

Abstract
Platelets are central mediators of thrombosis and hemostasis. At the site of vascular injury, platelet accumulation (i.e. adhesion and aggregation) constitutes the first wave of hemostasis. Blood coagulation, initiated by the coagulation cascades, is the second wave of thrombin generation and enhance phosphatidylserine exposure, can markedly potentiate cell-based thrombin generation and enhance blood coagulation. Recently, deposition of plasma fibronectin and other proteins onto the injured vessel wall has been identified as a new “protein wave of hemostasis” that occurs prior to platelet accumulation (i.e. the classical first wave of hemostasis). These three waves of hemostasis, in the event of atherosclerotic plaque rupture, may turn pathogenic, and cause uncontrolled vessel occlusion and thrombotic disorders (e.g. heart attack and stroke). Current anti-platelet therapies have significantly reduced cardiovascular mortality, however, on-treatment thrombotic events, thrombocytopenia, and bleeding complications are still major concerns that continue to motivate innovation and drive therapeutic advances. Emerging evidence has brought platelet adhesion molecules back into the spotlight as targets for the development of novel anti-thrombotic agents. These potential antiplatelet targets mainly include the platelet receptors glycoprotein (GP) Ib-IX-V complex, β3 integrins (αIIb subunit and PSI domain of β3 subunit) and GPVI. Numerous efforts have been made aiming to balance the efficacy of inhibiting thrombosis without compromising hemostasis. This mini-review will update the mechanisms of thrombosis and the current state of antiplatelet therapies, and will focus on platelet adhesion molecules and the novel anti-thrombotic therapies that target them.
Keywords: αIIbβ3, Anfibatide, GPIbα, GPVI, Hemostasis, Integrins, P-selectin, Stroke, Thrombosis
Background
Platelet adhesion, activation and aggregation are critical events in hemostasis and thrombosis [1–3]. Platelet adhesion molecules, αIIbβ3 integrin and the glycoprotein (GP) Ib-IX-V, are essential for these processes [4–6]. Other adhesion molecules, such as P-selectin, GPVI and cadherins, are also involved [7–10]. The important roles of adhesion molecules in normal hemostasis have been well demonstrated in bleeding disorders, for example, Glanzmann thrombasthenia (β3 integrin deficiency) [11] and Bernard-Soulier syndrome (GPIb-IX-V complex deficiency) [12]. However, under pathological conditions, excessive platelet function may lead to thrombotic diseases, such as myocardial infarction and ischemic stroke, which cause far more deaths each year than cancer or respiratory diseases [1, 2, 13–15]. Therefore, antiplatelet agents are vital for the treatment of thrombosis [16]. For over a decade, dual antiplatelet therapy with clopidogrel and aspirin has been considered a key treatment of patients with acute coronary syndrome [17, 18]. Nonetheless, some patients undergoing this combination therapy continue to suffer from recurrent thrombotic events, likely a result of platelet activation and aggregation occurring independently of ADP or thromboxane A2 receptor-mediated signalling pathways [17]. Thus, attenuating platelet adhesion appears to be a desirable strategy in effectively controlling pathological thrombosis [18]. Further understanding of the interactions between platelet adhesion molecules and their binding partners is therefore crucial in developing novel anti-thrombotic therapies. This review briefly summarizes the current knowledge of thrombosis and antiplatelet therapies, introduces a number of major platelet adhesion molecules, and highlights some recent advances in the new mechanisms of thrombosis, and anti-thrombotic therapies that are in clinical trials (unless otherwise indicated). There are several excellent available reviews regarding antiplatelet therapies, such as ADP antagonists (e.g. P2Y12 inhibitors), thromboxane antagonists and PAR-1/4 inhibitors [17, 18]. This mini-review will mainly focus on the therapeutic developments targeting platelet adhesion molecules.
Review
Arterial thrombosis and current state of antiplatelet therapies
Arterial thrombosis at the site of atherosclerotic plaque rupture may lead to uncontrolled vessel occlusion, resulting in life-threatening consequences (e.g. unstable angina, myocardial infarction and ischemic stroke) [1, 2, 13]. During plaque rupture, subendothelial matrix proteins, like collagen, von Willebrand factor (VWF), fibrinogen, fibronectin and laminin are exposed to circulation, leading to the rapid response of platelets [6]. Inappropriate platelet adhesion, activation and aggregation promote excessive platelet plug formation. Activated platelets can also provide negatively-charged surfaces that harbor coagulation factors and markedly potentiate cell-based thrombin generation and blood coagulation [1, 2, 19, 20]. The evolving concept of the “protein wave of hemostasis” indicates a potential role of platelet-released plasma fibronectin in thrombosis and hemostasis [21, 22]. Thus, platelets are key mediators of atherothrombosis, which are actively involved in all three waves of thrombus formation: protein wave, platelet accumulation, and blood coagulation [21, 23].
Current FDA-approved antiplatelet therapies (Fig. 1) mainly aim to (i) inhibit thromboxane A2 synthesis, which inhibits platelet activation (e.g. aspirin and triflusal); (ii) antagonize the function of platelet P2Y12 receptors, (e.g. clopidogrel, prasugrel, and ticagrelor); (iii) inhibit platelet integrin αIIbβ3 activity, which inhibits platelet aggregation, (e.g. abciximab, eptifibatide, and tirofiban); (iv) inhibit phosphodiesterase, which increases platelet cAMP/cGMP levels (e.g. dipyridamole and cilostazol) [24]. These antiplatelet drugs have significantly reduced cardiovascular deaths. However, limitations of current therapies, such as weak/poor inhibition of platelet function, excessive bleeding, thrombocytopenia and unexpected platelet activation are concerns that drive therapeutic advances [18, 25, 26]. In 2014, the FDA approved Vorapaxar, a novel antagonist of the thrombin receptor protease-activated receptor 1 (PAR1), which reduces the risk of heart attack and stroke in patients with atherosclerosis or peripheral arterial disease [27, 28]. However, Vorapaxar must not be used in patients who have histories of stroke, transient ischemic attack (TIA) or intracranial hemorrhage, since it increases the risk of intracranial bleeding [28, 29].
Fig. 1.
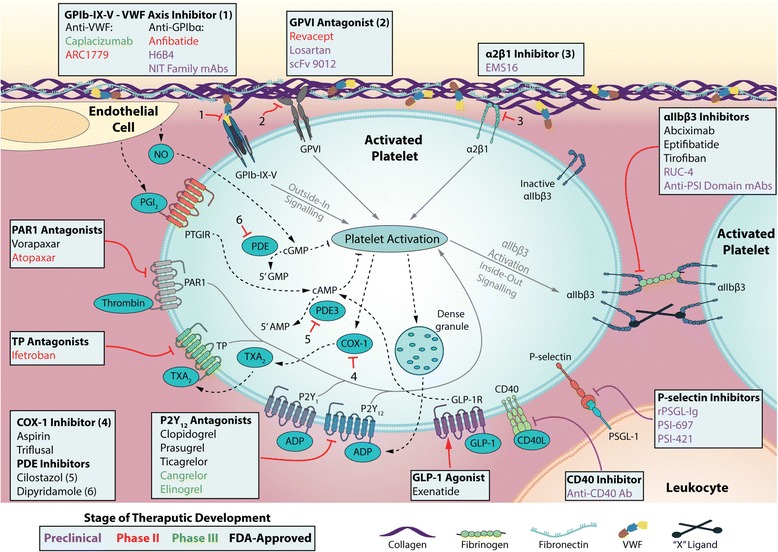
Current and novel antiplatlet therapies. Platelet adhesion to an injury site at a vessel wall is mediated by the exposure and binding of subendothelial matrix proteins (e.g. collagen, VWF, fibrinogen, and fibronectin) to glycoprotein (GP) receptors on the platelet surface. VWF binding to the GPIb-IX-V complex, collagen binding to platelet GPVI and integrin α2β1 receptors trigger a signal transduction process resulting in the local release of platelet activation agonists, such as thromboxane A2 and ADP. These agonists along with thrombin produced from coagulation cascades and activated platelets, bind to platelet surface bound G-coupled receptors inducing further platelet activation. Activation of platelet integrin αIIbβ3 induces platelet aggregation mediated by fibrinogen/VWF or the yet undetermined “X” ligands. Leukocyte-platelet adhesion can be driven by the interaction between platelet surface P-selectin and its counter-receptor PSGL-1 situated upon the leukocyte surface. Inhibition of platelet activation is mainly mediated by the PDE/PDE3 regulated degradation and PGI2, NO and GLP-1R regulated activation of cGMP or cAMP. Direct and indirect antithrombotic therapeutics are tabulated in the light colored boxes within the figure. The actions of antithrombotic therapies are depicted using red arrows, and some indirect antithrombotic agents (such as anti-atherosclerotic agents) are represented with purple arrows. Therapeutics, to name a few, listed in black, green, red and purple correspond to FDA-approved, phase III, phase II or preclinical development status, respectively. Numbered inhibitory arrows represent the actions of the correspondingly numbered therapies. Some other anti-platelet agents are not included, more information can be found in references 17, 18 and other publications. Abbreviations: COX-1 cyclooxygenase 1 GLP-1 glucagon-like peptide 1, GLP-1R glucagon-like peptide 1 receptor, PAR protease-activated receptor, PDE phosphodiesterase, PSGL-1 P-selectin glycoprotein ligand 1, TP thromboxane prostanoid receptor, TXA 2 thromboxane A2; VWF von Willebrand factor
Platelet adhesion molecules in hemostasis and thrombosis: novel mechanisms and therapeutic opportunities
Platelet adhesion molecules are proteins/receptors on the platelet surface that interact with other cells or the extracellular matrix, including the integrin family (e.g. α2β1, α5β1, α6β1, αLβ2, αIIbβ3, and ανβ3) [4, 30, 31], the immunoglobulin superfamily (e.g. GPVI, FcγRIIA, ICAM-2, PECAM-1, JAMs and Cadherin 6), the leucine-rich repeat family (LRR; e.g. GPIb-IX-V complex), and the C-type lectin receptor family (e.g. P-selectin and CLEC-2), etc. [32–34]. Recent evidence has shown that platelet adhesion molecules play key roles in a variety of pathophysiological processes [23], such as hemostasis and thrombosis [4, 33], immune responses [35, 36], inflammation [35–37], atherosclerosis [38–40], lymphatic vessel development [41–44], angiogenesis [45–47], miscarriage [48, 49], and tumor metastasis [50–52]. Platelets are versatile cells and the mechanisms of their diverse functions have emerged as hot research topics [23]. This review mainly focuses on their roles in thrombosis and as novel anti-thrombotic targets (Fig. 1).
The GPIb-IX-V complex: emerging targets of antiplatelet therapy
New insights into the GPIb-IX-V complex
Platelet GPIb-IX-V complex (LRR family protein) has approximately 50,000 copies/platelet. It is composed of one GPIbα subunit disulfide-linked to two molecules of GPIbβ, and non-covalently linked with GPIX and GPV in a 2:4:2:1 ratio [53]. GPIb-IX-V is a key platelet receptor in initiating platelet translocation and adhesion to the vessel wall during vascular injury, especially under high shear stress (e.g. in small or stenosed arteries) [54, 55]. Platelet translocation onto the subendothelium is mediated by the binding of GPIbα to the immobilized VWF, a multimeric adhesive protein secreted from activated endothelial cells and platelets. The crystal structure of the GPIbα N-terminal ligand-binding domain and the VWF A1 domain gives useful information regarding their interaction [56]. This interaction induces intracellular signalling events that can activate integrins, leading to platelet stable adhesion and subsequent platelet aggregation. Interestingly, platelet-derived VWF was recently shown not essential for hemostasis and thrombosis, but instead fosters thrombo-inflammatory diseases such as ischemic stroke in mice via a GPIb-dependent mechanism [57]. This suggests that targeting GPIbα-VWF may be a promising anti-thrombotic strategy, particularly in thrombo-inflammatory conditions.
Furthermore, GPIb-IX-V complex has a high affinity for thrombin [58, 59]. Two thrombin binding sites on GPIbα LRR C-terminal flank region have been revealed [58]. Consequently, thrombin can activate platelets via GPIbα in two ways [60]: accelerating the cleavage of PAR-1 and platelet activation [61], or direct signaling via GPIbα, particularly after cleaving GPV, which is generally considered a “brake” in GPIb-IX-V activation [62, 63]. It is currently unknown but it is reasonable to consider that targeting both VWF and thrombin binding sites of GPIbα might provide additional benefits in effectively controlling thrombosis.
GPIbα can also interact with multiple other ligands, leading to platelet activation (e.g. thrombospondin [64] and P-selectin), pro-coagulant activity (e.g. factors XI [65], XII [66], VIIa [67] and kininogen [68]), inflammatory responses (e.g. P-selectin [69, 70], αMβ2 [71]), arterial remodeling [72] and others. Recently, the antibody-GPIbα interaction in immune thrombocytopenia has been highlighted. Some anti-GPIbα antibodies cause platelet activation and desialylation (removal of sugars), followed by the clearance of desialylated platelets via Ashwell-Morell receptors on hepatocytes [73, 74].
Developing novel antiplatelet agents against GPIbα
Given the critical roles of GPIbα or GPIbα-VWF interactions in platelet adhesion, particularly under stenosis high-shear conditions, they are attractive targets in attenuating thrombosis [54, 75, 76]. Currently, two such agents are in active clinical trials. ALX-0081 (Caplacizumab), an anti-VWF humanized single-variable-domain immunoglobulin (Nanobody), binds to the A1 domains of VWF with high affinity [77]. The phase I and II clinical trials of ALX-0081 in patients with stable angina undergoing percutaneous coronary intervention (PCI) or high risk PCI patients have shown a promising antiplatelet effects, and a relatively safe profile [77, 78]. The phase III clinical trials will investigate its effects on acquired thrombotic thrombocytopenic purpura (TTP) [79–81]. ARC1779, an anti-VWF aptamer, was previously reported as an encouraging agent; however, the clinical trial of ARC1779 was prematurely terminated [82]. These VWF inhibitors may be useful candidates for TTP treatment.
A direct anti-GPIbα drug, Anfibatide, is purified from the snake venom of Agkistrodon acutus [83, 84]. Notably, Anfibatide inhibits both VWF and α-thrombin binding to GPIbα, representing a more potent anti-thrombotic effect [85]. In experimental models, Anfibatide inhibited platelet adhesion, aggregation and thrombus formation, without increasing bleeding time [83]. The phase II human clinical trials have also shown the promise of Anfibatide being utilized as a novel antiplatelet agent in cardiovascular diseases without significantly affecting hemostasis in patients with non-ST segment elevation myocardial infarction (unpublished data) [85]. Additionally, anti-GPIbα antibody displayed a strong protective effect in the mouse stroke models without inducing significant intracranial bleeding [86–88]. Anfibatide has also been shown as a candidate to treat ischemic stroke in experimental models [89] (the same may hold true for anti-VWF therapy) and deserves further investigation. There are some other preclinical agents targeting GPIbα that are under investigation, such as h6B4-Fab [90], GPG-290 [91], and anti-GPIbα NIT family monoclonal antibodies [92]. The generation of these novel antagonists is reaching the forefront of treatment against heart attack and stroke, although the efficacy and safety of these drugs remain to be further established or evaluated in human clinical trials. Notably, there are currently no clinically available direct GPIbα antagonists.
GPVI: a potential anti-thrombotic target
GPVI (immunoglobulin superfamily protein) is exclusively expressed on platelets and megakaryocytes. It is associated with the Fc receptor γ-chain, which contains an immunoreceptor tyrosine-based activation motif (ITAM). Cross-linking by ligands, such as collagen, leads to ITAM-dependent signalling, and platelet activation. A possible anti-thrombotic benefit of targeting PI3-kinase/Akt pathway on ITAM receptors was suggested [93]. Fibrin has also been identified as a new GPVI ligand [94]. The GPVI ectodomain interacts with immobilized fibrin, which amplifies thrombin generation, and promotes thrombus stabilization [94, 95].
The role of platelet GPVI in the pathogenesis of ischemic stroke has been gradually acknowledged [96–98]. Notably, platelet adhesion/activation can enhance infarct growth by promoting an inflammatory response [88, 99, 100]. GPVI-mediated platelet activation can lead to the release of interleukin-1α that drives cerebrovascular inflammation [100]. GPVI may be thus a potential antiplatelet target [97, 101, 102]. In animal models, anti-GPVI protected against thrombosis, ischemia-reperfusion injury [103] and stroke [104]. In phase I clinical trials, Revacept (the humanized Fc fusion protein of the GPVI ectodomain), inhibited collagen-induced human platelet aggregation [105]. Phase II trials of Revacept in patients with carotid artery stenosis, TIA, or stroke are ongoing [106]. The efficacy and safety of Revacept in these patients will be further determined. Some other GPVI targeted agents that are under preclinical development, such as Losartan [107] and scFv9012 [108], have been shown to inhibit the binding of GPVI to collagen.
Platelet integrin receptors
Integrins are heterodimeric transmembrane receptors, which are involved in cell-cell and cell-matrix interactions [30]. There are six different integrins on platelet surfaces: α2β1, α5β1, α6β1, αLβ2, αIIbβ3, and ανβ3. Platelet integrin αIIbβ3 is the dominant integrin expressed on platelets. Given the critical roles of αIIbβ3 integrin in mediating platelet aggregation, αIIbβ3 antagonists have been widely used for nearly two decades.
Integrin αIIbβ3 as anti-thrombotic targets: lessons and opportunities
Approximately 17 % of total platelet surface proteins are αIIbβ3 integrin, which contains both αIIb and β3 subunits [4]. Platelet “outside-in” signals are induced following platelet adhesion and platelet activation (e.g. GPIbα-VWF, GPVI/α2β1-collagen, P2Y12-ADP, PARs-thrombin), resulting in an increased Ca2+ influx and ultimately “inside-out” signaling. These “inside-out” signals further drive the conformational changes of αIIbβ3, from a low to high affinity state for binding to its ligands (e.g. fibrinogen/fibrin, VWF, fibronectin, thrombospondin, vitronectin and unidentified “X” ligands) [109–112].
Fibrinogen, a major prothrombotic ligand of αIIbβ3, has been documented to be required for platelet aggregation for over 50 years. However, platelet aggregation still occurs in the absence of fibrinogen and VWF, although in the absence of αIIbβ3, aggregation is abolished [5, 8, 21, 113–116]. The discovery of “fibrinogen-independent platelet aggregation” demonstrates that unidentified αIIbβ3 ligands also mediate platelet aggregation [5, 8, 21, 113, 116], and have the potential to be novel anti-thrombotic targets. Interestingly, some ligands (e.g. plasma fibronectin, vitronectin) may block prothrombotic ligand (e.g. fibrinogen)-αIIbβ3 interactions and attenuate thrombosis [21, 117].
Three FDA-approved αIIbβ3 antagonists are available: Abciximab (ReoPro), Eptifibatide (Integrilin) and Tirofiban (Aggrastat) [118–120]. Abciximab is a fragmented antibody that binds close to the ligand binding-pocket on αIIbβ3. Eptifibatide, isolated from snake venom, binds via a KGD sequence and is a competitive inhibitor for fibrinogen-αIIbβ3, whilst tirofiban is a small molecule RGD inhibitor. Currently, αIIbβ3 antagonists are used in patients undergoing PCI and significantly decrease the incidence of myocardial infarction and death [121]. However, these antagonists can induce further conformational changes in the β3 subunit that may have negative consequences, such as exposing previously hidden epitopes, and causing platelet activation [122]. αIIbβ3 antagonists are also associated with intracranial hemorrhage in patients with acute ischemic stroke [123]. Therefore, a safer and more specific on-target drug is required to provide better patient care. Recently, a novel αIIbβ3 antagonist, RUC-4 (a more potent and more soluble congener of RUC-2 that disrupts Mg2+ binding to the metal ion-dependent adhesion site of αIIbβ3), is suggested for prehospital therapy of myocardial infarction in animal models, without significantly priming the receptor to bind fibrinogen [124]. However, the possibility of increased bleeding with therapeutic doses of RUC-4 remains to be evaluated [124].
The plexin-semaphorin-integrin (PSI) domain, located near the N-terminus of the β3 subunit, is highly conserved across the integrin family in different species, and contains seven cysteine residues which have been implicated in regulating β2 integrin activation [125, 126]. Previous studies described a role for cysteine-derived thiol/disulfide groups in the conformational switches of the β3 integrin [127–130]. Disulfide bond remodeling is mediated primarily by thiol isomerase enzymatic activity, which is derived from active CXXC thioredoxin motifs and plays a role in the activation of αIIbβ3 [131]. Our group has recently identified that integrin PSI domain has endogenous thiol isomerase function and could be a novel target for anti-thrombotic therapy (unpublished data) [132]. We found that both CXXC motifs of β3 integrin PSI domain are required to maintain the optimal enzyme function, since mutations to one or both of the CXXC motifs decrease or abolish their protein disulphide isomerase (PDI)-like activity. We developed anti-PSI monoclonal antibodies and found that these antibodies cross-reacted with β3 PSI domains of human and other species and specifically inhibited the PDI-like activity, integrin activation and reduced PAC-1 binding to β3 integrin. Importantly, anti-PSI abrogated murine and human platelet aggregation in vitro and thrombus growth ex vivo and in vivo in both small and large vessels without significantly affecting bleeding time or platelet count. Thus, integrin PSI domain contains endogenous PDI activity and is a key regulator of integrin activation that can be a new target for therapy.
Interestingly, targeting activated platelets αIIbβ3 has been considered into the development of novel fibrinolytic drugs, which may allow effective thrombolysis and thromboprophylaxis [14, 133]. For example, scFvSCE5 (a single-chain urokinase plasminogen activator fused to a small recombinant antibody that binds activated αIIbβ3) directly targets thrombi and exerts an effective thrombolysis [133]. A chimeric platelet-targeted urokinase prodrug (composed of a single-chain version of the variable region of an anti-αIIbβ3 mAb and a thrombin-activatable, low-molecular-weight pro-uPA) selectively targets new thrombus formation [134].
Other platelet integrins: α2β1, α6β1 and α5β1
Other integrin receptors may also be considered as novel anti-thrombotic targets [16, 135]. Platelet α2β1 promotes stable platelet adhesion to collagen and may be a viable option, since overexpression of α2β1 in humans increases atherothrombotic risk, but lower level of α2β1 does not enhance bleeding risk [16]. Experimental evidence shows that α2β1 inhibitors (e.g. snake venom EMS-16) reduced pathological thrombus formation in vivo [136–138]. Platelet α6β1, the main receptor for laminin, plays a role in platelet adhesion/activation and arterial thrombosis, and may also be a new target [135]. Platelet α5β1, the major receptor for fibronectin, plays a supplementary role in platelet adhesion [139], but evidence is lacking regarding the anti-thrombotic benefits of antagonizing α5β1.
Other novel anti-thrombotic candidates: Glucagon-like peptide 1 receptor, P-selectin, CD40/CD40L, and Toll-like receptors
Strategies to target other platelet receptors beyond adhesive proteins have also been developed, such as P2Y12, PAR1, TP, 5HT2A antagonists [17, 140]. Interestingly, some chronic diseases, such as diabetes mellitus and atherosclerosis, are associated with arterial thrombosis [23, 141]. Recently, our group identified that a functional Glucagon-like peptide 1 receptor (GLP-1R) is expressed on human megakaryocytes and platelets [142]. Importantly, GLP-1R agonists (e.g. Exenatide), likely through increasing the intracellular cAMP levels, inhibit platelet function and thrombus formation [142]. This study provides important insights into why diabetic patients who are receiving GLP-1-targeted therapies have a reduced number of cardiovascular events [142, 143]. In addition, given the cross-talks between platelets and immune systems, thrombosis also intensively communicates with the inflammatory pathway [23]. Some anti-inflammatory/anti-atherosclerotic agents may therefore also indirectly inhibit thrombosis, especially in deep vein thrombosis [144]. For example, antagonists of P-selectin/PSGL-1, such as rPSGL-Ig [145], PSI-697 [146], PSI-421 [147], inhibit platelet-mediated leukocyte attachment and recruitment of procoagulant microparticles, and may represent a safe therapeutic intervention in accelerating thrombolysis [148]. Antagonists of CD40/CD40L [149], such as CD40 antibody, reduce atherosclerotic burden in a murine model [150]. In addition, as the important roles of Toll-like receptors in atherosclerosis are gradually recognized [151, 152], they may also be potential targets for the treatment of atherothrombosis.
Conclusions
Arterial thrombotic events, such as myocardial infarction and ischemic stroke, and venous thromboembolism, are three leading causes of morbidity and mortality worldwide [153]. Platelets play a central role in the pathogenesis of atherothrombosis, and contribute profoundly to the pathology of venous thrombosis [23]. Platelet adhesion molecules, act as the contacts between platelets and other cells or extracellular matrix proteins and, to a great extent, may determine the reactivity of platelets and thus are attractive anti-thrombotic targets (Fig. 1) [23]. Although evidence-based antiplatelet therapy has markedly improved patient care, on-treatment events and bleeding are still major concerns [17, 148].
Optimization of the use of currently available therapies, and improvements to the understanding of individual differences in response to anti-platelet treatments are still the most cost-effective treatment strategies [17, 148]. Additionally, improved understanding of the mechanisms of platelet accumulation has been critical for further developing novel antiplatelet therapies, such as the PAR1 antagonist Vorapaxar (recently approved by the FDA), GPIbα/VWF antagonists (e.g. ALX-0081 and Anfibatide; undergoing clinical trials), and GPVI antagonist (e.g. Revacept; undergoing clinical trials) (See section II. A-C). Another cost-effective strategy may be to repurpose already-established drugs by discovering novel mechanisms of action in anti-thrombotic diseases, such as the recently-identified GLP-1R agonist, Exenatide, an anti-diabetic drug that has potential anti-thrombotic effects [142, 154]. Future studies in the areas of atherothrombosis, inflammation, metabolic syndrome, diabetes, lipid metabolism and cancer-related thrombotic diseases in the next few years should advance our knowledge and the application of these and other new anti-platelet agents. Of note, clinical trials provide important evidence regarding the safety and efficacy of the treatments. However, difficulties such as narrow eligibility criteria, low enrollment of patients and the necessity to test the new drugs on top of the current dual antiplatelet therapy (e.g. aspirin and clopidogrel), may add complexity to the development of new drugs and also deserve our attention.
Acknowledgements
The authors would like to thank Dr. Richard O. Hynes, Dr. Zaverio M. Ruggeri, Dr. Denisa D. Wagner, and Dr. John Freedman for their long-term support for these research projects.
Declaration
Publication fees for this article have been funded by APSTH 2016.
This article has been published as part of Thrombosis Journal Volume 14 Supplement 1, 2016. The full contents of the supplement are available at https://thrombosisjournal.biomedcentral.com/articles/supplements/volume-14-supplement-1.
Funding
This work was supported by the Canadian Institutes of Health Research (MOP 119540, MOP 97918, and MOP 119551), Heart and Stroke Foundation of Canada (Ontario), Equipment Funds from Canada Foundation for Innovation, St. Michael’s Hospital, and Canadian Blood Services; and research Funds from CCOA Therapeutics Inc and Lee’s Pharmaceutical Holdings limited. X. R. X is a recipient of China National Scholarship award, Meredith & Malcolm Silver Scholarship in Cardiovascular Studies of Department of Laboratory Medicine and Pathobiology, and the Heart and Stroke/Richard Lewar Centre of Excellence Studentship award, University of Toronto. N.C. is a recipient of the Canadian Blood Services Postdoctoral Fellowship.
Availability of data and material
Not applicable.
Authors’ contributions
XRX and NC drafted the manuscript. MADN drew the figure. TM, TWS and RMPC contributed to preparation of the manuscript. XL, XD and BXL contributed to the original findings on the phase II human clinical trials of Anfibatide. PC and JX contributed to the original findings and further development of anti-PSI monoclonal antibodies. HN is the principal investigator who defined the topic and revised the manuscript. All of authors read, commented and approved the final manuscript.
Competing interests
J. X. is supported by the CCOA Therapeutics Inc. X. D. and B.X.L. are supported by the Lee’s Pharmaceutical Holdings limited. Some of the research fund of the projects is supported by CCOA Therapeutics and Lee’s Pharmaceutical Holdings limited. Canadian Blood Services have held the patents on the anti-GPIb NIT family monoclonal antibodies and anti-PSI monoclonal antibodies.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- ADP
Adenosine diphosphate
- GLP-1R
Glucagon-like peptide 1 receptor
- GP
Glycoprotein
- ITAM
Immunoreceptor tyrosine-based activation motif
- ITP
Idiopathic thrombocytopenic purpura
- LLR
Leucine-rich repeat
- PAR
Protease-activated receptor
- PCI
Percutaneous coronary intervention
- PDI
Protein disulphide isomerase
- PSI
Plexin-semaphorin-integrin
- TIA
Transient ischemic attack
- TTP
Thrombotic thrombocytopenic purpura
- VWF
von Willebrand factor
References
- 1.Ruggeri ZM. Platelets in atherothrombosis. Nat Med. 2002;8:1227–34. doi: 10.1038/nm1102-1227. [DOI] [PubMed] [Google Scholar]
- 2.Mackman N. Triggers, targets and treatments for thrombosis. Nature. 2008;451:914–8. doi: 10.1038/nature06797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Xu XR, Gallant RC, Ni H. Platelets, immune-mediated thrombocytopenias, and fetal hemorrhage. Thromb Res. 2016;141(Suppl 2):S76–9. doi: 10.1016/S0049-3848(16)30372-3. [DOI] [PubMed] [Google Scholar]
- 4.Ni H, Freedman J. Platelets in hemostasis and thrombosis: role of integrins and their ligands. Transfus Apher Sci. 2003;28:257–64. doi: 10.1016/S1473-0502(03)00044-2. [DOI] [PubMed] [Google Scholar]
- 5.Yang H, Reheman A, Chen P, Zhu G, Hynes RO, Freedman J, et al. Fibrinogen and von Willebrand factor-independent platelet aggregation in vitro and in vivo. J Thromb Haemost. 2006;4:2230–7. doi: 10.1111/j.1538-7836.2006.02116.x. [DOI] [PubMed] [Google Scholar]
- 6.Wang Y, Gallant RC, Ni H. Extracellular matrix proteins in the regulation of thrombus formation. Curr Opin Hematol. 2016;23:280–7. doi: 10.1097/MOH.0000000000000237. [DOI] [PubMed] [Google Scholar]
- 7.Moroi M, Jung SM, Okuma M, Shinmyozu K. A patient with platelets deficient in glycoprotein VI that lack both collagen-induced aggregation and adhesion. J Clin Invest. 1989;84:1440–5. doi: 10.1172/JCI114318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dunne E, Spring CM, Reheman A, Jin W, Berndt MC, Newman DK, et al. Cadherin 6 has a functional role in platelet aggregation and thrombus formation. Arterioscler Thromb Vasc Biol. 2012;32:1724–31. doi: 10.1161/ATVBAHA.112.250464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Palabrica T, Lobb R, Furie BC, Aronovitz M, Benjamin C, Hsu YM, et al. Leukocyte accumulation promoting fibrin deposition is mediated in vivo by P-selectin on adherent platelets. Nature. 1992;359:848–51. doi: 10.1038/359848a0. [DOI] [PubMed] [Google Scholar]
- 10.Yang H, Lang S, Zhai Z, Li L, Kahr WH, Chen P, et al. Fibrinogen is required for maintenance of platelet intracellular and cell-surface P-selectin expression. Blood. 2009;114:425–36. doi: 10.1182/blood-2008-03-145821. [DOI] [PubMed] [Google Scholar]
- 11.Nurden AT. Platelet membrane glycoproteins: a historical review. Semin Thromb Hemost. 2014;40:577–84. doi: 10.1055/s-0034-1383826. [DOI] [PubMed] [Google Scholar]
- 12.Lopez JA, Andrews RK, Afshar-Kharghan V, Berndt MC. Bernard-Soulier syndrome. Blood. 1998;91:4397–418. [PubMed] [Google Scholar]
- 13.Jackson SP. Arterial thrombosis--insidious, unpredictable and deadly. Nat Med. 2011;17:1423–36. doi: 10.1038/nm.2515. [DOI] [PubMed] [Google Scholar]
- 14.Reheman A, Xu X, Reddy EC, Ni H. Targeting activated platelets and fibrinolysis: hitting two birds with one stone. Circ Res. 2014;114:1070–3. doi: 10.1161/CIRCRESAHA.114.303600. [DOI] [PubMed] [Google Scholar]
- 15.Writing Group Members. Mozaffarian D, Benjamin EJ, Go AS, Arnett DK, Blaha MJ, et al. Heart disease and stroke statistics-2016 update: a report from the American Heart Association. Circulation. 2016;133:e38–e60. doi: 10.1161/CIR.0000000000000350. [DOI] [PubMed] [Google Scholar]
- 16.Michelson AD. Antiplatelet therapies for the treatment of cardiovascular disease. Nat Rev Drug Discov. 2010;9:154–69. doi: 10.1038/nrd2957. [DOI] [PubMed] [Google Scholar]
- 17.Franchi F, Angiolillo DJ. Novel antiplatelet agents in acute coronary syndrome. Nat Rev Cardiol. 2015;12:30–47. doi: 10.1038/nrcardio.2014.156. [DOI] [PubMed] [Google Scholar]
- 18.Gachet C. Antiplatelet drugs: which targets for which treatments? J Thromb Haemost. 2015;13(Suppl 1):S313–22. doi: 10.1111/jth.12947. [DOI] [PubMed] [Google Scholar]
- 19.Hou Y, Carrim N, Wang Y, Gallant RC, Marshall A, Ni H. Platelets in hemostasis and thrombosis: novel mechanisms of fibrinogen-independent platelet aggregation and fibronectin-mediated protein wave of hemostasis. J Biomed Res. 2015;29:437–444. doi: 10.7555/JBR.29.20150121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wang H, Bang KW, Blanchette VS, Nurden AT, Rand ML. Phosphatidylserine exposure, microparticle formation and mitochondrial depolarisation in Glanzmann thrombasthenia platelets. Thromb Haemost. 2014;111:1184–6. doi: 10.1160/TH13-08-0704. [DOI] [PubMed] [Google Scholar]
- 21.Wang Y, Reheman A, Spring CM, Kalantari J, Marshall AH, Wolberg AS, et al. Plasma fibronectin supports hemostasis and regulates thrombosis. J Clin Invest. 2014;124:4281–93. doi: 10.1172/JCI74630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang Y, Ni H. Fibronectin maintains the balance between hemostasis and thrombosis. Cell Mol Life Sci. 2016;73(17):3265–77. doi: 10.1007/s00018-016-2225-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Xu XR, Zhang D, Oswald BE, Carrim N, Wang X, Hou Y, et al. Platelets are versatile cells: New discoveries in hemostasis, thrombosis, immune responses, tumor metastasis and beyond. Crit Rev Clin Lab Sci. 2016. Published online:1–69. doi: 10.1080/10408363.2016.1200008. [DOI] [PubMed]
- 24.Metharom P, Berndt MC, Baker RI, Andrews RK. Current state and novel approaches of antiplatelet therapy. Arterioscler Thromb Vasc Biol. 2015;35:1327–38. doi: 10.1161/ATVBAHA.114.303413. [DOI] [PubMed] [Google Scholar]
- 25.Michelson AD. Advances in antiplatelet therapy. Hematology Am Soc Hematol Educ Program. 2011;2011:62–9. doi: 10.1182/asheducation-2011.1.62. [DOI] [PubMed] [Google Scholar]
- 26.Jackson SP, Schoenwaelder SM. Antiplatelet therapy: in search of the ‘magic bullet’. Nat Rev Drug Discov. 2003;2:775–89. doi: 10.1038/nrd1198. [DOI] [PubMed] [Google Scholar]
- 27.Morrow DA, Braunwald E, Bonaca MP, Ameriso SF, Dalby AJ, Fish MP, et al. Vorapaxar in the secondary prevention of atherothrombotic events. N Engl J Med. 2012;366:1404–13. doi: 10.1056/NEJMoa1200933. [DOI] [PubMed] [Google Scholar]
- 28.Adminstration USFaD. Drug Trials Snapshot Zontivity (vorapaxar). 2014. http://www.fda.gov/Drugs/InformationOnDrugs/ucm423935.htm. Accessed 15 June 2016.
- 29.ClinicalTrials.gov. 2011. https://clinicaltrials.gov/ct2/show/NCT00527943?term=Vorapaxar&rank=5. Accessed 15 June 2016.
- 30.Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110:673–87. doi: 10.1016/S0092-8674(02)00971-6. [DOI] [PubMed] [Google Scholar]
- 31.Mou Y, Ni H, Wilkins JA. The selective inhibition of beta 1 and beta 7 integrin-mediated lymphocyte adhesion by bacitracin. J Immunol. 1998;161:6323–9. [PubMed] [Google Scholar]
- 32.Clemetson KJ, Clemetson JM. Platelet receptors. In: Michelson AD, editor. Platelets. 3. Amsterdam: Academic Press/Elsevier; 2013. pp. 169–94. [Google Scholar]
- 33.Varga-Szabo D, Pleines I, Nieswandt B. Cell adhesion mechanisms in platelets. Arterioscler Thromb Vasc Biol. 2008;28:403–12. doi: 10.1161/ATVBAHA.107.150474. [DOI] [PubMed] [Google Scholar]
- 34.Tamura S, Suzuki-Inoue K, Tsukiji N, Shirai T, Sasaki T, Osada M, et al. Podoplanin-positive periarteriolar stromal cells promote megakaryocyte growth and proplatelet formation in mice by CLEC-2. Blood. 2016;127:1701–10. doi: 10.1182/blood-2015-08-663708. [DOI] [PubMed] [Google Scholar]
- 35.Li C, Li J, Li Y, Lang S, Yougbare I, Zhu G, et al. Crosstalk between platelets and the immune system: old systems with new discoveries. Adv Hematol. 2012;2012:384685. doi: 10.1155/2012/384685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Semple JW, Italiano JE, Jr, Freedman J. Platelets and the immune continuum. Nat Rev Immunol. 2011;11:264–74. doi: 10.1038/nri2956. [DOI] [PubMed] [Google Scholar]
- 37.Wagner DD, Burger PC. Platelets in inflammation and thrombosis. Arterioscler Thromb Vasc Biol. 2003;23:2131–7. doi: 10.1161/01.ATV.0000095974.95122.EC. [DOI] [PubMed] [Google Scholar]
- 38.Siegel-Axel D, Daub K, Seizer P, Lindemann S, Gawaz M. Platelet lipoprotein interplay: trigger of foam cell formation and driver of atherosclerosis. Cardiovasc Res. 2008;78:8–17. doi: 10.1093/cvr/cvn015. [DOI] [PubMed] [Google Scholar]
- 39.Lindemann S, Kramer B, Seizer P, Gawaz M. Platelets, inflammation and atherosclerosis. J Thromb Haemost. 2007;5(Suppl 1):203–211. doi: 10.1111/j.1538-7836.2007.02517.x. [DOI] [PubMed] [Google Scholar]
- 40.Murphy AJ, Bijl N, Yvan-Charvet L, Welch CB, Bhagwat N, Reheman A, et al. Cholesterol efflux in megakaryocyte progenitors suppresses platelet production and thrombocytosis. Nat Med. 2013;19:586–94. doi: 10.1038/nm.3150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hess PR, Rawnsley DR, Jakus Z, Yang Y, Sweet DT, Fu J, et al. Platelets mediate lymphovenous hemostasis to maintain blood-lymphatic separation throughout life. J Clin Invest. 2014;124:273–284. doi: 10.1172/JCI70422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Navarro-Nunez L, Langan SA, Nash GB, Watson SP. The physiological and pathophysiological roles of platelet CLEC-2. Thromb Haemost. 2013;109:991–8. doi: 10.1160/TH13-01-0060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Osada M, Inoue O, Ding G, Shirai T, Ichise H, Hirayama K, et al. Platelet activation receptor CLEC-2 regulates blood/lymphatic vessel separation by inhibiting proliferation, migration, and tube formation of lymphatic endothelial cells. J Biol Chem. 2012;287:22241–52. doi: 10.1074/jbc.M111.329987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Herzog BH, Fu J, Wilson SJ, Hess PR, Sen A, McDaniel JM, et al. Podoplanin maintains high endothelial venule integrity by interacting with platelet CLEC-2. Nature. 2013;502:105–9. doi: 10.1038/nature12501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Italiano JE, Jr, Richardson JL, Patel-Hett S, Battinelli E, Zaslavsky A, Short S, et al. Angiogenesis is regulated by a novel mechanism: pro- and antiangiogenic proteins are organized into separate platelet alpha granules and differentially released. Blood. 2008;111:1227–33. doi: 10.1182/blood-2007-09-113837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chatterjee M, Huang Z, Zhang W, Jiang L, Hultenby K, Zhu L, et al. Distinct platelet packaging, release, and surface expression of proangiogenic and antiangiogenic factors on different platelet stimuli. Blood. 2011;117:3907–11. doi: 10.1182/blood-2010-12-327007. [DOI] [PubMed] [Google Scholar]
- 47.Yougbare I, Lang S, Yang H, Chen P, Zhao X, Tai WS, et al. Maternal anti-platelet beta3 integrins impair angiogenesis and cause intracranial hemorrhage. J Clin Invest. 2015;125:1545–56. doi: 10.1172/JCI77820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li C, Piran S, Chen P, Lang S, Zarpellon A, Jin JW, et al. The maternal immune response to fetal platelet GPIbalpha causes frequent miscarriage in mice that can be prevented by intravenous IgG and anti-FcRn therapies. J Clin Invest. 2011;121:4537–47. doi: 10.1172/JCI57850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yougbare I, Wei-She T, Zdravic D, Chen P, Zhu G, Leong-Poi H, et al. Natural killer cells contribute to pathophysiology of placenta leading to miscarriage in fetal and neonatal alloimmune thrombocytopenia. Blood. 2015;126:2254. doi: 10.1182/blood-2015-08-663492. [DOI] [PubMed] [Google Scholar]
- 50.Labelle M, Hynes RO. The initial hours of metastasis: the importance of cooperative host-tumor cell interactions during hematogenous dissemination. Cancer Discov. 2012;2:1091–9. doi: 10.1158/2159-8290.CD-12-0329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Labelle M, Begum S, Hynes RO. Platelets guide the formation of early metastatic niches. Proc Natl Acad Sci U S A. 2014;111:E3053–61. doi: 10.1073/pnas.1411082111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Franco AT, Corken A, Ware J. Platelets at the interface of thrombosis, inflammation, and cancer. Blood. 2015;126:582–8. doi: 10.1182/blood-2014-08-531582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Luo SZ, Mo X, Afshar-Kharghan V, Srinivasan S, Lopez JA, Li R. Glycoprotein Ibalpha forms disulfide bonds with 2 glycoprotein Ibbeta subunits in the resting platelet. Blood. 2007;109:603–9. doi: 10.1182/blood-2006-05-024091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Jackson SP. The growing complexity of platelet aggregation. Blood. 2007;109:5087–95. doi: 10.1182/blood-2006-12-027698. [DOI] [PubMed] [Google Scholar]
- 55.Ruggeri ZM, Mendolicchio GL. Adhesion mechanisms in platelet function. Circ Res. 2007;100:1673–85. doi: 10.1161/01.RES.0000267878.97021.ab. [DOI] [PubMed] [Google Scholar]
- 56.Huizinga EG, Tsuji S, Romijn RA, Schiphorst ME, de Groot PG, Sixma JJ, et al. Structures of glycoprotein Ibalpha and its complex with von Willebrand factor A1 domain. Science. 2002;297:1176–9. doi: 10.1126/science.107355. [DOI] [PubMed] [Google Scholar]
- 57.Verhenne S, Denorme F, Libbrecht S, Vandenbulcke A, Pareyn I, Deckmyn H, et al. Platelet-derived VWF is not essential for normal thrombosis and hemostasis but fosters ischemic stroke injury in mice. Blood. 2015;126:1715–22. doi: 10.1182/blood-2015-03-632901. [DOI] [PubMed] [Google Scholar]
- 58.Dumas JJ, Kumar R, Seehra J, Somers WS, Mosyak L. Crystal structure of the GpIbalpha-thrombin complex essential for platelet aggregation. Science. 2003;301:222–6. doi: 10.1126/science.1083917. [DOI] [PubMed] [Google Scholar]
- 59.Celikel R, McClintock RA, Roberts JR, Mendolicchio GL, Ware J, Varughese KI, et al. Modulation of alpha-thrombin function by distinct interactions with platelet glycoprotein Ibalpha. Science. 2003;301:218–21. doi: 10.1126/science.1084183. [DOI] [PubMed] [Google Scholar]
- 60.Andrews RK, Berndt MC. The GPIb-IX-V Complex. In: Michelson AD, editor. Platelets. 3. Amsterdam: Academic Press/Elsevier; 2013. pp. 195–213. [Google Scholar]
- 61.De Candia E, Hall SW, Rutella S, Landolfi R, Andrews RK, De Cristofaro R. Binding of thrombin to glycoprotein Ib accelerates the hydrolysis of Par-1 on intact platelets. J Biol Chem. 2001;276:4692–8. doi: 10.1074/jbc.M008160200. [DOI] [PubMed] [Google Scholar]
- 62.Ramakrishnan V, DeGuzman F, Bao M, Hall SW, Leung LL, Phillips DR. A thrombin receptor function for platelet glycoprotein Ib-IX unmasked by cleavage of glycoprotein V. Proc Natl Acad Sci U S A. 2001;98:1823–8. doi: 10.1073/pnas.98.4.1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ni H, Ramakrishnan V, Ruggeri ZM, Papalia JM, Phillips DR, Wagner DD. Increased thrombogenesis and embolus formation in mice lacking glycoprotein V. Blood. 2001;98:368–73. doi: 10.1182/blood.V98.2.368. [DOI] [PubMed] [Google Scholar]
- 64.Jurk K, Clemetson KJ, de Groot PG, Brodde MF, Steiner M, Savion N, et al. Thrombospondin-1 mediates platelet adhesion at high shear via glycoprotein Ib (GPIb): an alternative/backup mechanism to von Willebrand factor. FASEB J. 2003;17:1490–2. doi: 10.1096/fj.02-0830fje. [DOI] [PubMed] [Google Scholar]
- 65.Baglia FA, Badellino KO, Li CQ, Lopez JA, Walsh PN. Factor XI binding to the platelet glycoprotein Ib-IX-V complex promotes factor XI activation by thrombin. J Biol Chem. 2002;277:1662–8. doi: 10.1074/jbc.M108319200. [DOI] [PubMed] [Google Scholar]
- 66.Bradford HN, Pixley RA, Colman RW. Human factor XII binding to the glycoprotein Ib-IX-V complex inhibits thrombin-induced platelet aggregation. J Biol Chem. 2000;275:22756–63. [DOI] [PubMed]
- 67.Weeterings C, de Groot PG, Adelmeijer J, Lisman T. The glycoprotein Ib-IX-V complex contributes to tissue factor-independent thrombin generation by recombinant factor VIIa on the activated platelet surface. Blood. 2008;112:3227–33. doi: 10.1182/blood-2008-02-139113. [DOI] [PubMed] [Google Scholar]
- 68.Chavakis T, Santoso S, Clemetson KJ, Sachs UJ, Isordia-Salas I, Pixley RA, et al. High molecular weight kininogen regulates platelet-leukocyte interactions by bridging Mac-1 and glycoprotein Ib. J Biol Chem. 2003;278:45375–81. doi: 10.1074/jbc.M304344200. [DOI] [PubMed] [Google Scholar]
- 69.Romo GM, Dong JF, Schade AJ, Gardiner EE, Kansas GS, Li CQ, et al. The glycoprotein Ib-IX-V complex is a platelet counterreceptor for P-selectin. J Exp Med. 1999;190:803–14. doi: 10.1084/jem.190.6.803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kaplan ZS, Zarpellon A, Alwis I, Yuan Y, McFadyen J, Ghasemzadeh M, et al. Thrombin-dependent intravascular leukocyte trafficking regulated by fibrin and the platelet receptors GPIb and PAR4. Nat Commun. 2015;6:7835. doi: 10.1038/ncomms8835. [DOI] [PubMed] [Google Scholar]
- 71.Simon DI, Chen Z, Xu H, Li CQ, Dong J, McIntire LV, et al. Platelet glycoprotein ibalpha is a counterreceptor for the leukocyte integrin Mac-1 (CD11b/CD18) J Exp Med. 2000;192:193–204. doi: 10.1084/jem.192.2.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Chandraratne S, von Bruehl ML, Pagel JI, Stark K, Kleinert E, Konrad I, et al. Critical role of platelet glycoprotein ibalpha in arterial remodeling. Arterioscler Thromb Vasc Biol. 2015;35:589–97. doi: 10.1161/ATVBAHA.114.304447. [DOI] [PubMed] [Google Scholar]
- 73.Li J, van der Wal DE, Zhu G, Xu M, Yougbare I, Ma L, et al. Desialylation is a mechanism of Fc-independent platelet clearance and a therapeutic target in immune thrombocytopenia. Nat Commun. 2015;6:7737. doi: 10.1038/ncomms8737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li J, Callum JL, Lin Y, Zhou Y, Zhu G, Ni H. Severe platelet desialylation in a patient with glycoprotein Ib/IX antibody-mediated immune thrombocytopenia and fatal pulmonary hemorrhage. Haematologica. 2014;99:e61–3. doi: 10.3324/haematol.2013.102897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gresele P, Momi S. Inhibitors of the interaction between von Willebrand factor and platelet GPIb/IX/V. Handb Exp Pharmacol. 2012;(210):287–309. doi: 10.1007/978-3-642-29423-5_12. [DOI] [PubMed] [Google Scholar]
- 76.Ulrichts H, Silence K, Schoolmeester A, de Jaegere P, Rossenu S, Roodt J, et al. Antithrombotic drug candidate ALX-0081 shows superior preclinical efficacy and safety compared with currently marketed antiplatelet drugs. Blood. 2011;118:757–65. doi: 10.1182/blood-2010-11-317859. [DOI] [PubMed] [Google Scholar]
- 77.Bartunek J, Barbato E, Heyndrickx G, Vanderheyden M, Wijns W, Holz JB. Novel antiplatelet agents: ALX-0081, a Nanobody directed towards von Willebrand factor. J Cardiovasc Transl Res. 2013;6:355–63. doi: 10.1007/s12265-012-9435-y. [DOI] [PubMed] [Google Scholar]
- 78.Muller O, Bartunek J, Hamilos M, Berza CT, Mangiacapra F, Ntalianis A, et al. von Willebrand factor inhibition improves endothelial function in patients with stable angina. J Cardiovasc Transl Res. 2013;6:364–70. doi: 10.1007/s12265-012-9422-3. [DOI] [PubMed] [Google Scholar]
- 79.Peyvandi F, Scully M, Kremer Hovinga JA, Cataland S, Knobl P, Wu H, et al. Caplacizumab for acquired thrombotic thrombocytopenic purpura. N Engl J Med. 2016;374:511–22. doi: 10.1056/NEJMoa1505533. [DOI] [PubMed] [Google Scholar]
- 80.Lammle B. Thrombotic microangiopathy: caplacizumab accelerates resolution of acute acquired TTP. Nat Rev Nephrol. 2016;12:259–60. doi: 10.1038/nrneph.2016.47. [DOI] [PubMed] [Google Scholar]
- 81.Von VA. Willebrand factor--a new target for TTP treatment? N Engl J Med. 2016;374:583–5. doi: 10.1056/NEJMe1515876. [DOI] [PubMed] [Google Scholar]
- 82.Blombery P, Scully M. Management of thrombotic thrombocytopenic purpura: current perspectives. J Blood Med. 2014;5:15–23. doi: 10.2147/JBM.S46458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lei X, Reheman A, Hou Y, Zhou H, Wang Y, Marshall AH, et al. Anfibatide, a novel GPIb complex antagonist, inhibits platelet adhesion and thrombus formation in vitro and in vivo in murine models of thrombosis. Thromb Haemost. 2014;111:279–89. doi: 10.1160/TH13-06-0490. [DOI] [PubMed] [Google Scholar]
- 84.Hou Y, Li BX, Dai X, Yang Z, Qian F, Zhang G, et al. The first in vitro and in vivo assessment of anfibatide, a novel glycoprotein ib antagonist, in mice and in a phase i human clinical trial. Blood. 2013;122:577. [Google Scholar]
- 85.Li B, Dai X, Yang Z, Qian F, Zhang G, Xu Z, et al. First ex vivo and in vivo assessment of anfibatide, a novel glycoprotein Ib-IV-V complex antagonist, in healthy human volunteers in phase I clinical trial. J Thromb Haemost. 2013;11(Suppl. 2):23. [Google Scholar]
- 86.Nieswandt B, Kleinschnitz C, Stoll G. Ischaemic stroke: a thrombo-inflammatory disease? J Physiol. 2011;589:4115–23. doi: 10.1113/jphysiol.2011.212886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Stoll G, Kleinschnitz C, Nieswandt B. Molecular mechanisms of thrombus formation in ischemic stroke: novel insights and targets for treatment. Blood. 2008;112:3555–62. doi: 10.1182/blood-2008-04-144758. [DOI] [PubMed] [Google Scholar]
- 88.Kleinschnitz C, Pozgajova M, Pham M, Bendszus M, Nieswandt B, Stoll G. Targeting platelets in acute experimental stroke: impact of glycoprotein Ib, VI, and IIb/IIIa blockade on infarct size, functional outcome, and intracranial bleeding. Circulation. 2007;115:2323–30. doi: 10.1161/CIRCULATIONAHA.107.691279. [DOI] [PubMed] [Google Scholar]
- 89.Li TT, Fan ML, Hou SX, Li XY, Barry DM, Jin H, Luo SY, Kong F, Lau LF, Dai XR, Zhang GH, Zhou LL. A novel snake venomderived GPIb antagonist, anfibatide, protects mice from acute experimental ischaemic stroke and reperfusion injury. Br J Pharmacol. [DOI] [PMC free article] [PubMed]
- 90.Fontayne A, Meiring M, Lamprecht S, Roodt J, Demarsin E, Barbeaux P, et al. The humanized anti-glycoprotein Ib monoclonal antibody h6B4-Fab is a potent and safe antithrombotic in a high shear arterial thrombosis model in baboons. Thromb Haemost. 2008;100:670–7. [PubMed] [Google Scholar]
- 91.Hennan JK, Swillo RE, Morgan GA, Leik CE, Brooks JM, Shaw GD, et al. Pharmacologic inhibition of platelet vWF-GPIb alpha interaction prevents coronary artery thrombosis. Thromb Haemost. 2006;95:469–75. doi: 10.1160/TH05-09-0640. [DOI] [PubMed] [Google Scholar]
- 92.Ni H, Zhu G. Novel monoclonal antibodies against platelet GPIb-alpha: potential anti-thrombotic drugs and research reagents for study of thrombosis and hemostasis. 2012. [Google Scholar]
- 93.Moroi AJ, Watson SP. Impact of the PI3-kinase/Akt pathway on ITAM and hemITAM receptors: haemostasis, platelet activation and antithrombotic therapy. Biochem Pharmacol. 2015;94:186–194. doi: 10.1016/j.bcp.2015.02.004. [DOI] [PubMed] [Google Scholar]
- 94.Alshehri OM, Hughes CE, Montague S, Watson SK, Frampton J, Bender M, et al. Fibrin activates GPVI in human and mouse platelets. Blood. 2015;126:1601–8. doi: 10.1182/blood-2015-04-641654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Mammadova-Bach E, Ollivier V, Loyau S, Schaff M, Dumont B, Favier R, et al. Platelet glycoprotein VI binds to polymerized fibrin and promotes thrombin generation. Blood. 2015;126:683–91. doi: 10.1182/blood-2015-02-629717. [DOI] [PubMed] [Google Scholar]
- 96.Bigalke B, Stellos K, Geisler T, Kremmer E, Seizer P, May AE, et al. Expression of platelet glycoprotein VI is associated with transient ischemic attack and stroke. Eur J Neurol. 2010;17:111–7. doi: 10.1111/j.1468-1331.2009.02754.x. [DOI] [PubMed] [Google Scholar]
- 97.Induruwa I, Jung SM, Warburton EA. Beyond antiplatelets: the role of glycoprotein VI in ischemic stroke. Int J Stroke. 2016 doi: 10.1177/1747493016654532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Al-Tamimi M, Gardiner EE, Thom JY, Shen Y, Cooper MN, Hankey GJ, et al. Soluble glycoprotein VI is raised in the plasma of patients with acute ischemic stroke. Stroke. 2011;42:498–500. doi: 10.1161/STROKEAHA.110.602532. [DOI] [PubMed] [Google Scholar]
- 99.Stoll G, Kleinschnitz C, Nieswandt B. Combating innate inflammation: a new paradigm for acute treatment of stroke? Ann N Y Acad Sci. 2010;1207:149–54. doi: 10.1111/j.1749-6632.2010.05730.x. [DOI] [PubMed] [Google Scholar]
- 100.Thornton P, McColl BW, Greenhalgh A, Denes A, Allan SM, Rothwell NJ. Platelet interleukin-1alpha drives cerebrovascular inflammation. Blood. 2010;115:3632–9. doi: 10.1182/blood-2009-11-252643. [DOI] [PubMed] [Google Scholar]
- 101.Stegner D, Haining EJ, Nieswandt B. Targeting glycoprotein VI and the immunoreceptor tyrosine-based activation motif signaling pathway. Arterioscler Thromb Vasc Biol. 2014;34:1615–20. doi: 10.1161/ATVBAHA.114.303408. [DOI] [PubMed] [Google Scholar]
- 102.Dutting S, Bender M, Nieswandt B. Platelet GPVI: a target for antithrombotic therapy?! Trends Pharmacol Sci. 2012;33:583–90. doi: 10.1016/j.tips.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 103.Pachel C, Mathes D, Arias-Loza AP, Heitzmann W, Nordbeck P, Deppermann C, et al. Inhibition of platelet GPVI protects against myocardial ischemia-reperfusion injury. Arterioscler Thromb Vasc Biol. 2016;36:629–35. doi: 10.1161/ATVBAHA.115.305873. [DOI] [PubMed] [Google Scholar]
- 104.Goebel S, Li Z, Vogelmann J, Holthoff HP, Degen H, Hermann DM, et al. The GPVI-Fc fusion protein Revacept improves cerebral infarct volume and functional outcome in stroke. PLoS One. 2013;8:e66960. doi: 10.1371/journal.pone.0066960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Ungerer M, Rosport K, Bultmann A, Piechatzek R, Uhland K, Schlieper P, et al. Novel antiplatelet drug revacept (Dimeric Glycoprotein VI-Fc) specifically and efficiently inhibited collagen-induced platelet aggregation without affecting general hemostasis in humans. Circulation. 2011;123:1891–9. doi: 10.1161/CIRCULATIONAHA.110.980623. [DOI] [PubMed] [Google Scholar]
- 106.ClinicalTrials.gov. 2015. https://clinicaltrials.gov/ct2/show/NCT01645306?term=Revacept&rank=1. Accessed 15 June 2016.
- 107.Ono K, Ueda H, Yoshizawa Y, Akazawa D, Tanimura R, Shimada I, et al. Structural basis for platelet antiaggregation by angiotensin II type 1 receptor antagonist losartan (DuP-753) via glycoprotein VI. J Med Chem. 2010;53:2087–93. doi: 10.1021/jm901534d. [DOI] [PubMed] [Google Scholar]
- 108.Muzard J, Bouabdelli M, Zahid M, Ollivier V, Lacapere JJ, Jandrot-Perrus M, et al. Design and humanization of a murine scFv that blocks human platelet glycoprotein VI in vitro. FEBS J. 2009;276:4207–22. doi: 10.1111/j.1742-4658.2009.07129.x. [DOI] [PubMed] [Google Scholar]
- 109.Takagi J, Petre BM, Walz T, Springer TA. Global conformational rearrangements in integrin extracellular domains in outside-in and inside-out signaling. Cell. 2002;110:599–611. doi: 10.1016/S0092-8674(02)00935-2. [DOI] [PubMed] [Google Scholar]
- 110.Li R, Mitra N, Gratkowski H, Vilaire G, Litvinov R, Nagasami C, et al. Activation of integrin alphaIIbbeta3 by modulation of transmembrane helix associations. Science. 2003;300:795–8. doi: 10.1126/science.1079441. [DOI] [PubMed] [Google Scholar]
- 111.Vinogradova O, Vaynberg J, Kong X, Haas TA, Plow EF, Qin J. Membrane-mediated structural transitions at the cytoplasmic face during integrin activation. Proc Natl Acad Sci U S A. 2004;101:4094–9. doi: 10.1073/pnas.0400742101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Vinogradova O, Velyvis A, Velyviene A, Hu B, Haas T, Plow E, et al. A structural mechanism of integrin alpha(IIb)beta(3) “inside-out” activation as regulated by its cytoplasmic face. Cell. 2002;110:587–97. doi: 10.1016/S0092-8674(02)00906-6. [DOI] [PubMed] [Google Scholar]
- 113.Ni H, Denis CV, Subbarao S, Degen JL, Sato TN, Hynes RO, et al. Persistence of platelet thrombus formation in arterioles of mice lacking both von Willebrand factor and fibrinogen. J Clin Invest. 2000;106:385–92. doi: 10.1172/JCI9896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Law DA, DeGuzman FR, Heiser P, Ministri-Madrid K, Killeen N, Phillips DR. Integrin cytoplasmic tyrosine motif is required for outside-in alphaIIbbeta3 signalling and platelet function. Nature. 1999;401:808–11. doi: 10.1038/44599. [DOI] [PubMed] [Google Scholar]
- 115.Hodivala-Dilke KM, McHugh KP, Tsakiris DA, Rayburn H, Crowley D, Ullman-Cullere M, et al. Beta3-integrin-deficient mice are a model for Glanzmann thrombasthenia showing placental defects and reduced survival. J Clin Invest. 1999;103:229–38. doi: 10.1172/JCI5487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Reheman A, Yang H, Zhu G, Jin W, He F, Spring CM, et al. Plasma fibronectin depletion enhances platelet aggregation and thrombus formation in mice lacking fibrinogen and von Willebrand factor. Blood. 2009;113:1809–17. doi: 10.1182/blood-2008-04-148361. [DOI] [PubMed] [Google Scholar]
- 117.Reheman A, Gross P, Yang H, Chen P, Allen D, Leytin V, et al. Vitronectin stabilizes thrombi and vessel occlusion but plays a dual role in platelet aggregation. J Thromb Haemost. 2005;3:875–83. doi: 10.1111/j.1538-7836.2005.01217.x. [DOI] [PubMed] [Google Scholar]
- 118.Topol EJ, Byzova TV, Plow EF. Platelet GPIIb-IIIa blockers. Lancet. 1999;353:227–31. doi: 10.1016/S0140-6736(98)11086-3. [DOI] [PubMed] [Google Scholar]
- 119.Phillips DR, Scarborough RM. Clinical pharmacology of eptifibatide. Am J Cardiol. 1997;80:11B–20B. doi: 10.1016/S0002-9149(97)00572-9. [DOI] [PubMed] [Google Scholar]
- 120.Egbertson MS, Chang CT, Duggan ME, Gould RJ, Halczenko W, Hartman GD, et al. Non-peptide fibrinogen receptor antagonists. 2. Optimization of a tyrosine template as a mimic for Arg-Gly-Asp. J Med Chem. 1994;37:2537–51. doi: 10.1021/jm00042a007. [DOI] [PubMed] [Google Scholar]
- 121.Schneider DJ. Anti-platelet therapy: glycoprotein IIb-IIIa antagonists. Br J Clin Pharmacol. 2011;72:672–82. doi: 10.1111/j.1365-2125.2010.03879.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Coller BS. alphaIIbbeta3: structure and function. J Thromb Haemost. 2015;13(Suppl 1):S17–25. doi: 10.1111/jth.12915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Ciccone A, Motto C, Abraha I, Cozzolino F, Santilli I. Glycoprotein IIb-IIIa inhibitors for acute ischaemic stroke. Cochrane Database Syst Rev. 2014;3:CD005208. doi: 10.1002/14651858.CD005208.pub3. [DOI] [PubMed] [Google Scholar]
- 124.Li J, Vootukuri S, Shang Y, Negri A, Jiang JK, Nedelman M, et al. RUC-4: a novel alphaIIbbeta3 antagonist for prehospital therapy of myocardial infarction. Arterioscler Thromb Vasc Biol. 2014;34:2321–9. doi: 10.1161/ATVBAHA.114.303724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Xiong JP, Stehle T, Goodman SL, Arnaout MA. A novel adaptation of the integrin PSI domain revealed from its crystal structure. J Biol Chem. 2004;279:40252–4. doi: 10.1074/jbc.C400362200. [DOI] [PubMed] [Google Scholar]
- 126.Zang Q, Springer TA. Amino acid residues in the PSI domain and cysteine-rich repeats of the integrin beta2 subunit that restrain activation of the integrin alpha(X)beta(2) J Biol Chem. 2001;276:6922–9. doi: 10.1074/jbc.M005868200. [DOI] [PubMed] [Google Scholar]
- 127.Ni H, Li A, Simonsen N, Wilkins JA. Integrin activation by dithiothreitol or Mn2+ induces a ligand-occupied conformation and exposure of a novel NH2-terminal regulatory site on the beta1 integrin chain. J Biol Chem. 1998;273:7981–7. doi: 10.1074/jbc.273.14.7981. [DOI] [PubMed] [Google Scholar]
- 128.Yan B, Smith JW. A redox site involved in integrin activation. J Biol Chem. 2000;275:39964–72. doi: 10.1074/jbc.M007041200. [DOI] [PubMed] [Google Scholar]
- 129.Essex DW, Li M. Redox control of platelet aggregation. Biochemistry. 2003;42:129–36. doi: 10.1021/bi0205045. [DOI] [PubMed] [Google Scholar]
- 130.Manickam N, Ahmad SS, Essex DW. Vicinal thiols are required for activation of the alphaIIbbeta3 platelet integrin. J Thromb Haemost. 2011;9:1207–15. doi: 10.1111/j.1538-7836.2011.04266.x. [DOI] [PubMed] [Google Scholar]
- 131.Wang L, Wu Y, Zhou J, Ahmad SS, Mutus B, Garbi N, et al. Platelet-derived ERp57 mediates platelet incorporation into a growing thrombus by regulation of the alphaIIbbeta3 integrin. Blood. 2013;122:3642–50. doi: 10.1182/blood-2013-06-506691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Carrim N, Zhu G, Reddy E, Xu M, Xu X, Wang Y, et al. Integrin PSI domain has endogenous thiol isomerase function and is a novel target for anti-thrombotic therapy. J Thromb Haemost. 2015;13(Supplement S2):60. [Google Scholar]
- 133.Wang X, Palasubramaniam J, Gkanatsas Y, Hohmann JD, Westein E, Kanojia R, et al. Towards effective and safe thrombolysis and thromboprophylaxis: preclinical testing of a novel antibody-targeted recombinant plasminogen activator directed against activated platelets. Circ Res. 2014;114:1083–93. doi: 10.1161/CIRCRESAHA.114.302514. [DOI] [PubMed] [Google Scholar]
- 134.Fuentes RE, Zaitsev S, Ahn HS, Hayes V, Kowalska M, Lambert MP, et al. A chimeric platelet-targeted urokinase prodrug selectively blocks new thrombus formation. J Clin Invest. 2016;126:483–94. doi: 10.1172/JCI81470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Schaff M, Tang C, Maurer E, Bourdon C, Receveur N, Eckly A, et al. Integrin alpha6beta1 is the main receptor for vascular laminins and plays a role in platelet adhesion, activation, and arterial thrombosis. Circulation. 2013;128:541–52. doi: 10.1161/CIRCULATIONAHA.112.000799. [DOI] [PubMed] [Google Scholar]
- 136.Miller MW, Basra S, Kulp DW, Billings PC, Choi S, Beavers MP, et al. Small-molecule inhibitors of integrin alpha2beta1 that prevent pathological thrombus formation via an allosteric mechanism. Proc Natl Acad Sci U S A. 2009;106:719–24. doi: 10.1073/pnas.0811622106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Marcinkiewicz C, Lobb RR, Marcinkiewicz MM, Daniel JL, Smith JB, Dangelmaier C, et al. Isolation and characterization of EMS16, a C-lectin type protein from Echis multisquamatus venom, a potent and selective inhibitor of the alpha2beta1 integrin. Biochemistry. 2000;39:9859–67. doi: 10.1021/bi000428a. [DOI] [PubMed] [Google Scholar]
- 138.Arlinghaus FT, Momic T, Ammar NA, Shai E, Spectre G, Varon D, et al. Identification of alpha2beta1 integrin inhibitor VP-i with anti-platelet properties in the venom of Vipera palaestinae. Toxicon. 2013;64:96–105. doi: 10.1016/j.toxicon.2013.01.001. [DOI] [PubMed] [Google Scholar]
- 139.Piotrowicz RS, Orchekowski RP, Nugent DJ, Yamada KY, Kunicki TJ. Glycoprotein Ic-IIa functions as an activation-independent fibronectin receptor on human platelets. J Cell Biol. 1988;106:1359–64. doi: 10.1083/jcb.106.4.1359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Przyklenk K, Frelinger AL, 3rd, Linden MD, Whittaker P, Li Y, Barnard MR, et al. Targeted inhibition of the serotonin 5HT2A receptor improves coronary patency in an in vivo model of recurrent thrombosis. J Thromb Haemost. 2010;8:331–40. doi: 10.1111/j.1538-7836.2009.03693.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Ni H. The platelet “sugar high” in diabetes. Blood. 2012;119:5949–51. doi: 10.1182/blood-2012-04-420794. [DOI] [PubMed] [Google Scholar]
- 142.Cameron-Vendrig A, Reheman A, Siraj MA, Xu XR, Wang Y, Lei X, et al. Glucagon-like peptide 1 receptor activation attenuates platelet aggregation and thrombosis. Diabetes. 2016;65:1714–23. doi: 10.2337/db15-1141. [DOI] [PubMed] [Google Scholar]
- 143.Monami M, Dicembrini I, Nardini C, Fiordelli I, Mannucci E. Effects of glucagon-like peptide-1 receptor agonists on cardiovascular risk: a meta-analysis of randomized clinical trials. Diabetes Obes Metab. 2014;16:38–47. doi: 10.1111/dom.12175. [DOI] [PubMed] [Google Scholar]
- 144.Martinod K, Wagner DD. Thrombosis: tangled up in NETs. Blood. 2014;123:2768–76. doi: 10.1182/blood-2013-10-463646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Kumar A, Villani MP, Patel UK, Keith JC, Jr, Schaub RG. Recombinant soluble form of PSGL-1 accelerates thrombolysis and prevents reocclusion in a porcine model. Circulation. 1999;99:1363–9. doi: 10.1161/01.CIR.99.10.1363. [DOI] [PubMed] [Google Scholar]
- 146.Bedard PW, Clerin V, Sushkova N, Tchernychev B, Antrilli T, Resmini C, et al. Characterization of the novel P-selectin inhibitor PSI-697 [2-(4-chlorobenzyl)-3-hydroxy-7,8,9,10-tetrahydrobenzo[h] quinoline-4-carboxylic acid] in vitro and in rodent models of vascular inflammation and thrombosis. J Pharmacol Exp Ther. 2008;324:497–506. doi: 10.1124/jpet.107.128124. [DOI] [PubMed] [Google Scholar]
- 147.Meier TR, Myers DD, Jr, Wrobleski SK, Zajkowski PJ, Hawley AE, Bedard PW, et al. Prophylactic P-selectin inhibition with PSI-421 promotes resolution of venous thrombosis without anticoagulation. Thromb Haemost. 2008;99:343–51. doi: 10.1160/TH07-10-0608. [DOI] [PubMed] [Google Scholar]
- 148.Kolandaivelu K, Bhatt DL. Novel antiplatelet therapies. In: Michelson AD, editor. Platelets. 3. Amsterdam: Academic Press/Elsevier; 2013. pp. 1185–1213. [Google Scholar]
- 149.Conde ID, Kleiman NS. Soluble CD40 ligand in acute coronary syndromes. N Engl J Med. 2003;348:2575–7. doi: 10.1056/NEJM200306193482516. [DOI] [PubMed] [Google Scholar]
- 150.Schonbeck U, Libby P. CD40 signaling and plaque instability. Circ Res. 2001;89:1092–1103. doi: 10.1161/hh2401.101272. [DOI] [PubMed] [Google Scholar]
- 151.Lin J, Kakkar V, Lu X. Essential roles of toll-like receptors in atherosclerosis. Curr Med Chem. 2016;23:431–54. doi: 10.2174/0929867323666151207111408. [DOI] [PubMed] [Google Scholar]
- 152.Hovland A, Jonasson L, Garred P, Yndestad A, Aukrust P, Lappegard KT, et al. The complement system and toll-like receptors as integrated players in the pathophysiology of atherosclerosis. Atherosclerosis. 2015;241:480–94. doi: 10.1016/j.atherosclerosis.2015.05.038. [DOI] [PubMed] [Google Scholar]
- 153.Beckman MG, Hooper WC, Critchley SE, Ortel TL. Venous thromboembolism: a public health concern. Am J Prev Med. 2010;38:S495–501. doi: 10.1016/j.amepre.2009.12.017. [DOI] [PubMed] [Google Scholar]
- 154.Husain M, Aameron-Vendrig A, Ni H. Methods for inhibiting platelet aggregation using glp-1 receptor agonists. Google Patents; 2014. WO2014066992.