

Abstract
Inflammatory bowel disease (IBD) arises by unknown environmental triggers in genetically susceptible individuals. Epigenetic regulation of gene expression may integrate internal and external influences and may thereby modulate disease susceptibility. Epigenetic modification may also affect the germ-line and in certain contexts can be inherited to offspring. This study investigates epigenetic alterations consequent to experimental murine colitis induced by dextran sodium sulphate (DSS), and their paternal transmission to offspring. Genome-wide methylome- and transcriptome-profiling of intestinal epithelial cells (IECs) and sperm cells of males of the F0 generation, which received either DSS and consequently developed colitis (F0DSS), or non-supplemented tap water (F0Ctrl) and hence remained healthy, and of their F1 offspring was performed using reduced representation bisulfite sequencing (RRBS) and RNA-sequencing (RNA-Seq), respectively. Offspring of F0DSS males exhibited aberrant methylation and expression patterns of multiple genes, including Igf1r and Nr4a2, which are involved in energy metabolism. Importantly, DSS colitis in F0DSS mice was associated with decreased body weight at baseline of their F1 offspring, and these F1 mice exhibited increased susceptibility to DSS-induced colitis compared to offspring from F0Ctrl males. This study hence demonstrates epigenetic transmissibility of metabolic and inflammatory traits resulting from experimental colitis.
The inflammatory bowel diseases (IBD) Crohn’s disease (CD) and ulcerative colitis (UC) are triggered by yet unknown environmental factors in genetically susceptible individuals1,2,3. IBD has dramatically risen in incidence over the last decades in the Western world, and more recently in countries adopting Western lifestyles1. Concordance rates in monozygotic twins of 40–50% and 10–15% in CD and UC4,5,6,7, respectively, indicate the relative contribution of genetic vs. environmental risks. In the last years genome-wide association studies (GWAS) identified thousands of single-nucleotide polymorphisms (SNPs) associated with complex diseases, and have been particularly successful in CD and UC8. It is estimated that only approximately 16% of IBD disease heritability can be explained by known common genetic risk factors9,10. While genetic interactions, which are not accounted for in these estimates, but which are consistent with observable data in human complex disease genetics11 and in model systems12, may create ‘phantom heritability’ and hence may possibly explain a large part of the current ‘missing heritability’13, additional, non-genetic, environmentally-imprinted disease triggers, e.g. epigenetic modifications, could contribute to the heritable fraction of disease risk.
Epigenetic alterations, including methylation, histone modifications and RNA-mediated effects, regulate gene expression, whereas the DNA sequence itself remains unaltered. DNA becomes predominantly methylated at cytosines (CpG sites or CpGs), but methylation at non-CpG sites has also been detected14,15,16. The highly dynamic nature, cell-specific patterns and genomic context of DNA methylation are of great importance in cellular and metabolic processes, development and disease17,18. Recent studies showed aberrant DNA methylation profiles in intestinal tissues of IBD patients19,20,21,22,23,24,25. Importantly, some epigenetic marks are acquired during periods of developmental plasticity and are not completely erased during epigenetic reprogramming of non-imprinted genes after fertilization allowing them to being passed on to the next generation26,27,28,29. Using this mechanism of inheritance, epigenomes of gametes serve as ‘messengers of ancestral exposures’ (epigenetic memory) and ‘inform’ offspring about prevailing environmental conditions30,31,32,33. Some of the recent studies report a maternal inheritance of epigenetic effects, which, however, are difficult to separate from in utero exposure or from other factors that can be passed on to the next generations via social or cultural inheritance systems30. In contrast, studying epigenetic inheritance in the paternal system has the advantage that sperm material likely serves as the only ‘carrier’ of epigenetic information to the next generation. However, in mammals, paternal and maternal methylomes go through active de-methylation at the majority of CpGs34. Only a few rare methylation sites can escape this process of ‘epigenetic reprogramming’ and can be passed on to the next generation35. Interestingly, a human cohort study investigating paternal epigenetic inheritance demonstrated that parental grandfathers, who were overfed in their childhood, had grandchildren with an increased mortality, a higher risk of diabetes and cardiovascular diseases36. Furthermore, a recent study investigating the role of DNA methylation in a trans-generational mouse model of undernourishment showed that under-nutrition during prenatal life perturbed epigenetic reprogramming of sperms with concomitant DNA methylation changes in the adult brain and liver tissues, which in turn were associated with metabolic disease (early-life adiposity, impaired pancreatic function, glucose intolerance) in offspring37. Moreover, a recent study in mice suggested that chronic paternal high fat diet (HFD) reprograms pancreatic β-cell function in their female offspring, which leads to impaired glucose-insulin homeostasis28. Similarly, a paternal low-protein diet can affect offspring metabolism by increasing expression of many hepatic genes involved in lipid and cholesterol biosynthesis, which correlated with aberrant cytosine methylation in offspring of males fed with a low-protein diet30. In addition, paternally inherited effects were linked to obesity38,39,40,41, diabetes29,42, glucose metabolism43, hepatic wound healing44, behavioral and stress responses45,46,47,48,49,50 and toxin-inherited effects51,52,53. These studies suggest that the paternal lifestyle permanently influenced by certain environmental factors can affect spermatogenesis and can induce inter-generational transmission of epigenetic marks modulating offspring’s metabolism, contributing to disease risk.
To the best of our knowledge, there is no study to date that has investigated an inter-generational transmission of epigenetic marks in the context of chronic intestinal inflammation. We established a colitis mouse model, which allowed us to elucidate the impact of a chronic environmentally-induced colitis on intestinal DNA methylation and the proposed transmission of aberrant methylation patterns to offspring.
Results
Offspring of DSS colitic males display a lower body weight and increased susceptibility to colitis
To decipher epigenetically altered marks that might contribute to the etiopathogenesis of colitis, we exposed wildtype C57Bl/6(N) male littermate mice at 5 weeks of age (Supplementary Fig. 1a) to three cycles of dextran sodium sulphate (DSS), which resulted in body weight loss and the induction of a chronic colitis in F0DSS mice compared to their F0Ctrl littermates that were meanwhile kept on tap water (Supplementary Fig. 1b). After recovering from the last DSS cycle, males of both groups were mated overnight with healthy female littermates (Supplementary Fig. 1a), before the colon was harvested for histology and length assessment, which is a measure for the extent of inflammation (Supplementary Fig. 1c–e). Next, we compared the offspring of DSS-treated males (‘F1DSS’) with the offspring of healthy males not exposed to DSS (F1Ctrl) (Supplementary Fig. 1f). The litter sizes between offspring of F0Ctrl and F0DSS mice were comparable (data not shown). However, the body weight of male and female offspring of F0DSS mice (‘F1DSS’) was significantly lower compared to offspring of F0Ctrl mice (‘F1Ctrl’; Fig. 1a). F1DSS mice at the age of 14 weeks exhibited a morphologically healthy colon (Fig. 1b). We next studied whether chronic intestinal inflammation in the paternal generation affects their offspring’s susceptibility to acute DSS colitis. For this purpose we applied 3.5% DSS to the drinking water of 7 week-old F1DSS and F1Ctrl mice for 6 consecutive days. Notably, F1DSS mice exhibited higher colitis scores in the proximal colon (Fig. 1c,d) and lower hematocrit levels (Fig. 1e) compared to offspring of healthy F0Ctrl mice. Interestingly, female offspring exhibited more pronounced effects compared to male offspring (Supplementary Fig. 2a–d). We noted no differences in the severity of inflammation in the distal colon of F1DSS compared to F1Ctrl mice, which, however, was severely inflamed which may have obfuscated our ability to discern modest differences (Supplementary Fig. 2e,f). Weight loss during DSS colitis and colonic length were comparable between F1DSS and F1Ctrl mice (Supplementary Fig. 2g,h). In summary, chronic DSS colitis in the F0 generation was associated with lower body weight and modestly increased susceptibility to acute DSS colitis in their F1 offspring, when compared to offspring from non-colitic mice.
Figure 1. Chronic experimental colitis leads to a lower body weight and increased susceptibility to inflammation in the offspring.
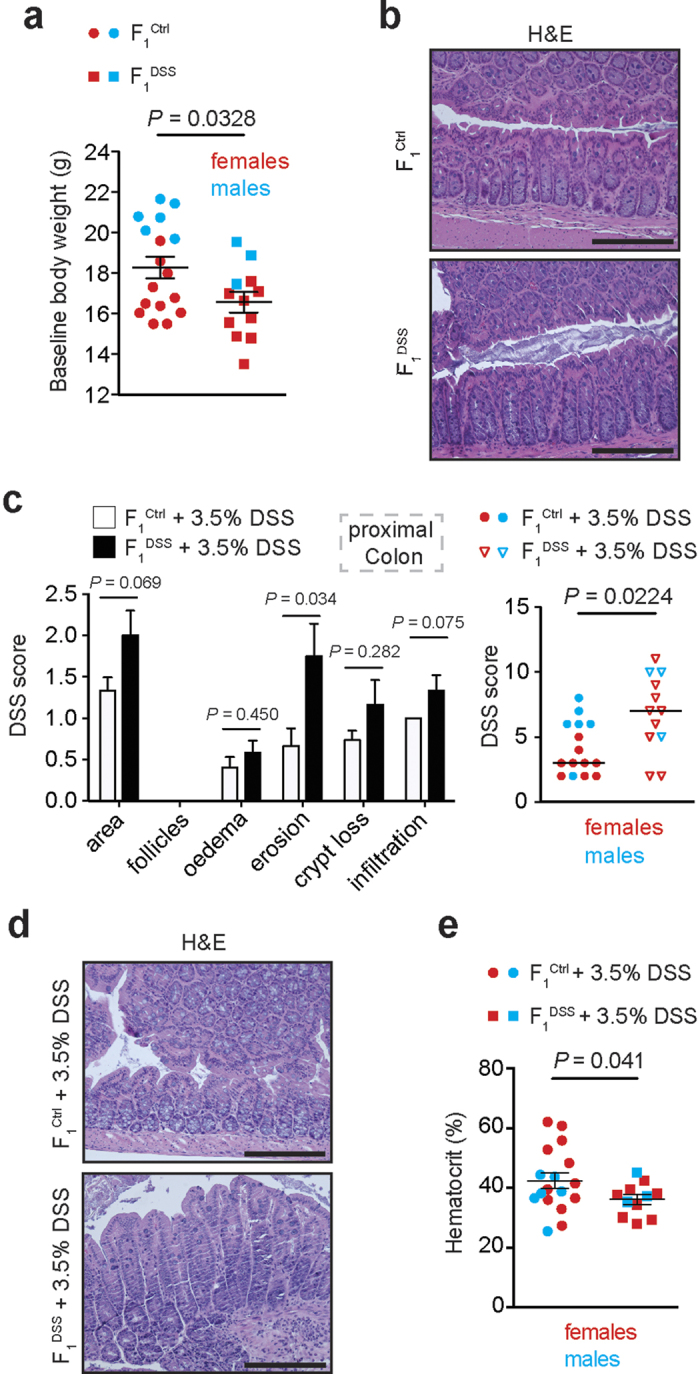
(a) Body weight comparison between 7 week-old female and male littermates that are offspring of either F0Ctrl males (F1Ctrl) or F0DSS males (F1DSS). Mice littermates of both groups were taken from average litter sizes: F1Ctrl and F1DSS mice originated from 3 and 2 different litters, respectively. F1Ctrl females, n = 11; F1DSS females, n = 9; F1Ctrl males, n = 6; F1DSS males, n = 3. Mean ± SEM; Unpaired, two-tailed Student’s t-test. (b) Representative H&E stainings of the distal colon of 13 week-old F1Ctrl and F1DSS mice at baseline conditions (scale bars, 200 μm). (c) DSS subscores (left) and the resulting cumulative score (right) of the proximal colon of F1Ctrl and F1DSS mice after induction of an acute colitis by adding 3.5% DSS in the drinking water of mice of both groups (F1 DSS susceptibility test). F1Ctrl females, n = 9; F1DSS females, n = 9; F1Ctrl males, n = 6; F1DSS males, n = 3. Mean ± SEM; Mann–Whitney U-test. (d) Representative H&E stainings of the proximal colon of F1Ctrl and F1DSS females after 6 days of treatment with 3.5% DSS (scale bars, 200μm). (e) Blood hematocrit value (in %) of F1Ctrl and F1DSS mice after 3.5% DSS treatment. F1Ctrl females, n = 11; F1DSS females, n = 8; F1Ctrl males, n = 6; F1DSS males, n = 3; Mean ± SEM; Unpaired, 1-tailed Student’s t-test.
Chronic inflammation induced by DSS leads to a differential DNA methylation pattern in sperm cells of the F0 and F1 generation
The reduced body weight and increased DSS-susceptibility of F1DSS mice suggested the involvement of epigenetic inheritance from F0DSS males to their offspring. In order to test this, we analyzed sperm cells together with fluorescence-activated cell sorted (FACS) EpCAM+ CD45− colonic intestinal epithelial cells (IECs), which are primarily affected in DSS-induced colitis54, from mice of the F1 and F0 generation (Supplementary Fig. 1f and Supplementary Fig. 3a) and subjected them to RNA-sequencing (RNA-Seq) and reduced representation bisulfite sequencing (RRBS) to assess their global mRNA expression and methylation profile, respectively. The normalized gene expression counts (Supplementary Fig. 3b) of epithelial (Actb, Cdh1/EpCAM) and sperm markers (Odf1, Smcp) confirmed the purity of samples used in this study (Supplementary Fig. 3c–h). Moreover, multi-dimensional scaling (MDS) analysis of the filtered and preprocessed methylome data showed clear differences between epithelial and sperm cells (Supplementary Fig. 4a).
Importantly, MDS analysis of the quality-controlled and filtered CpG sites in F0 and F1 sperm samples (F0: 820,122 CpG sites vs. F1: 727,273 CpG sites identified) pointed to stronger methylation differences in the F1 generation compared to the F0 generation (Fig. 2a,b). Comparing sperm samples of F0Ctrl and F0DSS mice resulted in 823 differentially methylated CpG sites with a methylation difference >0.20 (477 hypo-methylated and 346 hyper-methylated). Notably, 410 of these CpG sites could be annotated to either transcripts and/or promoters (Supplementary Data 1). Surprisingly, comparing the DNA methylation pattern of sperm cells of F1Ctrl mice with sperm cells of F1DSS mice resulted in 4,617 differentially methylated sites (2,429 hypo-methylated and 2,188 hyper-methylated) of which 1,680 CpG sites could be assigned to a specific gene (Supplementary Data 2). The quality controlled filtered CpGs in both generations were annotated to mainly regions outside of CpG islands (CGIs), inter-genic regions and regulatory regions of CTCF-binding sites (transcriptional repressor 11-zinc finger protein or CCCTC-binding factor) (Supplementary Fig. 4b,c).
Figure 2. DSS treatment affects gene expression in the F1 generation.
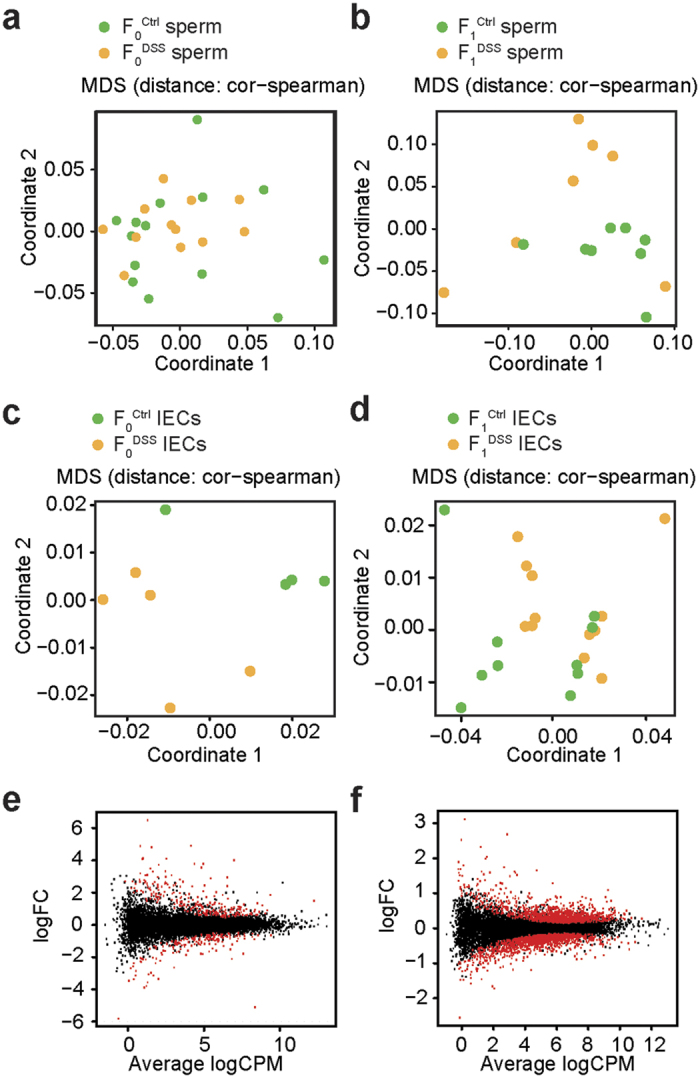
(a) MDS plot showing global effect of DSS treatment on methylation in sperms of F0Ctrl and F0DSS mice. (b) MDS plot showing global effect of DSS treatment in the F0 generation on methylation in sperms of F1Ctrl and F1DSS mice. (c) MDS plot showing global effect of DSS treatment on gene expression in colonic IECs of the F0 generation. (d) MDS plot showing global effect of DSS treatment in the F0 generation on gene expression of IECs in the F1 generation based on filtered high quality gene expression counts (P = 0.034, multivariate ANOVA). (e) MA plot showing the effect size and the significance (red colored dots) for F0Ctrl and F0DSS IECs. Differentially expressed genes (up-regulated or down-regulated) are highlighted in red color. (f ) MA plot showing the effect size and the significance for F1Ctrl and F1DSS IECs. Differentially expressed genes (up-regulated or down-regulated) are highlighted in red color.
We identified an overlap of 66 significantly differentially methylated genes (Supplementary Data 3) between sperm samples of mice of the F0 and F1 generation (37 hypo-methylated and 29 hyper-methylated genes). Importantly, this number is significantly larger than expected by chance in all 10,000 permutations (mean number after 10,000 permutations = 15.518 [3, 30]; Permutation based p-value = 0) (Supplementary Fig. 5a). The 50 most differentially methylated genes overlapping between sperm samples of both generations are shown as heatmaps in Supplementary Fig. 5b,c. Analyzing all 66 overlapping genes, ten CpG sites showed a differential methylation within the promoter or the transcription factor binding region of their annotated genes (Gm128, Pnpla1, Plekhg4, Hdac5, Tjp3, Ttc28, Pnpla1, Mir6991, Hmha1, Gm6484) and two CpGs were annotated to regulatory enhancers near Igf1r and Mcf2l. Moreover, seven differentially methylated sites were located at the same genomic location in both generations, two of which, were present within the intron or exon of the genes Mta1 and Zfp865, respectively. The remaining five sites (chr10:12,2960,510, chr15:37,868,157, chr17:7,882,215, chr4:136,419,115, chr19:16,938,303) were outside of CGIs (Supplementary Data 1 and Supplementary Data 2). Collectively, these data suggest that DSS-induced colitis in male mice affects the DNA methylation pattern in their and their offspring’s sperm cells.
Offspring of DSS-treated males display differential gene expression in colonic IECs at baseline conditions pointing to a dysregulation of the immune response
To explain if the differences in the sperm methylome contribute to a potential gene dysregulation in the F1DSS epithelium, which could possibly contribute to reduced body weight and increased susceptibility to colitis (Fig. 1a,c–e), we assessed the global mRNA expression profile of EpCAM+ CD45− colonic IECs of mice of the F0 and F1 generation with the latter not being exposed to DSS. MDS analysis of the mRNA expression profile not only showed a significant separation of F0DSS and F0Ctrl IECs (Fig. 2c) as expected, but notably also of F1DSS and F1Ctrl IECs (Fig. 2d). Comparative gene expression analysis of IECs of F0DSS and F0Ctrl IECs revealed 230 statistically significantly differentially expressed genes (Padjusted < 0.05; 13,198 tested genes with mean coverage of 30.83x) (Supplementary Data 4, Fig. 2e), whereas 2,358 differentially expressed genes were detected between F1DSS and F1Ctrl IECs (Padjusted < 0.05; Supplementary Data 5, Fig. 2f). The 50 most differentially expressed genes (based on their fold changes) of IECs of the F0 generation and F1 generation are shown as heatmaps in Fig. 3a,b, respectively. Gene set enrichment analysis for the differentially expressed genes in the F1 generation points to a strong deregulation of the immune response in F1DSS epithelium involving key genes of the type I IFN response (Tbkbp1, Gbp3, Gbp9, Ifit1, Ifit2, Ifit3, Oas2, Oas3, Isg15, Oasl2, Ddx60, Mx2). This analysis also identified key genes involved in immune pathways and pro-inflammatory processes (Cxcl3, Cxcl5, Cd8a, C4b, Igj, Nlrc5, Lbp), energy metabolism (Itln1) and vascular development (Ang4) (Fig. 3c). Taken together, offspring of male mice that experienced chronic colitis exhibited an altered intestinal epithelial transcriptome, which highlights a dysregulation of immunological and inflammatory pathways.
Figure 3. Differential gene expression in IECs of the F1 generation induced by chronic inflammation in the F0 generation.

(a) Heatmap showing hierarchical clustering of the top 50 differentially expressed genes (according to fold changes) in IECs between F0Ctrl and F0DSS mice. (b) Heatmap showing hierarchical clustering of the top 50 differentially expressed genes in IECs (according to fold changes) between F1Ctrl and F1DSS mice. Red and blue boxes indicate down-regulated and up-regulated genes, respectively. (c) Gene enrichment analysis of differentially expressed genes in the F1 generation (Padjusted < 0.05, absolute FC > 1). 1 = immune response, 2 = response to external biotic stimulus, 3 = defense response to other organism, 4 = response to other organism, 5 = response to biotic stimulus.
Differential DNA methylation in colonic epithelial cells
After finding significantly differentially methylated CpG sites in F0 sperm (which may shape future F1 somatic methylomes) affected by DSS treatment and dysregulated gene expression in their offspring, we analyzed the methylomes of EpCAM+ CD45− colonic IECs using RRBS. Despite observing a clear separation of the transcriptomes of IECs between F1Ctrl and F1DSS mice, MDS analysis of their methylomes (1,037,337 CpG sites identified) did not show a clear separation into two distinct clusters (Fig. 4a). However, comparing the methylomes of F1Ctrl and F1DSS IECs resulted in 219 differentially methylated CpG sites of which 115 CpGs could be annotated to transcripts and/or promoters (Supplementary Data 6). Consistently, we observed moderate global DNA methylation changes (20% cut-off) in IECs of F1DSS mice compared to F1Ctrl mice (Fig. 4b) indicating that DSS in F0 mice might have a small but significant effect on cytosine methylation in the next generation. For control purposes we also analyzed the methylomes of IECs of the F0 generation (904,007 CpG sites identified), which resulted in 2,107 CpG sites that are differentially methylated between F0Ctrl and F0DSS IECs (353 hypo-methylated and 1,754 hyper-methylated) (Fig. 4c). 866 of these sites could be annotated to transcripts and/or promoters (Supplementary Data 7). The 50 most differentially methylated sites (based on methylation difference) found in IECs of F0 and F1 mice are shown as heatmaps in Fig. 4d,e, respectively. The proportions of genomic region categories for quality-controlled CpGs in IECs of both generations were similar to what was previously observed in the sperm data (Supplementary Fig. 6a,b).
Figure 4. DSS-induced methylation changes are transmissible to offspring leading to a differential methylation profile in colonic epithelial cells.

(a) MDS showing global effect of DSS treatment on methylation in IECs of F1Ctrl and F1DSS mice. (b) Global methylation in F1 IECs is shown for methylated CpG sites in F1Ctrl (x-axis) compared to F1DSS mice (y-axis). Blue and red dots indicate significant CpG sites associated with P < 0.05 and a methylation difference >0.20. (c) MDS plot showing global effect of DSS treatment on methylation IECs in the F0 generation based on the filtered high quality methylation values for each covered position ranging from 0 to 1 (P = 0.001, multivariate ANOVA). (d) Heatmap showing hierarchical clustering of the top 50 differentially methylated genes between F0Ctrl and F0DSS IECs. The genes along with their genomic coordinates and associated site are also shown on the right column. (e) Heatmap showing hierarchical clustering of the top 50 differentially methylated genes between F1Ctrl and F1DSS mice. The genes along with their genomic coordinates and associated site are also shown on the right column. Blue and red boxes indicate high and low methylation, respectively.
An overlap of differentially methylated genes in F0 and F1 IECs resulted in 10 genes (two hypo-methylated and eight hyper-methylated in both generations) (Supplementary Data 8). Four of the ten sites showed a differential DNA methylation within the promoter or the transcription factor-binding region of their annotated genes in F1 IECs (associated with genes: Ick, Adcy6, Rbfox3 and Tspear). Only one differentially methylated site was located at the same inter-genic genomic location and was hypo-methylated in both generations (chr14: 32,700,214).
The DNA methylation pattern of a subset of paternally imprinted genes from the GeneImprint database55 was also checked in the epithelial and sperm samples to show that the bisulfite conversion for the RRBS protocol worked. As expected, we observed either no methylation (Igf2 and Dio3 genes), complete methylation (Gpr1, Peg10 and Magi2 genes) or partial methylation (Mest and Plagl1 genes) for these ‘control’ loci in IECs (Supplementary Fig. 7a–d), however, minor shifts were observed in the imprinting patterns of sperm cells (Supplementary Fig. 7e–h).
Finally, the overlap analysis of differential expression and differential DNA methylation in F1 IECs resulted in 13 genes (Supplementary Data 9, Supplementary Fig. 8a,b). 8,662 out of 10,000 permutations resulted in an overlap greater or equal to 13 (mean number after 10,000 permutations = 16.984 [3, 37]; Permutation based p-value = 0.86) (data not shown). In summary, these results suggest that DSS-induced colitis in F0 males affects the DNA methylation pattern of their germ-line and that intergenerational inheritance of these epigenetic modifications are associated with alterations of the IEC transcriptome and methylome in their offspring.
Patterns of epigenetic inheritance
To identify potential candidate genes with DNA methylation patterns that are passed on to the F1 generation, we screened for positional overlaps between differentially methylated sites in sperm samples of the F0 and F1 generation as well as of IECs of the F1 generation, which resulted in three inter-generationally inherited genes (Rbfox3, Msi2 and Ttc7) (Supplementary Data 3 and Supplementary Data 7, Fig. 5a). Interestingly, this number is significantly larger than expected by chance in 9,700 out of 10,000 permutations (mean number after 10,000 permutations = 0.677 [0, 6]; Permutation based p-value = 0.03) (Supplementary Fig. 9a). Furthermore, the overlap analysis, comparing differential DNA methylation in sperm samples of the F0 and F1 generation with the differential expression in F1 IECs, suggested 14 different candidate genes that may be epigenetically inherited (Supplementary Data 10, Fig. 5a). Nine genes were negatively regulated of which five were hypo-methylated and up-regulated (Sema6a, Tjp2, Hdac5, Arhgef3, Dock6) and four hyper-methylated and down-regulated (Ankrd13b, Blvrb, Nr4a2, Eya2). Five were positively regulated with four hypo-methylated and down-regulated (Edar, Bcl9l, Igf1r, Mta1) and one hyper-methylated and up-regulated (Nedd4l). The regional plots showing the differentially methylated sites for Igf1r, Nr4a2, Hdac5 and Mta1 are shown in Fig. 5b,c and Supplementary Fig. 9b,c.
Figure 5. Epigenetic alterations induced by chronic inflammation in the F0 generation were stably transmitted to offspring.

(a) Overlap of differentially methylated genes between sperm samples of F0 and F1 mice (same direction of effect) with differentially expressed genes in F1 IECs as well as with differentially methylated genes in F1 IECs. The genes and their associated methylation or expression values are listed in Supplementary Data 9, Supplementary Data 3 and Supplementary Data 7, respectively. (b,c) Regional browser-view plots for Igf1r (b) and Nr4a2 (c) genes that are differentially methylated (DM) in F0 and F1 sperm samples and differentially expressed in F1 IECs. The plots depict the EnsEMBL gene annotations, the log transformed coverage for each quality-controlled site and the median methylation for F1Ctrl and F1DSS sperm samples. Red arrows indicate significant DM sites in Igf1r (b) and Nr4a2 gene (c). (d) Gene enrichment analysis of the query candidate genes (colored in blue) that remain connected with a relationship confidence of 0.75. Other interacting genes expressed in epithelial tissue are shown as black dots. The displayed network shows predicted functional relationships between the most functionally related genes and the query input genes, which are also analyzed for gene ontology enrichment (Supplementary Data 12). The relationship confidence is supported by several functional genomic and expression datasets. The edges between genes are colored (light red to dark red) by the confidence of the predicted relationship (relationship confidence).
Next, we compared our F1 sperm DNA methylation data with the 17 validated hypo-methylated regions and their associated genes published in the study from Radford et al.37, who report that in utero undernourished pups displayed a lower body weight and suffered from metabolic disorders in their later lives. Interestingly, reporting a similar phenotype of lower body weight herein, we also identified three consistently hypo-methylated genes, i.e. Asap2 (also significantly up-regulated in F1 IECs), Wif1 and Rnf149. However, the associated CpG sites of these hypo-methylated genes were covered at different locations (Supplementary Data 11).
Finally, we studied the herein-identified candidate genes and their interactions by a systematic functional network analysis (Fig. 5d). Their enriched KEGG (Kyoto Encyclopedia of Genes and Genomes) pathways and Gene ontology (GO) biological process categories are shown in Supplementary Data 12, which highlight pathways involved in notch signaling, melanoma, prostate cancer and transcriptional misregulations affecting several, diverse biological processes like brain development, skeletal muscle development, B cell homeostasis, fear/behavioral defense response, abnormalities with cardiac function, and cell type specific apoptotic process.
Discussion
To the best of our knowledge, this is the first study investigating environmentally-induced epigenetic changes in the course of DSS-induced colitis in mice. We report that offspring of males that had developed and recovered from chronic DSS colitis (F1DSS) displayed a lower body weight at baseline conditions compared to offspring of healthy males (F1Ctrl). F1DSS mice were also more susceptible to experimental colitis but did not exhibit spontaneous intestinal inflammation.
We performed RRBS and RNA-Seq to identify differentially methylated genes and its impact on gene expression. Global DNA methylome analysis of sperm cells revealed that the sperm epigenome was affected by prior DSS-induced colitis. Similarly, the methylome and transcriptome of IECs of F1DSS mice at baseline showed differentially methylated and expressed genes compared to F1Ctrl mice. It had already been reported that unlike any other cell type, massive epigenetic changes take place during spermatogenesis26. Environmental triggers may also disturb imprinting of genes that can alter the sperm epigenome, resulting in compromised gene expression in the offspring56. Epigenetic modifications may also shape the response and phenotype of offspring to variable environmental conditions57,58. While several of the identified epigenetic alterations may not affect the expression of the associated genes, they may still be transmitted by a ‘silent carrier’ to the next generations without affecting the gene expression and resulting in an asymptomatic phenotype59,60.
Notably, we observed moderate global methylation changes (20% cut-off) between IECs of F1Ctrl and F1DSS mice indicating that DSS might have small but significant effects on cytosine methylation in the next generation. In agreement with our results, Carone et al.30 reported similar modest effects on cytosine methylations in offspring of males fed a low-protein diet. Strikingly, they found significant alterations in offspring’s gene expression of many hepatic genes involved in lipid and cholesterol biosynthesis30.
DSS is a large molecular entity (MW 36–50 kDa) that upon oral ingestion disrupts the intestinal epithelial surface and thereby induces colitis61. Since DSS is not present at appreciable levels in the circulation while ingested orally, cell membranes are impermeable to intact molecules, and dextran molecules are not supposed to directly or indirectly introduce methyl groups to DNA, direct DSS effects to the genome appear unlikely, although we cannot formally exclude this possibility. Altogether it is fair to conclude that the biological phenotypes in the F1 generation and the changes in the F0 sperm methylome, which are – at least partially – also present in F1 sperm, may indeed be a consequence of intestinal inflammation.
Notably, almost half of the CpG sites in our data were annotated to regulatory regions of CTCF binding sites supporting the role of chromatin modifications in regulating disease susceptibility and the low weight phenotype. Specifically, CTCF is involved in regulating chromatin structure and plays an important role as a transcriptional repressor of Igf2 (insulin-like growth factor 2)62,63,64,65. Small size at birth and food deprivation due to prenatal exposure to famine has been also linked to hyper-methylation of sensitive loci including the imprinted locus Igf266. Similarly, we identified Igf1r (insulin-like growth factor 1 receptor) and Nr4a2/Nurr1 (nuclear receptor subfamily 4, group A, member 2) to be dysregulated in the intestinal epithelium of F1DSS mice. Interestingly, Igf1r down-regulation has previously been associated with intrauterine and postnatal growth retardation67, growth failure, mental retardation and fetal abnormalities68. In the herein reported study, Igf1r was found to be hypo-methylated and down-regulated in F0DSS sperm cells as well as in F1DSS IECs and F1DSS sperm cells, suggesting a possible link between this gene’s function with the herein identified metabolic phenotype in the F1 generation. The other possible epigenetically inherited candidate gene identified in this study, Nr4a2, was previously shown to be strongly up-regulated in adipose tissue in human obesity and is also linked to the regulation of body weight69,70. Furthermore, functional in vitro analyses revealed that Nr4a2 is involved in stress response as well as immune development and function69,70. Specifically, Nr4a2 was hyper-methylated and down-regulated in F1DSS IECs and F1DSS sperm cells, which may possibly represent a link between our metabolic phenotype, differential DNA methylation and consequently modulation of adipose tissue expansion.
Furthermore, our gene set enrichment analysis of differentially expressed genes in F1DSS mice collectively suggests a dysregulation of the immune response in these mice, which is in agreement with the increased susceptibility to inflammation detected in F1DSS mice.
Altogether, we report that epigenetic modifications acquired during DSS-induced colitis were transmitted to offspring through the paternal germ-line and led to alterations in the methylation pattern of CpG sites situated in genes involved in energy metabolism, which in turn may be associated with a lower body weight and increased susceptibility to colitis in offspring of diseased mice. Moreover, this study provides a list of candidate genes that may contribute to a better understanding of epigenetically modulated pathways during or after chronic intestinal inflammation. As such, our study adds to a growing body of literature29,38,39,40,42,43,44,45,46,47,48,49,50,51,52,53, which suggests that paternal lifestyle and pathological conditions (e.g. chronic inflammation) can affect spermatogenesis and can induce intergenerational transmission of epigenetic marks modulating offspring’s metabolism, but also contributing to risk for disease.
Material and Methods
Mice
Newborn male and female C57Bl/6(N) littermate mice were purchased from Charles River Laboratories. For all mice used in this study it was requested that n ≥ 6 mice of the same gender came from the same litter. Mice were maintained under specific pathogen free (SPF) conditions and were kept with a strict 12-hour light/dark cycle at the ZVTA (Medical University Innsbruck). All mice received the same diet and were fed according to a strict schedule. At 13 weeks of age male littermates were mated overnight with female littermates. After mating, males were single housed and were allowed to refill their sperm reservoir for 4 days, before they were sacrificed at 13.5 weeks of age. The Austrian Ministry of Science and Research has approved all mouse protocols and all experiments were performed in accordance with institutional guidelines.
Induction of chronic colitis by dextran sodium sulphate (DSS)
Chronic experimental colitis was induced in 5-week-old male C57Bl/6(N) littermates (F0DSS) by administration of three cycles of 2% dextran sodium sulphate (DSS Reagent grade, MW 36–50 kDa, MP Biomedicals, LLC) in their drinking water for 5 consecutive days, followed by a 14-day tap water period each. In the meantime normal drinking water was administered to the other half of the litter (F0Ctrl). During DSS treatment the disease activity index was determined daily by combining scores of weight loss, consistency of stool and rectal bleeding71. 9 days after the last DSS administration (day 55 of colitis model), F0DSS and F0Ctrl male littermates were mated overnight with littermate females.
Inflammation susceptibility test by induction of an acute DSS colitis
Acute colitis was induced in 7-week-old offspring of F0Ctrl and F0DSS mice by administration of 3.5% DSS in their drinking water for 6 consecutive days. During DSS treatment the disease activity index was determined daily by combining scores of weight loss, consistency of stool and rectal bleeding. After sacrifice, histological evaluations and length measurements were performed on extracted colons.
Histology
Formalin-fixed and paraffin-embedded intestines were sectioned and stained with hematoxylin & eosin (H&E) as described previously72. Baseline inflammation in the small intestine was assessed on H&E stained sections using a score, which considers four criteria: mononuclear cell infiltrate (0–3), crypt hyperplasia (0–3), epithelial injury (0–3) and polymorphonuclear cell infiltrates (1–3). As described previously72, the area involved was incorporated in the score by adding 1 for <10%, 2 for 10–25%, 3 for 25–50% and 4 for >50% involvement. Histological scoring of DSS treated mice was performed as described previously71.
Isolation of intestinal epithelial cells (IECs)
Mice were euthanized at 13 weeks of age and the middle part of the colon was used for the isolation of intestinal epithelial cells (IECs). The colon parts were washed with ice-cold PBS after cut open longitudinally. Mucus was removed by shaking the intestine in 1x HBSS containing 10% FCS, 1 mM DTT, 2 mM EDTA for 10 min at 37 °C. Then the DTT concentration was increased to 2 mM and shaken another 10 min at 37 °C. After moderate vortexing for 3 × 4 min in 1x HBSS containing 10% FCS and 1 mM EDTA, intestinal epithelial cells (IECs) were poured through 70 μm cell strainer (BD Bioscience), centrifuged at 1,500 rpm for 10 min and resuspended in PBS for subsequent FACS analysis.
Sperm cell isolation
Sperm cells from sacrificed mice were isolated from the caudal epididymis and Vas deferens as described previously30. Briefly, sperm was allowed to release from punctured and incised caudal epididymis and Vas deferens by incubation for 30 min at 37 °C in M2 medium (Sigma-Aldrich). Released sperm was collected and determined by microscopy for motility and purity. After RNA isolation qRT-PCR of epididymis-specific (Myh11, Actb) and sperm-specific genes (Smcp, Odf1) was performed to assess purity of sperm preparations.
Hematocrit measurement
Mice were anesthetized using a ketamine/xylazine mixture and blood samples were taken from the heart using a heparinized needle and syringe. Hematocrit levels in the whole blood were measured with a veterinary animal blood counter (Scil vet abc, HORIBA Medical).
RNA sequencing (RNA-Seq)
RNA libraries were prepared using the TruSeq RNA Sample Preparation Kit v2 (Illumina) according to manufacturer’s instructions. Libraries were analyzed with an Agilent 2100 Bioanalyzer using the Agilent DNA 1000 Kit. All samples were sequenced using an Illumina HiSeq 2000 platform (Illumina) at an average of 40 million paired-end 100 bp reads (Supplementary Data 13). Analytical details are provided in Supplementary Methods.
Reduced Representation Bisulfite Sequencing (RRBS)
RRBS libraries of genomic DNA isolated from sperm cells and FAC-sorted EpCAM+ CD45− colonic IECs were prepared as described by Boyle and colleagues73 and Smallwood and Kelsey74 using 5mC sequencing adapters (Illumina) and PE1.0 and PE2.0 primers (Illumina) for single-sample sequencing and TruSeq adapters and primers (Illumina) for the multiplexed sequencing approach. The final amplification was performed in accordance to the conditions described by Gu and colleagues75. All libraries were sequenced using a HiSeq 2500 platform (Illumina) at an average of 127 million single-end 50 bp reads (Supplementary Data 14). Additional details and analytical methods are provided in Supplementary Methods.
Statistical analysis of in vivo model data
Data are presented as mean ± standard error of the mean (SEM). Statistical analysis was performed using Microsoft Excel (Microsoft) and GraphPad Prism 6 (GraphPad Software). If not stated otherwise, statistical significance was calculated using a two-tailed Student’s t-test or a Mann-Whitney U test and P < 0.05 was considered significant. Where more than two groups were compared, One-way ANOVA with Bonferroni’s post-hoc testing or Kruskal-Wallis-tests were performed.
Additional Information
How to cite this article: Tschurtschenthaler, M. et al. Paternal chronic colitis causes epigenetic inheritance of susceptibility to colitis. Sci. Rep. 6, 31640; doi: 10.1038/srep31640 (2016).
Supplementary Material
Acknowledgments
We are indebted to Markus Schilhabel, Melanie Friskovec, Catharina v.d. Lancken, Melanie Schlapkohl, Ina Clefsen (ICMB sequencing platform) for technological support and sequencing of RRBS libraries. The authors thank Dr. Sieghart Sopper (Medical University Innsbruck) for assistance and advice with cell sorting (FACS). We also thank Prof. Christoph Bock (CeMM Research Center for Molecular Medicine of the Austrian Academy of Sciences, Vienna, Austria) for his useful suggestions and critically reading the manuscript. This study was carried out as part of the Research Training Group “Genes, Environment and Inflammation”, supported by the Deutsche Forschungsgemeinschaft (RTG 1743/1) of which A.F. is the spokesperson, the European Research Council under the European Community’s Seventh Framework Programme (FP7/2007–2013)/ERC Grant agreement no. 260961 (A.K.), the Austrian Science Fund and Ministry of Science P21530-B18 and START Y446-B18 (A.K.), the Wellcome Trust (investigator award 106260/Z/14/Z) to A.K., the Cambridge Biomedical Research Centre (A.K.), a fellowship from the European Crohn’s and Colitis Organisation (M.T. and T.E.A.) and a DOC fellowship from the Austrian Academy of Sciences (J.K.).
Footnotes
Author Contributions M.T., P.K., F.-A.H., S. Szymczak, A.K. and A.F. designed the project; M.T., P.K., F.-A.H., T.E.A., M.R. and J.K. performed the molecular analyses and experiments; F.A.O., O.A., F.K. and S. Smallwood contributed new reagents and analytic tools; and M.T., P.K., S. Szymczak, A.K. and A.F., with contributions from all authors, wrote the paper. All authors approved the final version of the manuscript.
References
- Loftus E. V. Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 126, 1504–1517 (2004). [DOI] [PubMed] [Google Scholar]
- Burisch J. et al. Environmental factors in a population-based inception cohort of inflammatory bowel disease patients in Europe–an ECCO-EpiCom study. J Crohns Colitis 8, 607–616, doi: 10.1016/j.crohns.2013.11.021 (2014). [DOI] [PubMed] [Google Scholar]
- Kaser A., Zeissig S. & Blumberg R. S. Inflammatory bowel disease. Annu Rev Immunol 28, 573–621, doi: 10.1146/annurev-immunol-030409-101225 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breslin N. P., Todd A., Kilgallen C. & O’Morain C. Monozygotic twins with Crohn’s disease and ulcerative colitis: a unique case report. Gut 41, 557–560 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halfvarson J., Bodin L., Tysk C., Lindberg E. & Jarnerot G. Inflammatory bowel disease in a Swedish twin cohort: a long-term follow-up of concordance and clinical characteristics. Gastroenterology 124, 1767–1773, doi: S0016508503003858 [pii] (2003). [DOI] [PubMed] [Google Scholar]
- Orholm M., Binder V., Sorensen T. I., Rasmussen L. P. & Kyvik K. O. Concordance of inflammatory bowel disease among Danish twins. Results of a nationwide study. Scand J Gastroenterol 35, 1075–1081 (2000). [DOI] [PubMed] [Google Scholar]
- Petersen B. S. et al. Whole genome and exome sequencing of monozygotic twins discordant for Crohn’s disease. BMC Genomics 15, 564, doi: 10.1186/1471-2164-15-564 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jostins L. et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 491, 119–124, doi: 10.1038/nature11582 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson C. A. et al. Meta-analysis identifies 29 additional ulcerative colitis risk loci, increasing the number of confirmed associations to 47. Nat Genet 43, 246–252, doi: 10.1038/ng.764 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franke A. et al. Genome-wide association study for ulcerative colitis identifies risk loci at 7q22 and 22q13 (IL17REL). Nat Genet 42, 292–294, doi: 10.1038/ng.553 (2010). [DOI] [PubMed] [Google Scholar]
- Strange A. et al. A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat Genet 42, 985–990, doi: 10.1038/ng.694 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom J. S., Ehrenreich I. M., Loo W. T., Lite T. L. & Kruglyak L. Finding the sources of missing heritability in a yeast cross. Nature 494, 234–237, doi: 10.1038/nature11867 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuk O., Hechter E., Sunyaev S. R. & Lander E. S. The mystery of missing heritability: Genetic interactions create phantom heritability. Proc Natl Acad Sci USA 109, 1193–1198, doi: 10.1073/pnas.1119675109 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo W., Chung W. Y., Qian M., Pellegrini M. & Zhang M. Q. Characterizing the strand-specific distribution of non-CpG methylation in human pluripotent cells. Nucleic Acids Res 42, 3009–3016, doi: 10.1093/nar/gkt1306 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramsahoye B. H. et al. Non-CpG methylation is prevalent in embryonic stem cells and may be mediated by DNA methyltransferase 3a. Proc Natl Acad Sci USA 97, 5237–5242 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziller M. J. et al. Genomic distribution and inter-sample variation of non-CpG methylation across human cell types. Plos Genet 7, e1002389, doi: 10.1371/journal.pgen.1002389 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lister R. et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 462, 315–322, doi: 10.1038/nature08514 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kominsky D. J., Campbell E. L. & Colgan S. P. Metabolic shifts in immunity and inflammation. J Immunol 184, 4062–4068, doi: 10.4049/jimmunol.0903002 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooke J. et al. Mucosal genome-wide methylation changes in inflammatory bowel disease. Inflamm Bowel Dis 18, 2128–2137, doi: 10.1002/ibd.22942 (2012). [DOI] [PubMed] [Google Scholar]
- Harris R. A. et al. DNA methylation-associated colonic mucosal immune and defense responses in treatment-naive pediatric ulcerative colitis. Epigenetics 9, 1131–1137, doi: 10.4161/epi.29446 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasler R. et al. A functional methylome map of ulcerative colitis. Genome Res 22, 2130–2137, doi: 10.1101/gr.138347.112 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koizumi K. et al. Array-based identification of common DNA methylation alterations in ulcerative colitis. Int J Oncol 40, 983–994, doi: 10.3892/ijo.2011.1283 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Z. et al. Identification of disease-associated DNA methylation in B cells from Crohn’s disease and ulcerative colitis patients. Dig Dis Sci 57, 3145–3153, doi: 10.1007/s10620-012-2288-z (2012). [DOI] [PubMed] [Google Scholar]
- Nimmo E. R. et al. Genome-wide methylation profiling in Crohn’s disease identifies altered epigenetic regulation of key host defense mechanisms including the Th17 pathway. Inflamm Bowel Dis 18, 889–899, doi: 10.1002/ibd.21912 (2012). [DOI] [PubMed] [Google Scholar]
- McDermott E. et al. DNA Methylation Profiling in Inflammatory Bowel Disease Provides New Insights into Disease Pathogenesis. J Crohns Colitis 10, 77–86, doi: 10.1093/ecco-jcc/jjv176 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambrot R. et al. Low paternal dietary folate alters the mouse sperm epigenome and is associated with negative pregnancy outcomes. Nat Commun 4, 2889, doi: 10.1038/ncomms3889 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane N. et al. Resistance of IAPs to methylation reprogramming may provide a mechanism for epigenetic inheritance in the mouse. Genesis 35, 88–93, doi: 10.1002/gene.10168 (2003). [DOI] [PubMed] [Google Scholar]
- Ng S. F. et al. Chronic high-fat diet in fathers programs beta-cell dysfunction in female rat offspring. Nature 467, 963–966, doi: 10.1038/nature09491 (2010). [DOI] [PubMed] [Google Scholar]
- Wei Y. et al. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc Natl Acad Sci USA 111, 1873–1878, doi: 10.1073/pnas.1321195111 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carone B. R. et al. Paternally induced transgenerational environmental reprogramming of metabolic gene expression in mammals. Cell 143, 1084–1096, doi: 10.1016/j.cell.2010.12.008 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daxinger L. & Whitelaw E. Understanding transgenerational epigenetic inheritance via the gametes in mammals. Nat Rev Genet 13, 153–162, doi: 10.1038/nrg3188 (2012). [DOI] [PubMed] [Google Scholar]
- Rando O. J. & Verstrepen K. J. Timescales of genetic and epigenetic inheritance. Cell 128, 655–668, doi: 10.1016/j.cell.2007.01.023 (2007). [DOI] [PubMed] [Google Scholar]
- Rehan V. K. et al. Perinatal nicotine exposure induces asthma in second generation offspring. BMC Med 10, 129, doi: 10.1186/1741-7015-10-129 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L. et al. Programming and inheritance of parental DNA methylomes in mammals. Cell 157, 979–991, doi: 10.1016/j.cell.2014.04.017 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackett J. A. et al. Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine. Science 339, 448–452, doi: 10.1126/science.1229277 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaati G., Bygren L. O. & Edvinsson S. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period. Eur J Hum Genet 10, 682–688, doi: 10.1038/sj.ejhg.5200859 (2002). [DOI] [PubMed] [Google Scholar]
- Radford E. J. et al. In utero effects. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 345, 1255903, doi: 10.1126/science.1255903 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fullston T. et al. Paternal obesity initiates metabolic disturbances in two generations of mice with incomplete penetrance to the F2 generation and alters the transcriptional profile of testis and sperm microRNA content. FASEB J 27, 4226–4243, doi: 10.1096/fj.12-224048 (2013). [DOI] [PubMed] [Google Scholar]
- Martinez D. et al. In utero undernutrition in male mice programs liver lipid metabolism in the second-generation offspring involving altered Lxra DNA methylation. Cell Metab 19, 941–951, doi: 10.1016/j.cmet.2014.03.026 (2014). [DOI] [PubMed] [Google Scholar]
- Ost A. et al. Paternal diet defines offspring chromatin state and intergenerational obesity. Cell 159, 1352–1364, doi: 10.1016/j.cell.2014.11.005 (2014). [DOI] [PubMed] [Google Scholar]
- Huypens P. et al. Epigenetic germline inheritance of diet-induced obesity and insulin resistance. Nat Genet 48, 497–499, doi: 10.1038/ng.3527 (2016). [DOI] [PubMed] [Google Scholar]
- Linn T., Loewk E., Schneider K. & Federlin K. Spontaneous glucose intolerance in the progeny of low dose streptozotocin-induced diabetic mice. Diabetologia 36, 1245–1251 (1993). [DOI] [PubMed] [Google Scholar]
- Anderson L. M. et al. Preconceptional fasting of fathers alters serum glucose in offspring of mice. Nutrition 22, 327–331, doi: 10.1016/j.nut.2005.09.006 (2006). [DOI] [PubMed] [Google Scholar]
- Zeybel M. et al. Multigenerational epigenetic adaptation of the hepatic wound-healing response. Nat Med 18, 1369–1377, doi: 10.1038/nm.2893 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias B. G. & Ressler K. J. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nat Neurosci 17, 89–96, doi: 10.1038/nn.3594 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietz D. M. et al. Paternal transmission of stress-induced pathologies. Biol Psychiatry 70, 408–414, doi: 10.1016/j.biopsych.2011.05.005 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gapp K. et al. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat Neurosci 17, 667–669, doi: 10.1038/nn.3695 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gapp K. et al. Early life stress in fathers improves behavioural flexibility in their offspring. Nat Commun 5, 5466, doi: 10.1038/ncomms6466 (2014). [DOI] [PubMed] [Google Scholar]
- Morgan C. P. & Bale T. L. Early prenatal stress epigenetically programs dysmasculinization in second-generation offspring via the paternal lineage. J Neurosci 31, 11748–11755, doi: 10.1523/JNEUROSCI.1887-11.2011 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodgers A. B., Morgan C. P., Bronson S. L., Revello S. & Bale T. L. Paternal stress exposure alters sperm microRNA content and reprograms offspring HPA stress axis regulation. J Neurosci 33, 9003–9012, doi: 10.1523/JNEUROSCI.0914-13.2013 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerrero-Bosagna C., Settles M., Lucker B. & Skinner M. K. Epigenetic transgenerational actions of vinclozolin on promoter regions of the sperm epigenome. Plos One 5, doi: 10.1371/journal.pone.0013100 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skinner M. K. Environmental stress and epigenetic transgenerational inheritance. BMC Med 12, 153, doi: 10.1186/s12916-014-0153-y (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vassoler F. M., White S. L., Schmidt H. D., Sadri-Vakili G. & Pierce R. C. Epigenetic inheritance of a cocaine-resistance phenotype. Nat Neurosci 16, 42–47, doi: 10.1038/nn.3280 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ni J., Chen S. F. & Hollander D. Effects of dextran sulphate sodium on intestinal epithelial cells and intestinal lymphocytes. Gut 39, 234–241 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falls J. G., Pulford D. J., Wylie A. A. & Jirtle R. L. Genomic imprinting: implications for human disease. Am J Pathol 154, 635–647, doi: 10.1016/S0002-9440(10)65309-6 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li E., Beard C. & Jaenisch R. Role for DNA methylation in genomic imprinting. Nature 366, 362–365, doi: 10.1038/366362a0 (1993). [DOI] [PubMed] [Google Scholar]
- Skinner M. K., Anway M. D., Savenkova M. I., Gore A. C. & Crews D. Transgenerational epigenetic programming of the brain transcriptome and anxiety behavior. Plos One 3, e3745, doi: 10.1371/journal.pone.0003745 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews D. et al. Transgenerational epigenetic imprints on mate preference. Proc Natl Acad Sci USA 104, 5942–5946, doi: 10.1073/pnas.0610410104 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franklin T. B., Linder N., Russig H., Thony B. & Mansuy I. M. Influence of early stress on social abilities and serotonergic functions across generations in mice. Plos One 6, e21842, doi: 10.1371/journal.pone.0021842 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saavedra-Rodriguez L. & Feig L. A. Chronic social instability induces anxiety and defective social interactions across generations. Biol Psychiatry 73, 44–53, doi: 10.1016/j.biopsych.2012.06.035 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson M. E. et al. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. Plos One 5, e12238, doi: 10.1371/journal.pone.0012238 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohlsson R., Renkawitz R. & Lobanenkov V. CTCF is a uniquely versatile transcription regulator linked to epigenetics and disease. Trends Genet 17, 520–527 (2001). [DOI] [PubMed] [Google Scholar]
- Dunn K. L. & Davie J. R. The many roles of the transcriptional regulator CTCF. Biochem Cell Biol 81, 161–167, doi: 10.1139/o03-052 (2003). [DOI] [PubMed] [Google Scholar]
- Yang B. et al. A novel pathway links oxidative stress to loss of insulin growth factor-2 (IGF2) imprinting through NF-kappaB activation. Plos One 9, e88052, doi: 10.1371/journal.pone.0088052 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heijmans B. T. et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci USA 105, 17046–17049, doi: 10.1073/pnas.0806560105 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tobi E. W. et al. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum Mol Genet 18, 4046–4053, doi: 10.1093/hmg/ddp353 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walenkamp M. J., Losekoot M. & Wit J. M. Molecular IGF-1 and IGF-1 receptor defects: from genetics to clinical management. Endocr Dev 24, 128–137, doi: 10.1159/000342841 (2013). [DOI] [PubMed] [Google Scholar]
- Wit J. M. & Walenkamp M. J. Role of insulin-like growth factors in growth, development and feeding. World Rev Nutr Diet 106, 60–65, doi: 10.1159/000342546 (2013). [DOI] [PubMed] [Google Scholar]
- Veum V. L. et al. The nuclear receptors NUR77, NURR1 and NOR1 in obesity and during fat loss. Int J Obes (Lond) 36, 1195–1202, doi: 10.1038/ijo.2011.240 (2012). [DOI] [PubMed] [Google Scholar]
- Perez-Sieira S., Lopez M., Nogueiras R. & Tovar S. Regulation of NR4A by nutritional status, gender, postnatal development and hormonal deficiency. Sci Rep 4, 4264, doi: 10.1038/srep04264 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrett W. S. et al. Communicable ulcerative colitis induced by T-bet deficiency in the innate immune system. Cell 131, 33–45, doi: 10.1016/j.cell.2007.08.017 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaser A. et al. XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 134, 743–756, doi: 10.1016/j.cell.2008.07.021 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyle P. et al. Gel-free multiplexed reduced representation bisulfite sequencing for large-scale DNA methylation profiling. Genome Biol 13, R92, doi: 10.1186/gb-2012-13-10-r92 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smallwood S. A. & Kelsey G. Genome-wide analysis of DNA methylation in low cell numbers by reduced representation bisulfite sequencing. Methods Mol Biol 925, 187–197, doi: 10.1007/978-1-62703-011-3_12 (2012). [DOI] [PubMed] [Google Scholar]
- Gu H. et al. Preparation of reduced representation bisulfite sequencing libraries for genome-scale DNA methylation profiling. Nat Protoc 6, 468–481, doi: 10.1038/nprot.2010.190 (2011). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.