

Abstract
Background
Within the last 10 years Zika virus (ZIKV) has caused unprecedented epidemics of human disease in the nations and territories of the western Pacific and South America, and continues to escalate in both endemic and non-endemic regions. We evaluated the vector competence of Australian mosquitoes for ZIKV to assess their potential role in virus transmission.
Methodology/Principal Findings
Mosquitoes were exposed to infectious blood meals containing the prototype African ZIKV strain. After 14 days incubation at 28°C and high relative humidity, infection, dissemination and transmission rates were assessed. Infection in Culex annulirostris and Cx. sitiens could not be detected. 8% of Cx. quinquefasciatus were infected, but the virus did not disseminate in this species. Despite having infection rates > 50%, Aedes notoscriptus and Ae. vigilax did not transmit ZIKV. In contrast, Ae. aegypti had infection and transmission rates of 57% and 27%, respectively. In susceptibility trials, the virus dose required to infect 50% (ID50) of Ae. aegypti was106.4 tissue culture infectious dose50 (TCID50)/mL. Additionally, a threshold viral load within the mosquito of at least 105.1 TCID50 equivalents/mL had to be reached before virus transmission occurred.
Conclusions/Significance
We confirmed Ae. aegypti to be the most likely mosquito vector of ZIKV in Australia, although the restricted distribution of this species will limit the receptive zone to northern Queensland where this species occurs. Importantly, the role in ZIKV transmission of Culex and other Aedes spp. tested will be negligible. Despite being the implicated vector, the relatively high ID50 and need for a high titer disseminated infection in Ae. aegypti suggest that high mosquito population densities will be required to facilitate epidemic ZIKV transmission among the currently immunologically naïve human population in Australia.
Author Summary
Zika virus was first isolated in Uganda in 1947 and exists in a transmission cycle between mosquitoes and non-human primates or humans. Whilst most clinical infections result in a self-limiting febrile illness, Zika virus has recently been linked to neurological syndromes, such as Guillain-Barré syndrome and congenital birth defects. Since 2007, Zika virus has undergone a dramatic range expansion, causing epidemics in nations and territories of the western Pacific and South America. To assess the emergence and transmission risk of Zika virus emerging in Australia, we evaluated the ability of local mosquitoes to become infected with and transmit the prototype African Zika virus strain. In agreement with its substantiated role in Zika virus transmission overseas, Australian Aedes aegypti were shown to be competent vectors. Coupled with its anthropophilic feeding behavior, this species should be considered the primary potential Zika virus vector in Australia. Although other common Australian species, such as Ae. notoscriptus and Ae. vigilax, were readily infected, they did not transmit the virus. The species of Culex tested were either refractory to infection or had a low infection rate. We also demonstrated that the Zika virus dose necessary to infect Ae. aegypti was higher than virus levels reported in infected humans. Finally, a high threshold level of virus circulating through the mosquito body was required before Ae. aegypti transmitted the virus. These results suggest that an outbreak of Zika virus in Australia would require high mosquito population densities and a susceptible human population.
Introduction
Zika virus (ZIKV) was first isolated from a rhesus monkey in the Zika Forest of Uganda in 1947 during studies investigating the ecology of yellow fever virus [1]. It was subsequently isolated from Aedes africanus, indicating a mosquito-borne transmission cycle. It was not until almost 20 years later that human clinical disease attributed to ZIKV infection was recognized [2]. ZIKV circulates in a sylvatic transmission cycle between non-human primates and mosquitoes. Humans are only incidentally infected in this sylvatic cycle, but are the primary amplifying hosts during epidemics [3, 4]. Although 80% of infections remain asymptomatic, clinical disease caused by ZIKV is typical of many mosquito-borne viruses, and is characterized by fever, muscle and joint pain, headache, conjunctivitis, gastrointestinal manifestations and rash [2, 5, 6]. However, during its rapid expansion in the last 5 years, more severe disease manifestations have been recognized among those with Zika virus–like disease, most notably neurological symptoms, including Guillain-Barré syndrome, microcephaly and other central nervous system malformation in neonates [7, 8].
Based on serological evidence, ZIKV infection of humans has historically been restricted to Africa and Asia [9]. In 2007, an outbreak of ZIKV occurred on Yap Island in the western Pacific Ocean [5], signaling the beginning of an unprecedented range expansion of this virus. Since 2013, ZIKV has affected a number of countries and territories in the western Pacific, resulting in outbreaks of hundreds or thousands of suspected cases [10]. The 2013 epidemic in French Polynesia was the largest ever reported up to that point and resulted in an estimated 32,000 cases, representing 11.5% of the population [11]. In 2014 the virus was introduced into Brazil, from where it is believed to have subsequently spread to a number of countries in South America, causing over a million suspected cases [10, 12]. Between 2007 and June 2 2016, ZIKV was recognized in 63 countries or territories [13].
The risk of ZIKV spread to Australia is very high due to its close geographical proximity to the epidemic region in the western Pacific and high intake of travelers from this area. Indeed, between 2012 and June 3 2016, 60 people infected with ZIKV have traveled to Australia [14]. Should a viremic patient be bitten by a local mosquito, or transmit the virus sexually [15], there is potential for autochthonous transmission of the virus to occur, but this has not been reported to date. Clinical similarity of ZIKV disease with that caused by arboviruses, such as dengue (DENV) and chikungunya (CHIKV), could delay ZIKV identification or detection of transmission, and there are currently no established vaccines or therapeutics. Therefore, it is very important to estimate the risk of ZIKV establishment in Australia for the purposes of heightened public awareness, and for adequate and appropriate response of public health authorities.
In the current study we evaluated the ability of common Australian mosquito species to become infected with and transmit the African lineage of ZIKV. Although the Asian lineage is responsible for the recent activity in the Pacific and South America, we were not able to obtain an isolate to facilitate the assessment of the vector competence of local mosquito fauna for this ZIKV lineage in a manner that was timely for formulation of targeted control strategies. The sylvan vectors circulating ZIKV between primates in Africa are tree-hole inhabiting Aedes spp., of which Ae. africanus is the most important [4]. The primary urban vectors are Ae. aegypti and Ae. albopictus [9]. Of the species implicated abroad, only Ae. aegypti occurs on the Australian mainland, albeit with a distribution restricted to northern Queensland [16]. Ae. albopictus is currently restricted to the Torres Strait islands off northern Australia [17] and was not tested in the current experiments. In terms of other potential vectors, Australia has a number of other species that could potentially transmit ZIKV. For instance, Ae. notoscriptus is a widespread urban species throughout Australia and was shown to be a competent laboratory vector of yellow fever virus [18]. Another potential species is Ae. vigilax, which has a widespread coastal distribution, is a notorious biter and a highly competent laboratory vector of CHIKV [19]. Finally, it has been postulated that Culex spp., most notably Cx. quinquefasciatus, may play a role in transmission in South America [20].
Materials and Methods
Mosquitoes
Mosquitoes were collected as eggs or adults from several locations in Queensland. Adult Ae. vigilax, Ae. procax, Cx. annulirostris and Cx. sitiens were collected using Centers for Disease Control light traps (Model 512, John Hock Co., Gainesville, FL) baited with CO2 (1kg dry ice) from several suburbs in Brisbane, southeastern Queensland. Adult mosquitoes were transported to the laboratory and exposed to ZIKV within 5 h of collection.
The Ae. aegypti used were F4 generation females from eggs collected using ovitraps from Townsville, northern Queensland in March 2015. Eggs of Ae. notoscriptus and Cx. quinquefasciatus were collected using ovitraps and infusion buckets, respectively, from several suburbs in Brisbane. All larvae were reared at 26°C and 12:12 L:D. Both Aedes spp. were fed Hikari Cichlid Staple pellets (Kyorin Co. Ltd, Himeji, Japan). First and second instar larvae of Cx. quinquefasciatus were fed a 1:1 mixture of brewer’s yeast (Brewer’s Yeast, Healthy Life) and fish flakes (Wardley’s Tropical Fish Food Flakes, The Hartz Mountain Corporation, NJ), whilst third and fourth instars were fed Hikari Cichlid Staple pellets (Kyorin co. Ltd, Himeji, Japan). Adults were held for 3–7 d at 26°C 12:12 L:D, and high relative humidity, and fed on 15% honey water ad libitum. Mosquitoes were starved for 24 h prior to virus exposure.
Virus strain
The ZIKV strain (MR 766) was the prototype strain isolated from a rhesus macaque monkey in the Zika Forest, Uganda in 1947. The virus was sourced from the American Type Culture Collection (Manassas, VA, USA). It had been passaged 146 times in adult mouse brain, once in suckling mouse brain and three times in in African green monkey kidney (Vero) cells.
Exposure of mosquitoes to ZIKV
Mosquitoes were allowed to feed for 2 h on an infectious blood meal containing stock virus diluted in commercially available defibrinated sheep blood (Institute of Medical and Veterinary Science, Adelaide, Australia). The blood meal was housed within a Hemotek feeding apparatus (Discovery Workshops, Accrington, Lancashire, UK) that was fitted with pig intestine as the membrane. Due to sufficient numbers and relatively high feeding rates, Ae. aegypti and Ae. notoscriptus were exposed to serial 10-fold dilutions of ZIKV, with 106.7± 0.2 tissue culture infectious dose50 (TCID50)/mL the highest dose. The latter species was exposed to an additional blood feed at the highest dose. Due to limited numbers and low feeding rates, the other species were only exposed to blood meals containing a virus titer of 106.7± 0.2 TCID50/mL. Pre- and post- feeding samples of the blood/virus mixture were diluted 1:10 in growth media (GM; Opti-MEM (Gibco, Invitrogen Corporation, Grand Island, NY) containing 3% foetal bovine serum (FBS), antibiotics and antimycotics), and stored at -80°C.
Immediately following virus exposure, mosquitoes were anesthetized with CO2 and engorged mosquitoes were placed in 900 ml gauze-covered containers. All mosquitoes were maintained on 15% honey water at 28°C, high relative humidity and 12L:12D light cycle within an environmental growth cabinet.
Assessment of infection, dissemination and transmission
Mosquitoes were processed at day 14 post-exposure. Due to sufficient numbers, Ae. aegypti were also processed at 5, 7 and 10 d post exposure, whilst the additional Ae. notoscriptus were processed at day 7 post exposure. The ability for mosquitoes to become infected with and transmit ZIKV was assessed using a modified in vitro capillary tube technique [21]. Briefly, mosquitoes were anesthetized with CO2, and the legs and wings removed. The saliva was collected by inserting the proboscis of the mosquito into a capillary tube containing GM with 20% FBS. After 30 min, the contents of the capillary tube were expelled into 500 μl of GM with 3% FBS. The legs+wings and bodies were placed in separate 2 mL tubes containing 1 mL of GM with 3% FBS and a single 5 mm stainless steel bead. Detection of virus in the legs+wings indicates that the mosquito has developed an infection whereby the virus has escaped the midgut and disseminated throughout the hemocoel, thus bypassing the midgut escape barrier [22]. All samples were stored at -80°C.
Virus assays
Titration of initial blood/virus mixtures
Blood/virus mixtures were titrated as serial ten-fold dilutions in 96 well microtiter plates seeded with confluent monolayers of C6/36 cells. Plates were incubated at 28°C for 10 d before being fixed in PBS/acetone and stored at -20°C. Infection with ZIKV was detected using a cell culture enzyme immunoassay (CC-EIA; [23]) and the pan-flavivirus reactive monoclonal antibody 4G2 (provided by Roy Hall, University of Queensland, Australia).
Detection of ZIKV in mosquito components and saliva expectorates
Bodies and legs+wings were homogenized separately using a QIAGEN TissueLyser II (Qiagen, Hilden, Germany) and centrifuged at 14,000g for 5 min. Viral RNA was extracted from 140 μl of each homogenate and the saliva expectorates using the Qiagen BioRobot Universal System and QIAamp Virus BioRobot MDx Kit (Qiagen, Clifton Hill, Australia). All samples were analyzed for ZIKV RNA using a real-time TaqMan RT-PCR designed in the nonstructural protein 5 (NS5) gene. Primer and dual-labeled probe sequences (genome nucleotide positions corresponding to ZIKV MR766, GenBank accession number AY632535) were as follows: forward primer (Zika –F-2007) 5’-9845CCTCAAGGATGGGAGATCCA9864-3’, reverse primer (Zika-R-2007) 5’-9908 AGCTCGGCCAATCAGTTCAT9889-3’ and probe (Zika-FAM-2007) 5’FAM-9868 TGGTCCCTTGCCGCCACCA9886 TAMRA-3’. Primer and probe oligonucleotides were synthesized by Sigma-Aldrich (Australia). Separate synthetic primer and probe oligonucleotides were also designed to prepare in vitro transcribed RNA positive assay controls. The synthetic assay control sequences incorporated T7 promoter (bold), ZIKV TaqMan primer or probe (italics) and ubiquitin-conjugating enyzyme E2 D2 (UBE2D2) housekeeping gene (underlined) sequences (protocol adapted from Smith et al. [24]). The primer assay control (NS5-Zika-synPri) 5’-AAAATAATACGACTCACTATAGGG CCTCAAGGATGGGAGATCCAATGATCTGGCACGGGACCCTCCAA ATGAACTGATTGGCCGAGCT-3’ and probe assay control (NS5-Zika-synPro) 5’-AAAATAATACGACTCACTATAGGGTGAAGAGAATCCACAAGGAATTGAA TGGTCCCTTGCCGCCACCAACAGTGTTCAGCAGGTCCTGTTG-3’ synthetic assay control oligonucleotides were synthesized by GeneWorks Pty. Ltd. (Adelaide, Australia).
In vitro RNA transcription of 2–5μg of each synthetic assay control was performed for one hour at 37°C in a 100μl volume using the Riboprobe System-T7 kit (Promega, United States). Each RNA preparation was subjected to DNase treatment, DNase inactivation (TURBO DNA-free kit, Life Technologies, Unites States) and RNA purification using a spin column, according to the manufacturer’s instructions (RNeasy Mini Kit, QIAGEN, Australia). The DNase treatment, inactivation and RNA purification process was conducted twice to ensure sufficient reduction of amplifiable template DNA.
The ZIKV-NS5 real-time RT-PCR was performed using a Rotor-Gene 6000 real-time PCR machine (QIAGEN, Australia). Amplification of ZIKV RNA and detection of the 64 bp product took place in a 20 μL single-tube, Superscript III Platinum one-step qRT-PCR (Invitrogen, Carlsbad, CA). Reactions contained 0.4 μL Superscript III RT/Platinum Taq mix, 10.0 μL of 2X reaction mix, 650 nM primers, 250 nM dual-labeled probe, 50 nM ROX reference dye, and 5 μL of extracted sample RNA or serially diluted extracted ZIKV MR 766 RNA. Each amplification run included positive primer control (5 μl synthetic primer assay control RNA) and probe control (5 μl synthetic probe assay control RNA) reactions as previously described [24]. The cycling conditions consisted of one cycle at 50°C for 5 min, one cycle at 95°C for 2 min, and 40 cycles at 95°C for 3 s and 60°C for 30 s. The threshold cycle number (Ct) was determined for each sample and a Ct value ≥40 cycles was chosen to indicate no RNA detection.
The ZIKV stock was titrated in 96 well microtiter plates seeded with Vero African green monkey cells (ATCC, CCL-81) and the CC-EIA was used to determine TCID50/mL values [25]. Viral RNA was extracted as above and ten-fold dilutions were used to generate standard curves for the ZIKV-NS5 real-time RT-PCR. Each dilution (10−4–10−9) was run in duplicate and presented as a mean using Rotor-Gene software. Standard curves were plotted as Ct values versus –log TCID50/mL and were used to calculate ZIKV TCID50 equivalents/mL in each sample (S1 Fig).
Statistical analysis
The susceptibility of Ae. aegypti and Ae. notoscriptus to infection with ZIKV was calculated by probit analysis using SPSS release 16.0.0. Log-log models were assessed using the Pearson χ2 goodness-of-fit statistic and susceptibility to infection was expressed as ID50 ± 95% confidence intervals (CIs) and defined as the virus dose per mL at which 50% of mosquitoes tested positive for ZIKV infection in the TaqMan RT-PCR. Infection, dissemination and transmission rates between Ae. aegypti and the other species were analyzed using Fisher’s Exact Tests with two-tailed P-values (GraphPad Prism Version 6). ZIKV TCID50/mL equivalents were tested for differences between species, and within Ae. aegypti using the Kruskal-Wallis test (GraphPad Prism Version 6).
Results
Susceptibility of Ae. aegypti and Ae. notoscriptus to ZIKV infection
To assess their susceptibility to infection, Ae. aegypti and Ae. notoscriptus were exposed to ZIKV doses ranging from 103.9 to 106.8 TCID50/mL and their bodies tested for infection 14 d post exposure (Fig 1). Both species were susceptible to infection, with ID50s of 106.4 (106.0 and 107.1, 95% CL) TCID50/mL (χ2 = 2.49, df = 2, P = 0.288) and 106.6 (106.2 and 107.4, 95% CL) TCID50/mL (χ2 = 8.49, df = 2, P = 0.654) for Ae. aegypti and Ae. notoscriptus, respectively.
Fig 1. Susceptibility of Ae. aegypti and Ae. notoscriptus to ZIKV infection.

Percent infection rates in Ae. aegypti (circles) and Ae. notoscriptus (triangles) exposed to serial dilutions of ZIKV and tested at 14 d post-exposure.
Infection, dissemination and transmission rates in Australian mosquitoes
Five out of seven species, including all species of Aedes, were infected 14 days after being exposed to blood meals containing 106.7 ± 0.2 TCID50/mL of ZIKV (Table 1). Out of the 3 Culex spp. tested, Cx. annulirostris and Cx. sitiens were refractory to infection and only 2 out of 30 Cx. quinquefasciatus were infected but none developed a disseminated infection. On day 14 post exposure, infection rates in Ae. notoscriptus, Ae. procax and Ae. vigilax did not significantly differ (P > 0.05) from Ae. aegypti. With the exception of one cohort of Ae. notoscriptus, dissemination rates in these three species did not significantly differ (P > 0.05) from Ae. aegypti. However, Ae. aegypti was the only species that was able to transmit ZIKV, with transmission first observed at day 10 post exposure.
Table 1. Infection, dissemination and transmission rates in seven Australian mosquito species exposed to a blood meal containing 106.7 ± 0.2 TCID50/mL of ZIKV.
| Species | Day PE | % infectiona | % disseminationb | % dissemination/ infectionc | % transmissiond | % transmission/ disseminatione | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ae. aegypti | 5 | 40 | (10/25) | 8 | (2/25) | 20 | (2/10) | 0 | (0/25) | 0 | (0/0) |
| 7 | 52 | (13/25) | 36 | (9/25) | 69 | (9/13) | 0 | (0/25) | 0 | (0/0) | |
| 10 | 40 | (10/25) | 28 | (7/25) | 70 | (7/10) | 12 | (3/25) | 43 | (3/7) | |
| 14 | 57 | (17/30) | 40 | (12/30) | 71 | (12/17) | 27 | (8/30) | 67 | (8/12) | |
| Ae. notoscriptus | 7 | 72 | (18/25) | 8 | (2/25)* | 11 | (2/18)* | 0 | (0/25) | 0 | (0/0) |
| 14 | 60 | (18/30) | 0 | (0/30)* | 0 | (0/0) | 0 | (0/30)* | 0 | (0/0) | |
| 14 | 53 | (16/30) | 20 | (6/30) | 38 | (6/16) | 0 | (0/30)* | 0 | (0/0) | |
| Ae. procax | 14 | 33 | (2/6) | 17 | (1/6) | 50 | (1/2) | 0 | (0/6) | 0 | (0/0) |
| Ae. vigilax | 14 | 57 | (17/30) | 27 | (8/30) | 47 | (8/17) | 0 | (0/30)* | 0 | (0/0) |
| Cx. annulirostris | 14 | 0 | (0/30)* | 0 | (0/30)* | 0 | (0/0) | 0 | (0/30)* | 0 | (0/0) |
| Cx. quinquefasciatus | 14 | 7 | (2/30)* | 0 | (0/30)* | 0 | (0/0) | 0 | (0/30)* | 0 | (0/0) |
| Cx. sitiens | 14 | 0 | (0/11)* | 0 | (0/11)* | 0 | (0/0) | 0 | (0/11) | 0 | (0/11) |
aPercentage of mosquitoes containing virus in their bodies (number positive/number tested).
bPercentage of mosquitoes containing virus in their legs+wings (number positive/number tested).
cPercentage of infected mosquitoes containing virus in their legs+wings (number positive/number infected).
dPercentage of mosquitoes containing virus in the saliva expectorates (number positive/number tested).
ePercentage of mosquitoes with a disseminated infection containing virus in the saliva expectorates (number positive/number disseminated).
*Fisher’s exact test two-tailed P-value <0.05 for comparisons with Ae. aegypti.
Relative quantification of ZIKV in Australian mosquitoes
ZIKV titers in the mosquito bodies did not significantly differ among the four Aedes spp. tested (Fig 2). In contrast there was a significant difference (P = 0.0014) in the ZIKV titers in the legs+wings between these species. Importantly, transmission of ZIKV by Ae. aegypti on day 14 post exposure occurred only when the titer of ZIKV in the legs+wings was ≥ 105.6 TCID50 equivalents/mL, with all individuals at or above this threshold able to transmit ZIKV.
Fig 2. Replication of ZIKV in Australian species of Aedes.

The titer in TCID50 equivalents per mL of ZIKV in the bodies (A) and legs+wings (B) of four species of Aedes tested 14 days after ingesting an infectious blood meal containing 106.7± 0.2 TCID50/mL of ZIKV. Ae. notoscriptus A and B represents the two different blood feeds for this species. Each point on the plot represents an individual mosquito, and horizontal lines denote medians. Ae. aegypti had significantly higher (P < 0.001) legs+wings titer than Ae. notoscriptus B. The horizontal dashed line represents the threshold titer above which virus transmission occurred.
For the Ae.aegypti, there was an increase in the estimated virus titer on the different days tested post exposure, although the differences were not significant (P > 0.05; Fig 3A). Similarly, there was an increase in legs+wings titer over the various days, with the difference between days 5 and 14 being significant (P < 0.05; Fig 3B). The three mosquitoes that transmitted ZIKV on day 10 post exposure possessed legs+wings titers that were lower than the 105.6 TCID50 equivalents/mL required for transmission on day 14. The titer of the saliva expectorated did not significantly differ (P > 0.05) between mosquitoes sampled on days 10 and 14 post exposure (Fig 3C).
Fig 3. Replication of ZIKV in Ae. aegypti on different days following exposure to an infectious blood meal.
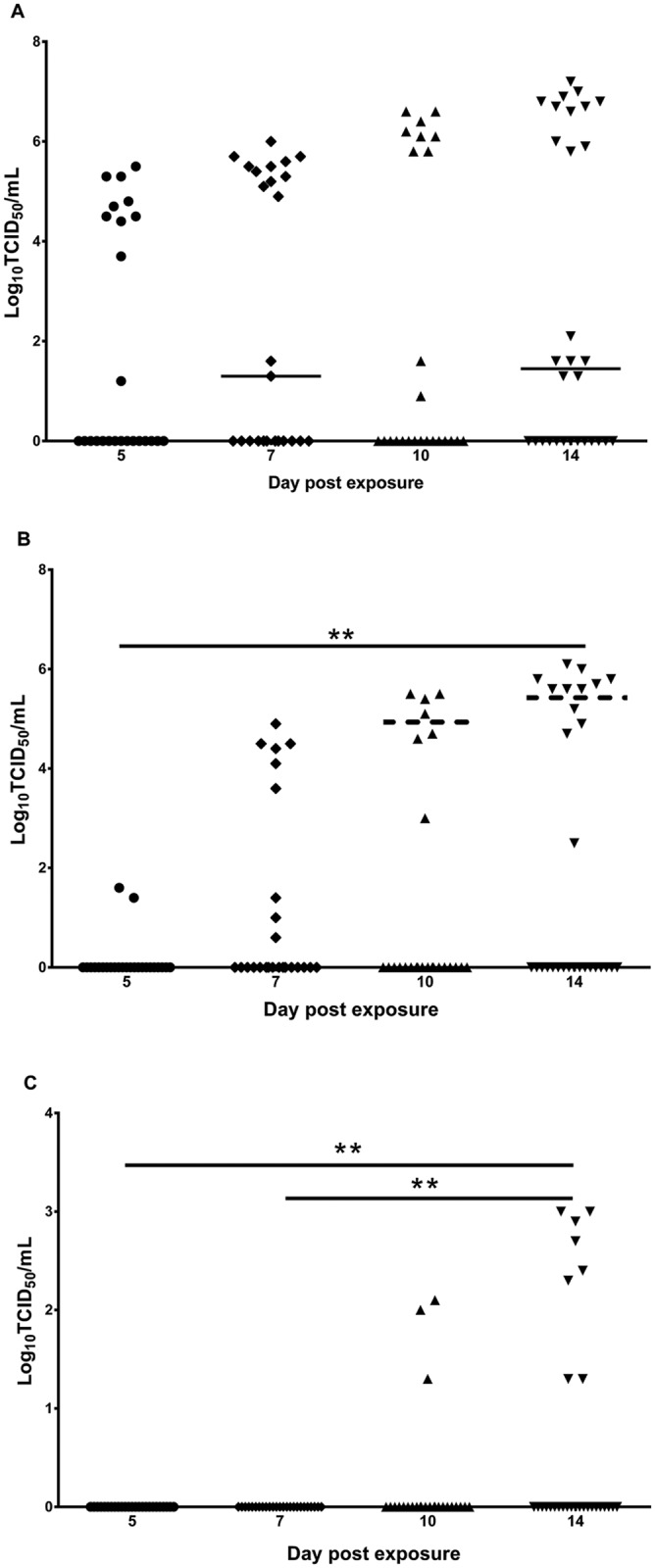
The titer in TCID50 equivalents/mL of ZIKV in the bodies (A), legs+wings (B) and saliva (C) of Ae. aegypti and tested on different days after ingesting an infectious blood meal containing 106.7± 0.2 TCID50/mL of ZIKV. Each point on the plot represents an individual mosquito, and bars denote medians. Solid lines represent significant differences P < 0.001 (**) between days for legs+wings and saliva titers. The horizontal dashed lines on days 10 and 14 represent the threshold titer above which virus transmission occurred.
Discussion
Commensurate with its role in transmission in Africa, the western Pacific and South America, we have demonstrated that a Townsville, Australia, population of Ae. aegypti is susceptible to infection and can transmit ZIKV. Transmission rates were higher than those recently reported for Brazilian and Senegalese strains of Ae. aegypti that were exposed to Asian and African lineages of ZIKV, respectively, but lower than those for a Singapore strain infected with MR 766 [26–28]. However, comparison of experimental vector competence outcomes between different laboratories should be viewed with caution due to differences in mosquito strain, virus strain, mosquito feeding method and virus assay used to analyze samples [29]. Regardless, we have confirmed that highly infected Ae. aegypti could be a potential source of transmission of ZIKV in Australia. Although Ae. aegypti was prevalent in eastern Australia during the first half of the 20th century, the distribution of this species is currently restricted to urban areas of northern Queensland [16]. Therefore, should the virus be introduced, this is the region that is most at risk of ZIKV transmission. However, if the distribution of Ae. aegypti expanded, or if Ae. albopictus, another potential vector [26], invaded the Australian mainland, then the ZIKV receptive zone would need to be redefined. This highlights the importance of surveillance for these container-inhabiting species to ensure that any expansion of these species is recognized and elimination programs initiated.
In addition to the intrinsic ability of Ae. aegypti to transmit ZIKV, this species exhibits a number of biological traits which dramatically elevate its role as a vector of this virus, as well as other viruses transmitted by this species. Jansen et al. [30] used a relatively simple vectorial capacity model [31] to assess the relative roles of Australian mosquitoes in the transmission of CHIKV. Vectorial capacity takes into account a number of factors, including mosquito density, host feeding patterns, survival, vector competence and the duration of the intrinsic incubation period of the virus. It was demonstrated that Ae. aegypti had the highest vectorial capacity even though other species, such as Ae. vigilax, had higher experimental transmission rates [19] and population densities compared to Ae. aegypti ([32]; Queensland Health, state government data). The reason for this was that a high proportion of Ae. aegypti obtain their blood meals from humans [33, 34], and they exhibit multiple host blood feeding behavior [35], whereby females probe/feed up to four times in a single gonotrophic cycle. This latter trait exposes them to more infected humans increasing the likelihood of consuming virus, as well as increasing the number of susceptible humans exposed to an infected mosquito. The majority of other mosquito species usually take only a single blood meal per gonotrophic cycle.
To assess the potential risk for local ZIKV transmission in Australia, we exposed 6 other mosquito species to ZIKV and evaluated their ability to transmit the virus. The results demonstrated that the Culex spp. were either refractory to infection or exhibited a low infection rate but did not transmit ZIKV, suggesting that they should not be considered potential vectors of ZIKV in Australia. Despite being susceptible to infection, the inability of Ae. notoscriptus, Ae. vigilax and Ae. procax to transmit the virus, indicate that these species would be unlikely to play a role in ZIKV transmission. However, given the potential for intraspecies variation in vector competence of arboviruses [36, 37], it is important that the vector competence of other Australian populations of these species for ZIKV be assessed using the prototype strain, as well as the Asian lineage viral variant(s) currently circulating in the western Pacific and South America, and recently imported into Africa.
The current study has highlighted aspects of the susceptibility, replication and transmission dynamics of ZIKV in the mosquito vector that may ultimately impact transmission cycles in the field. We demonstrated that the ID50 for Ae. aegypti was 106.4 TCID50/mL, which is considerably higher than that observed for other arboviruses in Australian populations of this species, including DENVs (≈ 105.5 TCID50/mL) and CHIKV (104.9 TCID50/mL) [19, 38]. The relatively low infection and transmission rates reported for some populations of Ae. aegypti [26, 27] would suggest a similarly high threshold for infection in this species. Paradoxically, the high ID50 required to infect Ae. aegypti in our study is potentially higher than the viremia values of 105 to 106 RNA copies per mL circulating in the blood of symptomatic patients during recent outbreaks in the western Pacific [39, 40]. This may simply reflect the differences inherent to quantifying infectious virus and viral RNA or it may hint at factors other than low susceptibility to infection of the mosquito vector. Such factors include high mosquito population density and high survival rates of infected mosquitoes, coupled with a naïve human population suffering high viral loads, and alternative modes of transmission, such as sexual transmission, all of which may contribute to epidemic transmission of ZIKV.
There are a number of intrinsic barriers that can influence the ability of an arbovirus to infect, disseminate within and be transmitted by mosquitoes [41] and the species tested in our study expressed one or more of these barriers. Based on their refractoriness to infection or low infection rate, it appears that the three species of Culex tested possess a midgut infection barrier. Similar to what has been observed previously with DENVs and CHIKV [19, 42], Ae. notoscriptus expresses a midgut escape barrier, as only 10% of mosquitoes tested at day 14 had a disseminated infection. With the exception of Ae. aegypti, the remaining Aedes spp. tested appeared to have a salivary infection/transmission barrier, as none of the individuals with a disseminated infection subsequently transmitted ZIKV. Interestingly, within the Ae. aegypti cohort, it was only those mosquitoes with a high disseminated infection titer of ≥ 105.1 TCID50/mL in the legs+wings that transmitted the virus. This suggests that a considerable quantity of virus is required to overcome the salivary gland barriers to transmission. Conversely, the other Aedes spp. had lower legs+wings titers of ≤ 104.8 TCID50/mL and this may explain why they did not transmit the virus. A correlation between the leg titer and the percentage of mosquitoes transmitting has previously been shown for dengue virus type 1 (DENV-1), whereby a threshold titer had to be reached before transmission occurred [43]. It was concluded that the high dissemination titers led to increased transmissibility by Ae. aegypti, which potentially resulted in a DENV-1 clade replacement event in Thailand. Future studies, using strains of ZIKV from the current epidemics and a considerably larger sample size, should examine whether a similar correlation exists between different viral lineages, proportion of each mosquito species infected, dissemination titers and the transmission rate. Importantly, this will reveal whether only a relatively small proportion of the mosquito population is likely to be contributing to the majority of virus transmission.
In conclusion, Australia possesses the two key elements for ZIKV to occur: a competent ZIKV vector and importation of the virus by infected travelers. The Australian distribution of Ae. aegypti is currently restricted to north Queensland, so this is the region that is receptive to ZIKV. It is paramount that container mosquito surveillance is maintained and, when resources are available, enhanced to ensure that Ae. aegypti and Ae. albopictus do not expand their range. There is also a need to maintain comprehensive testing of travelers from epidemic regions to ensure that suspected cases are diagnosed, particularly those residing in the ZIKV receptive zone. Indeed, there have already been several cases notified in northern Queensland where Ae. aegypti occurs, although control strategies implemented routinely for DENVs [44] may have assisted in preventing local transmission of the virus to date. Unfortunately, tracking of ZIKV infected travelers is compounded by the high number of asymptomatic infections and local transmission may be occurring before ZIKV is tested for in human or mosquito populations. Ultimately, a combination of case recognition, specific laboratory diagnostics, virus surveillance in mosquito populations, vector surveillance to detect incursions of potential vectors into uninfested locations and targeted mosquito control strategies will reduce the risk of an explosive ZIKV outbreak occurring in Australia.
Supporting Information
The standard curve was prepared using RNA extracted from ZIKV MR 766 virus stock as template. The x-axis represents the concentration of duplicate, serially diluted template (0.01 to 10000 tissue culture infectious doses equivalents/ml) and the y-axis plots the resultant real-time RT-PCR Ct values.
(TIF)
Acknowledgments
We thank Bruce Harrower, Peter Burtonclay, Tanya Constantino and Sean Moody for technical assistance. We also thank Roy Hall for providing the 4G2 monoclonal antibody and Ina Smith for assistance in developing the Zika virus TaqMan RT-PCR.
Data Availability
All relevant data are within the paper.
Funding Statement
This study was funded by an internal Forensic and Scientific Services research and development project grant (number RSS15-003). S. Ritchie is funded by National Health and Medical Research Council Senior Research Fellowship 1044698. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Dick GW, Kitchen SF, Haddow AJ. Zika virus. I. Isolations and serological specificity. Trans R Soc Trop Med Hyg. 1952;46: 509–520. [DOI] [PubMed] [Google Scholar]
- 2.Simpson DI. Zika virus infection in man. Trans R Soc Trop Med Hyg. 1964; 58: 335–338. [PubMed] [Google Scholar]
- 3.Boorman JP, Porterfield JS. A simple technique for infection of mosquitoes with viruses; transmission of Zika virus. Trans R Soc Trop Med Hyg. 1956;50: 238–242. [DOI] [PubMed] [Google Scholar]
- 4.Haddow AJ, Williams MC, Woodall JP, Simpson DI, Goma LK. Twelve isolations of Zika virus from Aedes (Stegomyia) africanus (Theobald) taken in and above a Uganda forest. Bull World Health Organ. 1964;31: 57–69. [PMC free article] [PubMed] [Google Scholar]
- 5.Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, Lanciotti RS, et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N Engl J Med. 2009;360: 2536–2543. 10.1056/NEJMoa0805715 [DOI] [PubMed] [Google Scholar]
- 6.Olson JG, Ksiazek TG, Suhandiman, Triwibowo. Zika virus, a cause of fever in Central Java, Indonesia. Trans R Soc Trop Med Hyg. 1981;75: 389–393. [DOI] [PubMed] [Google Scholar]
- 7.Cao-Lormeau VM, Blake A, Mons S, Lastere S, Roche C, Vanhomwegen J, et al. Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: a case-control study. Lancet. 2016;387: 1531–1539. 10.1016/S0140-6736(16)00562-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rasmussen SA, Jamieson DJ, Honein MA, Petersen LR. Zika virus and birth defects—reviewing the evidence for causality. N Engl J Med. 2016;374: 1981–1987. 10.1056/NEJMsr1604338 [DOI] [PubMed] [Google Scholar]
- 9.Hayes EB. Zika virus outside Africa. Emerg Infect Dis. 2009;15: 1347–50. 10.3201/eid1509.090442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Musso D, Gubler DJ. Zika Virus. Clin Microbiol Rev. 2016;29: 487–524. 10.1128/CMR.00072-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mallet HP, Vial AL, Musso D. Bilan de l’épidémie à virus Zika en Polynésie française, 2013–2014. Bulletin D’Iinformation Sanitaires, Epidemiologiques et Statistiques. 2015;13: 1–5. [Google Scholar]
- 12.Petersen LR, Jamieson DJ, Powers AM, Honein MA. Zika virus. N Engl J Med. 2016;374: 1552–1563. 10.1056/NEJMra1602113 [DOI] [PubMed] [Google Scholar]
- 13.World Health Organization. Situation Report—Zika Virus Microcephaly Guillain-Barre Syndrome 2 June 2016. Geneva: World Health Organization, 2016.
- 14.Department of Health Australian Government. SITREP #32—Zika. Canberra: Department of Health, Australian Government, 2016. doi: 10.1177/1363459315619021 26621264 [DOI]
- 15.Foy BD, Kobylinski KC, Chilson Foy JL, Blitvich BJ, Travassos da Rosa A, Haddow AD, et al. Probable non-vector-borne transmission of Zika virus, Colorado, USA. Emerg Infect Dis. 2011;17: 880–882. 10.3201/eid1705.101939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Beebe NW, Cooper RD, Mottram P, Sweeney AW. Australia's dengue risk driven by human adaptation to climate change. PLoS Negl Trop Dis. 2009;3: e429 10.1371/journal.pntd.0000429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Beebe NW, Ambrose L, Hill LA, Davis JB, Hapgood G, Cooper RD, et al. Tracing the tiger: population genetics provides valuable insights into the Aedes (Stegomyia) albopictus invasion of the Australasian Region. PLoS Negl Trop Dis. 2013;7: e2361 10.1371/journal.pntd.0002361 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.van den Hurk AF, McElroy K, Pyke AT, McGee CE, Hall-Mendelin S, Day A, et al. Vector competence of Australian mosquitoes for yellow fever virus. Am J Trop Med Hyg. 2011;85: 446–451. 10.4269/ajtmh.2011.11-0061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.van den Hurk AF, Hall-Mendelin S, Pyke AT, Smith GA, Mackenzie JS. Vector competence of Australian mosquitoes for chikungunya virus. Vector-borne and Zoonotic Dis. 2010;10: 489–495. [DOI] [PubMed] [Google Scholar]
- 20.Prada P. Research indicates another common mosquito may be able to carry Zika. Thomson Reuters. 3 March 2016. Available: http://www.reuters.com/article/us-health-zika-brazil-idUSKCN0W52AW. Accessed June 7 2016.
- 21.Aitken THG. An in vitro feeding technique for artificially demonstrating virus transmission by mosquitoes. Mosquito News. 1977;37: 130–133. [Google Scholar]
- 22.Turell MJ, Gargan TP II, Bailey CL. Replication and dissemination of Rift Valley fever virus in Culex pipiens. Am J Trop Med Hyg. 1984;33: 176–181. [DOI] [PubMed] [Google Scholar]
- 23.Broom AK, Hall RA, Johansen CA, Oliveira N, Howard MA, Lindsay MD, et al. Identification of Australian arboviruses in inoculated cell cultures using monoclonal antibodies in ELISA. Pathology. 1998;30: 286–288. [DOI] [PubMed] [Google Scholar]
- 24.Smith G, Smith I, Harrower B, Warrilow D, Bletchly C. A simple method for preparing synthetic controls for conventional and real-time PCR for the identification of endemic and exotic disease agents. J Virol Methods. 2006;135: 229–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Reed LJ, Meunch H. A simple method for estimating fifty percent end points. Am J Hyg. 1938;27: 493–497. [Google Scholar]
- 26.Chouin-Carneiro T, Vega-Rua A, Vazeille M, Yebakima A, Girod R, Goindin D, et al. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl Trop Dis. 2016;10: e0004543 10.1371/journal.pntd.0004543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Diagne CT, Diallo D, Faye O, Ba Y, Faye O, Gaye A, et al. Potential of selected Senegalese Aedes spp. mosquitoes (Diptera: Culicidae) to transmit Zika virus. BMC Infect Dis. 2015;15: 492 10.1186/s12879-015-1231-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li MI, Wong PS, Ng LC, Tan CH. Oral susceptibility of Singapore Aedes (Stegomyia) aegypti (Linnaeus) to Zika virus. PLoS Negl Trop Dis. 2012;6: e1792 10.1371/journal.pntd.0001792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Knox TB, Kay BH, Hall RA, Ryan PA. Enhanced vector competence of Aedes aegypti (Diptera: Culicidae) from the Torres Strait compared with mainland Australia for dengue 2 and 4 viruses. J Med Entomol. 2003;40: 950–956. [DOI] [PubMed] [Google Scholar]
- 30.Jansen CC, Williams CR, van den Hurk AF. The usual suspects: comparison of the relative roles of potential urban chikungunya virus vectors in Australia. PLoS One. 2015;10: e0134975 10.1371/journal.pone.0134975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Garrett-Jones C. Prognosis for interruption of malaria transmission through assessment of the mosquito's vectorial capacity. Nature. 1964;204:1173–1175. [DOI] [PubMed] [Google Scholar]
- 32.Frances SP, MacKenzie DO, Rowcliffe KL, Corcoran SK. Comparative field evaluation of repellent formulations containing deet and IR3535 against mosquitoes in Queensland, Australia. J Am Mosq Control Assoc. 2009;25:511–513. [DOI] [PubMed] [Google Scholar]
- 33.Jansen CC, Webb CE, Graham GC, Craig SB, Zborowski P, Ritchie SA, et al. Blood sources of mosquitoes collected from urban and peri-urban environments in eastern Australia with species-specific molecular analysis of avian blood meals. Am J Trop Med Hyg. 2009;81: 849–857. 10.4269/ajtmh.2009.09-0008 [DOI] [PubMed] [Google Scholar]
- 34.Scott TW, Chow E, Strickman D, Kittayapong P, Wirtz RA, Lorenz LH, et al. Blood-feeding patterns of Aedes aegypti (Diptera: Culicidae) collected in a rural Thai village. J Med Entomol. 1993;30: 922–927. [DOI] [PubMed] [Google Scholar]
- 35.Jetten TH, Focks DA. Potential changes in the distribution of dengue transmission under climate warming. Am J Trop Med Hyg. 1997;57: 285–297. [DOI] [PubMed] [Google Scholar]
- 36.Tabachnick WJ, Wallis GP, Aitken TH, Miller BR, Amato GD, Lorenz L, et al. Oral infection of Aedes aegypti with yellow fever virus: geographic variation and genetic considerations. Am J Trop Med Hyg. 1985;34: 1219–1224. [DOI] [PubMed] [Google Scholar]
- 37.Ye YH, Ng TS, Frentiu FD, Walker T, van den Hurk AF, O'Neill SL, et al. Comparative susceptibility of mosquito populations in north Queensland, Australia to oral infection with dengue virus. Am J Trop Med Hyg. 2014;90: 422–430. 10.4269/ajtmh.13-0186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ritchie SA, Pyke AT, Hall-Mendelin S, Day A, Mores CN, Christofferson RC, et al. An explosive epidemic of DENV-3 in Cairns, Australia. PLoS One. 2013;8: e68137 10.1371/journal.pone.0068137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lanciotti RS, Kosoy OL, Laven JJ, Velez JO, Lambert AJ, Johnson AJ, et al. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg Infect Dis. 2008;14: 1232–1239. 10.3201/eid1408.080287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Waehre T, Maagard A, Tappe D, Cadar D, Schmidt-Chanasit J. Zika virus infection after travel to Tahiti, December 2013. Emerg Infect Dis. 2014;20: 1412–1414. 10.3201/eid2008.140302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Black WC IV, Bennett KE, Gorrochótegui-Escalante N, Barillas-Mury CV, Fernández-Salas I, de Lourdes Muñoz M, et al. Flavivirus susceptibility in Aedes aegypti. Arch Med Res. 2002;33: 379–388. [DOI] [PubMed] [Google Scholar]
- 42.Pyke AT, Moore PR, Taylor CT, Hall-Mendelin S, Cameron JN, Hewitson GR, et al. Highly divergent dengue virus type 1 genotype sets a new distance record. Sci Rep. 2016;6: 22356 10.1038/srep22356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lambrechts L, Fansiri T, Pongsiri A, Thaisomboonsuk B, Klungthong C, Richardson JH, et al. Dengue-1 virus clade replacement in Thailand associated with enhanced mosquito transmission. J Virol. 2012;86: 1853–61. 10.1128/JVI.06458-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Department of Health, Queensland Government. Queensland Dengue Management Plan 2015–2020. Brisbane: Department of Health, Queensland Government, 2015. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The standard curve was prepared using RNA extracted from ZIKV MR 766 virus stock as template. The x-axis represents the concentration of duplicate, serially diluted template (0.01 to 10000 tissue culture infectious doses equivalents/ml) and the y-axis plots the resultant real-time RT-PCR Ct values.
(TIF)
Data Availability Statement
All relevant data are within the paper.