

Abstract
Infertility and subfertility represent major problems in domestic animals and humans, and the majority of embryonic loss occurs during the first month of gestation that involves pregnancy recognition and conceptus implantation. The critical genes and physiological pathways in the endometrium that mediate pregnancy establishment and success are not well understood. In study one, predominantly Angus heifers were classified based on fertility using serial embryo transfer to select animals with intrinsic differences in pregnancy loss. In each of the four rounds, a single in vitro-produced, high-quality embryo was transferred into heifers on Day 7 postestrus and pregnancy was determined on Days 28 and 42 by ultrasound and then terminated. Heifers were classified based on pregnancy success as high fertile (HF), subfertile (SF), or infertile (IF). In study two, fertility-classified heifers were resynchronized and bred with semen from a single high-fertility bull. Blood samples were collected every other day from Days 0 to 36 postmating. Pregnancy rate was determined on Day 28 by ultrasound and was higher in HF (70.4%) than in heifers with low fertility (36.8%; SF and IF). Progesterone concentrations in serum during the first 20 days postestrus were not different in nonpregnant heifers and also not different in pregnant heifers among fertility groups. In study three, a single in vivo-produced embryo was transferred into fertility-classified heifers on Day 7 postestrus. The uteri were flushed on Day 14 to recover embryos, and endometrial biopsies were obtained from the ipsilateral uterine horn. Embryo recovery rate and conceptus length and area were not different among the heifer groups. RNA was sequenced from the Day 14 endometrial biopsies of pregnant HF, SF, and IF heifers (n = 5 per group) and analyzed by edgeR-robust analysis. There were 26 differentially expressed genes (DEGs) in the HF compared to SF endometrium, 12 DEGs for SF compared to IF endometrium, and three DEGs between the HF and IF endometrium. Several of the DEG-encoded proteins are involved in immune responses and are expressed in B cells. Results indicate that preimplantation conceptus survival and growth to Day 14 is not compromised in SF and IF heifers. Thus, the observed difference in capacity for pregnancy success in these fertility-classified heifers is manifest between Days 14 and 28 when pregnancy recognition signaling and conceptus elongation and implantation must occur for the establishment of pregnancy.
Keywords: endometrium; pregnancy; ruminants (cows, sheep, llama, camel); uterus
INTRODUCTION
Infertility and subfertility are important and pervasive problems in agricultural animals and humans. In ruminants, embryo mortality is a major factor affecting fertility and thus production and economic efficiency [1–4]. There is a high degree of pregnancy loss in cattle between fertilization and term, ranging from 40% to 56% [4, 5]. The majority of embryonic loss (70%–80%) occurs in the first 3 wk of pregnancy in cattle [4–8]. Embryo mortality is greater in nonlactating cows than heifers [6], and early pregnancy loss is even greater in lactating dairy cattle and can approach 40% [1, 9, 10]. Infertility and subfertility also impact the embryo transfer (ET) industry [11]. Mean survival rate to calving following transfer of in vivo derived embryos from superovulated donors ranges from 31% to 60% [12], whereas in vitro produced (IVP) embryo survival rate is lower and ranges from 30% to 40% [3, 12]. Failure of the embryo to survive and establish pregnancy is due to both embryonic and maternal factors [13]. Many of the pregnancy losses observed in natural or assisted pregnancies can be attributed to inadequate uterine receptivity, which can be defined as the physiological state of the uterus when conceptus growth and implantation for the establishment of pregnancy is possible [14]. Understanding how embryo development is controlled is critical for determining ways to reduce the high rates of early embryonic mortality.
After fertilization (Day 0), the zona pellucida-enclosed bovine embryo enters the uterus at the morula stage on Days 4–5 of gestation and develops into a blastocyst. The spherical blastocyst hatches from the zona pellucida on Days 7–10 and continues to grow, changing from spherical to ovoid in shape between Days 12 and 14 during a transitory phase preceding elongation, after which it is termed a conceptus (embryo and associated extra-embryonic membranes) [15]. The conceptus grows from about 2 mm in length on Day 13 of gestation to 6 mm on Day 14, and reaches 60 mm by Day 16 to 20 cm or more by Day 19 [16]. After Days 16–17, the time of maternal recognition of pregnancy in cattle, the elongating conceptus begins the processes of implantation and placentation [17]. In both cattle and sheep, blastocyst growth into a conceptus and subsequent elongation has not been achieved in vitro and requires transfer into the uterus [18]. Progesterone action via the endometrium of the uterus is critical for conceptus growth and elongation in sheep and cattle [19–21]. Dynamic changes in endometrial gene expression occur between Days 7 and 13 that are regulated by progesterone in nonpregnant and pregnant cattle and are associated with the onset of conceptus elongation [20–23]. A prevailing theory is that the gene expression changes modify the intrauterine milieu for support of the survival and growth of the blastocyst into an ovoid conceptus and then elongated filamentous conceptus. Uterine secretions in the lumen are not well defined in cattle, but are a complex mixture of proteins, amino acids, sugars, lipids, and ions derived from genes expressed in the endometrium as well as selective transport of components (amino acids, glucose, and albumin and other proteins) from maternal blood. Endometrial epithelial secretions are particularly important for conceptus survival and growth in ruminants because uterine gland knockout ewes display recurrent early pregnancy loss due to defects in conceptus survival and elongation [24, 25]. Conceptus elongation is particularly critical for production of interferon tau (IFNT) [26, 27], the pregnancy recognition signal, that acts on the endometrium to sustain continued production of progesterone by the ovary and regulates genes implicated in implantation and placentation [14, 17, 28, 29]. Inadequate elongation of the conceptus results in low IFNT production, inability to maintain the corpus luteum (CL), and early pregnancy loss [30]. Although much information is known about embryo development into a blastocyst from in vitro systems [31], posthatching blastocyst survival and growth as well as conceptus elongation remain underinvestigated. Available evidence supports an unequivocal role for progesterone-dependent endometrial secretions of the uterus as primary regulators of conceptus survival, growth, and development throughout pregnancy [20, 32]. However, the essential endometrial genes and secretions that mediate survival and growth of the blastocyst and conceptus remain largely unknown in cattle [33, 34].
One of the major impediments to research on the physiology and genetics of early pregnancy success in cattle is the lack of animals with defined high and low rates of early pregnancy loss. McMillan and Donnison [35] summarized a unique approach for experimentally identifying high and low fertility in dairy heifers based on early pregnancy success. Contemporary yearling heifers (n = 155) received two IVP embryos on six separate occasions during a 26-mo period. Sixty days after transfer, pregnancy and the number of fetuses were determined ultrasonically and then pregnancies were terminated with the process being repeated six times. That approach identified 25 heifers with high (76%) and low (11%) aggregate pregnancy rates. Of note, a failure in the mechanism involved in conceptus elongation and maternal recognition of pregnancy was suggested to be the cause of early pregnancy loss in the low-fertility heifers [35, 36]. A similar approach was used here to fertility classify beef heifers based on natural variation in early pregnancy success. A series of studies were then conducted on fertility-classified heifers to begin determining the physiological and genetic factors underpinning early pregnancy loss in beef cattle.
MATERIALS AND METHODS
Animals
All animal procedures were conducted in accordance with the Guide for the Care and Use of Agriculture Animals in Research and Teaching and approved by the Institutional Animal Care and Use Committees of the USDA-ARS Fort Keogh Livestock and Range Research Laboratory, Washington State University, and the University of Missouri.
In Vitro Production of Embryos (Study One)
Embryos were produced in vitro at the University of Florida to obtain blastocysts for transfer into recipients. Oocytes obtained from genetically undefined ovaries (n = 1699–2800 per replicate) were fertilized with frozen-thawed spermatozoa pooled from three bulls of various breeds. The total number of bulls used in the eight replicates was 20 and represented Angus (n = 7), Limousin (n = 3), Brangus (n = 3), Simmental (n = 2), unknown (n = 2), Hereford (n = 2), and Polled Hereford (n = 1) sires.
The procedures for in vitro production were as follows. Cumulus oocyte complexes were obtained by cutting the surface of each ovary with a scalpel and vigorously rinsing the ovary through a bath of oocyte collection medium: tissue culture medium-199 with Hank salts (Hyclone Laboratories Inc.), 2% (v/v) of either bovine steer serum (Pel-Freez) or adult bovine serum (Thermo Scientific HyClone), 2 U/ml heparin, 100 U/ml penicillin-G, 0.1 mg/ml streptomycin, and 1 mM glutamine. Groups of 10 cumulus oocyte complexes were matured in 50 μl droplets of oocyte maturation medium (tissue culture medium-199 with Earle salts [Invitrogen], 10% [v/v] bovine steer serum or adult bovine serum, 2 μg/ml estradiol 17β, 20 μg/ml bovine follicle stimulating hormone [Bioniche Life Sciences], 22 μg/ml sodium pyruvate, 50 μg/ml gentamicin sulfate, and 1 mM glutamine) covered with mineral oil for 20 h at 38.5°C and in a humidified atmosphere of 5% (v/v) CO2. Sperm were purified by centrifugation over a Percoll (GE Healthcare) gradient consisting of 500 μl of a solution of 45% (v/v) Percoll, diluted 1:1 in HEPES-synthetic oviduct fluid fertilization medium (SOF) on top of 500 μl of a solution of 90% (v/v) Percoll [37]. Up to 250 matured oocytes were fertilized in a 35-mm dish with Percoll-purified sperm (1.0 × 106/ml) for 8–10 h at 38.5°C in 1.7 ml of SOF [38]. Cumulus cells were denuded after fertilization by vortexing in 600 μl HEPES-TALP containing 10 000 U/ml hyaluronidase. Putative zygotes were then cultured in 50 μl microdrops of SOF-bovine embryo 2 (SOF-BE2) covered with mineral oil at 38.5°C in a humidified atmosphere of 5% (v/v) O2, 5% (v/v) CO2, and 90% (v/v) N2. The cleavage rate was assessed at Day 3 postfertilization.
At Day 4 or 5 postfertilization, embryos were prepared for transport to Montana by placement of advanced embryos in groups of ∼50 per tube in polypropylene-stoppered 10 × 63 mm polystyrene tubes containing 1 ml SOF-BE2 supplemented with 1 mM HEPES, pH 7.5, and 50 μM dithiothreitol. Tubes had been equilibrated overnight in a humidified incubator at 38.5°C and 5% O2, 5% CO2, and 90% N2. After addition of embryos to the modified SOF-BE2, ∼1.5 ml mineral oil was layered over the medium and tubes were placed in a Biotherm portable incubator (Cryologic) at 38.5°C. Embryos were hand carried to Montana by an airliner on Day 5 or 6. Embryos were transferred to fresh 50 μl drops of SOF-BE2 and cultured at 38.5°C in a humidified atmosphere of 5% (v/v) O2, 5% (v/v) CO2, and 90% (v/v) N2 until Day 7 (embryos shipped on Day 4) or were allowed to develop in the shipping tubes until Day 7 (embryos shipped on Day 5).
Embryo Transfer and Pregnancy Determination by Ultrasonography (Study One)
Crossbred beef heifers (n = 275 Angus × Polled Hereford) were identified from a single population of cows at the USDA-ARS, Fort Keogh Livestock and Range Research Station and reared on a typical replacement heifer feedlot ration. Six heifers were culled at weaning based on extremely small body size. As illustrated in Figure 1, the remaining heifers were subjected to serial estrous synchronization and ET to classify heifers into fertility categories based on pregnancy outcome. The estrous cycles of heifers were synchronized beginning at approximately 14 mo of age when heifers weighed 368 ± 2.8 kg and had a body condition score of 5.9 ± 0.1 (scale of 1 to 9: 1 = emaciated and 9 = obese) to prepare heifers for ET on Day 7 after estrus (Day 0). Estrus was synchronized using the PG-6d-CIDR (prostaglandin F2 alpha-6 days-controlled intravaginal drug releasing device) protocol that includes prostaglandin F2 alpha (PGF) administration (Lutalyse; Zoetis Animal Health) on Day −12, an injection of GnRH (100 μg intramuscularly, Factrel; Zoetis Animal Health) concurrent with a intravaginal progesterone insert (i.e., CIDR) on Day −9, CIDR removal and PGF on Day −3, and estrus on Day 0. Estrus detection patches (Estrotect; Rockway, Inc.) were affixed to the tail head of each heifer to aid in visual detection of estrus. Heifers were observed for signs of estrus three times a day beginning 24 h after CIDR removal. Heifers that did not exhibit standing estrus received GnRH (100 μg intramuscularly, Factrel; Zoetis Animal Health) on Day 0. During each ET round, heifers were randomly assigned to one of two groups on consecutive days to facilitate animal handling. All heifers received an IVP embryo of high quality (blastocyst or expanded blastocyst) produced at the University of Florida on Day 7 after estrus or induced ovulation by one of two experienced technicians. Ultrasonography was used to identify the side and presence of the CL before ET by a single technician using an Aloka SSD 3500V and 7.5 MHz convex transducer (Aloka). The embryo was placed in the uterine horn ipsilateral to the ovary containing a CL using standard nonsurgical techniques. Heifers without a CL on Day 7 did not receive an embryo.
FIG. 1.

Experimental design and results for classification of fertility in beef heifers using serial embryo transfer (ET). See text for detailed description of results. Each row of numbers will not add up to 269 because of occasional death loss or removal of heifers from the study for health reasons. Further, 37 heifers did not have a CL at least once, and five heifers did not have a CL on two occasions. CIDR, controlled intravaginal drug releasing device; GnRH, gonadotropin releasing hormone; PGF2α, prostaglandin F2 alpha.
Heifers were diagnosed for pregnancy on Days 28 and 44 using transrectal ultrasonography with an Aloka SSD 500V and 7.5 MHz (Day 28) or 5.0 MHz (Day 42) linear transducer. Viability was confirmed at each ultrasound by the presence of a fetal heartbeat. At the second pregnancy diagnosis on Day 44, heifers received a single injection of PGF and were allowed a minimum of 30 days to exhibit another estrous cycle before being synchronized for another round of ET. After three or four rounds of ET and based on Day 28 pregnancy success, heifers were classified based on Day 28 pregnancy success as high fertile (HF; 100%), subfertile (SF; 25%–33%), or infertile (IF; 0%) (Fig. 1).
Blood Collection and Progesterone Radioimmunoassay for the Initial Round of ET (Study One)
Blood was collected from coccygeal vessels of heifers at the onset of synchronization (Days −22 and −12) for the initial round of ET to assess serum concentration of progesterone as an indicator of pubertal status. Blood samples were allowed to clot for 1 h at room temperature and then incubated at 4°C for approximately 24 h. Samples were centrifuged at 1200 × g for 25 min at 4°C, and serum was collected and stored at −20°C until radioimmunoassays (RIAs) were performed. The RIA was performed on serum samples to measure progesterone concentrations using the Coat a Count RIA kit (Siemens) [39]. Inter- and intra-assay CV for a sample with 1.5 ng/ml were 11.45% and 11.79%, respectively, and assay sensitivity was 0.08 ng/ml.
Estrous Synchronization and Artificial Insemination (Study Two)
Estrous synchronization and pregnancy determination.
Estrous cycles of fertility-classified heifers were synchronized using the PG-6d-CIDR protocol described in study one. Artificial insemination (AI) was performed at 12 and 24 h after onset of standing estrus with semen from a single bull of known high fertility. Pregnancy diagnosis was performed by transrectal ultrasonography (SonoSite EDGE equipped with a L52 10.0-5.0 MHz linear-array transducer; SonoSite Inc.) on Days 28 and 36 post-AI.
Blood sampling and serum progesterone determination.
Blood samples for determination of circulating progesterone concentrations were obtained by coccygeal venipuncture using evacuated tubes without anticoagulant (Becton Dickinson Vacutainer Systems) at the time of breeding (Day 0) and every other day from Days 0 to 28. Blood samples from heifers determined to be pregnant at Day 28 were also collected every other day from Day 28 to 36. Serum tubes were allowed to clot overnight at 4°C and then centrifuged at 3000 × g for 20 min at 4°C; serum was collected and frozen at −20°C until analyzed. Serum concentrations of progesterone were determined in triplicate 100 μl aliquots of sample using manufacturer (MP Biomedical) reagents and recommendations for the liquid-liquid phase double antibody precipitation assay (07-170105; MP Biomedicals) and validated as previously described [40]. Inter- and intra-assay coefficients of variation were <10%.
Embryo Transfer and Nonsurgical Uterine Flush on Day 14 (Study Three)
Synchronization and ET.
Estrous cycles of donor cows (n = 7) and fertility-classified heifers were tightly synchronized to optimize synchrony of embryos with the uterine environment of the recipient heifers. All females received PGF on Day −18 to presynchronized estrous cycles and increase ovulation response (follicular wave reset) to GnRH administration on Day −15. On Day −9, all females received an intravaginal progesterone insert (i.e., CIDR) to supplement blood progesterone to assist preparation of the uterus for pregnancy and an injection of 2.5 mg estradiol and 50 mg progesterone to regress all follicles larger than 0.5 mm and again reset follicular wave growth. Donor cows received twice daily injections of follicle stimulating hormone (Folltropin; Agtech, Inc.) in decreasing daily doses from Day −4.5 to −1 to stimulate multiple follicular development. The CIDRs were removed and PGF administered on Day −2.5 (recipients) and −1.5 (donors) to synchronize the estrus. Donors were inseminated 12 and 24 h after onset of estrus (Days 0 and 0.5) with semen from the same sire. Recipient heifers that did not exhibit estrus by Day 0 received GnRH and estrogen (1 mg estradiol benzoate). On Day 7, each heifer received an in vivo-produced embryo placed in the uterine horn ipsilateral to the ovary containing a CL (as determined by ultrasound) using standard nonsurgical techniques by a single technician. Ultrasonography was used to identify the side and presence of the CL before ET. Three heifers did not receive embryos due to excess fluid in their uterus or lack of a CL on their ovaries. Transferred embryos were of excellent quality and either a blastocyst (n = 53) or morula (n = 8) produced through superovulation of seven donor cows. All transfers were frozen-thawed direct transfer embryos blocked by donor and assigned to heifers equally across fertility classification group.
Nonsurgical uterine flush.
The uterus of each heifer was flushed to recover embryos at 7 days posttransfer using nonsurgical techniques. Briefly, a two-way luer-lock catheter (19982/0104; Minitube of America) was passed into the uterine horn ipsilateral to the CL and 20 ml of flush medium (Vigro complete flush medium; Agtech, Inc.) was placed into the uterine horn. After approximately 30 sec of gentle massage, the initial flush medium was recovered in a syringe, transferred to a Petri dish, and searched for a conceptus. If a conceptus was not observed in the initial flush, additional medium (1 L) was subsequently used to thoroughly flush the uterine horn. The flushes were accumulated and filtered through a 75 μm mesh nylon filter (VCI Filter; Agtech, Inc.) then rinsed with flush medium and searched again for an embryo.
Endometrial biopsy.
Following uterine flush, an endometrial biopsy was collected near the greater curvature of the uterine horn ipsilateral to the ovary containing the CL. Endometrial tissue was obtained using Jackson uterine biopsy forceps (GerMedUSA Inc.). The forcep was sterilized prior to biopsy collection using a hot bead sterilizer (Fine Science Tools Inc.). To collect the endometrial biopsy, the forcep was protected with a sanitary chemise (IMV Technologies), which was broken immediately before the biopsy tool was passed through the cervical os. The tip of the forceps was directed to the greater curvature of the ipsilateral uterine horn, and endometrial tissue (biopsy) was collected by closing the instrument jaw. The forcep was then removed, and the biopsy sections were immediately placed in 1.5 ml microcentrifuge tubes, frozen with liquid nitrogen, and stored at −80°C until RNA extraction.
Conceptus morphology and measurement.
Embryos were imaged on a Zeiss Discovery V8 stereomicroscope with an AxioCam ICC 1 and AxioVision version 4.6 software. Measurements of conceptus size (length and width) were collected using ImageJ (version 1.48; National Institutes of Health).
RNA Sequencing
RNA isolation.
Total RNA was extracted from Day 14 endometrial biopsies using the RNeasy Mini Kit (Qiagen). Briefly, frozen biopsy samples were disrupted and homogenized in RLT buffer with the use of a homogenizer (VDI 25; VWR International), and total RNA was purified following the manufacturer's instructions. To eliminate DNA contamination, RNA was treated with DNase I during RNA purification using the RNase-Free DNase Set (Qiagen). RNA concentration was determined by quantitative high-sensitivity RNA analysis on the Fragment Analyzer instrument (DNF-472; Advanced Analytical Technologies, Inc.). RNA library preparation and sequencing was conducted by the University of Missouri DNA Core facility.
Illumina TruSeq RNA library preparation and sequencing.
High-throughput sequencing services were performed at the University of Missouri DNA Core Facility. Libraries were constructed following the manufacturer's protocol with reagents supplied in Illumina's TruSeq mRNA stranded sample preparation kit. Briefly, the polyadenylated mRNA was purified from total RNA and fragmented. Double-stranded cDNA was generated from fragmented RNA, and the index-containing adapters were ligated. The final construct of each purified library was evaluated using the Fragment Analyzer instrument, quantified with the Qubit fluorimeter using the quant-iT HS dsDNA reagent kit (Invitrogen), and diluted according to Illumina's standard sequencing protocol for sequencing on an Illumina HiSeq 2500 sequencer.
RNA-Seq Data Analysis
Raw sequence reads (fastq) were trimmed of adapter sequences using cutadapt [41] followed by a windowed adaptive trimming step for base quality of the reads. The quality trimming was performed using Sickle (https://github.com/najoshi/sickle) with default parameters for paired-end reads. The read data was deposited in the Gene Expression Omnibus (GSE81449). The trimmed paired-end reads were then mapped to the bovine reference genome UMD3.1.1 using SubRead aligner [42] followed by quantifying the number of reads mapped to exon sequences of annotated genes using FeatureCounts [43]. Genes significantly differentially expressed (false discovery rate [FDR] P < 0.10) between samples were determined by fitting the read counts to a generalized linear model implemented in edgeR-robust [44]. The fold changes of differential expression were determined from log2(counts per million reads), and the length-normalized fragments per kilobase of exon per million reads was determined from the sum of exon lengths of each gene. All statistical analyses and plotting were performed in R. The Pearson correlation coefficients were measured in pairwise manner among samples to plot the correlation heat map of gene expressions. Conditional boxplots were generated using the Rlab package. The expression data of all genes across all the samples were used in exploratory maximum likelihood factor analysis by fitting the data to the factor analysis model using varimax rotation option of factanal in R. The plot was drawn using print method of R stats. Violin plots were generated using ggplot2.
Statistical Analyses
Statistical analyses were conducted using SAS (SAS Institute Inc.). Statistical significance was defined as P < 0.05.
Study two.
Pregnancy success to AI at Days 28 and 36 postinsemination was analyzed by logistic regression with Firth bias correction using the LOGISTIC procedure. The proportions of heifers pregnant at Days 28 and 36 were determined using the FREQ procedure. Progesterone concentrations were analyzed by ANOVA for repeated measures using the MIXED procedure. For the repeated measures, models included the effects of fertility classification, day of sample collection, the interaction between fertility classification and day of sample collection, and the random effect of heifer nested within fertility classification. For all analysis, orthogonal contrasts were used to compare groups with high (HF) versus low fertility (SF and IF) and to compare differences between the groups with low fertility (SF vs. IF).
Study three.
Day 14 embryo recovery rates were analyzed by logistic regression with Firth bias correction using the LOGISTIC procedure. The proportions of embryos recovered at Day 14 flush were determined using the FREQ procedure. Conceptus measurement data was subjected to least-squares ANOVA using the general linear models procedure. Conceptus length, area, and conception rate were tested for differences between fertility groups with embryo donor used as a covariate. Error terms used in the test of significance were identified according to the expectation of the mean squares for error. For all analysis, orthogonal contrasts were used to compare groups with high (HF) versus low fertility (SF and IF) and to compare the differences between the groups with low fertility (SF vs. IF).
RESULTS
Heifer Fertility Classification Using ET (Study One)
A total of 275 heifers were identified for the study, and six were culled due to extremely small size at weaning. Of the 269 heifers, four heifers were removed due to anovulation, death, health, or inability to pass the cervix. At the onset of synchronization for the first round of ET, 10 heifers were not cycling based on serum progesterone concentrations less than 1.0 ng per ml in two samples collected 10 days apart. Estrous cycles were induced in 8 of 10 of these heifers with synchronization based on serum progesterone concentration on Day 7 at ET. One of the anestrous heifers never began estrous cycles and was removed from the study; however, another anestrous heifer was pregnant each time she received an embryo and remained in the study.
In each round of ET, a single IVP embryo of high quality was transferred into heifers on Day 7 postestrus, and pregnancy determined on Days 28 and 44 by ultrasound followed by pregnancy termination on Day 44 (Fig. 1). Across the rounds of ET, 37 heifers did not have a CL at least once, and five heifers did not have a CL on two occasions. Pregnancy rates for each round of ET averaged 55% and 48% when diagnosed on Days 28 and 44, respectively. Overall, serial ET was conducted in 260 heifers, and four rounds of ET data were generated on 228 heifers and three rounds of ET data on 32 heifers. As illustrated in Figure 1, this approach was use to classify heifers as HF (100%, n = 30), SF (25%–33%, n = 53), or IF (0%, n = 12) based on Day 28 pregnancy success.
Pregnancy Rates after AI and Circulating Progesterone Concentrations (Study Two)
All HF and IF heifers and a subset of SF heifers were synchronized to estrus and bred 12 and 24 h postestrus with semen from a single bull of known high fertility. Pregnancy was determined on Days 28 and 36 by ultrasound. Blood samples were collected every other day beginning on Days 0 through 28 in all heifers and through Day 36 postmating in pregnant heifers. On Day 28, pregnancy rates were 70.4% in HF (19/27), 46.7% in SF (7/15), and 0% in IF (0/4) heifers. Orthogonal contrasts revealed that the Day 28 pregnancy rate was higher (P = 0.04) in HF than low fertile (SF and IF) heifers, but not different between SF and IF heifers (P = 0.24). On Day 36, pregnancy rates were 63.0% in HF, 40.0% in SF, and 0% in IF heifers. Day 36 pregnancy rates were not higher (P = 0.06) in HF than low fertile (SF and IF) heifers and not different (P = 0.30) between SF and IF heifers.
Progesterone concentrations during the first 20 days postinsemination were not different (P = 0.93) in all heifers regardless of fertility class (Fig. 2A). An effect of day (P < 0.0001) was detected because progesterone concentrations increased after ovulation with formation of a CL. Further, there was an effect of day (P < 0.0001), but not fertility class (P = 0.86), on progesterone concentrations for HF and SF heifers diagnosed as being pregnant through Day 36 (Fig. 2B). An interaction between fertility class and day of sample collection was observed (P = 0.04), which reflected the higher (P < 0.01) progesterone concentrations in SF than HF heifers on Day 36.
FIG. 2.
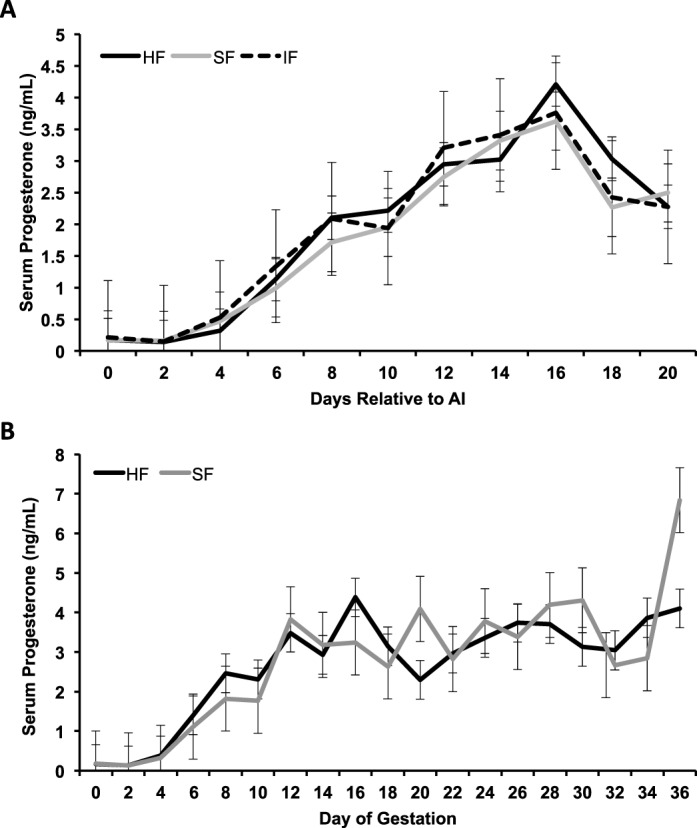
Circulating concentrations of progesterone in nonpregnant (A) and pregnant (B) fertility-classified heifers. Fertility-classified heifers were synchronized to estrus and bred by artificial insemination (AI) at 12 and 24 h postestrus with semen from a high-fertility bull. Pregnancy was determined on Days 28 and 35 by ultrasound. An effect of day (P < 0.0001) was detected because progesterone concentrations increased after ovulation. There was an effect of day (P < 0.0001), but not fertility class (P = 0.86), on progesterone concentrations for HF and SF heifers diagnosed pregnant through Day 36. Progesterone concentrations were higher in SF than HF heifers on Day 36 (P = 0.04, day × fertility class). Data is presented as least squares means (LSM) with the standard error of the mean (SEM).
Preimplantation Embryo Development in Fertility-Classified Heifers (Study Three)
Fertility-classified heifers were synchronized to estrus, and a single IVP embryo of high quality was transferred into HF (n = 28), SF (n = 17), and IF (n = 11) heifers on Day 7 postestrus. Conceptuses were recovered on Day 14 by nonsurgical uterine flush, and endometrial biopsies collected from the ipsilateral uterine horn. If recovered, conceptus length and area were determined. Embryo recovery rate was 60.7% for HF, 52.9% for SF, and 45.5% for IF heifers. Embryo donor had no effect (P = 0.94) on embryo recovery rate. Orthogonal contrasts found that embryo recovery rate was not different (P = 0.48) in HF as compared to low fertility (SF and IF) heifers and not different (P = 0.81) in SF than IF heifers. Gross conceptus morphology was not different among fertility-classifications, and a spherical to ovoid conceptus was recovered with normal morphology and obvious embryonic disk from HF, SF, and IF heifers (Fig. 3A). Conceptus length was not different (P = 0.47) in HF as compared to low fertility (SF and IF) heifers and not different (P = 0.73) in SF as compared to IF heifers (Fig. 3B). Similarly, conceptus area was not different (P = 0.82) between HF and low fertility (SF and IF) heifers and not different (P = 0.73) between SF and IF heifers (Fig. 3B).
FIG. 3.

Day 14 conceptus morphology and measurements. Fertility-classified heifers were synchronized to estrus and received a high-quality in vivo produced embryo on Day 7 postestrus. All heifers were nonsurgically flushed on Day 14 (7 days post-ET) to recover the conceptus. If present, conceptus length and width were determined under a microscope. A) Representative conceptus morphology recovered from high fertile (HF), subfertile (SF), and infertile (IF) heifers. Bars = 1 mm. B and C) Conceptus length and area were not different (P ≥ 0.60) among the fertility-classified heifers. Data is presented as least squares means (LSM) with the standard error of the mean (SEM).
RNA-Seq Analysis of Endometrial Biopsies (Study Three)
After nonsurgical flush on Day 14 (7 days post-ET), endometrial biopsies were obtained from the ipsilateral uterine horn. Histological analysis of selected endometrial biopsies determined that the biopsy method repeatedly obtained mostly intercaruncular endometrium with glands, stroma, and luminal epithelium (Fig. 4A).
FIG. 4.

Endometrial biopsy histology and RNA sequencing analysis from Day 14 pregnant heifers. Fertility-classified heifers were synchronized to estrus and received two high-quality in vivo produced embryos on Day 7 postestrus. All heifers were nonsurgically flushed on Day 14 (7 days post-ET) to recover the conceptus. If a conceptus was present in the uterine flush, an endometrial biopsy was obtained from the uterine horn ipsilateral to the corpus luteum (CL). Total RNA was extracted from five biopsies of pregnant high fertile (HF), subfertile (SF), and infertile (IF) heifers and sequenced. Normalized and log2 transformed read count data were produced with edgeR-robust analysis. A) Histological analysis of a representative endometrial biopsy. All biopsies were predominantly composed of intercaruncular endometrium. Sections were stained with hematoxylin and eosin. LE, luminal epithelium; GE, glandular epithelium; S, stroma. Bar = 500 μm. B) Pairwise correlation (Pearson) analysis of gene expression levels between endometrial biopsy samples. Each column represents one sample and shows the correlation to all samples (including itself) with red for lowest (0) distance and blue for the highest observed distance. C) Multidimensional scaling plot. A maximum likelihood factor plot of gene expression variation among the samples. The major factors (factor 1 and factor 2) that explain the expression changes among the samples are plotted in the x- and y-axis, respectively. The individual samples (n = 15) representing the three fertility groups (HF, SF, and IF) are shown with different colors (green, blue, and red, respectively).
Transcriptional profiling of Day 14 endometrial biopsies from pregnant heifers (n = 5 heifers per fertility class) was conducted using RNA sequencing. Sequencing of the libraries yielded more than 60 million quality reads for each sample. The reads used in data analysis for each sample were of minimum length 30 bp with sliding window Phred scores of at least 30 and were mapped to the reference genome at 96%–98% across samples with 70%–80% of all the pair reads mapped concordantly. Between 10% to 18% of paired reads mapped discordantly, and they likely represent reads aligning to splice or fusion sites. Less than 1% of the read pairs mapped to the genome as singletons with unmapped mates; those reads may represent repeat sequences, sequences from foreign sources, such as microbial contamination, or regions of the genome that have not been well assembled, and is not unusual [45].
A seed-and-vote mapping strategy was used to quantify read counts to individual genes annotated from the reference genome [42, 43]. On average, 63% of the mapped reads were associated with at least five reads to individual genes in each sample. About 14% of genes were not expressed in any samples based on absence of read counts. Such a result could be due to lack of coverage, but this is unlikely given the high coverage (>60 million) of reads per sample and consistency of zero-read mapping between samples. About 0.5% of mapped reads showed mapping of one read on average per gene per sample. Genes were then filtered that were associated with less than or equal to one read per million in each library prior to fitting the count data into the generalized linear model implemented in edgeR-robust [44]. Accordingly, 26 total differentially expressed genes (DEGs; >2-fold change, FDR P < 0.10) were identified for the endometrium from HF as compared to SF heifers (Table 1). There were 12 DEGs in the endometrium from SF as compared to IF heifers (Table 2), and only three DEGs when HF were compared to IF heifers (Table 3).
TABLE 1.
Differentially expressed genes in endometrial biopsies from high fertile (HF) and subfertile (SF) heifers.

Ig, immunoglobulin.
Log2 fold change (logFC) in HF as compared to SF endometrial biopsy samples.
FDR, false discovery rate.
Data are presented as mean fragments per kilobase of exon per million (FPKM) mapped reads.
TABLE 2.
Differentially expressed genes in endometrial biopsies from subfertile (SF) and infertile (IF) heifers.

Ig, immunoglobulin.
Log2 fold change (logFC) in SF as compared to IF endometrial biopsy samples.
FDR, false discovery rate.
Data are presented as mean fragments per kilobase of exon per million (FPKM) mapped reads.
TABLE 3.
Differentially expressed genes in endometrial biopsies from high fertile (HF) and infertile (IF) heifers.

Log2 fold change (logFC) in HF as compared to IF endometrial biopsy samples.
Data are presented as mean fragments per kilobase of exon per million (FPKM) mapped reads.
The low number of DEGs detected by edgeR-robust analysis suggested low overall variation in gene expression across samples, which was confirmed by high pairwise correlation between samples (Fig. 4B) and lack of sample clustering as illustrated in the multidimensional scaling plot (Fig. 4C). Expression values in each group were then partitioned into nonoverlapping bins and plotted as conditional box plots between groups (Supplemental Fig. S1A; Supplemental Data are available online at www.biolreprod.org). These plots indicate reduced gene expression changes in the endometria of IF as compared to HF or SF heifers and were not observed in HF as compared to SF heifers. The violin plots presented in Supplemental Figure S1B present distribution of log2 fold changes of expression relative to log2 counts per million reads in pairwise manner among samples. This analysis confirmed the reduced DEGs in IF as compared to HF and SF heifers.
The DEGs identified by edgeR-robust analysis of endometrial biopsy RNA sequencing are presented in Figure 5 and summarized in Tables 1–3 and Supplemental Tables S1–S3. Expression of 14 genes was higher and 12 genes was lower in endometrial biopsies of HF as compared to SF heifers (Table 1 and Supplemental Table S1). Many of the 14 genes that were more abundant in the HF endometrium encoded proteins with antimicrobial activity (TAP, MUC1) or were immunoglobulins. In SF as compared to IF heifers, there were 12 genes with reduced expression in the endometrial biopsies (Table 2 and Supplemental Table S2). All three DEGs were lower in abundance in endometrial biopsies of HF as compared to IF heifers (Table 3 and Supplemental Table S3). Of note, 10 of the DEGs overlapped between the fertility-classified heifer endometrial biopsies.
FIG. 5.

Venn diagram showing the number of unique or common transcripts between the endometrium of fertility-classified heifers (HF, high fertile; SF, subfertile; IF, infertile). Increased (red) and decreased (blue) genes are presented (>2-fold change and FDR P < 0.05).
DISCUSSION
The present study found that serial rounds of ET, each followed by pregnancy determination on Days 28 and 42, is an effective strategy to identify beef heifers with high and low aggregate rates of early pregnancy loss. One might assert that by chance rates of heifers always or never being pregnant 12.5% of the time after four rounds of ET would be correct if the pregnancy rate were a random binomial event; however, pregnancy is not a random binomial event and rather a cumulative physiological process in which each successive day of gestation is dependent upon the additive successful events up to that time. Similarly, McMillan and Donnison [35] utilized serial ET to classify dairy heifers as being high or low fertile. Failures in mechanisms involved in conceptus elongation and maternal recognition of pregnancy were hypothesized to be the major cause of pregnancy loss in the low-fertility dairy heifers [35, 36]. Indeed, the majority of pregnancy losses in beef and dairy heifers, nonlactating cows, and lactating cows occur during the first 3 to 4 wk of pregnancy [30, 46]. Studies in sheep, beef heifers, and dairy cattle established that an early or delayed rise in circulating levels of progesterone after ovulation can advance or retard conceptus elongation [20, 32, 47–50]. In the present study two, circulating levels of progesterone were not different in fertility-classified heifers, regardless of pregnancy status. This result agrees with findings from the analysis of both dairy heifers and beef heifers that were fertility classified by serial ET or AI, respectively [35, 51]. Thus, differences in circulating levels of progesterone are not a factor in the fertility differences for the beef heifers in the present study. Although increasing concentrations of progesterone after ovulation clearly advances conceptus elongation in both beef and dairy cattle, supplementation with progesterone during early pregnancy has equivocal efficacy in increasing embryonic survival [52, 53]. In fact, strategies to improve postovulatory progesterone concentration such as treatment with GnRH or human chorionic gonadotropin following AI have resulted in inconsistent benefits in pregnancy outcomes in lactating dairy cows [54–56]. Further, progesterone administration is unlikely to rescue development of embryos with inherent genetic defects or during gestation in high-producing dairy cows [57–59].
In study three, fertility-classified heifers received a single high-quality in vivo-produced embryo on Day 7 postestrus and were nonsurgically flushed to recover the embryo on Day 14. Embryo recovery rate and embryo morphology development (length and area) were not different among HF, SF, and IF heifers. The average embryo recovery rate on Day 14 was 53% across HF, SF, and IF heifers, which is not much lower than the 66% pregnancy rate found on Day 16 in beef heifers bred by AI [7]. The lower pregnancy rate in study three could be due to a failure of the nonsurgical flush procedure to recover embryos from some heifers and also the use of in vivo-produced embryo that was frozen and then thawed prior to transfer. The result of the present studies supports the idea that pregnancy loss in the SF and/or IF heifers must occur between Days 14 and 28, which encompasses the period of conceptus elongation, pregnancy recognition, and implantation for establishment of pregnancy [17, 20, 32]. Given the lack of differences in conceptus survival and development, it is not surprising that substantial differences in gene expression for the endometrial biopsies from Day 14 pregnant HF, SF, and IF heifers were not observed. Indeed, the transcriptome difference was also minimal in other studies of fertility-classified heifers that evaluated endometria from beef heifers on Day 13 postestrus [51], endometrial cytobrush biopsies from beef heifers on Day 14 postestrus [60], or endometrial biopsies from lactating dairy cows on Day 13 postestrus [61]. The lack of conserved differences could be attributed to a myriad of factors, including how fertility was classified, breed effects, and endometrial sampling. For instance, endometrium obtained at slaughter is generally a mixture of intercaruncular and caruncular endometrium, whereas the obtained biopsies in the present study are mostly intercaruncular endometrium. Of note, cytobrush sampling obtains mostly only luminal epithelium from both caruncular and intercaruncular endometrium.
Although large numbers of DEGs were not identified, RNA sequencing analysis did detect differences in endometrial gene expression among the fertility-classified heifers. The majority of genes that were more abundant in HF than SF, as well as IF than SF, endometrium encode factors involved in reproductive tract defense against pathogens. For instance, TAP was first identified in the bovine tracheal mucosa and found to be a member of a group of cysteine-rich, cationic, antimicrobial peptides found in animals, insects, and plants [62]. Purified TAP had antibacterial activity in vitro against Escherichia coli, Staphylococcus aureus, Klebsiella pneumonia, and Pseudomonas aeruginosa. In addition, TAP was active against Candida albicans, indicating a broad spectrum of activity. In the present study, MUC1 was also lower in SF heifers, and it forms part of the glycocalyx barrier that provides innate immune protection against bacterial infections [63]. Additionally, MUC1 regulates implantation in mice and perhaps other mammals. Walker and coworkers [64] found evidence that the embryo modulates the uterine immune system on Day 17 and induces expression of molecules in the endometrium that function to suppress the immune response and/or promote tolerance to the embryo. During this period of immune suppression, the endometrium would be expected to be susceptible to infections; therefore, it actively expresses specific molecules for defense against foreign pathogens such as upregulation of genes of the innate immune response, including antimicrobial response genes like TAP. It should be noted that none of the heifers used in the present study had evidence of metritis, and endometritis is not very prevalent in beef heifers.
Many of the down-regulated genes in SF as compared to HF endometrium encoded various immunoglobulins or a transcription factor (POU2AF1) that are expressed in B lymphocytes, which function in the humoral immunity component of the adaptive immune system by secreting antibodies. Thus, B cells are likely higher in the endometrium of HF than SF heifers. Pregnancy changes the population of lymphocytes and macrophages in the bovine uterus [65–67]. The numbers of both cell types are significantly reduced between early and midpregnancy in cattle [68]. By midpregnancy, virtually no lymphocytes or macrophages are found in the caruncular endometrium although they are still present in the intercaruncular endometrium [69]. The lymphocyte population in the early pregnant cattle uterus is composed primarily of B cells, T cells, and natural killer cells [70–72]. B lymphocytes are widely distributed throughout the endometrium, localizing in the stroma, myometrium, and the luminal and glandular epithelium. The B lymphocyte population in bovine endometrium was relatively large compared to the populations of γδT, CD4+, CD8+, and NK cells detected [70]. Although several genes were more abundant in the endometrium of SF than HF and SF than IF heifers, the function of most of those genes is unknown. Collectively, these results support the idea that the innate and adaptive immune system is different in the endometrium of SF as compared to that of HF and IF heifers, which could be involved in the observed fertility differences [67]. The lack of conserved differences in the endometrial transcriptome of the HF versus low fertility (SF and IF) heifers indicates that the biological mechanisms underlying subfertility and infertility are possibly different, and those differences may not manifest until after Day 14 as the conceptus begins rapid growth for elongation. Future experiments will need to explore differences in the endometrial secrotome because histotroph has a major influence on growth and development of the conceptus [14, 20, 32].
The present study and others [35, 36, 51, 60, 61] support the hypothesis that natural variation in pregnancy rates can be utilized in cattle to identify animals with innate differences in uterine competence to support growth and development of the conceptus for establishment of pregnancy. Although not addressed in the present study, it is also important to address the substantial loss of embryos that occurs prior to Day 7, particularly in lactating dairy cows, due to problems with oocyte competence, sperm transport, fertilization, and perhaps oviductal-uterine function [73, 74]. Studies of animals with natural variation in uterine competency for pregnancy could help define which genes and biological pathways in the endometrium are crucial to establish endometrial receptivity and support conceptus elongation in cattle. Further, the use of this animal model could discover genes and biomarkers that can be used to select animals for higher fertility and to diagnose subfertility and infertility.
Supplementary Material
ACKNOWLEDGMENT
The authors appreciate the assistance of the staff at the USDA-ARS Fort Keogh with animal handling and sample collection, William Rembert for collecting ovaries, the Chernin family and Central Packing (Center Hill, Florida) for donating ovarian tissue, Scott Randel of Southeastern Semen (Wellborn Florida) for donating semen, and Zoetis for the donation of synchronization products. They also appreciate the help of Drs. Mike Smith and Duane Keisler and the MU DNA Core facility of the University of Missouri-Columbia with completion of the studies.
Footnotes
This research was supported by Grant 1 R01 HD072898 from the Eunice Kennedy Shriver National Institute of Child Health and Human Development.
REFERENCES
- Diskin MG, Morris DG. Embryonic and early foetal losses in cattle and other ruminants Reprod Domest Anim 2008. 43 (Suppl 2): 260 267 [DOI] [PubMed] [Google Scholar]
- Santos JEP, Thatcher WW, Chebel RC, Cerri RLA, Galvao KN. The effect of embryonic death rates in cattle on the efficacy of estrus synchronization programs. Anim Reprod Sci. 2004;82:513–535. doi: 10.1016/j.anireprosci.2004.04.015. [DOI] [PubMed] [Google Scholar]
- Hansen PJ, Block J. Towards an embryocentric world: the current and potential uses of embryo technologies in dairy production. Reprod Fertil Dev. 2003;16:1–14. doi: 10.10371/RD03073. [DOI] [PubMed] [Google Scholar]
- Diskin MG, Murphy JJ, Sreenan JM. Embryo survival in dairy cows managed under pastoral conditions. Anim Reprod Sci. 2006;96:297–311. doi: 10.1016/j.anireprosci.2006.08.008. [DOI] [PubMed] [Google Scholar]
- Diskin MG, Parr MH, Morris DG. Embryo death in cattle: an update. Reprod Fertil Dev. 2011;24:244–251. doi: 10.1071/RD11914. [DOI] [PubMed] [Google Scholar]
- Berg DK, Van Leeuwen J, Beaumont S, Berg M, Pfeffer PL. Embryo loss in cattle between Days 7 and 16 of pregnancy. Theriogenology. 2010;73:250–260. doi: 10.1016/j.theriogenology.2009.09.005. [DOI] [PubMed] [Google Scholar]
- Diskin MG, Sreenan JM. Fertilization and embryonic mortality rates in beef heifers after artificial insemination. J Reprod Fertil. 1980;59:463–468. doi: 10.1530/jrf.0.0590463. [DOI] [PubMed] [Google Scholar]
- Roche JF, Bolandl MP, McGeady TA. Reproductive wastage following artificial insemination of heifers. Vet Rec. 1981;109:401–404. doi: 10.1136/vr.109.18.401. [DOI] [PubMed] [Google Scholar]
- Chebel RC, Santos JE, Reynolds JP, Cerri RL, Juchem SO, Overton M. Factors affecting conception rate after artificial insemination and pregnancy loss in lactating dairy cows. Anim Reprod Sci. 2004;84:239–255. doi: 10.1016/j.anireprosci.2003.12.012. [DOI] [PubMed] [Google Scholar]
- Moore K, Thatcher WW. Major advances associated with reproduction in dairy cattle. J Dairy Sci. 2006;89:1254–1266. doi: 10.3168/jds.S0022-0302(06)72194-4. [DOI] [PubMed] [Google Scholar]
- Looney CR, Nelson JS, Schneider HJ, Forrest DW. Improving fertility in beef cow recipients. Theriogenology. 2006;65:201–209. doi: 10.1016/j.theriogenology.2005.09.023. [DOI] [PubMed] [Google Scholar]
- McMillan WH. Statistical models predicting embryo survival to term in cattle after embryo transfer. Theriogenology. 1998;50:1053–1070. doi: 10.1016/s0093-691x(98)00207-6. [DOI] [PubMed] [Google Scholar]
- Miravet-Valenciano JA, Rincon-Bertolin A, Vilella F, Understanding Simon C. and improving endometrial receptivity. Curr Opin Obstet Gynecol. 2015;27:187–192. doi: 10.1097/GCO.0000000000000173. [DOI] [PubMed] [Google Scholar]
- Spencer TE, Hansen TR. Implantation and establishment of pregnancy in ruminants. Adv Anat Embryol Cell Biol. 2015;216:105–135. doi: 10.1007/978-3-319-15856-3_7. [DOI] [PubMed] [Google Scholar]
- Betteridge KJ, Flechon J-E. The anatomy and physiology of pre-attachment bovine embryos. Theriogenology. 1988;29:155–187. [Google Scholar]
- Betteridge KJ, Eaglesome MD, Randall GC, Collection Mitchell D. description and transfer of embryos from cattle 10–16 days after oestrus. J Reprod Fertil. 1980;59:205–216. doi: 10.1530/jrf.0.0590205. [DOI] [PubMed] [Google Scholar]
- Guillomot M, Reinaud P, Charpigny G, Martal J. Embryo-uterine interactions during early stages of pregnancy in domestic mammals. Reprod Nutr Dev. 1988;28:1629–1648. doi: 10.1051/rnd:19881008. [DOI] [PubMed] [Google Scholar]
- Fléchon J-E, Guillomot M, Charlier M, Fléchon B, Martal J. Experimental studies on the elongation of the ewe blastocyst. Reprod Nutr Dev. 1986;26:1017–1024. doi: 10.1051/rnd:19860609. [DOI] [PubMed] [Google Scholar]
- Satterfield MC, Bazer FW, Spencer TE. Progesterone regulation of preimplantation conceptus growth and galectin 15 (LGALS15) in the ovine uterus. Biol Reprod. 2006;75:289–296. doi: 10.1095/biolreprod.106.052944. [DOI] [PubMed] [Google Scholar]
- Spencer TE, Forde N, Lonergan P. The role of progesterone and conceptus-derived factors in uterine biology during early pregnancy in ruminants. J Dairy Sci. 2016;99:5941–5950. doi: 10.3168/jds.2015-10070. [DOI] [PubMed] [Google Scholar]
- Forde N, Lonergan P. Transcriptomic analysis of the bovine endometrium: what is required to establish uterine receptivity to implantation in cattle? J Reprod Dev. 2012;58:189–195. doi: 10.1262/jrd.2011-021. [DOI] [PubMed] [Google Scholar]
- Forde N, Carter F, Spencer TE, Bazer FW, Sandra O, Mansouri-Attia N, Okumu LA, McGettigan PA, Mehta JP, McBride R, O'Gaora P, Roche JF, et al. Conceptus-induced changes in the endometrial transcriptome: how soon does the cow know she is pregnant? Biol Reprod. 2011;85:144–156. doi: 10.1095/biolreprod.110.090019. [DOI] [PubMed] [Google Scholar]
- Forde N, Spencer TE, Bazer FW, Song G, Roche JF, Lonergan P. Effect of pregnancy and progesterone concentration on expression of genes encoding for transporters or secreted proteins in the bovine endometrium. Physiol Genomics. 2010;41:53–62. doi: 10.1152/physiolgenomics.00162.2009. [DOI] [PubMed] [Google Scholar]
- Gray CA, Bartol FF, Taylor KM, Wiley AA, Ramsey WS, Ott TL, Bazer FW, Spencer TE. Ovine uterine gland knock-out model: effects of gland ablation on the estrous cycle Biol Reprod 2000. 62 448 456 [DOI] [PubMed] [Google Scholar]
- Gray CA, Taylor KM, Ramsey WS, Hill JR, Bazer FW, Bartol FF, Spencer TE. Endometrial glands are required for preimplantation conceptus elongation and survival. Biol Reprod. 2001;64:1608–1613. doi: 10.1095/biolreprod64.6.1608. [DOI] [PubMed] [Google Scholar]
- Farin CE, Imakawa K, Roberts RM. In situ localization of mRNA for the interferon, ovine trophoblast protein-1, during early embryonic development of the sheep. Molecular Endocrinology. 1989;3:1099–1107. doi: 10.1210/mend-3-7-1099. [DOI] [PubMed] [Google Scholar]
- Farin CE, Imakawa K, Hansen TR, McDonnell JJ, Murphy CN, Farin PW, Roberts RM. Expression of trophoblastic interferon genes in sheep and cattle. Biol Reprod. 1990;43:210–218. doi: 10.1095/biolreprod43.2.210. [DOI] [PubMed] [Google Scholar]
- Roberts RM, Chen Y, Ezashi T, Walker AM. Interferons and the maternal-conceptus dialog in mammals. Semin Cell Dev Biol. 2008;19:170–177. doi: 10.1016/j.semcdb.2007.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer TE, Sandra O, Wolf E. Genes involved in conceptus-endometrial interactions in ruminants: insights from reductionism and thoughts on holistic approaches. Reproduction. 2008;135:165–179. doi: 10.1530/REP-07-0327. [DOI] [PubMed] [Google Scholar]
- Thatcher WW, Guzeloglu A, Mattos R, Binelli M, Hansen TR, Pru JK. Uterine-conceptus interactions and reproductive failure in cattle. Theriogenology. 2001;56:1435–1450. doi: 10.1016/s0093-691x(01)00645-8. [DOI] [PubMed] [Google Scholar]
- Lonergan P. State-of-the-art embryo technologies in cattle. Soc Reprod Fertil Suppl. 2007;64:315–325. doi: 10.5661/rdr-vi-315. [DOI] [PubMed] [Google Scholar]
- Lonergan P, Forde N, Spencer T. Role of progesterone in embryo development in cattle. Reprod Fertil Dev. 2016;28:66–74. doi: 10.1071/RD15326. [DOI] [PubMed] [Google Scholar]
- Bauersachs S, Mitko K, Ulbrich SE, Blum H, Wolf E. Transcriptome studies of bovine endometrium reveal molecular profiles characteristic for specific stages of estrous cycle and early pregnancy. Exp Clin Endocrinol Diabetes. 2008;116:371–384. doi: 10.1055/s-2008-1076714. [DOI] [PubMed] [Google Scholar]
- Ulbrich SE, Groebner AE, Bauersachs S. Transcriptional profiling to address molecular determinants of endometrial receptivity–lessons from studies in livestock species. Methods. 2013;59:108–115. doi: 10.1016/j.ymeth.2012.10.013. [DOI] [PubMed] [Google Scholar]
- McMillan WH, Donnison MJ. Understanding maternal contributions to fertility in recipient cattle: development of herds with contrasting pregnancy rates. Anim Reprod Sci. 1999;57:127–140. doi: 10.1016/s0378-4320(99)00063-9. [DOI] [PubMed] [Google Scholar]
- Peterson AJ, Lee R-F. Improving successful pregnancies after embryo transfer. Theriogenology. 2003;59:687–697. doi: 10.1016/s0093-691x(02)01248-7. [DOI] [PubMed] [Google Scholar]
- Parrish JJ, Krogenaes A, Susko-Parrish JL. Effect of bovine sperm separation by either swim-up or Percoll method on success of in vitro fertilization and early embryonic development. Theriogenology. 1995;44:859–869. doi: 10.1016/0093-691x(95)00271-9. [DOI] [PubMed] [Google Scholar]
- Sakatani M, Alvarez NV, Takahashi M, Hansen PJ. Consequences of physiological heat shock beginning at the zygote stage on embryonic development and expression of stress response genes in cattle. J Dairy Sci. 2012;95:3080–3091. doi: 10.3168/jds.2011-4986. [DOI] [PubMed] [Google Scholar]
- Bellow RA, Staigmiller RB, Wilson JM, Phelps DA, Darling A. Use of bovine FSH for superovulation and embryo production in beef heifers. Theriogenology. 1991;35:1069–1082. [Google Scholar]
- Pohler KG, Pereira MHC, Lopes FR, Lawrence JC, Keisler DH, Smith MF, Vasconcelos JLM, Green JA. Circulating concentrations of bovine pregnancy-associated glycoproteins and late embryonic mortality in lactating dairy herds. J Dairy Sci. 2015 doi: 10.3168/jds.2015-10192. [DOI] [PubMed] [Google Scholar]
- Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads EMBNet.journal [Online] 2011. 17 1: 10 12 [Google Scholar]
- Liao Y, Smyth GK, Shi W. The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013;41:e108–e108. doi: 10.1093/nar/gkt214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao Y, Smyth GK. Shi W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics. 2014;30:923–930. doi: 10.1093/bioinformatics/btt656. [DOI] [PubMed] [Google Scholar]
- Zhou X, Lindsay H, Robinson MD. Robustly detecting differential expression in RNA sequencing data using observation weights. Nucleic Acids Res. 2014;42:e91. doi: 10.1093/nar/gku310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitacre LK, Tizioto PC, Kim J, Sonstegard TS, Schroeder SG, Alexander LJ, Medrano JF, Schnabel RD, Taylor JF, Decker JE. What's in your next-generation sequence data? An exploration of unmapped DNA and RNA sequence reads from the bovine reference individual. BMC Genomics. 2015;16:1114. doi: 10.1186/s12864-015-2313-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sreenan JM, Diskin MG. Factors affecting pregnancy rate following embryo transfer in the cow. Theriogenology. 1987;27:99–113. [Google Scholar]
- Garrett JE, Geisert RD, Zavy MT, Morgan GL. Evidence for maternal regulation of early conceptus growth and development in beef cattle. J Reprod Fertil. 1988;84:437–446. doi: 10.1530/jrf.0.0840437. [DOI] [PubMed] [Google Scholar]
- Satterfield MC, Bazer FW, Spencer TE. Progesterone regulation of preimplantation conceptus growth and galectin 15 (LGALS15) in the ovine uterus. Biol Reprod. 2006;75:289–296. doi: 10.1095/biolreprod.106.052944. [DOI] [PubMed] [Google Scholar]
- Forde N, Beltman ME, Duffy GB, Duffy P, Mehta JP, O'Gaora P, Roche JF, Lonergan P, Crowe MA. Changes in the endometrial transcriptome during the bovine estrous cycle: effect of low circulating progesterone and consequences for conceptus elongation. Biol Reprod. 2011;84:266–278. doi: 10.1095/biolreprod.110.085910. [DOI] [PubMed] [Google Scholar]
- Forde N, Carter F, Fair T, Crowe MA, Evans ACO, Spencer TE, Bazer FW, McBride R, Boland MP, O'Gaora P, Lonergan P, Roche JF. Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol Reprod. 2009;81:784–794. doi: 10.1095/biolreprod.108.074336. [DOI] [PubMed] [Google Scholar]
- Minten MA, Bilby TR, Bruno RGS, Allen CC, Madsen CA, Wang Z, Sawyer JE, Tibary A, Neibergs HL, Geary TW, Bauersachs S, Spencer TE. Effects of fertility on gene expression and function of the bovine endometrium. PLoS One. 2013;8:e69444. doi: 10.1371/journal.pone.0069444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parr MH, Crowe MA, Lonergan P, Evans ACO, Rizos D, Diskin MG. Effect of exogenous progesterone supplementation in the early luteal phase post-insemination on pregnancy per artificial insemination in Holstein-Friesian cows. Anim Reprod Sci. 2014;150:7–14. doi: 10.1016/j.anireprosci.2014.08.008. [DOI] [PubMed] [Google Scholar]
- Beltman ME, Lonergan P, Diskin MG, Roche JF, Crowe MA. Effect of progesterone supplementation in the first week post conception on embryo survival in beef heifers. Theriogenology. 2009;71:1173–1179. doi: 10.1016/j.theriogenology.2008.12.014. [DOI] [PubMed] [Google Scholar]
- Santos JE, Thatcher WW, Pool L, Overton MW. Effect of human chorionic gonadotropin on luteal function and reproductive performance of high-producing lactating Holstein dairy cows. J Anim Sci. 2001;79:2881–2894. doi: 10.2527/2001.79112881x. [DOI] [PubMed] [Google Scholar]
- López-Gatius F, Santolaria P, Martino A, Delétang F, De Rensis F. The effects of GnRH treatment at the time of AI and 12 days later on reproductive performance of high producing dairy cows during the warm season in northeastern Spain. Theriogenology. 2006;65:820–830. doi: 10.1016/j.theriogenology.2005.07.002. [DOI] [PubMed] [Google Scholar]
- Nascimento AB, Bender RW, Souza AH, Ayres H, Araujo RR, Guenther JN, Sartori R, Wiltbank MC. Effect of treatment with human chorionic gonadotropin on day 5 after timed artificial insemination on fertility of lactating dairy cows. J Dairy Sci. 2013;96:2873–2882. doi: 10.3168/jds.2012-5895. [DOI] [PubMed] [Google Scholar]
- Mann GE, Fray MD, Lamming GE. Effects of time of progesterone supplementation on embryo development and interferon-τ production in the cow. Vet J. 2006;171:500–503. doi: 10.1016/j.tvjl.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Lonergan P, Woods A, Fair T, Carter F, Rizos D, Ward F, Quinn K, Evans A. Effect of embryo source and recipient progesterone environment on embryo development in cattle. Reprod Fertil Dev. 2007;19:861–868. doi: 10.1071/rd07089. [DOI] [PubMed] [Google Scholar]
- Wiltbank MC, Souza AH, Carvalho PD, Bender RW, Nascimento AB. Improving fertility to timed artificial insemination by manipulation of circulating progesterone concentrations in lactating dairy cattle. Reprod Fertil Dev. 2011;24:238–243. doi: 10.1071/RD11913. [DOI] [PubMed] [Google Scholar]
- Salilew-Wondim D, Hölker M, Rings F, Ghanem N, Ulas-Cinar M, Peippo J, Tholen E, Looft C, Schellander K, Tesfaye D. Bovine pretransfer endometrium and embryo transcriptome fingerprints as predictors of pregnancy success after embryo transfer. Physiol Genomics. 2010;42:201–218. doi: 10.1152/physiolgenomics.00047.2010. [DOI] [PubMed] [Google Scholar]
- Moore SG, Pryce JE, Hayes BJ, Chamberlain AJ, Kemper KE, Berry DP, McCabe M, Cormican P, Lonergan P, Fair T, Butler ST. Differentially expressed genes in endometrium and corpus luteum of Holstein cows selected for high and low fertility are enriched for sequence variants associated with fertility. Biol Reprod. 2015;94:19. doi: 10.1095/biolreprod.115.132951. [DOI] [PubMed] [Google Scholar]
- Diamond G, Zasloff M, Eck H, Brasseur M, Maloy WL, Bevins CL. Tracheal antimicrobial peptide, a cysteine-rich peptide from mammalian tracheal mucosa: peptide isolation and cloning of a cDNA. Proc Natl Acad Sci U S A. 1991;88:3952–3956. doi: 10.1073/pnas.88.9.3952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeSouza MM, Surveyor GA, Price RE, Julian J, Kardon R, Zhou X, Gendler S, Hilkens J, Carson DD. MUC1/episialin: a critical barrier in the female reproductive tract. J Reprod Immunol. 1999;45:127–158. doi: 10.1016/s0165-0378(99)00046-7. [DOI] [PubMed] [Google Scholar]
- Walker CG, Meier S, Littlejohn MD, Lehnert K, Roche JR, Mitchell MD. Modulation of the maternal immune system by the pre-implantation embryo. BMC Genomics. 2010;11:474. doi: 10.1186/1471-2164-11-474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- French LR, Northey DL. Inhibitory effect of the bovine conceptus on lymphocyte stimulation. J Anim Sci. 1983;57:456–465. doi: 10.2527/jas1983.572456x. [DOI] [PubMed] [Google Scholar]
- Skopets B, Li J, Thatcher WW, Roberts RM, Hansen PJ. Inhibition of lymphocyte proliferation by bovine trophoblast protein-1 (type I trophoblast interferon) and bovine interferon-alpha I1. Vet Immunol Immunopathol. 1992;34:81–96. doi: 10.1016/0165-2427(92)90153-h. [DOI] [PubMed] [Google Scholar]
- Fair T. The contribution of the maternal immune system to the establishment of pregnancy in cattle. Front Immunol. 2015;6:7. doi: 10.3389/fimmu.2015.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vander Wielen AL, King GJ. Intraepithelial lymphocytes in the bovine uterus during the oestrous cycle and early gestation. J Reprod Fertil. 1984;70:457–462. doi: 10.1530/jrf.0.0700457. [DOI] [PubMed] [Google Scholar]
- Low BG, Hansen PJ, Drost M, Gogolin-Ewens KJ. Expression of major histocompatibility complex antigens on the bovine placenta. J Reprod Fertil. 1990;90:235–243. doi: 10.1530/jrf.0.0900235. [DOI] [PubMed] [Google Scholar]
- Oliveira LJ, Mansouri-Attia N, Mansourri-Attia N, Fahey AG, Browne J, Forde N, Roche JF, Lonergan P, Fair T. Characterization of the Th profile of the bovine endometrium during the oestrous cycle and early pregnancy. PLoS One. 2013;8:e75571. doi: 10.1371/journal.pone.0075571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung ST, Derecka K, Mann GE, Flint AP, Wathes DC. Uterine lymphocyte distribution and interleukin expression during early pregnancy in cows. J Reprod Fertil. 2000;119:25–33. [PubMed] [Google Scholar]
- Mansouri-Attia N, Oliveira LJ, Forde N, Fahey AG, Browne JA, Roche JF, Sandra O, Reinaud P, Lonergan P, Fair T. Pivotal role for monocytes/macrophages and dendritic cells in maternal immune response to the developing embryo in cattle. Biol Reprod. 2012;87:123. doi: 10.1095/biolreprod.112.101121. [DOI] [PubMed] [Google Scholar]
- Santos JEP, Thatcher WW, Chebel RC, Cerri RLA, Galvão KN. The effect of embryonic death rates in cattle on the efficacy of estrus synchronization programs. Anim Reprod Sci. 2004;82–83:513–535. doi: 10.1016/j.anireprosci.2004.04.015. [DOI] [PubMed] [Google Scholar]
- Perkel KJ, Tscherner A, Merrill C, Lamarre J, Madan P. The ART of selecting the best embryo: A review of early embryonic mortality and bovine embryo viability assessment methods. Mol Reprod Dev. 2015;82:822–838. doi: 10.1002/mrd.22525. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.