

Supplemental digital content is available in the text.
Abstract
Acute cellular rejection (ACR) is a common complication in lung transplantation and associated with increased risk of chronic allograft dysfunction. MicroRNAs are critical controllers of cellular transcription whose expression can be altered in disease states. The purpose of this pilot study was to evaluate whether microRNA profiling of epithelial cells obtained from airway brushings can distinguish lung transplant patients with ACR from those without rejection. We studied 21 subjects (10 with ACR, 11 without ACR) and assessed the expression of over 700 microRNAs in their airway epithelium. We identified 117 differentially expressed microRNAs that robustly segregated the 2 groups and were uniformly downregulated in patients with ACR. Leveraging experimentally verified microRNA targets, we systematically mapped pathways and processes regulated by ACR-induced microRNAs and noted enrichment of programs involved in development, proliferation, migration, and repair. Collectively, our study suggests that ACR is associated with a distinct epithelial microRNA signature that can provide insight into the pathogenesis of acute rejection and potentially serve as a sensitive, minimally invasive biomarker tool for diagnostic and prognostic stratification of lung transplant patients.
The development of immunosuppressive medications, such as cyclosporine, has created the possibility for lung transplantation for individuals with end-stage lung disease.1 However, the level of immunosuppression must be tempered to avoid severe infectious complications associated with the immunocompromised state. Consequently, about a third of recipients proceed to develop acute cellular rejection (ACR) in the first year after lung transplantation.2 Identifying and effectively treating ACR is vital because of its adverse long-term impact on graft function. Indeed, the severity and frequency of ACR is directly proportional to the risk of developing bronchiolitis obliterans syndrome, the most common manifestation of chronic lung allograft dysfunction.1 Despite intensive research, the pathogenetic mechanisms leading to acute rejection remain poorly understood and have limited our ability diagnose this important condition at an early stage. This is of particular concern because up to 39% lung transplant patients may have clinically silent ACR. Moreover, transbronchial biopsy has a 72% to 84% sensitivity for detecting ACR and can result in serious complications.3,4 Therefore, understanding the molecular mechanisms of acute rejection and finding new, less invasive methods to improve the sensitivity of its diagnosis can yield significant long-term beneficial outcomes.
MicroRNAs are small RNAs approximately 23 nucleotides in length that are major posttranscriptional controllers regulating the expression of nearly a third of the genome.5 Each microRNA can affect multiple targets and broadly influence diverse cellular processes. In the lungs, microRNAs play critical roles during development and homeostasis.6 As the functional importance of microRNAs in maintaining health becomes recognized, there is increasing interest to define their role during disease states. For example, multiple studies now show direct evidence for microRNA dysregulation in various respiratory disorders.7,8
There is mounting evidence supporting a critical role for microRNAs as putative drivers and biomarkers of respiratory disorders.8 Studies of the airway epithelium, in particular, have found that microRNA expression may be a sensitive marker for various disease states.9-11 These intriguing findings illustrate the possibility of using molecular changes in the airway epithelium as surrogates for detecting lung diseases and gaining insight into their pathogenesis. Therefore, using freshly purified airway epithelial cells from airway brushings during bronchoscopy, we investigated whether ACR is associated with altered microRNA signatures in the airway epithelium of lung transplant patients. We hypothesized that an unbiased approach would identify previously unknown relationships between microRNA expression profiles and ACR and provide insights into putative molecular mechanisms of acute rejection.
MATERIALS AND METHODS
Study Population
Lung transplant patients undergoing bronchoscopic evaluation were enrolled into this study after providing informed consent. Samples were retrospectively collected from patients that had bronchoalveolar lavage (BAL) fluid collections showing no evidence of acute infection during the bronchoscopy. All patients had transbronchial biopsies that were evaluated by a dedicated lung pathologist. Patients in the “control” group had no evidence of ACR on pathological examination. Patients with ACR had various grades of vascular (A grade) and airway (B grade) inflammation. This study was approved by the Institutional Review Board at the University of Washington.
Airway Epithelial Brushing and RNA Collection
During the bronchoscopy, airway epithelial cells were collected from subsegmental airways by 4 to 6 passes with a cytologic brush (ConMed, Utica, NY). Cells were collected in cold saline and immediately transferred to the laboratory on ice for processing. To ensure a pure population of airway epithelial cells, inflammatory cells (approximately 3% of the total population) were first depleted using a biotinylated antihuman CD45 antibody (AbD Serotec, Raleigh, NC) as previously published.12 The purified airway epithelial cells were placed into TRIzol (Life Technologies, Grand Island, NY). RNA was processed using phase-lock tubes as per manufacturer's instructions (5PRIME, Inc, Gaithersburg, MD).
MicroRNA Profiling and Data Analysis
Following the manufacturer's protocols, cDNA was created using miRCURY LNA cDNA synthesis kit (Exiqon, Woburn, MA) and was subsequently used for microRNA profiling with the miRCURY LNA Universal RT human microRNA PCR panel (V2.0; Exiqon). This PCR-based panel evaluated 742 unique human microRNAs. A global normalization strategy was used in which each microRNA quantification cycle (Cq) was subtracted from the mean Cq of all microRNA probes.12,13 The variability of microRNA expression between control (n = 11) and ACR (n = 10) groups was evaluated by principal component analysis (PCA).14 Differential microRNA expression between control and ACR subjects was determined based on a nonparametric approach known as significance analysis of microarrays.15 Adjustment for multiple hypothesis testing was performed using false discovery rate analysis with a cutoff less than 0.05, signifying statistical significance.
To limit false-positive associations, we did not use in silico algorithms to predict microRNA targets. Instead, putative gene targets of differentially expressed microRNAs were identified using miRTarBase (version 4.5, mirtarbase.mbc.nctu.edu.tw), a manually curated database of over 38 000 experimentally validated human microRNA-target interactions. Interactions were classified as those with strong experimental evidence (validation with reporter assays, Western analysis) or weaker evidence (eg, microarray, proteomics).16 Gene targets were selected only if they had strong empirical evidence for interacting with at least 1 differentially expressed microRNA. An interaction network summarizing the relationships between differentially expressed microRNAs and their targets was developed using Cytoscape.17 Functional enrichment analysis of target genes whose expression may be modulated by the differentially expressed microRNAs was performed using Database for Annotation, Visualization and Integrated Discovery (version 6.7, david.abcc.ncifcrf.gov) and was based on multiple annotation and pathway resources including gene ontology, Kyoto Encyclopedia of Genes and Genomes, and Biocarta.18 Enrichment P values were adjusted for multiple comparisons using the Benjamini-Hochberg method.
RESULTS
Patient Demographics
We enrolled a total of 21 lung transplant patients in this study. The control group was composed of 11 subjects with no evidence of infection or rejection based on BAL cultures and transbronchial biopsy results. Ten patients had histopathologic evidence of ACR on biopsy, but no active infection by culture of BAL fluid (Table 1).
TABLE 1.
Patient demographics
Acute Rejection Profoundly Alters Airway Epithelial MicroRNA Expression Profiles
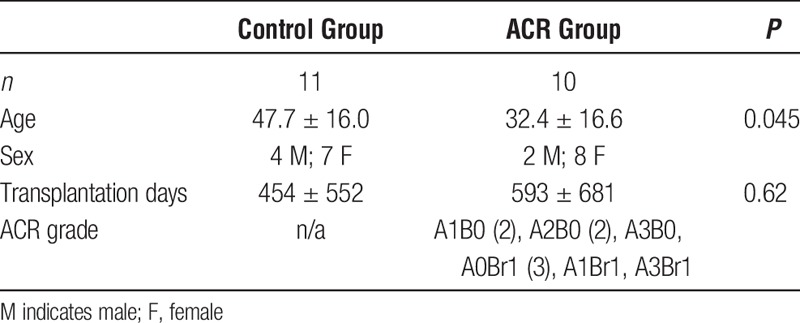
Initially, we applied PCA to assess whether variability across the entire microRNA dataset (n = 742 microRNAs) segregated patients with ACR from those without rejection (Figure 1A). We observed a reasonable distinction between the 2 groups, implying that acute rejection induces widespread alterations in microRNA expression and these patterns may be exploited to distinguish ACR patients from those without it. Next, we identified 117 differentially expressed microRNAs between ACR versus control lung transplant patients after adjustment for multiple comparisons using a false discovery rate less than 0.05 (Figure 1B; Table S1, SDC, http://links.lww.com/TXD/A16). Unexpectedly, all of these microRNAs were downregulated in the airway epithelium of patients suffering from acute rejection. Because microRNAs generally suppress transcription of their target genes, our results suggest that ACR activates a set of regulated airway epithelial genes through repression of specific microRNAs.
FIGURE 1.
Transcriptional patterns of airway epithelial microRNAs during ACR. A, Principal component analysis of the entire microRNA expression profiles (742 microRNAs) across 21 lung transplant recipients (n = 10 with ACR, n = 11 without rejection) yielded reasonable separation between the two patient groups. This implied the presence of significant global expression differences between the 2 groups. Each axis explains a proportion of the variability in microRNA expression, thereby determining the spatial location of the subjects. Note that for PCA, we used information across all 742 microRNAs, including many that were not differentially expressed between the 2 groups, thereby reducing power for segregating patients with and without ACR. B, Heatmap depiction of 117 differentially expressed microRNAs in ACR after adjustment for multiple testing using an FDR less than 0.05. Note that almost all microRNAs were downregulated in the airway epithelium of subjects with ACR, implying activation of their corresponding gene targets. FDR indicates false discovery rate.
Differentially Downregulated MicroRNAs in ACR Target a Limited set of Genes Mapping to Diverse Functional Pathways
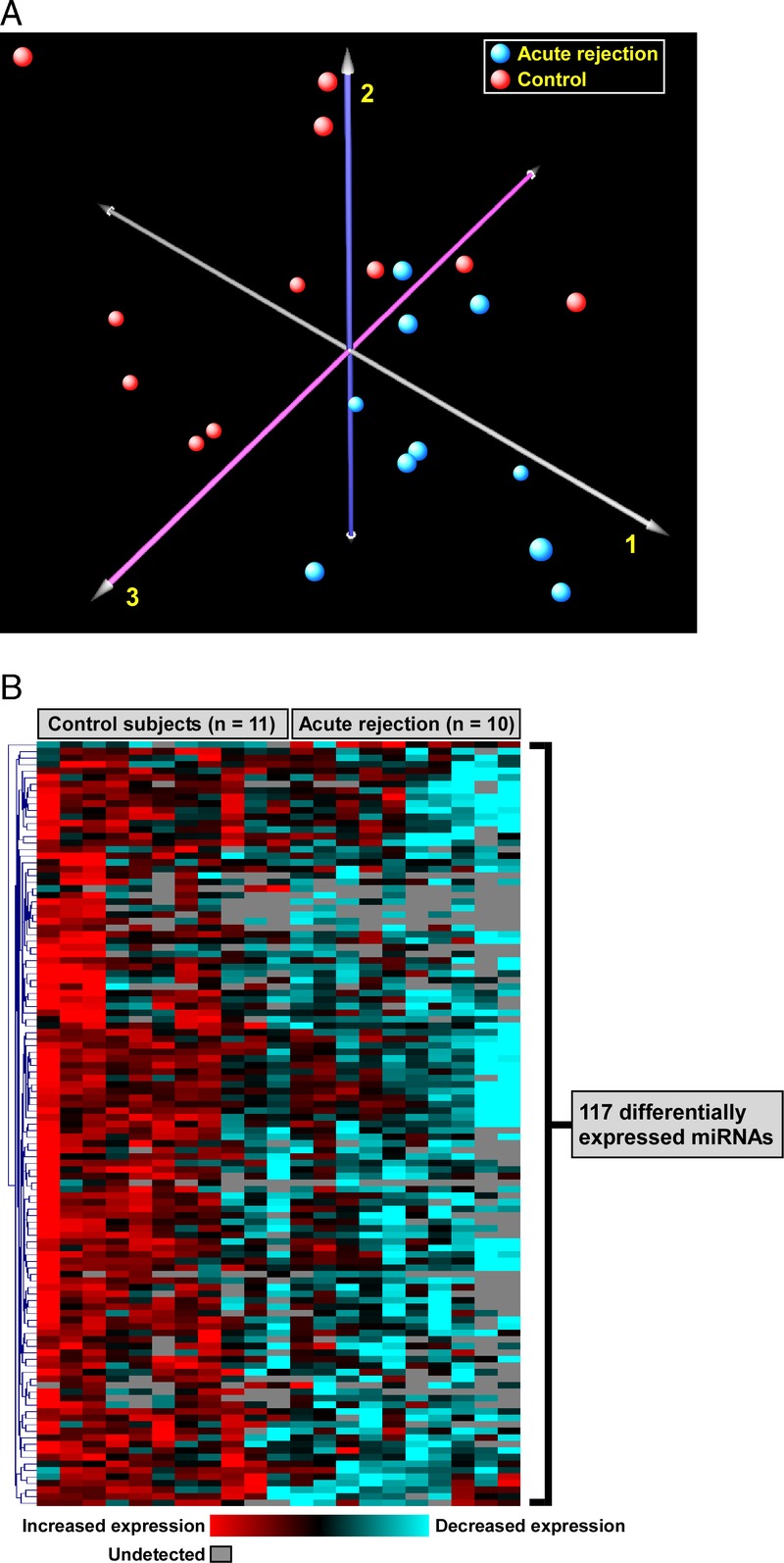
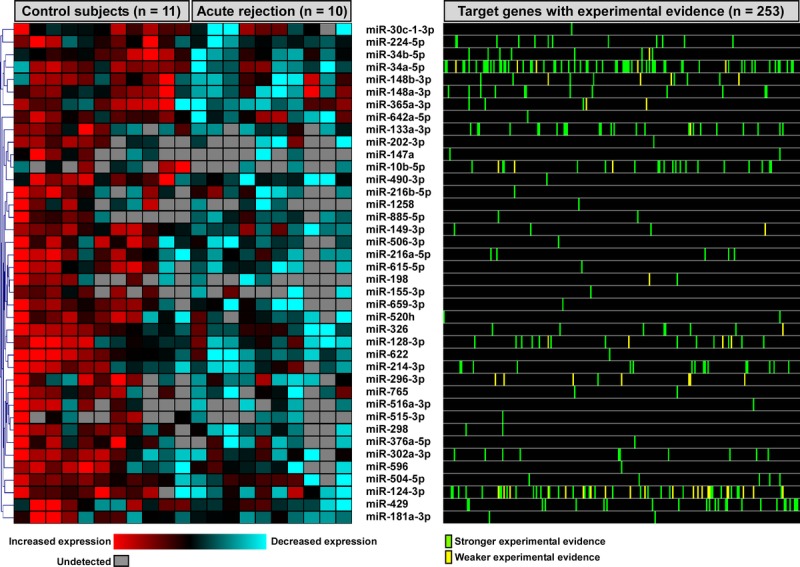
A number of computational methods have been developed to identify putative microRNA targets. However, these predictions are theoretical, and in many instances, may not translate into biologically verifiable effects. To enhance the biological relevance of our findings, we intersected the list of differentially expressed microRNAs in acute rejection with predicted gene targets that have been experimentally validated. Not surprisingly, of 117 downregulated microRNAs, only 40 had strong empirical evidence for regulating gene expression (Figure 2). This subset of 40 ACR-associated microRNAs influenced the expression of 253 genes based on previously published experimental results (Figure 2; Table S2, SDC, http://links.lww.com/TXD/A16). The interaction between differentially expressed microRNAs and their targets was explored using network analysis, revealing significant heterogeneity in the connectivity of microRNAs, with some interacting with very few genes, whereas others, such as miR-34a-5p and miR-124-3p, influencing a much wider repertoire of targets (Figure 3A). To gain an understanding of the biologic consequences of ACR-induced microRNA alterations in airway epithelial cells, we performed functional enrichment analysis of the 253 putative target genes. We identified a large number of overrepresented processes and pathways, many of which mapped to cellular proliferation, differentiation, migration as well as apoptosis, cancer, and developmental programs (Figure 3B; comprehensive list in Table S3, SDC, http://links.lww.com/TXD/A16).
FIGURE 2.
Linkage of differentially expressed microRNAs during ACR with their experimentally validated target genes. Of 117 ACR-associated microRNAs (Figure 1B), 40 microRNAs had strong empirical evidence for regulating a total of 253 gene products. The strength of experimental evidence was divided into strong (eg, reporter assay) or weak (eg, microarray-based); however, each candidate target gene was required to have at least 1 strong experimental association with a differentially expressed microRNA.
FIGURE 3.
Functional enrichment analysis of ACR-associated microRNA gene targets. A, Differentially expressed microRNAs in airway epithelium of patients with ACR were linked to 253 experimentally verified gene products, creating a complex interaction network (see also Figure 2). Note that some microRNAs, such as miR-34a-5p and miR-124-3p, modulated many targets, whereas others influenced very few genes. B, A representative summary of functional categories and pathways enriched among these 253 microRNA-regulated genes reveals that ACR is highlighted by activation of processes involved in proliferation, transformation, development, differentiation, repair, and migration.
DISCUSSION
Acute cellular rejection is a major complication of lung transplantation and contributes to reduced survival.2 This pilot study is the first report showing that acute rejection in lung transplant patients is associated with dysregulated microRNA expression in the airway epithelium. Our unbiased approach identified a panel of candidate microRNAs that distinguished patients with ACR from lung transplant recipients without rejection. Moreover, our findings suggest that repair and regeneration pathways regulated by these differentially expressed microRNAs are enhanced during acute rejection.
Finding more objective measurements of ACR may improve the diagnostic accuracy and interobserver differences between pathologists.19 Our study implies that microRNA profiling of the airway epithelium may be a useful tool to segregate lung transplant patients with and without ACR. Indeed, several studies have proven the utility of evaluating large airway epithelial expression of microRNAs to identify disease states of the smaller airways.9-11 This pilot study was not designed to differentiate the grade or severity of ACR, and larger studies are needed to further delineate specific molecular signatures within the airway epithelium that can more precisely separate various grades of rejection. However, our findings suggest that large airway epithelial cells may serve as informative sentinels for assessing disease states in lung transplant patients.
Dysregulation of microRNAs could reflect important changes within the lung epithelium in response to acute rejection. We noted that several differentially expressed microRNAs during ACR modulated many gene products, indicating that they yield widespread functional influence and may be potential therapeutic targets. For example, miR-124, one of the most densely connected network nodes (Figure 3A), regulates lung epithelial cell maturation20 and controls the proliferation and migration of pulmonary vascular fibroblasts.21 By focusing on validated targets of all differentially expressed microRNAs, we found that pathways associated with injury and repair, such as cellular proliferation, development, differentiation, and migration, were enriched during ACR (Figure 3B). Our results demonstrate that many microRNAs are specifically downregulated during ACR as part of lung epithelium's injury response and their suppression can lead to the activation of reparative and remodeling programs.
One such pathway enriched during ACR was the profibrotic TGF-β signaling cascade (adjusted P value 9.6 × 10−5). Because ACR is the largest risk factor for future development of obliterative bronchiolitis—a pathologic process characterized by peribronchiolar fibrosis22—our findings imply that although differential expression of some microRNAs may promote repair, they also shift the epithelial phenotype toward a fibroproliferative microenvironment with subsequent extracellular matrix deposition and remodeling. Indeed, microRNA-mediated dysregulation of the TGF-β pathway was recently reported in lung transplant patients with chronic obliterative bronchiolitis.23
To date, no studies have investigated microRNA profiles in the lung epithelium of transplanted patients. The bronchial epithelial cell transcriptome has been evaluated, and distinct signatures were identified that separate healthy controls from lung transplant patients providing further evidence that molecular changes in the epithelial compartment can be a used as informative biomarkers of disease state.24 Global transcriptional profiling of inflammatory cells within the airspaces have also been used to identify changes that segregated lung transplant patients with and without ACR.25,26 However, a limitation of this approach is that ACR is a T-cell–mediated process,22 and the observed molecular signatures may therefore largely reflect changes in cellular infiltration.26 Therefore, sampling the lung epithelium may be preferable in identifying unique signatures for differentiating various grades of ACR as well as distinguishing patients with infection from those with acute rejection.
Compared with transbronchial or surgical biopsies, airway brushing is minimally invasive and can circumvent complications, such as excessive bleeding, pneumothorax, and respiratory failure.26 In our pilot study, we evaluated microRNA signatures in a purified population of epithelial cells by including a rapid depletion step using an anti-CD45 antibody. However, cytologic brushing of the airways without purification yields an enriched epithelial cell population,24 and future studies should evaluate whether a leukocyte depletion step is necessary to provide an informative signatures for differentiating diseased states. Simplifying the process with less sample processing would be ideal in developing a diagnostic test that can be widely applicable across various institutions. Transbronchial biopsy is approximately 80% sensitive for diagnosing ACR,3,4 and the comparative sensitivity of airway epithelial brushings in identifying ACR needs to be determined using larger studies in the future.
Our study has several limitations. This was a pilot study with small sample size, and therefore our results must be validated independently using larger cohorts. However, the primary goal of this project was focused on determining whether ACR induces distinct changes in airway epithelium of lung transplant patients—a finding that is supported by our data. Another important limitation of our study was the heterogeneity of the patient population that likely contributes to significant biological variability. Furthermore, because we sampled large airway epithelial cells for microRNA profiling, where ACR was diagnosed based on small airway tissue, substantial biological discordance may be present between samples. Because the platform for measuring microRNAs was based on validated and highly sensitive quantitative PCR reactions, we did not independently confirm expression levels. Other platforms, such as those that are based on microarray technology, may have a larger coverage of microRNAs, but often require a preamplification step and may not be as sensitive and specific as qPCR. Because in silico prediction of microRNA targets may yield many false-positive associations, we limited the microRNA targets in our study to those genes with strong experimental validation, thereby increasing confidence in their putative biological roles.
In conclusion, this study demonstrates the feasibility of leveraging airway epithelial microRNA profiles to distinguish lung transplant patients with and without ACR while also identifying potential genes and pathways activated during rejection. Functionally, many of these microRNAs and their targets were enriched for wound repair and regeneration programs, which are consistent with the broader concept of airway epithelial injury during ACR. More comprehensive studies are needed to establish the sensitivity of this method in detecting ACR and delineating its various grades, differentiating ACR from infection, defining its utility to guide therapy, and identifying patients at risk for long-term complications, such as obliterative bronchiolitis. If independently validated, our findings offer proof-of-concept that microRNAs represent novel biomarkers for identifying ACR after lung transplantation and provide insight into potential molecular mechanisms leading to acute rejection.
Supplementary Material
ACKNOWLEDGMENTS
The authors would like to thank the subjects for agreeing to participate in this project, and Ellen McCown and Sharon Kelso for their help in enrolling patients.
Footnotes
Published online 20 November 2015.
Funding: This work was funded by the NHLBI/NIH grants HL084396 (P.C.), HL103868 (P.C.), and HL029594 (S.A.G.), the Institute of Translational Health Sciences (P.C.), and the Cystic Fibrosis Foundation (P.C.).
The authors declare no conflicts of interest.
P.C. and S.A.G. designed the study. P.C. and J.D.E. collected and processed the samples. P.C. and L.G. performed the experiments. S.A.G. analyzed the data. S.A.G. and P.C. wrote the article.
Supplemental digital content (SDC) is available for this article. Direct URL citations appear in the printed text, and links to the digital files are provided in the HTML text of this article on the journal's Web site (www.transplantationdirect.com).
REFERENCES
- 1. Martinu T, Chen D-F, Palmer SM. Acute rejection and humoral sensitization in lung transplant recipients. Proc Am Thorac Soc. 2009;6:54–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Christie JD, Edwards LB, Kucheryavaya AY, et al. The Registry of the International Society for Heart and Lung Transplantation: 29th Adult Lung and Heart-Lung Transplant Report-2012. J Heart Lung Transplant. 2012;31:1073–86. [DOI] [PubMed] [Google Scholar]
- 3. Trulock EP, Ettinger NA, Brunt EM, et al. The role of transbronchial lung biopsy in the treatment of lung transplant recipients. An analysis of 200 consecutive procedures. Chest. 1992;102:1049–54. [DOI] [PubMed] [Google Scholar]
- 4. Higenbottam T, Stewart S, Penketh A, et al. Transbronchial lung biopsy for the diagnosis of rejection in heart-lung transplant patients. Transplantation. 1988;46:532–9. [DOI] [PubMed] [Google Scholar]
- 5. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Nana-Sinkam SP, Hunter MG, Nuovo GJ, et al. Integrating the MicroRNome into the study of lung disease. Am J Respir Crit Care Med. 2009;179:4–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Rupani H, Sanchez-Elsner T, Howarth P. MicroRNAs and respiratory diseases. Eur Respir J. 2013;41:695–705. [DOI] [PubMed] [Google Scholar]
- 8. Booton R, Lindsay MA. Emerging role of MicroRNAs and long noncoding RNAs in respiratory disease. Chest. 2014;146:193–204. [DOI] [PubMed] [Google Scholar]
- 9. Perdomo C, Campbell JD, Gerrein J, et al. MicroRNA 4423 is a primate-specific regulator of airway epithelial cell differentiation and lung carcinogenesis. Proc Natl Acad Sci U S A. 2013;110:18946–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Schembri F, Sridhar S, Perdomo C, et al. MicroRNAs as modulators of smoking-induced gene expression changes in human airway epithelium. Proc Natl Acad Sci U S A. 2009;106:2319–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Solberg OD, Ostrin EJ, Love MI, et al. Airway epithelial miRNA expression is altered in asthma. Am J Respir Crit Care Med. 2012;186:965–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Chen P, Edelman JD, Gharib SA. Comparative evaluation of miRNA expression between in vitro and in vivo airway epithelium demonstrates widespread differences. Am J Pathol. 2013;183:1405–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Mestdagh P, Van Vlierberghe P, De Weer A, et al. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009;10:R64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Saeed AI, Sharov V, White J, et al. TM4: a free, open-source system for microarray data management and analysis. Biotechniques. 2003;34:374–8. [DOI] [PubMed] [Google Scholar]
- 15. Tusher VG, Tibshirani R, Chu G. Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A. 2001;98:5116–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hsu SD, Tseng YT, Shrestha S, et al. miRTarBase update 2014: an information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014;42:D78–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Dennis G, Jr, Sherman BT, Hosack DA, et al. DAVID: database for annotation, visualization, and integrated discovery. Genome Biol. 2003;4:P3. [PubMed] [Google Scholar]
- 19. Arcasoy SM, Berry G, Marboe CC, et al. Pathologic Interpretation of Transbronchial Biopsy for Acute Rejection of Lung Allograft Is Highly Variable. Am J Transplant. 2011;11:320–8. [DOI] [PubMed] [Google Scholar]
- 20. Wang Y, Huang C, Chintagari NR, et al. miR-124 regulates fetal pulmonary epithelial cell maturation. Am J Physiol Lung Cell Mol Physiol. 2015;309:L400–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wang D, Zhang H, Li M, et al. MicroRNA-124 controls the proliferative, migratory, and inflammatory phenotype of pulmonary vascular fibroblasts. Circ Res. 2014;114:67–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Martinu T, Howell DN, Palmer SM. Acute cellular rejection and humoral sensitization in lung transplant recipients. Semin Respir Crit Care Med. 2010;31:179–88. [DOI] [PubMed] [Google Scholar]
- 23. Xu Z, Ramachandran S, Gunasekaran M, et al. MicroRNA-144 dysregulates the transforming growth factor-β signaling cascade and contributes to the development of bronchiolitis obliterans syndrome after human lung transplantation. J Heart Lung Transplant. 2015;34:1154–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Skawran B, Dierich M, Steinemann D, et al. Bronchial epithelial cells as a new source for differential transcriptome analysis after lung transplantation. Eur J Cardiothorac Surg. 2009;36:715–21. [DOI] [PubMed] [Google Scholar]
- 25. Gimino VJ, Lande JD, Berryman TR, et al. Gene expression profiling of bronchoalveolar lavage cells in acute lung rejection. Am J Respir Crit Care Med. 2003;168:1237–42. [DOI] [PubMed] [Google Scholar]
- 26. Patil J, Lande JD, Li N, et al. Bronchoalveolar lavage cell gene expression in acute lung rejection: development of a diagnostic classifier. Transplantation. 2008;85:224–31. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.