. Biol Blood Marrow Transplant 2011; 17:1133–45; PMID:21536144; http://dx.doi.org/ 10.1016/j.bbmt.2011.03.011 [DOI] [PubMed] [Google Scholar]
- 26.Clave E, Douay C, Coman T, Busson M, Bompoint C, Moins-Teisserenc H, Glauzy S, Carmagnat M, Gorin NC, Toubert A et al.. Lenalidomide consolidation and maintenance therapy after autologous stem cell transplant for multiple myeloma induces persistent changes in T-cell homeostasis. Leuk Lymphoma 2014; 55:1788–95; PMID:24237448; http://dx.doi.org/ 10.3109/10428194.2013.865182 [DOI] [PubMed] [Google Scholar]
- 27.Minnema MC, van der Veer MS, Aarts T, Emmelot M, Mutis T, Lokhorst HM. Lenalidomide alone or in combination with dexamethasone is highly effective in patients with relapsed multiple myeloma following allogeneic stem cell transplantation and increases the frequency of CD4+Foxp3+ T cells. Leukemia 2009; 23:605–7; PMID:18784738; http://dx.doi.org/ 10.1038/leu.2008.247 [DOI] [PubMed] [Google Scholar]
- 28.Herth I, Witzens-Harig M, Beckhove P, Hose D, Meissner T, Neuber B, Engelhardt M, Haas J, Neben K, Ho AD et al.. Thalidomide maintenance therapy maturates the T cell compartment and compromises antigen-specific antitumor immunity in patients with multiple myeloma. Exp Hematol 2013; 41:231–40; PMID:23142220; http://dx.doi.org/ 10.1016/j.exphem.2012.10.018 [DOI] [PubMed] [Google Scholar]
- 29.Nachbaur D, Kropshofer G, Heitger A, Latzer K, Glassl H, Ludescher C, Nussbaumer W, Niederwieser D. Phenotypic and functional lymphocyte recovery after CD34+-enriched vs. non-T cell-depleted autologous peripheral blood stem cell transplantation. J Hematother Stem Cell Res 2000; 9:727–36; PMID:11091497; http://dx.doi.org/ 10.1089/15258160050196777 [DOI] [PubMed] [Google Scholar]
- 30.Peggs KS, Verfuerth S, Pizzey A, Khan N, Moss P, Goldstone AH, Yong K, Mackinnon S. Reconstitution of T-cell repertoire after autologous stem cell transplantation: influence of CD34 selection and cytomegalovirus infection. Biol Blood Marrow Transplant 2003; 9:198–205; PMID:12652471; http://dx.doi.org/ 10.1016/S1083-8791(03)70010-X [DOI] [PubMed] [Google Scholar]
- 31.van der Velden AM, Claessen AM, van Velzen-Blad H, Biesma DH, Rijkers GT. Development of T cell-mediated immunity after autologous stem cell transplantation: prolonged impairment of antigen-stimulated production of gamma-interferon. Bone Marrow Transplant 2007; 40:261–6; PMID:17563737; http://dx.doi.org/ 10.1038/sj.bmt.1705706 [DOI] [PubMed] [Google Scholar]
- 32.Wolniak KL, Goolsby CL, Chen YH, Chenn A, Singhal S, Jayesh Mehta, Peterson LA. Expansion of a clonal CD8+CD57+ large granular lymphocyte population after autologous stem cell transplant in multiple myeloma. Am J Clin Pathol 2013; 139:231–41; PMID:23355208; http://dx.doi.org/ 10.1309/AJCP1T0JPBLSLAQF [DOI] [PubMed] [Google Scholar]
- 33.Focosi D, Bestagno M, Burrone O, Petrini M. CD57+ T lymphocytes and functional immune deficiency. J Leukoc Biol 2010; 87:107–16; PMID:19880576; http://dx.doi.org/ 10.1189/jlb.0809566 [DOI] [PubMed] [Google Scholar]
- 34.Hundemer M, Schmidt S, Condomines M, Lupu A, Hose D, Moos M, Cremer F, Kleist C, Terness P, Belle S et al.. Identification of a new HLA-A2-restricted T-cell epitope within HM1.24 as immunotherapy target for multiple myeloma. Experimental Hematol 2006; 34:486–96; PMID:16569595; http://dx.doi.org/ 10.1016/j.exphem.2006.01.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Blade J, Samson D, Reece D, Apperley J, Bjorkstrand B, Gahrton G, Gertz M, Giralt S, Jagannath S, Vesole D. Criteria for evaluating disease response and progression in patients with multiple myeloma treated by high-dose therapy and haemopoietic stem cell transplantation. Myeloma Subcommittee of the EBMT. European Group for Blood and Marrow Transplant. Br J Haematol 1998; 102:1115-23; PMID:9753033; http://dx.doi.org/ 10.1046/j.1365-2141.1998.00930.x [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.