

SUMMARY
The recognition of a new family of rhodopsins in marine planktonic bacteria, proton-pumping proteorhodopsin, expanded the known phylogenetic range, environmental distribution, and sequence diversity of retinylidene photoproteins. At the time of this discovery, microbial ion-pumping rhodopsins were known solely in haloarchaea inhabiting extreme hypersaline environments. Shortly thereafter, proteorhodopsins and other light-activated energy-generating rhodopsins were recognized to be widespread among marine bacteria. The ubiquity of marine rhodopsin photosystems now challenges prior understanding of the nature and contributions of “heterotrophic” bacteria to biogeochemical carbon cycling and energy fluxes. Subsequent investigations have focused on the biophysics and biochemistry of these novel microbial rhodopsins, their distribution across the tree of life, evolutionary trajectories, and functional expression in nature. Later discoveries included the identification of proteorhodopsin genes in all three domains of life, the spectral tuning of rhodopsin variants to wavelengths prevailing in the sea, variable light-activated ion-pumping specificities among bacterial rhodopsin variants, and the widespread lateral gene transfer of biosynthetic genes for bacterial rhodopsins and their associated photopigments. Heterologous expression experiments with marine rhodopsin genes (and associated retinal chromophore genes) provided early evidence that light energy harvested by rhodopsins could be harnessed to provide biochemical energy. Importantly, some studies with native marine bacteria show that rhodopsin-containing bacteria use light to enhance growth or promote survival during starvation. We infer from the distribution of rhodopsin genes in diverse genomic contexts that different marine bacteria probably use rhodopsins to support light-dependent fitness strategies somewhere between these two extremes.
INTRODUCTION
Light is one of the most pervasive phenomena in nature, and life has had to cope with it from the very beginning. Like every other environmental factor, light can be a threat or an asset. Living beings have developed a variety of molecules to deal with both these aspects of light. Carotenoids were likely some of the earliest light-absorbing molecules appearing in evolution. They may have originated as a way to reinforce the structure of archaeal cell walls (1). Over evolutionary time, carotenoids were recruited for a variety of other cell functions, particularly for protection against blue and UV radiation and the oxygen radicals they generate.
Among proteins, a variety of molecules (e.g., sensory rhodopsins [SRs], phytochromes, photoactive yellow protein [PYP], phototropin [LOV], and blue-light sensing using FAD [BLUF]) act as light sensors that can modulate the behavior and life history of organisms and influence, for example, phototaxis, diurnal and seasonal rhythms, synthesis of protective pigments, and expression of particular genes. These molecules are beyond the scope of the present work (but see references 2 and 3). With regard to light-driven energy-generating mechanisms, only two general biological systems are known to be capable of net energy conservation: one is based on chlorophylls and the other on retinal molecules. These two systems have very distinct characteristics.
Chlorophyll systems are complex, being composed of dozens of proteins and pigments forming reaction centers, antenna complexes, and photosystems. Synthesis of the key pigment, chlorophyll, requires at least 16 specific metabolic steps with their corresponding enzymes. Moreover, cofactors such as Mg2+ are needed, imposing extra requirements on the organisms with such systems. Chlorophyll systems are present in the domains Bacteria and Eukarya and are very efficient at transforming light energy into reducing power as NADPH or NADH, as well as ATP during photosynthetic electron flow. Contemporary chlorophyll-based oxygenic photosynthesis, by supplying both energy and reducing power, using one of the most abundant and biologically compatible reductants on the planet (water), has evolved to become the most prevalent photosystem on Earth.
Retinal-based photosystems (e.g., rhodopsins), on the other hand, consist essentially of only one protein, an opsin, and one chromophore, retinal. The retinal has no cofactors and is produced in one metabolic step from the widely distributed carotenoid beta-carotene. Since light-absorbing photoprotective carotenoid metabolic pathways were present early in evolutionary history, their recruitment for retinal biosynthesis is relatively straightforward. Retinal-based photosystems are present in members of all three domains of life and have been modified to perform several different biological functions: proton pumping, ion pumping, light sensing and gene regulation, phototaxis, and vision.
According to their amino acid sequences, opsin proteins fall in two large groups: type I, found in Bacteria, Archaea, and Eukarya, and type II found only in animals (4, 5). The primary structures of these two types of rhodopsins are so divergent that it has been speculated that their secondary and tertiary structural similarities arise from convergent evolution. Results from a recent empirical test of this hypothesis using mutational analyses, however, contradicted central predictions of convergent evolution, implying that type I and II opsins most likely share an ancestor (6). Within type I opsins, Ihara et al. (7) concluded that many of the opsin genes found in haloarchaea were paralogous. Type II opsins are linked to a G protein located on the cytoplasmic side of the membrane, and they transform the light detected into a signal to this G protein. Thus, they are also called “G protein-coupled receptors.” Type II opsins are responsible for vision in animals (5, 8) and are beyond the scope of the present review.
The recognition of microbial rhodopsins as light-driven ion pumps began in the 1970s when Stoeckenius and Oesterhelt decided to study the colorful inhabitants of crystallizer ponds in solar salterns (9, 10). The photopigment they found in Halobacterium salinarum (Archaea) they named bacteriorhodopsin (BR), and its discovery initiated a long and detailed series of structural, biochemical, and biophysical studies (see references 4 and 11 for reviews). As a result, bacteriorhodopsin quickly became one of the most well-understood membrane proteins at the time and a model for generally understanding membrane protein interactions, structure, and activities. Moreover, a variety of physiological roles of rhodopsins in haloarchaea were uncovered (12–14). Another area of interest was exploiting such molecules for harnessing light energy for biotechnological applications (see, for example, references 15 and 16). After the initial discoveries of Stoeckenius and Oesterhelt, four other types of rhodopsins were found in haloarchaeal isolates (14, 17–19), further igniting research into microbial rhodopsins.
Rhodopsin-like proteins were later discovered in eukaryotic microorganisms, including algae and fungi. The discovery rate increased substantially at the turn of the century, since the sequencing of genomes revealed rhodopsin-like genes in many organisms. One of the more notable findings is that of the channelrhodopsins (ChRs) in the single-cell photosynthetic green alga Chlamydomonas reinhardtii strains 495 and CC-503 (20–22). Two different genes that acted as light-gated ion channels were present. These were similar to the sensory rhodopsins previously described in haloarchaea. However, since these proteins belonged to eukaryotes, the possibility of bioengineering them into human cells appeared. This approach has matured into a very successful neuron bioengineering technology: optogenetics (23, 24). When two different channelrhodopsins are introduced into brain cells, for example, they can be stimulated to depolarize the cell membrane by shining specific wavelengths of light on them. In this way, the triggering of the nervous impulse can be stimulated or inhibited in an extremely precise way (at the single-cell level). Thus, epileptic seizures have been prevented in mice via channelrhodopsin-enabled, light-directed mediation of nerve impulses. Many more channelrhodopsins have been found in algae by screening transcriptomes (25), while both phototaxis receptors and proton pumps have been found in fungi (26, 27).
The study of rhodopsins received another major impulse when metagenomic studies revealed the presence of a new family of rhodopsins, dubbed proteorhodopsin (PR), in marine bacteria (28). In short order following this discovery, microbial rhodopsins were suddenly recognized to be not just an exception but probably the rule, being present in perhaps half or more of all “heterotrophic” bacteria living in the surface ocean. The ubiquity of proteorhodopsins in bacterioplankton was recognized to potentially impact not only microbial survival ability but also carbon and energy fluxes in the world's oceans. It must be emphasized that this discovery occurred through the use of metagenomics and discovery-driven research. With most techniques and research approaches, what one finds in nature is what one is looking for; the rest remains invisible. The discovery of PR demonstrated the power of metagenomics, especially when combined with subsequent hypothesis testing at the physiological or biochemical level (in this case, experiments with heterologously expressed PR validated its retinal binding and light-driven proton-pumping activities [28]). The biochemistry and photobiology of this protein have also been studied in detail (29), and a blue-tuned PR variant (Med12BPR) was the first for which the crystal structure was determined (30).
In this review, we report on the diversity, evolution, physiology, and ecology of microbial type I rhodopsins, with particular focus on ion-pumping rhodopsins in Bacteria and Archaea in the marine environment. We only briefly allude to some important aspects of the biophysics and biochemistry of rhodopsins relevant to the physiology and ecology of Bacteria and Archaea, since excellent comprehensive presentations on the structure, biophysics, and molecular mechanism of rhodopsins are already available (8, 31–34). For sensory rhodopsins, we refer to insightful reviews (2, 8). Nevertheless, to allow assessment of the genetic diversity of rhodopsins in marine microorganisms, we carried out a phylogenetic analysis of the full range of type I rhodopsins, including also sequences from eukaryotes and from nonmarine environments. We therefore also briefly mention some features of such rhodopsins. Most recently, a wealth of new information on the microbial ecology of rhodopsins has resulted from studies of proton-pumping proteorhodopsins in marine bacteria, and these rhodopsins will be a central focus point of this review, with some discussion also of more recently discovered rhodopsin types in marine viruses and single-celled eukaryotes.
TYPE I RHODOPSIN CONSERVATION AND DIVERSITY
Rhodopsins are based on the covalent union of a retinal chromophore molecule to an opsin protein (apoprotein). The opsin amino acid chain folds to form seven alpha-helices linked by short, apparently unorganized, sections (Fig. 1). This structure is conserved across all domains of life. The amino acid and nucleotide sequences, however, are very different between diverse life forms. For proteorhodopsin, this transmembrane apoprotein is encoded by the prd gene (the gene nomenclature follows that proposed upon original experimental validation of PR functioning through mutational analysis of the gene encoding proteorhodopsin in Vibrio sp. strain AND4 [35]) and requires the binding of the chromophore cofactor retinal to become functional (Fig. 1). Retinal is produced through cleavage of beta-carotene, and Sabehi et al. (36) identified a potential pathway for retinal biosynthesis in marine bacteria. Sabehi et al. (37) thereafter demonstrated the function of the enzyme encoded by blh (bacteriorhodopsin-related-protein-like homolog protein) in cleaving beta-carotene to retinal after expressing it in Escherichia coli. Synthesis of beta-carotene is mediated by the carotenoid biosynthesis genes crtEBIY from isoprenoid precursors. While basic isoprenoid synthesis pathways are found in a large number of living organisms, the ability to synthesize specific carotenoids is more limited, which has implications for how PR phototrophy is distributed among divergent microbial taxa (see “Lateral Transfer of Rhodopsin Genes” below).
FIG 1.
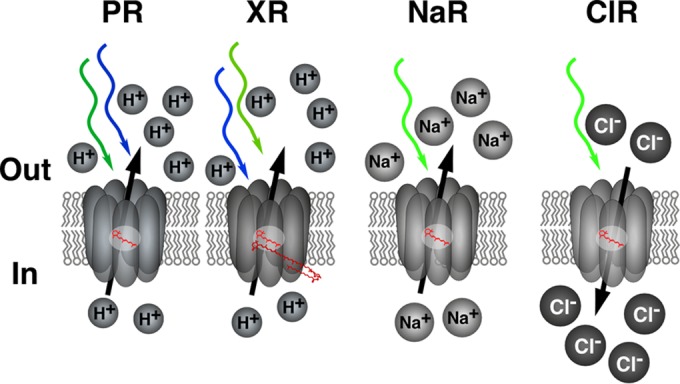
Schematic representation of different proton, sodium, and chloride rhodopsin pumps. Colored arrows show the different wavelengths at which each rhodopsin absorbs. In the case of proteorhodopsin (PR), the arrows represent the absorption of blue-tuned (∼490-nm) and green-tuned (∼530-nm) rhodopsins. For xanthorhodopsin (XR), the arrows represent absorption at different wavelengths (∼470 and 520 nm) due to XR having an additional carotenoid antenna molecule. Peak absorption maxima and ranges of absorption for green- and blue-tuned PRs in the visible light range are detailed in Fig. 3, top panel. NaR, sodium-pumping rhodopsin; ClR, chloride-pumping rhodopsin. (Redrawn and modified from reference 32.)
Rhodopsin Types
Type I opsins have been found in Bacteria (Proteobacteria, Bacteroidetes, Cyanobacteria, Firmicutes, Actinobacteria, Deinococcus-Thermus, Chloroflexi, Planctomycetes, and Ktedonobacteria), Euryarchaeota (especially haloarchaea), and Eukarya (Alveolata, including dinoflagellates, diatoms, and cryptophytes; fungi, including both Basidiomycota and Ascomycota; and Viridiplantae, including Glaucophyta, Chlorophyta, and Streptophyta) (see reviews by Sharma et al. [38] and Ernst et al. [5] for overviews). New types of rhodopsins are constantly being discovered, and the distribution of recognized types is extended to taxa where they were previously not observed. Thus, it is challenging to present a full and coherent picture of the diversity of microbial rhodopsins. Moreover, since amino acid sequences have diverged considerably, alignments for phylogenetic analyses are delicate, and inferred trees are sensitive to which sequences are included. There is as yet no comprehensive phylogenetic tree of rhodopsins in the literature, although a database (MicRhoDE) for phylogenetic analysis of microbial rhodopsins was presented in June 2015 (39). Figure 2 shows our best effort to build a robust tree with all the different types of type I rhodopsins known in March 2016. It must be kept in mind that future discoveries may change some aspects of the topology of the tree.
FIG 2.

Phylogenetic relationships of rhodopsins. An unrooted tree of rhodopsins from the three domains of life is shown. A total of 756 rhodopsin sequences were downloaded from RefSeq (http://www.ncbi.nlm.nih.gov/refseq/; release 75, 14 March 2016), following identification using the PFAM for bacteriorhodopsin-like protein (Bac_rhodopsin, PF01036). The sequences were aligned using MUSCLE and further edited with Gblocks to eliminate highly divergent regions. The tree was constructed by the maximum-likelihood method using the LG model of amino acid substitution rates with empirical amino acid frequencies and the gamma model of rate heterogeneity (PROTGAMMALGF) in RAxML. Bootstrap support values higher than 50% are shown above branches based on 100 pseudoreplicates. Only the region of the peptide that spans the PFAM protein family specific for rhodopsin was considered in the alignment. GenBank accession numbers are shown in parentheses for the individual sequences. Blue asterisks indicate individual organisms for which there is experimental evidence for the function of the rhodopsin; in the case of collapsed branches (marked in red), asterisks indicate that there is experimental evidence for at least one organism. Numbers within circles are the number of sequences in the collapsed branches. The scale bar represents substitutions per site.
The tree shows two main clusters of type I rhodopsins (Fig. 2), which was already noted by Sharma et al. (38). Cluster A has mostly archaeal sequences but also (at least) three eukaryal clusters and a few bacterial sequences. Cluster B has mostly bacterial sequences but also includes a few archaeal sequences and (at least) one eukaryal cluster. It is tempting to compare this clustering to that of the rRNA tree, where Bacteria branched off earlier, while Archaea and Eukarya separated from each other later. The sequences found in the “wrong” cluster (for example bacterial sequences in cluster A) might be the result of lateral gene transfer (LGT) across domains (see Genomics of Marine Bacterial and Archaeal Rhodopsins below). The first major branching, therefore, seems linked to phylogeny. An ancient rhodopsin gene was inherited by both branches and then evolved separately in Bacteria on the one hand and in Archaea and Eukarya on the other. LGT events then occurred in both directions. Determining the relative importance of vertical inheritance versus LGT remains an important future challenge for understanding type I rhodopsin evolutionary history.
Within each major cluster (clusters A and B), rhodopsins form smaller clusters with different functions. In many cases, bootstrap support for these clusters is low, so that associations may change when more rhodopsins are discovered. Moreover, biochemical evidence of the specific activities and ion specificities has been obtained for only a limited number of rhodopsins (see “Proton-pumping rhodopsins” and “Chloride and sodium ion-pumping rhodopsins” below). Furthermore, the physiological functions and roles of these photoproteins have been determined for only a very few microorganisms (see Experimental Assessment of Rhodopsin Photoheterotrophy in Heterologous Hosts and Native Aquatic Bacteria below). The remaining sequences are assumed to carry out the same function as those in the same cluster. However, physiological, biochemical, and biophysical experiments are necessary to confidently assign a function to any newly discovered rhodopsin.
One interesting aspect is that while many microorganisms lack rhodopsin genes, others have several, and in these cases, they may encode rhodopsins with different functions. For example, Halobacterium salinarum has two sensory rhodopsins (SR I and SR II), a halorhodopsin (HR), and bacteriorhodopsin. In a similar way, genes encoding distinct rhodopsins are present in some marine bacteria, such as Dokdonia eikasta NBRC 100814T (Flavobacteriaceae family, Bacteroidetes phylum; isolate previously called Krokinobacter eikastus NBRC 100814T), Dokdonia PRO95, and Gillisia limnaea R-8282T, that contain a sodium-pumping rhodopsin (NaR) and a regular proton pump (40–42), where some of the NaRs were originally identified as a novel group of xanthorhodopsin (XR)-like rhodopsins (41, 42). Notably, Nonlabens marinus S1-08T (Flavobacteriaceae) carries genes encoding three functionally and structurally distinct rhodopsins: chloride-, sodium-, and proton-pumping rhodopsins (43). Another example is the freshwater green alga C. reinhardtii CC-503, which has at least five rhodopsin genes, encoding one rhodopsin (Cop), two channelrhodopsins (ChR1 and ChR2), and two rhodopsins linked to histidine kinases (HKR1 and HKR2) (44). The Salinibacter ruber M31T genome, in turn, encodes four rhodopsins, one related to those from bacteria (xanthorhodopsin) and the others clearly having archaeal origins, i.e., an inward-directed chloride pump and two SR Is (45). Strikingly, the marine cryptophyte alga Guillardia theta CCMP2712 and the dinoflagellate Oxyrrhis marina CCMP1788 genomes carry over 50 and 40 rhodopsin genes, respectively (46–49). Among the G. theta proteins are at least 11 light-gated ion channels (also called channelrhodopsins). In contrast to the cation-conducting channelrhodopsins (CCRs) previously discovered in C. reinhardtii and many other green (chlorophyte) algae, there are two channelrhodopsins in G. theta that belong to a new family, namely, light-gated anion channels (ACRs) (48). Recently, CCRs that are structurally distinct from chlorophyte CCRs have also been identified in the cryptophyte G. theta, apparently having converged on cation channel function independently (49). Intriguingly, so far only CCRs have been found in chlorophyte algae, while ACRs as well as CCRs have been discovered in cryptophyte algae. Whether this fact will hold when more algae are examined, however, is unknown.
Phylogenetic Distribution of Microbial Rhodopsins According to Biochemical Functions
In cluster B, three different functions have evolved (at least that are known today): proton pumps (proteorhodopsins, xanthorhodopsins, and actinorhodopsins), halogen ion pumps, and sodium ion pumps. Some of these can also have dual functionalities (40, 50). Cluster A, in contrast, contains several more functions, including proton pumps (bacteriorhodopsins, proteorhodopsins), halogen and other ion pumps (halorhodopsins), light sensors (sensory rhodopsins I, II, and III), light-gated channels (channelrhodopsins), and light-sensing two-component systems (enzymerhodopsins, HKRI histidine kinase). The last two functions have appeared only in Eukarya. The appearance of these functional clusters seems best explained by gene duplication followed by specialization of the paralogous genes. It is interesting that, apparently, this process occurred in the two large clusters independently.
The most straightforward function carried out by rhodopsins is pumping ions across the cell membrane. This can be achieved by a single rhodopsin unit (i.e., opsin with bound retinal). A photon of the appropriate wavelength is absorbed by the retinal, and the conformational changes induced result in one ion being absorbed on one side and one ion released on the other side of the membrane. Rhodopsins pumping protons, chloride, or sodium ions exist in Bacteria, Archaea, and Eukarya. Sensory rhodopsins are functionally more complex.
Proton-pumping rhodopsins.
The proton gradient across the cell membrane is the main contributor to the proton motive force, the primary source of energy for the cell, fueling processes such as flagellar motility or transport of solutes. One of the most important functions of this proton gradient is tied to membrane-bound ATPase, which generates ATP while dissipating the gradient. There are different ways to generate this gradient. In respiring cells, the protons are pumped out of the cell by electron transport chains. In fermenting cells, ATP is generally generated by substrate-level phosphorylation, and then the ATPase functions in reverse and consumes ATP to pump protons out of the cell. Microbial rhodopsins also contribute to generate this proton gradient by using light energy. The relative contribution of rhodopsins to the gradient varies depending on the particular microorganism and the growth conditions (see Experimental Assessment of Rhodopsin Photoheterotrophy in Heterologous Hosts and Native Aquatic Bacteria below). In any case, proton pumping is one of the most metabolically central functions of rhodopsins.
The electron flow in chlorophyll-based photosystems generates a proton motive force. It is remarkable that the retinal-driven proton-pumping systems achieve proton pumping with one single protein (for a discussion of how this proton gradient is used by bacteria, see Experimental Assessment of Rhodopsin Photoheterotrophy in Heterologous Hosts and Native Aquatic Bacteria below). It is thus tempting to think that proton pumping mediated by rhodopsins might have been the first mechanism that evolved to convert light energy into useful biochemical energy in early cellular life forms. A possible association with early life confers an additional interest to the study of the biochemistry, physiology, and ecology of rhodopsins.
Proton-pumping rhodopsins are found in both clusters A and B. In cluster A, the bacteriorhodopsins are found only in haloarchaea (Fig. 2). Bacteriorhodopsin from H. salinarum was in fact the first rhodopsin discovered (10) and remains the best studied with respect to its biochemistry and photobiology. In cluster B, proton-pumping rhodopsins have experienced a spectacular radiation. The largest cluster includes the proteorhodopsins that are widespread in aquatic environments. These rhodopsins are found in Proteobacteria, Bacteroidetes, and two clusters of marine Euryarchaeota. Proteorhodopsins are very well known due to the fact that they are the most widely distributed rhodopsins in the oceans, making them the most abundant rhodopsins on our planet (51). All proteorhodopsins studied so far primarily absorb visible light in the blue and green range. Most sequences group in two clusters, one with Proteobacteria and the other with Bacteroidetes sequences (Fig. 2).
The rhodopsins in Proteobacteria were the first to be discovered, and this is the origin of the name proteorhodopsin. The PR gene was found in a fosmid from an environmental metagenome that also had the small-subunit rRNA (SSU rRNA) gene of SAR86, an uncultured member of the Gammaproteobacteria (28) (Table 1). This early application of metagenomics, and downstream biochemical study, resulted in assigning a novel function to a member of the plankton that was known to be relatively abundant (through SSU rRNA gene surveys) but that was not in culture and so was unknown with respect to its metabolism. Since then, PR genes have been found in many other uncultured members of the Alpha- and Gammaproteobacteria (52–54) (Table 1) and also in cultured isolates from Alphaproteobacteria (55), Betaproteobacteria (56–58), and Gammaproteobacteria (59–61); some bacterial isolates, such as S. ruber M31T and N. marinus S1-08T, even have two or more different rhodopsins (Table 2). Of particular relevance was the discovery of the PR gene in members of the most abundant heterotrophic bacterial clade in the surface ocean, the SAR11 clade, represented by the isolate “Candidatus Pelagibacter ubique” HTCC1062 (55). The finding of PR in genetically tractable members of the genus Vibrio, i.e., Vibrio sp. AND4 and Vibrio campbellii BAA-1116, substantially extended the physiological repertoire of this genus, which otherwise was known for containing several notorious pathogens (35). Subsequent analyses of available genome sequences revealed the PR photosystem in additional Vibrio genomes, i.e., those of V. campbellii PEL22A (62) and V. splendidus ATCC 33789 and V. caribbeanicus BAA-2122 (63) (Table 2).
TABLE 1.
Rhodopsins from environmental samplesa
| Sampling site(s) | No. of rhodopsins | Taxon (clade) | Methodology | Rhodopsin | Reference |
|---|---|---|---|---|---|
| Monterey Bay, East Pacific | 1 | Gammaproteobacteria (SAR86) | Metagenomics, BACs | PR | 28 |
| Monterey Bay, East Pacific; HOT, Antarctica | 6 | Proteobacteria | PR gene clone sequencing | PR | 96 |
| East Mediterranean Sea; Red Sea | 30 | Proteobacteria | PR gene clone sequencing | PR | 94 |
| HOTS, Pacific; East Pacific; Antarctica | 5 | Alphaproteobacteria, Gammaproteobacteria | Metagenomics, genomic library, BACs | PR | 52 |
| Red Sea; Pacific | 2 | Gammaproteobacteria (SAR86) | Metagenomics, BACs | PR | 36 |
| HOTS, Pacific | 17 | Archaea (marine group II Euryarchaeota) | Metagenomics, fosmid library | PR | 69 |
| Gulf of Maine, Northwest Atlantic | 2 | Bacteroidetes | Cell sorting, SAGs | PR | 68 |
| HOTS, Pacific | 16 | Planctomycetales | Metagenomics, fosmid and BAC library sequencing | PR | 56 |
| Monterey Bay, East Pacific | 16 | Betaproteobacteria | Metagenomics, fosmid and BAC library sequencing | PR | 56 |
| Atlantic to Pacific | 2,674 | Multipleb | Metagenomics. sequencing | PR | 84 |
| Freshwater, Panama | 56 | Multiple | Metagenomics. sequencing | PR | 84 |
| North Atlantic | 91 | Multiple | PR gene clone sequencing, qPCR | PR | 92 |
| Freshwater (multiple) | 66 | Alphaproteobacteria (LD12), Bacteroidetes | PR gene clone sequencing | PR | 163 |
| Great Lakes, Canada | 69 | Actinobacteria | Metagenomics. sequencing | ActR | 164 |
| China Sea | 140 | Bacteroidetes | PR gene clone sequencing qPCR | PR | 104 |
| Chukchi and Beaufort Sea, Arctic Ocean | 54 | Multiple | PR gene clone sequencing, qPCR | PR | 105 |
| Lakes, Canada | 69 | Actinobacteria (acI, Luna) | ActR gene clone sequencing | ActR | 82 |
| North Sea | 117 | Alphaproteobacteria, Gammaproteobacteria, Bacteroidetes | PR gene clone sequencing, qPCR | PR | 93 |
| Antarctica, sea ice | 52 | Proteobacteria, Bacteroidetes | PR gene clone sequencing | PR | 165 |
| West Mediterranean Sea | 39 | Archaea | Metagenomics, sequencing, fosmid library | PR | 70 |
| Freshwater lakes | 119 | Actinobacteria (multiple) | Cell sorting, SAGs | ActR, PR | 81 |
| High mountain lake | 91 | Multiple | PR gene clone sequencing | PR | 166 |
| Puget Sound, Pacific | 1 | Archaea (marine group II Euryarchaeota) | Metagenomics, sequencing (genome assembly) | PR | 71 |
| Leaf surfaces, Israel | 156 | Multiple | Metagenomics, sequencing | Multiple | 167 |
| Northeast Pacific Ocean | 3 | Diatoms, haptophytes | (Meta)transcriptomics | XR | 154 |
| Gulf of Maine; Mediterranean; Pacific; Atlantic | 26 | Proteobacteria, Bacteroidetes, Archaea | Cell sorting, SAGs | PR | 54 |
| Red Sea | 12 | Bacteria, Archaea, virus | Metagenomics, sequencing | Multiple | 103 |
| Sargasso Sea | 782 | Proteobacteria, Bacteroidetes | Metagenomics, sequencing | PR | 64 |
| Beaufort Sea, Arctic Ocean | 187 | Proteobacteria, Bacteroidetes | PR gene clone sequencing, qPCR | PR | 106 |
| Arctic permafrost | 1 | Firmicutes (Exiguobacterium) | BR gene clone sequencing | BR | 72 |
Abbreviations: BACs, bacterial artificial chromosomes; qPCR, quantitative PCR; SAGs, single amplified genomes; PR, proteorhodopsin; ActR, actinorhodopsin; XR, xanthorhodopsin; BR, bacteriorhodopsin.
Bacteria from multiple classes or phyla.
TABLE 2.
Cultured microbial isolates with rhodopsins
| Strain(s); species or taxon | Taxon (clade) | Geographic origin | Rhodopsin typea | Reference |
|---|---|---|---|---|
| HTCC2207; SAR92 | Gammaproteobacteria | Oregon, East Pacific | PR | 53 |
| IMCC3088; MO60/NOR5 | Gammaproteobacteria | South Korea, Yellow Sea | PR | 60 |
| IMCC2047; Alteromonadales | Gammaproteobacteria | South Korea, Yellow Sea | PR | 61 |
| AND4; Vibrio sp. | Gammaproteobacteria | Andaman Sea | PR | 35 |
| BAA-1116; Vibrio campbellii | Gammaproteobacteria | Marine | PR | 142 |
| PEL22A; Vibrio campbellii | Gammaproteobacteria | Abrolhos Bank, South Atlantic | PR | 62 |
| BAA-2122; Vibrio caribbeanicus | Gammaproteobacteria | Curaçao, Caribbean Sea | PR | 63 |
| ATCC 33789; Vibrio splendidus | Gammaproteobacteria | Seawater, Hawaii, Pacific | PR | 63 |
| 10 strains; SAR11 | Alphaproteobacteria | Oregon, East Pacific | PR | 55 |
| 11 strains; SAR11 | Alphaproteobacteria | Oregon coast; BATSd | PR | 102 |
| IMCC1322; Puniceispirillum marinum | Alphaproteobacteria (SAR116) | South Korea, West Pacific | PR | 59 |
| HTCC2181; OM43 | Betaproteobacteria | Oregon, East Pacific | XR | 57 |
| MOLA814 | Betaproteobacteria | Beaufort Sea, Arctic Ocean | PR | 58 |
| MED134, MED152; Flavobacteriia | Bacteroidetes | West Mediterranean Sea | PR | 65 |
| 38 strains; Flavobacteriia | Bacteroidetes | Japan, West Pacific | PR | 67 |
| M31; Salinibacter ruber | Bacteroidetes | Solar saltern | XR, HR, 2 SR | 75 |
| DSW-6; Nonlabens dokdonensis | Bacteroidetes | South Korea, West Pacific | NaR, PR | 87 |
| NBRC 100814; Dokdonia eikasta | Bacteroidetes | West Pacific | NaR, PR | 40 |
| R-8282; Gillisia limnaeab | Bacteroidetes | Lake Fryxell, Antarctica | NaR, PR | 41 |
| PRO95; Dokdonia sp. | Bacteroidetes | North Sea | NaR, PR | 42, 168 |
| S1-08; Nonlabens marinus | Bacteroidetes | Northwestern Pacific Ocean | NaR, ClR, PR | 43 |
| 5 luna mixed cultures | Actinobacteria | Great Lakes, Canada | ActR | 82 |
| JL-3; Exiguobacterium sp. | Firmicutes | Freshwater lake | SRc | 74 |
| CCMP 1788; Oxyrrhis marina (and others) | Dinoflagellate (multiple) | Caribbean Sea | PR | 47 |
| UNC 1102; Pseudo-nitzschia granii | Diatoms | Indian Ocean | XR | 155 |
| Organic lake phycodnavirus | Phycodnaviridae | Antarctica | PR | 156 |
| Phaeocystis globosa virus (PGV) | Phycodnaviridae | Antarctica | PR | 156 |
PR, proteorhodopsin; XR, xanthorhodopsin; HR, halorhodopsin, SR, sensory rhodopsin; NaR, sodium-pumping rhodopsin; ClR, chloride-pumping rhodopsin.
The genome of the strain carries a third rhodopsin gene, likely encoding a truncated NaR containing only 120 amino acids.
Rhodopsin pumps protons when overexpressed in E. coli.
Bermuda Atlantic Time Series study site.
Venter et al. (64) suggested that Bacteroidetes also had PRs, based on the taxonomic affiliation of an RNA polymerase subunit gene on a PR-containing scaffold. This finding was subsequently confirmed by genome analyses of cultured Bacteroidetes large genome fragments or isolates (56, 65). Bacteroidetes in general, and members of the class Flavobacteriia in particular, frequently constitute an important fraction of marine bacterioplankton (66). Later, screening of a set of Flavobacteriia showed that 38 out of 75 isolates carried the PR gene (67) (Table 2). Further, flavobacterial PRs were found in two single amplified genomes retrieved from a marine environment (68).
Two subclusters of archaeal PRs are embedded within the bacterial PR cluster (Archaea clades A and B [Fig. 2]). Analysis of large insert DNA fosmid libraries made from microorganisms in the North Pacific Subtropical Gyre led to the discovery of PR genes in marine group II Euryarchaeota (69). These bacterial-like PRs were restricted to Euryarchaea inhabiting the photic zone, while other euryarchaeal lineages residing in deeper waters lacked PR genes. Ghai et al. (70) found that a few of the rhodopsins in the deep chlorophyll maximum belonged to Euryarchaeota. Analysis of metagenomes from Puget Sound (Pacific Ocean, coastal Washington), led Iverson et al. (71) to report that Euryarchaeota encode two distinct rhodopsins, which they defined as marine group II Euryarchaeota rhodopsin clade A and clade B (Fig. 2). The clade A Euryarchaeota PRs are phylogenetically related to PRs from Proteobacteria and correspond to the PRs originally reported by Frigaard et al. (69). Clade B Euryarchaeota PRs are distinct but were found also among PR sequences from the Global Ocean Sampling (GOS) sampling (71).
One small rhodopsin cluster consists of sequences related to the Gram-positive genus Exiguobacterium. E. sibiricum 255-15 was isolated from a 3-million-year old permafrost sample in Siberia. Its rhodopsin (ESR) is distantly related to marine rhodopsins and was able to function in E. coli, pumping protons upon illumination with green light (72, 73). In contrast, Exiguobacterium sp. strain JL-3 was isolated from Jinniu Lake, China. Characterization of its proteorhodopsin suggested that it might be a sensory rhodopsin despite a 90% similarity to the PR from E. sibiricum (74).
Another cluster includes the xanthorhodopsins. Xanthorhodopsin was initially found in S. ruber M31T (75). In addition to retinal, this proton pump is associated with a particular carotenoid molecule named salinixanthin (Fig. 1). This carotenoid expands the range of wavelengths that can be absorbed and thereby transfer energy to the retinal. The structure of this complex has attracted attention from crystallographers (76), largely due to interest in its unique accessory pigments. The S. ruber sequence is related to many other sequences from Planctomycetes, Proteobacteria, and a subcluster of the eukaryotic Alveolata, including diatoms such as Fragillariopsis and dinoflagellates such as Oxyrrhis, Alexandrium, and Polarella.
The large xanthorhodopsin-like cluster contains several poorly defined subclusters. One of them includes related sequences from several cyanobacteria. The Gloeobacter violaceus PCC 7421 gene has been cloned in E. coli, where it was expressed and could be complemented with both retinal and the salinixanthin from S. ruber M31T (77). This suggests that this clade of rhodopsins may function in a similar way to that of S. ruber. Moreover, similar to salinixanthin, the carotenoid echinenone that is naturally present in G. violaceus binds to the rhodopsin through the carotenoid 4-keto group and thereby also functions as a light-harvesting antenna (78). Another subcluster includes sequences from Roseiflexus RS 1 and several members of the genus Thermus. These bacteria share the same high-temperature environments, but they belong to different phyla (Chloroflexus and Deinococcus-Thermus, respectively).
Pyrocystis lunula, one of the dinoflagellates whose PR falls in the large xanthorhodopsin cluster, has been demonstrated to have a rhodopsin related to the proton-pumping type (79). In O. marina CCMP1788, two types of rhodopsins were detected in an expressed sequence tag (EST) library (47). One was related to the eukaryotic sequences in cluster A discussed above, including Cyanophora, Guillardia, and others. This included two different sequences, presumably representing two genes, that were scarcely represented in the EST library. The other group included over 40 different clusters, and some of them were the most abundant in the EST library. Based on the amino acid sequence, the rhodopsins in this group were assumed to act as proton pumps. Together with the closeness to xanthorhodopsins (Fig. 2), this suggests that the Alveolata rhodopsins in this cluster likely act as proton pumps. Slamovits et al. (47) suggested an interesting function for these proton-pumping PRs. If the proteins are in the membranes of food vacuoles, they could act to acidify the interior, a necessary condition for the food digestion in the vacuoles. Thus, they point out, a former phototroph (the dinoflagellate), which had lost the capacity for photosynthesis, had become a heterotroph thanks to a return to the use of light with PR. This idea could possibly be further extended to organelles in other organisms.
The sequences from Actinobacteria in freshwater environments have been named actinorhodopsins (ActR) (80–83). In view of the tree in Fig. 2, where the ActR are embedded in the xanthorhodopsin cluster and bootstrap values are low, a name for ActR different from xanthorhodopsin may be unwarranted. These ActR were first reported in the lakes sampled in the frame of the GOS expedition (84). Subsequent analyses extended the findings of ActR to many freshwater environments and to Actinobacteria in culture, suggesting that the globally abundant acI lineage may also carry ActR (82). Evidence for ActR in the acI cluster was finally provided through analysis of single amplified genomes in temperate lakes (81). The function of these ActR awaits experimental confirmation.
Chloride and sodium ion-pumping rhodopsins.
Other known ion-pumping rhodopsins include those that pump anions inward or cations outward across the cell membrane. It has been experimentally shown that a proton-pumping bacteriorhodopsin can be converted into a chloride pump by changing a single amino acid (85). Thus, the conversion of proton pumps into chloride ion pumps in some cases could lead to relatively rapid functional transition in evolution.
Chloride ion pumps are found in Bacteria and Archaea living in hypersaline systems. These rhodopsins have arisen several times over the course of evolution, with those found in marine bacteria (43) being structurally and evolutionarily distinct from those found in haloarchaea in high-salinity habitats. Their abundance in the environment remains largely unexplored. Since sodium and chloride are the two most abundant ions in marine environments, having such pumps may be helpful in maintaining osmotic balance, as well as in generating energy via a chloride or sodium chemiosmotic potential.
Cluster A includes two rather different types of halogen pumps. Halorhodopsins are exclusively archaeal, although the bacterium S. ruber M31T has a homologous gene. These were the first chloride ion pumps discovered, and their structure and function are very well characterized (86). A second type is the channelrhodopsins found in the eukaryal group Chlorophyta and Cryptophyta. These rhodopsins act as light-gated ion channels. In proton-pumping rhodopsins, the transfer of protons is a two-step process. The proton taken up on the inside of the cell membrane is not the proton released outside the cell. The two parts are physically separated, likely to avoid back flow. Channelrhodopsins, on the other hand, consist of a channel that allows ions to move through when opened in response to the appropriate light signal. Thus, not only protons and chloride but also sodium, potassium, lithium, and calcium can be transferred from one side of the membrane to the other. Channelrhodopsins have an additional protein domain attached to the carboxyl end of the opsin in the cytoplasmic part of the membrane (22).
Cluster B includes a substantial number of chloride and sodium ion pumps. The currently known sequences form two statistically well supported sequence clusters (Fig. 2), one containing rhodopsins pumping sodium and another containing rhodopsins pumping chloride. The sodium pumps have been found in Bacteroidetes, as well as in members of the Deinococcus-Thermus phylum isolated from very different environments (40, 87, 88). Interestingly, sodium pumps have been shown to have dual functionality, transporting protons and halogens depending on the chemical environment (40, 50, 89). It will be exciting to discern how such variability in function could influence the physiology of bacteria in their natural environment. The chloride pumps have so far been found in marine Alphaproteobacteria and in one member of Bacteroidetes (43).
Sensory rhodopsins.
Sensory rhodopsins are found only in cluster A (Fig. 1). The simplest type is found in Archaea, and at least three slightly different types have been defined. Type I sensory rhodopsins (SR I) are used by Archaea to move toward the light and have been found in haloarchaea and S. ruber (90). Type II sensory rhodopsins (SR II), in contrast, are used to move toward the dark and have thus been named phoborhodopsins; these have been found only in haloarchaea (90). A third type of sensory rhodopsins (subsequently termed SR III) was first proposed based on study of rhodopsins in Haloarcula marismortui ATCC 43049 (19).
The photoprotein of Anabaena strain PCC7120 was the first sensory rhodopsin described in Bacteria (91). This peptide fits within a larger group that includes a number of cyanobacteria. Thus, the proteins in the cyanobacteria Cyanothece, Chroococcidiopsis, Gloeocapsa, Leptolyngbya, and Chamaesiphon might also be sensory rhodopsins (labeled “Cyanobacteria sensory rhodopsin” in Fig. 2). Interestingly, the protein in Anabaena is associated with a soluble protein that likely transduces the signal from the rhodopsin sensor. This is different from the archaeal sensory rhodopsins discussed above, where the signal is transduced to another membrane-bound protein.
Eukarya have another type of sensory rhodopsin that is also localized within cluster A. These rhodopsins are the most complex within type I rhodopsins, since the peptides accommodate, in addition to the conserved rhodopsin domain, a histidine kinase and a response regulator domain. In effect, the structure of these genes is the classical structure of two-component systems triggered by light. These rhodopsins have been referred to as “enzymerhodopsins” and have been found in cyanobacteria and some algae (2). One consistent clade according to the bootstrap value includes enzymerhodopsins from different prasinophytes, i.e., Micromonas, Bathycoccus, and Ostreococcus, three of the most abundant algal genera in coastal marine waters. A clade with low support includes sequences from the freshwater green alga Chlamydomonas and the marine cryptophyte Guillardia. The cyanobacterial enzymerhodopsin is phylogenetically very different from those of eukaryotic algae. Notice also that the C. reinhardtii sequence with accession number EDO99289 is also a His kinase and does not cluster with the other enzymerhodopsins (Fig. 2).
ENVIRONMENTAL DISTRIBUTION OF BACTERIAL RHODOPSINS
Following the initial discovery of a rhodopsin gene in a SAR86 genomic fragment recovered off the coast of California (28), the exploration of rhodopsin gene diversity in a variety of marine habitats accelerated (Table 1). A large number of studies reported on the presence and diversity of rhodopsins in seawater from the Northern Hemisphere, while fewer studies were conducted in waters south of the equator. Quantitative estimates of the proportion of bacteria in surface waters carrying rhodopsins vary broadly, from 15% to over 70% (84, 92–94), compared to that of bacteria carrying genes in pathways associated with aerobic anoxygenic photosynthesis (AAP) in bacteriochlorophyll-containing cells which are in up to 30% of planktonic bacteria (95). In a recent survey of published microbial metagenome studies, Finkel et al. (51) estimated that on average over 60% of the microbial cells in the 0.1- to 0.8-μm size fraction (as determined by pore size of filters to collect microorganisms) carry rhodopsin genes, while around 20% carry genes for the AAP photosynthetic reaction centers. At present, it is clear that major portions of marine bacteria in sunlit environments carry rhodopsin genes. Still, variability in precise proportions between studies can likely be attributed both to differences in methodologies for quantification and to actual differences driven by environmental factors.
Biogeography of Rhodopsin Spectral Tuning
Given that rhodopsins function in part as light energy-absorbing molecular entities, it seems logical that spectral tuning of rhodopsins to the light fields (i.e., wavelength and light intensity distribution) prevailing in the natural environment might play an important role in their ecological distribution (Fig. 3). Early functional surveys of proteorhodopsin genes showed that the SAR86 PR had an absorbance maximum in the green light range (520 nm), coinciding with the available photosynthetically active radiation (PAR) in oceanic surface water (28). Soon thereafter, Béjà et al. (96) described a divergence of PRs with absorption maxima in the range from blue (around 490 nm) to green (around 540 nm) light and deduced that these PRs represent spectral tuning to the dominant light field in deeper and shallow oceanic waters, respectively (Fig. 3). These findings sparked research into determining the genetic and biochemical basis for spectral tuning, which in turn provided a basis for interpreting ecological patterns of distributions of distinct PRs. Accordingly, site-directed mutagenesis demonstrated the primary role of PR amino acid 105 in the retinal binding pocket for spectral tuning, so that, e.g., Leu gives green- and Gln gives blue-tuned PRs, although other amino acids at this position can also provide similar tuning effects (97). In Flavobacteriia, for example, PRs primarily contain a hydrophobic side chain amino acid (Met) at this position instead of the “canonical” Leu, which also results in an absorbance maximum in the green light (535 nm) (65). Amino acid residues at other positions were subsequently shown to contribute to spectral fine-tuning (98–100).
FIG 3.
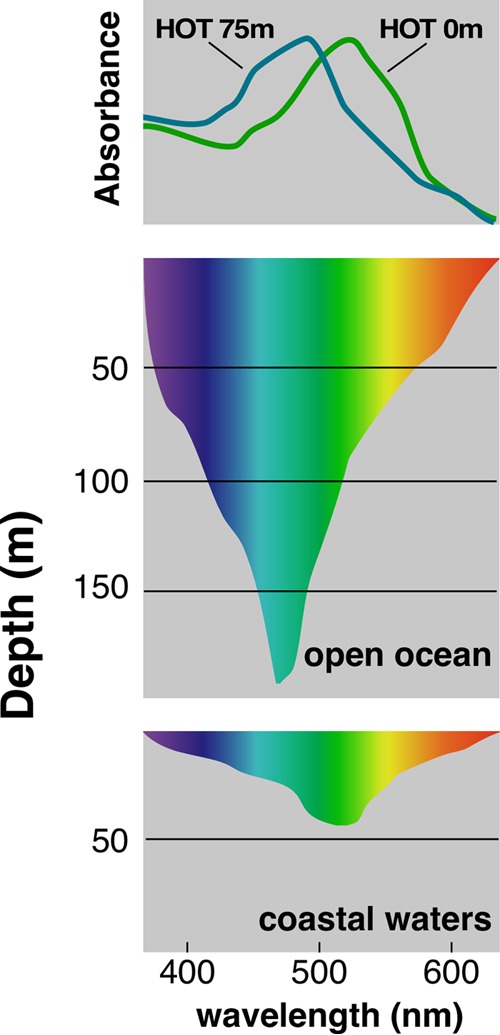
Absorption spectra of PRs in relation to light attenuation patterns in different marine environments. Top panel, spectra of green-tuned PR (GPR) and blue-tuned PR (BPR) collected from different depths at the Hawaii Ocean time series (HOT) station ALOHA. Absorption spectra were obtained following heterologous expression in E. coli. Middle and lower panels, typical light attenuation in open ocean and coastal environments, respectively. (Inspired by reference 169.)
Sabehi et al. (101) elaborated on the original suggestion of distinct depth distributions of differentially tuned rhodopsins. They found that in the Eastern Mediterranean Sea, under stratified summer conditions, green-tuned rhodopsins were dominant in the surface and blue-tuned rhodopsins greatly dominated at the 55-m depth. However, during winter, the distributions of the two tuning types were relatively similar throughout the mixed layer (with roughly one-third of rhodopsins being green-tuned). In the Sargasso Sea, in contrast, nearly all rhodopsins consisted of blue-tuned variants irrespective of season and depth (101). Further insight into the spatial distribution of rhodopsin tuning variants was provided by the GOS expedition, where Rusch et al. (84) investigated the diversity and spatial distribution of rhodopsins across a transect from the northwest Atlantic to the Pacific. Among the total of 1,874 rhodopsin sequences encoding the primary tuning residue, green-tuned rhodopsins were relatively more abundant in temperate coastal waters, whereas blue-tuned rhodopsins dominated in warmer open ocean water (84). They thus suggested that, analogous to the light depth distribution (96), the spatial distribution of rhodopsin tuning variants might reflect changing spectral properties of the light along the coastal-open ocean transition (Fig. 3). Consistent with these findings, PCR screening showed that PRs in SAR11 isolates from the Oregon coast were either green- or blue-tuned, while all isolates from the Sargasso Sea had blue-tuned rhodopsins (102). In comparison, among a total of 117 PRs from the North Sea affiliated with Proteobacteria and Bacteroidetes, nearly all PRs (97%) were green-tuned (93), while a majority (71%) of bacterial and archaeal rhodopsins in the Red Sea surface waters were putatively blue-tuned (103).
Rhodopsin Diversity and Distributional Surveys
In the first large survey of rhodopsins in DNA from marine prokaryotes, using PCR with degenerate PR primers applied to Mediterranean and Red Sea samples, Sabehi et al. (94) uncovered a large phylogenetic diversity of rhodopsins, with some rhodopsin clades (groups) consisting of sequences from both seas and others apparently found primarily in one of the seas. Curiously, at this time, it remained unknown if distinct PR variants represented “functionally divergent paralogous genes, derived from the same or similar species, or orthologous proteorhodopsins that are distributed among divergent planktonic microbial taxa” (94). This issue was soon after resolved by findings that in most (but not all) cases, divergent PRs belonged to different microbial taxa (Tables 1 and 2) (interestingly, marine prokaryotes containing several rhodopsins have now been found, but in these cases the rhodopsins are mostly distinct; see “Rhodopsin Types” above). Thus, the first report on metagenome shotgun sequencing of microbial DNA from the Sargasso Sea substantially extended the range of phylogenetic diversity of PRs that can be found at a single geographic location, by a claimed study of 782 rhodopsin genes, of which several were distant from known PRs or other rhodopsins (64). The actual number of new rhodopsins found is uncertain, however, since partial gene sequences mapped onto a scaffold phylogeny were used to estimate PR diversity. It is probable that different nodes in their phylogenetic tree were actually derived from different portions of identical molecules, hence overinflating their diversity claims. In the expanded GOS expedition data set, Rusch et al. (84) found that in several SAR11 PR clades, similar PR sequences were found in the Atlantic and Pacific Oceans (see also “Biogeography of Rhodopsin Spectral Tuning” above).
To obtain further knowledge on the spatial structuring of rhodopsins, Campbell et al. (92) investigated the distribution of PR genes in the North Atlantic Ocean. They first characterized PR gene diversity in a sample from the Sargasso Sea using clone library construction from PCR products obtained by degenerate PR gene primers. Among the total of 27 PR gene clusters recorded, quantitative PCR (qPCR) analysis of four distinct clusters of PRs (three alphaproteobacterial, of which SAR11 and HOT2C01 PRs were particularly abundant, and one flavobacterial) indicated that roughly 50% of bacteria in the Sargasso Sea contained these PRs. However, these PRs were much less abundant further northeast in the Atlantic. Interestingly, there was an overall negative correlation between PR gene abundance and chlorophyll a concentrations (but not light) in the surface samples and also in the depth profiles (92). In the latter there was also a negative correlation between PR genes and inorganic nutrients. Still, some variability in the relationship between these environmental variables and the four distinct PR types was observed. Using a similar approach, but focusing on flavobacterial PRs, Zhao et al. (104) recorded that PRs were structured essentially according to clone library sampling site in the eastern and southern China Sea. Moreover, qPCR showed a higher abundance of these PR genes in offshore waters than in coastal waters (overall, these flavobacterial PRs were represented in up to 1.6% of total bacteria [104], which is similar to the up to 3.0% found with primers specific for PRs from an uncultured flavobacterial group in the Sargasso Sea [92]).
Distinct spatial distributions of PRs have also been found in PR gene clone libraries from the Arctic compared to samples from lower latitudes (105), and Arctic PRs were also different from PRs previously reported from Antarctica by Béjà et al. (96). Moreover, qPCR analyses showed that from summer to winter, the abundance of PR gene-containing bacteria largely changed in relation to total bacterial abundance, and the proportion of photoheterotrophs (PR- and bacteriochlorophyll-containing bacteria alike) remained stable across these seasons, despite extreme differences in day length (105). A relatively stable abundance of PR genes during dark winter months in the Arctic was confirmed in the Amundsen Gulf of the southeastern Beaufort Sea (106). Still, analysis of PR gene clone libraries showed that PR diversity changed from early to late winter (with notable changes in PR genes affiliated with Bacteroidetes and Alphaproteobacteria, while gammaproteobacterial PRs were consistently abundant).
Taken together, so far, few studies have comprehensively investigated spatial variability in PR diversity and prevalence distributions. It will be intriguing to learn about the biogeography of rhodopsins from the summed global oceanic GOS, Tara Oceans, and Malaspina 2010 expeditions. Even fewer studies have investigated temporal dynamics in PR gene distributions (see, e.g., references 101, 105, 106, and 107). An important task for the future is therefore to determine dynamics of rhodopsin-containing bacterial populations in relation to rhodopsin gene expression patterns over different ranges of spatiotemporal resolution. An intriguing question that remains to be answered is whether expression of rhodopsin genes might provide sufficient ecological benefits that in turn determine the seasonal dynamics and activity of particular bacterial populations.
Environmental Rhodopsin Gene Expression Analyses
Analyses of expression of rhodopsins have been carried out in a quite limited number of marine environments. Yet, the studies done so far show that rhodopsins are widely expressed in both coastal and open ocean waters. After the original finding of the proteorhodopsin gene in the ocean (28), analysis of membrane preparations from Monterey Bay surface water using spectroscopic analysis revealed photochemical activity consistent with an abundance of expressed and functional PR protein in native marine bacterioplankton (96). Further north along the Pacific coast of the United States, expressed PR protein was detected by mass spectrometry (MS) in seawater samples off Oregon, with mass spectra similar to those of PRs in cultured SAR11 bacteria (55). Pioneering metatranscriptomic analyses from the Hawaiian Ocean time-series station ALOHA in the North Pacific Subtropical Gyre showed PR to be among the highly expressed gene products (108, 109). Subsequent extended metatranscriptomic analyses at the ALOHA station showed that PR genes were highly expressed in the photic zone (25- to 125-m depth) but not in the mesopelagic at 500 m (110). Notably, rhodopsin genes were found at up to nearly two orders of magnitude higher relative abundances in the transcript pool than in the gene pool. A majority of transcripts belonged to the SAR11 clade (around 60%), with contributions up to 10% also of Bacteroidetes, SAR86, and euryarchaeal PRs (see Fig. S5 in reference 110). In particular among “Ca. Pelagibacter”-related populations, PR genes were among the most highly expressed transcripts. Expression of SAR11 and SAR86 clade PR genes has also been detected in the Red Sea (103).
Further detail on PR gene expression levels was recently provided by Satinsky et al. (111), who carried out a quantitative inventory of microbial gene expression patterns at an outer continental shelf station in Amazon River Plume waters where a phytoplankton bloom was occurring. The PR gene accounted for ∼1% of total transcripts and was the most abundantly expressed of all genes defined as biogeochemically relevant, in both the free-living and particle-attached fractions of the prokaryote community (∼2 × 109 transcripts liter−1). Interestingly, transcription in the latter fraction, where Flavobacteriia and SAR116 clade bacteria were dominant, was around 3-fold higher on a per-gene basis; SAR11 clade bacteria had a similar per-gene expression ratio in the free-living and particle-attached fractions.
Given that phytoplankton gene expression regulation is linked to diurnal variations in light and darkness, the influence of such variations on rhodopsin-associated metabolisms has attracted some attention. Comparisons of gene expression patterns in day-night samples collected at the station ALOHA showed a relatively higher abundance of PR transcripts during the day (109). Similarly, in Delaware coastal waters an elevated abundance of PR genes belonging to SAR11 clade bacteria and Flavobacteriia was observed during daylight hours (112). A follow-up study in the same waters confirmed that SAR11 PR gene expression is higher (∼2.5-fold) during the day at different seasons (113). Using metatranscriptomic analysis on samples collected in southeast U.S. coastal waters, it was recorded that, in general, PR genes are abundantly expressed by a multitude of taxa (114). Surprisingly, though, a subsequent study in the same waters showed significant differences in PR gene expression between day and night samples for only 2 out of 27 taxa carrying the PR gene (115).
Comprehensive monitoring of diurnal changes in microbial transcript abundance over 48 h in the Pacific off Monterey Bay revealed pronounced temporal variability in a multitude of genes involved in different metabolic pathways in the PR-containing lineages SAR11, SAR86, and marine group II Archaea. In contrast to genes belonging to phytoplankton, the overall changes in gene expression levels in these bacteria were not directly linked to light intensity or time of day (116). Nevertheless, detailed analyses of the same data set showed pronounced peaks in expression of the glyoxylate shunt gene encoding isocitrate lyase in the tricarboxylic acid (TCA) cycle coinciding with highest solar irradiance, which were suggested to be tightly linked to PR phototrophy (117). Even more pronounced diurnal dynamics in isocitrate lyase gene expression in a wide variety of Proteobacteria clades (e.g., SAR11 and SAR86) were then found using automated sampling in both California coastal waters and the North Pacific Subtropical Gyre (118). Using the same automated sampler, with sampling extended over 3 days at station ALOHA, pronounced diurnal cycles in gene expression patterns of both photoautotrophs and photoheterotrophs (including both PR- and bacteriochlorophyll-containing bacteria) were observed (119). Notably, significant diurnal changes in PR expression were recorded for SAR11, SAR116, SAR86, and SAR324 clade bacteria, with peaks in expression levels near dawn.
There are as yet no analyses of rhodopsin gene expression in marine bacterioplankton across broad seasonal/annual scales. The one exception so far is work in the Arctic, showing a relatively stable presence of PRs in the gene pool, which was in stark contrast to very pronounced changes in PR gene expression levels (106). Accordingly, expression of the investigated Alpha- and Gammaproteobacteria and Bacteroidetes PR types decreased from December to very low levels in winter (i.e., from January to March, the period dominated by darkness), followed by peaks in expression in spring (early April and June) (106).
Interestingly, experiments and expression analyses on surface ocean bacterioplankton have suggested links between the proton motive force created by PR phototrophy and the stimulation of nutrient uptake through various membrane transporters. Conducting tracer experiments in the Atlantic, Mary et al. (120) suggested that an observed 23% increase in amino acid uptake in the light among low-nucleic-acid cells sorted by flow cytometry (60% of which were SAR11 cells) could be attributed largely to PR phototrophy in these bacteria (in contrast, high-nucleic-acid cells showed no light-stimulated uptake). In a subsequent study in the Atlantic, Gómez-Pereira et al. (121) confirmed that the uptake of radiolabeled ATP (either the phosphate or adenosine moieties) as well as methionine by SAR11 bacteria (and Prochlorococcus) was stimulated by around 30% in the light compared to darkness.
The exact molecular mechanisms underlying such light-induced substrate uptake by SAR11 clade bacteria or other PR photoheterotrophs remain unknown. Yet, Morris et al. (122) proposed a link between PR and nutrient uptake membrane transporters (in their case TonB transporters, which utilize the proton motive force), based on observing concomitantly elevated expression levels of these membrane proteins in South Atlantic surface waters. Dominance of membrane transport proteins that rely on ATP or directly on the proton gradient across the membrane (i.e., ATP-binding cassette [ABC] and tripartite ATP-independent periplasmic [TRAP] transporters) has also been found in proteomes of marine bacterioplankton from Pacific coastal waters off Oregon (123). Incidentally, at the Pacific time-series station ALOHA, “Ca. Pelagibacter” strain HTCC7211 PR genes were abundantly expressed along with high relative levels of genes encoding components of the C4-dicarboxylate transport (Dct) system. These secondary membrane transporters are directly dependent on the electrochemical proton gradient, rather than ATP, for transporting carbon compounds like mannitol and malate, indicating a potential link between PR proton pumping and nutrient transport into the cell (110). Such links have been experimentally shown in starving cells of “Ca. Pelagibacter ubique” HTCC1062, where taurine uptake was 65% higher in the light than in darkness (124).
GENOMICS OF MARINE BACTERIAL AND ARCHAEAL RHODOPSINS
In the marine environment, rhodopsin genes are distributed in a wide variety of microbial taxa and habitats. As already mentioned, the main branching in the rhodopsin tree seems coincidental with deep branching in the rRNA tree: Bacteria form the deepest root, with Archaea and Eukarya forming a subsequent bifurcation. Moreover, many rhodopsin genes fall within taxonomically coherent clusters. While most microorganisms have inherited rhodopsin genes from their ancestors, there are substantial mismatches between the phylogenetic assignments of the rhodopsin compared to the SSU rRNA genes for several microorganisms. This suggests that lateral gene transfer of rhodopsin genes is a fairly frequent process. In this section we examine the acquisition of rhodopsin genes by LGT, consider the consequences of incorporating this gene in different genomic backgrounds, and finally explore how its expression might be regulated.
Lateral Transfer of Rhodopsin Genes
An important consequence of the genetic simplicity of the rhodopsin photosystem is the relative ease with which lateral gene transfer might occur (38, 56, 69, 125). Thus, inconsistencies between the phylogeny of taxonomic markers, such as the SSU rRNA gene, and the rhodopsin and carotenoid genes are frequently observed (36, 56). A particularly striking example is that of S. ruber M31T (45). This bacterium belongs to a group close to the basal branching of the Bacteroidetes and lives exclusively in hypersaline aquatic environments, particularly in crystallizer ponds of solar salterns. S. ruber shares this environment with the haloarchaeon Haloquadratum walsbyi. They are both found in very large concentrations, sometimes reaching 108 cells ml−1. S. ruber has four rhodopsin genes, encoding xanthorhodopsin (a proton pump), halorhodopsin (a chloride pump), and two sensory rhodopsins (45). Except for the xanthorhodopsin gene, the other three genes cluster together with homologous genes from haloarchaea, which is parsimoniously explained by LGT between these two coexisting microorganisms (45). There are many other examples of transfers across domains. For example the PR genes from group II Euryarchaeota (Archaea clades A and B) are highly similar to PR genes from Proteobacteria but only distantly related to known archaeal PR genes, indicative of LGT from Bacteria to Archaea (69).
LGT has also occurred between phyla. For example, there are Gammaproteobacteria genomes carrying PR genes clustering with Actinobacteria (81). Further, there is differentiation within lower-level taxa, such as the Gammaproteobacteria genera Vibrio and Glaciecola, where some members have PRs clustering with Alphaproteobacteria and others have PRs clustering with Gammaproteobacteria (63). Curiously, though, Bacteroidetes PRs cluster together and are consistently distinct from PRs in other taxa, such as Proteobacteria (Fig. 2). Thus, PR genes appear to have moved within Proteobacteria (e.g., between Alpha- and Gammaproteobacteria and maybe from Proteobacteria to other distant taxa) but not to or from Bacteroidetes.
LGT events can also be detected among closely related genera. For example, among Bacteroidetes, the PR in Dokdonia sp. strain MED134 is closer to the PR in Polaribacter sp. strain MED152 than to the PR in the more closely related Dokdonia sp. strain PRO95 (126). In fact, within the Bacteroidetes family Flavobacteriaceae, several studies report that the PR phylogeny is not in all cases congruent with their SSU rRNA gene phylogeny, indicating past LGT events (42, 67, 126). These and other instances show that LGT of rhodopsin genes has occurred relatively frequently between both closely and distantly related taxa. It should be noted that the potential transfer frequency between closely related taxa is harder to distinguish, and thus it may be more frequent than realized so far.
LGT can also be observed in natural assemblages. Thus, an analysis of a set of 119 single amplified genomes (SAGs) from freshwater bacteria containing both PR and SSU rRNA genes showed that in a large majority of the SAGs the phylogeny of the PR gene was congruent with that of the SSU rRNA gene, while in around 5% of the SAGs it was not, implying that in the latter case they were acquired through LGT (81). Interestingly, this analysis of SAGs also resulted in the first evidence for recombination between ActR and PR genes, resulting in composite rhodopsins in Deltaproteobacteria and Betaproteobacteria SAGs, giving another twist to the occurrence of LGT of rhodopsin genes.
An important consideration in relation to the often-observed indications for LGT of PR is that this system requires the presence of the carotenoid synthesis genes. Otherwise, retinal will not be made and the PR will not be functional. In some bacteria, the genes for beta-carotene synthesis are already present (for example, in many Bacteroidetes genomes). In these genomes, the PR gene is flanked only by blh (for cleaving beta-carotene to retinal) (Fig. 4), and as a consequence, the transfer of these blh-prd tandem genes among bacteria with the ability for beta-carotene synthesis should be relatively straightforward (56, 126).
FIG 4.

Arrangement of rhodopsin and carotenoid synthesis genes in different bacteria. (A) Carotenoid synthesis genes (crt) and the gene for synthesizing the chromophore retinal (blh) are in red. Rhodopsin variants are shown in green. In blue are genes whose products are known to respond to light. tRNA in black might be involved in lateral gene transfer. Labels above genes denote gene products. (B) Gene arrangement in the Flavobacteriia single amplified genome (SAG) MS024-2A; the PR is next to the conserved cluster of highly expressed genes that includes ribosomal protein, RNA polymerase subunit alpha, and transcription factors genes, in addition to Sec secretion pathway and TCA cycle genes.
In other cases, the carotenoid synthesis genes must be transferred with the blh-prd tandem. An excellent example is that of the family Vibrionaceae (63). Most genomes in the family do not have carotenoid synthesis genes or PR. The Vibrio genomes with PR, however, have a gene cluster consisting of blh, the four crtEBIY genes, and prd (Fig. 4). Importantly, PR is present only in vibrios that also have the idi gene somewhere in the genome; idi encodes an enzyme involved in the synthesis of carotenoid precursors (either a type 1 or a type 2 isopentenyl-diphosphate delta-isomerase). Curiously, neither of the pathogens Vibrio cholerae and V. splendidus carries the idi gene, except the one strain, V. splendidus ATCC 33789, that also has the prd-blh gene cassette, and in this strain the idi gene is positioned immediately upstream from the PR gene, suggesting that it may have been transferred along with the prd-blh cassette (63). The association of the prd-crtEIBY-blh gene cluster with idi was also observed in genome fragments from the ocean (37, 56). In effect, one-third of the environmental genome fragments analyzed by McCarren and DeLong (56) carried the PR gene linked with the necessary set of crt genes. This arrangement was found in both Proteobacteria and Planctomycetes but not in planktonic Euryarchaea (56, 69). Thus, in many bacteria these six genes make up a “gene cassette” that can confer phototrophic potential to new hosts with relative ease if transferred laterally (Fig. 4). This indicates that the gene arrangement next to the rhodopsin gene and the gene set in the new host would determine the likelihood for successful outcomes of LGT events. This further suggests that LGT is possible even when the receptor cell does not have the carotenoid synthesis genes. Thus, the general genome architecture is highly likely to have implications for the extent of LGT of rhodopsin phototrophy between different bacteria.
Extraordinary verification of the potential ease of transfer between organisms came from experiments in which genome fragments containing PR gene cassettes were recombined into the standard laboratory model heterotrophic bacterium E. coli, where they were expressed to yield photosystems that were functional in the receiving host (127). An intriguing but yet-unresolved issue in the context of lateral gene transfer is how the genes for PR phototrophy, once acquired, are incorporated into the metabolic and genetic regulatory network so that adequate expression and physiological integration in relation to relevant environmental conditions are obtained.
Regulation of Rhodopsin Gene Expression
The benefits provided by expressing rhodopsin genes likely depend on the level of sophistication of the regulation of their expression. The genetic and genomic elements in different bacteria that contribute to the regulation of rhodopsin gene expression, or rhodopsin photoheterotrophy in general, remain largely unexplored but presumably involve a multitude of layers.
Genome context.
The role of genome architecture and gene location is increasingly recognized as an important factor in regulation of cell metabolism in eukaryotes and, more recently, also in prokaryotes (128). In microorganisms, the PR gene is found in very different phylogenetic and physiological contexts. However, there are some examples where a notable degree of synteny exists. Thus, in several genomes, the PR gene is in the vicinity of one of the longest conserved gene clusters in the bacterial world. Since this gene cluster contains genes that are part of the translational machinery, including a number of ribosomal protein genes and the gene for the alpha subunit of RNA polymerase, it is highly expressed in actively growing bacteria (129). Highly active genes tend to be in loops that extend into the cytoplasm closer to enzymes, transcription factors, and ribosomes (128). It is known that the cytoplasm of all organisms from all three domains of life is not homogenous (130). Such is also the case of the expression of highly active genes, which occurs in subcellular locations referred to as “transcription factories” (131). In these locations, RNA polymerases gather along with transcription factors and the genes that are being expressed. Therefore, the location of the rhodopsin gene close to such clusters likely contributes to expressing the rhodopsin gene at adequate levels in relation to the most actively expressed genes.
Genomes with the PR in this genetic context include the following: the environmental DNA fragments HOT2C01 (Alphaproteobacteria) and EBAC31A08 (Gammaproteobacteria), where the PR gene is only 12 genes away from the cluster (52); the flavobacterial SAGs MS024-2A and MS024-3C, which are abundant in surface ocean samples (132), where the gene is right next to the cluster (Fig. 4B); and some flavobacterial isolates such as Leeuwenhoekiella strain Hel-I-48 (eight genes away) and Mesoflavibacter zeaxanthinifaciens S86 (seven genes away). In other bacteria, the distance is a bit longer, such as in Polaribacter isolates and SAGs, for example, where the PR gene is about 35 genes away, and in S. ruber, in which it is around 25 genes away. Therefore, placement of the PR gene in this location seems to be fairly common in environmentally abundant bacteria.
Other locations are also conserved in several bacteria. For example, in the genus Dokdonia (Flavobacteriaceae), the rhodopsin gene is next to a genomic island that is flanked by a gene with a conserved BLUF domain, whose product detects blue light and therefore might be a light regulator. There is also a tRNA, which is usually the flanking gene of genomic islands (126).
A comparison of assemblies of SAR11 metagenomic fragments with the genome of “Ca. Pelagibacter ubique” HTCC1062 showed that of the 10 genes surrounding the PR gene, eight consistently conserved synteny, while two genes from HTCC1062 were consistently missing from the metagenome. These results show that there is a certain conservation of the location of PR in SAR11 from the Pacific Ocean (133). The consistency of this and other cases of synteny needs to be determined by examining more examples, but we foresee that analyses of gene placement for the regulation of the rhodopsin phototrophy is a promising avenue of research.
Promoters, light sensors, and transcription factors.
Another, more fine-tuned mode of regulation occurs more locally at the level of individual prd genes. González et al. (126) found a strong promoter sequence in the upstream region of the PR gene in Dokdonia sp. MED134. This sequence is recognized by sigma factor RpoD (σ70), which is the main sigma factor initiating the transcription of housekeeping genes in most bacteria, including all Bacteroidetes. A strong promoter sequence upstream of the PR gene would ensure relatively high levels and adequate timing of the PR gene expression. Two pieces of evidence are consistent with a regulation by a strong promoter. First, the PR gene in Dokdonia sp. MED134 was one of the relatively few genes in this bacterium predicted to be highly expressed based on codon bias (126), a prediction shared with the usual highly expressed genes involved in ribosome synthesis, for example. Second, a high expression of PR in Dokdonia sp. MED134 was confirmed experimentally (117), which also showed its expression depends on the growth phase (117).
Microbial cells have several proteins with light-sensing domains, such as BLUF, PAS, EAL, GAF, GGDEF, phytochromes, or cryptochromes, that play different roles in the regulation of metabolism and growth. It would be interesting to ask whether microbes with rhodopsins have more or fewer such genes than bacteria without rhodopsins. Analysis of the genomes of Polaribacter sp. MED152 and Dokdonia sp. MED134 showed that these PR-containing bacteria carry a considerable number of light-sensing genes with a variety of architectures (134); such an abundance of light sensing genes was not observed in Alpha- or Gammaproteobacteria.
Almost nothing is known about other possible regulation mechanisms of rhodopsin photoheterotrophy, e.g., two-component systems or riboswitches. One intriguing possibility is that small RNAs may be involved, since Kimura et al. (135) found that some small RNAs were highly expressed in the light together with the PR genes of Dokdonia MED134.
EXPERIMENTAL ASSESSMENT OF RHODOPSIN PHOTOHETEROTROPHY IN HETEROLOGOUS HOSTS AND NATIVE AQUATIC BACTERIA
The first experimental work on PR-containing cultured marine bacterial members of the SAR11 and SAR92 clades did not identify significant phenotypic properties that might be expected for rhodopsin-containing bacteria grown in light, such as elevated growth rates or yields (53, 55). Notably, these organisms were grown in seawater, so their specific carbon sources, or other factors that might limit their growth, were unknown at the time. The simplistic perspective that higher growth rates or yields are the only physiological or ecological advantages provided by light-driven energy generation raised doubts about the ecological relevance of PR phototrophy in marine bacteria (136, 137). Currently, new knowledge is accumulating on how light energy harvested by PR is utilized, resulting from ecophysiological response experiments with different bacteria. Thus, a picture is emerging where the benefit of PR phototrophy can be realized through a multitude of physiological and ecological strategies. This is not so surprising, given the great diversity of phylogenetic, genomic, and physiological backgrounds in which PR is found. This also, however, poses challenges to broad generalizations or oversimplifications regarding the significance of rhodopsins for the global energy budgets. Current understanding of the physiological strategies and ecological roles of rhodopsins among different bacteria can be further advanced by defining the potential principal ecophysiological strategies for how rhodopsin phototrophy is utilized: (i) for promoting survival during conditions of starvation, (ii) for stimulating growth (note that these first two are principal components of ecological fitness), and (iii) for promoting general physiological efficiencies that might provide less readily detected fitness benefits, for example, by enhancing synergistic community interactions. It should be recognized that these are potential extremes on a continuum of life history strategies within which PR phototrophy might benefit bacteria, and it would be no surprise if the details in the benefits of PR phototrophy and its regulation were as diverse as the bacterial community itself. In the following we provide insights into the benefits of rhodopsin phototrophy gained from experiments with expression of PR in heterologous hosts followed by experimental evidence for the use of PR in marine bacteria naturally harboring the PR photosystem.
Clues to the Mechanisms of Proteorhodopsin Photophysiology from Expression of Proteorhodopsin in Heterologous Hosts
Expression of PR in heterologous bacterial hosts has been an important approach to investigate both biochemical characteristics of PR and potential effects of light-driven proton pumping on bacterial physiology. Recently, a functional metagenomics assay that allows detecting pH changes from light-induced proton pumping by E. coli cells carrying fosmids with environmental DNA encoding rhodopsins was presented (138). Analyses of heterologously expressed rhodopsins have provided mechanistic understanding of the potential use for rhodopsin phototrophy in different aspects of cellular bioenergetics.
Given the central role of the proton motive force for driving motility in E. coli, Walter et al. (139) determined the bioenergetic potential of PR light harvesting by quantifying swimming and flagellar rotation speeds in E. coli cells expressing PR (compared to wild-type E. coli without the PR gene). Under standard aerobic growth conditions, light exposure had no effect. However, in cells under respiratory stress (from the poison azide, which inhibits respiration, or from low-oxygen conditions), light exposure considerably stimulated the flagellar rotation speeds in the E. coli with PR (139). Interestingly, the relative contribution of PR light harvesting to total proton motive force increased with increasing respiratory stress, ultimately essentially covering the cellular energy demands, and even led to improved survival under such stress (i.e., having potential to replace the proton motive force driven by active respiration).
Further exploring the potential for light energy harvesting through PR, Martinez et al. (127) screened fosmid libraries from metagenomic DNA from the ALOHA station in the Central North Pacific Gyre, using E. coli as host bacterium, for the presence of expressed retinal synthesis and PR genes that could generate a functional PR photosystem. Orange or red colony pigmentation of the E. coli host was used as a possible indication of recombinants with the PR photosystem, which resulted in two distinct clones with confirmed insertions of the crtEIBY, blh, and prd genes. Mutational analysis confirmed the function of each of the PR photosystem genes, and assays showed light-induced proton pumping across the cell membrane in both clones (but not in the mutants devoid of functional PR). Moreover, the E. coli recombinants with functional PR were able to synthesize 29% more ATP in the light than in darkness, thanks to the activity of proton-ATP synthase (127). This represented the first direct measure of photophosphorylation in bacteria containing PR.
The ability to use a PR-derived proton motive force for driving distinct enzymatic reactions was further investigated in E. coli BL21 carrying PR and [NiFe]-hydrogenase genes on plasmids (16). Under illumination, coexpression of the two genes increased production of hydrogen gas by 30% compared to that with hydrogenase gene expression only. Further, increasing the light intensity from 70 to 130 μmol photons m−2 s−1 caused a yield improvement up to 80%. Subsequent work on fine-tuning the genetic setup in relation to defined growth conditions has resulted in the construction of an E. coli MG1655 ΔhemD mutant that under anaerobic conditions reaches 10% higher growth yields upon growth with lactose (but not glucose) in the light compared to darkness, thanks to the expression of PR carried on a plasmid (140). Interestingly, the authors noted that light stimulation was found only when using a codon-optimized SAR86 PR gene (i.e., from a gammaproteobacterium like E. coli), while tests with four other PRs from Flavobacteriia and Alphaproteobacteria were unsuccessful.
Johnson et al. investigated the benefit of PR proton pumping for physiological functions in another heterotrophic Gammaproteobacteria species, Shewanella oneidensis MR-1, in part also to explore potential practical aspects of metabolic engineering (15). The PR gene from a SAR86 clade bacterium expressed from a broad-host-range vector in S. oneidensis resulted in an estimated 40,000 PR molecules per cell, and light exposure of starving cells caused similar levels of energized cells as amendment of the medium with lactate as an organic matter source under anaerobic conditions. Moreover, cultures exposed to light maintained viability over 6 days of stationary-phase starvation, while viability in dark cultures decreased to levels only 25% of those for cultures exposed to light. Measures of currents generated by cells colonizing the electrode in an electrochemical chamber showed that the oxidation current increased with increasing light intensity from 0.7 mW cm−2 to 9.0 mW cm−2. These findings potentially indicate that the PR-generated proton motive force increases the rate of uptake of lactate. In a complementary study, PR was shown to provide sufficient proton motive force to run substrate-level phosphorylation under anaerobic growth with lactate in an S. oneidensis MR-1 ATP synthase mutant (141).
Collectively, these studies support the long-held view that light-driven PR proton pumping can provide important benefits to bacteria, both by the immediate strengthening of a variety of cellular processes directly dependent on the proton motive force and by providing the driving force for production of ATP. These studies also raise a series of questions relevant to understanding the mechanisms for proton motive force generation and/or increase in ATP production rates and, in particular, how adequate signaling is obtained to generate a required balance between the generation of ATP, reducing power, and cellular building blocks. Future work on PR gene expression in heterologous hosts could provide novel insights into how these complex processes are regulated.
Increased Subsistence of Naturally Proteorhodopsin-Containing Bacteria under Starvation
Gómez-Consarnau et al. (35) investigated the growth and light responses of a PR-containing member of the genus Vibrio, strain AND4, which is closely related to the taxonomically described species V. campbellii. Vibrio sp. AND4 prefers growth under eutrophic conditions and does not show improved growth yields in the light compared to darkness (Fig. 5). However, during extended periods of starvation (up to 2 weeks), biomass levels and bacterial abundance remain 40 to 60% higher in the light (35) (Fig. 5). Parallel analyses of a PR gene deletion mutant and a mutant strain complemented with the PR gene in trans provided the first direct evidence for a role of PR in mediating the light responses in bacteria naturally carrying the PR gene (wherefore we have chosen to use their designated gene name, prd, for the PR gene sensu stricto). Regrowth experiments after starvation periods in light or darkness with the wild-type and mutant strains, where the most rapid growth responses were found for the wild-type strain starved in the light, confirmed the fitness benefit for promoting survival of PR light energy harvesting under starvation.
FIG 5.
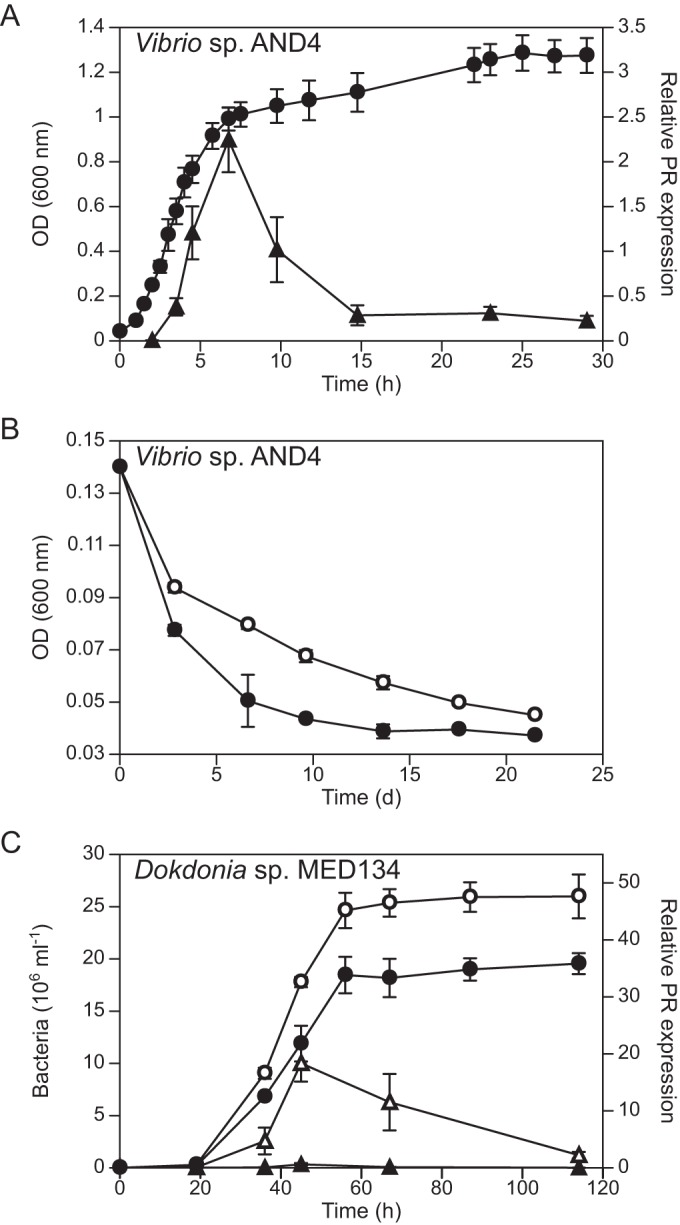
Temporal dynamics in bacterial abundance and proteorhodopsin gene expression. (A) Growth of Vibrio sp. AND4 in Zobell rich medium as measured by optical density (circles) and relative expression of the PR gene (triangles). Relative expression values were obtained by quantitative real-time PCR; the housekeeping genes rpoD and rpoZ were used for normalization. (Redrawn from reference 63.) (B) Dynamics in optical density of Vibrio sp. AND4 during starvation in constant light (open circles) and in darkness (closed circles). (Redrawn from reference 35.) (C) Growth of the flavobacterial isolate Dokdonia sp. MED134 in the light (open circles) and in the dark (closed circles) in seawater medium with complex organic matter as a carbon source. Also shown is relative expression of the PR gene in the light (open triangles) and in the dark (closed triangles). Error bars denote the standard deviations from three biological replicates; if not visible, error bars are hidden in the symbols (for gene expression in panel C, error bars denote standard error from biological triplicates). (Redrawn from reference 117.)
Further work on Vibrio sp. AND4 and V. campbellii BAA-1116 has provided detail on the effects of PR phototrophy during starvation (63, 142). Wang et al. (142) characterized the function of PR in V. campbellii BAA-1116, establishing its light-induced proton pumping ability and demonstrating a 61% increase in cellular levels of ATP concentration upon light exposure compared to dark conditions and compared to those in a mutant lacking the PR gene. Starvation experiments with cells in nutrient-poor artificial seawater revealed that when challenged with respiratory stress induced by the poison sodium azide, cell viability was substantially higher in the light than in darkness (142). This emphasizes the importance of photophosphorylation for sustaining viability under nutrient-poor conditions.
In Vibrio sp. AND4, improved survival under starvation in the light was observed only for cells in late stationary phase and not for actively growing cells. This is consistent with low levels of PR gene expression during exponential growth phase and a pronounced peak in expression right at the shift between active growth and stationary phase in this strain (63) (Fig. 5). For comparison, in BAA-1116, PR gene expression peaks later and well after entry into stationary phase. Curiously, in BAA-1116, survival is better in darkness than in light, possibly due to damage by light stress or induction of prophage (142). Another difference between these closely related strains is that the PR gene expression is induced by nutrient limitation in AND4 (63) but is induced by light in BAA-1116 (142). Mutational analysis further clarified that the gene encoding the alternative sigma factor RpoS, a major regulator of cellular adjustments to stationary phase, positively regulates PR gene expression in BAA-1116 (142). Quantitative PCR analyses further showed that rpoS gene expression precedes PR gene expression in AND4 (63). Light-induced responses have been investigated also in the copiotrophic Photobacterium angustum S14, another member of the family Vibrionaceae. Like Vibrio sp. AND4, this bacterium reaches equal cell yields in the light and in darkness when growing in rich medium (18 mM carbon as glucose) in batch cultures. However, using the continuous-culture approach to quantify physiological responses under nutrient limitation, light-exposed cultures (12-h light and 12-h dark cycles) showed rearrangements of cell stoichiometry and elevated estimated yield values in rich medium (even though direct measures of cell abundance were higher in darkness, which is possibly attributable to fluctuations in flow rates), whereas no influence of light in cultures with 5-fold-diluted medium were detected (143).
Steindler et al. (124) investigated a wide variety of cellular properties during growth and in stationary phase with carbon starvation in the PR-containing strain “Ca. Pelagibacter ubique” HTCC1062, a member of the cosmopolitan SAR11 clade. During exponential growth, cells were similarly comma shaped in both light and dark, and while cell shape remained stable after entry into stationary phase in the light, cells in the dark became smaller and more unevenly shaped (and connected by pili). Light exposure further affected the oxygen consumption patterns, so that in late exponential phase and into stationary phase, oxygen concentrations in the seawater medium decreased less in the light than in the dark, although cell abundances under the two light conditions were similar at around 5 × 108 cells ml−1. At different times in stationary phase, experiments to measure ATP content per cell upon short-term (5-min) light exposure were carried out (124). This showed that ATP per cell increased around 17% in the light compared to darkness in early stationary phase, and values increased to around 50% later into stationary phase (similar to the 61% observed in Vibrio sp. BAA-1116 [142]). This suggests that the relative importance of photophosphorylation as a means of obtaining energy increases with the severity of starvation. Further, light resulted in 65% higher uptake rates of the N- and S-containing organic acid taurine, providing first evidence that light exposure in PR-containing bacteria can stimulate ATP-dependent transport of resources into the cell (124). Microarray analysis was used to determine changes in whole-genome gene expression patterns induced by light. Nearly 10% of the genes (n = 132) were differentially expressed, around half of which were upregulated in the light. These included transcriptional regulators and, for example, genes in glycolysis and the glyoxylate shunt. In contrast, the pilin gene and several genes in the pathway of oxidative phosphorylation were upregulated in the dark. Notably, the PR gene also was upregulated 2-fold in the dark, but overall the PR gene expression appears to be relatively stable in this SAR11 clade representative (in comparison to SAR11 PR transcripts in nature). These consistent results, from measures of oxygen consumption to gene expression patterns, support the conclusion that oxidative phosphorylation (i.e., respiration) rates were reduced in carbon-starved “Ca. Pelagibacter ubique” cells in the light (124), thanks to PR phototrophy providing ATP.
Taken together, the analyses of bacteria with such different life strategies as the oligotrophic SAR11 clade representative “Ca. Pelagibacter ubique” HTCC1062 and the copiotrophic Vibrio sp. AND4 appear to indicate principal similarities in how bacteria use PR phototrophy to promote survival under starvation. Nevertheless, future direct comparisons of distinct bacteria under controlled experimental conditions are likely to uncover exciting variations in the efficiency and molecular regulation of how rhodopsin phototrophy contributes to promoting fitness under starvation conditions.
Light-Stimulated Growth of Naturally Proteorhodopsin-Containing Bacteria
Growth stimulation by light.
Light-mediated growth stimulation has currently been reported for five different bacteria, of which four are members of the class Flavobacteriia (Bacteroidetes) and one is a Gammaproteobacteria (Fig. 5). Laboratory experiments with the naturally PR-containing isolate Dokdonia sp. MED134 (Flavobacteriaceae, Bacteroidetes) were the first to show that PR light harvesting could result in improved growth of marine bacteria (65). Growth stimulation was strongest for cultures growing under exposure to green light, in accordance with the wavelength absorbance maximum of the MED134 PR. Moreover, stimulation of growth was most pronounced in seawater with low concentrations of organic carbon, where over 4-fold-higher cell yields in the light compared to darkness were recorded (65). Subsequent experimental analyses on Dokdonia sp. MED134 confirmed that the relative benefit of PR phototrophy was larger in seawater with low concentrations of dissolved organic carbon (DOC), showing the stimulation of light on both cell yields and growth rates (135). This study provided further evidence for the role of PR in mediating the positive growth response to light, using a specific inhibitor of the retinal biosynthesis pathway, 2-(4-methylphenoxy)triethylamine hydrochloride (MPTA); while comparable cell yields were obtained with or without MPTA in the dark, growth in seawater with MPTA abolished the positive light response.
It is generally recognized that both the quantity and the quality of DOC are likely to influence bacterial growth (144). Palovaara et al. (117) compared the phototrophic growth response of Dokdonia sp. MED134 in seawater with low concentrations of either a mixed organic carbon source (yeast extract and peptone [YEP]) or the single compound alanine. This showed that the relative benefit of PR light energy harvesting was notably more pronounced during growth with only alanine, emphasizing the role of DOC quality or composition in conjunction with light for determining fundamental growth characteristics of PR-containing bacteria (117).
In the first experiments with another flavobacterial isolate, Polaribacter sp. MED152, growth in rich medium (1:8 strength marine broth) was similar in light and darkness (134). Still, subsequent analyses in growth media with lower levels of organic matter and nutrients showed that this Flavobacteriia representative also can use its PR phototrophy machinery to achieve higher growth yields in the light (145).
Work on the PR-containing psychrophilic sea ice bacterium Psychroflexus torquis ATCC 700755 (Flavobacteriia) has provided further intriguing knowledge on how light affects distinct bacteria (146). In this bacterium, growth is generally stimulated under illumination compared to darkness, with better growth achieved at very low light levels of 3.7 μmol photons m−2 s−1 than at 27.7 μmol photons m−2 s−1. The growth response at such low light levels is remarkable but was suggested to be in accordance with light levels attainable in sea ice. It should be noted that there are at present no studies that in a comprehensive way have investigated PR phototrophy responses under different light intensities (but see reference 35). Importantly, the light stimulation of growth in P. torquis is salinity dependent, so that the strongest responses to light are found at salinities lower (17.5) or higher (70.0) than the salinity for optimal growth (35.0). The PR gene expression in this strain is stable across growth phases and different light conditions (as normalized against the SSU rRNA gene expression). Nevertheless, the relative abundance of PR protein in the cells, as determined from liquid chromatography (LC)-MS, varied in relation to both light and salinity conditions; the highest abundance was found in the light at intermediate salinities (146). The distinct patterns of gene expression compared to relative protein abundance suggested that PR in P. torquis ATCC 700755 is under posttranscriptional regulation. Further detailed experiments on P. torquis uncovered pronounced proteome rearrangements under distinct light and salinity conditions (147). Notably, proteins involved in central metabolic pathways increased under light compared to dark conditions, complemented by anaplerotic CO2 fixation enzymes. Moreover, light significantly influenced the relative abundance of a range of membrane transporters, in particular TonB-dependent transporters (147). These findings suggest that PR light energy harvesting allows P. torquis to adapt to the dynamic life conditions in sea ice.
Stimulation of growth by light is also observed in the flavobacterial isolate N. marinus S1-08, which contains three phylogenetically and functionally distinct rhodopsins (i.e., proton-, sodium-, and chloride-pumping varieties) (43). In low-carbon seawater medium (0.14 mM carbon), its growth was around 30% higher in the light than in the dark. Still, it remains an intriguing issue to unravel whether this growth response is specifically determined by any particular of the rhodopsins or whether it results from the concerted action of all three rhodopsins.
In a manner similar to that in these Flavobacteriia, growth of the proteorhodopsin-containing marine gammaproteobacterium HIMB30, which contains the genetic machinery for carbon fixation (complete Calvin cycle and carboxysome components) but not chlorophyll, is also improved in the light (148). In this strain, cell yields in seawater enriched with low-micromolar concentrations of glucose doubled under continuous light compared to darkness; under 12:12-h light-dark cycles, yields increased by around 30%.
Anaplerotic reactions and central metabolism during light-stimulated growth.
The findings of an ample set of genes encoding anaplerotic enzymes in the genome analysis of Polaribacter sp. MED152 inspired the first measures of the potential stimulation of bicarbonate fixation through PR phototrophy in an otherwise heterotrophic marine bacterium (134). This showed that CO2 fixation rates were nearly twice as high in the light than in darkness. Still, per-cell rates of fixation were very low (<0.001 fg C cell−1 h−1), possibly reflecting that the bacteria were grown in rich medium (full-strength marine broth diluted 1:8 with seawater) where PR phototrophy did not have an influence on bacterial growth yields. Subsequent comparative genome analysis of Bacteroidetes genomes revealed that PR-containing Bacteroidetes have a higher number of genes involved in anaplerotic CO2 fixation than Bacteroidetes without PR (149). Expanded work on the photophysiology of anaplerotic reactions in Dokdonia sp. MED134 was carried out during growth in seawater cultures with relatively low DOC concentrations (i.e., enrichment with 0.3 to 0.7 mM C in the form of yeast extract and peptone [YEP] or alanine) (117). Under these conditions, per-cell CO2 fixation rates of 0.02 to 0.47 fg C cell−1 h−1 were over 100-fold higher than those for MED152 growing in rich medium (i.e., around 30 mM C). Anaplerotic CO2 fixation in MED134 thus contributed 24 to 31% of cell carbon in the light, with the highest levels in the cultures with alanine, while levels were below 14% in the dark (117).
Mechanism of proteorhodopsin phototrophy: insights from gene expression analyses of rhodopsin-containing bacterial isolates.
In Dokdonia sp. MED134, PR gene expression levels are substantially higher upon growth in the light than with growth in darkness (65, 117, 135, 150) (Fig. 5). Results for genes involved in the synthesis of the PR cofactor retinal are more varied, with some genes being upregulated in the light and some remaining stable (135). Comparison of genome-wide gene expression patterns showed that an overall 20% of genes carried in the genome of MED134 (n = 601) are differentially expressed in light and dark cultures, with around half showing higher relative expression levels in the light (compared to 10% in “Ca. Pelagibacter ubique” HTCC1062 [124]). Notably, a histidine kinase was highly upregulated in the light, indicating its possible involvement in regulating the PR light response (135). Also, one cryptochrome and two putative DNA photolyase/cryptochrome genes were more highly expressed in light, while other genes containing light-sensing domains were equally expressed in light and darkness. Further, in accordance with the general stimulation of growth, transcript abundance was significantly higher in the light for several genes involved in central metabolic pathways (e.g., glycolysis and the TCA cycle) and genes for membrane transporters (e.g., Na+-dependent alanine or phosphate transporters). Genes involved in anaplerotic reactions (e.g., bicarbonate uptake and CO2 fixation through carboxylating enzymes) were not upregulated in the light; no differences in ATP synthase transcript abundance were found (135).
Detailed monitoring of relative gene expression patterns of a selection of genes in MED134 by qPCR showed that PR gene expression levels in the light vary considerably over time (20- to 100-fold), with peak values during late exponential phase (117) (Fig. 5). PR gene expression was detected at all growth phases also in the dark, but at levels 10 to 30 times lower than in the light (Fig. 5). Further, a number of genes encoding enzymes involved in bicarbonate acquisition and anaplerotic CO2 fixation were more highly expressed in the light, although the ranges of response for these genes were much smaller than for the PR gene. Remarkably, the two genes in the glyoxylate shunt, encoding isocitrate lyase and malate synthase, showed very pronounced dynamics. The relative expression of these genes was very low and relatively stable across growth phases in both the light and darkness in seawater with mixed DOC in the form of YEP. A similar (lack of a) pattern was observed during growth with the single carbon compound alanine in darkness. However, in light-exposed seawater cultures with alanine, the two glyoxylate shunt genes changed nearly 200-fold over time, with around 400-fold higher values in the light than in darkness from late exponential phase onwards. This suggested that glyoxylate shunt regulation in MED134 is an important component of the PR phototrophy response in a manner strongly dependent on the quality of available DOC.
Curiously, in the flavobacterial isolate Dokdonia sp. PRO95, which has a PR gene that is 76% identical to that of Dokdonia sp. MED134, growth is not stimulated by light. In this strain, PR gene expression levels normalized to the SSU rRNA gene were similar in the light and in the dark (93). It should be noted, though, that the original work on PRO95 was done in seawater media with 10× higher, or more, concentrations of organic carbon (9.7 and 121 mM carbon) than in the seawater used for experiments with MED134. Nevertheless, subsequent analysis in artificial seawater amended with only 0.14 mM carbon confirmed that PRO95 (which also turns out to have a rhodopsin affiliated with sodium-pumping “NDQ motif” rhodopsins) does not respond to light by increasing growth yields, even though light induction of proton pumping was recorded under these conditions (42). Genome-wide gene expression analysis of PRO95 cells in half-strength marine broth or artificial seawater showed that rhodopsins as well as associated retinal synthesis genes were expressed under these conditions. Gómez-Consarnau et al. (150) recently found compelling indications from gene expression analyses that differences in growth responses to light between the closely related Dokdonia sp. strains MED134 and DSW-1T on the one hand and strain PRO95 on the other hand are linked to vitamin B1 acquisition. Thus, the authors suggested that the former two species, which are vitamin B1 auxotrophs, efficiently use light to take up vitamin B1 from their surroundings, while the latter species, which is a vitamin B1 autotroph, is inefficient in vitamin B1 uptake and thus needs to spend substantial energy on vitamin synthesis (150). These findings provide an important impetus to investigate the ecological settings under which rhodopsin photoheterotrophy provides fitness benefits.
Proton-Pumping Rhodopsins in Chlorophyll-Containing Microbes and Associated Viruses
There are several marine microorganisms that have both rhodopsin genes and genes for synthesis of chlorophyll. These include a few bacteria with bacteriochlorophyll a, such as Fulvimarina pelagi HTCC2506T, which carries a xanthorhodopsin-like photoprotein that putatively is involved in proton translocation (151). It is true also of several chlorophyll a-containing eukaryotic phytoplankton in the Alveolata and Viridiplantae, as it is for several cyanobacteria. In many cases, the rhodopsins in these organisms seem to be used as sensory proteins or as ion channels (for example, in Chlamydomonas) and not as proton pumps. However, in a few cases there has been experimental demonstration of proton-pumping rhodopsins in chlorophyll a-based photoautotrophs that harbor these photoproteins.
Analysis of transcripts from Long Island Sound revealed that dinoflagellates can carry and express a gene for a xanthorhodopsin-like photoprotein with genetic features consistent with proton pumping (152). This was immediately followed by a report on a proton-pumping rhodopsin in a specific marine eukaryote species, the heterotrophic dinoflagellate Oxyrrhis marina. O. marina CCMP1788 contains both a sensory rhodopsin related to other eukaryotic rhodopsins and a rhodopsin phylogenetically affiliated with proton-pumping bacterial xanthorhodopsins (47). The latter rhodopsin was highly expressed via typical eukaryote transcriptional regulatory mechanisms in O. marina and was found in several other dinoflagellates as well. Slamovits et al. (47) further reported that the photosynthetic dinoflagellate Karlodinium micrum contains another rhodopsin affiliated with PRs in Proteobacteria. The authors speculated that proton-pumping rhodopsins could contribute to the acidification of feeding vacuoles in phagotrophic dinoflagellates. Subsequent analyses in the photosynthetic dinoflagellate Prorocentrum donghaiens CCMAXU-364 showed that xanthorhodopsin gene expression is regulated by light and influenced by the light spectrum and intensity (153). Recent work on O. marina CCMP1795 showed that light induces rhodopsin expression and promotes survival under starvation (79), supposedly by providing energy for maintenance of basic cell integrity.
An exciting development in marine microbial oceanography is the recent indication that rhodopsins may be expressed in diatom and Phaeocystis lineages in the open ocean but not in coastal areas (154). These findings resulted from studies investigating adaptations of phytoplankton to iron deficiency in the surface ocean, where the independency of rhodopsins from iron cofactors, in contrast to chlorophyll reaction centers, might provide a selective advantage. Further exploration of genomes and transcriptomes from isolated phytoplankton from different seas revealed rhodopsin expression in a wide variety of phytoplankton species (155). Indeed, laboratory experiments showed that the rhodopsin gene and protein expression in the open ocean diatom Pseudo-nitzschia granii UWOSP1E was regulated by iron, with highest levels under iron-limited growth (155). The authors thus conjectured that rhodopsins could “influence carbon cycling indirectly by maintaining seed populations of PR-containing diatoms under chronic iron stress.”
Genes predicted to encode a distinct cluster of rhodopsins have been found in viruses with large genomes that infect unicellular eukaryotes (156), and by using these rhodopsins as queries, they were also found in metagenome libraries from the Global Ocean Survey. Analysis of conserved amino acids for rhodopsin functioning indicated that these were sensory rhodopsins, thus potentially affecting, for example, phototaxis of infected photosynthetic protists, such as Phaeocystis (156). Such virus-associated rhodopsins have also been found in Red Sea metagenomes (103). Taking these findings together, future analyses of the way in which rhodopsin- and chlorophyll-based systems for gathering light energy are coordinated promise to provide fascinating novel insights into the factors that control the ecology of marine planktonic microorganisms under different environmental conditions.
TOWARD QUANTIFICATION OF MARINE PHOTOHETEROTROPHY
A large number of studies show that bulk activities of heterotrophic bacteria (here broadly referring to nonphotosynthetic prokaryotes) in aquatic environments are affected by light (120, 157–160). Moreover, laboratory experiments indicate that light can influence the structure of bacterioplankton assemblages (161). The current knowledge of the general influence of light on heterotrophic bacterioplankton activity was comprehensively reviewed by Ruiz-González et al. (162). Many unknowns remain, however, especially with respect to the differential effects of light on oxygenic phototrophs versus photoheterotrophic microorganisms with respect to productivity and nutrient cycling. For example, and importantly, the degree to which bacterioplankton growth and survival are indirectly influenced by light effects on phytoplankton photosynthesis and dissolved organic matter production, compared to direct light effects on the photoheterotrophic metabolism of rhodopsin- or bacteriochlorophyll-containing microbes, remains to be determined.
While current studies have provided a rich perspective on the diversity and function of marine microbial rhodopsins, it is essential to obtain more quantitative constraints on the contribution of photoheterotrophy-mediated controls on biogeochemical fluxes of carbon and energy. An initial modeling approach applied to photoheterotrophic bacteria compared net benefits of light harvesting compared to photosystem maintenance costs for bacteriochlorophyll- and rhodopsin-containing bacteria (137). While this was a useful exercise, many of the model assumptions about light-harvesting efficiencies and other parameters were not well constrained, and the great physiological variability known to be represented in opsin-containing microbes was not incorporated in the model. This initial analysis concluded that light harvesting could be beneficial to bacteriochlorophyll-containing bacteria but not to bacteria with rhodopsins (except under high light intensities or for bacteria with a large number of rhodopsins in their membranes). While this conclusion largely contradicts experimental data with rhodopsin-containing bacteria (see Experimental Assessment of Rhodopsin Photoheterotrophy in Heterologous Hosts and Native Aquatic Bacteria above), such model-based analyses could be further explored to determine how different parameter values (e.g., number of photosynthetic units per cell and energy yield per light unit) affect the potential net benefit to rhodopsin-containing bacteria. For example, the use of accessory pigments in addition to retinal (as found in xanthorhodopsin) may have a large effect on photosynthetic cross sections and light utilization efficiencies. It is quite likely that model parameter values differ significantly between bacteria that use rhodopsins for survival and bacteria in which light stimulates growth, and parameters such as carbon sources utilized, life history strategies, and other biologically relevant traits may differ also.
We recognize that there are important gaps in knowledge on the kinetics of a range of biochemical and physiological processes that ultimately determine the energetic benefit of rhodopsin light harvesting. Still, there are a few studies that provide actual estimates of a number of variables in marine bacteria of relevance to quantitatively assess the potential for light harvesting. Thus, Béjà et al. (96) estimated that SAR86 bacteria may contain 2.4 × 104 PR molecules per cell. The corresponding estimate was 1.0 × 104 PR molecules per cell for SAR11 bacteria (55). For Flavobacteriia, Yoshizawa et al. (67) estimated that Winogradskyella sp. strain PG2 in culture has approximately 5.2 × 104 PR molecules per cell (cell size, 1.2 μm long and 0.5 μm wide) or 1.6 × 104 PR molecules per cell for the size of these bacteria in the sea. The functional analyses of a diverse subset of eight Flavobacteriia isolates represented the first direct measures of light-driven proton pumping by PR in naturally PR-containing marine bacteria, indicating a rate of 124 protons per PR molecule per minute (67). Subsequent studies on aquatic PR-containing bacteria have confirmed this finding (146). In a heterologous expression system, S. oneidensis MR-1 produced 4.0 × 104 PR molecules per cell while growing anaerobically in minimal medium (15). In another heterologous expression system, Martinez et al. (127) found that starved E. coli harboring recombinant PR retrieved 2.2 × 105 molecules of ATP from 5 min of illumination, which was compared to 9 × 105 molecules of ATP from heterotrophic utilization of succinate under similar conditions. Similarly, modeling work by Walter et al. (139) suggested that under respiratory stress “PR is able to maintain E. coli cellular pmf [proton motive force] near this maximum potential (VPR ∼ −0.2V) with sufficiently bright illumination (KM ∼ 60mW/cm2),” which is similar to the corresponding values for E. coli aerobic respiration. Recently, a surprisingly low estimate of around 300 PR molecules per cell in membranes in E. coli was reported to yield a 10% increase in growth under light with specific growth conditions (140).
CONCLUDING REMARKS
While microbial rhodopsins span a wide range of divergent amino acid sequences, their tertiary structure and general biochemical function are remarkably conserved. However, rhodopsins are distributed among microbes belonging to a very wide variety of taxa, having vastly different genomic backgrounds and distinct physiologies. This suggests that the ecophysiological roles of rhodopsins will differ substantially among the variety of rhodopsin-containing bacteria. Accordingly, rhodopsin genes are found in the genomes of Bacteria and Archaea having very different life strategies. For example, among heterotrophic bacteria, PR genes are present in members of the widespread SAR11 clade, which are oligotrophic bacteria with the smallest known genome size for free-living bacteria. At the other end of the spectrum, PR genes are found in copiotrophs in the genus Vibrio with severalfold-larger genomes.
Therefore, it seems clear that the light-driven proton pumping of PR serves a continuum of functions, from promoting long-term starvation survival and short-term persistence under low-nutrient conditions to growth rate and yield enhancement. As such, promotion of starvation survival and stimulation of growth are the two extremes by which PR phototrophy could benefit marine bacteria. This provides the framework necessary for interpreting seemingly conflicting results from experiments with different bacteria or analyses of light responses of bacterioplankton in their native marine environment. In fact, one should expect fundamentally different modes of regulation of metabolism and physiology depending on whether light energy harvested by rhodopsins is used for growth or for survival.
There are cases where light does not immediately stimulate growth or long-term survival under starvation of rhodopsin-containing bacteria. In such bacteria, rhodopsins probably are providing more subtle physiological or ecological benefits. Further, one can envision significant interactions between rhodopsins and other gene systems, whereby rhodopsin genes may affect the regulation and function of other genes in the same genome. For example, PR phototrophy may cause some genes to become unnecessary, and therefore these genes may be lost through evolution. The function of some other genes, such as genes for transporters that utilize the chemiosmotic proton motive force, may be enhanced by the activity of the rhodopsin, and in this case, these genes might be favorably selected for. Ingenious analyses, using alternative experimental approaches, with different rhodopsin-containing microbes will be necessary to reveal ecological benefits, as well as the evolutionary trajectories that have led to the widespread distribution and maintenance of PR photosystems among highly divergent bacterial groups.
Upon the initial discovery of PR, Béjà et al. (28) speculated that PR could potentially drive an unrecognized kind of photosynthesis. RuBisCO, the key enzyme that fixes CO2 in the Calvin cycle, has been found in strain IMCC2047 (Gammaproteobacteria), which also has a rhodopsin (61). Still, rhodopsin-driven proton pumping does not directly generate the reductant necessary for CO2 fixation, as does, for example, oxygenic photosynthesis. In order to support net CO2 fixation, then, reducing power would have to be obtained from exogenous chemical sources. There is as yet no direct evidence for metabolisms where rhodopsin-based energy generation is combined with generation of reducing power. Thus, to date, rhodopsin-based light energy capture is known to support photoheterotrophy, not net photoautotrophy. Yet, untangling the intricate regulation of mechanisms for ATP, reducing power, and cell carbon acquisition in cells that contain multiple routes for production of these essential components will be a promising venue for future research.
Another intriguing venue of research will concern the physiology and regulation in those microorganisms with several rhodopsin genes. The case of C. reinhardtii strains is particularly complex, but there are several bacteria with three or four rhodopsin genes. Some of them clearly have different functions, such as the three genes in N. marinus S1-08T, but even in this well-known case, the regulation has not been well studied. Other cases involve microorganisms with both chlorophyll- and retinal-based systems for using light energy. The regulation in this case must be fascinating.
Even in photoheterotrophic microorganisms with a single rhodopsin proton pump, many aspects remain unknown. There is evidence that both the concentration and the quality of organic matter will result in different expression and benefits of the PR. It is likely that the response will also depend on inorganic nutrients, temperature, light quality and quantity, oxygen concentration, and many other environmental factors that have yet to be explored.
In addition, the functions of many rhodopsins are still unknown, and the ecological and physiological roles of others, such as chloride or sodium pumps, remain to be explored experimentally. There is also undoubtedly a considerable amount of rhodopsin diversity remaining to be discovered. For example, how extensive are additional rhodopsin-associated accessory pigments acting like those of xanthorhodopsin antennae in the oceans? Are there more complex antennae intermediate between xanthorhodopsin and those of chlorophyll systems? How many clusters are we missing in the rhodopsin tree? How extensive has lateral gene transfer been? We can foresee a substantial number of novel rhodopsins being discovered in the near future and hope that, eventually, a less confusing nomenclature will be achieved, once the natural diversity of rhodopsins has been mapped satisfactorily.
An even more challenging yet critical objective is to determine how rhodopsin-based light harvesting affects the carbon cycling and ecosystem energy fluxes globally and how PR-containing bacteria might react to increasing CO2 concentrations and climate change relative to strictly heterotrophic bacteria. All these questions herald future discoveries and insights that are sure to arise from ecologically grounded rhodopsin research. Over the past few decades, many studies have fruitfully leveraged interdisciplinary research on rhodopsins carried by microorganisms in the marine environment by combining both applied and discovery-driven research, an approach that certainly can be usefully extended well into the future.
ACKNOWLEDGMENTS
We gratefully acknowledge past and present members of our research groups for making our research on rhodopsins possible. The insightful corrections, comments, and criticism from two anonymous reviewers greatly improved the manuscript.
This work was supported by grants from the Göran Gustafsson Foundation for Research in Natural Sciences and Medicine, the Swedish Research Council VR, and the Swedish Research Council FORMAS strong research program EcoChange to J.P., the Gordon and Betty Moore Foundation (3777), NSF C-MORE grant EF0424599, and the Simons Foundation SCOPE (329108) to E.F.D., and the Spanish Ministry of Economy and Competitiveness grant “EcoBGM” CTM2013-48292-C3 to J.M.G. and C.P.-A.
We declare no conflict of interest.
Biographies

Jarone Pinhassi is a Professor of Microbiology at the Department of Biology and Environmental Science at Linnaeus University, Kalmar, Sweden. Dr. Pinhassi received his M.S. in biology (1994) and his Ph.D. in microbiology (1999) at Umeå University, Sweden. He was a postdoctoral fellow at the Kinneret Lake Laboratory, Israel, and at the Marine Sciences Institute (CSIC, Barcelona, Spain). The central focus of his research is on the biodiversity and community structure of planktonic bacteria and the critical role they play in regulating nutrient turnover and energy fluxes in the marine ecosystem. For his research on light energy harvesting through rhodopsins in marine bacteria, Dr. Pinhassi is a recipient of a prize from King Carl XVI Gustaf's 50-year foundation for science, technology, and environment and was awarded the Göran Gustafsson prize in molecular biology from the Göran Gustafsson Foundation for Research in Natural Sciences and Medicine.

Edward F. DeLong received his Bachelor of Science degree in bacteriology at the University of California, Davis, and his Ph.D. in marine biology at the University of California, San Diego. He is currently Professor of Oceanography at the University of Hawaii, Manoa. He serves as Co-Director for both the Center for Microbial Oceanography: Research and Education (C-MORE) and the Simons Collaboration on Ocean Processes and Ecology (SCOPE). Dr. DeLong is a Fellow in the American Academy of Microbiology, the American Academy of Arts and Science, and the U.S. National Academy of Science. In 2016 he was elected incoming Vice President of the International Society for Microbial Ecology. His scientific interests focus primarily on questions in marine microbial genomics, biogeochemistry, ecology, and evolution. A large part of Dr. DeLong's efforts have been devoted to the study of microbes and microbial processes in the ocean, combining both laboratory- and field-based approaches.

Oded Béjà is a Professor in the Department of Biology and Environmental Science at the Technion-Israel Institute of Technology, Haifa, Israel. Dr. Béjà received his M.Sc. (1993) from the Department of Membrane Research and Biophysics and his Ph.D. (1998) from the Department of Biological Chemistry, both at The Weizmann Institute of Science, Rehovot, Israel. He was a postdoctoral fellow at the Monterey Bay Aquarium Research Institute (MBARI), Moss Landing, CA, USA, working with Dr. Edward DeLong. He is currently exploring the metabolism of planktonic microbes using novel molecular biology techniques, along with functional genomics and bioinformatics. Oded Béjà holds the Louis and Lyra Richmond Chair in Life Sciences at the Technion.

José M. González received his Ph.D. in microbiology from the University of Georgia, Athens, GA, USA. He was a postdoctoral fellow at the Department of Marine Sciences at the same university. Currently, he is Associate Professor of Microbiology at the University of La Laguna, Spain. His research interest is in using genomics for studying the microbial ecology of bacterioplankton. Dr. González has focused on research on the Roseobacter group, its quantification in the marine environment, and its physiology and metabolism by using several approaches. For the past decade, emphasis has been on the adaptations in Bacteroidetes that contribute to their success in the marine environment.

Carlos Pedrós-Alió graduated in biology at the Autonomous University of Barcelona and got his Ph.D. in bacteriology at the University of Wisconsin—Madison. After a postdoctoral stay at the Autonomous University, he became an Assistant Professor of Microbiology. He moved to the Marine Sciences Institute (CSIC, Barcelona) in 1989, where he was a Research Professor since 2000. In 2016, he moved to the National Center for Biotechnology (CSIC, Madrid). Dr. Pedrós-Alió's interest is in understanding the ecology of aquatic microorganisms. Around 2005, he started to use genomics as a tool to generate hypotheses that could later be tested experimentally. He also likes to study extreme environments such as hypersaline systems, thermal springs, or polar waters. Another interest is in finding the mechanisms maintaining a large number of rare bacteria in aquatic ecosystems. He is also interested in outreach, relationships between art and science, biology of spirituality, fiction writing, and birding.
REFERENCES
- 1.Vershinin A. 1999. Biological functions of carotenoids—diversity and evolution. Biofactors 10:99–104. doi: 10.1002/biof.5520100203. [DOI] [PubMed] [Google Scholar]
- 2.Hegemann P. 2008. Algal sensory photoreceptors. Annu Rev Plant Biol 59:167–189. doi: 10.1146/annurev.arplant.59.032607.092847. [DOI] [PubMed] [Google Scholar]
- 3.Bryant DA, Frigaard NU. 2006. Prokaryotic photosynthesis and phototrophy illuminated. Trends Microbiol 14:488–496. doi: 10.1016/j.tim.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 4.Spudich JL, Yang CS, Jung KH, Spudich EN. 2000. Retinylidene proteins: structures and functions from archaea to humans. Annu Rev Cell Dev Biol 16:365–392. doi: 10.1146/annurev.cellbio.16.1.365. [DOI] [PubMed] [Google Scholar]
- 5.Ernst OP, Lodowski DT, Elstner M, Hegemann P, Brown LS, Kandori H. 2014. Microbial and animal rhodopsins: structures, functions, and molecular mechanisms. Chem Rev 114:126–163. doi: 10.1021/cr4003769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mackin KA, Roy RA, Theobald DL. 2014. An empirical test of convergent evolution in rhodopsins. Mol Biol Evol 31:83–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ihara K, Umemura T, Katagiri I, Kitajima-Ihara T, Sugiyama Y, Kimura Y, Mukohata Y. 1999. Evolution of the archaeal rhodopsins: evolution rate changes by gene duplication and functional differentiation. J Mol Biol 285:163–174. doi: 10.1006/jmbi.1998.2286. [DOI] [PubMed] [Google Scholar]
- 8.Spudich JL, Jung K-H. 2005. Microbial rhodopsins: phylogenetic and functional diversity, p 1–23. In Briggs WR, Spudich JL (ed), Handbook of photosensory receptors. Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim, Germany. [Google Scholar]
- 9.Grote M, O'Malley MA. 2011. Enlightening the life sciences: the history of halobacterial and microbial rhodopsin research. FEMS Microbiol Rev 35:1082–1099. doi: 10.1111/j.1574-6976.2011.00281.x. [DOI] [PubMed] [Google Scholar]
- 10.Oesterhelt D, Stoeckenius W. 1971. Rhodopsin-like protein from purple membrane of Halobacterium halobium. Nat New Biol 233:149–152. [DOI] [PubMed] [Google Scholar]
- 11.Neutze R, Pebay-Peyroula E, Edman K, Royant A, Navarro J, Landau EM. 2002. Bacteriorhodopsin: a high-resolution structural view of vectorial proton transport. Biochim Biophys Acta 1565:144–167. doi: 10.1016/S0005-2736(02)00566-7. [DOI] [PubMed] [Google Scholar]
- 12.Brock TD, Petersen S. 1976. Some effects of light on viability of rhodopsin-containing Halobacteria. Arch Microbiol 109:199–200. doi: 10.1007/BF00425136. [DOI] [PubMed] [Google Scholar]
- 13.Oesterhelt D, Krippahl G. 1983. Phototropic growth of halobacteria and its use for isolation of photosynthetically-deficient mutants. Ann Inst Pasteur Microbiol 134B:137–150. [DOI] [PubMed] [Google Scholar]
- 14.Spudich JL, Bogomolni RA. 1984. Mechanism of colour discrimination by a bacterial sensory rhodopsin. Nature 312:509–513. doi: 10.1038/312509a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Johnson ET, Baron DB, Naranjo B, Bond DR, Schmidt-Dannert C, Gralnick JA. 2010. Enhancement of survival and electricity production in an engineered bacterium by light-driven proton pumping. Appl Environ Microbiol 76:4123–4129. doi: 10.1128/AEM.02425-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim JYH, Jo BH, Jo Y, Cha HJ. 2012. Improved production of biohydrogen in light-powered Escherichia coli by co-expression of proteorhodopsin and heterologous hydrogenase. Microb Cell Fact 11:2. doi: 10.1186/1475-2859-11-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Matsuno-Yagi A, Mukohata Y. 1980. ATP synthesis linked to light-dependent proton uptake in a rad mutant strain of Halobacterium lacking bacteriorhodopsin. Arch Biochem Biophys 199:297–303. doi: 10.1016/0003-9861(80)90284-2. [DOI] [PubMed] [Google Scholar]
- 18.Tomioka H, Takahashi T, Kamo N, Kobatake Y. 1986. Flash spectroscopic indentification of a fourth rhodopsin-like pigment in Halobacterium halohium. Biochem Biophys Res Commun 139:389–395. doi: 10.1016/S0006-291X(86)80003-1. [DOI] [PubMed] [Google Scholar]
- 19.Fu H-Y, Lin Y-C, Chang Y-N, Tseng H, Huang C-C, Liu K-C, Huang C-S, Su C-W, Weng RC, Lee Y-Y, Ng WV, Yang C-S. 2010. A novel six-rhodopsin system in a single archaeon. J Bacteriol 192:5866–5873. doi: 10.1128/JB.00642-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sineshchekov OA, Jung K-H, Spudich JL. 2002. Two rhodopsins mediate phototaxis to low- and high-intensity light in Chlamydomonas reinhardtii. Proc Natl Acad Sci U S A 99:8689–8694. doi: 10.1073/pnas.122243399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nagel G, Ollig D, Fuhrmann M, Kateriya S, Mustl AM, Bamberg E, Hegemann P. 2002. Channelrhodopsin-1: a light-gated proton channel in green algae. Science 296:2395–2398. doi: 10.1126/science.1072068. [DOI] [PubMed] [Google Scholar]
- 22.Nagel G, Szellas T, Huhn W, Kateriya S, Adeishvili N, Berthold P, Ollig D, Hegemann P, Bamberg E. 2003. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc Natl Acad Sci U S A 100:13940–13945. doi: 10.1073/pnas.1936192100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pastrana E. 2011. Optogenetics: controlling cell function with light. Nat Methods 8:24–25. [Google Scholar]
- 24.Deisseroth K. 2011. Optogenetics. Nat Methods 8:26–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Klapoetke NC, Murata Y, Kim SS, Pulver SR, Birdsey-Benson A, Cho YK, Morimoto TK, Chuong AS, Carpenter EJ, Tian ZJ, Wang J, Xie YL, Yan ZX, Zhang Y, Chow BY, Surek B, Melkonian M, Jayaraman V, Constantine-Paton M, Wong GKS, Boyden ES. 2014. Independent optical excitation of distinct neural populations. Nat Methods 11:338–346. doi: 10.1038/nmeth.2836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Avelar GM, Schumacher RI, Zaini PA, Leonard G, Richards TA, Gomes SL. 2014. A rhodopsin-guanylyl cyclase gene fusion functions in visual perception in a fungus. Curr Biol 24:1234–1240. doi: 10.1016/j.cub.2014.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fan Y, Solomon P, Oliver RP, Brown LS. 2011. Photochemical characterization of a novel fungal rhodopsin from Phaeosphaeria nodorum. Biochim Biophys Acta 1807:1457–1466. doi: 10.1016/j.bbabio.2011.07.005. [DOI] [PubMed] [Google Scholar]
- 28.Béjà O, Aravind L, Koonin EV, Suzuki MT, Hadd A, Nguyen LP, Jovanovich SB, Gates CM, Feldman RA, Spudich JL, Spudich EN, DeLong EF. 2000. Bacterial rhodopsin: evidence for a new type of phototrophy in the sea. Science 289:1902–1906. doi: 10.1126/science.289.5486.1902. [DOI] [PubMed] [Google Scholar]
- 29.Bamann C, Bamberg E, Wachtveitl J, Glaubitz C. 2014. Proteorhodopsin. Biochim Biophys Acta 1837:614–625. doi: 10.1016/j.bbabio.2013.09.010. [DOI] [PubMed] [Google Scholar]
- 30.Ran T, Ozorowski G, Gao Y, Sineshchekov OA, Wang W, Spudich JL, Luecke H. 2013. Cross-protomer interaction with the photoactive site in oligomeric proteorhodopsin complexes. Acta Crystallogr D Biol Crystallogr 69:1965–1980. doi: 10.1107/S0907444913017575. [DOI] [PubMed] [Google Scholar]
- 31.Inoue K, Kato Y, Kandori H. 2015. Light-driven ion-translocating rhodopsins in marine bacteria. Trends Microbiol 23:91–98. doi: 10.1016/j.tim.2014.10.009. [DOI] [PubMed] [Google Scholar]
- 32.Béjà O, Lanyi JK. 2014. Nature's toolkit for microbial rhodopsin ion pumps. Proc Natl Acad Sci U S A 111:6538–6539. doi: 10.1073/pnas.1405093111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Béjà O, Pinhassi J, Spudich JL. 2013. Proteorhodopsins: widespread microbial light-driven proton pumps, p 280–285. In Levin SA. (ed), Encyclopedia of biodiversity, 2nd ed, vol 6 Academic Press, Waltham, MA. [Google Scholar]
- 34.Brown LS. 2014. Eubacterial rhodopsins—unique photosensors and diverse ion pumps. Biochim Biophys Acta 1837:553–561. doi: 10.1016/j.bbabio.2013.05.006. [DOI] [PubMed] [Google Scholar]
- 35.Gómez-Consarnau L, Akram N, Lindell K, Pedersen A, Neutze R, Milton DL, González JM, Pinhassi J. 2010. Proteorhodopsin phototrophy promotes survival of marine bacteria during starvation. PLoS Biol 8:e1000358. doi: 10.1371/journal.pbio.1000358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sabehi G, Béjà O, Suzuki MT, Preston CM, DeLong EF. 2004. Different SAR86 subgroups harbour divergent proteorhodopsins. Environ Microbiol 6:903–910. doi: 10.1111/j.1462-2920.2004.00676.x. [DOI] [PubMed] [Google Scholar]
- 37.Sabehi G, Loy A, Jung K-H, Partha R, Spudich JL, Isaacson T, Hirschberg J, Wagner M, Béjà O. 2005. New insights into metabolic properties of marine bacteria encoding proteorhodopsins. PLoS Biol 3:e273. doi: 10.1371/journal.pbio.0030273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sharma AK, Spudich JL, Doolittle WF. 2006. Microbial rhodopsins: functional versatility and genetic mobility. Trends Microbiol 14:463–469. doi: 10.1016/j.tim.2006.09.006. [DOI] [PubMed] [Google Scholar]
- 39.Boeuf D, Audic S, Brillet-Guéguen L, Caron C, Jeanthon C. 18 August 2015. MicRhoDE: a curated database for the analysis of microbial rhodopsin diversity and evolution. Database doi: 10.1093/database/bav1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Inoue K, Ono H, Abe-Yoshizumi R, Yoshizawa S, Ito H, Kogure K, Kandori H. 2013. A light-driven sodium ion pump in marine bacteria. Nat Commun 4:1678. doi: 10.1038/ncomms2689. [DOI] [PubMed] [Google Scholar]
- 41.Riedel T, Held B, Nolan M, Lucas S, Lapidus A, Tice H, Del Rio TG, Cheng JF, Han C, Tapia R, Goodwin LA, Pitluck S, Liolios K, Mavromatis K, Pagani I, Ivanova N, Mikhailova N, Pati A, Chen A, Palaniappan K, Land M, Rohde M, Tindall BJ, Detter JC, Goker M, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Kyrpides NC, Klenk HP, Woyke T. 2012. Genome sequence of the Antarctic rhodopsins-containing flavobacterium Gillisia limnaea type strain (R-8282(T)). Stand Genomic Sci 7:107–119. doi: 10.4056/sigs.3216895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Riedel T, Gómez-Consarnau L, Tomasch J, Martin M, Jarek M, González JM, Spring S, Rohlfs M, Brinkhoff T, Cypionka H, Goker M, Fiebig A, Klein J, Goesmann A, Fuhrman JA, Wagner-Dobler I. 2013. Genomics and physiology of a marine flavobacterium encoding a proteorhodopsin and a xanthorhodopsin-like protein. PLoS One 8:e57487. doi: 10.1371/journal.pone.0057487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yoshizawa S, Kumagai Y, Kim H, Ogura Y, Hayashi T, Iwasaki W, DeLong EF, Kogure K. 2014. Functional characterization of flavobacteria rhodopsins reveals a unique class of light-driven chloride pump in bacteria. Proc Natl Acad Sci U S A 111:6732–6737. doi: 10.1073/pnas.1403051111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, Kianianmomeni A, Prigge M, Berndt A, Cushman J, Polle J, Magnuson J, Hegemann P, Deisseroth K. 2011. The microbial opsin family of optogenetic tools. Cell 147:1446–1457. doi: 10.1016/j.cell.2011.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mongodin EF, Nelson KE, Daugherty S, DeBoy RT, Wister J, Khouri H, Weidman J, Walsh DA, Papke RT, Perez GS, Sharma AK, Nesbo CL, MacLeod D, Bapteste E, Doolittle WF, Charlebois RL, Legault B, Rodríguez-Valera F. 2005. The genome of Salinibacter ruber: convergence and gene exchange among hyperhalophilic bacteria and archaea. Proc Natl Acad Sci U S A 102:18147–18152. doi: 10.1073/pnas.0509073102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Curtis BA, Tanifuji G, Burki F, Gruber A, Irimia M, Maruyama S, Arias MC, Ball SG, Gile GH, Hirakawa Y, Hopkins JF, Kuo A, Rensing SA, Schmutz J, Symeonidi A, Elias M, Eveleigh RJM, Herman EK, Klute MJ, Nakayama T, Obornik M, Reyes-Prieto A, Armbrust EV, Aves SJ, Beiko RG, Coutinho P, Dacks JB, Durnford DG, Fast NM, Green BR, Grisdale CJ, Hempel F, Henrissat B, Hoppner MP, Ishida K-I, Kim E, Koreny L, Kroth PG, Liu Y, Malik S-B, Maier UG, McRose D, Mock T, Neilson JAD, Onodera NT, Poole AM, Pritham EJ, Richards TA, Rocap G, Roy SW, et al. . 2012. Algal genomes reveal evolutionary mosaicism and the fate of nucleomorphs. Nature 492:59–65. doi: 10.1038/nature11681. [DOI] [PubMed] [Google Scholar]
- 47.Slamovits CH, Okamoto N, Burri L, James ER, Keeling PJ. 2011. A bacterial proteorhodopsin proton pump in marine eukaryotes. Nat Commun 2:183. doi: 10.1038/ncomms1188. [DOI] [PubMed] [Google Scholar]
- 48.Govorunova EG, Sineshchekov OA, Janz R, Liu X, Spudich JL. 2015. Natural light-gated anion channels: a family of microbial rhodopsins for advanced optogenetics. Science 349:647–650. doi: 10.1126/science.aaa7484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Govorunova EG, Sineshchekov OA, Spudich JL. 2016. Structurally distinct cation channelrhodopsins from cryptophyte algae. Biophys J 110:2302–2304. doi: 10.1016/j.bpj.2016.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.da Silva GFZ, Goblirsch BR, Tsai A-L, Spudich JL. 2015. Cation-specific conformations in a dual-function ion-pumping microbial rhodopsin. Biochemistry 54:3950–3959. doi: 10.1021/bi501386d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Finkel OM, Béjà O, Belkin S. 2013. Global abundance of microbial rhodopsins. ISME J 7:448–451. doi: 10.1038/ismej.2012.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.de la Torre JR, Christianson LM, Béjà O, Suzuki MT, Karl DM, Heidelberg J, DeLong EF. 2003. Proteorhodopsin genes are distributed among divergent marine bacterial taxa. Proc Natl Acad Sci U S A 100:12830–12835. doi: 10.1073/pnas.2133554100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Stingl U, Desiderio RA, Cho JC, Vergin KL, Giovannoni SJ. 2007. The SAR92 clade: an abundant coastal clade of culturable marine bacteria possessing proteorhodopsin. Appl Environ Microbiol 73:2290–2296. doi: 10.1128/AEM.02559-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Swan BK, Tupper B, Sczyrba A, Lauro FM, Martínez-García M, González JM, Luo HW, Wright JJ, Landry ZC, Hanson NW, Thompson BP, Poulton NJ, Schwientek P, Acinas SG, Giovannoni SJ, Moran MA, Hallam SJ, Cavicchioli R, Woyke T, Stepanauskas R. 2013. Prevalent genome streamlining and latitudinal divergence of planktonic bacteria in the surface ocean. Proc Natl Acad Sci U S A 110:11463–11468. doi: 10.1073/pnas.1304246110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Giovannoni SJ, Bibbs L, Cho J-L, Stapels MD, Desiderio R, Vergin KL, Rappé MS, Laney S, Wilhelm LJ, Tripp HJ, Mathur EJ, Barofsky DF. 2005. Proteorhodopsin in the ubiquitous marine bacterium SAR11. Nature 438:82–85. doi: 10.1038/nature04032. [DOI] [PubMed] [Google Scholar]
- 56.McCarren J, Delong EF. 2007. Proteorhodopsin photosystem gene clusters exhibit co-evolutionary trends and shared ancestry among diverse marine microbial phyla. Environ Microbiol 9:846–858. doi: 10.1111/j.1462-2920.2006.01203.x. [DOI] [PubMed] [Google Scholar]
- 57.Giovannoni SJ, Hayakawa DH, Tripp HJ, Stingl U, Givan SA, Cho JC, Oh HM, Kitner JB, Vergin KL, Rappé MS. 2008. The small genome of an abundant coastal ocean methylotroph. Environ Microbiol 10:1771–1782. doi: 10.1111/j.1462-2920.2008.01598.x. [DOI] [PubMed] [Google Scholar]
- 58.Courties A, Riedel T, Jarek M, Intertaglia L, Lebaron P, Suzuki MT. 2013. Genome sequence of strain MOLA814, a proteorhodopsin-containing representative of the Betaproteobacteria common in the ocean. Genome Announc 1:e01062-13. doi: 10.1128/genomeA.01062-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Oh HM, Kwon KK, Kang I, Kang SG, Lee JH, Kim SJ, Cho JC. 2010. Complete genome sequence of “Candidatus Puniceispirillum marinum” IMCC1322, a representative of the SAR116 clade in the Alphaproteobacteria. J Bacteriol 192:3240–3241. doi: 10.1128/JB.00347-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Jang Y, Oh H-M, Kang I, Lee K, Yang S-J, Cho J-C. 2011. Genome sequence of strain IMCC3088, a proteorhodopsin-containing marine bacterium belonging to the OM60/NOR5 clade. J Bacteriol 193:3415–3416. doi: 10.1128/JB.05111-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kang I, Kang D, Oh HM, Kim H, Kim HJ, Kang TW, Kim SY, Cho JC. 2011. Genome sequence of strain IMCC2047, a novel marine member of the Gammaproteobacteria. J Bacteriol 193:3688–3689. doi: 10.1128/JB.05226-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Amaral GRS, Silva BSD, Santos EO, Dias GM, Lopes RM, Edwards RA, Thompson CC, Thompson FL. 2012. Genome sequence of the bacterioplanktonic, mixotrophic Vibrio campbellii strain PEL22A, isolated in the Abrolhos Bank. J Bacteriol 194:2759–2760. doi: 10.1128/JB.00377-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Akram N, Palovaara J, Forsberg J, Lindh MV, Milton DL, Luo H, González JM, Pinhassi J. 2013. Regulation of proteorhodopsin gene expression by nutrient limitation in the marine bacterium Vibrio sp. AND4. Environ Microbiol 15:1400–1415. doi: 10.1111/1462-2920.12085. [DOI] [PubMed] [Google Scholar]
- 64.Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA, Wu D, Paulsen I, Nelson KE, Nelson W, Fouts DE, Levy S, Knap AH, Lomas MW, Nealson K, White O, Peterson J, Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers Y-H, Smith HO. 2004. Environmental genome shotgun sequencing of the Sargasso Sea. Science 304:66–74. doi: 10.1126/science.1093857. [DOI] [PubMed] [Google Scholar]
- 65.Gómez-Consarnau L, González JM, Coll-Lladó M, Gourdon P, Pascher T, Neutze R, Pedrós-Alió C, Pinhassi J. 2007. Light stimulates growth of proteorhodopsin-containing marine flavobacteria. Nature 445:210–213. doi: 10.1038/nature05381. [DOI] [PubMed] [Google Scholar]
- 66.Alonso C, Warnecke F, Amann R, Pernthaler J. 2007. High local and global diversity of Flavobacteria in marine plankton. Environ Microbiol 9:1253–1266. doi: 10.1111/j.1462-2920.2007.01244.x. [DOI] [PubMed] [Google Scholar]
- 67.Yoshizawa S, Kawanabe A, Ito H, Kandori H, Kogure K. 2012. Diversity and functional analysis of proteorhodopsin in marine Flavobacteria. Environ Microbiol 14:1240–1248. doi: 10.1111/j.1462-2920.2012.02702.x. [DOI] [PubMed] [Google Scholar]
- 68.Stepanauskas R, Sieracki ME. 2007. Matching phylogeny and metabolism in the uncultured marine bacteria, one cell at a time. Proc Natl Acad Sci U S A 104:9052–9057. doi: 10.1073/pnas.0700496104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Frigaard N-U, Martinez A, Mincer TJ, DeLong EF. 2006. Proteorhodopsin lateral gene transfer between marine planktonic Bacteria and Archaea. Nature 439:847–850. doi: 10.1038/nature04435. [DOI] [PubMed] [Google Scholar]
- 70.Ghai R, Martín-Cuadrado AB, Molto AG, Heredia IG, Cabrera R, Martin J, Verdú M, Deschamps P, Moreira D, López-García P, Mira A, Rodríguez-Valera F. 2010. Metagenome of the Mediterranean deep chlorophyll maximum studied by direct and fosmid library 454 pyrosequencing. ISME J 4:1154–1166. doi: 10.1038/ismej.2010.44. [DOI] [PubMed] [Google Scholar]
- 71.Iverson V, Morris RM, Frazar CD, Berthiaume CT, Morales RL, Armbrust EV. 2012. Untangling genomes from metagenomes: revealing an uncultured class of marine Euryarchaeota. Science 335:587–590. doi: 10.1126/science.1212665. [DOI] [PubMed] [Google Scholar]
- 72.Petrovskaya LE, Lukashev EP, Chupin VV, Sychev SV, Lyukmanova EN, Kryukova EA, Ziganshin RH, Spirina EV, Rivkina EM, Khatypov RA, Erokhina LG, Gilichinsky DA, Shuvalov VA, Kirpichnikov MP. 2010. Predicted bacteriorhodopsin from Exiguobacterium sibiricum is a functional proton pump. FEBS Lett 584:4193–4196. doi: 10.1016/j.febslet.2010.09.005. [DOI] [PubMed] [Google Scholar]
- 73.Gushchin I, Chervakov P, Kuzmichev P, Popov AN, Round E, Borshchevskiy V, Ishchenko A, Petrovskaya L, Chupin V, Dolgikh DA, Arseniev AA, Kirpichnikov M, Gordeliy V. 2013. Structural insights into the proton pumping by unusual proteorhodopsin from nonmarine bacteria. Proc Natl Acad Sci U S A 110:12631–12636. doi: 10.1073/pnas.1221629110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhu WJ, Lan YL, Lou XD, Han N, Ran TT, Xu LL, Xu DQ, Wang WW. 2013. Isolation of proteorhodopsin-bearing bacterium JL-3 from fresh water and characterization of the proteorhodopsin. FEMS Microbiol Lett 344:10–17. doi: 10.1111/1574-6968.12144. [DOI] [PubMed] [Google Scholar]
- 75.Balashov SP, Imasheva ES, Boichenko VA, Antón J, Wang JM, Lanyi JK. 2005. Xanthorhodopsin: a proton pump with a light-harvesting antenna. Science 309:2061–2064. doi: 10.1126/science.1118046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Luecke H, Schobert B, Stagno J, Imasheva ES, Wang JM, Balashov SP, Lanyi JK. 2008. Crystallographic structure of xanthorhodopsin, the light-driven proton pump with a dual chromophore. Proc Natl Acad Sci U S A 105:16561–16565. doi: 10.1073/pnas.0807162105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Imasheva ES, Balashov SP, Choi AR, Jung KH, Lanyi JK. 2009. Reconstitution of Gloeobacter violaceus rhodopsin with a light-harvesting carotenoid antenna. Biochemistry 48:10948–10955. doi: 10.1021/bi901552x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Balashov SP, Imasheva ES, Choi AR, Jung K-H, Liaaen-Jensen S, Lanyi JK. 2010. Reconstitution of Gloeobacter rhodopsin with echinenone: role of the 4-keto group. Biochemistry 49:9792–9799. doi: 10.1021/bi1014166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Guo ZL, Zhang H, Lin SJ. 2014. Light-promoted rhodopsin expression and starvation survival in the marine dinoflagellate Oxyrrhis marina. PLoS One 9:e114941. doi: 10.1371/journal.pone.0114941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Jezberová J, Jezbera J, Hahn MW. 2013. Insights into variability of actinorhodopsin genes of the LG1 cluster in two different freshwater habitats. PLoS One 8:e68542. doi: 10.1371/journal.pone.0068542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Martínez-García M, Swan BK, Poulton NJ, Gomez ML, Masland D, Sieracki ME, Stepanauskas R. 2012. High-throughput single-cell sequencing identifies photoheterotrophs and chemoautotrophs in freshwater bacterioplankton. ISME J 6:113–123. doi: 10.1038/ismej.2011.84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sharma AK, Sommerfeld K, Bullerjahn GS, Matteson AR, Wilhelm SW, Jezbera J, Brandt U, Doolittle WF, Hahn MW. 2009. Actinorhodopsin genes discovered in diverse freshwater habitats and among cultivated freshwater Actinobacteria. ISME J 3:726–737. doi: 10.1038/ismej.2009.13. [DOI] [PubMed] [Google Scholar]
- 83.Salka I, Wurzbacher C, Garcia SL, Labrenz M, Jurgens K, Grossart HP. 2014. Distribution of acI-actinorhodopsin genes in Baltic Sea salinity gradients indicates adaptation of facultative freshwater photoheterotrophs to brackish waters. Environ Microbiol 16:586–597. doi: 10.1111/1462-2920.12185. [DOI] [PubMed] [Google Scholar]
- 84.Rusch DB, Halpern AL, Sutton G, Heidelberg KB, Williamson S, Yooseph S, Wu DY, Eisen JA, Hoffman JM, Remington K, Beeson K, Tran B, Smith H, Baden-Tillson H, Stewart C, Thorpe J, Freeman J, Andrews-Pfannkoch C, Venter JE, Li K, Kravitz S, Heidelberg JF, Utterback T, Rogers YH, Falcón LI, Souza V, Bonilla-Rosso G, Eguiarte LE, Karl DM, Sathyendranath S, Platt T, Bermingham E, Gallardo V, Tamayo-Castillo G, Ferrari MR, Strausberg RL, Nealson K, Friedman R, Frazier M, Venter JC. 2007. The Sorcerer II Global Ocean Sampling expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol 5:e77. doi: 10.1371/journal.pbio.0050077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sasaki J, Brown LS, Chon YS, Kandori H, Maeda A, Needleman R, Lanyi JK. 1995. Conversion of bacteriorhodopsin into a chloride ion pump. Science 269:73–75. doi: 10.1126/science.7604281. [DOI] [PubMed] [Google Scholar]
- 86.Lanyi JK. 1986. Halorhodopsin: a light-driven chloride ion pump. Annu Rev Biophys Biophys Chem 15:11–28. doi: 10.1146/annurev.bb.15.060186.000303. [DOI] [PubMed] [Google Scholar]
- 87.Kwon SK, Kim BK, Song JY, Kwak MJ, Lee CH, Yoon JH, Oh TK, Kim JF. 2013. Genomic makeup of the marine flavobacterium Nonlabens (Donghaeana) dokdonensis and identification of a novel class of rhodopsins. Genome Biol Evol 5:187–199. doi: 10.1093/gbe/evs134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kwon S-K, Lee HG, Kwak M-J, Kim JF. 2016. Genome sequence of the marine flavobacterium Croceitalea dokdonensis DOKDO 023 that contains proton- and sodium-pumping rhodopsins. Mar Genomics 26:1–3. doi: 10.1016/j.margen.2015.11.011. [DOI] [PubMed] [Google Scholar]
- 89.Li H, Sineshchekov OA, da Silva GFZ, Spudich JL. 2015. In vitro demonstration of dual light-driven Na+/H+ pumping by a microbial rhodopsin. Biophys J 109:1446–1453. doi: 10.1016/j.bpj.2015.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Engelhard M, Schmies G, Wegener AA. 2003. Archaeal phototaxis, p 1–39. In Batschauer A. (ed), Photoreceptors and light signalling. Royal Society of Chemistry, Cambridge, United Kingdom. [Google Scholar]
- 91.Jung KH, Trivedi VD, Spudich JL. 2003. Demonstration of a sensory rhodopsin in eubacteria. Mol Microbiol 47:1513–1522. doi: 10.1046/j.1365-2958.2003.03395.x. [DOI] [PubMed] [Google Scholar]
- 92.Campbell BJ, Waidner LA, Cottrell MT, Kirchman DL. 2008. Abundant proteorhodopsin genes in the North Atlantic Ocean. Environ Microbiol 10:99–109. doi: 10.1111/j.1462-2920.2007.01436.x. [DOI] [PubMed] [Google Scholar]
- 93.Riedel T, Tomasch J, Buchholz I, Jacobs J, Kollenberg M, Gerdts G, Wichels A, Brinkhoff T, Cypionka H, Wagner-Döbler I. 2010. Constitutive expression of the proteorhodopsin gene by a flavobacterium strain representative of the proteorhodopsin-producing microbial community in the North Sea. Appl Environ Microbiol 76:3187–3197. doi: 10.1128/AEM.02971-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sabehi G, Massana R, Bielawski JP, Rosenberg M, DeLong EF, Béjà O. 2003. Novel proteorhodopsin variants from the Mediterranean and Red Seas. Environ Microbiol 5:842–849. doi: 10.1046/j.1462-2920.2003.00493.x. [DOI] [PubMed] [Google Scholar]
- 95.Béjà O, Suzuki MT. 2008. Photoheterotrophic marine prokaryotes, p 131–157. In Kirchman DL. (ed), Microbial ecology of the oceans. John Wiley & Sons, Inc., New York, NY. [Google Scholar]
- 96.Béjà O, Spudich EN, Spudich JL, Leclerc M, DeLong EF. 2001. Proteorhodopsin phototrophy in the ocean. Nature 411:786–789. doi: 10.1038/35081051. [DOI] [PubMed] [Google Scholar]
- 97.Man D, Wang W, Sabehi G, Aravind L, Post AF, Massana R, Spudich EN, Spudich JL, Béjà O. 2003. Diversification and spectral tuning in marine proteorhodopsins. EMBO J 22:1725–1731. doi: 10.1093/emboj/cdg183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Man-Aharonovich D, Sabehi G, Sineshchekov OA, Spudich EN, Spudich JL, Béjà O. 2004. Characterization of RS29, a blue-green proteorhodopsin variant from the Red Sea. Photoch Photobio Sci 3:459–462. doi: 10.1039/B316071H. [DOI] [PubMed] [Google Scholar]
- 99.Kim SY, Waschuk SA, Brown LS, Jung KH. 2008. Screening and characterization of proteorhodopsin color-tuning mutations in Escherichia coli with endogenous retinal synthesis. Biochim Biophys Acta 1777:504–513. doi: 10.1016/j.bbabio.2008.03.010. [DOI] [PubMed] [Google Scholar]
- 100.Yamada K, Kawanabe A, Kandori H. 2010. Importance of alanine at position 178 in proteorhodopsin for absorption of prevalent ambient light in the marine environment. Biochemistry 49:2416–2423. doi: 10.1021/bi9020204. [DOI] [PubMed] [Google Scholar]
- 101.Sabehi G, Kirkup BC, Rozenberg M, Stambler N, Polz MF, Béjà O. 2007. Adaptation and spectral tuning in divergent marine proteorhodopsins from the eastern Mediterranean and the Sargasso Seas. ISME J 1:48–55. doi: 10.1038/ismej.2007.10. [DOI] [PubMed] [Google Scholar]
- 102.Stingl U, Tripp HJ, Giovannoni SJ. 2007. Improvements of high-throughput culturing yielded novel SAR11 strains and other abundant marine bacteria from the Oregon coast and the Bermuda Atlantic Time Series study site. ISME J 1:361–371. [DOI] [PubMed] [Google Scholar]
- 103.Philosof A, Béjà O. 2013. Bacterial, archaeal and viral-like rhodopsins from the Red Sea. Environ Microbiol Rep 5:475–482. doi: 10.1111/1758-2229.12037. [DOI] [PubMed] [Google Scholar]
- 104.Zhao M, Chen F, Jiao N. 2009. Genetic diversity and abundance of flavobacterial proteorhodopsin in China seas. Appl Environ Microbiol 75:529–533. doi: 10.1128/AEM.01114-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Cottrell MT, Kirchman DL. 2009. Photoheterotrophic microbes in the Arctic Ocean in summer and winter. Appl Environ Microbiol 75:4958–4966. doi: 10.1128/AEM.00117-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Nguyen D, Maranger R, Balagué V, Coll-Lladó M, Lovejoy C, Pedrós-Alió C. 2015. Winter diversity and expression of proteorhodopsin genes in a polar ocean. ISME J 9:1835–1845. doi: 10.1038/ismej.2015.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Rich VI, Pham VD, Eppley J, Shi YM, DeLong EF. 2011. Time-series analyses of Monterey Bay coastal microbial picoplankton using a ‘genome proxy’ microarray. Environ Microbiol 13:116–134. doi: 10.1111/j.1462-2920.2010.02314.x. [DOI] [PubMed] [Google Scholar]
- 108.Frias-Lopez J, Shi Y, Tyson GW, Coleman ML, Schuster SC, Chisholm SW, DeLong EF. 2008. Microbial community gene expression in ocean surface waters. Proc Natl Acad Sci U S A 105:3805–3810. doi: 10.1073/pnas.0708897105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Poretsky RS, Hewson I, Sun SL, Allen AE, Zehr JP, Moran MA. 2009. Comparative day/night metatranscriptomic analysis of microbial communities in the North Pacific subtropical gyre. Environ Microbiol 11:1358–1375. doi: 10.1111/j.1462-2920.2008.01863.x. [DOI] [PubMed] [Google Scholar]
- 110.Shi YM, Tyson GW, Eppley JM, DeLong EF. 2011. Integrated metatranscriptomic and metagenomic analyses of stratified microbial assemblages in the open ocean. ISME J 5:999–1013. doi: 10.1038/ismej.2010.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Satinsky BM, Crump BC, Smith CB, Sharma S, Zielinski BL, Doherty M, Meng J, Sun S, Medeiros PM, Paul JH, Coles VJ, Yager PL, Moran MA. 2014. Microspatial gene expression patterns in the Amazon River Plume. Proc Natl Acad Sci U S A 111:11085–11090. doi: 10.1073/pnas.1402782111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Lami R, Cottrell MT, Campbell BJ, Kirchman DL. 2009. Light-dependent growth and proteorhodopsin expression by Flavobacteria and SAR11 in experiments with Delaware coastal waters. Environ Microbiol 11:3201–3209. doi: 10.1111/j.1462-2920.2009.02028.x. [DOI] [PubMed] [Google Scholar]
- 113.Lami R, Kirchman DL. 2014. Diurnal expression of SAR11 proteorhodopsin and 16S rRNA genes in coastal North Atlantic waters. Aquat Microb Ecol 73:185–194. doi: 10.3354/ame01716. [DOI] [Google Scholar]
- 114.Gifford S, Sharma AK, Booth M, Moran MA. 2013. Expression patterns reveal niche diversification in a marine microbial assemblage. ISME J 7:281–298. doi: 10.1038/ismej.2012.96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Gifford SM, Sharma S, Moran MA. 2014. Linking activity and function to ecosystem dynamics in a coastal bacterioplankton community. Front Microbiol 5:185. doi: 10.3389/fmicb.2014.00185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Ottesen EA, Young CR, Eppley JM, Chavez FP, Scholin CA, DeLong EF. 2013. Pattern and synchrony of gene expression among sympatric marine microbial populations. Proc Natl Acad Sci U S A 110:E488–E497. doi: 10.1073/pnas.1222099110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Palovaara J, Akram N, Baltar F, Bunse C, Forsberg J, Pedrós-Alió C, González JM, Pinhassi J. 2014. Stimulation of growth by proteorhodopsin phototrophy involves regulation of central metabolic pathways in marine planktonic bacteria. Proc Natl Acad Sci U S A 111:E3650–E3658. doi: 10.1073/pnas.1402617111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Aylward FO, Eppley JM, Smith JM, Chavez FP, Scholin CA, DeLong EF. 2015. Microbial community transcriptional networks are conserved in three domains at ocean basin scales. Proc Natl Acad Sci U S A 112:5443–5448. doi: 10.1073/pnas.1502883112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Ottesen EA, Young CR, Gifford SM, Eppley JM, Marin R III, Schuster SC, Scholin CA, DeLong EF. 2014. Multispecies diel transcriptional oscillations in open ocean heterotrophic bacterial assemblages. Science 345:207–212. doi: 10.1126/science.1252476. [DOI] [PubMed] [Google Scholar]
- 120.Mary I, Tarran GA, Warwick PE, Terry MJ, Scanlan DJ, Burkill PH, Zubkov MV. 2008. Light enhanced amino acid uptake by dominant bacterioplankton groups in surface waters of the Atlantic Ocean. FEMS Microbiol Ecol 63:36–45. doi: 10.1111/j.1574-6941.2007.00414.x. [DOI] [PubMed] [Google Scholar]
- 121.Gómez-Pereira PR, Hartmann M, Grob C, Tarran GA, Matin AP, Fuchs BM, Scanlan DJ, Zubkov MV. 2013. Comparable light stimulation of organic nutrient uptake by SAR11 and Prochlorococcus in the North Atlantic subtropical gyre. ISME J 7:603–614. doi: 10.1038/ismej.2012.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Morris RM, Nunn BL, Frazar C, Goodlett DR, Ting YS, Rocap G. 2010. Comparative metaproteomics reveals ocean-scale shifts in microbial nutrient utilization and energy transduction. ISME J 4:673–685. doi: 10.1038/ismej.2010.4. [DOI] [PubMed] [Google Scholar]
- 123.Sowell SM, Abraham PE, Shah M, Verberkmoes NC, Smith DP, Barofsky DF, Giovannoni SJ. 2011. Environmental proteomics of microbial plankton in a highly productive coastal upwelling system. ISME J 5:856–865. doi: 10.1038/ismej.2010.168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Steindler L, Schwalbach MS, Smith DP, Chan F, Giovannoni SJ. 2011. Energy starved Candidatus Pelagibacter ubique substitutes light-mediated ATP production for endogenous carbon respiration. PLoS One 6:e19725. doi: 10.1371/journal.pone.0019725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Gärtner W, Losi A. 2003. Crossing the borders: archaeal rhodopsins go bacterial. Trends Microbiol 11:405–407. doi: 10.1016/S0966-842X(03)00203-8. [DOI] [PubMed] [Google Scholar]
- 126.González JM, Pinhassi J, Fernández-Gómez B, Coll-Lladó M, González-Velázquez M, Puigbò P, Jaenicke S, Gómez-Consarnau L, Fernàndez-Guerra A, Goesmann A, Pedrós-Alió C. 2011. Genomics of the proteorhodopsin-containing marine flavobacterium Dokdonia sp. strain MED134. Appl Environ Microbiol 77:8676–8686. doi: 10.1128/AEM.06152-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Martinez A, Bradley AS, Waldbauer JR, Summons RE, DeLong EF. 2007. Proteorhodopsin photosystem gene expression enables photophosphorylation in a heterologous host. Proc Natl Acad Sci U S A 104:5590–5595. doi: 10.1073/pnas.0611470104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Güell M, Yus E, Lluch-Senar M, Serrano L. 2011. Bacterial transcriptomics: what is beyond the RNA horiz-ome? Nat Rev Microbiol 9:658–669. doi: 10.1038/nrmicro2620. [DOI] [PubMed] [Google Scholar]
- 129.Rogozin IB, Makarova KS, Murvai J, Czabarka E, Wolf YI, Tatusov RL, Szekely LA, Koonin EV. 2002. Connected gene neighborhoods in prokaryotic genomes. Nucleic Acids Res 30:2212–2223. doi: 10.1093/nar/30.10.2212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Lawrence JG. 2002. Shared strategies in gene organization among prokaryotes and eukaryotes. Cell 110:407–413. doi: 10.1016/S0092-8674(02)00900-5. [DOI] [PubMed] [Google Scholar]
- 131.Jin DJ, Cabrera JE. 2006. Coupling the distribution of RNA polymerase to global gene regulation and the dynamic structure of the bacterial nucleoid in Escherichia coli. J Struct Biol 156:284–291. doi: 10.1016/j.jsb.2006.07.005. [DOI] [PubMed] [Google Scholar]
- 132.Woyke T, Xie G, Copeland A, González JM, Han C, Kiss H, Saw JH, Senin P, Yang C, Chatterji S, Cheng JF, Eisen JA, Sieracki ME, Stepanauskas R. 2009. Assembling the marine metagenome, one cell at a time. PLoS One 4:e5299. doi: 10.1371/journal.pone.0005299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Wilhelm LJ, Tripp HJ, Givan SA, Smith DP, Giovannoni SJ. 2007. Natural variation in SAR11 marine bacterioplankton genomes inferred from metagenomic data. Biol Direct 2:27. doi: 10.1186/1745-6150-2-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.González JM, Fernández-Gómez B, Fernàndez-Guerra A, Gómez-Consarnau L, Sánchez O, Coll-Lladó M, del Campo J, Escudero L, Rodríguez-Martínez R, Alonso-Sáez L, Latasa M, Paulsen I, Nedashkovskaya OI, Lekunberri I, Pinhassi J, Pedrós-Alió C. 2008. Genome analysis of the proteorhodopsin-containing marine bacterium Polaribacter sp. MED152 (Flavobacteria). Proc Natl Acad Sci U S A 105:8724–8729. doi: 10.1073/pnas.0712027105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kimura H, Young CR, Martinez A, DeLong EF. 2011. Light-induced transcriptional responses associated with proteorhodopsin-enhanced growth in a marine flavobacterium. ISME J 5:1641–1651. doi: 10.1038/ismej.2011.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Fuhrman JA, Schwalbach MS, Stingl U. 2008. Proteorhodopsins: an array of physiological roles? Nat Rev Microbiol 6:488–494. doi: 10.1038/nrmicro1893. [DOI] [PubMed] [Google Scholar]
- 137.Kirchman DL, Hanson TE. 2013. Bioenergetics of photoheterotrophic bacteria in the oceans. Environ Microbiol Rep 5:188–199. doi: 10.1111/j.1758-2229.2012.00367.x. [DOI] [PubMed] [Google Scholar]
- 138.Pushkarev A, Béjà O. 2016. Functional metagenomic screen reveals new and diverse microbial rhodopsins. ISME J doi: 10.1038/ismej.2016.1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Walter JM, Greenfield D, Bustamante C, Liphardt J. 2007. Light-powering Escherichia coli with proteorhodopsin. Proc Natl Acad Sci U S A 104:2408–2412. doi: 10.1073/pnas.0611035104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Wang Y, Li Y, Xu T, Shi ZY, Wu Q. 2015. Experimental evidence for growth advantage and metabolic shift stimulated by photophosphorylation of proteorhodopsin expressed in Escherichia coli at anaerobic condition. Biotechnol Bioeng 112:947–956. doi: 10.1002/bit.25504. [DOI] [PubMed] [Google Scholar]
- 141.Hunt KA, Flynn JM, Naranjo B, Shikhare ID, Gralnick JA. 2010. Substrate-level phosphorylation is the primary source of energy conservation during anaerobic respiration of Shewanella oneidensis strain MR-1. J Bacteriol 192:3345–3351. doi: 10.1128/JB.00090-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Wang Z, O'Shaughnessy TJ, Soto CM, Rahbar AM, Robertson KL, Lebedev N, Vora GJ. 2012. Function and regulation of Vibrio campbellii proteorhodopsin: acquired phototrophy in a classical organoheterotroph. PLoS One 7:e38749. doi: 10.1371/journal.pone.0038749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Courties A, Riedel T, Rapaport A, Lebaron P, Suzuki MT. 2015. Light-driven increase in carbon yield is linked to maintenance in the proteorhodopsin-containing Photobacterium angustum S14. Front Microbiol 6:688. doi: 10.3389/fmicb.2015.00688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Kirchman DL. 2003. The contribution of monomers and other low-molecular weight compounds to the flux of dissolved organic matter in aquatic ecosystems, p 217–241. In Findlay SEG, Sinsabaugh RL (ed), Aquatic ecosystems: interactivity of dissolved organic matter. Elsevier Science, New York, NY. [Google Scholar]
- 145.Fernández-Gómez B. 2012. Ecology of marine Bacteroidetes: a genomics approach. Ph.D. thesis Universidad de Las Palmas de Gran Canaria, Las Palmas de Gran Canaria, Spain. [Google Scholar]
- 146.Feng S, Powell SM, Wilson R, Bowman JP. 2013. Light-stimulated growth of proteorhodopsin-bearing sea-ice psychrophile Psychroflexus torquis is salinity dependent. ISME J 7:2206–2213. doi: 10.1038/ismej.2013.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Feng S, Powell SM, Wilson R, Bowman JP. 2015. Proteomic insight into functional changes of proteorhodopsin-containing bacterial species Psychroflexus torquis under different illumination and salinity levels. J Proteome Res 14:3848–3858. doi: 10.1021/acs.jproteome.5b00241. [DOI] [PubMed] [Google Scholar]
- 148.Michelou VK, Rappé MS. 2014. The effect of light on the physiology of HIMB30, a proteorhodopsin-containing marine gammaproteobacterium possessing the genetic machinery for carbon fixation, poster 69. ASLO Ocean Sci Meet, Honolulu, HI, USA. [Google Scholar]
- 149.Fernández-Gómez B, Richter M, Schuler M, Pinhassi J, Acinas SG, González JM, Pedrós-Alió C. 2013. Ecology of marine Bacteroidetes: a comparative genomics approach. ISME J 7:1026–1037. doi: 10.1038/ismej.2012.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Gómez-Consarnau L, González JM, Riedel T, Jaenicke S, Wagner-Döbler I, Sañudo-Wilhelmy SA, Fuhrman JA. 2016. Proteorhodopsin light-enhanced growth linked to vitamin-B1 acquisition in marine Flavobacteria. ISME J 10:1102–1112. doi: 10.1038/ismej.2015.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Kang I, Oh HM, Lim SI, Ferriera S, Giovannoni SJ, Cho JC. 2010. Genome sequence of Fulvimarina pelagi HTCC2506(T), a Mn(II)-oxidizing alphaproteobacterium possessing an aerobic anoxygenic photosynthetic gene cluster and xanthorhodopsin. J Bacteriol 192:4798–4799. doi: 10.1128/JB.00761-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Lin S, Zhang H, Zhuang Y, Tran B, Gill J. 2010. Spliced leader-based metatranscriptomic analyses lead to recognition of hidden genomic features in dinoflagellates. Proc Natl Acad Sci U S A 107:20033–20038. doi: 10.1073/pnas.1007246107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Shi X, Li L, Guo C, Lin X, Li M, Lin S. 2015. Rhodopsin gene expression regulated by the light dark cycle, light spectrum and light intensity in the dinoflagellate Prorocentrum. Front Microbiol 6:555. doi: 10.3389/fmicb.2015.00555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Marchetti A, Schruth DM, Durkin CA, Parker MS, Kodner RB, Berthiaume CT, Morales R, Allen AE, Armbrust EV. 2012. Comparative metatranscriptomics identifies molecular bases for the physiological responses of phytoplankton to varying iron availability. Proc Natl Acad Sci U S A 109:E317–E325. doi: 10.1073/pnas.1118408109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Marchetti A, Catlett D, Hopkinson BM, Ellis K, Cassar N. 2015. Marine diatom proteorhodopsins and their potential role in coping with low iron availability. ISME J 9:2745–2748. doi: 10.1038/ismej.2015.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Yutin N, Koonin EV. 2012. Proteorhodopsin genes in giant viruses. Biol Direct 7:34. doi: 10.1186/1745-6150-7-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Alonso-Sáez L, Gasol JM, Lefort T, Hofer J, Sommaruga R. 2006. Effect of natural sunlight on bacterial activity and differential sensitivity of natural bacterioplankton groups in northwestern Mediterranean coastal waters. Appl Environ Microbiol 72:5806–5813. doi: 10.1128/AEM.00597-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Church MJ, Ducklow HW, Karl DM. 2004. Light dependence of [3H]leucine incorporation in the oligotrophic North Pacific Ocean. Appl Environ Microbiol 70:4079–4087. doi: 10.1128/AEM.70.7.4079-4087.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Straza TRA, Kirchman DL. 2011. Single-cell response of bacterial groups to light and other environmental factors in the Delaware Bay, USA. Aquat Microb Ecol 62:267–277. doi: 10.3354/ame01469. [DOI] [Google Scholar]
- 160.Michelou VK, Cottrell MT, Kirchman DL. 2007. Light-stimulated bacterial production and amino acid assimilation by cyanobacteria and other microbes in the North Atlantic Ocean. Appl Environ Microbiol 73:5539–5546. doi: 10.1128/AEM.00212-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Schwalbach MS, Brown M, Fuhrman JA. 2005. Impact of light on marine bacterioplankton community structure. Aquat Microb Ecol 39:235–245. doi: 10.3354/ame039235. [DOI] [Google Scholar]
- 162.Ruiz-González C, Simó R, Sommaruga R, Gasol JM. 2013. Away from darkness: a review on the effects of solar radiation on heterotrophic bacterioplankton activity. Front Microbiol 4:131. doi: 10.3389/fmicb.2013.00131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Atamna-Ismaeel N, Sabehi G, Sharon I, Witzel KP, Labrenz M, Jürgens K, Barkay T, Stomp M, Huisman J, Béjà O. 2008. Widespread distribution of proteorhodopsins in freshwater and brackish ecosystems. ISME J 2:656–662. doi: 10.1038/ismej.2008.27. [DOI] [PubMed] [Google Scholar]
- 164.Sharma AK, Zhaxybayeva O, Papke RT, Doolittle WF. 2008. Actinorhodopsins: proteorhodopsin-like gene sequences found predominantly in non-marine environments. Environ Microbiol 10:1039–1056. doi: 10.1111/j.1462-2920.2007.01525.x. [DOI] [PubMed] [Google Scholar]
- 165.Koh EY, Atamna-Ismaeel N, Martin A, Cowie ROM, Béjà O, Davy SK, Maas EW, Ryan KG. 2010. Proteorhodopsin-bearing bacteria in Antarctic sea ice. Appl Environ Microbiol 76:5918–5925. doi: 10.1128/AEM.00562-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Bohorquez LC, Ruiz-Pérez CA, Zambrano MM. 2012. Proteorhodopsin-like genes present in thermoacidophilic high-mountain microbial communities. Appl Environ Microbiol 78:7813–7817. doi: 10.1128/AEM.01683-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Atamna-Ismaeel N, Finkel OM, Glaser F, Sharon I, Schneider R, Post AF, Spudich JL, von Mering C, Vorholt JA, Iluz D, Béjà O, Belkin S. 2012. Microbial rhodopsins on leaf surfaces of terrestrial plants. Environ Microbiol 14:140–146. doi: 10.1111/j.1462-2920.2011.02554.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Bertsova YV, Bogachev AV, Skulachev VP. 2015. Proteorhodopsin from Dokdonia sp. PRO95 is a light-driven Na+-pump. Biochemistry 80:449–454. [DOI] [PubMed] [Google Scholar]
- 169.Levine JS, MacNichol EF Jr. 1982. Color vision in fishes. Sci Am 246:140–149. doi: 10.1038/scientificamerican0282-140. [DOI] [Google Scholar]