

Abstract
MicroRNAs (miRNAs) have been identified as a new class of regulatory molecules that influence many biological functions, including metabolism, adipocyte differentiation. To determine the role of adipogenic miRNAs in the adipocyte differentiation process, we used microarray technology to monitor miRNA levels in human adipose-derived mesenchymal stem cells (hMSCs-Ad), human stromal vascular cells (SVCs) and differentiated adipocytes. 79 miRNAs were found to be differentially expressed, most of which are located in obesity related chromosomal regions but have not been previously linked to adipocyte differentiation process. A systematic search was made for relevant studies in academic data bases, involving the Gene Expression Omnibus (GEO) ArrayExpress, Pubmed and Embase database. Eight studies on human adipocyte differentiation or obesity were included in the final analysis. After combining our microarray data with meta-analysis of published microarray data, we detected 42 differently expressed miRNAs (meta-signature miRNAs) in mature adipocytes compared to SVCs or hMSCs-Ad. Our study shows meta-signature miRNAs specific for adipogenesis, several of which are correlated with key gene targets demonstrating functional relationships to pathways in BMP signaling pathway, Cell differentiation, Wnt signaling, insulin receptor signaling pathway, MAPK signaling, Cell cycle and lipid metabolic process. Our study shows that the first evidence of hsa-let-7 family, hsa-miR-15a-5p, hsa-miR-27a-3p, hsa-miR-106b-5p, hsa-miR-148a-3p and hsa-miR-26b-5p got a great weight in adipogenesis. We concluded that meta-signature miRNAs involved in adipocyte differentiation and provided pathophysiological roles and novel insight into obesity and its related metabolic diseases.
Keywords: miRNA, adipocyte, adipogenesis, meta-analysis, obesity
INTRODUCTION
Obesity has become a major worldwide health problem associated with metabolic and cardiovascular disorders. Obesity is a progressive and multifactorial pathology characterized by excessive expansion of white adipose tissue, which due to an increase in adipocyte hyperplasia and hypertrophy [1]. Since obesity-related hyperplasia and hypertrophy are often associated with various metabolic disorders, proper understanding of the molecular events regulating adipogenesis and lipid metabolism can provide valuable information for exploiting comprehensive and effective therapeutic strategies against obesity.
MicroRNAs (miRNAs), small non-coding RNAs of 18–25 nucleotides in length, which were seen to control gene expression in virtually obesity or other metabolism syndrome, were abundantly investigated relating to proliferation and differentiation of adipogenesis. Indeed, the role of some miRNAs has been described in obesity. For example, miR-103, miR-107, and miR-143 regulate liver insulin sensitivity in diet-induced obesity through different mechanisms [2, 3]. The involvement of miRNAs in obesity pathogenesis is well established, as they can behave as pro-adipogenic or adipogenic suppressor genes depending on the cellular function of their targets [4].
Emerging evidence shows that miRNAs may directly or indirectly modulate adipocyte differentiation. In our previous study, we found that miR-148a, as a biomarker of obesity, promoted adipogenesis by inhibiting Wnt1 [5]. miR-146b as a positive regulator of accelerated adipocyte differentiation through modulation of KLF7 [6]. Recent findings [7] have suggested that miR-26b inhibits adipogenic differentiation and overexpression of miR-1908 inhibited adipogenic differentiation [8]. However, to date, there is no meta-analysis to interpret the biology of miRNA and its contribution to obesity development which may provide early promising therapeutic targets and effective control of obesity.
In the present study, using a miRNA array strategy, we identified 79 differentially expressed miRNAs, most of which located in obesity related chromosomal regions but have not been previously linked to adipocyte differentiation process. Additionally, to overcome the limitations in current researches, we performed meta-analysis applying the robust rank aggregation method [9], followed by pathway analysis, to identify miRNA regulation in adipogenesis and the pathways that key miRNAs may impact. Identification of miRNA meta-signature and involved pathways would provide the potential target and novel insight into obesity and its related metabolic diseases.
RESULTS
miRNA microarray results
miRNA microarray analysis was performed to identify differentially expressed miRNAs in mature adipocytes and then compared to SVCs and hMSCs-Ad. A larger number of microRNAs (79 miRNAs, 45 up-regulated miRNAs and 34 down-regulated miRNAs) were significantly up- or down-regulated in mature adipocytes compared to SVCs and hMSCs-Ad (Figure 1, Supplementary Table 1). Our data showed that differentially expressed miRNAs were overlapping either in mature adipocytes versus SVCs or in mature adipocytes versus hMSCs-Ad (Supplementary Table 1, P< 0.05). Between mature adipocytes and SVCs/hMSCs-Ad, miR-146b-5p and miR-335 were found to be differentially expressed with a fold change >10 (Supplementary Table 1). Whereas, miR-424 was differentially expressed with a fold change of more than 10 in mature adipocytes when compared to hMSCs-Ad, and the expression level was about 9.2-fold when compared to SVCs. Of all differentially expressed miRNAs, miR-1275, miR-155 and miR-1268 were down-regulated in mature adipocytes over 10-fold when compared with SVCs and hMSCs-Ad. We identified that miR-148a, miR-26b, miR-132, miR-365 and miR-1908 were highly expressed in mature adipocytes with over 5-fold compared to SVCs/hMSCs-Ad. Additionally, miR-93 and miR-720 were lowly expressed with 5-fold in mature adipocytes compared to SVCs/hMSCs-Ad. Among these differentially expressed miRNAs, our group did further research for the adipogenic miRNAs in vivo and in vitro, including miR-148a [5], miR-26b [10], miR-146b [6] and miR-1275.
Figure 1. Heatmap of results of microRNA microarray analysis.

Up-regulated microRNAs are shown in red, and downregulated microRNAs are shown in green. The fold-changes (log2 transformed) of miRNA expression in differentiated versus SVCs or undifferentiated hMSCs-A. Each group (A1-A4, PA1-PA4 and M1-M4) is pooled from three samples. A: mature adipocyte; SVCs (PA: preadipocyte): human stromal vascular cells; M: undifferentiated hMSCs-Ad.
PCR validation of significant differentially expressed miRNAs expression
To confirm the microarray hybridization results, qRT-PCR was performed on 8 up-regulated miRNAs (miR-335, miR-146b-5p, miR-26b, miR-30b, miR-21, miR-378, miR-143 and miR-148a) [10] and 3 down-regulated miRNA (miR-155,miR-221, and miR-1275), chosen on the basis of their levels of expression on the microarray and their biological significance. qRT-PCR was performed in triplicate on the diluted cDNA and the experiments were conducted twice on differentiated hMSCs-Ad/mature adipocytes and hMSCs-Ad/SVCs. The qRT-PCR results were closely mirrored with the microarray analysis showing P-value <0.05 (Figure 2).
Figure 2. mRNAs expression in mature adipocytes versus SVCs or hMSCs-Ad.SVCs or hMSCs-Ad was grown to confluence and adipogenic differentiation was initiated as described in Research Design and Methods.

Expression of miRNAs during adipocyte differentiation was quantitated by TaqMan miR-based qRT-PCR. The bar is the mean ± SD of four independent experiments that were analyzed using Independent T-Test vs. undifferentiated SVCs or hMSCs-Ad. SVCs: human stromal vascular cells; hMSCs-Ad: undifferentiated hMSC-Ad; Ad: mature adipocyte.
The location of differentially expressed miRNAs in obesity gene map
To elucidate the correlation pattern between miRNA and obesity, the chromosomal regions of differentially expressed miRNAs were performed using Genomic map view analysis. We compared the chromosomal regions of differentially expressed miRNAs with obesity gene map published in 2005 [11]. As shown in Table 1, there were about 65% (51/79) miRNAs located in obesity related chromosomal regions. Our results indicated that these differentially expressed miRNAs have a close relationship with obesity.
Table 1. Chromosome of Adipocytes specific miRNAs distribution compared with obese gene map.
| miRNA Name | Localization | Population | Phenotypes |
|---|---|---|---|
| hsa-miR-34a | 1p36.22 | >168 pairs | Skinfolds, uprailiac |
| hsa-miR-128-1 | 2q21.3 | 453 subjects, 99 families | Abdominal visceral fat |
| hsa-miR-26b | 2q35 | 453 subjects, 99 families | Abdominal visceral fat |
| hsa-miR-128-2 | 3p22.3 | 377 pairs | Body fat (%) |
| hsa-miR-26a-1 | 3p22.2 | 580 families | BMI |
| hsa-miR-143 | 5q32 | 453 subjects, 99 families | Abdominal total fat |
| hsa-miR-145 | 5q32 | 453 subjects, 99 families | Abdominal total fat |
| hsa-miR-378 | 5q32 | 453 subjects, 99 families | Abdominal total fat |
| hsa-miR-25 | 7q22.1 | 261 subjects, 27 pedigrees, 1297 subjects, 260 families | High-density lipoprotein, in triglyceridesBMI |
| hsa-miR-93 | |||
| hsa-miR-106b | |||
| hsa-miR-335 | 7q32.2 | 545 pairs | BMI |
| hsa-miR-29a | 7q32.3 | 3027 subjects, 401 families, 317 sibships | BMI |
| hsa-miR-320a | 8p21.3 | 994 subjects, 37 pedigrees | BMI |
| hsa-miR-1915 | 10p12.31 | 667 subjects, 244 families; 862 subjects, 170 families |
Obesity (in whites and African Americans) Obesity (inAfrican Americans, in European Americans) |
| hsa-miR-605 | 10q21.1 | 672 subjects, 28 pedigrees; 862 subjects, 170 families |
Leptin, Obesity |
| hsa-miR-146b | 10q24.32 | 447 subjects, 109 pedigrees | BMI > 27 |
| hsa-miR-130a | 11q12.1 | 369 subjects, 89 families | Obesity (in white children and adolescents) |
| hsa-miR-125b-1 | 11q24.1 | 430 subjects, 27 sibpairs, 27 pedigrees;1526 pairs;1778 sibships;994 subjects, 37 pedigrees | BMI |
| hsa-let-7a-2 | |||
| hsa-miR-100 | |||
| hsa-miR-196a-2 | 12q13.13 | 729 subjects, 275 families | Waist-to-hip ratio |
| hsa-miR-26a-2 | 12q14.1 | 514 subjects, 99 families, 347 sibships | Fat intake |
| hsa-let-7i | |||
| hsa-miR-16-1 | 13q14.2 | 3027 subjects, 401 families, 317 sibships | BMI |
| hsa-miR-17 | 13q31.3 | 1297 subjects, 260 families; 1297 subjects, 260 families; 1312 subjects, 696 families | BMI, paternal, Waist circumference, paternal; BMI; Factor central obesity |
| hsa-miR-18a | |||
| hsa-miR-20a | |||
| hsa-miR-19b-1 | |||
| hsa-miR-92a-1 | |||
| hsa-miR-1260 | 14q24.3 | 672 subjects, 28 pedigrees | Leptin |
| hsa-miR-1469 | 15q26.2 | 336 sibpairs, 609 relative pairs | Fat-free mass |
| hsa-miR-365-1 | 16p13.12 | 893 sibpairs | BMI (in whites) |
| hsa-miR-22 | 17p13.3 | 729 subjects, 275 families | Abdominal subcutaneous fat |
| hsa-miR-132 | 17p13.3 | ||
| hsa-miR-195 | 17p13.1 | 478 subjects, 10 families | asp levels |
| hsa-miR-423 | 17q11.2 | 470 subjects, 10 families | BMI |
| hsa-miR-365-2 | 17q11.2 | ||
| hsa-miR-152 | 17q21.32 | 521 subjects, 156 families | Abdominal subcutaneous fat |
| hsa-miR-196a-1 | 17q21.32 | ||
| hsa-miR-638 | 19p13.2 | 522 subjects, 99 families, 364 sibpairs | Skinfolds, sum of eight (in whites), Leptin (in whites), Body fat (%) (in whites) |
| hsa-miR-199a-1-5p | 19p13.2 | ||
| hsa-miR-199a-1-3p | 19p13.2 | ||
| hsa-miR-24-2 | 19p13.13 | 506 subjects, 115 pedigrees | Age adiposity rebound |
| hsa-miR-27a | 19p13.13 | ||
| hsa-miR-23a | 19p13.13 | ||
| hsa-miR-155 | 21q21.3 | 1510 subjects, 509 families | Factor central obesity |
| hsa-let-7a-3 | 22q13.31 | >168 pairs | Body weight |
| hsa-let-7b | 22q13.31 | ||
| hsa-miR-221 | Xp11.3 | 1148 subjects, 133 families, 190 European-American families (940 members), 43 African-American families (208 members) | Waist-to-hip ratio (in European Americans and African Americans) |
| hsa-miR-222 |
Note: The Human Obesity Gene Map: The 2005 Update
Study selection and data extraction
We retrieved publications by the term of “adipocyte OR fat cell OR adipogenesis OR MSC (mesenchymal stem cells) OR obesity OR adipocyte differentiation” AND “miRNA OR microRNA” AND human”. Database searches initially yielded a total of 526 publications and 8 studies met the inclusion criteria (Figure 3, Table 2). Our research was included in the final analysis because the lists of ranked miRNA were suitable for the inclusion criteria. Most of the studies were published between 2010 and 2014. 3 of the researches came from America and 5 of the researches came from Europe.
Figure 3. Flowchart of study selection.
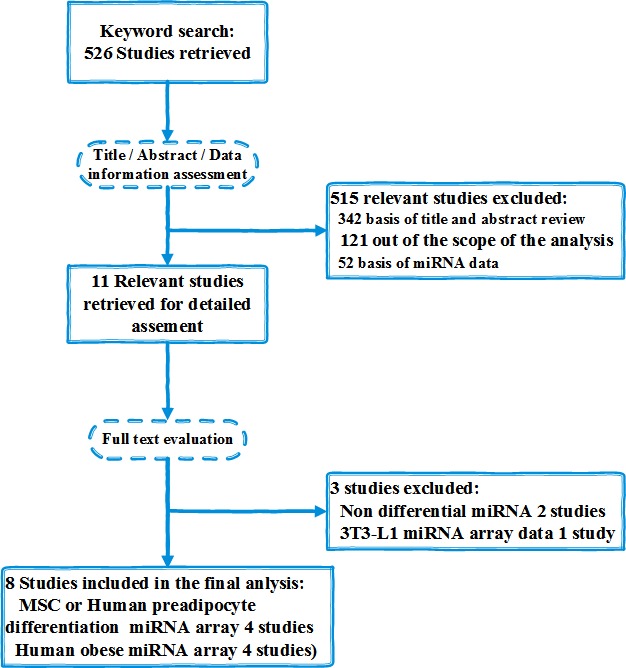
Table 2. Baseline characteristics of eligible studies.
| ID | link | panle | Group | Country | Fold change |
|---|---|---|---|---|---|
| [12] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/20057369 | GPL7162 CEINGE_Exiqon_Human miRNA Microarray [208002V8.1] | nondiabetic severely obese Vs. nonobese adults | Italy | No |
| [13] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/25518011 | GPL16384 [miRNA-3_0] Affymetrix Multispecies miRNA-3 Array | obese Vs. lean visceral adipocye exosomes | USA | Yes |
| [14] | http://www.ncbi.nlm.nih.gov/pubmed/22688341 | [miRNA-1_0] Affymetrix miRNA Array | obese (n = 30) Vs. nonobese (n = 26) | Sweden | Yes |
| [15] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/23493574 | GPL8786 [miRNA-1_0] Affymetrix miRNA Array | adipocytes derived from PCOS patients vs. control | USA | Yes |
| [18] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/20126310 | GPL7731 Agilent-019118 Human miRNA Microarray 2.0 G4470B | obese Vs. normal | Spain | Yes |
| [16] | http://www.ncbi.nlm.nih.gov/pubmed/25356868 | miRCURY LNA microRNA Array, 6th generation - hsa, mmu&rno [miRBase 17.0] | AD day 13 Vs AD day 0 adipocyte | Denmark | Yes |
| [17] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/21756067 | GPL11107 mirVana 1.3K v1.4 201205 | adipocyte differentiation | Norway | no |
| [19] | https://www.gcbi.com.cn/gclib/html/pubmed/detail/20887899# | GPL8180 Illumina Mouse v1 MicroRNA expression beadchip | adipocyte differentiation | USA | YES |
Differently expressed miRNAs in meta-publications
As shown in Supplementary Table 2, Martinelli et al. [12] found 41 differentially expressed miRNAs in subcutaneous adipose tissue (SAT) from nondiabetic severely obese (n = 20) and non-obese adults (n = 8). Ferrante et al. [13] developed techniques to quantify and characterize exosomes shed by adipocytes from seven obese (age: 12–17.5 y, BMI: 33–50 kg/m2) and five lean (age: 11–19 y, BMI: 22–25 kg/m2) subjects and found 89 differentially expressed miRNAs between obese and lean visceral adipocyte. Arner et al. [14] identified 20 differentially expressed miRNAs in intact adipose tissue from obese (n = 30) and non-obese (n = 26) subjects. Chen et al. [15] detected 28 differentially expressed miRNAs in adipocytes of PCOS (polycystic ovary syndrome) patients and matched control subjects. Alajez et al. [16]used telomerase immortalized human bone marrow-derived stromal cell line (hMSC-TERT) to identify the differentially expressed miRNA; they identified 36 up-regulated miRNAs and 2 down-regulated miRNAs during adipocytic differentiation. Skårn et al. [17] found 66 differentially expressed miRNAs in adipogenesis. Ortega et al. [18] isolated fat cells from both lean (BMI<25.0Kg/m2, n=3) and obese (BMI>30.0Kg/m2, n=3) subjects for miRNA array, and identified 5up-regulated miRNAs and 7 down-regulated miRNAs between lean and obese subjects. Mikkelsen et al. [19] got the data from hMSCs-Ad adipogenesis, which used the same cell line in our study, and detected 17 up-regulated miRNAs and 19 down-regulated miRNAs in mature adipocytes compared to hMSCs-Ad.
Our study found 79 miRNAs (45 up-regulated miRNAs and 34 down-regulated miRNAs) were significantly up- or down-regulated in mature adipocytes compared to SVCs and hMSCs-Ad (Figure 1, Supplementary Table 1). We detected 42 meta-signature miRNAs from these miRNA array data, which were differently expressed miRNAs in mature adipocytes compared to SVCs or MSC (Table 3). According to the sources of cells, we detected the meta-signature miRNAs from human adipocytes and MSC, respectively. We also identified 17 meta-signature differently expressed miRNAs in obese and lean subjects (Table 4).
Table 3. Adipocyte differentiation related meta-signature miRNAs.
| miRNA name | Our study typeD15 Vs D0 | other studies type | Reference ID |
|---|---|---|---|
| hsa-let-7a | UP | UP | [17] |
| hsa-let-7c | UP | UP | [17] |
| hsa-let-7d | UP | UP | [17] |
| hsa-let-7e | UP | UP | [17] |
| hsa-let-7f | UP | UP | [17] |
| hsa-miR-10b | UP | UP | [12] [16] |
| hsa-miR-1201 | UP | UP | [15] |
| hsa-miR-132 | UP | UP | [12] |
| hsa-miR-137 | UP | UP | [19] |
| hsa-miR-152 | UP | UP | [16] |
| hsa-miR-154 | UP | UP | [15] |
| hsa-miR-15a | UP | UP | [16] |
| hsa-miR-181a | UP | UP | [12),[17] |
| hsa-miR-193b* | UP | UP | [16] |
| hsa-miR-26a | UP | UP | [16), [17] |
| hsa-miR-26b | UP | UP | [16] |
| hsa-miR-30a | UP | UP | [16),[19] |
| hsa-miR-30a* | UP | UP | [19] |
| hsa-miR-30b | UP | UP | [16] |
| hsa-miR-30c | UP | UP | [16] |
| hsa-miR-30d | UP | UP | [12] [15],[16],[19] |
| hsa-miR-365 | UP | UP | [16] |
| hsa-miR-374a | UP | UP | [16] |
| hsa-miR-374b | UP | UP | [16] |
| hsa-miR-378 | UP | UP | [19] |
| hsa-miR-378* | UP | UP | [19] |
| hsa-miR-498 | UP | UP | [12), [13] |
| hsa-miR-642 | UP | UP | [12), [19] |
| hsa-miR-663 | UP | UP | [12] |
| hsa-miR-18a | DOWN | DOWN | [12] |
| hsa-miR-27a | DOWN | DOWN | [17] |
| hsa-miR-92a | DOWN | DOWN | [17] |
| hsa-miR-106b | DOWN | DOWN | [17] |
| hsa-miR-130b | DOWN | DOWN | [17] |
| hsa-miR-155 | DOWN | DOWN | [13] |
| hsa-miR-221 | DOWN | DOWN | [17] |
| hsa-miR-222 | DOWN | DOWN | [13] |
| hsa-miR-218-2* | DOWN | DOWN | [13] |
| hsa-miR-494 | DOWN | DOWN | [15] |
| hsa-miR-629* | DOWN | DOWN | [13] |
| hsa-miR-758 | DOWN | DOWN | [13] |
| hsa-miR-1825 | DOWN | DOWN | [21] |
Table 4. Obese related meta-signature miRNAs.
| miRNA name | type D15 Vs D0 | obese vs normal | Reference ID |
|---|---|---|---|
| hsa-miR-92a | DOWN | DOWN | [14] |
| hsa-miR-758 | DOWN | DOWN | [13] |
| hsa-miR-663 | UP | UP | [12] |
| hsa-miR-642 | UP | UP | [12] |
| hsa-miR-629* | DOWN | DOWN | [13] |
| hsa-miR-498 | UP | UP | [12], [13] |
| hsa-miR-484 | DOWN | DOWN | [18] |
| hsa-miR-30d | UP | UP | [12] |
| hsa-miR-218-2* | DOWN | DOWN | [13] |
| hsa-miR-199a-5p | UP | UP | [18] |
| hsa-miR-185 | DOWN | DOWN | [18] |
| hsa-miR-181a | UP | UP | [12] |
| hsa-miR-132 | UP | UP | [12] |
| hsa-miR-130b | DOWN | DOWN | [18] |
| hsa-miR-10b | UP | UP | [12] |
| hsa-miR-106b | DOWN | DOWN | [12] |
| hsa-let-7i | DOWN | DOWN | [14] |
GO and pathway analysis
To determine which GO and pathways of meta-signature miRNAs are involved in adipogenesis, the predicted gene targets were compared against pathways in the Go and Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The predicted targets were found to be significantly over represented in cell differentiation, cell cycle, cell proliferation and Wnt signaling pathway of GO terms (Figure 4, Supplementary Table 3). The KEGG enrichment pathway analysis of our microarray data revealed that almost all of the meta-signature miRNAs were particularly involved in Wnt signaling, MAPK signaling, mTOR signaling and ECM-receptor interaction. Several genes in the Wnt signaling set (WNT4; LRP6; DAAM2; FRAT2;SKP1; PRKX; FZD3; FZD9; NFATC1; NFAT5; CHP; PPP3CA) were also targeted by multiple miRNAs as part of a complex regulatory network (Figure 5, Supplementary Table 4).
Figure 4. Bioinformatic analysis target genes of meta-signature miRNAs.

GeneCodis web tool was used to perform statistical analysis of over represented GO terms to predict target genes of meta-signature miRNAs. Cellular processes were sorted by score (−log [P value]). The highly positive score set included genes involved in cell differentiation, cell cycle, cell proliferation and Wnt signaling pathway of GO terms.
Figure 5. Pathway analyses for predicting target genes of meta-signature miRNAsin Kyoto Encyclopedia of Genes and Genomes (KEGG) database.

Almost all of the meta-signature miRNAs were particularly involved in Wnt signaling, MAPK signaling, mTOR signaling and ECM-receptor interaction.
miRNA-GO-NetWork
To identify which miRNAs play a critical role in adipogenesis, miRNA-Go-NetWork was analyzed by bioinformatics analysis. As shown in Fig 6, hsa-miR-15a-5p, hsa-miR-106b-5p, hsa-miR-181a-5p, hsa-let-7 family, hsa-miR-27a-3p, hsa-miR-130b -3p, hsa-miR-152/148a-3p and hsa-miR-26b-5p got the highest degree means, which indicated that these miRNAs had more weight in adipogenesis than others.
Figure 6. GO-NETwork miRNA-target gene network analysis.

Red box nodes represented miRNAs and green cycle nodes represented target genes. Edges indicated the inhibitive effect of miRNAs on target genes.
DISCUSSION
In this study, we reported the differential miRNA expression in mature adipocyte compared with SVCs or hMSCs-Ad and most of differentially expressed miRNAs located in obesity related chromosomal regions. The expression pattern of differential miRNAs between SVCs Vs. mature adipocyte and hMSCs-Ad Vs. mature adipocyte are similar, indicating that these differential miRNAs were indeed involved in adipogenesis. Our results are in line with the study from Alajez et.al. [16], describing 36 up-regulated miRNAs and 2 down-regulated miRNAs in mature adipocytes compared to hMSCs-TERT.
miR-146b-5p, miR-335, miR-424, miR-1275, miR-155, miR-1268, miR-148a, miR-26b, miR-132, miR-365, miR-1908, miR-93 and miR-720 were identified more than 3-fold changes in mature adipocyte compared with SVCs or hMSCs-Ad. More importantly, the highest fold change was observed in miR-146b in mature adipocyte. Ahn et al. reported that miR-146b identified as a positive modulator of adipocyte differentiation via suppression of sirtuin 1 (SIRT1) [20]. Interestingly, our previous study reported that miR-146b is highly expressed in the adipose tissue of obese mouse models such as DIO, diet-induced obesity, ob/ob, and db/db. This finding has implicated miR-146b as a positive regulator of accelerated adipocyte differentiation through modulation of SIRT1 and KLF7 [6]. Indeed, our previous study found that miR-148a promotes hMSCs-Ad to differentiate to mature adipocyte by targeting Wnt1 [5]. Additionally, our group [7] has also suggested that inhibited miR-26b inhibits adipogenic differentiation in human preadipocytes, and miR-335, as a potential adipogenic miRNA, is involved in adipose tissue inflammation and is highly expressed in mature 3T3-L1 [21]. Therefore, our present and previous studies bring in valuable information with respect to human obesity pathology because we have demonstrated that miR-148a, miR-146b, miR-26b and miR-335 are dysregulated in the process of adipocyte differentiation.
Since obesity-related hyperplasia and hypertrophy are often associated with various metabolic disorders, the detail understandings of the molecular events regulating adipogenesis and lipid metabolism are important. Ortega et al. [18] isolated fat cells from both lean (BMI<25.0Kg/m2, n=3) and obese (BMI>30.0Kg/m2, n=3) subjects for miRNA array and then differentiated to mature adipocyte, and identified 5 up-regulated miRNAs and 7 down-regulated miRNAs between lean and obese subjects. In addition, 65% (51/79) differentially expressed miRNAs from our data located in obesity related chromosomal regions which was published in 2005 (11), indicating that these differentially expressed miRNAs have a close relationship with obesity.
Although, miRNAs play an important role in the progression of obesity and adipogenesis, different microarray techniques, technical approaches, tissue or cellular models, and small sample size led to inconsistent findings in previous studies. Thus, the present observation reported that 42 differentially expressed miRNAs (meta-signature miRNAs) in mature adipocytes compared to SVCs or hMSCs-Ad. The number of meta-signature miRNAs studies reported varied greatly but at least 12 deregulated miRNAs were reported in each study. Ortega et al. [18] identified that miR-484 and miR-130b were lowly expressed in human subcutaneous adipose from obese and type 2 diabetes mellitus (DM), miR-199a-5p, miR-221, miR-125b and miR-1229 were lowly expressed, indicating that these miRNAs play a key role in obesity. However, this study identified miR-143 was down regulated in mature adipocyte, which conflicted with other studies [22]. Furthermore, this microarray data was not verified by qRT-PCR, thus, this result may be influenced by experimental error, individual difference and species variation.
The KEGG enrichment pathway analysis of our microarray data revealed that almost all of the meta-signature miRNAs were particularly involved in Wnt signaling, MAPK signaling, mTOR signaling and ECM-receptor interaction. Wnt signaling was first recognized as a possible negative regulator of adipogenesis when Wnt1 expression decreased significantly during adipocyte differentiation progress [23]. In particular, our study found that miR-148a through regulating Wnt1, but not Wnt10b, promotes hMSCs-Ad differentiation [5]. ERK, p38 and JNK signaling pathway belong to MAPK signaling, which are intracellular signaling pathways that play a pivotal role in many essential cellular processes such as proliferation and adipocyte differentiation [24]. Importantly, Esau et al. [25] reported that miR-143 promoted adipocyte differentiation by inhibiting ERK5. mTOR signaling belongs to the phosphoinositide 3-kinase (PI3K)-related kinase family, and promotes adipogenesis in white adipocytes [26]. Previous study reported that miR-17-92 cluster has been reported to be upregulated during the clonal expansion stage of adipocyte differentiation, positively regulating adipogenesis by targeting the tumor suppressor RB2/p130 [27] and knockdown of miR-106b and miR-93 significantly induced the expression of brown fat-specific genes and promoted the accumulation of lipid-droplet in differentiating brown adipocytes. In addition, ectopic expression of miR-106b and miR-93 suppressed the mRNA level of Ucp1 [28]. miR-93 is a highly expressed PCOS patients' adipocyte tissue and downregulates GLUT4 gene expression, suggesting that it plays a role in the IR of PCOS [15]. Interestingly, let-7-mediated repression of adipogenesis was also found to promote osteogenesis [29]. In the present study, we found that hsa-miR-15a-5p, hsa-miR-106b-5p, hsa-miR-181a-5p, hsa-let-7 family, hsa-miR-27a-3p, hsa-miR-130b-3p, hsa-miR-152/148a-3p and hsa-miR-26b-5p got high degree means, which indicated that these miRNAs had a great weight in adipogenesis than others.
We concluded that meta-signature miRNAs involved in adipocyte differentiation and provided pathophysiological roles and novel insight into obesity and its related metabolic diseases.
MATERIALS AND METHODS
Cell culture and adipocyte differentiation
Human stromal vascular cells (SVCs, also called “primary” preadipocytes, Cat.No.7210, ScienCell Research Laboratories, San Diego, CA) and Human adipose-derived mesenchymal stem cells (hMSCs-Ad; Cat.No.7510, ScienCell Research Laboratories, San Diego, CA) were maintained in Preadipocyte Medium (PAM) (Cat.No.7211, ScienCell Research Laboratories) and Mesenchymal Stem Cell Medium (MSCM) (Cat.No.7501, ScienCell Research Laboratories) supplemented with 5% fetal bovine serum, 1% mesenchymal stem cell growth supplement (Cat. No. 7501) and 1% penicillin/streptomycin solution at 37°C in a humidified atmosphere under 5% CO2. To induce differentiation, HMSC-Ad cells were incubated in serum-free MSCM supplemented with 50 nM insulin, 100 nM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine, and 100 μM rosiglitazone (Day 0) and the medium was replaced every 2 days over 4 days. Thereafter, cells were incubated in serum-free MSCM supplemented with 50 nM insulin and replaced every 2 days until lipid accumulated in cells.
miRNA microarray
RNA was labeled and hybridized to μParaflo® human miRNA microarrays following the manufacturer's instructions by LC Sciences (MRA-1001, miRHuman_14). Each chip contains 894 specific microRNA probes (repeated four times for each probe) and 50 controlled probes (according to the purpose of experiment, each probe is repeated 4-16 times). miRNA microarray analysis was performed using RNA samples from human preadipocyte, human adipose tissue-derived mesenchymal stem cells (hMSCs-Ad) and mature adipocyte.
RNA isolation and analysis of miRNAs expression by RT-quantitative PCR (qPCR)
Total RNA was prepared from SVCs or hMSCs-Ad at varying intervals after induction of adipocyte differentiation using TRIzol (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol, followed by DNase treatment (TaKaRa, Japan). The quality and concentration of RNA was assessed by Nanodrop2.0 (Thermo Fisher Scientific, Waltham, MA). 240 ng of total RNA was reverse transcribed using the high capacity reverse transcription kit (Applied Biosystems, Life Technologies) according to the manufacturer's protocol. One microliter of cDNA was then amplified by real-time qPCR using the Taqman universal PCR master mix (Applied Biosystems) and specific primers in an Applied 7500 QPCR System (Applied Biosystems). miRNA expression levels were normalized using the snoU6 miRNA expression level as internal controls. Each sample was measured in triplicate, and the gene expression levels were calculated using the 2−ΔΔct method.
Study selection and data extraction
A systematic literature search was performed for the identification of adipogenic or obesity miRNA expression profiling studies using a two-level search strategy. First, we undertook a web-based search in Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/) using search term (“adipocyte OR adipogenesis OR fat cell OR adipocyte differentiation OR obesity”[Mesh]) AND (“MicroRNAs”[Mesh]) AND “Humans”[Mesh]. To perform a comprehensive retrieval, searching in ArrayExpress (www.ebi.ac.uk/arrayexpress), Pubmed and Embase database were also performed. Second, the reference lists of all relevant and existing studies were reviewed through a manual search for further identification of potential relevant studies.
Abstracts were screened carefully and full texts of relevant potential abstracts were evaluated. Studies with original experimental design that analyzed the miRNA expression profiling in human between adipose tissues or human adipocytes were included. Meanwhile, studies were not eligible for meta-analysis if they met the following selection criteria: 1) using human preadipocytes or SVCs or hMSCs-Ad or hMSC cell, 2) preselected candidate genes research, 3) differentiating to mature adipocyte.
The lists of statistically significant expressed miRNAs were extracted from publications. Authors were contacted when the lists could not be obtained. All miRNA names were standardized through miRBase version 20.0. miRNAs that cannot be related to either −3p or −5p in miRBase were designated with hsa-miR-*, such as hsa-miR-210. Those identified as dead entry in miRBase retained their names mentioned in the literatures.
Integrative identification of miRNA targets
The meta-signature miRNAs were selected for target prediction by using TargetScan database version 6.2 [30] and miRanda (http://www.microrna.org) were used to predict the target consensus sequences for meta-miRNA.
Enrichment analysis
To identify the pathways of predicted miRNA targets, Kyoto Encyclopedia of Genes and Genomes (KEGG), Panther pathways and Gene Ontology terms were carried out with GeneCodis web tool (http://genecodis.dacya.ucm.es/) [31]. Use the KEGG database to build the network of genes in accordance with the relationship among the genes, proteins and compounds in the database. GO analysis was applied to analyze the main function of the target genes of differential expression miRNA according to the Gene Ontology which is the key functional classification of NCBI [32]. Enrichment provides a measure of the significance of the function: as the enrichment increases, the corresponding function is more specific, which helps us to find those GOs with more concrete function description in adipogenesis.
miRNA-GO network
MiRNA-GO network is built according to the relationship of miRNAs and significant GOs. In the miRNA-GO network, the circle represents GO and the shape of red square represents miRNAs, and their relationship is represented by one edge. The center of the network is represented by degree. Degree of miRNA means the number of links one miRNA regulates GOs. The core miRNAs in the network always have the highest degrees.
Statistical analysis
The lists of miRNAs were extracted based on statistical test P-values (<0.05 was considered significant). The representatives of replicate experiments were shown in the figures and the data were shown as the means ± SEM. The statistical analyses were performed using independent Student's t-tests. In case of false positive results, Bonferroni correction was performed.
SUPPLEMENTARY MATERIAL
Footnotes
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
GRANT SUPPORT
This study was supported by grants from the National Key Basic Research Program of China (2013CB530604), Key project of the National Natural Science Foundation of China (81330067), National Natural Science Foundation of China (81200642, 81270928, 81370964,81500674), Program for Innovative Research Teams of Jiangsu Province (LJ201108), National Natural Science Foundation of Jiangsu Province (BK20150082), Nanjing Technological Development Program (201104013, ZKX14042, YKK15165), Key Project supported by Medical Science and technology developmental Foundation, Nanjing Department of Health (JQX13012) and Science and Technology Development Fund of Nanjing Medical University (2014 NJMUZD041).
Authors' contributions
Author contributions include the following: XR.G., CB.J., CM.S. conceived and designed the experiments .CM.S., FY.H., XH.G, LH.Y., X.W., J.W. and XW.C. performed the experiments and interpreted the data. CM.S. and CB.J. contributed to the get the meta data, XR.G. and CB.J. contributed to the funding. CM.S. wrote the manuscript.
REFERENCES
- 1.Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell. 2014;156:20–44. doi: 10.1016/j.cell.2013.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Trajkovski M, Hausser J, Soutschek J, Bhat B, Akin A, Zavolan M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature. 2011;474:649–53. doi: 10.1038/nature10112. [DOI] [PubMed] [Google Scholar]
- 3.Jordan SD, Kruger M, Willmes DM, Redemann N, Wunderlich FT, Bronneke HS, Merkwirth C, Kashkar H, Olkkonen VM, Böttger T, Braun T, Seibler J, Brüning JC. Obesity-induced overexpression of miRNA-143 inhibits insulin-stimulated AKT activation and impairs glucose metabolism. Nat Cell Biol. 2011;13:434–46. doi: 10.1038/ncb2211. [DOI] [PubMed] [Google Scholar]
- 4.Lin Q, Gao Z, Alarcon RM, Ye J, Yun Z. A role of miR-27 in the regulation of adipogenesis. FEBS J. 2009;276:2348–58. doi: 10.1111/j.1742-4658.2009.06967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shi C, Zhang M, Tong M, Yang L, Pang L, Chen L, Xu G, Chi X, Hong Q, Ni Y, Ji C, Guo X. miR-148a is Associated with Obesity and Modulates Adipocyte Differentiation of Mesenchymal Stem Cells through Wnt Signaling. Sci Rep. 2015;5:9930. doi: 10.1038/srep09930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen L, Dai YM, Ji CB, Yang L, Shi CM, Xu GF, Pang LX, Huang FY, Zhang CM, Guo XR. MiR-146b is a regulator of human visceral preadipocyte proliferation and differentiation and its expression is altered in human obesity. Mol Cell Endocrinol. 2014;393:65–74. doi: 10.1016/j.mce.2014.05.022. [DOI] [PubMed] [Google Scholar]
- 7.Song G, Xu G, Ji C, Shi C, Shen Y, Chen L, Zhu L, Yang L, Zhao Y, Guo X. The role of microRNA-26b in human adipocyte differentiation and proliferation. Gene. 2014;533:481–7. doi: 10.1016/j.gene.2013.10.011. [DOI] [PubMed] [Google Scholar]
- 8.Yang L, Shi CM, Chen L, Pang LX, Xu GF, Gu N, Zhu LJ, Guo XR, Ni YH, Ji CB. The biological effects of hsa-miR-1908 in human adipocytes. Mol Biol Rep. 2015;42:927–35. doi: 10.1007/s11033-014-3830-1. [DOI] [PubMed] [Google Scholar]
- 9.Kolde R, Laur S, Adler P, Vilo J. Robust rank aggregation for gene list integration and meta-analysis. Bioinformatics. 2012;28:573–80. doi: 10.1093/bioinformatics/btr709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xu G, Ji C, Song G, Zhao C, Shi C, Song L, Chen L, Yang L, Huang F, Pang L, Zhang N, Zhao Y, Guo X. MiR-26b modulates insulin sensitivity in adipocytes by interrupting the PTEN/PI3K/AKT pathway. Int J Obes (Lond) 2015;39:1523–30. doi: 10.1038/ijo.2015.95. [DOI] [PubMed] [Google Scholar]
- 11.Rankinen T, Zuberi A, Chagnon YC, Weisnagel SJ, Argyropoulos G, Walts B, Pérusse L, Bouchard C. The human obesity gene map: the 2005 update. Obesity (Silver Spring) 2006;14:529–644. doi: 10.1038/oby.2006.71. [DOI] [PubMed] [Google Scholar]
- 12.Martinelli R, Nardelli C, Pilone V, Buonomo T, Liguori R, Castano I, Buono P, Masone S, Persico G, Forestieri P, Pastore L, Sacchetti L. miR-519d overexpression is associated with human obesity. Obesity (Silver Spring) 2010;18:2170–6. doi: 10.1038/oby.2009.474. [DOI] [PubMed] [Google Scholar]
- 13.Ferrante SC, Nadler EP, Pillai DK, Hubal MJ, Wang Z, Wang JM, Gordish-Dressman H, Koeck E, Sevilla S, Wiles AA, Freishtat RJ. Adipocyte-derived exosomal miRNAs: a novel mechanism for obesity-related disease. Pediatr Res. 2015;77:447–54. doi: 10.1038/pr.2014.202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Arner E, Mejhert N, Kulyte A, Balwierz PJ, Pachkov M, Cormont M, Lorente-Cebrián S, Ehrlund A, Laurencikiene J, Hedén P, Dahlman-Wright K, Tanti JF, Hayashizaki Y, et al. Adipose tissue microRNAs as regulators of CCL2 production in human obesity. Diabetes. 2012;61:1986–93. doi: 10.2337/db11-1508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen YH, Heneidi S, Lee JM, Layman LC, Stepp DW, Gamboa GM, Chen BS, Chazenbalk G, Azziz R. miRNA-93 inhibits GLUT4 and is overexpressed in adipose tissue of polycystic ovary syndrome patients and women with insulin resistance. Diabetes. 2013;62:2278–86. doi: 10.2337/db12-0963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hamam D, Ali D, Vishnubalaji R, Hamam R, Al-Nbaheen M, Chen L, Kassem M, Aldahmash A, Alajez NM. microRNA-320/RUNX2 axis regulates adipocytic differentiation of human mesenchymal (skeletal) stem cells. Cell Death Dis. 2014;5:e1499. doi: 10.1038/cddis.2014.462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Skarn M, Namlos HM, Noordhuis P, Wang MY, Meza-Zepeda LA, Myklebost O. Adipocyte differentiation of human bone marrow-derived stromal cells is modulated by microRNA-155, microRNA-221, and microRNA-222. Stem Cells Dev. 2012;21:873–83. doi: 10.1089/scd.2010.0503. [DOI] [PubMed] [Google Scholar]
- 18.Ortega FJ, Moreno-Navarrete JM, Pardo G, Sabater M, Hummel M, Ferrer A, Rodriguez-Hermosa JI, Ruiz B, Ricart W, Peral B, Fernández-Real JM. MiRNA expression profile of human subcutaneous adipose and during adipocyte differentiation. PLoS One. 2010;5:e9022. doi: 10.1371/journal.pone.0009022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mikkelsen TS, Xu Z, Zhang X, Wang L, Gimble JM, Lander ES, Rosen ED. Comparative epigenomic analysis of murine and human adipogenesis. Cell. 2010;143:156–69. doi: 10.1016/j.cell.2010.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ahn J, Lee H, Jung CH, Jeon TI, Ha TY. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol Med. 2013;5:1602–12. doi: 10.1002/emmm.201302647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhu L, Chen L, Shi CM, Xu GF, Xu LL, Zhu LL, Guo XR, Ni Y, Cui Y, Ji C. MiR-335, an adipogenesis-related microRNA, is involved in adipose tissue inflammation. Cell Biochem Biophys. 2014;68:283–90. doi: 10.1007/s12013-013-9708-3. [DOI] [PubMed] [Google Scholar]
- 22.Zhu L, Shi C, Ji C, Xu G, Chen L, Yang L, Fu Z, Cui X, Lu Y, Guo X. FFAs and adipokine-mediated regulation of hsa-miR-143 expression in human adipocytes. Mol Biol Rep. 2013;40:5669–75. doi: 10.1007/s11033-013-2668-2. [DOI] [PubMed] [Google Scholar]
- 23.Wildwater M, Sander N, de Vreede G, van den Heuvel S. Cell shape and Wnt signaling redundantly control the division axis of C. elegans epithelial stem cells. Development. 2011;138:4375–85. doi: 10.1007/s11033-013-2668-2. [DOI] [PubMed] [Google Scholar]
- 24.Bost F, Aouadi M, Caron L, Binetruy B. The role of MAPKs in adipocyte differentiation and obesity. Biochimie. 2005;87:51–6. doi: 10.1016/j.biochi.2004.10.018. [DOI] [PubMed] [Google Scholar]
- 25.Esau C, Kang X, Peralta E, Hanson E, Marcusson EG, Ravichandran LV, Sun Y, Koo S, Perera RJ, Jain R, Dean NM, Freier SM, Bennett CF, et al. MicroRNA-143 regulates adipocyte differentiation. J Biol Chem. 2004;279:52361–5. doi: 10.1074/jbc.C400438200. [DOI] [PubMed] [Google Scholar]
- 26.Xiang X, Zhao J, Xu G, Li Y, Zhang W. mTOR and the differentiation of mesenchymal stem cells. Acta Biochim Biophys Sin (Shanghai) 2011;43:501–10. doi: 10.1093/abbs/gmr041. [DOI] [PubMed] [Google Scholar]
- 27.Wang J, Kong J, Qi Y, Quigg R, Li X. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc Natl Acad Sci USA. 2008;105:2889–94. doi: 10.1073/pnas.0800178105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wu Y, Zuo J, Zhang Y, Xie Y, Hu F, Chen L, Liu B, Liu F. Identification of miR-106b-93 as a negative regulator of brown adipocyte differentiation. Biochem Biophys Res Commun. 2013;438:575–80. doi: 10.1016/j.bbrc.2013.08.016. [DOI] [PubMed] [Google Scholar]
- 29.Wei J, Li H, Wang S, Li T, Fan J, Liang X, Li J, Han Q, Zhu L, Fan L, Zhao R. let-7 enhances osteogenesis and bone formation while repressing adipogenesis of human stromal/mesenchymal stem cells by regulating HMGA2. Stem Cells Dev. 2014;23:1452–63. doi: 10.1089/scd.2013.0600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell. 2007;27:91–105. doi: 10.1016/j.molcel.2007.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nogales-Cadenas R, Carmona-Saez P, Vazquez M, Vicente C, Yang X, Tirado F, Carazo JM, Pascual-Montano A. GeneCodis: interpreting gene lists through enrichment analysis and integration of diverse biological information. Nucleic Acids Res. 2009;37:W317–22. doi: 10.1093/nar/gkp416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gene Ontology C. The Gene Ontology (GO) project in 2006. Nucleic Acids Res. 2006;34:D322–6. doi: 10.1093/nar/gkj021. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.