

Abstract
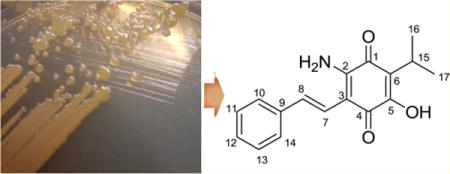
Lumiquinone A (1), an unusual aminobenzoquinone member within the phenylpropanoid class of natural products, together with the known compound 3,5-dihydroxy-4-isopropyl-trans-stilbene (2), was isolated from the entomopathogenic bacterium Photorhabdus luminescens TT01. On the basis of the analysis of extensive 2D NMR and high-resolution ESI-QTOF-MS spectral data, the structure of 1 was determined to be a 2-amino-5-hydroxy-1,4-benzoquinone substituted with (E)-2-phenylvinyl and isopropyl functional groups. Free α-aminomalonate medium supplementation significantly enhanced production of 1 relative to 2 in a dose-dependent manner, suggesting that promiscuous polyketide synthase processing of malonate- versus α-aminomalonate-derived substrates represents a competitive route for polyketide structural diversification. Metabolites 1 and 2 were active against Bacillus subtilis and Saccharomyces cerevisiae.
Graphical abstract
Natural products from microorganisms have provided various chemical templates of clinically useful lead compounds in the pharmaceutical industry.1,2 However, utilization of traditional microbial sources and discovery techniques have led to the repetitive reisolation of known natural products, resulting in declining discoveries of new chemical diversity.3 Chemical investigation of diverse bacteria–host symbioses has emerged as a powerful approach for the discovery of untapped bioactive molecules within defined ecological contexts.4–9 Many of these molecules play key roles in regulating their complex symbiotic relationships and providing chemical defenses against competing animals and microbes.
Photorhabdus luminescens, an insect pathogenic (entomopathogenic) Gram-negative Gammaproteobacterium, is a bacterial symbiont of Heterorhabditis nematode hosts.10,11 Free-living infective juvenile (IJ) stage nematodes parasitize insect larvae and release their mutualistic bacteria into the insect’s circulatory system. The bacteria then secrete various protein toxins, enzymes, and small molecules that participate in insect virulence, nematode mutualism, and competitor defense. The P. luminescens genome encodes an interesting assortment of known and unknown biosynthetic gene clusters responsible for the synthesis of diverse secondary metabolites.12,13
During the course of our metabolite profiling of P. luminescens in cultivation media supplemented with L-proline, an amino acid nutrient signal that enhances growth and production of some secondary metabolites in P. luminescens,14 we identified a novel phenylpropanoid containing a 2-amino-1,4-benzoquinone moiety, which we named lumiquinone A (1). We also analyzed production of a major LC/MS peak ([M + H]+ m/z 255) corresponding to the known biosynthetically related compound 3,5-dihydroxy-4-isopropyl-trans-stilbene (2). NMR and mass spectral data of 2 were consistent with previously described data for this known Photorhabdus metabolite (Figure S7).15 Here, we describe the isolation, structural characterization, substrate feeding studies, and proposed biosynthesis of 1, which provides new insights for competitive polyketide extender unit incorporation.
Lumiquinone A (1) was obtained as a pale blue pigment with an assigned molecular formula of C17H17NO3 based on HR-ESI-QTOF-MS data (obsd [M + H]+ m/z 284.1289, calcd 284.1287, Figure S1). The UV–vis spectrum of 1 revealed absorption maxima at 204, 336, and 584 nm, suggesting an aminobenzoquinone-like chromophore (Figure 1 and Figure S2).16 The 1H NMR spectrum recorded in chloroform-d1 displayed three distinct aromatic methines [(δH 7.46, 2H, d, J = 7.9 Hz), (δH 7.34, 2H, t, J = 7.3 Hz), and (δH 7.26, 1H, overlapped with chloroform-d1)] and two olefinic proton signals [(δH 7.15, 1H, d, J = 16.7 Hz) and (δH 6.84, 1H, d, J = 16.7 Hz)], suggesting the presence of a monosubstituted phenyl group and one set of (E)-alkenyl hydrogens, respectively (Table 1; NMR spectral data are shown in Figures S3–S6).
Figure 1.
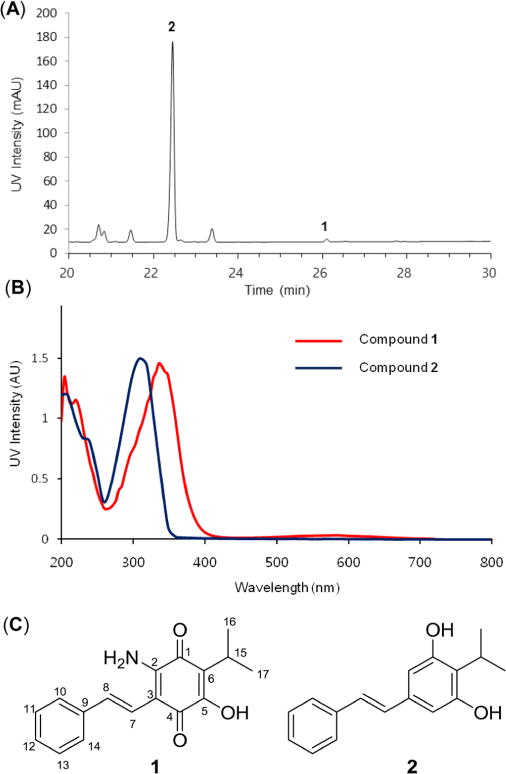
Production of compounds 1 and 2 from P. luminescens TT01. (a) HPLC chromatogram of ethyl acetate-soluble fraction of P. luminescens TT01 detected by 310 nm. (b) UV traces of compounds 1 and 2. (c) Structures of compounds 1 and 2.
Table 1.
NMR Spectral Data of Lumiquinone A (1) in Chloroform-d1 (δH 7.25, δC 77.1)
| no. | δCa | type | δHb | mult (J in Hz) | HMBC |
|---|---|---|---|---|---|
| 1 | 181.8 | C | |||
| 2 | 145.6 | C | |||
| 3 | 105.1 | C | |||
| 4 | 179.1 | C | |||
| 5 | 153.1 | C | |||
| 6 | 120.5 | C | |||
| 7 | 118.2 | CH | 6.84 | d (16.7) | C-2, C-4, C-8, C-9 |
| 8 | 132.4 | CH | 7.15 | d (16.7) | C-3, C-10, C-14 |
| 9 | 137.3 | C | |||
| 10 | 126.2 | CH | 7.46 | d (7.9) | C-8, C-12, C-14 |
| 11 | 128.6 | CH | 7.34 | t (7.3) | C-9, C-13 |
| 12 | 127.8 | CH | 7.26 | mc | C-10, C-14 |
| 13 | 128.6 | CH | 7.34 | t (7.3) | C-9, C-11 |
| 14 | 126.2 | CH | 7.46 | d (7.9) | C-8, C-10, C-12 |
| 15 | 24.1 | CH | 3.17 | m | C-1, C-5, C-6, C-16, C-17 |
| 16 | 19.8 | CH3 | 1.23 | d (7.1) | C-6, C-15, C-17 |
| 17 | 19.8 | CH3 | 1.23 | d (7.1) | C-6, C-15, C-16 |
Chemical shifts of 13C were determined by HSQC and HMBC.
600 MHz.
Overlapping signal.
The phenyl identity was supported by sequential 1H–1H COSY NMR correlations from both H-10 and H-14 at δH 7.46, which were shifted slightly downfield due to the conjugated alkenyl π-bond, through the aromatic methine H-12 at δH 7.26. Establishment of the (E)-2-phenylvinyl moiety was deduced by the analysis of 1H–13C HMBC correlations from both H-10 and H-14 to olefinic carbon C-8 (δC 132.4) and from H-11, H-13, and H-7 (δH 6.84) to quarternary aromatic carbon C-9 (δC 137.3), along with long-range three-bond COSY cross-peaks between olefinic H-8 (δH 7.15) and aromatic methines H-10 and H-14 at the ortho-positions of the phenyl ring (Figure 2). Moreover, the presence of a methine H-15 at δH 3.17 and two methyl signals H-16 and H-17 at δH 1.23 observed in the 1H NMR spectrum suggested another spin system. COSY cross-peaks between H-16, H-17, and H-15 of this spin system allowed the assignment of the isopropyl substituent (Figure 2).
Figure 2.
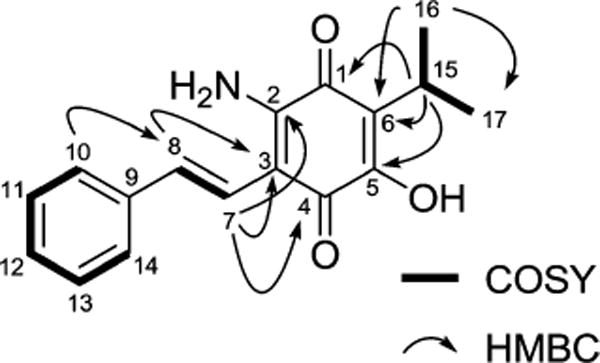
Key COSY and HMBC correlations of 1.
Construction of the 1,4-benzoquinone ring system was established by the HMBC correlations from olefinic H-7 (δH 6.84) to two quaternary carbons, C-2 (δC 145.6) and C-4 (δC 179.1), and from methine H-15 (δH 3.17) to two additional quaternary carbons, C1 (δC 181.8) and C5 (δC 153.1), indicating the presence of a tetrasubstituted 1,4-benzoquinone (Figure 2). Finally, the attachments of primary amine (−NH2) and hydroxy (−OH) functionalities, which are positioned at two deshielded carbons, C-2 and C-5, respectively, were determined by the comparison with 13C chemical shift values of previously reported compounds (Figure S8).17
The novel structure of lumiquinone A (1) consists of an isopropyl-substituted 2-amino-5-hydroxyl-1,4-benzoquinone ring fused with an (E)-2-phenylvinyl group. Within the 1,4-benzoquinone family, our reported structure belongs to the aminobenzoquinone group. The rare isopropyl-substituted aminobenzoquinone moiety has not been reported from microbial sources, but shares some benzoquinone derivatives from plant sources. For example, phytochemical thymoquinone is an isopropyl-substituted benzoquinone member from Nigella savita and harbors antioxidant, analgesic, anticancer, and anticonvulsant effects in animal models.18 Tricyclic abietane diterpenes, such as royleanones,19 are additional isopropyl-substituted benzoquinone members reported from plant Plectranthus species; however, their isopropyl group is derived from an isoprene unit in contrast to the leucine-derived unit in lumiquinone biosynthesis. Moreover, to the best of our knowledge, 1,4-aminobenzoquinones fused with an (E)-2-phenylvinyl group have not been reported for any natural or synthetic molecule described to date.
Compounds 1 and 2 were tested for their antimicrobial activities against several indicator strains, including the Gram-positive bacterium Bacillus subtilis NCIB3610, Gram-negative bacteria Escherichia coli Nissle1917 and their producer Photorhabdus luminescens TT01, and the yeast strain Saccharomyces cerevisiae using paper disk diffusion assays (Figure S9). As described in the literature,20,21 compound 2 revealed inhibitory activities against B. subtilits (zone of inhibition: 10 mm at a concentration of 50 μg/disk) and S. cerevisiae (zone of inhibition: 6 mm at a concentration of 50 μg/disk) as expected, while compound 1 had very weak antibacterial activity against B. subtilis (zone of inhibition: 4 mm at a concentration of 50 μg/disk). Neither 1 nor 2 showed antibacterial activity against the Gram-negative bacteria.
The bacterial biosynthesis of stilbene 2, which is distinct from plant stilbene biosynthesis, proceeds through cyclohexanedione intermediates and was nicely outlined and revised by Bode and co-workers.22,23 Cinnamoyl-CoA and 3-methylbutanoyl-CoA substrates derived from phenylalanine and leucine, respectively, are each subjected to one round of polyketide extension and processing using malonyl-CoA as substrate (Figure 4). In the proposed biosynthesis of stilbene 2, the resulting intermediates are ultimately cyclized by a novel ketosynthase homologue termed DarB (2,5-dialkylresorcinol biosynthetic enzyme) through dual Claisen condensation–Michael addition reactions to liberate a carboxylated cyclohexanedione. Decarboxylation and oxidation by homologue DarA led to aromatic resorcinol 2.22,23 A parallel biosynthetic route incorporating an α-aminomalonate instead of a malonate substrate at either of the polyketide extension sites could explain the biosynthesis of lumiquinone A (1) (Figure 4). Although the proposed aminobenzene intermediate was not detected under the conditions of our experiment, an oxidation event could lead to aminobenzoquinone 1.
Figure 4.
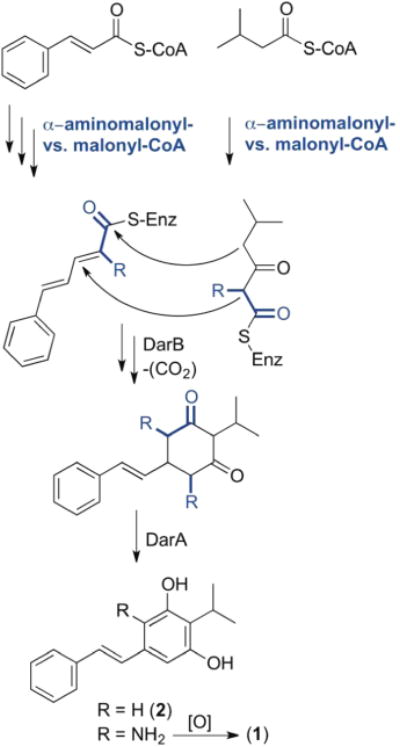
Proposed biosynthesis of lumiquinone A (1) focusing on competitive malonyl- versus α-aminomalonyl-polyketide diversification.
To gain support for our hypothesis, we initially searched the P. luminescens TT01 genome for ZmaG and ZmaI homologues, which sequentially convert a serine-linked carrier protein to an α-aminomalonyl carrier protein polyketide extender unit in zwittermicin biosynthesis.26 However, we did not detect good protein candidates for this conversion. We considered alternative possibilities, such as a direct oxidation of free serine, which could be initiated by a serine dehydrogenase, converting serine to α-aminomalonate semialdehyde. We then searched for homologues of the E. coli serine dehydrogenase27 and tentatively identified P. luminescens serine dehydrogenase Plu2233 (global alignment: identity/similarity/gap, 73.5/83.5/0.4%) clustered directly next to the phenylalanine ammonia lyase (StlA, Plu2234), which converts phenylalanine to cinnamate, initiating stilbene biosynthesis. Oxidation of α-aminomalonate semialdehyde to α-aminomalonate and subsequent activation (α-aminomalonyl-CoA or α-aminomalonyl carrier protein) could be an alternative competitive polyketide extender route for polyketide structural diversification. To test this hypothesis, we supplemented cultures with varying concentrations of free α-aminomalonate and, as expected, observed a dose-dependent stimulation of lumiquinone 1 (Figure 3). In contrast, an inverse relationship for the production of 2 at higher α-aminomalonate concentrations indicated a competitive metabolic divergence point between stilbene 2 and lumiquinone 1. Additionally, we fed 2-13C-labeled malonate to the culture broth of P. luminescens TT01 and analyzed its incorporation in compounds 1 and 2 by HRESI-QTOF-MS (Figure S10). Under our rich medium cultivation conditions, we observed a 6.2% normalized increase for the singly 13C-labeled lumiquinone 1 and an 8.4% increase for the doubly 13C-labeled stilbene 2 compared with a nonlabeled control, further supporting the proposed lumiquinone biosynthesis.
Figure 3.
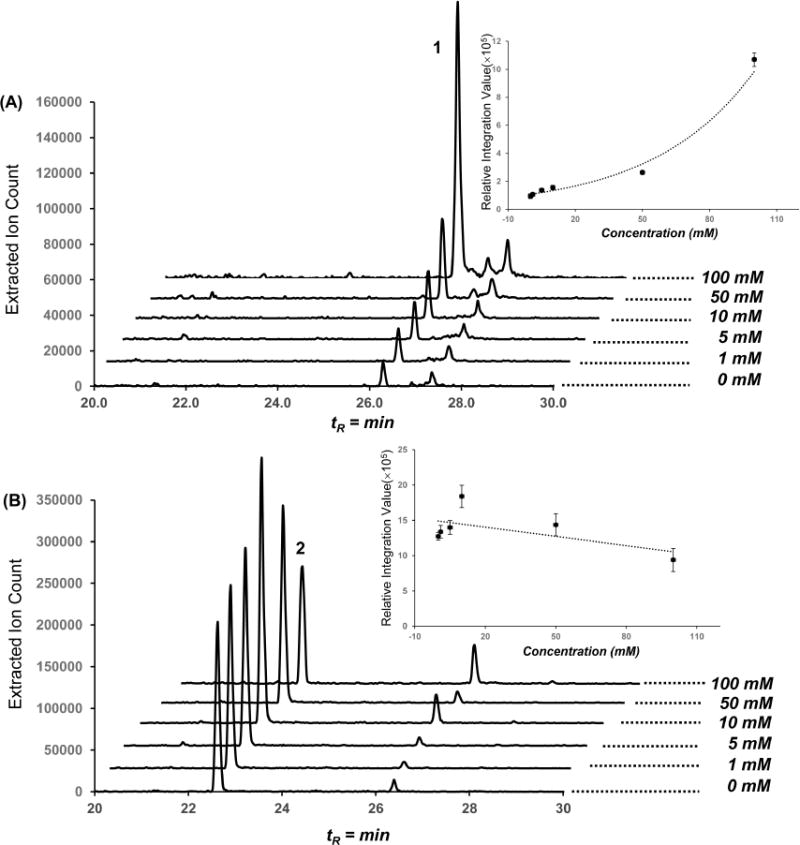
Production of 1 and 2 in dose-dependent 2-aminomalonic acid feeding experiments. HPLC traces with the extracted m/z 284 (A) and m/z 255 (B) ion signals from P. luminescens TT01 culture broth fed with varying concentrations of 2-aminomalonic acid. 2-Aminomalonic acid dose–response curve for 1 and 2 determined by product peak integration values. Error bars represent 95% standard deviation confidence limits.
Biosynthesis of the isopropyl-substituted aminobenzoquinone moiety of lumiquinone A (1) is distinct from the aminobenzoquinones found in geldanamycin and related ansamycin family antibiotics and anticancer molecules. Geldanamycin features an aminobenzoquinone macrocyclic lactam, which is synthesized by a polyketide synthase assembly line initiated with a 3-amino-5-hydroxybenzoic acid (AHBA) starter unit.24 In geldanamycin biosynthesis, the aminobenzoquinone moiety can result from spontaneous oxidation after hydroxylation of the AHBA moiety.25 While the biosynthetic pathways differ, it is likely that a similar spontaneous oxidation could occur in the formation of lumiquinone’s aminobenzoquinone moiety.
The bacterial stilbene biosynthetic pathway has proven to be relatively flexible for product diversification. For example, it was previously demonstrated that P. luminescens TT01 harbored a separate gene cluster for the synthesis of the nonstandard amino acid dihydrophenylalanine, which served as a phenylalanine competitor substrate leading to dihydrostilbene biosynthesis.28,29 By analogy to related systems, these modifications containing readily oxidizable structural features were suggested to contribute to oxidative stress management. Here, competitive α-aminomalonate incorporation activates the resorcinol ring, facilitating its oxidation to the aminobenzoquinone scaffold, and may similarly contribute to oxidative stress reduction. Collectively, our data support that free α-aminomalonate can serve as a competitive extender substrate for polyketide diversification in select metabolic pathways.
EXPERIMENTAL SECTION
General Experimental Procedures
UV/vis spectra were obtained on an Agilent Cary 300 UV/vis spectrophotometer. NMR spectra were obtained on an Agilent 600 MHz NMR system equipped with a cryoprobe. Electrospray ionization mass spectral (ESIMS) data were measured on an Agilent 6120 Quadrupole HPLC/MS system with a Phenomenex Kinetex C18 (2) 5 μm column (4.6 × 250 mm). High-resolution ESIMS data were obtained using an Agilent iFunnel 6550 Q-TOF (quadrupole time-of-flight) MS instrument. Flash column chromatography was performed using Lichroprep RP-18 (40–63 μm, Merck, NJ, USA). The Agilent 1260 Infinity system with a Phenomenex Luna C18 (2) 5 μm column (4.6 × 150 mm) and a photodiode array (PDA) detector was used for sample analysis. Reversed-phase HPLC was performed using an Agilent Prepstar HPLC system with an Agilent Polaris C18-A 5 μm (21.2 × 250 mm) column for sample separation and Phenomenex Luna C18 or C8 (2) 10 μm columns (10.0 × 250 mm) for compound purification.
Small-Scale Cultivation and Metabolite Analysis
Photorhabdus luminescens subsp. laumondii strain TT0130 was grown on Luria–Bertani (LB) agar plates (tryptone 10 g, yeast extract 5 g, sodium chloride 10 g, and agar 18 g per 1 L) at 30 °C for 4 days. A single colony was inoculated into 5 mL of LB broth supplemented with L-proline (72 mM) and incubated at 30 °C for 3 days at 250 rpm. The 5 mL culture broth was extracted with ethyl acetate (2 × 5 mL), and the ethyl acetate-soluble fraction was evaporated under reduced pressure. The organic crude materials, which were dissolved in 200 μL of 100% methanol, were then analyzed by an HPLC/ESIMS system (Phenomenex kinetics C18 column, 250 × 4.6 mm, 5 μm; flow rate: 0.7 mL/min; mobile phase composition: water and acetonitrile containing 0.1% formic acid; 30 min gradient elution from 10% aqueous acetonitrile to 100% acetonitrile, followed by a 5 min isocratic step at 100% acetonitrile). Lumiquinone A (1) and 3,5-dihydroxy-4-isopropyl-trans-stilbene (2) eluted at tR = 26 min ([M + H]+ m/z 284) and at tR = 21 min ([M + H]+ m/z 255), respectively.
Larger Scale Cultivation and Extraction
P. luminescens TT01 was grown in 6 × 5 mL of LB broth supplemented with L-proline (72 mM) at 30 °C for 2 days. The 5 mL cultures were transferred into 6 × 4 L Elenmeyer flasks containing 2 L of the same media to give a total of 12 L of culture broth. After 3 days, the whole culture was extracted with 2 × 12 L of ethyl acetate, and the ethyl acetate-soluble layer was dried in vacuo to yield a crude extract (1.5 g).
Compound Isolation and Purification
The crude extract (1.5 g) was subjected to flash C18 column chromatography with step gradients (40%, 60%, and 80% methanol in water). The 80% methanol fraction was fractionated on a preparative C18 HPLC system (60–100% acetonitrile in water for 30 min), and 1 min fractions were collected using a fraction collector. HPLC fraction 22 was then purified via semipreparative C18 HPLC to give compound 1 (tR = 28.08 min, 0.5 mg) with a gradient system of 10–100% acetonitrile in water for 30 min. HPLC fraction 20 was subjected to C8 HPLC with a gradient system of 60–100% methanol in water for 30 min to yield compound 2 (tR = 25.12 min, 6 mg).
Lumiquinone A (1): pale blue pigment; UV (CH3OH) λmax (log ε) 584 (2.23), 336 (3.92), 204 (3.88) nm; 1H and 13C NMR spectra, see Table 1; HR-ESI-TOF-MS [M + H]+ m/z 284.1289 (calcd for C17H18NO3, 284.1287, 0.7 ppm error).
3,5-Dihydroxy-4-isopropyl-trans-stilbene (2): amorphous solid; 1H NMR (CDCl3, 600 MHz) δ 7.47 (2H, d, J = 7.5 Hz, H-10 and H-14), 7.35 (2H, t, J = 7.5 Hz, H-11 and H-13), 7.25 (1H, t, J = 7.3 Hz, H-12), 6.99 (1H, d, J = 16.0 Hz, H-8), 6.90 (1H, d, J = 16.0 Hz, H-7), 6.50 (2H, s, H-2 and H-4), 4.75 (2H, s, OH), 3.53–3.38 (1H, m, H-15), 1.38 (6H, d, J = 7.0 Hz, H-16 and H-17); positive ESIMS m/z 255 [M + H]+, 277 [M + Na]+.
Paper Disk Test for Antimicrobial Activities
B. subtilis, E. coli, and P. luminescens TT01 were grown in 5 mL of LB broth, and S. cerevisiae was incubated in 5 mL of YM broth (yeast extract 5 g and malt extract 30 g per 1 L distilled water) media at 30 °C for 18 h. The cultures were diluted 1:100 with sterilized water, and 200 μL of culture broths were added to the agar plates. Compounds 1 and 2 (10 μL of each sample with a concentration of 5 mg/mL) were added to sterile paper disks (7 mm). DMSO and ampicillin (1 mg/mL) were used as negative and positive controls, respectively. Paper disks soaked with samples were placed onto LB and YM agar plates for the inhibitory activities against microorganisms and were incubated at 30 °C for 24 h. The zone of inhibition was determined by measuring the distance from the center of the disk to the end of the clear zone.
2-Aminomalonic Acid Feeding Experiment
P. luminescens TT01 was grown in 5 mL of LB liquid media at 30 °C for 24 h. A 25 μL amount of culture broth was then added to 5 mL of fresh LB media supplemented with filter-sterilized 2-aminomalonic acid (Santa Cruz Biotechnology) at five different final culture concentrations (0, 1, 5, 10, 50, and 100 mM) and incubated at 30 °C for 72 h. The growth media was then extracted with ethyl acetate (2 × 5 mL) and dried under reduced pressure. The crude extracts were then analyzed using an HPLC/ESIMS system (Phenomenex kinetics C18 column, 250 × 4.6 mm, 5 μm; flow rate: 0.7 mL/min; mobile phase composition: water and acetonitrile containing 0.1% formic acid; 30 min gradient elution from 10% aqueous acetonitrile to 100% acetonitrile, followed by a 5 min isocratic step at 100% acetonitrile). The feeding experiments were conducted in triplicate.
Supplementary Material
Acknowledgments
This work was supported by the National Institutes of Health (National Institute of General Medical Sciences Grant R00-GM097096). We also thank the Searle Scholars Program for general laboratory support (Grant 13-SSP-210).
Footnotes
Supporting Information
Supplementary figures include NMR (1H, gCOSY, gHSQC, and gHMBC), HR-ESI-TOF-MS, and UV spectral data for new compound 1. These are available free of charge via the Internet at http://pubs.acs.org.
Notes
The authors declare no competing financial interest.
References
- 1.Cragg GM, Newman DJ. Biochim Biophys Acta Gen Subj. 2013;1830:3670–3695. doi: 10.1016/j.bbagen.2013.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Butler MS, Robertson AAB, Cooper MA. Nat Prod Rep. 2014;31:1612–1661. doi: 10.1039/c4np00064a. [DOI] [PubMed] [Google Scholar]
- 3.Pettit RK. Appl Microbiol Biotechnol. 2009;83:19–25. doi: 10.1007/s00253-009-1916-9. [DOI] [PubMed] [Google Scholar]
- 4.Crawford JM, Clardy J. Chem Commun. 2011;47:7559–7566. doi: 10.1039/c1cc11574j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hentschel U, Piel J, Degnan SM, Taylor MW. Nat Rev Microbiol. 2012;10:641–654. doi: 10.1038/nrmicro2839. [DOI] [PubMed] [Google Scholar]
- 6.Schmidt EW, Donia MS, McIntosh JA, Fricke WF, Ravel J. J Nat Prod. 2012;75:295–304. doi: 10.1021/np200665k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rath CM, Dorrestein PC. Curr Opin Microbiol. 2012;15:147–154. doi: 10.1016/j.mib.2011.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Scherlach K, Graupner K, Hertweck C. Annu Rev Microbiol. 2013;67:375–397. doi: 10.1146/annurev-micro-092412-155702. [DOI] [PubMed] [Google Scholar]
- 9.Ramadhar TR, Beemelmanns C, Currie CR, Clardy J. J Antibiot. 2014;67:53–58. doi: 10.1038/ja.2013.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Waterfield NR, Ciche T, Clarke D. Annu Rev Microbiol. 2009;63:557–574. doi: 10.1146/annurev.micro.091208.073507. [DOI] [PubMed] [Google Scholar]
- 11.Murfin KE, Dillman AR, Foster JM, Bulgheresi S, Slatko BE, Sternberg PW, Goodrich-Blair H. Biol Bull. 2012;223:85–102. doi: 10.1086/BBLv223n1p85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brachmann AO, Bode HB. Adv Biochem Eng Biotechnol. 2013;135:123–155. doi: 10.1007/10_2013_192. [DOI] [PubMed] [Google Scholar]
- 13.Vizcaino MI, Guo X, Crawford JM. J Ind Microbiol Biotechnol. 2014;41:285–299. doi: 10.1007/s10295-013-1356-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Crawford JM, Kontnik R, Clardy J. Curr Biol. 2010;20:69–74. doi: 10.1016/j.cub.2009.10.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Paul VJ, Frautschy S, Fenical W, Nealson KH. J Chem Ecol. 1981;7:589–597. doi: 10.1007/BF00987707. [DOI] [PubMed] [Google Scholar]
- 16.Muroi M, Tanida S, Asai M, Kishi T. J Antibiot. 1980;33:1449–1456. doi: 10.7164/antibiotics.33.1449. [DOI] [PubMed] [Google Scholar]
- 17.Enríquez RG, Ortiz B, Alducin E, Walls F, Gnecco D, Yu M, Reynolds WF. Nat Prod Lett. 1995;6:103–109. [Google Scholar]
- 18.Gali-Muhtasib H, Roessner A, Schneider-Stock R. Int J Biochem Cell Biol. 2006;38:1249–1253. doi: 10.1016/j.biocel.2005.10.009. [DOI] [PubMed] [Google Scholar]
- 19.Rijo P, Duarte A, Francisco AP, Semedo-Lemsaddek T, Simões MF. Phytother Res. 2014;28:76–81. doi: 10.1002/ptr.4961. [DOI] [PubMed] [Google Scholar]
- 20.Hu K, Webster JM. FEMS Microbiol Lett. 2000;189:219–223. doi: 10.1111/j.1574-6968.2000.tb09234.x. [DOI] [PubMed] [Google Scholar]
- 21.Kumar SN, Nambisan B. Appl Biochem Biotechnol. 2014;172:741–754. doi: 10.1007/s12010-013-0567-6. [DOI] [PubMed] [Google Scholar]
- 22.Joyce SA, Brachmann AO, Glazer I, Lango L, Schwar G, Clarke DJ, Bode HB. Angew Chem, Int Ed. 2008;47:1942–1945. doi: 10.1002/anie.200705148. [DOI] [PubMed] [Google Scholar]
- 23.Fuchs SW, Bozhüyük KAJ, Kresovic D, Grundmann F, Dill V, Brachmann AO, Waterfield NR, Bode HB. Angew Chem, Int Ed. 2013;52:4108–4112. doi: 10.1002/anie.201210116. [DOI] [PubMed] [Google Scholar]
- 24.Kim CG, Kirschning A, Bergon P, Zhou P, Su E, Sauerbrei B, Ning S, Ahn Y, Breuer M, Leistner E, Floss HG. J Am Chem Soc. 1996;118:7486–7491. [Google Scholar]
- 25.Lee D, Lee K, Cai XF, Dat NT, Boovanahalli SK, Lee M, Shin JC, Kim W, Jeong JK, Lee JS, Lee CH, Lee JH, Hong YS, Lee J. J ChemBioChem. 2006;7:246–248. doi: 10.1002/cbic.200500441. [DOI] [PubMed] [Google Scholar]
- 26.Chan YA, Boyne MT, 2nd, Podevels AM, Klimowicz AK, Handelsman J, Kelleher NL, Thomas MG. Proc Natl Acad Sci USA. 2006;103:14349–14354. doi: 10.1073/pnas.0603748103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yamazawa R, Nakajima Y, Mushiake K, Yoshimoto T, Ito K. J Biochem. 2011;149:701–712. doi: 10.1093/jb/mvr024. [DOI] [PubMed] [Google Scholar]
- 28.Kontnik R, Crawford JM, Clardy J. ACS Chem Biol. 2010;5:659–665. doi: 10.1021/cb100117k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Crawford JM, Mahlstedt SA, Malcolmson SJ, Clardy J, Walsh CT. Chem Biol. 2011;18:1102–1112. doi: 10.1016/j.chembiol.2011.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fischer-Le Saux M, Viallard V, Brunel B, Normand P, Boemare NE. Int J Syst Bacteriol. 1999;49(Pt 4):1645–1656. doi: 10.1099/00207713-49-4-1645. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.