

Abstract
Background
The olfactomedin-like domain (OLFML) is present in at least four families of proteins, including OLFML2A and OLFML2B, which are expressed in adult rat retina cells. However, no expression of their orthologous has ever been reported in human and baboon.
Objective
The aim of this study was to investigate the expression of OLFML2A and OLFML2B in ocular tissues of baboons (Papio hamadryas) and humans, as a key to elucidate OLFML function in eye physiology.
Methods
OLFML2A and OLFML2B cDNA detection in ocular tissues of these species was performed by RT-PCR. The amplicons were cloned and sequenced, phylogenetically analyzed and their proteins products were confirmed by immunofluorescence assays.
Results
OLFML2A and OLFML2B transcripts were found in human cornea, lens and retina and in baboon cornea, lens, iris and retina. The baboon OLFML2A and OLFML2B ORF sequences have 96% similarity with their human’s orthologous. OLFML2A and OLFML2B evolution fits the hypothesis of purifying selection. Phylogenetic analysis shows clear orthology in OLFML2A genes, while OLFML2B orthology is not clear.
Conclusions
Expression of OLFML2A and OLFML2B in human and baboon ocular tissues, including their high similarity, make the baboon a powerful model to deduce the physiological and/or metabolic function of these proteins in the eye.
Keywords: Olfactomedin, Eye, Old world monkey, Baboon, OLFML2
Background
The olfactomedin (OLFM) family is a group of glycoproteins originally identified in bullfrogs (Rana catesbeiana) as the major component of the olfactory mucus layer, which surrounds the chemosensory dendrites of olfactory neurons [1]. Subsequently these proteins were found in the brain of species ranging from Caenorhabditis elegans to Homo sapiens [2]. OLFM shares a domain in the C-terminal of ~250 amino acids known as OLF [3]. Accordingly, in this domain, the OLF family was classified into seven subfamilies by phylogenetic analysis (as roman numerals from I to VII) [4]. Biological functions of proteins, which posses the OLF domain, remain for the most part elusive. A growing body of evidence indicates that these proteins may play very important roles in normal development and pathology [5]. For example, mutations in the OLF domain of myocilin were closely associated with primary open angle glaucoma [6]. Noelin-1 (OLFM-1) was found to play an important role in vertebrate neural crest development [7] and it is involved in frog neurogenesis (Xenopus laevis) [8]. Olfactomedin-likes (OLFML) are other members of the OLF family. Some of their members are the glycoproteins OLFML2A and OLFML2B, also known as photomedin-1 and photomedin-2, respectively, which were described in mice at 2005 [3]. OLFML2A and OLFML2B proteins are members of the subfamily IV [2]. The human OLFML2A and OLFML2B genes are located on chromosomes 9q33.3 and 1q23.3, respectively. These two genes are composed of at least 8 exons, and have a length of 37.3 and 40.7 kb, respectively [9].
In a mouse, OLFML2A and OLFML2B cDNAs have open reading frames (ORF) encoding 681 and 746 amino acid (aa) residues, respectively. Both proteins have a signal sequence at their N-terminal followed by two tandem CXCXCX9C motifs, a putative coiled-coil region, a serine/threonine-rich region, an OLF domain in their C-terminal, and two or three potential N-glycosylation sites. Both genes are expressed in the adult retina of mouse where they show mutually exclusive expression patterns. OLFML2A was predominantly detected in the photoreceptor layer, while OLFML2B is present in ganglion cells and inner nuclear layers, the inner segment of photoreceptor layer, and retinal-pigmented-epithelium [3]. Currently, the function of OLFML2 (A and B) proteins are still not clear and it is unknown if they are expressed in the retina of other mammals. Based on the above, the aims of this study were: (1) clone and sequence OLFML2A and OLFML2B cDNAs from the retina of baboons (Papio hamadryas) and humans; and (2) identify the cell layers where these proteins are expressed as a key to elucidate OLFML function in eye physiology.
Methods
Baboon’s biological specimens
Animal protocols were designed and developed according to the ethical guidelines of the Institutional Animal Care and Use Committee of the Texas Institute of Biomedical Research (TIBR). The baboon (Papio hamadryas) colony is preserved at the Southwest National Primate Research Center in San Antonio, Texas, USA. All the animals have the same diet and share similar environmental conditions. Baboons were gang-housed and fed ad libitum, a standard low-fat chow diet (Harlan Tecklad 15% monkey diet, 8715). The complete eyes of three adult female baboons (15, 16 and 18 years old) were collected. One eye was frozen in liquid nitrogen for RNA extraction and the other was included in formaldehyde at 4% for the immunofluorescence assays.
Human’s biological specimens
Biopsies from human eyes were collected at the Department of Ophthalmology of the “Dr. José Eleuterio González” University Hospital of the Universidad Autónoma de Nuevo Leon, in Monterrey, Mexico. Specimens came from programmed eye surgery procedures where ocular tissues were removed by medical indication. All patients signed an informed consent according to the ethics committee guidelines of the institution. Biological samples were immediately immersed in RNA later solution (Ambion Inc., Austin, TX, USA) for RNA extraction and stored at −70 °C until their use. Paraffin-embedded biopsies, were used in immunofluorescence assays, these biopsies were provided by Pathology Department of the Instituto de Oftalmología Fundación de Asistencia Privada Conde de Valenciana IAP, at Mexico City. The characteristics of human eyes specimens are shown in Table 1.
Table 1.
Characteristics of human eye tissues
| Age (years) | Diagnostic/procedure | Eye tissues collected | Assay | Source |
|---|---|---|---|---|
| 47 | Enucleated eye for a traumatic penetrating injury | Retina, sclera and uvea | RT-PCR | 1 |
| 82 | Penetrating keratoplasty and cataract extraction for a cornea stroma scar and dislocated cataract | Lens, vitreous and corneal endothelium | RT-PCR | 1 |
| 71 | Enucleated eye for an orbit carcinoma, infective keratitis and neovascular glaucoma | Optic nerve | RT-PCR | 1 |
| 7 | Strabismus surgery (medical rectus recession) | Conjunctiva | RT-PCR | 1 |
| 13 | Strabismus surgery (inferior oblique muscle myectomy) | Inferior oblique muscle, conjunctiva and tendon | RT-PCR | 1 |
| 2 | Retinoblastoma | Conjunctiva, retina, choroid and sclera | IHF | 2 |
| 35 | Enucleated eye for a traumatic penetrating injury | Retina, uvea and sclera | IHF | 2 |
| 63 | Chorodial melanoma | Retina, uvea and sclera | IHF | 2 |
1 Department of Ophthalmology of the Hospital Universitario “Dr. Jose Eleuterio González of the UANL, 2 Department of Pathology of the Instituto de Oftalmología. Fundación de Asistencia Privada Conde de Valenciana IAP
Reverse transcription and polymerase chain reaction
Each ocular piece from three adult baboons was dissected. The retina, cornea, lens, sclera, iris, choroid and optic nerve were separated. Total RNA was extracted from the eye tissues samples with Trizol reagent according to the manufacturer’s instructions (ThermoFisher Scientific Walthman, MA, USA). RNA was treated with RQ1 DNAse (Promega, Madison, WI, USA) for 15 min at 37 °C to remove traces of genomic DNA. For the assessment of RNA purity and integrity, we used standard methods of spectrophotometery and gel electrophoresis, respectively. Complementary DNA (cDNA) was synthesized using total RNA (1 µg), High Capacity cDNA Reverse Transcription kit (ThermoFisher Scientific) and oligo (dT) 12–18 primer (ThermoFisher Scientific) in 60 µL of total volume reaction. A primer set to amplify baboon OLFML2 A and B transcripts was designed using human sequences as templates; accesion numbers NM_182487 and AK316154, respectively. Such design was performed using an online tool [10], for OLFML2A the sense primer: 5′-CAGGCAGAGCGGGCGAAG-3′ and the anti sense primer: 5′-AATATTTGCGGACTGGGTCA-3′; while for OLFML2B, sense primer: 5′-AAGGGGCTGAGGACACTCTT-3′ and anti sense primer: 5′-GGAGGATGAGACCAGCACAT-3′. PCR was carried out using 100 ng of cDNA, 0.4 µM of each primer and GoTaq PCR master mix kit (Promega, Valencia, CA, USA). The amplification reaction was carried out in a thermal cycler Veriti 96-Well Thermal Cycler (ThermoFisher Scientific). The amplification used program was as follows: an initial denaturation step of 4 min at 94 °C, 40 cycles of 30 s each at 94 °C, 30 s at 60 °C, 90 s at 72 °C, and finally an elongation step of 6 min at 72 °C. The PCR products were visualized on 0.8% agarose gels stained with ethidium bromide and visualized under UV light.
Molecular cloning and sequence analysis
The amplified products were cloned in the 3.5-kb XL-TOPO vector and transformed into electrocompetent Escherichia coli bacteria strain Top 10 according to the manufacturer’s specifications (Invitrogen, Carlsbad, CA, USA). Positive clones were sequenced using Big Dye Terminator Cycle Sequencing Kit v3.1 using specific oligos and/or M13 universal primers. The reactions were analyzed in the ABI PRISM 3100 Genetic Analyzer using the Sequencing Analysis Software v5.3 (Applied Biosystems, Foster city, CA, USA). The information obtained from the sequencing assays was subjected to a BLAST test to determine identity.
Phylogenetic analysis
The sequences obtained from clones were aligned with the human orthologous reported gene (GenBank: NM_182487 and AK316154) using the CLUSTAL W program [11] followed by manual corrections in case of need. Protein sequences were derived by conceptual translation of the coding sequences. From amino acidic sequences, a phylogenetic tree was built with MEGA 6.06 software [12] using the maximum likelihood (ML), neighbor-joining (NJ) and UPGMA methods; then a bootstrap test was done with 1000 replicates [13]. Sequences used in this study are listed in Table 2.
Table 2.
Primate OLFML2A and OLFML2B sequences from NCBI GenBank used in this study
Seeking to identify the evolutionary forces that underlie the process of divergence in the OLFML2A and OLFML2B primate genes, we tested the hypothesis of positive or adaptive evolution (dN > dS), purifying selection (dN < dS), and neutrality (dN = dS). For this purpose, first, we calculated the non-synonymous (causes an amino acid change) dN and synonymous (does not cause an amino acid change) dS distances respectively, by the the Li-Wu-Luo method (Kimura 2-parameters) [14] from OLFML2′ coding sequences from apes, OWM and NWM with their lemur counterpart. Second, we tested whether dN is significantly greater, lower or equal, respectively, than dS using a codon-based Z test of selection as implemented in MEGA 6.06 software [12]. Differences were considered statistically significant at a P < 0.05.
Immunofluorescence assays
Ocular tissues were fixed in 4% paraformaldehyde (Sigma, St. Louis, MO, USA). These tissues were then dehydrated in a graded series of ethanol (50%, 30 min; 70%, 30 min; 95%, 30 min; 100%, 30 min) and cleared with xylene (Merck, Whitehouse Station, NJ, USA) for 30 min; tissues were then infiltrated with paraffin wax at 56 °C for 3 h. Serial transverse tissue sections (5 microns) were taken using a Leica RM-2135 microtome (Leica, Solms, Germany) and mounted into charged slides with 5% Gelatin (Bio-Rad, Hercules, CA, USA). The sections were cleared with citrosolv for 15 min and rehydrated with increasing serial ethanol concentrations (100, 100, 96, 96, 70, 70, and 50%) for five minutes each concentration and equilibrated in distillated water for 2 min. Exposure of the epitopes was performed by incubating the preparations with citrate buffer (10 mM sodium citrate, 0.05% Triton, pH 6). The sections were then blocked with 5% non-fat dry milk (Bio-Rad, Hercules, CA, USA); and treated with the reducing agent sodium borohydride in 0.01% in TBS 1X. The primary antibody incubation was overnight using a rabbit polyclonal anti-human OLFML2A antibody (Abcam ab75882, at a dilution 1:500), rabbit polyclonal anti-human OLFML2B antibody (Abcam ab75355, at a dilution 1:500). Double labeling was performed with both anti-OLFML2 antibodies and antibodies for neurons mouse monoclonal anti-human neuronal β-tubulin 3 beta chain (Covance MMS-453P, at a dilution 1:250) and astrocytes mouse monoclonal anti-human GFAP (anti-glial fibrillary acidic protein, Millipore MAB360, at a dilution 1:300). The antibodies were diluted in TTBS buffer with 1% skim milk. The primary antibody was omitted, for further negative controls. After primary antibody incubation, slides were washed with TTBS 1X buffer at room temperature for 10 min and repeating the process five times. Then the sections were incubated for 2 h in darkness at room temperature with secondary antibodies Cy3® goat anti-rabbit IgG (Life technologies, A10520, at a dilution 1:4000), Cy5® goat anti-rabbit IgG (Life technologies, A10523, at a dilution 1:4000), goat anti-mouse IgG FITC (Life technologies, 62-6511, at a dilution 1:250). The sections were washed five times with TTBS 1X for 5 min each time, subsequently placed with DAPI (Sigma, St. Louis, MO, USA, D-9542, at a final concentration of 0.1 µg/ml) for 20 min. The labeled sections were then washed three times with TTBS 1X for 5 min each time and mounted with Vectashield (Vector Laboratories, H-1000). Also, the labeled sections were examined using a LSM 780 confocal microscope (Carl Zeiss, Göttin-gen, Germany) and lasers at excitation wavelengths of 550, 650 and 495 nm, respectively. Digital images were brightness and contrast balanced.
Results
RT-PCR, molecular cloning and sequence analysis
The OLFML2A and OLFML2B coding sequences were as expected (1947 and 2262 bp, respectively) and no other bands were detected as possible isoforms. In human samples, we detected amplification in retina, cornea and lens. While in baboon, amplification was detected in retina, cornea, lens and iris (Table 3; Fig. 1). The novel baboon OLFML2A and OLFML2B mRNA sequences were deposited in the GenBank database under the accession numbers KU587785 and KU587786, respectively. Such sequences have full CDS, which codifies for predicted proteins of 648 and 753 amino acids in length, respectively. The baboon OLFML2A and OLFML2B CDSs nucleotide sequences have 96% similarity with the human orthologous. In the amino acidic sequence, the similarity between baboon and humans is 98%.
Table 3.
Tissue distribution of OLFML2A and OLFML2B mRNAs and proteins in baboon eye
| Primate | mRNA | Tissue | ||||
|---|---|---|---|---|---|---|
| Retina | Cornea | Lens | Sclera | Iris | ||
| Baboon | OLFML2A | √ | √ | √ | X | √ |
| OLFML2B | √ | √ | √ | X | √ | |
| Human | OLFML2A | √ | √ | √ | X | X |
| OLFML2B | √ | √ | √ | X | X | |
| Condition | Protein | Cell type or layer (of retina) | |||||
|---|---|---|---|---|---|---|---|
| Ganglion cell layer (GCL) | Inner plexiform layer (IPL) | Inner nuclear layer (INL) | Outer nuclear layer (ONL) | Rods layer (RL) | Pigmented epithelium (PE) | ||
| Baboon | OLFML2A | √ | √ | √ | √ | √ | √ |
| OLFML2B | √ | √ | √ | √ | √ | X | |
| Human | OLFML2A | √ | X | √ | √ | X | X |
| OLFML2B | √ | X | √ | √ | X | X | |
Fig. 1.
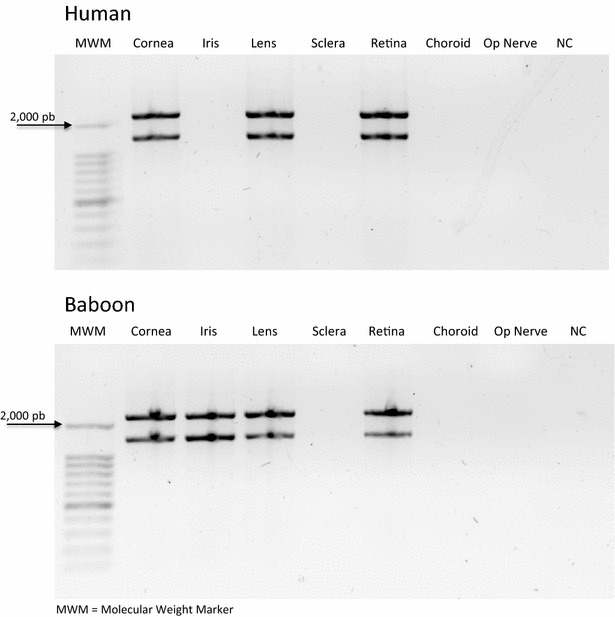
0.8% agarose gel. RT-PCR products were visualized and no other bans bans were detected as possible transcript variants. OLFML2A and OLFML2B were as expected 1947 and 2262 respectively
Two phylogenetic trees were built, one for each protein. The OLFML2A tree (Fig. 2) shows four clades in a lineage-specific manner. These clades match in a lineage-specfic manner, they correspond to apes, OWM, NWM, and lemur (out-group). It confirms the orthology between primate OLFML2A genes. While the OLFML2B tree (Fig. 3) shows the same clades, but they are not in a lineage-specific manner. NWM act as out-grop, thus orthology is not clear. Bootstrap values are shown on the tree’s branches. Similar results were obtained using maximum likelihood (ML), neighbor-joining (NJ) and UPGMA phylogenetic methods. We confirmed that OLFML2A and OLFML2B evolution fit the hypothesis of purifying selection (dN > dS, P < 0.05). See Table 4.
Fig. 2.
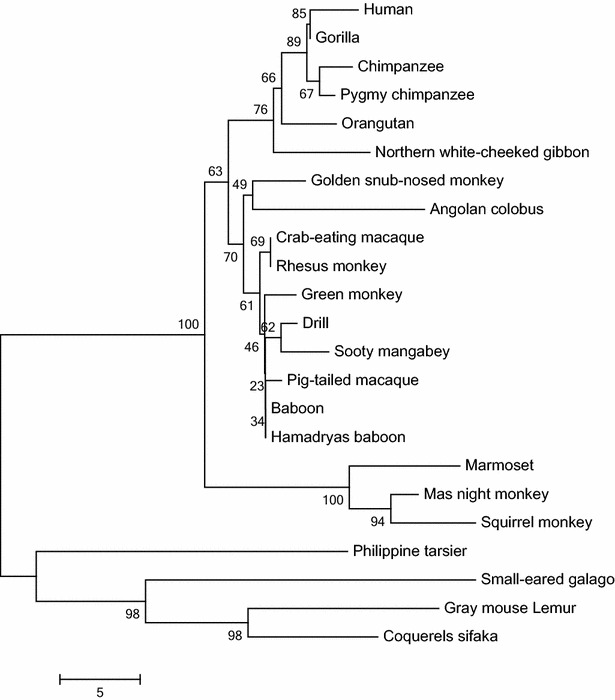
Phylogenetic tree of OLFML2A proteins from various primates. The tree was built using MEGA version 6.06 by the ML, NJ and UPGMA methods and further bootstrap analysis of 1000 replicas. The numbers in the branches indicate Bootstrap value and below the branches length. Clades are in linage specific manner, apes, OWM, NWM and lemur (out-group)
Fig. 3.
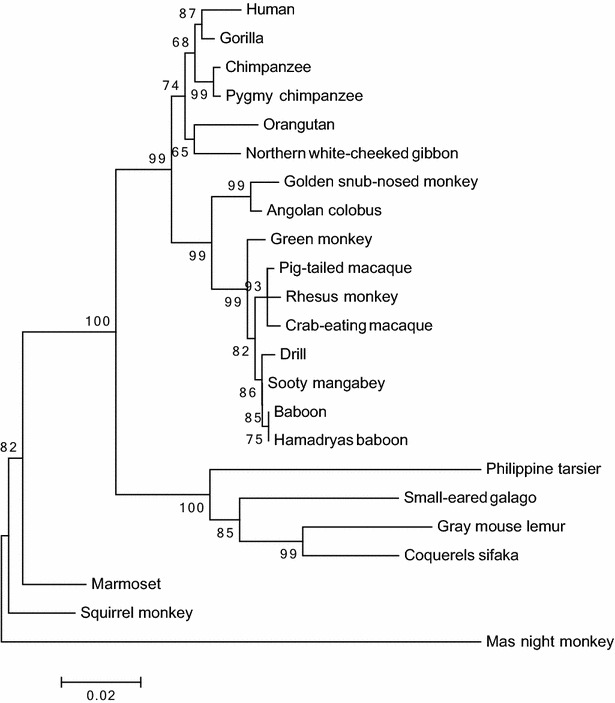
Phylogenetic tree of OLFML2B proteins from various primates. The tree was built using MEGA version 6.06 by the ML, NJ and UPGMA methods and further bootstrap analysis of 1000 replicas. Numbers in the branches indicate Bootstrap value and below the branches length. Clades are not in linage specific, NWM looks like out-group
Table 4.
Evolutionary forces that underlie the process of divergence in the OLFML2A and OLFML2B primate genes
| Species | OLFML2A | OLFML2B | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| dN | dS | dN = dS | P value | dN | dS | dN = dS | P value | |||
| dN > dS | dN < dS | dN > dS | dN < dS | |||||||
| Apes | ||||||||||
| Human | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Chimpanzee | 0.005 | 0.036 | 1.000 | 1.000 | 0.0001 | 0.009 | 0.031 | 1.000 | 1.000 | 0.0001 |
| Pygmy chimpanzee | 0.004 | 0.031 | 1.000 | 1.000 | 0.0001 | 0.009 | 0.031 | 1.000 | 1.000 | 0.0001 |
| Gorilla | 0.003 | 0.026 | 1.000 | 1.000 | 0.0001 | 0.007 | 0.031 | 1.000 | 1.000 | 0.0001 |
| Orangutan | 0.007 | 0.074 | 1.000 | 1.000 | 0.0001 | 0.015 | 0.048 | 1.000 | 1.000 | 0.0001 |
| Old world monkeys (OWM) | ||||||||||
| Hamadryas baboon | 0.008 | 0.135 | 1.000 | 1.000 | 0.0001 | 0.021 | 0.071 | 1.000 | 1.000 | 0.0001 |
| Baboon | 0.008 | 0.144 | 1.000 | 1.000 | 0.0001 | 0.021 | 0.071 | 1.000 | 1.000 | 0.0001 |
| Rhesus monkey | 0.008 | 0.138 | 1.000 | 1.000 | 0.0001 | 0.023 | 0.073 | 1.000 | 1.000 | 0.0001 |
| Crab-eating macaque | 0.008 | 0.138 | 1.000 | 1.000 | 0.0001 | 0.024 | 0.074 | 1.000 | 1.000 | 0.0001 |
| Pig-tailed macaque | 0.009 | 0.144 | 1.000 | 1.000 | 0.0001 | 0.023 | 0.076 | 1.000 | 1.000 | 0.0001 |
| Sooty mangabey | 0.011 | 0.141 | 1.000 | 1.000 | 0.0001 | 0.019 | 0.066 | 1.000 | 1.000 | 0.0001 |
| Green monkey | 0.010 | 0.141 | 1.000 | 1.000 | 0.0001 | 0.020 | 0.075 | 1.000 | 1.000 | 0.0001 |
| Angolan colobus | 0.019 | 0.151 | 1.000 | 1.000 | 0.0001 | 0.021 | 0.079 | 1.000 | 1.000 | 0.0001 |
| Northern white-cheeked gibbon | 0.008 | 0.088 | 1.000 | 1.000 | 0.0001 | 0.013 | 0.046 | 1.000 | 1.000 | 0.0001 |
| Golden snub-nosed monkey | 0.011 | 0.135 | 1.000 | 1.000 | 0.0001 | 0.022 | 0.073 | 1.000 | 1.000 | 0.0001 |
| Drill | 0.010 | 0.135 | 1.000 | 1.000 | 0.0001 | 0.021 | 0.073 | 1.000 | 1.000 | 0.0001 |
| New world monkeys (NWM) | ||||||||||
| Ma’s night monkey | 0.018 | 0.229 | 1.000 | 1.000 | 0.0001 | 0.128 | 0.241 | 1.000 | 1.000 | 0.0001 |
| Marmoset | 0.022 | 0.247 | 1.000 | 1.000 | 0.0001 | 0.034 | 0.145 | 1.000 | 1.000 | 0.0001 |
| Squirrel monkey | 0.022 | 0.221 | 1.000 | 1.000 | 0.0001 | 0.035 | 0.137 | 1.000 | 1.000 | 0.0001 |
| Prosimians | ||||||||||
| Gray mouse lemur | 0.050 | 0.280 | 1.000 | 1.000 | 0.0001 | 0.060 | 0.327 | 1.000 | 1.000 | 0.0001 |
| Coquerel’s sifaka | 0.044 | 0.329 | 1.000 | 1.000 | 0.0001 | 0.054 | 0.232 | 1.000 | 1.000 | 0.0001 |
| Small-eared galago | 0.050 | 0.402 | 1.000 | 1.000 | 0.0001 | 0.057 | 0.297 | 1.000 | 1.000 | 0.0001 |
| Philippine tarsier | 0.045 | 0.392 | 1.000 | 1.000 | 0.0001 | 0.066 | 0.242 | 1.000 | 1.000 | 0.0001 |
Localization of OLFML2A and OLFML2B proteins in the eye of baboons and humans by immunofluorescence assays
After identifying the ocular tissues which express both mRNAs, and in an effort to determine the cell type of the retina that expresses the genes, we performed the immunoreactivity (IR) analyses in baboon and human retina. IR signal of baboon retina for OLFML2A and OLFML2B are shown in Figs. 4 and 5 respectively. While in humans IR of OLFML2A and OLFML2B is observed in Fig. 6, also see Table 3. More analysis is required in normal human tissue. Only in baboon retina we did a double-immunolocalizaction of OLFML2A and OLFML2B with β-tubulin 3 beta chain, which is a cytoskeletal protein that is currently a neuronal cell marker in the developing and mature human nervous system to diferenciate retinal ganglion cells and astrocytes (Figs. 4, 5).
Fig. 4.
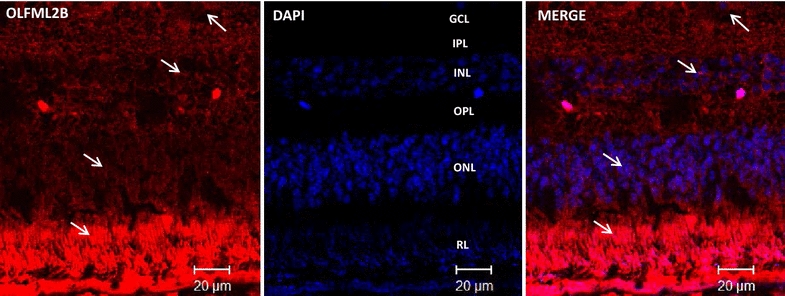
OLFML2A immunodetection in the retina of adult baboons. Confocal images of double stained retina sections to identify cells expressing OLFML2A (red 1st Ab: rabbit polyclonal anti-human OLFML2A 1:500; 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000), β-Tubulin (1st Ab: mouse monoclonal anti-mammal Tubulin 3 beta chain 1:250, 2nd Ab: goat anti mouse IgG FITC 1:250) and Glial Fibrillary Acid Protein (1st Ab: mouse monoclonal anti-GFAP 1:300, 2nd Ab: goat anti mouse IgG FITC 1:250) in baboon retina. Cells nuclei were labeled with DAPI (blue). GCL ganglion cell layer, IPL inner plexiform layer, INL inner nuclear layer, ONL outer nuclear layer, RL rod layer, PE pigmented epithelium
Fig. 5.
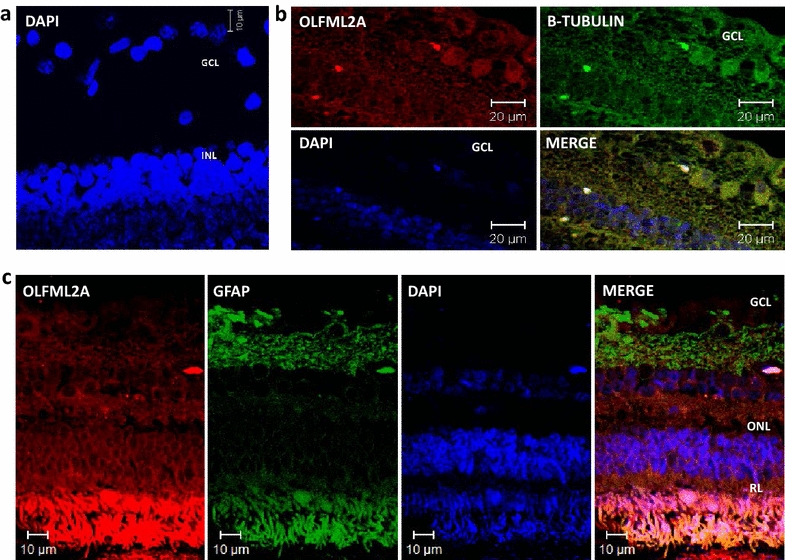
OLFML2A and OLFML2B double stained immunoreactivity in baboon retina. a Negative Control of baboon retina sections wherein the incubation with the 1st Ab is omitted and incubated with only 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000. b Confocal images of double stained retina sections to identify cells expressing OLFML2A (red, 1st Ab: rabbit polyclonal anti-human OLFML2A 1:500; 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000) and β-Tubulin (green, 1st Ab: mouse monoclonal anti-mammal Tubulin 3 beta chain 1:250) in baboon retina. c Confocal images of double stained retina sections to identify cells expressing OLFML2A (red, 1st Ab: rabbit polyclonal anti-human OLFML2A 1:500; 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000) and Glial Fibrillary Acid Protein (green, 1st Ab: mouse monoclonal anti-GFAP 1:300, 2nd Ab: goat anti mouse IgG FITC 1:250) in baboon retina. Cells nuclei were labeled with DAPI (blue). GCL ganglion cell layer, INL inner nuclear layer, ONL outer nuclear layer, PL photoreceptor layer
Fig. 6.
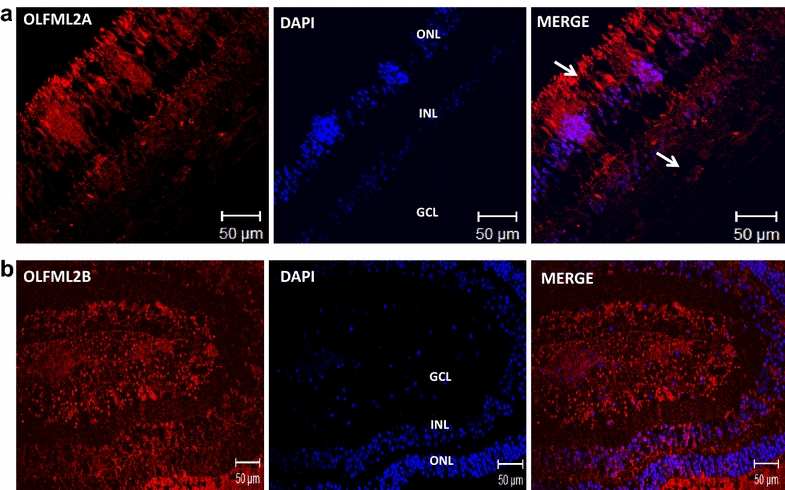
OLFML2A and OLFML2B immunoreactivity in human retina. a Confocal image of stained retina sections to identify cells expressing OLFML2A (red, 1st Ab: rabbit polyclonal anti-human OLFML2A 1:500; 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000). b Confocal image of stained retina seccions to identify cells expressing OLFML2B (red, 1st Ab: rabbit polyclonal anti-human OLFML2B 1:500; 2nd Ab: goat anti-rabbit IgG-Cy3® 1:4000). Cells nuclei were labeled with DAPI (blue). GCL ganglion cell layer, INL inner nuclear layer, ONL outer nuclear layer, PL photoreceptor layer
Discussion
Olfactomedin was originally identified as the major component of the mucus layer that surrounds the chemosensory dendrites of olfactory neurons [15]. Subsequently, a vast numbers of proteins that share a ~250 amino acid domain homologous to olfactomedin were discovered in animals ranging from nematodes to humans [4]. One of these proteins was the olfactomedin like 2 proteins (OLFML2A and OLFML2B), also known as photomedins (−1 and −2, respectively), which were first identified and characterized in mouse retina [3]. But so far, it is not known if some primates such as baboon and human express these photomedins, whereas some olfactomedin-proteins like myocilin are associated with eye diseases such as glaucoma. Based on the above in the present study we cloned, sequenced and characterized the olfactomedin-like 2 cDNAs (OLFML2A and OLFML2B) in baboon (Papio hamadryas) and human from different ocular tissues. This is the first study that identifies expression of these genes in the eye of primates. Nevertheless, in mouse retina by northern blot analysis two RNA transcripts for OLFML2A (5 and 3.5 kb) and one for OLFML2B (3.5 kb) have been reported [15]. The authors suggest that these two RNA species for OLFML2A could be due to alternative splicing [3]. Also in human podocyte cells have been reported two OLFML2A mRNA variants [9]. However, we did not find in our study other transcripts of genes that may indicate the presence of isoforms derived by alternative splicing. This may be due to differences in the animal model studied; the tissues, and also the RNA transcripts found in mouse were not cloned and sequenced [3].
However, it is well known that some members of the subfamily of olfactomedin are expressed in the eye [3, 15, 16]. Similarly, the expression pattern suggests that mechanisms of regulation of gene expression are similar in the two species. It also suggests that the OLFML2A and OLFML2B genes might have similar physiological effects. However more studies are needed. The most extensively studied olfactomedin protein to date is myocilin (MYOC), which was first discovered in human trabecular meshwork cells [17, 18]. Several studies suggest that MYOC plays an important role in regulation in ocular hypertensin. Ocular hypertension is a major risk factor for glaucoma, leadin cause of blindness [15, 19]. Trabecular meshwork is a connective tissue that regulates the outflow at the iridocorneal angle of the eye and, hence, controls intraocular pressure [19], aqueous humor is continually produced by the ciliary body and it is in direct contact with the anterior surface of the lens, iris, and corneal endothelial cells, before draining out of the eye via the trabecular meshwork [20]. MYOC expression was observed in cornea, ciliary body, iris, sclera, optic nerve and retina, in human and mouse eye [21–23]. It is known that these tissues get their nutrientes from the aqueous humor and also export their metabolites, allowing an exchange with neighboring tissues [20]; it is also known that glaucoma is a group of progressive neurodegenerative multifactorial diseases, characterized by the loss of retinal ganglion cells (RGCs), optic nerve excavation, and axonal degeneration leading to irreversible vision loss [24]. Interestingly, we find the expression of photomedins (Table 3) in some of the tissues that express MYOC (cornea, lens, iris, and retina). The function of MYOC is still not known [19], however its been reported that MYOC may interact with other olfactomedin knows as optimedin (OLFM3), these wo proteins are expressed in human trabecular meswork and retina, and may be involved in glaucoma disease [25]. It would be interesting to study the interaction of myocilin and photomedins and see the correlation with these proteins in ocular pathologies.
The functional roles of the olfactomedin proteins in the retina are still not known [17]. Olfactomedins appear to be critical mediators for development of nervous systems and hematopoiesis [19]. Some others members are identified to be associated with human disease processes like glaucoma and cancer [4, 19]. Overexpression studies and inhibition of protein expression in zebrafish embryos showed that Noelin (olfactomedin 1), has profound effect on eye development, eye size, the projection field of retinal ganglion cells to the optic tectum, and extension and branching of retinal ganglion cell axons [19, 26]. Further studies showed in zebrafish that Noelin promotes retinal ganglion cell axon growth [27]. OLFM1 and OLFM2 are preferentially expressed in the developing retinal ganglion cells [16] in rat and mouse. In zebrafish eye OLFM2 was detected in the retinal ganglion cell layer and the inner nuclear layer [28]. OLFML2 in humans was found by RT-PCR in corneal endothelium, uvea, lens and retina-RPE. In baboon it was found in cornea, lens, iris, and retina-RPE [17]. Others authors have reported the expression of OLFM4 in mouse Müller glial cells. In the retina, OLFML2A was selectively expressed in the outer segment of photoreceptor cells and OLFML2B was expressed in all retinal neurons in a mouse. These proteins bound to other proteins like chondroitin sulphate-E and heparin suggest that photomedins-1 and -2 are extracellular proteins capable of binding of proteoglycans [3]. OLFML3 may play a possible role in angiogenesis in ocular tissues and it has been proposed that this protein may play a role in anterior segment and retinal diseases [17].
Positive selection (dN > dS) implies that the substitutions, mostly non-synonymous, are functional and benefit the organism, conferring some evolutionary advantage. While purifying selection (dN < dS) indicates that evolutionary pressure has been relaxed. The dN and dS rates show that the evolutionary force, actually acting on these, is the purification of the selection (P < 0.05). It fits the hypothesis that purifying of the selection is a clue that these genes are functional in the studied species, because there are not functional genes that do not fit this hypothesis. Similar expression profiles of human and baboon OLFML2A and OLFML2B genes, suggests that they have similar binding sites for known transcriptional factors. The phylogenetic relationship between NWM, OWM and apes OLFML2A proteins was determined to evaluate their evolution in primates. The phylogenetic tree (Fig. 1) shows three clades in a linage-specific manner. These clades correspond to NWM, OWM and apes, finally galago (out-group). The tree’s topology, branch length, and bootstrap values are similar using either phylogenetic method (ML/NJ/UPGMA). This confirms a clear orthology within the OLFML2A gene. While OLFML2B orthology is not clear (Fig. 2). The tree’s topology does not fit in a lineage-specific manner. It could be for many reasons, such as Ma’s night monkey sequence is shorter than the rest or it may be because more species should be included in the study.
Given these finding together, olfactomedins play essential roles in development and cell differentiation, also their effects are mediated through intercellular interactions, sometimes with other proteins or extracellular matrix components [15, 19], and some olfactomedin are implicated in important pathologies.
OLFML2A and OLFML2B seem to play an important role in ocular tissues, however the functions of these olfactomedins are still unknown. Therefore, further studies are needed to elucidate the role of these proteins in embryonic development, investigate its biological function, their protein interactions and diseases.
Conclusions
The function of olfactomedin proteins in the eye, especially OLFML2A and OLFML2B, is still unknown; a lot of work is needed to clarify their actual role. Due to the high similarity between baboon and human olfactomedin expression, the baboon is a powerful model to deduce the physiological functions of these proteins in the eye.
Authors’ contributions
DCPI, MDGR, HABS, DPR, RGA, LEMD, MDMF, OBQ, GRA, MLM, CAD and IPRS conceived and designed the experiments. JMH, KMH, DCPI, HAGS, MDGR and IPRS participated in the baboon eye collection; JMH, KMH, RGG and VMBD enrolled patients from their respective institutes; DCPI, MDGR, HABS and IPRS performed the RT-PCR experiments; RGA, HABS, MDGR and IPRS performed the phylogenetic analysis; DCPI, MLM, CAD and IPRS performed the immunofluorescence experiments. DCPI, MDGR, HABS, RGA, LEMD, MDMF and IPRS contributed to the writing of the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The present work was supported by the Grant Number 16769 from the Mexican Council of Science and Technology, CONACyT. The authors gratefully acknowledge the critical reading of the manuscript by Dr. Sergio Lozano-Rodriguez.
Competing interests
The authors declare that they have no competing interests.
Availability of data and material
The sequences obtained from baboon clones were deposited in the GenBank with the access number NM_182487 for OLML2A and AK316154 for OLFML2B.
Ethics approval and consent to participate
Animal protocols were designed and developed according to the ethical guidelines of the Institutional Animal Care and Use Committee of the Texas Institute of Biomedical Research (TIBR).
Biopsies from human eyes were collected at the Department of Ophthalmology of the “Dr. José Eleuterio González” University Hospital of the Universidad Autónoma de Nuevo Leon, in Monterrey, Mexico. Specimens came from programmed eye surgery procedures where ocular tissues were removed by medical indication. All patients signed an informed consent according to the ethics committee guidelines of the institution under the following registration number OF10-005.
Abbreviations
- OLF
olfactomedin
- OLFML2A
olfactomedin like 2A
- OLFML2B
olfactomedin like 2B
- OLFML3
olfactomedin like 3
- OLFML-1
olfactomedin like 1
- aa
aminoacids
- nt
nucleotides
- bp
base pair
- kb
kilo base
- °C
celsius
- RT-PCR
reverse transcription-polymerase chain reaction
- cDNA
complementary deoxyribonucleic acid
- RNA
ribonucleic acid
- mRNA
messenger RNA
- CDS
coding DNA sequence
- min
minutes
- sec
seconds
- µl
microliter
- mM
mili molar
- nm
nanometer
- IgG
inmmunoglobulin G
- UV
ultraviolet
- OWM
old world monkey
- NWM
new world monkey
- GCL
ganglion cell layer
- IPL
inner plexiform layer
- INL
inner nuclear layer
- ONL
outer nuclear layer
- RL
rod layer
- PE
pigmented epithelium
Contributor Information
Diana Cristina Pérez-Ibave, Email: dianics83@gmail.com.
Rafael González-Alvarez, Email: rgonzaleza5@hotmail.com.
Margarita de La Luz Martinez-Fierro, Email: margaritamf@uaz.edu.mx.
Gabriel Ruiz-Ayma, Email: ayma.gabriel@gmail.com.
Maricela Luna-Muñoz, Email: lunam@unam.mx.
Laura Elia Martínez-De-Villarreal, Email: lauraeliamv@gmail.com.
María De Lourdes Garza-Rodríguez, Email: lulugarza87@gmail.com.
Diana Reséndez-Pérez, Email: diaresendez@gmail.com.
Jibran Mohamed-Noriega, Email: jibran.mohamed@gmail.com.
Raquel Garza-Guajardo, Email: rgguajardo@yahoo.com.
Víctor Manuel Bautista-De-Lucío, Email: vbautistal@institutodeoftalmologia.org.
Karim Mohamed-Noriega, Email: drkmohamed1@gmail.com.
Oralia Barboza-Quintana, Email: oralibarboza@gmail.com.
Carlos Arámburo-De-La-Hoz, Email: aramburo@unam.mx.
Hugo Alberto Barrera-Saldaña, Email: habarrera@gmail.com.
Irám Pablo Rodríguez-Sánchez, Email: iramrodriguez@gmail.com.
References
- 1.Yokoe H, Anholt RR. Molecular cloning of olfactomedin, an extracellular matrix protein specific to olfactory neuroepithelium. Proc Natl Acad Sci USA. 1993;90(10):4655–4659. doi: 10.1073/pnas.90.10.4655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mukhopadhyay A, Talukdar S, Bhattacharjee A, Ray K. Bioinformatic approaches for identification and characterization of olfactomedin related genes with a potential role in pathogenesis of ocular disorders. Mol Vis. 2004;10:304–314. [PubMed] [Google Scholar]
- 3.Furutani Y, Manabe R, Tsutsui K, Yamada T, Sugimoto N, Fukuda S, et al. Identification and characterization of photomedins: novel olfactomedin-domain-containing proteins with chondroitin sulphate-E-binding activity. Biochem J. 2005;389(Pt 3):675–684. doi: 10.1042/BJ20050120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zeng LC, Han ZG, Ma WJ. Elucidation of subfamily segregation and intramolecular coevolution of the olfactomedin-like proteins by comprehensive phylogenetic analysis and gene expression pattern assessment. FEBS Lett. 2005;579(25):5443–5453. doi: 10.1016/j.febslet.2005.08.064. [DOI] [PubMed] [Google Scholar]
- 5.Tomarev SI, Nakaya N. Olfactomedin domain-containing proteins: possible mechanisms of action and functions in normal development and pathology. Mol Neurobiol. 2009;40(2):122–138. doi: 10.1007/s12035-009-8076-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Adam MF, Belmouden A, Binisti P, Brezin AP, Valtot F, Bechetoille A, et al. Recurrent mutations in a single exon encoding the evolutionarily conserved olfactomedin-homology domain of TIGR in familial open-angle glaucoma. Hum Mol Genet. 1997;6(12):2091–2097. doi: 10.1093/hmg/6.12.2091. [DOI] [PubMed] [Google Scholar]
- 7.Barembaum M, Moreno TA, LaBonne C, Sechrist J, Bronner-Fraser M. Noelin-1 is a secreted glycoprotein involved in generation of the neural crest. Nat Cell Biol. 2000;2(4):219–225. doi: 10.1038/35008643. [DOI] [PubMed] [Google Scholar]
- 8.Moreno TA, Bronner-Fraser M. The secreted glycoprotein Noelin-1 promotes neurogenesis in Xenopus. Dev Biol. 2001;240(2):340–360. doi: 10.1006/dbio.2001.0472. [DOI] [PubMed] [Google Scholar]
- 9.Sistani L, Rodriguez PQ, Hultenby K, Uhlen M, Betsholtz C, Jalanko H, et al. Neuronal proteins are novel components of podocyte major processes and their expression in glomerular crescents supports their role in crescent formation. Kidney Int. 2013;83(1):63–71. doi: 10.1038/ki.2012.321. [DOI] [PubMed] [Google Scholar]
- 10.Boutros R, Stokes N, Bekaert M, Teeling EC. UniPrime2: a web service providing easier Universal Primer design. Nucleic Acids Res. 2009;37(4):W209–W213. doi: 10.1093/nar/gkp269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22(22):4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 14.Zhang J, Rosenberg HF, Nei M. Positive Darwinian selection after gene duplication in primate ribonuclease genes. Proc Natl Acad Sci USA. 1998;95(7):3708–3713. doi: 10.1073/pnas.95.7.3708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kulkarni NH, Karavanich CA, Atchley WR, Anholt RR. Characterization and differential expression of a human gene family of olfactomedin-related proteins. Genet Res. 2000;76(1):41–50. doi: 10.1017/S0016672300004584. [DOI] [PubMed] [Google Scholar]
- 16.Sultana A, Nakaya N, Senatorov VV, Tomarev SI. Olfactomedin 2: expression in the eye and interaction with other olfactomedin domain-containing proteins. Invest Ophthalmol Vis Sci. 2011;52(5):2584–2592. doi: 10.1167/iovs.10-6356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rodriguez-Sanchez IP, Garza-Rodriguez ML, Mohamed-Noriega K, Voruganti VS, Tejero ME, Delgado-Enciso I, et al. Olfactomedin-like 3 (OLFML3) gene expression in baboon and human ocular tissues: cornea, lens, uvea, and retina. J Med Primatol. 2013;42(3):105–111. doi: 10.1111/jmp.12037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stone EM, Fingert JH, Alward WL, Nguyen TD, Polansky JR, Sunden SL, et al. Identification of a gene that causes primary open angle glaucoma. Science. 1997;275(5300):668–670. doi: 10.1126/science.275.5300.668. [DOI] [PubMed] [Google Scholar]
- 19.Anholt RR. Olfactomedin proteins: central players in development and disease. Front Cell Dev Biol. 2014;2:6. doi: 10.3389/fcell.2014.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kim EB, Kim HK, Hyon JY, Wee WR, Shin YJ. Oxidative stress levels in aqueous humor from high myopic patients. Korean J Ophthalmol. 2016;30(3):172–179. doi: 10.3341/kjo.2016.30.3.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Swiderski RE, Ross JL, Fingert JH, Clark AF, Alward WL, Stone EM, et al. Localization of MYOC transcripts in human eye and optic nerve by in situ hybridization. Invest Ophthalmol Vis Sci. 2000;41(11):3420–3428. [PubMed] [Google Scholar]
- 22.Takahashi H, Noda S, Imamura Y, Nagasawa A, Kubota R, Mashima Y, et al. Mouse myocilin (Myoc) gene expression in ocular tissues. Biochem Biophys Res Commun. 1998;248(1):104–109. doi: 10.1006/bbrc.1998.8917. [DOI] [PubMed] [Google Scholar]
- 23.Noda S, Mashima Y, Obazawa M, Kubota R, Oguchi Y, Kudoh J, et al. Myocilin expression in the astrocytes of the optic nerve head. Biochem Biophys Res Commun. 2000;276(3):1129–1135. doi: 10.1006/bbrc.2000.3591. [DOI] [PubMed] [Google Scholar]
- 24.Madeira MH, Ortin-Martinez A, Nadal-Nicolas F, Ambrosio AF, Vidal-Sanz M, Agudo-Barriuso M, et al. Caffeine administration prevents retinal neuroinflammation and loss of retinal ganglion cells in an animal model of glaucoma. Sci Rep. 2016;6:27532. doi: 10.1038/srep27532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Torrado M, Trivedi R, Zinovieva R, Karavanova I, Tomarev SI. Optimedin: a novel olfactomedin-related protein that interacts with myocilin. Hum Mol Genet. 2002;11(11):1291–1301. doi: 10.1093/hmg/11.11.1291. [DOI] [PubMed] [Google Scholar]
- 26.Nakaya N, Lee HS, Takada Y, Tzchori I, Tomarev SI. Zebrafish olfactomedin 1 regulates retinal axon elongation in vivo and is a modulator of Wnt signaling pathway. J Neurosci. 2008;28(31):7900–7910. doi: 10.1523/JNEUROSCI.0617-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nakaya N, Sultana A, Lee HS, Tomarev SI. Olfactomedin 1 interacts with the Nogo A receptor complex to regulate axon growth. J Biol Chem. 2012;287(44):37171–37184. doi: 10.1074/jbc.M112.389916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lee JA, Anholt RR, Cole GJ. Olfactomedin-2 mediates development of the anterior central nervous system and head structures in zebrafish. Mech Dev. 2008;125(1–2):167–181. doi: 10.1016/j.mod.2007.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]