

ABSTRACT
Obesity is a serious health problem worldwide since it is a major risk factor for chronic diseases such as type II diabetes. Obesity is the result of hyperplasia (associated with increased adipogenesis) and hypertrophy (associated with decreased adipogenesis) of the adipose tissue. Therefore, understanding the molecular mechanisms underlying the process of adipocyte differentiation is relevant to delineate new therapeutic strategies for treatment of obesity. As in all differentiation processes, temporal patterns of transcription are exquisitely controlled, allowing the acquisition and maintenance of the adipocyte phenotype. The genome is spatially organized; therefore decoding local features of the chromatin language alone does not suffice to understand how cell type-specific gene expression patterns are generated. Elucidating how nuclear architecture is built during the process of adipogenesis is thus an indispensable step to gain insight in how gene expression is regulated to achieve the adipocyte phenotype. Here we will summarize the recent advances in our understanding of the organization of nuclear architecture as progenitor cells differentiate in adipocytes, and the questions that still remained to be answered.
keywords: adipogenesis, chromosome territory, nuclear lamina, nucleoskeleton
The pandemic of obesity has led scientists to focus their attention on the study of adipose tissue and the development of fat cells. During the last 2 decades it has been demonstrated that adipocytes release a variety of factors, including cytokines, chemokines, and many other biologically active molecules, generically called adipokines, that have led adipose tissue to be regarded as an active endocrine organ. Adipokines signal to organs of metabolic importance including brain, liver, skeletal muscle, and the immune system.1,2 In this way, the adipose tissue plays a central role in modulating the homeostasis of lipid and glucose metabolism, blood pressure, and inflammation. In obese individuals, the secretion of adipokines is deregulated3 and adipose tissue is infiltrated by a higher number of macrophages compared to normal tissue.4,5 These events lead to a state of chronic inflammation and metabolic syndrome.6,7 Conversely, lipodystrophy, a disorder characterized by selective total or partial loss of body fat, is also accompanied by metabolic consequences similar to those observed in obesity, including insulin resistance, dyslipidemia, hepatic and myocellular steatosis and increased risk of diabetes and atherosclerosis,8 reinforcing the notion that adipose tissue plays a key role in the control of the homeostasis of whole-body metabolism.
In mammals, adipocytes have been classified into 2 distinct types: white, and brown adipose cells. White adipocytes express cell type-selective machinery for triglyceride synthesis from lipoprotein-derived fatty acids, as well as for hormone-stimulated glucose uptake and lipolysis. In addition, they produce adipokines such as leptin, resistin, adiponectin, and TNFα that modulate systemic metabolism, a function shared with brown fat cells.9,10 The development of methods for separating white adipose tissue (WAT) into adipocyte and stromal-vascular fractions (SVF),11 led to the discovery that SVF is the source of adipocyte precursor cells.12 However, the characterization of this cell population remained elusive for several decades and much effort has been done to uncover it. Sengenes et al showed by FACS analysis performed on crude SVF cultured under adipogenic conditions, that the progressive accumulation of lipid droplets was associated with a selective enrichment of the CD34+/CD31- cell population.13 Rodeheffer et al. identified a subpopulation of early adipocyte progenitor cells resident in the WAT vascular stroma as Lin−:CD29+:CD34+: Sca-1+:CD24+ that constitutes in total approximately 0.08% of the cells in the SVF.14 Using genetically tagged mice, Tang et al. demonstrated that most adipocytes derive from a pool of proliferating and renewing adipogenic progenitors that are already committed either prenatally or early in postnatal life.15 These progenitors reside in the mural cell compartment of the adipose vasculature, but not in the vasculature of other tissues,15 an observation supported by early studies using electron microscopy.16,17 The local microenvironment is a crucial determinant of progenitor fate, function, and maintenance.18 Thus, the adipose vasculature appears to function as a progenitor niche, possibly providing signals for adipocyte development.
In contrast to WAT, the main role of BAT is to metabolize fatty acids to maintain body temperature, by dissipating chemical energy in the form of heat. This function depends on its high mitochondrial content and the ability to uncouple cellular respiration through the action of uncoupling protein-1 (UCP-1).6,10,19 There is evidence that brown adipocytes originate from a cell lineage distinct from that of white adipose cells.20-23 Microarray analysis of primary brown adipocytes showed the expression of a myogenic transcriptional signature.20 Further, myogenic factor 5 (Myf5), previously thought to be expressed only in skeletal muscle precursors, is also expressed in BAT committed precursors.22 Myf5 positive myoblast-like cells differentiate into brown adipocytes through the action of the transcriptional regulators PRMD16 and C/EBPβ.21,22,24 BAT was considered insignificant in adult humans; however recent studies have shown that brown adipocytes are dispersed throughout adult adipose tissue.25-27 The amount of BAT is inversely correlated with body-mass index, especially in older people. BAT depots respond to changes in temperature, but this response is significantly lower in the overweight or obese than in lean subjects suggesting a potential role of BAT in adult human metabolism.28-30 Mitochondrial function, which is key in non-shivering thermogenesis in brown adipose cells, is also diminished in white adipocytes at the onset of obesity,31 exemplifying a negative relationship between obesity and mitochondrial biogenesis that potentially contributes to the development of diabetes.32,33 Therefore, the balance between BAT and WAT affects energy balance, and any factor that may alter the equilibrium between them may possibly facilitate the development of obesity and loss of metabolic homeostasis.
However, adipose tissue is even more complex than previously thought. It was recently shown that in addition to classical white and brown adipocytes, there is at least a third cell type: beige or brite (brown-in-white) cells.34-37 They are present in patches dispersed in WAT, and do not derive from the same lineage as the classical brown fat cells. The existence of a progenitor cell that differentiates into beige cells is still controversial. It has been shown that chronic treatment of precursors of WAT with PPARγ ligands leads to the generation of beige cells that express PGC-1α and respond to norepinephrine by increasing UCP-1 gene expression.35 This effect of PPAR agonists is possibly due to an increase in the half-life of the PRDM16 protein.38 However, beige adipose cells are devoid of expression of myocyte-associated genes and transcription factors present in classical brown adipocytes.35 It has also been proposed that browning of WAT upon exposure of mice to cold is a consequence of transdifferentiation of white into beige adipocytes, since neither adipocyte number nor DNA content increases in white fat depots.36 It was proposed that BAT and inducible beige fat cells participate in non-shivering thermogenesis, an event crucial for mammals as a defense against cold and obesity, under the control of the sympathetic nervous system and β-adrenergic signaling. Interestingly, it has been recently reported that white and beige fat cells have the ability to respond to low temperatures by activating a thermogenic gene program in a cell-autonomous manner.39 This activation in isolated cells is independent of the cAMP/PKA/CREB pathway downstream of the β-adrenergic receptors.39 Bone morphogenic proteins (BMPs) regulate both the formation and the thermogenic activity of BAT.40,41 Disruption of BMPs signaling in BAT causes a severe paucity of BAT and a compensatory sympathetic response in WAT by increasing the number of beige cells.42 This compensatory mechanism is sufficient to maintain the homeostasis of body temperature and to generate resistance to diet-induced obesity.42 Importantly, adipose cells with the molecular signature of beige cells are also present in fat depots in humans.43,44 Thus beige cells may represent a potential target for therapeutic intervention for the treatment of obesity and metabolic syndrome. Unequivocally, adipose tissue is very complex and heterogeneous; therefore its deregulation can deeply affect whole-body metabolic homeostasis.
Adipose tissue is not static and approximately 10% of our body's fat cells are regenerated each year.45 In order to support adipose dynamics in the adult, proliferative adipocyte precursor cells must exist and be poised to respond to metabolic demands. During adipogenesis, as in any differentiation process, temporal patterns of transcription are exquisitely controlled upon activation of different signaling pathways that lead to the binding of transcription factors to target genes as well as factors that coordinately modify the chromatin landscape as adipogenesis proceeds, allowing the acquisition and maintenance of the adipocyte phenotype.10,46,47 All cells of an individual possess the same genomic information; yet we do not know how the precise pattern of gene expression is established and maintained in order to delineate the final phenotype of a cell. During the last decades a great deal of effort has been focused on deciphering the histone code to elucidate how the architecture of nuclear chromatin is organized.48,49 Covalent histone modifications and their interplay with DNA methylation sites, as well as histone variants and chromatin remodeling events, have emerged as major players, yielding distinct modifications of chromatin which allow or prevent transcription.48,50 Control of epigenetic signatures is not only involved in a cell differentiation process such as adipogenesis,51 but also in re- or de-programming during development.52,53 The in vitro reprogramming of somatic cells to pluripotent stem cells (SC) is also accompanied by genome-wide remodeling of chromatin modifications from a somatic to an SC-like state,54 although the temporal sequence of these changes remains elusive. Nevertheless, understanding transcriptional regulation and decoding the epigenetic code also are not enough to understand how cell type-specific gene expression patterns are generated throughout development or during any process of cell differentiation in an adult organism. In this regard, knowledge of the architecture of nuclear compartments is also critical to understand how gene expression as well as other nuclear functions, such as chromatin replication and DNA repair are regulated.55
One of the current pressing questions is how gene expression is integrated into the architectural framework of the cell nucleus, delineating the goal of “epigenomics”: the functional integration of epigenetics and nuclear architecture.55 In the path toward epigenomics, we must become familiar with the complex and dynamic architecture of the nucleus in interphase. The nucleus is highly compartmentalized, but in contrast to compartments in the cytoplasm, nuclear compartments are not limited by membranes. This characteristic favors their highly dynamic assembly-disassembly status, dependent on the cell cycle phase and the transcriptional state of the cell. In the interphase nucleus, the nuclear compartments include the nuclear lamina, the still controversial nucleoskeleton or nuclear matrix, the chromosome territories and nuclear bodies (Fig. 1A). Much effort is being directed toward understanding how gene expression is integrated into the landscape of nuclear architecture. Here we review the first steps taken to dissect the complex organization of the nucleus during the process of adipocyte differentiation.
Figure 1.
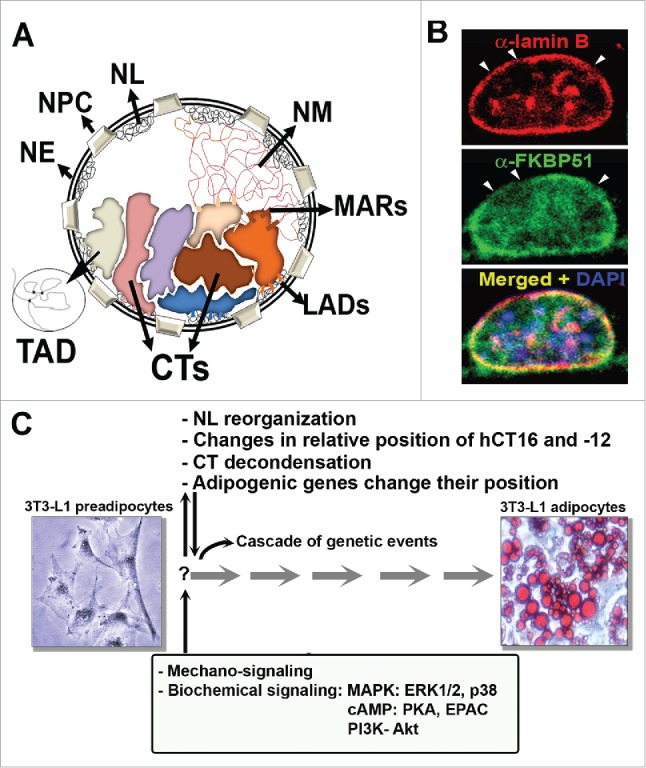
Nuclear architecture during adipogenesis. (A) Schematic representation of the compartments of the nucleus in interphase: NE: nuclear envelope, NPC: nuclear pore complex, NL: nuclear lamina, NM: nuclear matrix, CTs: chromosome territories, LADs: lamin attachment domains, MARs: Matrix attachment regions; TADs: topologically associating domains. (B) Reorganization of the NL during adipocyte differentiation. 3T3L1 preadipocytes grown on coverslips were induced to differentiate for 24h. Indirect immunofluorescence and confocal microscopy imaging shows lamin B (red), FKBP51 (green) and chromatin stained with DAPI (blue), as described.98 Observe the discontinuous staining of lamin B (arrow heads) due to the reorganization of the NL. (C) Summary of the events of nuclear reorganization that were described during adipogenesis. Images depict 3T3-L1 preadipocytes and adipocytes, the latter with lipid vesicles stained with Oil Red O, as described.98
Nuclear lamina and its importance in the adipocyte
The nuclear lamina (NL) is a filamentous protein mesh-work that lines the nucleoplasmic surface of the nuclear envelope (NE) interacting with inner nuclear membrane proteins and nuclear pores (Fig. 1A),56,57 reviewed in.58 The NL consists of a polymeric assembly of nuclear lamins, the A-type (LA and LC) and the B-type lamins (LB1 and LB2), respectively.59 LA and LC are derived from a single gene (LMNA) by alternative splicing and are expressed only in differentiated cells. On the other hand, LB1 and LB2 are encoded by LMNB1 and LMNB2, respectively, and at least one of them is expressed in all cells throughout development.60 The lamins bind to other NE proteins, including some belonging to the nuclear pore complex (NPC) and the inner nuclear membrane, interactions that are functionally important in regulating the proper assembly of the NE.61 Although the major fraction of lamins is found in the NL, they are also located throughout the nucleoplasm.62,63 The NL is thought to provide a structural framework for the NE, contributing to the size, shape and mechanical stability of the nucleus. However, lamins participate not only in the architectural organization of the nucleus, but also in its cellular integrity, possibly through the Linker of Nucleus and Cytoskeleton (LINC) complex.64 LINC is formed by lamins that interact with the inner nuclear membrane-bound SUN proteins, which in turn associate with the nuclear-cytoskeleton linker proteins in the outer nuclear membrane, i.e. nesprins. Nesprin-1 and −2 contain a binding domain for direct interaction with actin, and nesprin-3 for plectrin, an intermediate filament binding protein. The NL through LINC constitute an interconnected network that is involved in the control of nuclear positioning, mechano-signal transduction, mitosis as well as in the organization of the cytoskeleton.65 In addition, the NL also participates in DNA replication and repair, RNA polymerase II transcription, and the epigenetic control of chromatin remodeling.63 The NL provides anchoring sites for interphase chromosomes at the nuclear periphery. In this regard, a high resolution map of the interaction sites of the entire genome with the NL components in human fibroblasts shows that genome-lamina interactions occur through more than 1,300 sharply defined large domains 0.1-10 megabases in size, known as lamin associated domains (LADs).66 These LADs are characterized by low gene-expression levels, indicating that they represent a repressive chromatin environment.66 Using 3-dimensional DNA-immunoFISH, it was demonstrated that repositioning of chromosomal regions to the nuclear lamina is dependent on breakdown and reformation of the NE during mitosis.67
The NL was long considered to be a nuclear domain that was not permissive for transcription, since genes positioned at the NL and even tethered to it were silenced.67,68 However, more recent studies have shown that some transcriptionally active genes are associated with the NL via nucleoporins at the nucleoplasmic face of the NPCs and/or are relocated upon transcriptional activation to nuclear pores.69-72 H3K27Me3, which marks repressed gene promoters, is enriched at the nuclear periphery in embryonic stem cells (ESC).73 In contrast, H3K27Me3 was less concentrated at the nuclear periphery of differentiated cells, demonstrating that the nuclear periphery is an epigenetically dynamic compartment that might possess distinct epigenetic marks in pluripotent ESCs compared to differentiated cells.73 Furthermore, it was recently shown that FKBP52, a component of the Hsp90•Hsp70•p23 heterocomplex bound to steroid receptors,74 concentrates in a perinuclear structure associated with Hsp90 and p23 in undifferentiated murine neuroblastoma cells (N2a) and embryonic hippocampal neurons.75 Upon cell differentiation, this structure disassembles and the perinuclear area becomes transcriptionally active.75 Therefore, in the nuclear periphery different subdomains can be dynamically established with epigenetic marks as well as signaling molecules; some are repressive and enriched in facultative heterochromatin, while others are permissive for transcription to occur when the proper signal is received by the cell.
The importance of NL is highlighted by the existence of a group of pathologies known as laminopathies that affect specific tissues and are caused either by mutations in the lamin A/C (LMNA) gene or by mutations in the FACE-1 gene affecting the correct post-translational processing of prelamin A (reviewed in76). It has been proposed that mutations that affect lamins might disrupt their binding to yet unidentified tissue-specific partner proteins to generate pathology in a particular tissue (reviewed in65). Laminopathies affecting the adipose tissue are characterized by lipodystrophies with selective and variable loss of adipose tissue accompanied by metabolic complications including insulin resistance, type 2 diabetes, hypertriglyceridemia, and liver steatosis. These laminopathies include Dunningan-type familial partial lipodystrophy (DFPLD), partial lipodystrophy with mandibuloacral dysplasia (MAD), both associated with mutations in LMNA gene; Berardinelli-Seip congenital generalized lipodystrophy (CGL); and some cases with Barraquer-Simons acquired partial lipodystrophy (APL) associated with mutations in lamin B2.77 Lipodystrophy can also be acquired, for example, in patients under treatment for human immunodeficiency virus.78 Most of the DFDLD mutations in LMNA are missense within the 3´end of the gene, downstream of the nuclear localization signal, and it has been proposed that these mutations may alter the interactions of lamin A with transcription factors or other DNA binding molecules.79
The accumulation of the lamin A precursor has also been described in lipodystrophic cells.80 The transcription factor sterol regulatory element binding protein 1 (SRBP1) that is present in adipocytes interacts with pre-lamin A in fibroblasts obtained from lipodystrophic patients, as well as in fibroblasts from normal subjects forced to accumulate pre-lamin A by treatment with farnesylation inhibitors.80 It was proposed that the lack of proper lamina maturation determines the sequestration of SRBP1 at the nuclear rim, thus decreasing the pool of active SRBP1 that normally activates PPARγ, a key regulator of the acquisition and maintenance of the adipose phenotype,81,82 thus impairing preadipocyte differentiation.80 It is noteworthy that mutations in PPARγ are also responsible for other forms of partial lipodystrophy.83-85 Retention of SREBP-1 at the NL and reduced PPARγ expression were also observed in 3T3-L1 preadipocytes treated with some of the HIV protease inhibitors (i.e., Indinavir) employed in treatment of viral infection, drugs that have been reported to cause lipodystrophy as a side-effect.86 Therefore, SREBP-1 retention at the nuclear rim of lipodystrophic cells can be associated with the presence of increased levels of pre-lamin A irrespective of the occurrence of LMNA mutations, findings that help to explain the pathophysiology of both inherited and acquired lipodystrophies. Furthermore, overexpression of both wild-type or of mutated lamin A-R482A (present in DFPLD) in 3T3- L1 preadipocytes inhibits lipid accumulation, triglyceride synthesis and expression of adipogenic markers, such as PPARγ2.87 In contrast, embryonic fibroblasts lacking A-type lamins accumulated more intracellular lipids, exhibited elevated de novo triglyceride synthesis, and increased basal activation of Akt1, a well known mediator of insulin signaling.87 Therefore, A-type lamins may act as inhibitors of adipocyte differentiation when their expression level is inappropriate, consistent with the proposal that mutations that cause lypodystrophy may reflect a “gain of function” in these proteins, resulting in higher binding affinity to pro-adipogenic transcription factors such as SREBP-1, that are sequestered at the NL and are thus prevented from activating their target gene(s).
Ultrastructural studies suggest the presence of a dispersed lamin network throughout the nucleus.88,89 The existence of a dispersed, veil-like nucleoplasmic lamin network was also shown using GFP-tagged lamins and bleaching techniques, which showed that a considerable fraction of intranuclear lamins, visible as diffuse nucleoplasmic fluorescence, is stably integrated in the nuclear interior.62,90 The exact molecular structure of this lamin veil is unknown; it possibly offers the spatial possibility for lamins to interact with chromatin or to bind to nuclear histone proteins as shown in vitro.91,92 Therefore, although the role of this fine lamin network in cellular processes is so far unclear, these findings suggest they provide a scaffold for transcription and DNA replication. Interestingly, cell lines transfected with GFP-lamins with mutations similar to those found in Dunnigan's type lipodystrophy patients showed that these mutated lamins do not incorporate properly into the nucleoplasmic veil,93 and in this manner possibly contribute to the abnormalities found in this type of lipodystrophy.
The nuclear lamina is reorganized at the onset of adipogenesis
Analysis of the expression level of lamin A and the NE transmembrane protein emerin at the onset of differentiation of 3T3F442A preadipocytes showed that, while lamin A expression progressively decreases, emerin expression increases.94 Emerin participates in the control of β-catenin95 whose sustained activation inhibits the process of adipogenesis.96 Increased expression of emerin controls the efficient redistribution of β-catenin from the nucleus to the cytoplasm, facilitating its proteasomal degradation and consequently allowing the process of adipocyte differentiation to proceed.94 Interestingly, it was demonstrated that the NL is fragmented at the early stages of adipogenesis97,98 (Fig. 1B), event that is accompanied by the loss of not only lamin A, but also C, B1, and emerin at the nuclear rim.97 Later on, upon maturation of the adipose cell (day 18 post-induction of adipogenesis) lamins A, C and B1 increase at the nuclear rim independently of their low level of protein expression.97 In contrast, lamin B2 remains at the nuclear rim throughout the process of adipogenesis.97
Since the NL participates in the control of many aspects of nuclear events as already described, it was proposed that the decreased presence of most lamin subtypes at the nuclear rim and the fragmentation of the NL results in enhanced plasticity of the nucleus.97 In support of this, we have recently reported that the high molecular weight immunophilin (IMM) FKBP51, a member of the glucocorticoid receptor (GR)•Hsp90•Hsp70•p23 heterocomplex required for proper signaling of steroid nuclear receptors,74 translocates from mitochondria to the nucleus at the onset of adipogenesis.98,99 In the nucleus, FKBP51 not only co-localizes with lamin B in the fragmented pattern of the lamina (Fig. 1B), but also interacts with lamin B.98 The transient relocalization of FKBP51 from mitochondria to the nucleus depends on an increase in intracellular cAMP and PKA activation.98 Interestingly, PKA-cα also translocates to the nucleus, and concentrates in the NL possibly through its interaction with FKBP51.98 Several phosphorylation sites, including those for the cyclin B1-(CCNB1)-CDC2 complex, PKC, and PKA are important in nuclear lamina disassembly.100,101 Therefore, we propose that enrichment of PKA-cα in the NL may facilitate its reorganization through phosphorylation of lamins during the process of adipogenesis. We hypothesize that the accumulation of PKA-cα in the NL may be also involved in the control of gene expression at the onset of adipogenesis possibly by regulating the phosphorylation of transcription factors enriched in this nuclear compartment, similar to a mechanism shown for the control of AP-1 transcriptional activity upon the sequestration of c-fos in the NL in an ERK1/2 dependent manner.102 Nuclear dynamics during adipogenesis can be analyzed in 3T3-L1 preadipocytes treated with dexamethasone, isobutyl-methylxantine (IBMX) and insulin to induce the adipogenic program.103 Notably, co-treatment of the preadipocytes with dexamethasone and IBMX, which increases cAMP levels and activates PKA, also increases the presence of GR in the nuclear periphery, where it exhibits a high level of co-localization with FKBP51. This observation raises the possibility that, dependent on cAMP-signaling, the nuclear bioavailability of GR may be regulated by FKBP51 which retains GR in the NL at the early stages of adipogenesis; in this manner, the IMM may participate in regulating the expression of GR target genes.98 When the adipocyte phenotype is achieved, and possibly upon maturation of adipocytes, the fraction of cells expressing lamins A, C and B1 at the nuclear rim increases, though overall lamin A/C protein levels remain low,97 suggesting a mechanism that concentrates lamins in the nuclear periphery. Uncovering the functional importance of such parallel changes in regulation of nuclear architecture will provide insight into molecular mechanisms that control adipogenesis as well as the pathophysiology of adipose tissue.
The organization and dynamics of the NL is also a field of intensive research in the context of its interconnections with the NE, the cytoskeleton, and mechano-signal transduction. Mechanical stress exerted at the outer cell surface causes changes in nuclear shape, possibly through the LINC complex.104 In this way, interactions between the plasma membrane and the cytoskeleton may regulate gene expression in response to mechanical stress initiated at the cell surface, with the advantage that mechano-transduction propagation is faster than chemically based diffusion.105,106 There is evidence that some NE proteins are involved in gene regulation in response to mechanical stress. In this regard, emerin knockout mouse embryonic fibroblasts subjected to mechanical stress are defective in expressing mechano-sensitive genes.107 However, disruption of the LINC complex does not always deregulate mechano-sensitive genes in response to mechanical stress,108 raising the possibility that other signaling pathways may act synergistically with mechano-transduction. Tissues can be classified as being soft and stiff based on their capacity for bearing low or high levels of mechanical stress, respectively. Fat tissue, like brain and bone marrow, belongs to the group of soft tissues. In line with this classification, stem cells cultured on matrices of different elasticity have different capacities to differentiate into various cell types, as demonstrated for muscle stem cells.109 This suggests that differentiation is mechanosensitive. It has been recently shown that matrix stiffness directly influences lamin A protein levels.110 Bone marrow-derived mesenchymal stem cells (MSCs) have a high lamin A: lamin B ratio that possibly reflects their osteogenic niche origin.114 When MSC are grown on a soft matrix, the percentage of cells that differentiate in adipose cells is higher than the percentage obtained when MSCs are grown on a stiff matrix (8% versus 1%, respectively).114 Interestingly, knockdown of lamin A in MSCs greatly favored their differentiation in adipose cells (∼20%) on a soft matrix.110 These observations are in line with the fact that embryonic fibroblasts lacking A-type lamins accumulate more intracellular lipids,87 and that during the process of 3T3-L1 preadipocyte differentiation there is a decrease in lamin A/C and B1.97 Overall, tissue stiffness and mechanical stress-dependent changes in lamin A levels, accompanied by differential expression of NE transmembrane proteins,111 may contribute to the architectural organization of the nucleus that is ultimately required for the control of cell fate choice. As we learn more of the dynamic reorganization of the NL and the cytoskeleton during adipogenesis as a model of cell differentiation, an integrated and better understanding of both the mechano-dependent and -independent mechanisms of control of gene expression will be achieved.
Does the nuclear matrix play a role in adipogenesis?
The nuclear matrix (NM) (Fig. 1A) is defined as the non-chromatin structure of the nucleus and is readily observed in extracted cells under the electron microscope,112 although its existence is still a topic under discussion. The main feature of this non-chromatin structure is its fibrogranular ribonucleoprotein (RNP) network.113 NM preparations usually retain approximately 70% of the total nuclear RNA114 and RNAse treatment destroys this nuclear structure.89,115,116 The internal nuclear matrix is connected to the lamina and fills the nuclear interior.117 Chromatin forms loops that were first inferred from measurements of nuclear sedimentation in ethidium bromide118 and confirmed by their direct microscopic visualization after the stripping of histones.119 Chromatin loops that range in size from 30 to 110 kb in somatic cells120,121 and from 20 to 50 kb in sperm,119,122 are formed by attachments of chromatin to the NM. Chromatin interacts with the NM through DNA sequences called matrix-attachment regions (MARs) or scaffold-attachment regions (SARs) that have 150-200 bp subregions that can become single stranded under torsional stress.123 Studies performed in Drosophila have shown that MARs are interspersed in the genome at intervals of 26,000-112,000 kb,124 consistent with the estimated sizes of chromatin loops in flies and mammals.120,125 MARs bind to specific NM and NL proteins, and one of the best characterized MAR-binding protein is SATB1, which is preferentially expressed in thymocytes.126 Binding of MARs to the nuclear matrix usually enhances transcription, but it can sometimes repress gene expression.127 The absence of SATB1 causes the de-repression of at least 2% of all genes, leading to the blockade of normal T cell development.128 This example of cell-type specificity of chromatin loop architecture in relation to developmentally programmed gene expression demonstrates the importance of nuclear matrix-chromatin interactions in cells. To our knowledge, there is no specific MAR-binding protein described for preadipocytes and adipocytes. The finding of specific MAR-binding protein(s) in adipose precursor cells will be relevant to shed light on the mechanisms that control the repression or activation of genes for precursor cells to acquire the adipocyte phonotype.
The protein composition of the nuclear matrix is complex and analysis by 2D gel electrophoresis revealed 200 major protein spots. A subset of these proteins is cell-type specific, and the expression of some proteins of the NM correlates with malignat transformation.129,130 Lamins can be detected at sites along nuclear filaments,89 but they do not seem to form the structural core of the NM. NuMA (nuclear mitotic apparatus protein) is a good example of a NM protein, although its primary function during interphase is not fully defined yet. NuMA shows homology to some structural filament-forming proteins such as cytokeratins, nuclear lamins and myosin heavy chain.131,132 Overexpression of NuMA lacking the nuclear localization signal results in cytoplasmic aggregates,133 whereas overexpression of full-length NuMA leads to the formation of a lattice-like structure in the nucleus.133,134 NuMA can self-assemble into multi-arm oligomers in vitro.134 Further, it is resistant to detergent extraction and DNAse treatment, but a fraction of NuMA also interacts with chromatin through its binding to MARs.135 Such interaction may have an impact in chromatin organization, as shown by changes in chromatin epigenetic markers H4K20Me and acetyl-H4 upon silencing of NuMA.136 Different nuclear factors have been shown to be associated with the NM, e.g. histone deacetylases, steroid hormone receptors, oncogene proteins like c-myb,117 proteins of the SWI/SNF complex,137 and the mitotic scaffold associated protein Sc II.138 It has been proposed that actively transcribing RNA polymerases are located on the NM near actively transcribed genes, together with bound transcription factors, facilitating their accessibility for binding to the promoter and regulating the expression of their target genes.139-141
We showed that a fraction of C/EBPβ, a transcription factor required for proper adipogenesis,142,143 is associated with the NM.144 The non-histone nuclear protein Heterochromatin Protein (HP)1α interacts with C/EBPβ, restraining its transcriptional capacity.144 When we mapped the subnuclear domains where HP1α -C/EBPβ complexes localize, they were distributed mainly in heterochromatic domains, were also found in euchromatin but were completely absent in the NM.144 Thus, it is likely that the matrix-associated fraction of C/EBPβ that does not interact with HP1α represents the potentially active pool of C/EBPβ that regulates specific genes such as c/ebpα, which is activated by C/EBPβ at the onset of adipogenesis.144 It is of note that active genes can be clustered in specialized nuclear domains termed transcription factories145-147; thus the C/EBPβ pool associated with the NM may efficiently regulate clusters of C/EBP target genes. Furthermore, the fraction of C/EBPβ associated with the NM is also sensitive to treatment with the histone deacetylase inhibitor Trichostatin A, suggesting that acetylation of C/EBPβ, and possibly of other nuclear factors, may participate in the regulation of the equilibrium of C/EBPβ bound to chromatin and the nuclear matrix compartments.144,148
We also found that the high molecular weight immunophilin FKBP51 interacts with the NM, depending on the presence of ribonucleoproteins and/or RNA.98 FKBP51 has been shown to be a negative regulator of the transcriptional capacity of the GR as well as the mineralocorticoid receptor (MR).149,150 Recent studies have revealed a dramatic and dynamic modulation of the chromatin landscape during the first hours of adipocyte differentiation.151-155 These changes coincide with cooperative binding of early adipogenic transcription factors to transcription factor “hotspots.”154 GR is one of the transcription factors that binds to more than 4,000 sites, and its binding appears to be highly correlated with hypersensitivity to DNase I treatment, suggesting that an open chromatin structure may be required for GR binding, and/or that GR may be involved in early chromatin remodeling without massive gene transcriptional activation.153,154 By analogy, one can speculate that in the nuclear matrix compartment FKBP51 may interact with co-repressors, co-activators and/or components of the chromatin remodeling machinery, and in this way exert its control on GR/MR-dependent gene expression.
Does the genome's 3D organization undergo modifications during adipogenesis?
Mammalian cell nuclei with diameters in the range of 10-20 μm contain approximately 2 to 4 m of DNA in G1 and in G2, respectively. DNA is wrapped around histone octamers that constitute the nucleosomes, the first level of chromatin organization.156 Chromatin threads were named “chromosomes” by W. Waldeyer in 1,888,157 referring symply to the staining properties of the structures observed. By the end of the 19th and the beginning of the 20th century, Theodor Boveri formulated the hypothesis that each interphase chromosome occupies a distinct portion of the nuclear volume, introducing the term and concept of chromosome territory (CT) (Fig. 1A).158 Almost a century later, the existence of chromosome territories was demonstrated in pioneering microlaser experiments by Thomas and Christoph Cremer.162 They used laser-UV-microirradiation to induce local genome damage, followed by pulse-labeling of damaged DNA with 3H-thymidine, and autoradiography to detect 3H-thymidine incorporation in nuclei fixed immediately after the pulse. They thus predicted that inflicting DNA damage within a small volume of the nucleus would yield different results depending on how chromosomes were arranged. If chromosomes occupied distinct territories, localized damage would affect only a small subset of chromosomes, whereas if the chromatin fibers of each chromosome were randomly distributed throughout the nucleus, many of them would be damaged. They showed that only a subset of the chromosomes was damaged, providing the first experimental evidence for the existence of CT.159 Later, fluorescent in situ hybridization (FISH) techniques and the generation of chromosome specific painting probes for a large number of species allowed the direct visualization of individual CTs and their study in the interphase nucleus.160 The importance of the existence of CTs is reinforced their conservation during evolution over several hundred million years,161-164 demonstrating that the radial arrangement of chromatin is a fundamental feature of nuclear architecture. The impressive evolutionary conservation argues in favor of an adaptative value, yet the functional implications of non-random radial arrangements are not fully understood.
It is not known how the position of the CT is established in the nucleus. Several studies reported a preferential positioning of gene-rich CTs toward the center of the nucleus, and of gene-poor CTs toward the nuclear periphery.165-169 In nuclei of cultured human fibroblasts, which have a shape resembling a flat cylinder or ellipsoid, a size-dependent distribution of CTs was described.166,169 CT organization is maintained despite the extraction of more than 90% of the histones and other soluble proteins in DNA-rich nuclear matrix preparations.170 The NM possibly provides a scaffold or a basic structure to support CTs.116 It was shown that complete extraction of the internal NM components by RNase treatment followed by 2M NaCl results in disruption of higher order CT architecture, supporting a role of NM in the conformation/maintenance of the CTs architecture.116 The NM associations suggest that CTs may be constrained with a limited degree of mobility but with enough plasticity to allow motion and possibly dynamic shape changes.171,172 However, despite disruption of CTs upon removal of the nuclear matrix, the DNA remains predominantly inside the residual nuclear structure, and a small amount of DNA can extend past the nuclear lamina boundary forming a faint DNA halo.116 These results suggest that the nuclear lamina also plays a role in the anchoring of chromosomal DNA and that specific territorial arrangements require an intact NM.116 Indeed, in human cells, genome-wide mapping of lamin B1 binding sites has shown that this protein is not evenly bound to all chromosomes, and that the number of lamin B1 interactions per chromosome correlates with their radial positioning.66 Moreover, modification of the expression of lamins influences the radial positioning of some peripheral CTs.173 Lamins interact with DNA at MARs and LADs as well as at specific chromatin structures, such as centromeres and telomeres (reviewed in174). It has been demonstrated that lamin A/C and the lamin B receptor (LBR), a NE transmembrane protein, not only tethers heterochromatin to the nuclear periphery but also mediates control of gene expression during cell differentiation.175 Since lamins are ubiquitously distributed, it has been proposed that cell-type specific combination of NE transmembrane proteins may help to drive the spatial distribution of chromosomes, generating a genome organization distinctive to each cell type (reviewed in65). However, it will be necessary to explore whether cell-type specific components of the NM may be partners of NE transmembrane proteins in such a complex and whether it has a central function. In summary, the coordinated action of chromatin interactions with components of the NM, lamins, as well as with tissue specific NE transmembrane proteins could control the precise positioning of chromosomes in the interphase nucleus.
Does the genome's 3D organization undergo modifications during adipogenesis? Kuroda et al examined the relative and radial position of the CTs of human chromosomes 12 and 16 during differentiation of preadipocytes obtained from healthy female donors undergoing cosmetic liposuction procedures.176 There are 2 key aspects to consider with respect to CT positioning: the absolute radial location within the nucleus and the position of CTs relative to one another. An increased size of CT16, as well as a close association between territories of chromosomes 12 and 16 due to changes in the relative positioning between them were found in differentiated adipocytes, an association not observed in preadipocytes.176 This change in the relative positioning of CTs 12 and 16 may play a role in the t(12;16)(q13.3;p11.2) translocation present in 95-98% of myxoid and rounded cell liposarcomas. This chromosomal translocation creates a chimeric oncogene comprising part of the TLS/FUS gene found at 16p11.2 and part of the CHOP gene found at 12q13.3.177-180 The resultant fusion protein is crucial because it deregulates the expression of the adipocyte differentiation gene DOL54.181 In spite of not knowing the molecular mechanism that leads to specific translocations in tumor cells, close juxtaposition of interphase chromosomes may play an important role, as shown in chronic myeloid and promyelocytic leukemias.182-185 Although further studies are required to elucidate the mechanism underlying the adipocyte-specific changes in position of chromosomes 12 and 16, this study provided the first evidence of the relative repositioning of CTs during adipogenesis.
A non-random positioning of CTs was also demonstrated during the differentiation of mesenchymal stem cells (MSC) derived from porcine bone marrow, with the most dramatic change in position observed for CT4 moving from the nuclear periphery toward the nuclear interior.186 In addition, using 3D FISH analysis, it was shown that the architecture of CT changed during the course of MSC adipocyte differentiation. In undifferentiated MSCs, CTs are condensed with gene signals buried inside them. In contrast, in adipose-differentiated MSC, CTs are decondensed, exhibiting approximately twice the size compared to nuclei prior induction of adipogenesis; they also display loops of chromatin emanating from the CT with the gene signal at the end of the loop, oriented toward the nuclear interior.186 The nuclear positions of genes and the CTs carrying these gene loci have also been examined. The nuclear position of genes involved in adipogenesis is altered: genes that become more internally located in the nucleus are actively transcribed, while genes that move toward the nuclear periphery are down–regulated.186 More dramatic differences in localization were found for gene loci than for CTs, revealing genes moving from the nuclear periphery to the interior (PPARG, FABP4, GATA2), from an intermediate position to the nuclear interior (SREBF1, CEBPB) and from a peripheral to an intermediate location (CREB).186 Thus 6 out of 7 genes required for proper differentiation significantly change their nuclear location during adipogenesis. An interesting gene behavior is exhibited by GATA2, whose expression is restricted to preadipocytes where it exerts an antiadipogenic effect by the direct suppression of PPARγ.187 GATA2 relocates from a peripheral nuclear location to the interior at day 7 post-induction of adipogenesis of MSC, coinciding with its highest level of expression.186 By day 14, its expression falls and the gene localizes in the nuclear periphery again, consistent with the concept that the nuclear periphery acts as a transcriptional repressive domain. Furthermore, when MSC are differentiated, PPARG, SREBF1, FABP4 and GATA2 reside in a different compartment relative to their parent chromosome, showing that genes associated with adipogenesis loop out from their CT during differentiation and suggesting that this event is possibly associated with their increased expression.186
Insight into how chromatin is organized within CTs has been advanced by the development of chromosome conformation capture (3C)188 and derivative methods (4C, 5C, HI-C and single cell HI-C)189-193 which allow delineation of the organization of chromosomes in the 3-dimensional space of the interphase nucleus. With this information, when we zoom into chromosomal structure, we find that chromosomes are comprised of topologically associating domains (TADs) (Fig. 1A) defined as regions of high local contact frequency, separated by boundaries where contacts are relatively infrequent.194-196 In spite of TADs being too small to be visualized by microscopy-based methods, data obtained by FISH analysis is consistent with data obtained by 3C and related techniques.194,195,197 Mammalian genomes contain approximately 2000 TADs, and the CCCTC-binding factor and cohesin mediate their establishment and/or maintenance.194,197-199 Several chromatin associated proteins, transcription factors and cofactors participate in chromatin interactions of TADs that preferentially are intra-chromosomal rather than inter-chromosomal.192,196,200 Over one million interactions between loci have been detected genome-wide; these contacts make possible the interaction between enhancers and promoters that are kb distances in a linear sense but are in close physical proximity.201 The 3D organization of enhancers and promoters facilitates the interaction between protein complexes bound at enhancers with those bound at promoters, and can thereby modulate gene transcription in a cell-type specific manner. The same TAD can be found in different cell types or differentiation steps, but may be located in a different nuclear compartment (active vs. inactive), as part of a mechanism of control of cell-type or differentiation dependent gene expression. By 3C assay, the PPARG promoter is positioned in proximity to the promoters of genes encoding adipokines such as leptin and adiponectin, as well as of genes that encode lipid droplet-associated proteins.202 This positioning event takes place at early time points post-induction of adipogenesis, when these genes are not actively expressed, but when changes occur in chromatin accessibility and occupancy of the PPARγ promoter by transcription factors responsible for its activation.151,154,203 PPARG is not actively expressed at this early time, possibly because it requires chromatin remodeling events that take place later as differentiation progresses.153,154,204 The intergenic interactions of the PPARG promoter are dependent on C/EBPβ, since knockdown of this transcription factor, required for proper differentiation, abrogates the interactions.202 Furthermore, pharmacological inhibition of PKA or knockdown of the catalytic subunits of the kinase also prevent the intergenic interactions of the PPARG promoter, indicating that genome reorganization, at least for the PPARG loci, depends on c-AMP-PKA signaling.202
Once the organization of the genome is established in white, brown or beige adipocytes, the chromatin becomes stable, based on the fact that during browning of human white adipose cells by long-term exposure to PPARγ agonists, considerable changes in gene expression are found without major changes in the chromatin landscape.205 Superenhancers have been defined as clusters of enhancers that constitute regulatory nodes controlling expression of genes defining cell identity.206 When PPARγ-binding sites located within 12.5 kb from each other were ranked according to the intensity of their Mediator complex subunit 1 signal, over 1100 PPARγ superenhancers were identified in beige adipocytes, and 324 of them were exclusively present in beige adipose cells.205 These particular PPARγ superenhancers are in the vicinity of beige-selective genes which encode functionally important metabolic regulators involved in fatty acid degradation and β-oxidation. Thus during the reprogramming events of white adipose to beige cells, although no major changes in chromatin take place, the selective activation of PPARγ superenhancers seems to be required.205 Overall the physical association of coordinately regulated genes facilitates their control, suggesting that active co-regulated genes generate nuclear hot spots for precise and efficient control of transcription.
Future perspectives
When adipogenesis is triggered the architecture of the nucleus of progenitor cells undergoes substantial modifications (Fig. 1C). The NL is reorganized, CTs decondensed and change their relative position, adipogenic genes such as C/EBPβ and PPARγ undergo repositioning, accompanied by dramatic changes in chromatin accessibility, promoters occupancy and superenhancers formation among many other modifications. However, many questions are still waiting for answers, to gain insight not only into adipogenesis, but into cell differentiation processes in general. How are nuclear compartments established to produce a precise pattern of gene expression? Is this a cause or a consequence of the differentiation program? It has been extensively studied all the signaling pathways that control the process of adipogenesis (reviewed in10,143), but how do biochemical and mechanical signaling are coordinately transduced in the organization of nuclear compartments in the adipose or any other cell fate determination? It is known that active genes are preferentially localized in euchromatin, and in many cases even loop out of their CT. In contrast, when genes that normally reside in euchromatin translocate to centromeric heterochromatin or are preferentially positioned at the nuclear periphery during cell differentiation, they are silenced. How are these events regulated? Which signals required for cells to differentiate are transduced by changes of the nuclear architecture? We still know very little about the 3D organization of the genome during adipogenesis, but undoubtedly, future studies will unveil the relationships between the architecture of the genome and the control of gene expression, as part of the mechanism that controls the acquisition of the phenotype of the different types of adipose cells. These studies will depend, in part, on new technologies to prove or disprove the proposed models. After all, Boveri's hypothesis on the existence of CTs waited for almost a century for experimental verification. Studying the dynamics of nuclear architecture during adipogenesis will increase our knowledge of the mechanisms that control cell differentiation, while enhancing our ability to understand adipose metabolism in health and disease.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We are very thankful to Prof. Jessica Schwartz for her constructive suggestions and comments on the manuscript.
Funding
This work was supported by grants from Agencia Nacional de Promoción Científica y Tecnológica (PICT2012-2612 to G.P.P, and PICT2013-1745 to J.T.), and Fundación René Barón. N.M.G. is a recipient of a doctoral fellowship and N.L.C of a post-doctoral fellowship from CONICET. M.I.R.C. is a recipient of a doctoral fellowship from Agencia Nacional de Promoción Científica y Tecnológica.
References
- [1].Schwartz MW, Porte D Jr. Diabetes, obesity, and the brain. Science 2005; 307:375-9; PMID:15662002; http://dx.doi.org/ 10.1126/science.1104344 [DOI] [PubMed] [Google Scholar]
- [2].Olefsky JM. Fat talks, liver and muscle listen. Cell 2008; 134:914-6; PMID:18805083; http://dx.doi.org/ 10.1016/j.cell.2008.09.001 [DOI] [PubMed] [Google Scholar]
- [3].Galic S, Oakhill JS, Steinberg GR. Adipose tissue as an endocrine organ. Mol Cell Endocrinol 2010; 316:129-39; PMID:19723556; http://dx.doi.org/ 10.1016/j.mce.2009.08.018 [DOI] [PubMed] [Google Scholar]
- [4].Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest 2003; 112:1796-808; PMID:14679176; http://dx.doi.org/ 10.1172/JCI200319246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Lumeng CN, Deyoung SM, Bodzin JL, Saltiel AR. Increased inflammatory properties of adipose tissue macrophages recruited during diet-induced obesity. Diabetes 2007; 56:16-23; PMID:17192460; http://dx.doi.org/ 10.2337/db06-1076 [DOI] [PubMed] [Google Scholar]
- [6].Spiegelman BM, Flier JS. Obesity and the regulation of energy balance. Cell 2001; 104:531-43; PMID:11239410; http://dx.doi.org/ 10.1016/S0092-8674(01)00240-9 [DOI] [PubMed] [Google Scholar]
- [7].Cao H. Adipocytokines in obesity and metabolic disease. J Endocrinol 2014; 220:T47-59; PMID:24403378; http://dx.doi.org/ 10.1530/JOE-13-0339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Garg A, Agarwal AK. Lipodystrophies: disorders of adipose tissue biology. Biochim Biophys Acta 2009; 1791:507-13; PMID:19162222; http://dx.doi.org/ 10.1016/j.bbalip.2008.12.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Waki H, Tontonoz P. Endocrine functions of adipose tissue. Annu Rev Pathol 2007; 2:31-56; PMID:18039092; http://dx.doi.org/ 10.1146/annurev.pathol.2.010506.091859 [DOI] [PubMed] [Google Scholar]
- [10].Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell 2014; 156:20-44; PMID:24439368; http://dx.doi.org/ 10.1016/j.cell.2013.12.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Rodbell M. Metabolism of Isolated Fat Cells. I. Effects of Hormones on Glucose Metabolism and Lipolysis. J Biol Chem 1964; 239:375-80; PMID:14169133 [PubMed] [Google Scholar]
- [12].Hollenberg CH, Vost A. Regulation of DNA synthesis in fat cells and stromal elements from rat adipose tissue. J Clin Invest 1969; 47:2485-98; PMID:4304653; http://dx.doi.org/ 10.1172/JCI105930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Sengenes C, Lolmede K, Zakaroff-Girard A, Busse R, Bouloumie A. Preadipocytes in the human subcutaneous adipose tissue display distinct features from the adult mesenchymal and hematopoietic stem cells. J Cell Physiol 2005; 205:114-22; PMID:15880450; http://dx.doi.org/ 10.1002/jcp.20381 [DOI] [PubMed] [Google Scholar]
- [14].Rodeheffer MS, Birsoy K, Friedman JM. Identification of white adipocyte progenitor cells in vivo. Cell 2008; 135:240-9; PMID:18835024; http://dx.doi.org/ 10.1016/j.cell.2008.09.036 [DOI] [PubMed] [Google Scholar]
- [15].Tang W, Zeve D, Suh JM, Bosnakovski D, Kyba M, Hammer RE, Tallquist MD, Graff JM. White fat progenitor cells reside in the adipose vasculature. Science 2008; 322:583-6; PMID:18801968; http://dx.doi.org/ 10.1126/science.1156232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Iyama K, Ohzono K, Usuku G. Electron microscopical studies on the genesis of white adipocytes: differentiation of immature pericytes into adipocytes in transplanted preadipose tissue. Virchows Arch B Cell Pathol Incl Mol Pathol 1979; 31:143-55; PMID:42211; http://dx.doi.org/ 10.1007/BF02889932 [DOI] [PubMed] [Google Scholar]
- [17].Cinti S, Cigolini M, Bosello O, Bjorntorp P. A morphological study of the adipocyte precursor. J Submicrosc Cytol 1984; 16:243-51; PMID:6325721 [PubMed] [Google Scholar]
- [18].Jones DL, Wagers AJ. No place like home: anatomy and function of the stem cell niche. Nat Rev Mol Cell Biol 2008; 9:11-21; PMID:18097443; http://dx.doi.org/ 10.1038/nrm2319 [DOI] [PubMed] [Google Scholar]
- [19].Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev 2004; 84:277-359; PMID:14715917; http://dx.doi.org/ 10.1152/physrev.00015.2003 [DOI] [PubMed] [Google Scholar]
- [20].Timmons JA, Wennmalm K, Larsson O, Walden TB, Lassmann T, Petrovic N, Hamilton DL, Gimeno RE, Wahlestedt C, Baar K, et al.. Myogenic gene expression signature establishes that brown and white adipocytes originate from distinct cell lineages. Proc Natl Acad Sci U S A 2007; 104:4401-6; PMID:17360536; http://dx.doi.org/ 10.1073/pnas.0610615104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Seale P, Kajimura S, Yang W, Chin S, Rohas LM, Uldry M, Tavernier G, Langin D, Spiegelman BM. Transcriptional control of brown fat determination by PRDM16. Cell Metab 2007; 6:38-54; PMID:17618855; http://dx.doi.org/ 10.1016/j.cmet.2007.06.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Seale P, Bjork B, Yang W, Kajimura S, Chin S, Kuang S, Scime A, Devarakonda S, Conroe HM, Erdjument-Bromage H, et al.. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008; 454:961-7; PMID:18719582; http://dx.doi.org/ 10.1038/nature07182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Seale P, Kajimura S, Spiegelman BM. Transcriptional control of brown adipocyte development and physiological function–of mice and men. Genes Dev 2009; 23:788-97; PMID:19339685; http://dx.doi.org/ 10.1101/gad.1779209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Seale P, Conroe HM, Estall J, Kajimura S, Frontini A, Ishibashi J, Cohen P, Cinti S, Spiegelman BM. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest 2011; 121:96-105; PMID:21123942; http://dx.doi.org/ 10.1172/JCI44271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Hany TF, Gharehpapagh E, Kamel EM, Buck A, Himms-Hagen J, von Schulthess GK. Brown adipose tissue: a factor to consider in symmetrical tracer uptake in the neck and upper chest region. Eur J Nucl Med Mol Imaging 2002; 29:1393-8; PMID:12271425; http://dx.doi.org/ 10.1007/s00259-002-0902-6 [DOI] [PubMed] [Google Scholar]
- [26].Yeung HW, Grewal RK, Gonen M, Schoder H, Larson SM. Patterns of (18)F-FDG uptake in adipose tissue and muscle: a potential source of false-positives for PET. J Nucl Med 2003; 44:789-96; PMID:NOT_FOUND17473055 [PubMed] [Google Scholar]
- [27].Nedergaard J, Bengtsson T, Cannon B. Unexpected evidence for active brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab 2007; 293:E444-52; PMID:17473055; http://dx.doi.org/ 10.1152/ajpendo.00691.2006 [DOI] [PubMed] [Google Scholar]
- [28].van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, Schrauwen P, Teule GJ. Cold-activated brown adipose tissue in healthy men. N Engl J Med 2009; 360:1500-8; PMID:19357405; http://dx.doi.org/ 10.1056/NEJMoa0808718 [DOI] [PubMed] [Google Scholar]
- [29].Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, Taittonen M, Laine J, Savisto NJ, Enerback S, et al.. Functional brown adipose tissue in healthy adults. N Engl J Med 2009; 360:1518-25; PMID:19357407; http://dx.doi.org/ 10.1056/NEJMoa0808949 [DOI] [PubMed] [Google Scholar]
- [30].Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng YH, Doria A, et al.. Identification and importance of brown adipose tissue in adult humans. N Engl J Med 2009; 360:1509-17; PMID:19357406; http://dx.doi.org/ 10.1056/NEJMoa0810780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Wilson-Fritch L, Nicoloro S, Chouinard M, Lazar MA, Chui PC, Leszyk J, Straubhaar J, Czech MP, Corvera S. Mitochondrial remodeling in adipose tissue associated with obesity and treatment with rosiglitazone. J Clin Invest 2004; 114:1281-9; PMID:15520860; http://dx.doi.org/ 10.1172/JCI21752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Sparks LM, Xie H, Koza RA, Mynatt R, Hulver MW, Bray GA, Smith SR. A high-fat diet coordinately downregulates genes required for mitochondrial oxidative phosphorylation in skeletal muscle. Diabetes 2005; 54:926-33; http://dx.doi.org/ 10.2337/diabetes.54.7.1926 [DOI] [PubMed] [Google Scholar]
- [33].Abdul-Ghani MA, DeFronzo RA. Mitochondrial dysfunction, insulin resistance, and type 2 diabetes mellitus. Curr Diab Rep 2008; 8:173-8; PMID:18625112; http://dx.doi.org/ 10.1007/s11892-008-0030-1 [DOI] [PubMed] [Google Scholar]
- [34].Vegiopoulos A, Muller-Decker K, Strzoda D, Schmitt I, Chichelnitskiy E, Ostertag A, Berriel Diaz M, Rozman J, Hrabe de Angelis M, Nusing RM, et al.. Cyclooxygenase-2 controls energy homeostasis in mice by de novo recruitment of brown adipocytes. Science 2010; 328:1158-61; PMID:20448152; http://dx.doi.org/ 10.1126/science.1186034 [DOI] [PubMed] [Google Scholar]
- [35].Petrovic N, Walden TB, Shabalina IG, Timmons JA, Cannon B, Nedergaard J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J Biol Chem 2010; 285:7153-64; PMID:20028987; http://dx.doi.org/ 10.1074/jbc.M109.053942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Barbatelli G, Murano I, Madsen L, Hao Q, Jimenez M, Kristiansen K, Giacobino JP, De Matteis R, Cinti S. The emergence of cold-induced brown adipocytes in mouse white fat depots is determined predominantly by white to brown adipocyte transdifferentiation. Am J Physiol Endocrinol Metab 2010; 298:E1244-53; PMID:20354155; http://dx.doi.org/ 10.1152/ajpendo.00600.2009 [DOI] [PubMed] [Google Scholar]
- [37].Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, Khandekar M, Virtanen KA, Nuutila P, Schaart G, et al.. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012; 150:366-76; PMID:22796012; http://dx.doi.org/ 10.1016/j.cell.2012.05.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Ohno H, Shinoda K, Spiegelman BM, Kajimura S. PPARgamma agonists induce a white-to-brown fat conversion through stabilization of PRDM16 protein. Cell Metab 2012; 15:395-404; PMID:22405074; http://dx.doi.org/ 10.1016/j.cmet.2012.01.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Ye L, Wu J, Cohen P, Kazak L, Khandekar MJ, Jedrychowski MP, Zeng X, Gygi SP, Spiegelman BM. Fat cells directly sense temperature to activate thermogenesis. Proc Natl Acad Sci U S A 2013; 110:12480-5; PMID:23818608; http://dx.doi.org/ 10.1073/pnas.1310261110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Tseng YH, Kokkotou E, Schulz TJ, Huang TL, Winnay JN, Taniguchi CM, Tran TT, Suzuki R, Espinoza DO, Yamamoto Y, et al.. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 2008; 454:1000-4; PMID:18719589; http://dx.doi.org/ 10.1038/nature07221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Whittle AJ, Carobbio S, Martins L, Slawik M, Hondares E, Vazquez MJ, Morgan D, Csikasz RI, Gallego R, Rodriguez-Cuenca S, et al.. BMP8B increases brown adipose tissue thermogenesis through both central and peripheral actions. Cell 2012; 149:871-85; PMID:22579288; http://dx.doi.org/ 10.1016/j.cell.2012.02.066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Schulz TJ, Huang P, Huang TL, Xue R, McDougall LE, Townsend KL, Cypess AM, Mishina Y, Gussoni E, Tseng YH. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 2013; 495:379-83; PMID:23485971; http://dx.doi.org/ 10.1038/nature11943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Cypess AM, White AP, Vernochet C, Schulz TJ, Xue R, Sass CA, Huang TL, Roberts-Toler C, Weiner LS, et al.. Anatomical localization, gene expression profiling and functional characterization of adult human neck brown fat. Nat Med 2013; 19:635-9; PMID:23603815; http://dx.doi.org/ 10.1038/nm.3112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Lidell ME, Betz MJ, Dahlqvist Leinhard O, Heglind M, Elander L, Slawik M, Mussack T, Nilsson D, Romu T, Nuutila P, et al.. Evidence for two types of brown adipose tissue in humans. Nat Med 2013; 19:31-4; http://dx.doi.org/ 10.1038/nm.3017 [DOI] [PubMed] [Google Scholar]
- [45].Spalding KL, Arner E, Westermark PO, Bernard S, Buchholz BA, Bergmann O, Blomqvist L, Hoffstedt J, Naslund E, Britton T, et al.. Dynamics of fat cell turnover in humans. Nature 2008; 453:783-7; PMID:18454136; http://dx.doi.org/ 10.1038/nature06902 [DOI] [PubMed] [Google Scholar]
- [46].Siersbaek R, Nielsen R, Mandrup S. Transcriptional networks and chromatin remodeling controlling adipogenesis. Trends Endocrinol Metab 2012; 23:56-64; PMID:22079269; http://dx.doi.org/ 10.1016/j.tem.2011.10.001 [DOI] [PubMed] [Google Scholar]
- [47].Algire C, Medrikova D, Herzig S. White and brown adipose stem cells: from signaling to clinical implications. Biochim Biophys Acta 2013; 1831:896-904; PMID:23051608; http://dx.doi.org/ 10.1016/j.bbalip.2012.10.001 [DOI] [PubMed] [Google Scholar]
- [48].Jenuwein T, Allis CD. Translating the histone code. Science 2001; 293:1074-80; PMID:11498575; http://dx.doi.org/ 10.1126/science.1063127 [DOI] [PubMed] [Google Scholar]
- [49].Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Cell 2007; 128:635-8; PMID:17320500; http://dx.doi.org/ 10.1016/j.cell.2007.02.006 [DOI] [PubMed] [Google Scholar]
- [50].Munshi A, Shafi G, Aliya N, Jyothy A. Histone modifications dictate specific biological readouts. J Genet Genomics 2009; 36:75-88; PMID:19232306; http://dx.doi.org/ 10.1016/S1673-8527(08)60094-6 [DOI] [PubMed] [Google Scholar]
- [51].Sugii S, Evans RM. Epigenetic codes of PPARgamma in metabolic disease. FEBS Lett 2011; 585:2121-8; PMID:21605560; http://dx.doi.org/ 10.1016/j.febslet.2011.05.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Mikkelsen TS, Ku M, Jaffe DB, Issac B, Lieberman E, Giannoukos G, Alvarez P, Brockman W, Kim TK, Koche RP, et al.. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 2007; 448:553-60; PMID:17603471; http://dx.doi.org/ 10.1038/nature06008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Meissner A, Mikkelsen TS, Gu H, Wernig M, Hanna J, Sivachenko A, Zhang X, Bernstein BE, Nusbaum C, Jaffe DB, et al.. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature 2008; 454:766-70; PMID:18600261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Mikkelsen TS, Hanna J, Zhang X, Ku M, Wernig M, Schorderet P, Bernstein BE, Jaenisch R, Lander ES, Meissner A. Dissecting direct reprogramming through integrative genomic analysis. Nature 2008; 454:49-55; PMID:18509334; http://dx.doi.org/ 10.1038/nature07056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Cremer T, Zakhartchenko V. Nuclear architecture in developmental biology and cell specialisation. Reprod Fertil Dev 2011; 23:94-106; PMID:21366985; http://dx.doi.org/ 10.1071/RD10249 [DOI] [PubMed] [Google Scholar]
- [56].Fawcett DW. On the occurrence of a fibrous lamina on the inner aspect of the nuclear envelope in certain cells of vertebrates. Am J Anat 1966; 119:129-45; PMID:6007824; http://dx.doi.org/ 10.1002/aja.1001190108 [DOI] [PubMed] [Google Scholar]
- [57].Patrizi G, Poger M. The ultrastructure of the nuclear periphery. The zonula nucleum limitans. J Ultrastruct Res 1967; 17:127-36; PMID:6017352; http://dx.doi.org/ 10.1016/S0022-5320(67)80025-X [DOI] [PubMed] [Google Scholar]
- [58].Ho CY, Lammerding J. Lamins at a glance. J Cell Sci 2012; 125:2087-93; PMID:22669459; http://dx.doi.org/ 10.1242/jcs.087288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].McKeon FD, Kirschner MW, Caput D. Homologies in both primary and secondary structure between nuclear envelope and intermediate filament proteins. Nature 1986; 319:463-8; PMID:3453101; http://dx.doi.org/ 10.1038/319463a0 [DOI] [PubMed] [Google Scholar]
- [60].Dechat T, Pfleghaar K, Sengupta K, Shimi T, Shumaker DK, Solimando L, Goldman RD. Nuclear lamins: major factors in the structural organization and function of the nucleus and chromatin. Genes Dev 2008; 22:32-53; http://dx.doi.org/ 10.1101/gad.1652708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Aaronson RP, Blobel G. Isolation of nuclear pore complexes in association with a lamina. Proc Natl Acad Sci U S A 1975; 72:007-11; http://dx.doi.org/ 10.1073/pnas.72.3.1007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Moir RD, Yoon M, Khuon S, Goldman RD. Nuclear lamins A and B1: different pathways of assembly during nuclear envelope formation in living cells. J Cell Biol 2000; 151:1155-68; PMID:11121432; http://dx.doi.org/ 10.1083/jcb.151.6.1155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Shimi T, Pfleghaar K, Kojima S, Pack CG, Solovei I, Goldman AE, Adam SA, Shumaker DK, Kinjo M, Cremer T, et al.. The A- and B-type nuclear lamin networks: microdomains involved in chromatin organization and transcription. Genes Dev 2008; 22:409-21; http://dx.doi.org/ 10.1101/gad.1735208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Crisp M, Liu Q, Roux K, Rattner JB, Shanahan C, Burke B, Stahl PD, Hodzic D. Coupling of the nucleus and cytoplasm: role of the LINC complex. J Cell Biol 2006; 172:41-53; PMID:16380439; http://dx.doi.org/ 10.1083/jcb.200509124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].de Las Heras JI, Meinke P, Batrakou DG, Srsen V, Zuleger N, Kerr AR, Schirmer EC. Tissue specificity in the nuclear envelope supports its functional complexity. Nucleus 2014; 4:40-77; PMID:NOT_FOUND18463634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Guelen L, Pagie L, Brasset E, Meuleman W, Faza MB, Talhout W, Eussen BH, de Klein A, Wessels L, de Laat W, et al.. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008; 453:948-51; PMID:18463634; http://dx.doi.org/ 10.1038/nature06947 [DOI] [PubMed] [Google Scholar]
- [67].Reddy KL, Zullo JM, Bertolino E, Singh H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature 2008; 452:243-7; PMID:18272965; http://dx.doi.org/ 10.1038/nature06727 [DOI] [PubMed] [Google Scholar]
- [68].Kosak ST, Skok JA, Medina KL, Riblet R, Le Beau MM, Fisher AG, Singh H. Subnuclear compartmentalization of immunoglobulin loci during lymphocyte development. Science 2002; 296:158-62; PMID:11935030; http://dx.doi.org/ 10.1126/science.1068768 [DOI] [PubMed] [Google Scholar]
- [69].Rodriguez-Navarro S, Fischer T, Luo MJ, Antunez O, Brettschneider S, Lechner J, Perez-Ortin JE, Reed R, Hurt E. Sus1, a functional component of the SAGA histone acetylase complex and the nuclear pore-associated mRNA export machinery. Cell 2004; 116:75-86; PMID:14718168; http://dx.doi.org/ 10.1016/S0092-8674(03)01025-0 [DOI] [PubMed] [Google Scholar]
- [70].Taddei A, Van Houwe G, Hediger F, Kalck V, Cubizolles F, Schober H, Gasser SM. Nuclear pore association confers optimal expression levels for an inducible yeast gene. Nature 2006; 441:774-8; PMID:16760983; http://dx.doi.org/ 10.1038/nature04845 [DOI] [PubMed] [Google Scholar]
- [71].Cabal GG, Genovesio A, Rodriguez-Navarro S, Zimmer C, Gadal O, Lesne A, Buc H, Feuerbach-Fournier F, Olivo-Marin JC, Hurt EC, et al.. SAGA interacting factors confine sub-diffusion of transcribed genes to the nuclear envelope. Nature 2006; 441:770-3; PMID:16760982; http://dx.doi.org/ 10.1038/nature04752 [DOI] [PubMed] [Google Scholar]
- [72].Brown CR, Kennedy CJ, Delmar VA, Forbes DJ, Silver PA. Global histone acetylation induces functional genomic reorganization at mammalian nuclear pore complexes. Genes Dev 2008; 22:627-39; PMID:18316479; http://dx.doi.org/ 10.1101/gad.1632708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Luo L, Gassman KL, Petell LM, Wilson CL, Bewersdorf J, Shopland LS. The nuclear periphery of embryonic stem cells is a transcriptionally permissive and repressive compartment. J Cell Sci 2009; 122:3729-37; PMID:19773359; http://dx.doi.org/ 10.1242/jcs.052555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Galigniana MD, Echeverria PC, Erlejman AG, Piwien-Pilipuk G. Role of molecular chaperones and TPR-domain proteins in the cytoplasmic transport of steroid receptors and their passage through the nuclear pore. Nucleus 2010; 1:299-308; PMID:21113270; http://dx.doi.org/ 10.4161/nucl.1.4.11743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Quinta HR, Maschi D, Gomez-Sanchez C, Piwien-Pilipuk G, Galigniana MD. Subcellular rearrangement of hsp90-binding immunophilins accompanies neuronal differentiation and neurite outgrowth. J Neurochem 2010; 115:716-34; PMID:20796173; http://dx.doi.org/ 10.1111/j.1471-4159.2010.06970.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Shimi T, Butin-Israeli V, Adam SA, Goldman RD. Nuclear lamins in cell regulation and disease. Cold Spring Harb Symp Quant Biol 2010; 75:25-31; http://dx.doi.org/ 10.1101/sqb.2010.75.045 [DOI] [PubMed] [Google Scholar]
- [77].Hegele RA, Joy TR, Al-Attar SA, Rutt BK. Thematic review series: Adipocyte Biology. Lipodystrophies: windows on adipose biology and metabolism. J Lipid Res 2007; 48:433-44; http://dx.doi.org/ 10.1194/jlr.R700004-JLR200 [DOI] [PubMed] [Google Scholar]
- [78].Garg A. Acquired and inherited lipodystrophies. N Engl J Med 2004; 350:1220-34; PMID:15028826; http://dx.doi.org/ 10.1056/NEJMra025261 [DOI] [PubMed] [Google Scholar]
- [79].Hegele R. LMNA mutation position predicts organ system involvement in laminopathies. Clin Genet 2005; 68:1-4; http://dx.doi.org/ 10.1111/j.1399-0004.2005.00447.x [DOI] [PubMed] [Google Scholar]
- [80].Capanni C, Mattioli E, Columbaro M, Lucarelli E, Parnaik VK, Novelli G, Wehnert M, Cenni V, Maraldi NM, Squarzoni S, et al.. Altered pre-lamin A processing is a common mechanism leading to lipodystrophy. Hum Mol Genet 2005; 14:1489-502; PMID:15843404; http://dx.doi.org/ 10.1093/hmg/ddi158 [DOI] [PubMed] [Google Scholar]
- [81].Tontonoz P, Hu E, Spiegelman BM. Stimulation of adipogenesis in fibroblasts by PPARy2 by a lipid-activated transcription factor. Cell 1994; 79:147-1156; http://dx.doi.org/ 10.1016/0092-8674(94)90006-X [DOI] [PubMed] [Google Scholar]
- [82].Tontonoz P, Spiegelman BM. Fat and beyond: the diverse biology of PPARgamma. Annu Rev Biochem 2008; 77:289-312; PMID:18518822; http://dx.doi.org/ 10.1146/annurev.biochem.77.061307.091829 [DOI] [PubMed] [Google Scholar]
- [83].Hegele RA, Cao H, Frankowski C, Mathews ST, Leff T. PPARG F388L, a transactivation-deficient mutant, in familial partial lipodystrophy. Diabetes 2002; 51:3586-90; PMID:12453919; http://dx.doi.org/ 10.2337/diabetes.51.12.3586 [DOI] [PubMed] [Google Scholar]
- [84].Agarwal AK, Garg A. A novel heterozygous mutation in peroxisome proliferator-activated receptor-gamma gene in a patient with familial partial lipodystrophy. J Clin Endocrinol Metab 2002; 87:408-11; PMID:11788685 [DOI] [PubMed] [Google Scholar]
- [85].Al-Shali K, Cao H, Knoers N, Hermus AR, Tack CJ, Hegele RA. A single-base mutation in the peroxisome proliferator-activated receptor gamma4 promoter associated with altered in vitro expression and partial lipodystrophy. J Clin Endocrinol Metab 2004; 89:5655-60; PMID:15531525; http://dx.doi.org/ 10.1210/jc.2004-0280 [DOI] [PubMed] [Google Scholar]
- [86].Caron M, Auclair M, Sterlingot H, Kornprobst M, Capeau J. Some HIV protease inhibitors alter lamin A/C maturation and stability, SREBP-1 nuclear localization and adipocyte differentiation. Aids 2003; 17:2437-44; PMID:14600514; http://dx.doi.org/ 10.1097/00002030-200311210-00005 [DOI] [PubMed] [Google Scholar]
- [87].Boguslavsky RL, Stewart CL, Worman HJ. Nuclear lamin A inhibits adipocyte differentiation: implications for Dunnigan-type familial partial lipodystrophy. Hum Mol Genet 2006; 15:653-63; PMID:16415042; http://dx.doi.org/ 10.1093/hmg/ddi480 [DOI] [PubMed] [Google Scholar]
- [88].Jackson DA, Cook PR. Visualization of a filamentous nucleoskeleton with a 23 nm axial repeat. Embo J 1988; 7:3667-77; PMID:3208744 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Hozak P, Sasseville AM, Raymond Y, Cook PR. Lamin proteins form an internal nucleoskeleton as well as a peripheral lamina in human cells. J Cell Sci 1995; 108(Pt 2):635-44; PMID:7769007 [DOI] [PubMed] [Google Scholar]
- [90].Broers JL, Machiels BM, van Eys GJ, Kuijpers HJ, Manders EM, van Driel R, Ramaekers FC. Dynamics of the nuclear lamina as monitored by GFP-tagged A-type lamins. J Cell Sci 1999; 112(Pt 20):3463-75; PMID:10504295 [DOI] [PubMed] [Google Scholar]
- [91].Taniura H, Glass C, Gerace L. A chromatin binding site in the tail domain of nuclear lamins that interacts with core histones. J Cell Biol 1995; 131:33-44; PMID:7559784; http://dx.doi.org/ 10.1083/jcb.131.1.33 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Goldberg M, Harel A, Brandeis M, Rechsteiner T, Richmond TJ, Weiss AM, Gruenbaum Y. The tail domain of lamin Dm0 binds histones H2A and H2B. Proc Natl Acad Sci U S A 1999; 96:2852-7; PMID:10077600; http://dx.doi.org/ 10.1073/pnas.96.6.2852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Broers JL, Kuijpers HJ, Ostlund C, Worman HJ, Endert J, Ramaekers FC. Both lamin A and lamin C mutations cause lamina instability as well as loss of internal nuclear lamin organization. Exp Cell Res 2005; 304:582-92; PMID:15748902; http://dx.doi.org/ 10.1016/j.yexcr.2004.11.020 [DOI] [PubMed] [Google Scholar]
- [94].Tilgner K, Wojciechowicz K, Jahoda C, Hutchison C, Markiewicz E. Dynamic complexes of A-type lamins and emerin influence adipogenic capacity of the cell via nucleocytoplasmic distribution of beta-catenin. J Cell Sci 2009; 122:401-13; PMID:19126678; http://dx.doi.org/ 10.1242/jcs.026179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Markiewicz E, Tilgner K, Barker N, van de Wetering M, Clevers H, Dorobek M, Hausmanowa-Petrusewicz I, Ramaekers FC, Broers JL, Blankesteijn WM, et al.. The inner nuclear membrane protein emerin regulates beta-catenin activity by restricting its accumulation in the nucleus. Embo J 2006; 25:275-85; http://dx.doi.org/ 10.1038/sj.emboj.7601230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA. Inhibition of adipogenesis by Wnt signaling. Science 2000; 289:950-953; PMID:10937998; http://dx.doi.org/ 10.1126/science.289.5481.950 [DOI] [PubMed] [Google Scholar]
- [97].Verstraeten VL, Renes J, Ramaekers FC, Kamps M, Kuijpers HJ, Verheyen F, Wabitsch M, Steijlen PM, van Steensel MA, Broers JL. Reorganization of the nuclear lamina and cytoskeleton in adipogenesis. Histochem Cell Biol 2011; 135:251-61; PMID:21350821; http://dx.doi.org/ 10.1007/s00418-011-0792-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Toneatto J, Guber S, Charo NL, Susperreguy S, Schwartz J, Galigniana MD, Piwien-Pilipuk G. Dynamic mitochondrial-nuclear redistribution of the immunophilin FKBP51 is regulated by the PKA signaling pathway to control gene expression during adipocyte differentiation. J Cell Sci 2013; 126:5357-68; PMID:24101724; http://dx.doi.org/ 10.1242/jcs.125799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Toneatto J, Charo NL, Galigniana NM, Piwien-Pilipuk G. Adipogenesis is under surveillance of Hsp90 and the high molecular weight Immunophilin FKBP51. Adipocyte 2015; 4:29-47; http://dx.doi.org/ 10.1080/21623945.2015.1049401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].D'Angelo MA, Hetzer MW. The role of the nuclear envelope in cellular organization. Cell Mol Life Sci 2006; 63:316-32; PMID:16389459; http://dx.doi.org/ 10.1007/s00018-005-5361-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Stuurman N. Identification of a conserved phosphorylation site modulating nuclear lamin polymerization. FEBS Lett 1997; 401:171-4; PMID:9013881; http://dx.doi.org/ 10.1016/S0014-5793(96)01464-0 [DOI] [PubMed] [Google Scholar]
- [102].Gonzalez JM, Navarro-Puche A, Casar B, Crespo P, Andres V. Fast regulation of AP-1 activity through interaction of lamin A/C, ERK1/2, and c-Fos at the nuclear envelope. J Cell Biol 2008; 183:653-66; PMID:19015316; http://dx.doi.org/ 10.1083/jcb.200805049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Gregoire FM, Smas CM, Sul HS. Understanding adipocyte differentiation. Physiol Rev 1998; 78:783-809; PMID:9674695 [DOI] [PubMed] [Google Scholar]
- [104].Maniotis AJ, Chen CS, Ingber DE. Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc Natl Acad Sci U S A 1997; 94:849-54; PMID:9023345; http://dx.doi.org/ 10.1073/pnas.94.3.849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Wang N, Butler JP, Ingber DE. Mechanotransduction across the cell surface and through the cytoskeleton. Science 1993; 260:1124-7; PMID:7684161; http://dx.doi.org/ 10.1126/science.7684161 [DOI] [PubMed] [Google Scholar]
- [106].Wang N, Tytell JD, Ingber DE. Mechanotransduction at a distance: mechanically coupling the extracellular matrix with the nucleus. Nat Rev Mol Cell Biol 2009; 10:75-82; PMID:19197334; http://dx.doi.org/ 10.1038/nrm2594 [DOI] [PubMed] [Google Scholar]
- [107].Lammerding J, Schulze PC, Takahashi T, Kozlov S, Sullivan T, Kamm RD, Stewart CL, Lee RT. Lamin A/C deficiency causes defective nuclear mechanics and mechanotransduction. J Clin Invest 2004; 113:370-8; PMID:14755334; http://dx.doi.org/ 10.1172/JCI200419670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Lombardi ML, Jaalouk DE, Shanahan CM, Burke B, Roux KJ, Lammerding J. The interaction between nesprins and sun proteins at the nuclear envelope is critical for force transmission between the nucleus and cytoskeleton. J Biol Chem 2011; 286:26743-53; PMID:21652697; http://dx.doi.org/ 10.1074/jbc.M111.233700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Gilbert PM, Havenstrite KL, Magnusson KE, Sacco A, Leonardi NA, Kraft P, Nguyen NK, Thrun S, Lutolf MP, Blau HM. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science 2010; 329:1078-81; PMID:20647425; http://dx.doi.org/ 10.1126/science.1191035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Swift J, Ivanovska IL, Buxboim A, Harada T, Dingal PC, Pinter J, Pajerowski JD, Spinler KR, Shin JW, Tewari M, et al.. Nuclear lamin-A scales with tissue stiffness and enhances matrix-directed differentiation. Science 2013; 341:1240104; PMID:23990565; http://dx.doi.org/ 10.1126/science.1240104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [111].Zuleger N, Boyle S, Kelly DA, de las Heras JI, Lazou V, Korfali N, Batrakou DG, Randles KN, Morris GE, Harrison DJ, et al.. Specific nuclear envelope transmembrane proteins can promote the location of chromosomes to and from the nuclear periphery. Genome Biol 2013; 14:R14; PMID:23414781; http://dx.doi.org/ 10.1186/gb-2013-14-2-r14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Fawcet DW. An Atlas of Fine Structure: The Cell, Its Organelles and Inclusions, W. B. Saunders Co. (ed). Ch. 2, pp. 6–7. Philadelphia, 1966. [Google Scholar]
- [113].Steele WJ, Busch H. Studies on the ribonucleic acid components of the nuclear ribonucleoprotein network. Biochim Biophys Acta 1966; 129:54-67; PMID:5970076; http://dx.doi.org/ 10.1016/0005-2787(66)90008-6 [DOI] [PubMed] [Google Scholar]
- [114].He DC, Nickerson JA, Penman S. Core filaments of the nuclear matrix. J Cell Biol 1990; 110:569-80; PMID:2307700; http://dx.doi.org/ 10.1083/jcb.110.3.569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Fey EG, Ornelles DA, Penman S. Association of RNA with the cytoskeleton and the nuclear matrix. J Cell Sci Suppl 1986; 5:9-119; PMID:NOT_FOUND10444063 [DOI] [PubMed] [Google Scholar]
- [116].Ma H, Siegel AJ, Berezney R. Association of chromosome territories with the nuclear matrix. Disruption of human chromosome territories correlates with the release of a subset of nuclear matrix proteins. J Cell Biol 1999; 146:531-42; PMID:10444063; http://dx.doi.org/ 10.1083/jcb.146.3.531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Nickerson J. Experimental observations of a nuclear matrix. J Cell Sci 2001; 114:463-74; PMID:11171316 [DOI] [PubMed] [Google Scholar]
- [118].Cook PR, Brazell IA, Jost E. Characterization of nuclear structures containing superhelical DNA. J Cell Sci 1976; 22:303-24; PMID:1002771 [DOI] [PubMed] [Google Scholar]
- [119].Birkenmeier EH, Gwynn B, Howard S, Jerry J, Gordon JI, Landschulz WH, McKnight SL. Tissue-specific expression, developmental regulation, and genetic mapping of the gene encoding CCAAT/enhancer binding protein. Genes & Dev. 1989; 3:1146-1156; PMID:2792758 [DOI] [PubMed] [Google Scholar]
- [120].Vogelstein B, Pardoll DM, Coffey DS. Supercoiled loops and eucaryotic DNA replicaton. Cell 1980; 22:79-85; PMID:7428042; http://dx.doi.org/ 10.1016/0092-8674(80)90156-7 [DOI] [PubMed] [Google Scholar]
- [121].Linnemann AK, Platts AE, Krawetz SA. Differential nuclear scaffold/matrix attachment marks expressed genes. Hum Mol Genet 2009; 18:645-54; PMID:19017725; http://dx.doi.org/ 10.1093/hmg/ddn394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Barone JG, De Lara J, Cummings KB, Ward WS. DNA organization in human spermatozoa. J Androl 1994; 15:139-44; PMID:8056637 [PubMed] [Google Scholar]
- [123].Bode J, Kohwi Y, Dickinson L, Joh T, Klehr D, Mielke C, Kohwi-Shigematsu T. Biological significance of unwinding capability of nuclear matrix-associating DNAs. Science 1992; 255:195-7; PMID:1553545; http://dx.doi.org/ 10.1126/science.1553545 [DOI] [PubMed] [Google Scholar]
- [124].Mirkovitch J, Spierer P, Laemmli UK. Genes and loops in 320,000 base-pairs of the Drosophila melanogaster chromosome. J Mol Biol 1986; 190:255-8; PMID:3098982; http://dx.doi.org/ 10.1016/0022-2836(86)90296-2 [DOI] [PubMed] [Google Scholar]
- [125].Benyajati C, Worcel A. Isolation, characterization, and structure of the folded interphase genome of Drosophila melanogaster. Cell 1976; 9:393-407; PMID:825231; http://dx.doi.org/ 10.1016/0092-8674(76)90084-2 [DOI] [PubMed] [Google Scholar]
- [126].Dickinson LA, Joh T, Kohwi Y, Kohwi-Shigematsu T. A tissue-specific MAR/SAR DNA-binding protein with unusual binding site recognition. Cell 1992; 70:631-45; PMID:1505028; http://dx.doi.org/ 10.1016/0092-8674(92)90432-C [DOI] [PubMed] [Google Scholar]
- [127].Boulikas T. Chromatin domains and prediction of MAR sequences. Int Rev Cytol 1995; 162A:279-388; PMID:8575883 [DOI] [PubMed] [Google Scholar]
- [128].Alvarez JD, Yasui DH, Niida H, Joh T, Loh DY, Kohwi-Shigematsu T. The MAR-binding protein SATB1 orchestrates temporal and spatial expression of multiple genes during T-cell development. Genes Dev 2000; 14:521-35; PMID:10716941 [PMC free article] [PubMed] [Google Scholar]
- [129].Fey EG, Penman S. Nuclear matrix proteins reflect cell type of origin in cultured human cells. Proc Natl Acad Sci U S A 1988; 85:21-5; PMID:3277171; http://dx.doi.org/ 10.1073/pnas.85.1.121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [130].Getzenberg RH, Pienta KJ, Huang EY, Coffey DS. Identification of nuclear matrix proteins in the cancer and normal rat prostate. Cancer Res 1991; 51:514-20; PMID:1898713 [PubMed] [Google Scholar]
- [131].Compton DA, Szilak I, Cleveland DW. Primary structure of NuMA, an intranuclear protein that defines a novel pathway for segregation of proteins at mitosis. J Cell Biol 1992; 116:1395-408; PMID:1541636; http://dx.doi.org/ 10.1083/jcb.116.6.1395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Yang CH, Lambie EJ, Snyder M. NuMA: an unusually long coiled-coil related protein in the mammalian nucleus. J Cell Biol 1992; 116:1303-17; PMID:1541630; http://dx.doi.org/ 10.1083/jcb.116.6.1303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Gueth-Hallonet C, Wang J, Harborth J, Weber K, Osborn M. Induction of a regular nuclear lattice by overexpression of NuMA. Exp Cell Res 1998; 243:434-52; PMID:9743603; http://dx.doi.org/ 10.1006/excr.1998.4178 [DOI] [PubMed] [Google Scholar]
- [134].Harborth J, Wang J, Gueth-Hallonet C, Weber K, Osborn M. Self assembly of NuMA: multiarm oligomers as structural units of a nuclear lattice. Embo J 1999; 18:689-700; http://dx.doi.org/ 10.1093/emboj/18.6.1689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Luderus ME, den Blaauwen JL, de Smit OJ, Compton DA, van Driel R. Binding of matrix attachment regions to lamin polymers involves single-stranded regions and the minor groove. Mol Cell Biol 1994; 14:6297-305; PMID:8065361; http://dx.doi.org/ 10.1128/MCB.14.9.6297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [136].Abad PC, Lewis J, Mian IS, Knowles DW, Sturgis J, Badve S, Xie J, Lelievre SA. NuMA influences higher order chromatin organization in human mammary epithelium. Mol Biol Cell 2007; 18:348-61; PMID:17108325; http://dx.doi.org/ 10.1091/mbc.E06-06-0551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Reyes JC, Muchardt C, Yaniv M. Components of the human SWI/SNF complex are enriched in active chromatin and are associated with the nuclear matrix. J Cell Biol 1997; 137:263-74; PMID:9128241; http://dx.doi.org/ 10.1083/jcb.137.2.263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Saitoh N, Goldberg IG, Wood ER, Earnshaw WC. ScII: an abundant chromosome scaffold protein is a member of a family of putative ATPases with an unusual predicted tertiary structure. J Cell Biol 1994; 127:303-18; PMID:7929577; http://dx.doi.org/ 10.1083/jcb.127.2.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [139].Cook PR. The organization of replication and transcription. Science 1999; 284:1790-1795; PMID:10364545; http://dx.doi.org/ 10.1126/science.284.5421.1790 [DOI] [PubMed] [Google Scholar]
- [140].Berezney R, Mortillaro MJ, Ma H, Wei X, Samarabandu J. The nuclear matrix: a structural milieu for genomic function. Int Rev Cytol 1995; 162A:1-65; PMID:8575878 [DOI] [PubMed] [Google Scholar]
- [141].Sutherland H, Bickmore WA. Transcription factories: gene expression in unions? Nat Rev Genet 2009; 10:457-66; PMID:19506577; http://dx.doi.org/ 10.1038/nrg2592 [DOI] [PubMed] [Google Scholar]
- [142].Tanaka T, Akira S, Yoshida K, Umemoto M, Yoneda Y, Shirafuji N, Fujiwara H, Suematsu S, Yoshida N, Kishimoto T. Targeted disruption of the NF-IL6 gene discloses its essential role in bacteria killing and tumor cytotoxicity by macrophages. Cell 1995; 80:353-361; PMID:7530603; http://dx.doi.org/ 10.1016/0092-8674(95)90418-2 [DOI] [PubMed] [Google Scholar]
- [143].Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol 2006; 7:885-96; PMID:17139329; http://dx.doi.org/ 10.1038/nrm2066 [DOI] [PubMed] [Google Scholar]
- [144].Susperreguy S, Prendes LP, Desbats MA, Charo NL, Brown K, MacDougald OA, Kerppola T, Schwartz J, Piwien-Pilipuk G. Visualization by BiFC of different C/EBPbeta dimers and their interaction with HP1alpha reveals a differential subnuclear distribution of complexes in living cells. Exp Cell Res 2011; 317:706-23; PMID:21122806; http://dx.doi.org/ 10.1016/j.yexcr.2010.11.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [145].Eskiw CH, Rapp A, Carter DR, Cook PR. RNA polymerase II activity is located on the surface of protein-rich transcription factories. J Cell Sci 2008; 121:1999-2007; PMID:18495842; http://dx.doi.org/ 10.1242/jcs.027250 [DOI] [PubMed] [Google Scholar]
- [146].Xu M, Cook PR. Similar active genes cluster in specialized transcription factories. J Cell Biol 2008; 181:615-23; PMID:18490511; http://dx.doi.org/ 10.1083/jcb.200710053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [147].Cook PR. A model for all genomes: the role of transcription factories. J Mol Biol 2010; 395:1-10; PMID:19852969; http://dx.doi.org/ 10.1016/j.jmb.2009.10.031 [DOI] [PubMed] [Google Scholar]
- [148].Cesena TI, Cardinaux JR, Kwok R, Schwartz J. CCAAT/enhancer-binding protein (C/EBP) beta is acetylated at multiple lysines: acetylation of C/EBPbeta at lysine 39 modulates its ability to activate transcription. J Biol Chem 2007; 282:956-67; PMID:17110376; http://dx.doi.org/ 10.1074/jbc.M511451200 [DOI] [PubMed] [Google Scholar]
- [149].Wochnik GM, Ruegg J, Abel GA, Schmidt U, Holsboer F, Rein T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J Biol Chem 2005; 280:4609-16; PMID:15591061; http://dx.doi.org/ 10.1074/jbc.M407498200 [DOI] [PubMed] [Google Scholar]
- [150].Gallo LI, Ghini AA, Piwien Pilipuk G, Galigniana MD. Differential recruitment of tetratricorpeptide repeat domain immunophilins to the mineralocorticoid receptor influences both heat-shock protein 90-dependent retrotransport and hormone-dependent transcriptional activity. Biochemistry 2007; 46:14044-57; PMID:18001136; http://dx.doi.org/ 10.1021/bi701372c [DOI] [PubMed] [Google Scholar]
- [151].Lefterova MI, Zhang Y, Steger DJ, Schupp M, Schug J, Cristancho A, Feng D, Zhuo D, Stoeckert CJ Jr., Liu XS, et al.. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev 2008; 22:941-52; http://dx.doi.org/ 10.1101/gad.1709008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Nielsen R, Pedersen TA, Hagenbeek D, Moulos P, Siersbaek R, Megens E, Denissov S, Borgesen M, Francoijs KJ, Mandrup S, et al.. Genome-wide profiling of PPARgamma:RXR and RNA polymerase II occupancy reveals temporal activation of distinct metabolic pathways and changes in RXR dimer composition during adipogenesis. Genes Dev 2008; 22:2953-67; PMID:18981474; http://dx.doi.org/ 10.1101/gad.501108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [153].Steger DJ, Grant GR, Schupp M, Tomaru T, Lefterova MI, Schug J, Manduchi E, Stoeckert CJ Jr., Lazar MA. Propagation of adipogenic signals through an epigenomic transition state. Genes Dev 2010; 24:1035-44; PMID:20478996; http://dx.doi.org/ 10.1101/gad.1907110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [154].Siersbaek R, Nielsen R, John S, Sung MH, Baek S, Loft A, Hager GL, Mandrup S. Extensive chromatin remodelling and establishment of transcription factor ‘hotspots’ during early adipogenesis. Embo J 2011; 30:1459-72; PMID:21427703; http://dx.doi.org/ 10.1038/emboj.2011.65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [155].Siersbaek R, Nielsen R, Mandrup S. PPARgamma in adipocyte differentiation and metabolism–novel insights from genome-wide studies. FEBS Lett 2010; 584:3242-9; PMID:20542036; http://dx.doi.org/ 10.1016/j.febslet.2010.06.010 [DOI] [PubMed] [Google Scholar]
- [156].Luger K, Dechassa ML, Tremethick DJ. New insights into nucleosome and chromatin structure: an ordered state or a disordered affair? Nat Rev Mol Cell Biol 2012; 13:436-47; PMID:22722606; http://dx.doi.org/ 10.1038/nrm3382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [157].Waldeyer W. Karyokinese und ihre Bezlehung zu den Befruchtungsvorgängen Archiv für mikroskopische. Anatomie 1888; 32:1-22; PMID:NOT_FOUND499376 [Google Scholar]
- [158].Boveri T. Die Blastomerenkerne von Ascaris megalocephala und die Theorie der Chromosomen individualität. Archiv Zellforsch 1909; 3:181-268; PMID:NOT_FOUND499376 [Google Scholar]
- [159].Zorn C, Cremer C, Cremer T, Zimmer J. Unscheduled DNA synthesis after partial UV irradiation of the cell nucleus. Distribution in interphase and metaphase. Exp Cell Res 1979; 124:111-9; PMID:499376; http://dx.doi.org/ 10.1016/0014-4827(79)90261-1 [DOI] [PubMed] [Google Scholar]
- [160].Cremer T, Cremer M. Chromosome territories. Cold Spring Harb Perspect Biol 2010; 2:a003889; PMID:20300217; http://dx.doi.org/ 10.1101/cshperspect.a003889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [161].Habermann FA, Cremer M, Walter J, Kreth G, von Hase J, Bauer K, Wienberg J, Cremer C, Cremer T, Solovei I. Arrangements of macro- and microchromosomes in chicken cells. Chromosome Res 2001; 9:569-84; PMID:11721954; http://dx.doi.org/ 10.1023/A:1012447318535 [DOI] [PubMed] [Google Scholar]
- [162].Tanabe H, Muller S, Neusser M, von Hase J, Calcagno E, Cremer M, Solovei I, Cremer C, Cremer T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc Natl Acad Sci U S A 2002; 99:4424-9; PMID:11930003; http://dx.doi.org/ 10.1073/pnas.072618599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Federico C, Scavo C, Cantarella CD, Motta S, Saccone S, Bernardi G. Gene-rich and gene-poor chromosomal regions have different locations in the interphase nuclei of cold-blooded vertebrates. Chromosoma 2006; 115:123-8; PMID:16404627; http://dx.doi.org/ 10.1007/s00412-005-0039-z [DOI] [PubMed] [Google Scholar]
- [164].Neusser M, Schubel V, Koch A, Cremer T, Muller S. Evolutionarily conserved, cell type and species-specific higher order chromatin arrangements in interphase nuclei of primates. Chromosoma 2007; 116:307-20; PMID:17318634; http://dx.doi.org/ 10.1007/s00412-007-0099-3 [DOI] [PubMed] [Google Scholar]
- [165].Croft JA, Bridger JM, Boyle S, Perry P, Teague P, Bickmore WA. Differences in the localization and morphology of chromosomes in the human nucleus. J Cell Biol 1999; 145:1119-31; PMID:10366586; http://dx.doi.org/ 10.1083/jcb.145.6.1119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [166].Cremer M, von Hase J, Volm T, Brero A, Kreth G, Walter J, Fischer C, Solovei I, Cremer C, Cremer T. Non-random radial higher-order chromatin arrangements in nuclei of diploid human cells. Chromosome Res 2001; 9:51-67; http://dx.doi.org/ 10.1023/A:1012495201697 [DOI] [PubMed] [Google Scholar]
- [167].Cremer M, Kupper K, Wagler B, Wizelman L, von Hase J, Weiland Y, Kreja L, Diebold J, Speicher MR, Cremer T. Inheritance of gene density-related higher order chromatin arrangements in normal and tumor cell nuclei. J Cell Biol 2003; 162:809-20; PMID:12952935; http://dx.doi.org/ 10.1083/jcb.200304096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [168].Bridger JM, Boyle S, Kill IR, Bickmore WA. Re-modelling of nuclear architecture in quiescent and senescent human fibroblasts. Curr Biol 2000; 10:49-52; http://dx.doi.org/ 10.1016/S0960-9822(00)00312-2 [DOI] [PubMed] [Google Scholar]
- [169].Bolzer A, Kreth G, Solovei I, Koehler D, Saracoglu K, Fauth C, Muller S, Eils R, Cremer C, Speicher MR, et al.. Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol 2005; 3:e157; PMID:15839726; http://dx.doi.org/ 10.1371/journal.pbio.0030157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [170].Berezney R, Buchholtz LA. Isolation and characterization of rat liver nuclear matrices containing high molecular weight deoxyribonucleic acid. Biochemistry 1981; 20:4995-5002; PMID:7295663; http://dx.doi.org/ 10.1021/bi00520a028 [DOI] [PubMed] [Google Scholar]
- [171].Shelby RD, Hahn KM, Sullivan KF. Dynamic elastic behavior of alpha-satellite DNA domains visualized in situ in living human cells. J Cell Biol 1996; 135:545-57; PMID:8909532; http://dx.doi.org/ 10.1083/jcb.135.3.545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [172].Zink D, Cremer T, Saffrich R, Fischer R, Trendelenburg MF, Ansorge W, Stelzer EH. Structure and dynamics of human interphase chromosome territories in vivo. Hum Genet 1998; 102:241-51; PMID:9521598; http://dx.doi.org/ 10.1007/s004390050686 [DOI] [PubMed] [Google Scholar]
- [173].Malhas A, Lee CF, Sanders R, Saunders NJ, Vaux DJ. Defects in lamin B1 expression or processing affect interphase chromosome position and gene expression. J Cell Biol 2007; 176:593-603; PMID:17312019; http://dx.doi.org/ 10.1083/jcb.200607054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [174].Zuleger N, Robson MI, Schirmer EC. The nuclear envelope as a chromatin organizer. Nucleus 2011; 2:339-49; PMID:21970986; http://dx.doi.org/ 10.4161/nucl.2.5.17846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [175].Solovei I, Wang AS, Thanisch K, Schmidt CS, Krebs S, Zwerger M, Cohen TV, Devys D, Foisner R, Peichl L, et al.. LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate differentiation. Cell 2013; 152:584-98; PMID:23374351; http://dx.doi.org/ 10.1016/j.cell.2013.01.009 [DOI] [PubMed] [Google Scholar]
- [176].Kuroda M, Tanabe H, Yoshida K, Oikawa K, Saito A, Kiyuna T, Mizusawa H, Mukai K. Alteration of chromosome positioning during adipocyte differentiation. J Cell Sci 2004; 117:5897-903; PMID:15537832; http://dx.doi.org/ 10.1242/jcs.01508 [DOI] [PubMed] [Google Scholar]
- [177].Eneroth M, Mandahl N, Heim S, Willen H, Rydholm A, Alberts KA, Mitelman F. Localization of the chromosomal breakpoints of the t(12;16) in liposarcoma to subbands 12q13.3 and 16p11.2. Cancer Genet Cytogenet 1990; 48:101-7; PMID:2372777; http://dx.doi.org/ 10.1016/0165-4608(90)90222-V [DOI] [PubMed] [Google Scholar]
- [178].Aman P, Ron D, Mandahl N, Fioretos T, Heim S, Arheden K, Willen H, Rydholm A, Mitelman F. Rearrangement of the transcription factor gene CHOP in myxoid liposarcomas with t(12;16)(q13;p11). Genes Chromosomes Cancer 1992; 5:278-85; PMID:1283316; http://dx.doi.org/ 10.1002/gcc.2870050403 [DOI] [PubMed] [Google Scholar]
- [179].Crozat A, Aman P, Mandahl N, Ron D. Fusion of CHOP to a novel RNA-binding protein in human myxoid liposarcoma. Nature 1993; 363:640-4; PMID:8510758; http://dx.doi.org/ 10.1038/363640a0 [DOI] [PubMed] [Google Scholar]
- [180].Rabbitts TH, Forster A, Larson R, Nathan P. Fusion of the dominant negative transcription regulator CHOP with a novel gene FUS by translocation t(12;16) in malignant liposarcoma. Nat Genet 1993; 4:15-80; http://dx.doi.org/ 10.1038/ng0693-175 [DOI] [PubMed] [Google Scholar]
- [181].Kuroda M, Wang X, Sok J, Yin Y, Chung P, Giannotti JW, Jacobs KA, Fitz LJ, Murtha-Riel P, Turner KJ, et al.. Induction of a secreted protein by the myxoid liposarcoma oncogene. Proc Natl Acad Sci U S A 1999; 96:025-30; PMID:NOT_FOUND10654081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [182].Kozubek S, Lukasova E, Mareckova A, Skalnikova M, Kozubek M, Bartova E, Kroha V, Krahulcova E, Slotova J. The topological organization of chromosomes 9 and 22 in cell nuclei has a determinative role in the induction of t(9,22) translocations and in the pathogenesis of t(9,22) leukemias. Chromosoma 1999; 108:426-35; PMID:10654081; http://dx.doi.org/ 10.1007/s004120050394 [DOI] [PubMed] [Google Scholar]
- [183].Neves H, Ramos C, da Silva MG, Parreira A, Parreira L. The nuclear topography of ABL, BCR, PML, and RARalpha genes: evidence for gene proximity in specific phases of the cell cycle and stages of hematopoietic differentiation. Blood 1999; 93:197-207; PMID:NOT_FOUND12575798 [PubMed] [Google Scholar]
- [184].Bickmore WA, Teague P. Influences of chromosome size, gene density and nuclear position on the frequency of constitutional translocations in the human population. Chromosome Res 2002; 10:707-15; PMID:12575798; http://dx.doi.org/ 10.1023/A:1021589031769 [DOI] [PubMed] [Google Scholar]
- [185].Meaburn KJ, Misteli T, Soutoglou E. Spatial genome organization in the formation of chromosomal translocations. Semin Cancer Biol 2007; 17:0-90; http://dx.doi.org/ 10.1016/j.semcancer.2006.10.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [186].Szczerbal I, Foster HA, Bridger JM. The spatial repositioning of adipogenesis genes is correlated with their expression status in a porcine mesenchymal stem cell adipogenesis model system. Chromosoma 2009; 118:647-63; PMID:19585140; http://dx.doi.org/ 10.1007/s00412-009-0225-5 [DOI] [PubMed] [Google Scholar]
- [187].Tong Q, Dalgin G, Xu H, Ting CN, Leiden JM, Hotamisligil GS. Function of GATA transcription factors in preadipocyte-adipocyte transition. Science 2000; 290:134-8; PMID:11021798; http://dx.doi.org/ 10.1126/science.290.5489.134 [DOI] [PubMed] [Google Scholar]
- [188].Dekker J, Rippe K, Dekker M, Kleckner N. Capturing chromosome conformation. Science 2002; 295:1306-11; PMID:11847345; http://dx.doi.org/ 10.1126/science.1067799 [DOI] [PubMed] [Google Scholar]
- [189].Simonis M, Klous P, Splinter E, Moshkin Y, Willemsen R, de Wit E, van Steensel B, de Laat W. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nat Genet 2006; 38:1348-54; PMID:17033623; http://dx.doi.org/ 10.1038/ng1896 [DOI] [PubMed] [Google Scholar]
- [190].Zhao Z, Tavoosidana G, Sjolinder M, Gondor A, Mariano P, Wang S, Kanduri C, Lezcano M, Sandhu KS, Singh U, et al.. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nat Genet 2006; 38:1341-7; PMID:17033624; http://dx.doi.org/ 10.1038/ng1891 [DOI] [PubMed] [Google Scholar]
- [191].Dostie J, Richmond TA, Arnaout RA, Selzer RR, Lee WL, Honan TA, Rubio ED, Krumm A, Lamb J, Nusbaum C, et al.. Chromosome Conformation Capture Carbon Copy (5C): a massively parallel solution for mapping interactions between genomic elements. Genome Res 2006; 16:1299-309; PMID:16954542; http://dx.doi.org/ 10.1101/gr.5571506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [192].Lieberman-Aiden E, van Berkum NL, Williams L, Imakaev M, Ragoczy T, Telling A, Amit I, Lajoie BR, Sabo PJ, Dorschner MO, et al.. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009; 326:289-93; PMID:19815776; http://dx.doi.org/ 10.1126/science.1181369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [193].Nagano T, Lubling Y, Stevens TJ, Schoenfelder S, Yaffe E, Dean W, Laue ED, Tanay A, Fraser P. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature 2013; 502:59-64; PMID:24067610; http://dx.doi.org/ 10.1038/nature12593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [194].Dixon JR, Selvaraj S, Yue F, Kim A, Li Y, Shen Y, Hu M, Liu JS, Ren B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012; 485:376-80; PMID:22495300; http://dx.doi.org/ 10.1038/nature11082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [195].Nora EP, Lajoie BR, Schulz EG, Giorgetti L, Okamoto I, Servant N, Piolot T, van Berkum NL, Meisig J, Sedat J, et al.. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature 2012; 485:381-5; PMID:22495304; http://dx.doi.org/ 10.1038/nature11049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [196].Gorkin DU, Leung D, Ren B. The 3D genome in transcriptional regulation and pluripotency. Cell Stem Cell 2014; 14:62-75; http://dx.doi.org/ 10.1016/j.stem.2014.05.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [197].Sofueva S, Yaffe E, Chan WC, Georgopoulou D, Vietri Rudan M, Mira-Bontenbal H, Pollard SM, Schroth GP, Tanay A, Hadjur S. Cohesin-mediated interactions organize chromosomal domain architecture. Embo J 2013; 32:119-29; http://dx.doi.org/ 10.1038/emboj.2013.237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [198].Merkenschlager M, Odom DT. CTCF and cohesin: linking gene regulatory elements with their targets. Cell 2013; 152:1285-97; PMID:23498937; http://dx.doi.org/ 10.1016/j.cell.2013.02.029 [DOI] [PubMed] [Google Scholar]
- [199].Zuin J, Dixon JR, van der Reijden MI, Ye Z, Kolovos P, Brouwer RW, van de Corput MP, van de Werken HJ, Knoch TA, van IWF, et al.. Cohesin and CTCF differentially affect chromatin architecture and gene expression in human cells. Proc Natl Acad Sci U S A 2013; 111:996-1001; PMID:24335803; http://dx.doi.org/ 10.1073/pnas.1317788111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [200].Van Bortle K, Corces VG. Nuclear organization and genome function. Annu Rev Cell Dev Biol 2012; 28:163-87; PMID:22905954; http://dx.doi.org/ 10.1146/annurev-cellbio-101011-155824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [201].Jin F, Li Y, Dixon JR, Selvaraj S, Ye Z, Lee AY, Yen CA, Schmitt AD, Espinoza CA, Ren B. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature 2013; 503:290-4; PMID:24141950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [202].LeBlanc SE, Wu Q, Barutcu AR, Xiao H, Ohkawa Y, Imbalzano AN. The PPARgamma locus makes long-range chromatin interactions with selected tissue-specific gene loci during adipocyte differentiation in a protein kinase A dependent manner. PLoS One 2014; 9:e86140; PMID:24465921; http://dx.doi.org/ 10.1371/journal.pone.0086140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [203].Xiao H, Leblanc SE, Wu Q, Konda S, Salma N, Marfella CG, Ohkawa Y, Imbalzano AN. Chromatin accessibility and transcription factor binding at the PPARgamma2 promoter during adipogenesis is protein kinase A-dependent. J Cell Physiol 2011; 226:86-93; PMID:20625991; http://dx.doi.org/ 10.1002/jcp.22308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [204].Salma N, Xiao H, Mueller E, Imbalzano AN. Temporal recruitment of transcription factors and SWI/SNF chromatin-remodeling enzymes during adipogenic induction of the peroxisome proliferator-activated receptor gamma nuclear hormone receptor. Mol Cell Biol 2004; 24:4651-63; PMID:15143161; http://dx.doi.org/ 10.1128/MCB.24.11.4651-4663.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [205].Loft A, Forss I, Siersbaek MS, Schmidt SF, Larsen AS, Madsen JG, Pisani DF, Nielsen R, Aagaard MM, Mathison A, et al.. Browning of human adipocytes requires KLF11 and reprogramming of PPARgamma superenhancers. Genes Dev 2014; 29:7-22; PMID:25504365; http://dx.doi.org/ 10.1101/gad.250829.114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [206].Whyte WA, Orlando DA, Hnisz D, Abraham BJ, Lin CY, Kagey MH, Rahl PB, Lee TI, Young RA. Master transcription factors and mediator establish super-enhancers at key cell identity genes. Cell 2013; 153:307-191. [DOI] [PMC free article] [PubMed] [Google Scholar]