

Abstract
In a number of human cancers, NTN1 upregulation inhibits apoptosis induced by its so‐called dependence receptors DCC and UNC5H, thus promoting tumor progression. In other cancers however, the selective inhibition of this dependence receptor death pathway relies on the silencing of pro‐apoptotic effector proteins. We show here that a substantial fraction of human breast tumors exhibits simultaneous DNA methylation‐dependent loss of expression of NTN1 and of DAPK1, a serine threonine kinase known to transduce the netrin‐1 dependence receptor pro‐apoptotic pathway. The inhibition of DNA methylation by drugs such as decitabine restores the expression of both NTN1 and DAPK1 in netrin‐1‐low cancer cells. Furthermore, a combination of decitabine with NTN1 silencing strategies or with an anti‐netrin‐1 neutralizing antibody potentiates tumor cell death and efficiently blocks tumor growth in different animal models. Thus, combining DNA methylation inhibitors with netrin‐1 neutralizing agents may be a valuable strategy for combating cancer.
Keywords: apoptosis, breast cancer, decitabine, dependence receptor, DNA methylation
Subject Categories: Cancer; Chromatin, Epigenetics, Genomics & Functional Genomics
Introduction
Recent research focusing on a specific functional family of receptors, namely the dependence receptors (DRs) (Mehlen et al, 1998; Llambi et al, 2001), has highlighted their implication in inhibiting tumor progression. Indeed, in contrast to most cellular receptors, a dual role characterizes these transmembrane receptors: in the presence of their respective ligands, they provide a typical positive signal (promoting cell survival, migration, proliferation, etc.), while the absence of ligand triggers a cascade of signaling events leading to apoptotic cell death. The DR protein family currently contains approximately twenty known members, and one of their most exhaustively studied ligands is Netrin‐1. This DR ligand was the first secreted attractive axon guidance cue to be described in the early nineties, since then, the role of netrin‐1 in many biological functions has been established (Cirulli & Yebra, 2007; Mehlen & Guenebeaud, 2010; Ramkhelawon et al, 2014). Netrin‐1 receptors, namely, are the Deleted in Colorectal Carcinoma (DCC), the Uncoordinated‐5‐Homologs (UNC5H1‐4/A–D). The ability of these receptors to trigger apoptosis in settings under ligand‐limited conditions was shown to be a constrain for impede tumor progression. Indeed, the inactivation of UNC5C, and DCC, or the mutation of the DCC‐inducing apoptosis domain is associated with tumor development and progression in mouse models (Bernet et al, 2007; Castets et al, 2012; Krimpenfort et al, 2012). Furthermore, in line with these observations, DCC and UNC5H are silenced in many cancers, either by loss of heterozygosity or epigenetic mechanisms (Hedrick et al, 1994; Bernet et al, 2007; Shin et al, 2007). Alternatively, in some types of cancers, an upregulation of NTN1 provides a similar tumor growth selective advantage by abolishing their dependency on netrin‐1 availability in the micro‐environment (Fitamant et al, 2008). This upregulation is particularly marked in some cases of aggressive breast cancer and in breast metastasis (Fitamant et al, 2008; Harter et al, 2014). The reliance on netrin‐1 for tumor growth was rapidly perceived as an opportunity for therapeutic intervention, since it was speculated that the disruption of the binding of netrin‐1 to its receptors should induce apoptotic cell death in vitro and tumor growth inhibition in vivo. Consistently, the silencing of netrin‐1 or the development of biological agents interfering with netrin‐1/DRs interactions has been shown to efficiently reduce tumor growth and metastasis in different animal models (Fitamant et al, 2008; Delloye‐Bourgeois et al, 2009a,b; Paradisi et al, 2013; Grandin et al, 2016). Moreover, an anti‐netrin‐1 antibody is under preclinical evaluation and should be assessed in early clinical trials in 2016.
Nevertheless, a substantial fraction of human tumors appears to conserve the expression of netrin‐1 receptors without upregulating NTN1 expression, suggesting that the downstream DR pathways may be impaired (Shin et al, 2007; Mian et al, 2011; Krimpenfort et al, 2012). In cancers, epigenetic modifications are frequently associated with an increase in the expression of anti‐apoptotic proteins and with the inactivation of factors inducing apoptosis (Baylin & Ohm, 2006). In this context, previous studies have reported that DAPK1, a serine threonine kinase responsible for UNC5H‐induced apoptosis, is downregulated in various cancers (Llambi et al, 2005; Guenebeaud et al, 2010). Furthermore, mechanistic analyses have demonstrated a direct relationship between the hypermethylation of the CpG island (CGi) located within the DAPK1 promoter region and its downregulation (Raval et al, 2007; Pulling et al, 2009; Mian et al, 2011; Kilinc et al, 2012). Finally, it was shown that treatment with decitabine (5‐aza‐2′‐deoxycytidine, DAC) inhibits the DNA methylation of the DAPK1 promoter and restores DAPK1 expression in lung cancer cell lines (Tang et al, 2004).
Inhibition of DNA methylation may prove to be a promising approach for combating cancer (Yang et al, 2010; Rodriguez‐Paredes & Esteller, 2011a; Dawson & Kouzarides, 2012; Tsai et al, 2012). Concordantly, DAC showed anti‐leukemic effects in several clinical trials and has been approved by FDA for the treatment of myelodysplastic syndromes and recent studies indicated that DAC shows some effects in preclinical models (Tsai et al, 2012). Here, we report that treatment with DAC restores the pro‐apoptotic machinery linked to the netrin‐1/DR‐mediated cell death signaling pathway in breast cancer cell lines and patient‐derived xenografts and sensitize cancer cells to netrin‐1 neutralizing agents. We thus provide evidence that combining DNA methylation inhibitors with a netrin‐1 neutralizing antibody empowers tumor cell death in vitro and tumor growth inhibition in mice.
Results
Epigenetic downregulation of netrin‐1 is associated with the epigenetic downregulation of DAPK1, in human breast cancers
In order to study the mechanisms underlying inhibition of netrin‐1/DRs‐mediated apoptosis, which in a fraction of tumors is neither dependent on netrin‐1 upregulation nor DR silencing, we investigated DAPK1 expression and DNA methylation in breast tumors.
We thus examined differentially methylated regions (DMRs) associated with malignant transformation in breast cancer samples (n = 92) available from The Cancer Genome Atlas (TCGA) by comparing paired normal/tumoral data obtained from the HumanMethylation450K Array (HM450, Illumina) and by high‐throughput sequencing of polyadenylated RNA (RNAseq). This comparison focused on the level of DNA methylation within the 5′‐end of NTN1 (position −765 to + 1,300 relative to the transcription start site, TSS) and DAPK1 (+ 365 and + 838 relative to the TSS) and revealed that these regions were hypermethylated (threshold = 2) in about 30% of tumoral samples compared with their normal counterparts (Fig 1A and B). Furthermore, NTN1 was downregulated (Fig 1C, fold change (FC) ≤ 0.5) in 43% of cases and the “NTN1‐low” samples were hypermethylated (Fig 1G; P = 3 × 10−2, two‐sided Mann–Whitney test) when compared with samples exhibiting no NTN1 downregulation (FC ≥ 1.3). Using the same approach for DAPK1, we observed that samples exhibiting DAPK1 downregulation (29% of the samples, Fig 1D) were also hypermethylated (P = 3 × 10−4) when compared with the other samples (Fig 1H). These epigenetic modifications did not seem to be independent events since in NTN1‐hypermethylated samples (n = 23, FC ≥ 2), DAPK1 was also hypermethylated (mean FC = 2.22), while in NTN1‐hypomethylated samples (n = 13, FC < 0.7), DAPK1 was not hypermethylated (mean FC = 0.93). The relationship between DAPK1/NTN1 downregulation and DNA hypermethylation was also observed in a larger number of breast cancer samples (n = 807) available on TCGA data portal. Indeed, the mean percentage of CpG methylation (Fig 1E and F) of DAPK1 and NTN1 were inversely correlated with their levels of expression (Pearson's r = −0.32, P < 10−18 and Pearson's r = −0.14, P = 6.7 × 10−5, respectively), suggesting that DNA methylation represses DAPK1 and NTN1 transcription in human breast tumors.
Figure 1. NTN1 and DAPK1 are hypermethylated and downregulated in human breast cancers.
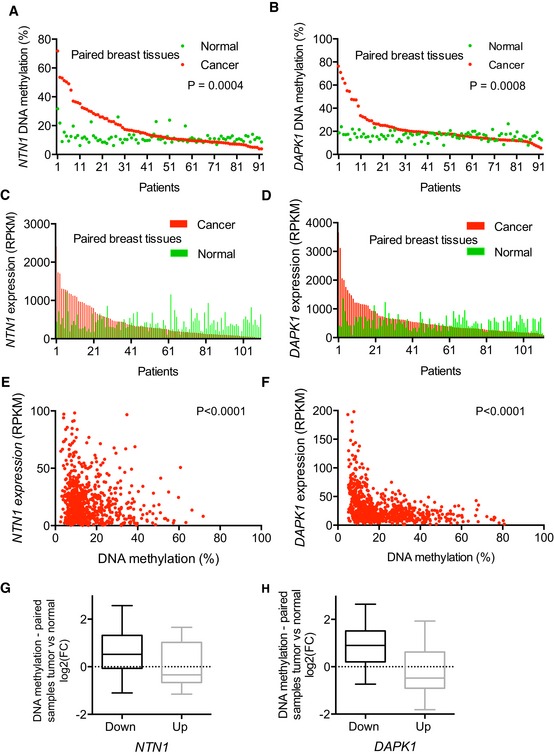
-
A, BDNA methylation level of NTN1 (A) and DAPK1 (B) 5′ regions (Illumina's HumanMethylation450K Array (HM450) from The Cancer Genome Atlas breast cohort) in paired breast tissues (normal: green circles, tumor: red circles), n = 92, Wilcoxon matched‐pairs signed rank test, P = 0.004 and P = 0.0008.
-
C, DNTN1 (D) and DAPK1 (G) gene expression in paired breast tissues (normal: green bars, tumor: red bars), RNAseq from TCGA breast cohort, n = 112 and 114, respectively.
-
E, FCorrelation between NTN1 (E) and DAPK1 (F) gene expression and DNA methylation in the breast cancer cohort (TCGA, n = 807). Pearson correlation, P = 6.7.10−5, r = −0.14 for NTN1 (A) and P < 10−16, r = −0.32 for DAPK1 (B), respectively.
-
G, HTumor/normal DNA methylation ratio of NTN1 (G) and DAPK1 (H) in human breast tumors (data extracted from TCGA cohort, paired samples) according to gene expression (downregulated FC ≤ 0.5, down, n = 33, or upregulated FC ≥ 1.3, up, n = 16). P = 3 × 10−2 and P = 3 × 10−4 two‐sided Mann–Whitney test, for NTN1 and DAPK1, respectively.
Although we cannot excluded a bias due to stromal contamination, it should be noted that, in paired samples, while hypermethylation was observed in tumoral tissues, very low levels of DNA methylation were measured in their normal counterparts (Fig 1A and B). For both genes, a region (not represented on HM450) located at the 3′‐end of the CGis (Fig 2A, light gray boxes) was selected for the quantitative analysis of DNA methylation of breast cancer biopsies (tumor bank—Centre Léon Bérard) by bisulfite pyrosequencing. The analysis indicated that the mean percentage of CpG methylation of the DAPK1 and NTN1 pyrosequenced regions were inversely correlated (Pearson's r = −0.66, P = 0.003, and Pearson's r = −0.55, P = 0.008, respectively) with their levels of expression (Fig EV1A and B). Altogether, these data suggested that DNA methylation is involved in the downregulation of NTN1 and DAPK1 in human breast cancers.
Figure 2. DNA methylation and demethylation in mammary cell lines.
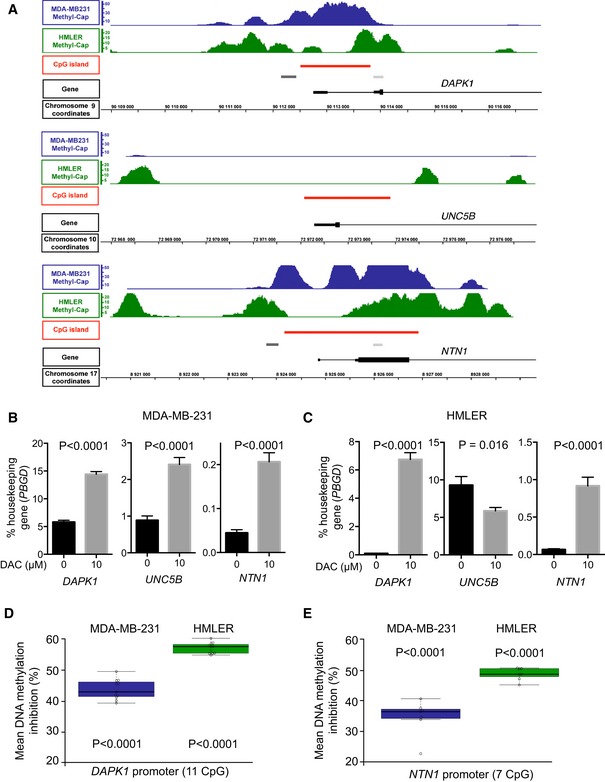
-
AMethyl‐Cap‐seq read density profiles of the 5′ end of DAPK1, UNC5B, and NTN1 in MDA‐MB‐231 (blue) and HMLER (green) cells. Red boxes represent the CpG islands (CGis); light gray boxes the regions analyzed by bisulfite PCR sequencing; dark gray boxes represent the regions analyzed by parallel sequencing of amplicons from bisulfite modified DNA; and black boxes represent the exons and UTR. Chromosome coordinates of each gene are given (black lines).
-
B, CGene expression was measured by qRT–PCR after 72 h for MDA‐MB‐231 (B) and HMLER cells (C) treated daily with DAC (10 μM). PBGD expression level was used as an internal control. Data are expressed as mean ± s.e.m. of at least 3 independent experiments. ****P < 0.0001, two‐tailed unpaired Student's t‐test.
-
D, EMeasurement of DNA hypomethylation of the DAPK1 (D) and NTN1 (E) promoters after decitabine treatment of MDA‐MB‐231 and HMLER cells. Over 1960 sequences were analyzed per group in 2 independent experiments. ****P < 0.0001, two‐way ANOVA and post hoc Tukey test.
Figure EV1. NTN1 and DAPK1 are concomitantly altered in human breast cancers.
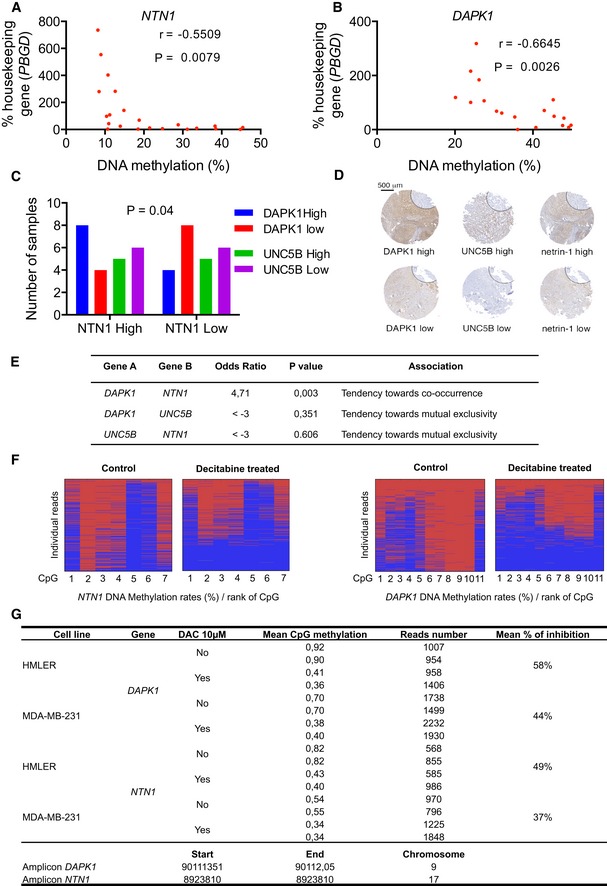
- Bisulfite PCR sequencing (4 CpGs analyzed, region: light gray boxes in Fig 1A) indicated that DNA methylation of the NTN1 CpG island (CGi) was inversely correlated with its expression. Pearson correlation, P = 0.008, r = −0.55, n = 18.
- Bisulfite PCR sequencing (3 CpGs analyzed, region: light gray boxes in Fig 2A) indicated that DNA methylation of the DAPK1 CGi was inversely correlated with its expression. Pearson correlation, P = 0.003, r = −0.66, n = 22.
- Tissue microarrays (70 paraffin embedded sections) from human breast carcinomas were immuno‐stained with antibodies against DAPK1, UNC5B, and netrin‐1. Samples were classified in quartiles according to the level of netrin‐1 expression. The levels of DAPK1 and UNC5B expression (index constructed from the percentage of sections exhibiting a positive staining) in the first and fourth quartiles of netrin‐1 expressing groups were compared using a chi‐squared test.
- Representative staining corresponding to low and high levels of expression is shown for each antibody, and expression levels were determined from the percentage of sections exhibiting a positive staining. As a control, staining of the samples using a non‐related isotype antibody was performed.
- Quantification of the presence or absence of alterations in gene A was associated with the presence or absence of alterations in gene B in human breast tumors which was determined from cBioPortal web site, z‐score threshold ± 2.2
- Individual methylation plots. Red rectangles represent methylated CpG and blue rectangles unmethylated CpG.
- Mean DNA methylation inhibition of DAPK1 and NTN1 in MDA‐MB‐231 and HMLER cells upon DAC treatment.
To determine whether this concomitant change in DAPK1 and NTN1 expression was also observed at the protein levels, DAPK1, UNC5B, and netrin‐1 were measured by immunohistochemistry (IHC) using tissue microarrays (70 sections) from human breast ductal carcinoma (Super Bio Chips). This analysis revealed, that, netrin‐1‐low samples also exhibited low levels of DAPK1 (χ2 test, P = 0.04). In contrast, UNC5B levels were the same, irrespective of netrin‐1 levels (Fig EV1C and D). This result was validated in the TCGA cohort of breast tumors samples (n = 807), where a correlation between DAPK1 and NTN1 expression was observed (odd ratio = 4.71, P = 0.03, Fig EV1E).
In order to establish an in vitro experimental model mimicking the DNA methylation alterations observed in breast cancer tissues, DNA methylation patterns of two cancer cell lines were determined by parallel sequencing. Pull‐down assays were conducted using the MDA‐MB‐231 cell line derived from human breast cancer, and the HMLER cell line generated through the in vitro transformation of human mammary cells (Elenbaas et al, 2001; Morel et al, 2008). Methylated DNA fragments were selected using a recombinant protein containing the Methyl‐CpG‐binding domain of MBD2 of MBD2 (Methyl‐Cap‐seq), from MDA‐MB‐231 cell line derived from human breast cancer, and the HMLER cell line constructed from in vitro transformation of human mammary cells (Elenbaas et al, 2001; Morel et al, 2008). Data obtained indicated that the 5′‐end CpGis of DAPK1 and NTN1 were methylated in the two cancer cell lines (Fig 2A). Furthermore, the treatment of MDA‐MB‐231 and HMLER cells with decitabine (DAC) resulted in a significant upregulation of DAPK1 and NTN1 mRNA (Fig 2B and C) and in the inhibition of DNA methylation within the DAPK1 and NTN1 promoter regions (Figs 2D and E, and EV1F and G).
Parallel sequencing of DMRs (Fig 2A, dark gray boxes) validated their methylation status obtained from Methyl‐Cap‐seq experiments and indicated that DAC treatments reduced their level of methylation by half, in vitro (Fig 2D and E). Of note, DAC treatment of MDA‐MB‐231 cells resulted in the upregulation of UNC5B, although its promoter was not methylated in any of the cell lines tested (Fig 2B and C), suggesting an indirect regulatory mechanism. However, this upregulation was not observed for the other netrin‐1‐specific receptors, UNC5A, UNC5C, and DCC (Fig EV2A and B). Altogether, these data strongly suggest that DNA hypermethylation is involved in the transcriptional silencing of DAPK1 and NTN1 in human breast cancer cells.
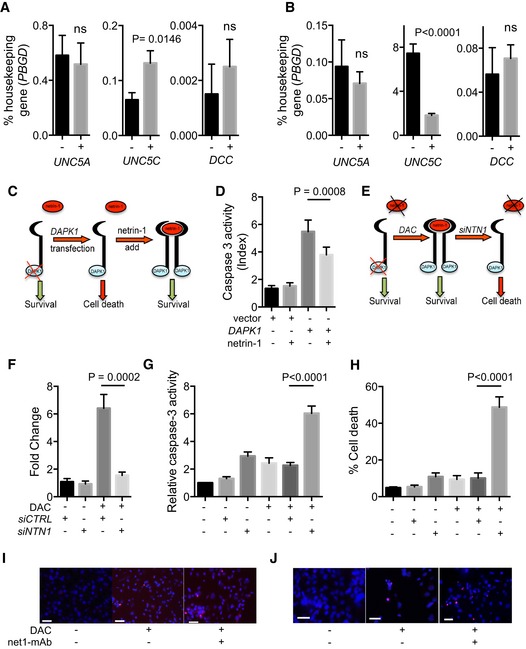
Figure EV2. The NTN1 DR apoptotic pathway, mechanism, and specificity.
-
A, BOther NTN1 DR gene expression in breast cell lines, and impact of DAPK1 and NTN1 on the induction of apoptotic cell death in vitro. Gene expression was measured by qRT–PCR after 72 h in MDA‐MB‐231 (A) and HMLER (B) cells treated daily with 10 μM decitabine (DAC). The level of PBGD expression was used as an internal control. Data are represented as mean ± s.e.m. for to 3 independent experiments. ****P < 0.0001, two‐tailed unpaired Student's t‐test.
-
CSchematic representation of the cellular effect on HMLER cells of DAPK1 overexpression and/or recombinant netrin‐1 treatment.
-
DCaspase‐3 activation upon DAPK1 overexpression in HMLER and its reversion by the addition of netrin‐1. ***P < 0.0001, one‐way ANOVA.
-
ESchematic representation of DAC and effect of NTN1 siRNA on HMLER cells.
-
FEffect of NTN1 siRNA (siNTN1) on the expression of NTN1 by qRT–PCR. The level of PBGD expression was used as an internal control. ***P < 0.0001, one‐way ANOVA.
-
G, HsiNTN1 triggers apoptosis and cell death in hypomethylated HMLER cells. Cells were treated with DAC (10 μM, 72 h), and/or siNTN1 (30 pmol, 48 h), and caspase‐3 activity (G) and cellular mortality (H) were measured. As a control, a scramble siRNA was used. Data (G, H) are expressed as mean ± s.e.m. of at least 3 independent experiments. ****P < 0.0001, one‐way ANOVA.
-
I, JRepresentative images of TUNEL experiments shown in Fig 3, in MDA‐MB‐231 and HMLER cells, respectively. Scale bars = 50 μm.
Decitabine resensitizes cancer cells to netrin‐1 interference in vitro
We thus hypothesized that the pharmacological targeting of the DNA methylation machinery may restore functional netrin‐1/DR pathways and resensitize netrin‐1‐low cells to netrin‐1 interference. We investigated whether the forced expression of DAPK1 in the DAPK1‐negative HMLER cells re‐established pro‐apoptotic pathways (Fig EV2C and D). As expected, an increase in caspase‐3 activity was observed in DAPK1 transfected cells. Furthermore, this pro‐apoptotic effect was partially reversed by adding recombinant netrin‐1 (Fig EV2D). We next analyzed cell death by conducting viability assays and caspase‐3 activity assays in HMLER cells transfected with NTN1 siRNA and treated with DAC. While transfections had a mild effect on HMLER cells per se, treatment with DAC strongly potentiated the netrin‐1 deprivation‐induced cell death (Fig EV2F–H).
Keeping in mind the therapeutic perspective of our study, we assessed the effect of combining DAC with a human anti‐netrin‐1 antibody, net1‐mAb (Grandin et al, 2016). MDA‐MB‐231 and HMLER cells were treated with DAC or net1‐mAb, or a combination of both drugs. As anticipated, the cell lines were resistant to net1‐mAb alone, while, as previously observed (Lund et al, 2011) (Rodriguez‐Paredes & Esteller, 2011b), DAC induced cell death in most of the cells tested, as evidenced by DNA fragmentation (Figs 3A and B, and EV2I and J) and viability assays (Fig 3C). The addition of net1‐mAb to DAC‐treated cells significantly enhanced apoptosis (Fig 3A–C). Moreover, this effect was blocked by the concomitant addition of recombinant netrin‐1 (Fig 3A–C), indicating that net1‐mAb‐induced cell death was specifically linked to netrin‐1 neutralization. To confirm that the pro‐apoptotic activity observed upon the combined net1‐mAb and DAC treatment was not linked to an intrinsic property of DAC, MDA‐MB‐231 and HMLER cell lines were treated with the 5‐azacytidine (Aza), a DNA methylation inhibitor. This treatment resulted in similar gene expression modifications and cell death effects as DAC, when combined with the net1‐mAb (Appendix Fig S1). The functional consequences of this combination treatment were also evaluated in additional human breast cancer cell lines. The highest upregulation of both NTN1 and DAPK1 was observed when treating cells with 10 μM of DAC (Fig EV3A–E), we thus used this concentration in the combined DAC and net1‐mAb treatment to investigate the induction of apoptosis (Fig EV3F). A similar potentiation of cell death by combining DAC and net1‐mAb was observed in cell lines (AU565, SKBR3, and MDA‐MB‐157) exhibiting an upregulation of at least one of 3 genes, NTN1, UNC5B, or DAPK1 (Fig EV3C–E).
Figure 3. Netrin‐1 neutralizing antibody net1‐mAb triggers apoptosis in hypomethylated MDA‐MB‐231 and HMLER breast cancer cell lines.
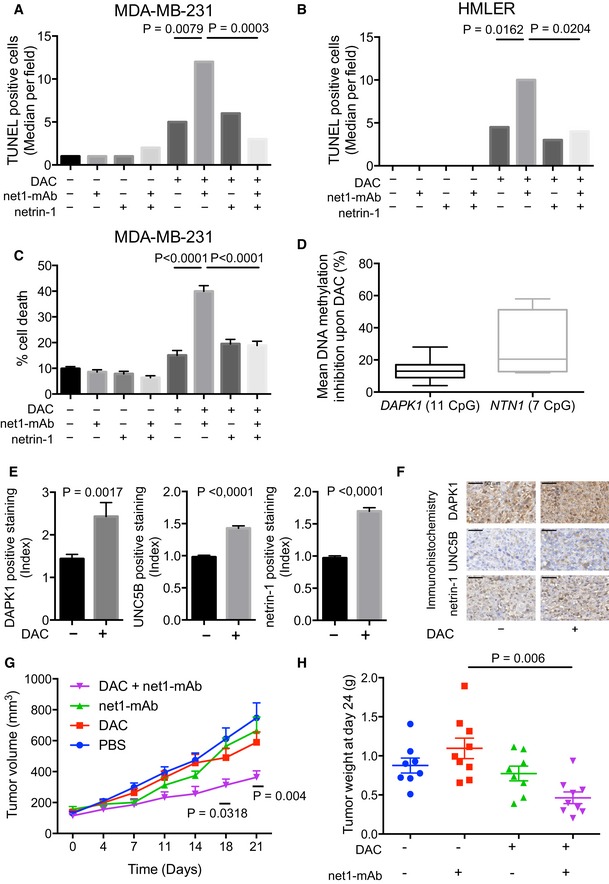
-
A, BCells were treated with DAC (10 μM, 72 h), and/or net1‐mAb (10 μg/ml, 48 h) and/or recombinant netrin‐1 (5 μg/ml, 48 h). TUNEL assays from 3 independent experiments. ***P < 0.0001 one‐way ANOVA.
-
CCellular mortality was assessed using the toxilight assay. Data represent mean ± s.e.m. from 3 independent experiments. ****P < 0.0001, one‐way ANOVA.
-
D–HMDA‐MB‐231 cells were implanted into the mammary gland fat pad of immuno‐compromised mice. When tumors reached 100 mm3, mice were injected subcutaneously with decitabine (0.4 mg/kg) or PBS and/or intraperitoneally with net1‐mAb (10 mg/kg). (D) Loss of DNA methylation of the NTN1 and DAPK1 promoters in decitabine treated tumors, compared with PBS treated tumors, after one week of treatment. > 1,700 sequences were analyzed per group in 2 independent experiments. ****P < 0.0001, two‐way ANOVA and post hoc Tukey‐test. (E, F) After one week of treatment, tumors from xenografted mice were fixed in formalin, embedded in paraffin, and sliced into 4 μm sections. (E) Levels of DAPK1, UNC5B, and netrin‐1 were measured in 4 independent tumors per treatment group by immunohistochemistry staining, and expressed as a percentage of total tumor surface normalized against the control PBS‐group. Data represented as means ± s.e.m. ****P < 0.0001, two‐way ANOVA and post hoc Tukey‐test. (F) Representative tumor sections corresponding to MDA‐MB‐231 xenografts, scale bars = 50 μm. (G) Tumor volumes were measured twice a week. The statistical significance of the differences obtained between the control PBS‐group and treated (DAC + net1‐mAb) group was determined by two‐way ANOVA and post hoc Tukey‐test. ***P < 0.0001. Error bars = s.e.m. n = 9 mice per group. (H) Tumor weights were measured 3 days after the end of the experiment. The statistical significance of the differences obtained between the groups was determined by one‐way ANOVA, ***P < 0.0001. Error bars = s.e.m. n = 9 mice per group.
Figure EV3. Netrin‐1 neutralizing antibody net1‐mAb triggers apoptosis in DAC‐treated breast cancer cell lines.
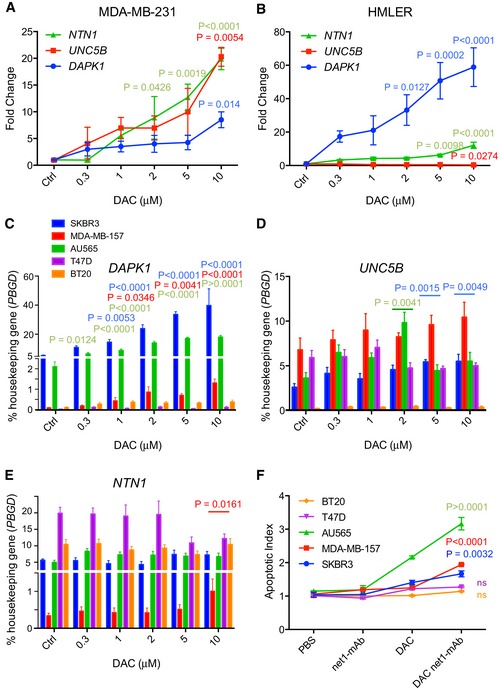
-
A–EGene expression was measured by qRT–PCR after 72 h in human breast cancer cell lines: MDA‐MB‐231 (A), HMLER (B) and SKBR3, MDA‐MB‐157, AU565, T47D, and BT20 (C–E) treated daily with DAC at final concentrations of 0.3, 1, 2, 5, and 10 μM. The level of PBGD expression was used as an internal control. Data are expressed as mean ± s.e.m. of at least 3 independent experiments. ****P < 0.0001, one‐way ANOVA. Colors of the stars correspond to the gene analyzed (A and B) or to the cell lines analyzed (C–F).
-
FComparison of cellular mortality in DAC‐treated cells versus DAC + net1‐mAb treated cells assessed using the caspase cleavage assay. Data are expressed as mean ± s.e.m. of at least 3 independent. ****P < 0.0001, two‐way ANOVA and post hoc Tukey test. ns = not significant.
The combination of the netrin‐1 neutralizing antibody with decitabine inhibits tumor growth and metastasis in animal models
We next investigated whether the inhibition of DNA methylation could also re‐establish the netrin‐1/DRs‐mediated pro‐apoptotic pathways in vivo. Nude mice were engrafted with MDA‐MB‐231 cells in the mammary fat pad as an orthotopic site for breast cancer. When tumors were palpable (≈100 mm3), DAC was subcutaneously injected at a therapeutic dose (0.5 mg/kg). Parallel sequencing of amplicons derived from DAPK1 and NTN1 CGis indicated that these genes were also hypomethylated (ranging from 5 to 27% depending of the CpG analyzed) in tumors from DAC‐treated mice (Fig 3D). Furthermore, we observed that DAC treatments were associated with the re‐expression of DAPK1, netrin‐1, and UNC5B by the IHC staining of tumor sections (Fig 3E and F).
To evaluate the effect on tumor growth, MDA‐MB‐231 orthotopic xenografts were performed, and mice were treated by intraperitoneal injections of net1‐mAb, or subcutaneous injections of DAC, or a combination of both treatments for a period of 21 days (Fig 3G and H). While no significant difference was observed between the control groups and mice treated either with net1‐mAb or DAC alone, the combination of both drugs strongly inhibited tumor growth (Fig 3G) and led to a reduction in tumor weight (Fig 3H). Furthermore, similar results were obtained using the NET1‐mAb antibody, which is scheduled for human trials in 2016 (Grandin et al, 2016) (Appendix Fig S2A). Interestingly, the inhibitory effect of the combined treatment on tumor growth was associated with a marked increase in tumor cell death (Appendix Fig S2B and C).
To gain further insights into the mechanisms underlying the tumor suppressive activity of netrin‐1 neutralization following DAC treatment, cancer cell proliferation and tumor angiogenesis were analyzed in treated tumors. A small reduction in the proliferative rate (Ki67 staining) was determined in MDA‐MB‐231 xenografts from mice treated with DAC; however, the differences between treated samples and controls were not statistically significant (Appendix Fig S3A and B). Staining of sections from MDA‐MB‐231 xenografts with an antibody directed against CD31 (also known as platelet endothelial cell adhesion molecule) revealed that the anti‐netrin‐1 antibody treatment led to a reduction in angiogenesis. This finding strengthens a previous report, which suggested the implication of netrin‐1 during angiogenesis (Bongo & Peng, 2014). However, no statistically significant differences were observed between mice treated with the anti‐netrin‐1 antibody alone, where no inhibition in tumor growth was detected, and the mice treated with a combination of DAC and net1‐mAb, where tumor growth was profoundly affected (Appendix Fig S3A). Hence, the inhibitory effect of the combination of DAC and net1‐mAb on tumor growth is likely related to their pro‐apoptotic effects, rather than to a change in proliferation or in angiogenesis.
Finally, the effect of this combination treatment on metastasis formation was evaluated in a model previously used for monitoring tumor dissemination (Walker et al, 2004; Delloye‐Bourgeois et al, 2009b). MDA‐MB‐231 cells previously treated with control IgG1, DAC, and net1‐mAb alone or in combination, were seeded onto the chorioallantoic membrane (CAM) of 10 days old chicken embryos. Data obtained indicated that CAMs grafted with cells previously treated with DAC and the net1‐mAb exhibited a reduction in the formation of lung metastasis (Appendix Fig S4).
To move toward a model closer to the human pathology, we analyzed the effect of combining DAC and the netrin‐1 antibody in mice model bearing a patient‐derived tumor (PDX model). Consistently with the data obtained from mice engrafted with cell lines, DAC treatments induced the hypomethylation of DAPK1 and NTN1 in our PDX (Fig 4A) and increased the levels of DAPK1, UNC5B, and netrin‐1 (Fig 4B and C). Furthermore, IHC staining on tumor sections indicated that the combination treatment increased the apoptotic rate (Fig 4D and E), while it was not correlated with tumor cell proliferation or angiogenesis (Appendix Fig S3C and D). Similarly to the data obtained in MDA‐MB‐231 engrafted cells, the combined treatment reduced the growth of the PDX tumor when compared to the effects of monotherapies (Fig 4F).
Figure 4. DAC treatment triggers upregulation of genes in the netrin‐1/dependence receptor signaling pathway, and, when combined with net1‐mAb, induces apoptosis and tumor growth inhibition in a mouse model bearing a patient‐derived breast tumor.
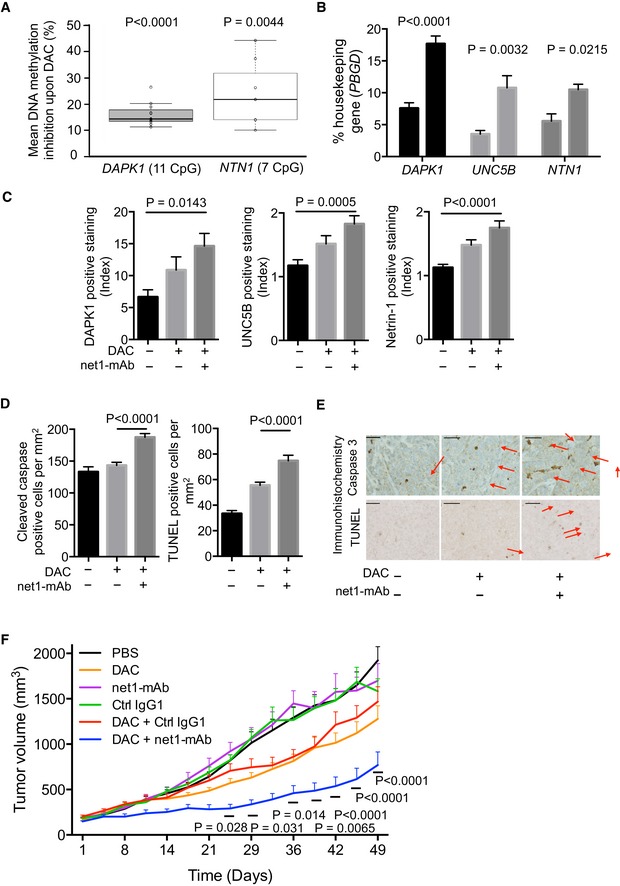
- Loss of DNA methylation of the NTN1 and DAPK1 promoters in decitabine‐treated PDX tumors, compared with the control PBS group. The percentage of mean DNA methylation of the 11 DAPK1‐CpGs was 94% (550 amplicons analyzed), while that of the 7 NTN1‐CpGs was 64% (213 amplicons analyzed). Two‐way ANOVA and post hoc Tukey test.
- Expression of DAPK1, UNC5B, and NTN1 was measured by qRT–PCR in PDX tumors after 7 days of in vivo DAC treatment (0.4 mg/kg). The level of PBGD expression was used as an internal control. Data are expressed as mean ± s.e.m. for at least 3 grafts per group. ****P < 0.0001, two‐way ANOVA and post hoc Tukey test.
- Levels of DAPK1, UNC5B, and netrin‐1 were measured in at least 3 independent paraffin embedded xenografts per treatment group by immunohistochemistry. Data are expressed as the percentage of the total tumor surface normalized against the median tumor surface of the PBS group. ****P < 0.0001, one‐way ANOVA. Error bars = s.e.m.
- Cleaved caspase‐3 and DNA fragmentation (TUNEL) median number of cells per mm2 were measured in treated xenografts. Data represent mean ± s.e.m. from at least 3 tumors per group. ****P < 0.0001, one‐way ANOVA.
- Representative sections of the xenograft PDX analyzed in (D). Scale bars = 50 μm. Arrows indicate positive staining.
- The combination of net1‐mAb and DAC reduces human breast tumor growth in immuno‐compromised mice. After anesthesia, mice were engrafted in the interscapular area with a ≈ 60 mm3 patient‐derived tumor. When the tumors reached 120–150 mm3, mice were injected subcutaneously with decitabine (0.4 mg/kg) or PBS and/or intraperitoneally with net1‐mAb (10 mg/kg) or with a human IgG1 control isotype antibody (Ctrl IgG1, 10 mg/kg) from day 1 to day 21. Tumor volumes were measured twice a week; n = 7 mice per group. The statistical significance of the differences obtained between the DAC + Ctrl IgG1 group and the DAC + net1‐mAb group was determined by two‐way ANOVA2 and post hoc Tukey test, ****P < 0.0001. Error bars = s.e.m.
Tumor growth inhibition induced by combining decitabine with netrin‐1 interference is directly mediated by UNC5B and DAPK1
The potential reversibility of epigenetic DNA modifications raised the possibility that inhibition of DNA methylation might induce the re‐expression of these genes, and thus, resensitize cells to several pathways resulting in apoptosis. Therefore, we studied gene re‐expression following DAC treatment in MDA‐MB‐231 and HMLER cell lines, by RNAseq. Indeed, the 626 upregulated genes upon DAC treatments (FC untreated/treated cells > 2) both in MDA‐MB‐231 and HMLER cells were investigated for their Gene Ontology term and their KEGG pathway contributions using the WEB‐based GEne SeT AnaLysis Toolkit (Wang et al, 2013). Enriched Gene Ontology terms were of various nature and included response to molecules of bacterial origin, response to lipopolysaccharide, biological regulation, cell proliferation, and cytokine activity. Enriched KEGG pathways included KEGG osteoclast differentiation, KEGG MAPK signaling pathway, KEGG rheumatoid arthritis, KEGG cytokine–cytokine receptor interaction, pathways in cancer, and NOD‐like receptor signaling pathway (Appendix Fig S5). Genes upregulated upon DAC treatment were associated with many biological functions; however, the profiles of transcriptomic changes suggested that no other pro‐apoptotic pathway participated in the synergistic effect of net1‐mAb and DAC on cell death observed herein.
It has been reported (Qi et al, 2015) that the addition of netrin‐1 to the culture medium of human liver cancer, glioblastoma, and embryonic kidney cell lines induces the expression of the Yes‐associated protein (YAP), of TAZ, a transcriptional coactivator with a PDZ‐binding motif (WWTR1), and of the connective tissue growth factor (CTGF), the transcription of which is initiated by YAP/TAZ. The induction of YAP/TAZ following the addition netrin‐1 was correlated with an increase in cell proliferation and following the addition of netrin‐1 neutralization by specific antibodies, with a decrease in the proliferation and migration of the cell lines analyzed (Qi et al, 2015). Therefore, we verified whether, in DAC‐treated cells, netrin‐1 neutralization leads to the downregulation of the YAP/TAZ signaling pathway, which, in turn, could participate in the anti‐tumoral effects associated with the combined treatment. The effect of the net1‐mAb on the expression of YAP, TAZ, and CTGF, in MDA‐MB‐231 and HMLER cells treated with DAC, was thus investigated. In MDA‐MB‐231 cells, DAC treatment increased the level of TAZ and CTGF expression, while net1‐mAb treatment only induced a slight increase in TAZ expression. In contrast, in HMLER cells, the addition of net1‐mAb only induced the expression of CTGF in DAC‐treated cells, while the expression of YAP and TAZ was not significantly modified (Appendix Fig S6A and B). Collectively, these observations did not seem to be in favor of major role of the YAP/TAZ‐mediated signals in the response of cancer cells to the combined DAC/+net1‐mAB treatment. In a similar context, p53 alterations were reported to cooperate with YAP signaling pathways during cancer cell proliferation (Di Agostino et al, 2016), raising the question of the implication of TP53 in the apoptotic effect observed. In the panel of cells analyzed, T47D cells exhibited no alteration in p53 and did not respond to the combined treatment (Fig EV3F), while other cell lines, mutated (MDA‐MB‐231), blocked (HMLER), or wild type (H460) responded to the combined DAC/+net1‐mAb treatment (Grandin et al, 2016). Taken together, these data suggested that TP53 does not play a determinant role in the tumor growth inhibitory effect conferred by the combination treatment.
Similarly to the data reported by Roulois et al (2015) in colorectal cancer cells, DAC treatment of MDA‐MB‐231 and HMLER cells induced the interferon regulatory factor 7 (IRF7), a key player in the type I interferon (IFN)‐dependent immune responses (Appendix Fig S6C and D). We thus investigated the effect of this transcriptional regulator on the expression of DAPK1, NTN1, or UNC5B upon DAC treatment in MDA‐MB‐231 and HMLER cells. The transient transfection of these two cell lines with a siRNA targeting IRF7 strongly reduced IRF7 expression and prevented its upregulation upon DAC treatment (Appendix Fig S6C and D). However, IRF7 mRNA depletion failed to abrogate DAC induction of DAPK1, NTN1, and UNC5B upregulation, suggesting that IRF7 and more generally the interferon (IFN)‐like response is not implicated in regulating the expression of these genes following DAC treatment.
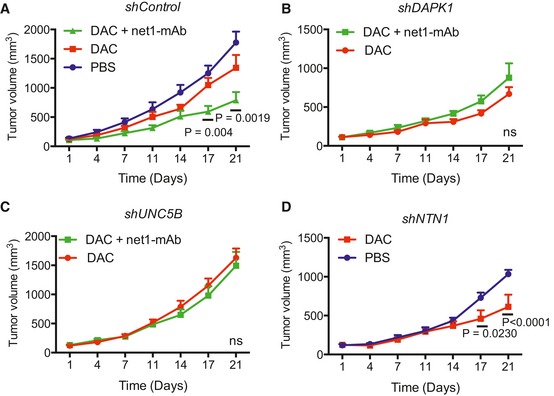
Our final working hypothesis was, therefore, that the tumor growth inhibitory effects conferred by the combination treatment were mediated through the pro‐apoptotic DR pathway. To validate this hypothesis, we generated MDA‐MB‐231 cells stably expressing shRNAs targeting DAPK1, NTN1, or UNC5B transcripts. qRT–PCR analyses indicated that the DAC‐induced expression of DAPK1, NTN1, and UNC5B was efficiently counteracted by gene silencing in the corresponding MDA‐MB‐231‐shDAPK1, MDA‐MB‐231‐shNTN1, and MDA‐MB‐231‐shUNC5B cells (Fig EV4A). We then investigated the responses of the MDA‐MB‐231‐shRNA cells, when engrafted in mammary fat pad and treated or not with the combination treatment (DAC and the net1‐mAb). After validating the levels of DAPK1, UNC5B, and netrin‐1 by IHC (Fig EV4B–D), anti‐active caspase‐3 IHC staining was performed on tumor sections and revealed that the silencing of DAPK1 or UNC5B fully prevents the activation of caspase‐3 mediated by the combination treatment, while DAC monotherapy resulted in a marked increase in caspase‐3 activity when NTN1 was silenced. Furthermore, the silencing of DAPK1 or UNC5B mRNA prevented the inhibition of tumor growth induced by the combination treatment, further confirming our working hypothesis (Fig 5A–C). Moreover, NTN1 silencing was sufficient to induce tumor growth inhibition following DAC monotherapy (Fig 5D). Taken together, these data demonstrate that DAPK1 and UNC5B are the key players involved in the response to the combined DAC and the anti‐netrin‐1 antibody treatment.
Figure EV4. Response of MDA‐MB‐231 cell lines stably transfected with shRNA targeting DAPK1, UNC5B, and NTN1 to treatments combining DAC and net1‐mAb.
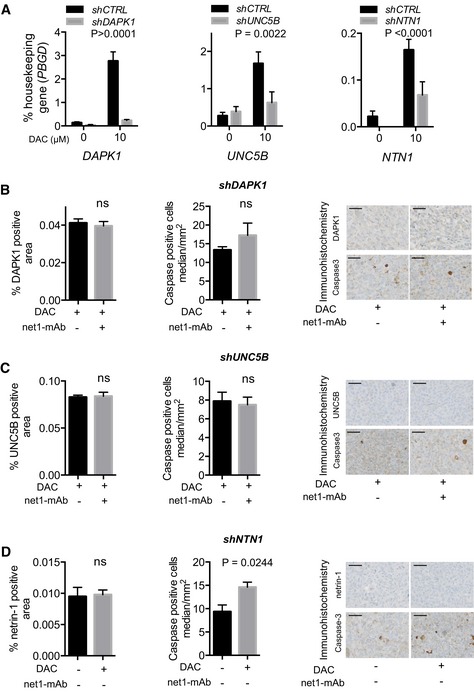
-
AGene expression was measured by qRT–PCR after 72 h in cell lines stably transfected with shRNA. Cells were treated daily with 10 μM DAC. The level of PBGD expression was used as an internal control. Data are expressed as mean ± s.e.m. of at least 3 independent experiments. ****P < 0.0001, one‐way ANOVA.
-
B–DLevels of DAPK1, UNC5B, and netrin‐1 were measured by immunohistochemistry in paraffin embedded xenografts of MDA‐MB‐231 stably transfected with shRNA. Left panels: levels of DAPK1 (B), UNC5B (C), and netrin‐1 (D) in xenografts of MDA‐MB‐231 shDAPK1, shUNC5B, and shNTN1, respectively. Protein levels are expressed as the percentage of total tumor surface; P‐values from at least 3 tumors per group. Middle panels: median number of apoptotic cells per mm2; values from at least 3 tumors per group. Error bars = s.e.m. Mann–Whitney test. ns = not significant. Right panels: representative sections of in vivo protein levels and induction of apoptosis in MDA‐MB‐231 tumors cells depleted of key genes from the netrin‐1 dependence receptor pathway. Scale bars = 50 μm.
Figure 5. Sensitivity of MDA‐MB‐231 cell lines stably transfected with shRNA targeting DAPK1,UNC5B, and NTN1 to treatments combining DAC and net1‐mAb.
-
A–DStably transfected MDA‐MB‐231 cells bearing a control (A), DAPK1 (B), UNC5B (C), or NTN1 shRNA (D) were injected into the mammary fat pad of immuno‐compromised mice. When tumors reached 100–120 mm3, mice were injected subcutaneously with DAC (0.4 mg/kg) or PBS and/or intraperitoneally with net1‐mAb (10 mg/kg). Tumor volumes were measured twice a week, n = 8 mice per group. The statistical significance of the differences obtained between DAC group and DAC + net1‐mAb group for shControl, DAC group and DAC + net1‐mAb group for shDAPK1 and shUNC5B, and PBS group and DAC group for shNTN1, respectively, was determined by two‐way ANOVA and post hoc Tukey test. ****P < 0.0001, ns = not significant. Error bars = s.e.m.
Discussion
The results presented herein provide a proof of concept that inhibition of DNA methylation can sensitize solid tumors to antibodies mediating tumor cell apoptosis. We previously proposed that in netrin‐1‐low tumors, inhibition of the netrin‐1 dependence receptor (DR) pathway may occur either through the downregulation of the receptors or of key signaling pathway partners in colorectal cancer and neuroblastoma (Bernet et al, 2007; Zhu et al, 2013). Interestingly, in the panel of breast cancers analyzed in the current study, low levels of NTN1 expression are associated with the hypermethylation of the CpG island located in the 5′‐end of this gene. Furthermore, this correlation between hypermethylation and gene silencing was also found for DAPK1, an essential partner in the apoptotic netrin‐1/DR pathway. These features were confirmed in breast cancer cell lines, since NTN1 was downregulated and methylated in HMLER and MDA‐MB‐231 cells, and DAPK1 was methylated and downregulated in HMLER cells.
In addition we showed, both in vitro and in xenograft models, that combining inhibition of DNA methylation with an anti‐netrin‐1 antibody resulted in the re‐expression of netrin‐1 and DAPK1 and led to tumor cell death and tumor growth inhibition. The correlation between NTN1 and DAPK1 silencing and DNA hypermethylation does not exclude that their upregulation in DAC‐treated cells might be dependent on additional factors. For example, UNC5B, while unmethylated, was upregulated in DAC‐treated MDA‐MB‐231 cells, and it was previously shown that in breast cancer cells, the downregulation of some methylated genes is dependent on the deposition of the methyl‐CpG binding domain protein 2 (MBD2) within their promoter sequence (Devailly et al, 2015).
It is intriguing that, in the PDX model, the administration of DAC as a monotherapy only resulted in a slight tumor growth inhibitory effect. It is unlikely that the resistance to DAC treatment, used as a monotherapeutic agent, may have resulted from an over‐ or under‐dosage, since several reports have shown that similar DAC doses are efficient in reducing tumor formation in some mouse models, including ApcMin‐induced intestinal neoplasia (Tsai et al, 2012), HRAS‐G12V‐transformed human epithelial kidney, leukemic cells, breast cancer cell lines, and engrafted patient‐derived tumors (Mian et al, 2011; Kilinc et al, 2012). Furthermore, DNA hypomethylation was observed following DAC treatment in MDA‐MB‐231 xenograft tumors and PDX, indicating that DNA methylation was efficiently inhibited by DAC, in vivo. Alternatively, these data would suggest that tumor‐growing conditions can overcome, at least partially, the anti‐tumor effect of DAC. Experimental approaches (Rodriguez‐Paredes & Esteller, 2011b; Yang et al, 2012; Azad et al, 2013) and clinical trials (ClinicalTrials.gov http://www.clinicaltrials.gov) using combinations of DAC with other epigenetic modifiers and/or cytotoxic agents provide interesting strategies for tumor growth reduction. However, these approaches have overlooked an important aspect of DAC, which is the re‐expression of potentially masked targets and pathways. The data reported here provide not only evidence for the importance of ligand/DRs pairs in the regulation of tumor development but also advocate a new strategy based on the specific targeting of genes re‐expressed following a DNA hypomethylating treatment. Finally, inhibitors of DNA methylation could “prime” tumors to netrin‐1 addiction, and the subsequent treatment with a combination of inhibitors of DNA methylation and an anti‐netrin‐1 antibody could result in a powerful therapeutic strategy.
Materials and Methods
Tumor samples
Human breast cancer samples were provided by the tumor bank of the Centre Léon Bérard (Lyon, France). Fresh tissue samples were obtained during breast surgery, prior to any systemic therapy, snap‐frozen in liquid nitrogen, and stored for scientific research in a biological resources repository, according to the French National Ethical Guidelines. Tissue microarrays of paraffin embedded breast tumor sections were obtained from Super Bio Chips (Cliniscience, Nanterre, France).
Cell lines and treatments
The HMLER, MDA‐MB‐157, MDA‐MB‐231, and MDA‐MB‐231‐Luc breast cancer cell lines (Cell Biolabs, San Diego, CA, USA) were maintained in Dulbecco's Minimum Essential Medium F12 Glutamax (DMEM‐F12 Glutamax) (Life Technologies). In addition, human EGF 10 ng/ml (Promocell, Heidelberg, Germany), hydrocortisone 0.5 μg/ml, puromycin 0.5 μg/ml (InVitrogen), and insulin 10 μg/ml (InVitrogen) were added to the medium of HMLER cells. AU565 and T47D cell lines were maintained in RPMI medium (Life Technologies), supplemented with insulin 0.2 U/ml (InVitrogen) for T47D cells. SKBR3 cells were maintained in McCoy's medium (Life Technologies). All of these cell lines were supplemented with 10% FBS (Lonza, Basel, Switzerland) and 1% penicillin/streptomycin (InVitrogen, Carlsbad, CA, USA). Twenty‐four hours after plating, cells were grown for 3 days in fresh medium containing various concentrations of decitabine (5‐aza‐2′‐deoxycytidine, DAC) (Sigma‐Aldrich) or 2 μM 5‐azacytidine (Sigma‐Aldrich), renewed every day. The cells were then cultured, under serum deprived conditions for 48 h, in the presence of the net1‐mAb anti‐netrin‐1 antibody (10 μg/ml) (Netris Pharma, Lyon, France), as well as in the presence or not of the recombinant Flag‐tagged netrin‐1 (5 μg/ml) (Adipogen).
Enforced gene expression and siRNA experiment
The pcDNA3.1 vector coding for DAPK1 (Llambi et al, 2005 ) was used to enforce the expression of DAPK1 in HMLER cells. Empty plasmids or plasmids containing a HA‐tagged DAPK construct were transfected into HMLER cells using Lipofectamine 2000 (Invitrogen). NTN1 siRNA has been previously described (Delloye‐Bourgeois et al, 2009a) and was transfected using Lipofectamine 2000 into HMLER cells, which were previously treated or not with decitabine for 72 h. IRF7 siRNA was transfected using Lipofectamine 2000, and transfected cells were treated for 72 h with DAC. Scramble siRNA (Sigma) was used as a control.
Stable shRNA transfection of cell lines
Control, DAPK1, UNC5B, and NTN1 shRNA plasmids (Sigma Mission shRNA) were transfected into subconfluent cells using Lipofectamine 2000, according to the manufacturer's protocol. After transfection, puromycin (2 μg/ml) was added to the fresh medium as a selection factor. The selection of transfected cells occurred over a 5‐day time‐course. Cells were then trypsinized and diluted to obtain 0–2 clones per well in 96‐well plates. Cell selection using puromycin (2 μg/ml) was conducted throughout the shRNA transfection, selection, and all of the subsequent experiments, in order to conserve gene downregulation. Following their amplifications, cells were treated or not with DAC, and tested for their level of DAPK1/UNC5B/NTN1 expression before and after DAC treatment. Clones that exhibited a “normal” proliferation rate and a low level of expression of the gene of interest were selected.
Caspase‐3 activity and viability assays
Caspase‐3 activity was measured as previously described (Llambi et al, 2005), (Delloye‐Bourgeois et al, 2009a) using the Ac‐DEVD‐AFC substrate assay (Gentaur Biovision, Brussel, Belgium). Alternatively, the percentage of cell death was measured by acridine orange and DAPI staining, using the NucleoCounter NC‐3000 system (ChemoMetec A/S, Allerød, Denmark).
Fluorescent caspase‐3 activity
The apoptotic index was measured using the CellPlayer 96‐Well Kinetic Caspase 3/7 Apoptosis Kit, according to the manufacturer's protocol (Essen bioscience, Hertfordshire, UK). Three thousands cells were plated in 96‐well plates, treated with DAC (10 μM final) or vehicle (PBS) for 48 h. The cells were then incubated in serum free medium enriched with a kinetic apoptosis reagent (1/5,000e), with net1‐mAb (10 μg/ml) and/or with recombinant netrin‐1 (5 μg/ml), and with DAC or PBS. Cells were placed in an IncuCyte FLR or ZOOM with a 10 × objective in a standard cell culture incubator at 37°C and 5% CO2 for 48 h. As a marker of proliferation, and to correct for differential proliferation of cells, the total number of DNA‐containing objects was counted at the end of the experimental time‐course, using Vybrant Green. This number was used to calculate the “apoptotic index”, defined as the ratio of the number of caspase‐3/7 positive objects to the total number of DNA‐containing objects, as recommended by the manufacturer's protocol.
TUNEL assays
For the detection of DNA fragmentation, cells were cultured on coverslips, and, after treatments, fixed in 4% paraformaldehyde for 20 min. TUNEL assays (terminal deoxynucleodityl‐transferase‐mediated dUTP‐biotin nick end labeling) were then performed using 300 U/ml TUNEL enzyme and 6 μM biotinylated dUTP, according to the manufacturer's guidelines (Roche Diagnostics, Meylan, France).
Mouse model of xenografts
Five‐week‐old female athymic Swissnu/nu mice were obtained from Charles River (Ecully, France) and were housed in a specific pathogen‐free animal facility. MDA‐MB‐231 or MDA‐MB‐231‐Luc shRNA cells (2 × 106) were resuspended in 200 μl PBS and implanted into the mammary gland fat pad. When tumors reached an approximate volume of 100 mm3, 10 mg/kg of net1‐mAb (Netris Pharma, France) or an equal volume of PBS (Life Technologies) was injected intraperitoneally, twice a week for 3 weeks. DAC was then injected subcutaneously into the left flank 3 times a week for 3 weeks (dosage: 0.5 mg/kg; vehicle: PBS) or replaced by PBS in the control groups. Tumor volumes were assessed twice a week with a caliper and calculated with the formula V = ½ (length × width2). Furthermore, a patient‐derived xenograft, PDX‐HBC‐146, was generated and housed in the tumor model laboratory (LMT) of the Centre Léon Bérard. Briefly, following isoflurane anesthesia, mice were engrafted in the interscapular area with a ≈ 60 mm3 patient‐derived tumor. When tumors reached 120–150 mm3, mice were injected subcutaneously with DAC (0.5 mg/kg) or PBS 3 times a week for 3 weeks and/or intraperitoneally with net1‐mAb (10 mg/kg) or with a human IgG1 control isotype antibody (Ctrl IgG1, 10 mg/kg, Evitria, Switzerland) twice a week for 3 weeks. Concomitantly, in order to evaluate the effects of the treatments on DNA methylation, cell death, cell proliferation, tumor angiogenesis, and protein expression, some xenografted mice were treated for only one week (as described above) before tumor extraction for further analysis. All experiments were performed in accordance with the relevant guidelines and regulations required by the Animal Ethics Committee (accreditation of laboratory animal care by CECCAP, ENS Lyon‐PBES).
Chicken model for tumor cell dissemination
As previously described (Delloye‐Bourgeois et al, 2009b), MDA‐MB‐231 cells were initially treated for 3 days with DAC 10 μM or PBS (vehicle) and then for 2 days with Control Iso‐mAb or net1‐mAb. After trypsinization, 2 × 106 cells were suspended in 100 μl of PBS/Matrigel (1:1) and seeded on the chorioallantoic membrane (CAM) of 10‐day‐old chicks. On day 17, lungs were harvested from the tumor‐bearing embryos and genomic DNA was extracted with a NucleoSpin Tissue kit (Macherey‐Nagel). Metastasis was quantified by PCR‐based detection of the human Alu sequence, using primers for avian repeated element‐specific sequences as controls (see Appendix Table S1). For both couples of primers, metastasis was assessed by polymerase activation at 95°C for 2 min followed by 40 cycles at 95°C for 30 s, 63°C for 30 s, and 72°C for 30 s. Genomic DNA extracted from chicken breast and lungs of healthy chick embryos was used to determine the threshold between neuroblastoma (NB) cell‐invaded and non‐invaded lungs. All experiments were performed in accordance with the relevant guidelines and regulations required by the Animal Ethics Committee (accreditation of laboratory animal care by CECCAP, ENS Lyon‐PBES).
Immunohistochemistry analysis of xenografted cell lines and patient biopsies
Immunohistochemistry staining was performed on an automated immunostainer (Ventana Discovery XT, Roche, Meylan, France) using the DABmap Kit according to the manufacturer's instructions. Tissue samples were fixed in 10% buffered formalin and embedded in paraffin. Following antigen unmasking (citrate buffer pH 7.3, 98°C for 35 min), immunostainings were performed with a polyclonal rat anti‐mouse netrin‐1 antibody (R&D), a rabbit polyclonal anti‐DAPK1 (Acris), or an UNC5B antibody (Sigma), using the Novolink kit (Leica) for revelation. Apoptotic cell staining was performed using a rabbit cleaved caspase‐3‐specific antibody (Cell Signaling) and the In Situ Cell Death Detection kit POD (Roche). Angiogenesis was measured using a rabbit polyclonal CD31 (Platelet Endothelial Cell Adhesion Molecule‐1) antibody (ANASPEC), and the rate of cell proliferation was determined using a mouse monoclonal MIB‐1‐specific antibody (DAKO). Image analysis was performed using a light microscope (Eclipse E400, Nikon France, Champigny, France) equipped with a tri‐CDD video camera (Sony, Japan). Quantitative values were determined by morphometric analysis (Histolab, Microvision Instruments, Evry, France) of at least 40 fields per staining at a ×200 magnification for the xenograft tumor sections, and one field per staining of the total tissue section at ×40 magnification for the tissue microarray. The total surface occupied by the tumor was automatically measured, and the positively stained at each surface was expressed as a percentage of the total surface for each field analyzed.
RNA isolation
Total RNA was extracted from biopsies and xenografts using the TRIzol‐Reagent (Ambion, Life Technologies) and from cell lines using the Nucleospin RNAII kit (Macherey‐Nagel, Hoerdt, Germany), according to the manufacturer's instructions. For RNA‐seq experiments, RNA purity, integrity, and quantification were assessed using agarose gel electrophoresis and a NanoDrop 1000 (Thermo Scientific, Wilmington, DE, USA). Pools of three to five independent extractions were sent for high‐throughput sequencing to the Beijing Genomics Institute (BGI, Hong‐Kong, China).
Reverse transcription and quantitative real‐time RT–PCR
One microgram of RNA was reverse‐transcribed, using the iScript cDNA Synthesis Kit (Bio‐Rad, Ivry, France). Quantitative RT–PCR was performed using a Mini opticon (Bio‐Rad) and the SYBR supermix qPCR kit (Bio‐Rad). Polymerase was activated at 95°C for 3 min, followed by 45 cycles of amplification and 30 s of cooling. Moreover, gene expression profiles of human samples and cell lines were validated using 3 other standard housekeeping genes, namely PBGD, GAPDH, and MBD2, to confirm the results. Primer sequences for the genes targeted are shown in Appendix Table S1.
DNA extraction
Biopsy samples and xenografted cell lines were cryogrinded in liquid nitrogen. DNA was then extracted and purified using the Nucleospin tissue DNA extraction kit (Macherey‐Nagel), according to the manufacturer's protocol. DNA from cell lines was directly extracted using the standard protocol (Auriol et al, 2005).
Bisulfite treatment of genomic DNA
Two hundred nanograms of genomic DNA was added to 1.8 μg of standard plasmid DNA (pGL3 Basic) and converted using the Epitect Bisulfite kit (Qiagen), according to the manufacturer's protocol.
Amplification of bisulfite converted DNA and pyrosequencing
Sets of biotinylated NTN1, DAPK1, and primers were designed within the promoter region (Fig 1D). As a control, non‐modified and modified GAPDH sets of primers were used to assess the efficiency of the bisulfite DNA conversion. Modified DNA was amplified in a total volume of 50 μl using the Hotstartaq (Qiagen) kit, in the presence of 1 mM MgCl2 for the DAPK1 and NTN1 primers.
Thermal profiles were as follows: 95°C for 10 min followed by 50 cycles of 95°C for 30 s, 50°C (NTN1) or 52°C (DAPK1) for 30 s and 1 min 30 s (NTN1 and DAPK1) of extension followed by a 10‐min final elongation. The primer sequences are shown in Appendix Table S1. The quality and quantity of the PCR product were confirmed on a 2% agarose gel.
PCR products were then pyrosequenced using the Pyromark kit (Qiagen). Reverse single‐stranded biotinylated templates were isolated using the PyroMark Vacuum Prep WorkStation (Qiagen). Forty microliters of PCR product was added to 38 μl of binding buffer (Qiagen) and 2 μl streptavidin sepharose high‐performance beads (GE Healthcare®). The mixtures were shaken for 10 min at 200 g (revolution per minute). After agitation, beads covered with biotinylated DNA were collected and retained on the filter probes by permanent vacuum. The filter probes were successively immerged in different baths: in ethanol 70% for 5 s, in PyroMark denaturation solution for 5 s, and in PyroMark wash buffer 1× for 15 s (Qiagen). The vacuum was then turned off, and the beads fixing DNA strands were released into a 96‐well plate containing 25 μl of annealing buffer with 0.3 μM of sequencing primer in each well. The sequencing plate was kept at 80°C for 2 min and at room temperature for 5 min. Pyrosequencing reactions were performed in a PyroMark Q96 system using PyroGold reagents (Qiagen). Results were analyzed using the PyroMark Software.
DNA methylation analysis
Bisulfite sequencing, used to determine the CpG methylation patterns of DAPK1 and NTN1 5′UTR regions (Appendix Table S1), was performed as described (Beygo et al, 2013). Briefly, after a first amplification using sequence‐specific primers, PCR fragments were tagged in a second amplification step and sequenced using the Roche/454 GS junior system according to the manufacturer's protocol (Roche emPCR Amplification Method Manual—Lib‐A and Roche Sequencing Method Manual). Data were analyzed using Amplikyzer (https://pypi.python.org/pypi/amplikyzer/0.97).
Methyl‐Capture‐sequencing (Methyl‐Cap‐seq)
Genomic DNAs (1 μg) were sheared to an average length of 300–600 bp. Methylated DNA fragments were isolated using beads containing the Methyl‐CpG‐binding domain of MBD2, according to manufacturer's recommendations (MethylMiner, InVitrogen). After sequencing, using the Illumina 2000 high‐throughput sequencing technology by BGI service (Beijing, China), 30–40 million 50‐bp reads were obtained for each input and bound fraction and analyzed using R and bioconductor packages.
RNA‐seq analysis
Reads were aligned on the UCSC Homo sapiens hg19 genome using TopHat2. Differential expression analyses were performed as described (Kim et al, 2013), using the Galaxy server (https://usegalaxy.org/). Only genes with at least 1 read per million (RPM) were kept for subsequent analyses. Enriched Gene Ontology terms and KEGG pathways were identified using Gene Set Enrichment Analysis (Subramanian et al, 2005) with genes preranked according to their fold change induced by DAC treatment.
Data deposition
The MeDP and the RNAseq data from this publication have been submitted to the GEO database the accession number GSE80177.
Author contributions
MG contributed to the experimental design and performed the work, with the help of PM, GD, BG, YB, AG, CF, IP, JGD. YB provided some scientific insight and technical support on 454 Junior DNA methylation sequencing experiments and their analysis. NG conducted immunohistochemical staining of xenografted murine samples. AB has provided the net1‐mAb antibody and technical support. ZH and AG provided some scientific insight and technical support on pyrosequencing experiments and their analysis. MG participated in the writing of the manuscript. PM and RD proposed the project, experimental design, and wrote the manuscript.
Conflict of interest
PM and AB declare having a conflict of interest in this study as co‐founders and shareholders of Netris Pharma.
The paper explained.
Problem
In a substantial part of human cancers, netrin‐1 (NTN1) is upregulated and this upregulation is inhibiting apoptosis induced by its so‐called dependence receptors, DCC and UNC5H, and thus promotes tumor progression. However, in other cancers, the selective inhibition of this dependence receptor death pathway relies on the silencing of pro‐apoptotic effector proteins. A large fraction of human breast tumors exhibits simultaneous DNA methylation‐dependent loss of expression of NTN1 and of DAPK1, a serine threonine kinase known to transduce the netrin‐1 dependence receptor pro‐apoptotic pathway.
Results
Results described in this manuscript propose that the inhibition of DNA methylation by drugs such as decitabine restores the expression of both NTN1 and DAPK1, in netrin‐1‐low cancer cells. Combination of decitabine with NTN1 silencing strategies or with an anti‐netrin‐1 neutralizing antibody potentiates tumor cell death and inhibits tumor growth in different animal models including patient‐derived xenografts.
Impact
With more than 500,000 death worldwide in 2012, breast cancer is one of the most frequent cancer and represents a therapeutic challenge. Our data suggest that combining DNA methylation inhibitors with netrin‐1 neutralizing agents could be a valuable strategy for combating netrin‐1 low breast tumors cancer, which may represent 40% of breast cancers.
Supporting information
Appendix
Expanded View Figures PDF
Review Process File
Acknowledgements
We are grateful to Drs. A Puisieux and AP Morel for the HMLER cells, and to Sandrine Viala for her nice and efficient work. We thank Dr B. Manship for editing of this manuscript and helpful discussions. This work was supported by institutional grants from the CNRS, the University of Lyon, the Centre Léon Bérard, the Ligue Contre le Cancer, the INCA, the ANR, the ERC, the EC FP7 Hermione‐2man and from the Fondation Bettencourt. MG was supported by a fellowship grant from the LabEx DEVweCAN.
EMBO Mol Med (2016) 8: 863–877
References
- Auriol E, Billard LM, Magdinier F, Dante R (2005) Specific binding of the methyl binding domain protein 2 at the BRCA1‐NBR2 locus. Nucleic Acids Res 33: 4243–4254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azad N, Zahnow CA, Rudin CM, Baylin SB (2013) The future of epigenetic therapy in solid tumours–lessons from the past. Nat Rev Clin Oncol 10: 256–266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baylin SB, Ohm JE (2006) Epigenetic gene silencing in cancer – a mechanism for early oncogenic pathway addiction? Nat Rev Cancer 6: 107–116 [DOI] [PubMed] [Google Scholar]
- Bernet A, Mazelin L, Coissieux MM, Gadot N, Ackerman SL, Scoazec JY, Mehlen P (2007) Inactivation of the UNC5C Netrin‐1 receptor is associated with tumor progression in colorectal malignancies. Gastroenterology 133: 1840–1848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beygo J, Citro V, Sparago A, De Crescenzo A, Cerrato F, Heitmann M, Rademacher K, Guala A, Enklaar T, Anichini C et al (2013) The molecular function and clinical phenotype of partial deletions of the IGF2/H19 imprinting control region depends on the spatial arrangement of the remaining CTCF‐binding sites. Hum Mol Genet 22: 544–557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bongo JB, Peng DQ (2014) The neuroimmune guidance cue netrin‐1: a new therapeutic target in cardiovascular disease. J Cardiol 63: 95–98 [DOI] [PubMed] [Google Scholar]
- Castets M, Broutier L, Molin Y, Brevet M, Chazot G, Gadot N, Paquet A, Mazelin L, Jarrosson‐Wuilleme L, Scoazec JY et al (2012) DCC constrains tumour progression via its dependence receptor activity. Nature 482: 534–537 [DOI] [PubMed] [Google Scholar]
- Cirulli V, Yebra M (2007) Netrins: beyond the brain. Nat Rev Mol Cell Biol 8: 296–306 [DOI] [PubMed] [Google Scholar]
- Dawson MA, Kouzarides T (2012) Cancer epigenetics: from mechanism to therapy. Cell 150: 12–27 [DOI] [PubMed] [Google Scholar]
- Delloye‐Bourgeois C, Brambilla E, Coissieux MM, Guenebeaud C, Pedeux R, Firlej V, Cabon F, Brambilla C, Mehlen P, Bernet A (2009a) Interference with netrin‐1 and tumor cell death in non‐small cell lung cancer. J Natl Cancer Inst 101: 237–247 [DOI] [PubMed] [Google Scholar]
- Delloye‐Bourgeois C, Fitamant J, Paradisi A, Cappellen D, Douc‐Rasy S, Raquin MA, Stupack D, Nakagawara A, Rousseau R, Combaret V et al (2009b) Netrin‐1 acts as a survival factor for aggressive neuroblastoma. J Exp Med 206: 833–847 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devailly G, Grandin M, Perriaud L, Mathot P, Delcros JG, Bidet Y, Morel AP, Bignon JY, Puisieux A, Mehlen P et al (2015) Dynamics of MBD2 deposition across methylated DNA regions during malignant transformation of human mammary epithelial cells. Nucleic Acids Res 43: 5838–5854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Agostino S, Sorrentino G, Ingallina E, Valenti F, Ferraiuolo M, Bicciato S, Piazza S, Strano S, Del Sal G, Blandino G (2016) YAP enhances the pro‐proliferative transcriptional activity of mutant p53 proteins. EMBO Rep 17: 188–201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elenbaas B, Spirio L, Koerner F, Fleming MD, Zimonjic DB, Donaher JL, Popescu NC, Hahn WC, Weinberg RA (2001) Human breast cancer cells generated by oncogenic transformation of primary mammary epithelial cells. Genes Dev 15: 50–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitamant J, Guenebeaud C, Coissieux MM, Guix C, Treilleux I, Scoazec JY, Bachelot T, Bernet A, Mehlen P (2008) Netrin‐1 expression confers a selective advantage for tumor cell survival in metastatic breast cancer. Proc Natl Acad Sci USA 105: 4850–4855 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grandin M, Meier M, Delcros JG, Nikodemus D, Reuten R, Patel TR, Goldschneider D, Orriss G, Krahn N, Boussouar A et al (2016) Structural decoding of the Netrin‐1/UNC5 interaction and its therapeutical implications in cancers. Cancer Cell 29: 173–185 [DOI] [PubMed] [Google Scholar]
- Guenebeaud C, Goldschneider D, Castets M, Guix C, Chazot G, Delloye‐Bourgeois C, Eisenberg‐Lerner A, Shohat G, Zhang M, Laudet V et al (2010) The dependence receptor UNC5H2/B triggers apoptosis via PP2A‐mediated dephosphorylation of DAP kinase. Mol Cell 40: 863–876 [DOI] [PubMed] [Google Scholar]
- Harter PN, Zinke J, Scholz A, Tichy J, Zachskorn C, Kvasnicka HM, Goeppert B, Delloye‐Bourgeois C, Hattingen E, Senft C et al (2014) Netrin‐1 expression is an independent prognostic factor for poor patient survival in brain metastases. PLoS One 9: e92311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedrick L, Cho KR, Fearon ER, Wu TC, Kinzler KW, Vogelstein B (1994) The DCC gene product in cellular differentiation and colorectal tumorigenesis. Genes Dev 8: 1174–1183 [DOI] [PubMed] [Google Scholar]
- Kilinc D, Ozdemir O, Ozdemir S, Korgali E, Koksal B, Uslu A, Gultekin YE (2012) Alterations in promoter methylation status of tumor suppressor HIC1, SFRP2, and DAPK1 genes in prostate carcinomas. DNA Cell Biol 31: 826–832 [DOI] [PubMed] [Google Scholar]
- Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL (2013) TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol 14: R36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krimpenfort P, Song JY, Proost N, Zevenhoven J, Jonkers J, Berns A (2012) Deleted in colorectal carcinoma suppresses metastasis in p53‐deficient mammary tumours. Nature 482: 538–541 [DOI] [PubMed] [Google Scholar]
- Llambi F, Causeret F, Bloch‐Gallego E, Mehlen P (2001) Netrin‐1 acts as a survival factor via its receptors UNC5H and DCC. EMBO J 20: 2715–2722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llambi F, Lourenco FC, Gozuacik D, Guix C, Pays L, Del Rio G, Kimchi A, Mehlen P (2005) The dependence receptor UNC5H2 mediates apoptosis through DAP‐kinase. EMBO J 24: 1192–1201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund P, Kotova I, Kedinger V, Khanwalkar H, Voltz E, Hahn WC, Gronemeyer H (2011) Transformation‐dependent silencing of tumor‐selective apoptosis‐inducing TRAIL by DNA hypermethylation is antagonized by decitabine. Mol Cancer Ther 10: 1611–1623 [DOI] [PubMed] [Google Scholar]
- Mehlen P, Rabizadeh S, Snipas SJ, Assa‐Munt N, Salvesen GS, Bredesen DE (1998) The DCC gene product induces apoptosis by a mechanism requiring receptor proteolysis. Nature 395: 801–804 [DOI] [PubMed] [Google Scholar]
- Mehlen P, Guenebeaud C (2010) Netrin‐1 and its dependence receptors as original targets for cancer therapy. Curr Opin Oncol 22: 46–54 [DOI] [PubMed] [Google Scholar]
- Mian OY, Wang SZ, Zhu SZ, Gnanapragasam MN, Graham L, Bear HD, Ginder GD (2011) Methyl‐binding domain protein 2‐dependent proliferation and survival of breast cancer cells. Mol Cancer Res 9: 1152–1162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morel AP, Lievre M, Thomas C, Hinkal G, Ansieau S, Puisieux A (2008) Generation of breast cancer stem cells through epithelial‐mesenchymal transition. PLoS One 3: e2888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paradisi A, Creveaux M, Gibert B, Devailly G, Redoulez E, Neves D, Cleyssac E, Treilleux I, Klein C, Niederfellner G et al (2013) Combining chemotherapeutic agents and netrin‐1 interference potentiates cancer cell death. EMBO Mol Med 5: 1821–1834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulling LC, Grimes MJ, Damiani LA, Juri DE, Do K, Tellez CS, Belinsky SA (2009) Dual promoter regulation of death‐associated protein kinase gene leads to differentially silenced transcripts by methylation in cancer. Carcinogenesis 30: 2023–2030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi Q, Li DY, Luo HR, Guan KL, Ye K (2015) Netrin‐1 exerts oncogenic activities through enhancing Yes‐associated protein stability. Proc Natl Acad Sci USA 112: 7255–7260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramkhelawon B, Hennessy EJ, Menager M, Ray TD, Sheedy FJ, Hutchison S, Wanschel A, Oldebeken S, Geoffrion M, Spiro W et al (2014) Netrin‐1 promotes adipose tissue macrophage retention and insulin resistance in obesity. Nat Med 20: 377–384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raval A, Tanner SM, Byrd JC, Angerman EB, Perko JD, Chen SS, Hackanson B, Grever MR, Lucas DM, Matkovic JJ et al (2007) Downregulation of death‐associated protein kinase 1 (DAPK1) in chronic lymphocytic leukemia. Cell 129: 879–890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez‐Paredes M, Esteller M (2011a) Cancer epigenetics reaches mainstream oncology. Nat Med 17: 330–339 [DOI] [PubMed] [Google Scholar]
- Rodriguez‐Paredes M, Esteller M (2011b) A combined epigenetic therapy equals the efficacy of conventional chemotherapy in refractory advanced non‐small cell lung cancer. Cancer Discov 1: 557–559 [DOI] [PubMed] [Google Scholar]
- Roulois D, Loo Yau H, Singhania R, Wang Y, Danesh A, Shen SY, Han H, Liang G, Jones PA, Pugh TJ et al (2015) DNA‐demethylating agents target colorectal cancer cells by inducing viral mimicry by endogenous transcripts. Cell 162: 961–973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin SK, Nagasaka T, Jung BH, Matsubara N, Kim WH, Carethers JM, Boland CR, Goel A (2007) Epigenetic and genetic alterations in Netrin‐1 receptors UNC5C and DCC in human colon cancer. Gastroenterology 133: 1849–1857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES et al (2005) Gene set enrichment analysis: a knowledge‐based approach for interpreting genome‐wide expression profiles. Proc Natl Acad Sci USA 102: 15545–15550 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang X, Wu W, Sun SY, Wistuba II, Hong WK, Mao L (2004) Hypermethylation of the death‐associated protein kinase promoter attenuates the sensitivity to TRAIL‐induced apoptosis in human non‐small cell lung cancer cells. Mol Cancer Res 2: 685–691 [PubMed] [Google Scholar]
- Tsai HC, Li H, Van Neste L, Cai Y, Robert C, Rassool FV, Shin JJ, Harbom KM, Beaty R, Pappou E et al (2012) Transient low doses of DNA‐demethylating agents exert durable antitumor effects on hematological and epithelial tumor cells. Cancer Cell 21: 430–446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker JA, Hughes DA, Hedges DJ, Anders BA, Laborde ME, Shewale J, Sinha SK, Batzer MA (2004) Quantitative PCR for DNA identification based on genome‐specific interspersed repetitive elements. Genomics 83: 518–527 [DOI] [PubMed] [Google Scholar]
- Wang J, Duncan D, Shi Z, Zhang B (2013) WEB‐based GEne SeT AnaLysis Toolkit (WebGestalt): update 2013. Nucleic Acids Res 41: W77–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X, Lay F, Han H, Jones PA (2010) Targeting DNA methylation for epigenetic therapy. Trends Pharmacol Sci 31: 536–546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang D, Torres CM, Bardhan K, Zimmerman M, McGaha TL, Liu K (2012) Decitabine and vorinostat cooperate to sensitize colon carcinoma cells to Fas ligand‐induced apoptosis in vitro and tumor suppression in vivo. J Immunol 188: 4441–4449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Y, Li Y, Haraguchi S, Yu M, Ohira M, Ozaki T, Nakagawa A, Ushijima T, Isogai E, Koseki H et al (2013) Dependence receptor UNC5D mediates nerve growth factor depletion‐induced neuroblastoma regression. J Clin Investig 123: 2935–2947 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix
Expanded View Figures PDF
Review Process File