

Abstract
Background
Bacteria treated with different classes of antibiotics exhibit changes in susceptibility to successive antibiotic treatments. This study was designed to evaluate the influence of sequential antibiotic treatments on the development of antibiotic resistance in Klebsiella pneumoniae associated with β-lactamase and efflux pump activities.
Methods
The antibiotic susceptibility, β-lactamase activity, and efflux activity were determined in K. pneumoniae grown at 37 °C by adding initial (0 h) and second antibiotics (8 or 12 h). Treatments include control (CON; no first and second antibiotic addition), no initial antibiotic addition followed by 1 MIC ciprofloxacin addition (CON-CIP), no initial antibiotic addition followed by 1 MIC meropenem addition (CON-MER), initial 1/4 MIC ciprofloxacin addition followed by no antibiotic addition (1/4CIP-CON), initial 1/4 MIC ciprofloxacin addition followed by 1 MIC ciprofloxacin addition (1/4CIP-CIP), and initial 1/4 MIC ciprofloxacin addition followed by 1 MIC meropenem addition (1/4CIP-MER).
Results
Compared to the CON, the initial addition of 1/4 MIC ciprofloxacin inhibited the growth of K. pneumoniae throughout the incubation period. The ciprofloxacin treatments (CON-CIP and 1/4CIP-CIP) showed significant reduction in the number of K. pneumoniae cells compared to meropenem (CON-MER and 1/4CIP-MER). The 1/4CIP-CIP achieved a further 1 log reduction of K. pneumoniae, when compared to the 1/4CIP-CON and 1/CIP-MER. The increase in sensitivity of K. pneumoniae to cefotaxime, kanamycin, levofloxacin, nalidixic acid was observed for CON-CIP. Noticeable cross-resistance pattern was observed at the 1/4CIP-CIP, showing the increased resistance of K. pneumoniae to chloramphenicol, ciprofloxacin, kanamycin, levofloxacin, nalidixic acid norfloxacin, sulphamethoxazole/trimethoprim, and tetracycline. The levels of β-lactamase activities were estimated to be 8.4 μmol/min/ml for CON, 7.7 μmol/min/ml for 1/4CIP-CON and as low as 2.9 μmol/min/ml for CON-CIP. Compared to the absence of phenylalanine-arginine-β-naphthylamide (PAβN), the fluorescence intensity of EtBr was increased in K. pneumoniae cells treated at the CON, CON-CIP, and CON-MER in the presence of PAβN. However, the efflux pump activity remained in K. pneumoniae cells treated at the 1/CIP, 1/CIP–CIP, and 1/CIP-MER in the presence of PAβN.
Conclusion
The results suggest that the pre-exposed antibiotic history, treatment order, and concentrations influenced the development of multiple antibiotic resistant associated with β-lactamase and efflux pump activities. This study highlights the importance of antibiotic treatment conditions, which would be taken into consideration when new antibiotic strategy is designed to prevent antibiotic resistance.
Keywords: β-Lactamase, Efflux pump system, Meropenem, Ciprofloxacin, Antibiotic resistance
Background
The overuse, underuse, and misuse of antibiotics have become major causes of the development of antibiotic resistance in bacteria [1]. The emergence and spread of antibiotic-resistant bacteria has been a growing concern over the last decade, which can lead to serious clinical and public health problems. Klebsiella pneumoniae, an opportunistic pathogen, is mainly responsible for nosocomial infections with high morbidity and mortality [2]. Recently, the rapid emergence of extended-spectrum β-lactamase (ESBL) producing K. pneumoniae has significantly increased the risk of developing serious nosocomial and community-acquired infections worldwide [3, 4]. Furthermore, multidrug-resistant K. pneumoniae strains can cause treatment failure with current antibiotic therapy [2].
The proposed resistance mechanisms of K. pneumoniae against different classes of antibiotics include release of antibiotic-inactivating enzymes, modification of antibiotic target sites, change in membrane permeability, activation of efflux pump systems, and alteration of metabolic pathways [2, 5]. Among these mechanisms, the enzymatic degradation and efflux pump systems play an important role in the development of multidrug resistance in K. pneumoniae [6]. K. pneumoniae strains produce enzymes, including extended-spectrum β-lactamases, metallo-β-lactamases, oxacillinases, and carbapenemases, that can degrade β-lactam antibiotics. The efflux pumps, belonging to the resistance-nodulation-division (RND) family, can extrude amphiphilic and charged antibiotics such as β-lactams, fluoroquinolones, and aminoglycosides [2].
The sequential and combination antibiotic therapies have currently been used to reduce not only the evolution of multidrug resistant bacteria but also the levels of antibiotics used in the treatment of bacterial infection [7–9]. The decreased selection pressure occurs when antibiotics are treated in appropriate order, which can prevent the emergence and spread of multidrug resistance [10]. From the practical viewpoint of antibiotic effectiveness, however, there is an important challenging question of whether the treatment history can cause potential carry-over effects on the additional antibiotic therapy. The pre-exposure to antibiotics influences the acquisition of resistance to second-line antibiotics [11]. The beneficial and adverse effects of current therapeutic approaches for treating bacterial infections are still under debate. Therefore, the objective of this study was to evaluate the impact of sequential antibiotic treatments on the antibiotic susceptibility and resistance mechanisms of K. pneumoniae in association with β-lactamase and efflux pump activities.
Methods
Bacterial strain and culture condition
Strain of K. pneumoniae ATCC 23357 was purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). The strain was cultured in trypticase soy broth (TSB; BD, Becton, Dickinson and Co., Sparks, MD, USA) at 37 °C for 18 h. After cultivation, the culture was centrifuged at 3000×g for 20 min at 4 °C. The harvested cells were washed twice with phosphate-buffered saline (PBS, pH 7.2) and diluted to 108 CFU/ml for assays.
Antibiotic susceptibility assay
The susceptibility of K. pneumoniae to ciprofloxacin and meropenem was evaluated according to the Clinical Laboratory Standards Institute (CLSI) procedure with slight modification [12]. All antibiotics used in this study were purchased from Sigma Chemicals (St. Louis, MO, USA). Antibiotic stock solutions were prepared by dissolving in glacial acetic acid for ciprofloxacin and distilled water for meropenem to obtain a final concentration of 10.24 mg/ml. The stock solutions of ciprofloxacin and meropenem (100 μl each) were serially (1:2) diluted to concentrations ranging from 2 to 0 μg/ml with TSB in 96-well flat-bottomed polystyrene microtiter plates (BD Falcon, San Jose, CA, USA). The test strain was inoculated at 105 CFU/ml and incubated at 37 °C for 18 h. Minimum inhibitory concentrations (MICs) of ciprofloxacin and meropenem were defined as the lowest concentration of each antibiotic at which no visible cell growth was observed.
Dynamic time-kill curve analysis
Time-kill curve assay was carried out to determine the antibiotic activities of ciprofloxacin and meropenem against K. pneumoniae using a sequential treatment scheme as described in Table 1. The initial population (105 CFU/ml) of K. pneumoniae was inoculated at 37 °C in TSB treated with (Treatments 1, 2, and 3) and without antibiotic (Treatments 4, 5, and 6). After being reached an optical density (OD) of 0.5, the non-adapted and ciprofloxacin-adapted K. pneumoniae cells were further treated with no (Treatments 1 and 4) and the increased concentrations of ciprofloxacin (Treatments 2 and 5) and meropenem (Treatments 3 and 6). The survival curves were measured at 600 nm at every 4 h interval throughout the incubation period. The cultured cells were collected for further analyses, including bacterial enumeration, disk diffusion, lactamase activity, and cartwheel tests.
Table 1.
Sequential antibiotic treatments used in this study
| Treatment | Antibiotic addition (h) | Abbreviation | Description | |
|---|---|---|---|---|
| 0 h | 8 or 12 h | |||
| 1 | No (0) | No (8) | CON | No first and second antibiotic addition |
| 2 | No (0) | 1 MIC CIP (8) | CON-CIP | No initial antibiotic addition followed by 1 MIC ciprofloxacin addition |
| 3 | No (0) | 1 MIC MER (8) | CON-MER | No initial antibiotic addition followed by 1 MIC meropenem addition |
| 4 | 1/4 MIC CIP (0) | No (12) | 1/4CIP-CON | Initial 1/4 MIC ciprofloxacin followed by no antibiotic addition |
| 5 | 1/4 MIC CIP (0) | 1 MIC CIP (12) | 1/4CIP-CIP | Initial 1/4 MIC ciprofloxacin followed by 1 MIC ciprofloxacin addition |
| 6 | 1/4 MIC CIP (0) | 1 MIC MER (12) | 1/4CIP-MER | Initial 1/4 MIC ciprofloxacin followed by 1 MIC meropenem addition |
CON, CIP, and MER denote no antibiotic addition, ciprofloxacin, and meropenem, respectively
Microbial analysis
The collected cells were serially diluted (1:10) with PBS and then plated on trypticase soy agar (TSA) using an Autoplate Spiral Plating System (Spiral Biotech Inc., Norwood, MA, USA). The plates were incubated at 37 °C for 24–48 h to enumerate viable cells using a QCount Colony Counter (Spiral Biotech Inc.). Log reduction (N TRT/N CON) was calculated for each treatment as compared to the control (CON). N CON and N TRT represent the numbers of control and treatments after incubation, respectively.
Disk-diffusion assay
The antibiotic susceptibility of cultured K. pneumoniae was determined by the disk-diffusion test. The cultured cells were evenly spread-plated on TSA and then allowed to dry for 5 min. Antibiotic disks (Becton, Dickinson and Company, NJ, USA), including cefotaxime (CTX; 30 μg), chloramphenicol (CHL; 30 μg), ciprofloxacin (CIP; 5 μg), kanamycin (KAN; 30 μg), levofloxacin (LEV; 5 μg), nalidixic acid (NAL; 30 μg), norfloxacin (NOR; 10 μg), sulphamethoxazole/trimethoprim (S/T; 25 μg), and tetracycline (TET; 30 μg), were placed on the surface of the agar using sterilized forceps. After incubation at 38 °C for 24 h, the diameters of inhibition zone were measured using a metric ruler.
β-Lactamase activity assay
The β-lactamase activity was evaluated using a nitrocefin hydrolysis assay [13]. The cultured cells were centrifuged at 3000×g for 20 min at 4 °C to collect cell-free supernatants. The cell-free supernatants were mixed with 20 μl of 1.5 mM nitrocefin (Biovision, Inc., CA, USA) and incubated at 37 °C. The absorbance was measured every 5 min up to 1 h at 515 nm [14].
Ethidium bromide (EtBr)-agar cartwheel assay
The cultured K. pneumoniae cells were centrifuged at 3000×g for 20 min at 4 °C, rinsed twice with PBS, and then suspended in PBS with and without efflux pump inhibitor (phenylalanine-arginine-β-naphthylamide; PAβN, 8 μg/ml) [15, 16]. The prepared cells were swabbed on EtBr-agar plates containing EtBr (1 μg/ml) and incubated at 37 °C for 18 h. After incubation, the swabbed EtBr-agar plates exposed to UV light were photographed using a Gel-doc XR System (Bio-Rad, Hertfordshire, UK).
Statistical analysis
All analyses were conducted in duplicates for three replicates. Data were analyzed by the Statistical Analysis System (SAS) software. The General Linear Model (GLM) and least significant difference (LSD) procedures were used to compare treatments at p < 0.05.
Results
Effect of serial antibiotic treatments on the viability and antibiotic susceptibility
The MIC values of ciprofloxacin and meropenem were 0.03 and 0.06 μg/ml, respectively. The survival of K. pneumoniae was observed during the sequential antibiotic treatments (Fig. 1). The growth of K. pneumoniae was retarded by 1/4 MIC ciprofloxacin, showing that the non- and ciprofloxacin-treated K. pneumoniae cells reached OD of 0.5 after 8 and 12 h of incubation, respectively. The antibiotic effects of ciprofloxacin and meropenem on the growth of K. pneumoniae was noticeable in the non-adapted cells (CON-CIP and CON-MER) compared to the ciprofloxacin-adapted cells (1/4CIP-CIP and 1/4CIP-MER). The number of viable K. pneumoniae cells under 1/4CIP-CIP was significantly reduced by 1.8 log, followed by CON-CIP and 1/4CIP-MER (Fig. 2). The least reduction in K. pneumoniae cells was observed at the CON-MER. The reduction rate increased with increasing the concentration of CIP to 0.03 μg/ml (1/4CIP-CIP), while no significant reduction was observed for the 1/4CIP-MER switched from CIP to MER at the second treatment. The antibiotic resistance acquisition was evaluated in K. pneumoniae exposed to sequential antibiotic treatments (Fig. 3). The susceptibilities of K. pneumoniae to different classes of antibiotics varied in the treatments. The 1/4CIP-CIP treatment exhibited the decreased susceptibility to all antibiotics with exception of CTX, showing small clear zones (<10 cm). The enhanced susceptibility to all antibiotics tested was observed at the CON-CIP treatment, while the decreased susceptibilities to CIP and TET were observed at the treatments, CON-MER, 1/4CIP-CON, and 1/4CIP-MER. The treatments, CON-MER and 1/4CIP-MER, showed the similar resistance pattern as the CON.
Fig. 1.
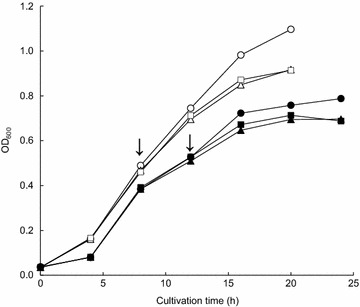
Survival of Klebsiella pneumonia exposed to sequential antibiotics (CON, open circle; CON-CIP, open triangle; CON-MER, open square; 1/4CIP-CON, closed circle; 1/4CIP-CIP, closed triangle; 1/4CIP-MER, closed square). Arrow indicates the addition time
Fig. 2.
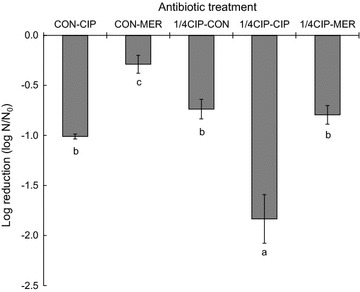
Reduction of Klebsiella pneumonia exposed to sequential antibiotics. N CON and N TRT represent the numbers of control and treatments after incubation, respectively. Log reduction with different letters (a–c) are significantly different at p < 0.05
Fig. 3.
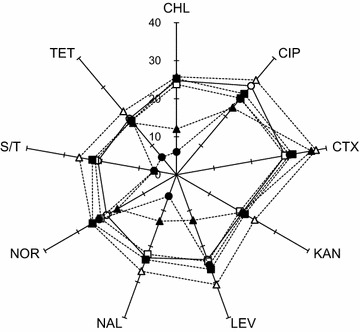
Radar plot of antibiotic resistance profiles (disk diffusion in cm) of Klebsiella pneumonia exposed to sequential antibiotics (CON, open circle; CON-CIP, open triangle; CON-MER, open square; 1/4CIP-CON, closed circle; 1/4CIP-CIP, closed triangle; 1/4CIP-MER, closed square). Antibiotic discs include chloramphenicol (CHL; 30 μg), ciprofloxacin (CIP; 5 μg), cefotaxime (CTX; 30 μg), kanamycin (KAN; 30 μg), levofloxacin (LEV; 5 μg), nalidixic acid (NAL; 30 μg), norfloxacin (NOR; 10 μg), sulphamethoxazole/trimethoprim (S/T; 25 μg), and tetracycline (TET; 30 μg)
Effect of sequential antibiotic treatments on β-lactamase and efflux pump activities
The β-lactamase activities were measured in K. pneumoniae cultured at sequential antibiotic treatments (Fig. 4). The highest β-lactamase activity was observed for the CON (8.4 μmol/min/ml), followed by 1/4CIP-CON (7.7 μmol/min/ml) and CON-MER (6.0 μmol/min/ml).
Fig. 4.
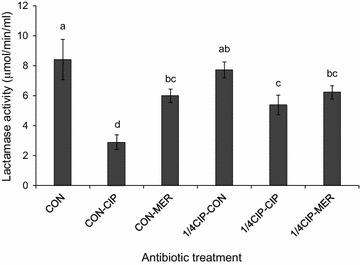
Hydrolyzing activity of the β-lactamases produced from Klebsiella pneumonia exposed to sequential antibiotics (CON, CON-CIP, CON-MER, 1/4CIP-CON, 1/4CIP-CIP, and 1/4CIP-MER). Means with different letters (a–d) are significantly different at p < 0.05
The efflux activity of K. pneumoniae in the absence and presence of efflux pump inhibitor (EPI; PAβN) was evaluated on the EtBr-agar plates (Fig. 5). The efflux pump activity of K. pneumoniae was observed at all treatments in the absence of PAβN, showing the decreased fluorescence intensity (Fig. 5a). The increased EtBr accumulation was observed in the presence of PAβN at the CON, CON-CIP, and CON-MER, while relatively low EtBr accumulation was observed at the 1/4CIP-CON, 1/4CIP-CIP, and 1/4CIP-MER (Fig. 5b). The efflux pump activity in K. pneumoniae cells treated at the 1/4CIP-CIP, 1/4CIP-CIP, and 1/4CIP-MER was not reduced by the PAβN, suggesting that the EPI-insensitive efflux systems were activated in K. pneumoniae cells under sublethal concentration of ciprofloxacin.
Fig. 5.

Accumulation and efflux activity of Klebsiella pneumonia on EtBr agar plates containing without (a) and with (b) efflux pump inhibitor (PAβN). (1 CON; 2 CON-CIP; 3 CON-MER; 4, 1/4CIP-CON; 5 1/4CIP-CIP; 6 1/4CIP-MER)
Discussion
This study describes the effect of sequential antibiotic treatments on the development of antibiotic resistance in K. pneumoniae. Pathogenic bacteria exposed to different levels and various classes of antibiotics can exhibit various susceptibility to additional antibiotic treatments. However, few studies have been focused on the development of antibiotic resistance during the sequential antibiotic treatments [11]. Thus, this study highlights the pre-exposed antibiotic history as an important factor for the evolution of antibiotic resistance.
The CON-CIP was more effectively reduced the number of K. pneumoniae cells than the CON-MER, while no significant difference was observed between 1/4CIP-CON and 1/4CIP-MER (Fig. 2). The results indicate that meropenem was less effective against K. pneumoniae than ciprofloxacin. The lack of antibacterial activity of meropenem might be due to its short elimination half-life [17]. The successive ciprofloxacin treatment (1/4CIP-CIP) showed strong antibacterial activity against K. pneumoniae (Fig. 2). However, the cells exposed to 1/4CIP-CIP treatment induced were significantly resistant to different classes of antibiotics such as CHL, CIP, LEV, NAL, and S/T, known as cross-resistance induction (Fig. 3). This suggests that the sequential single-antibiotic therapy at different levels can lead to the emergence of antibiotic resistant bacteria due to the phenotypic adaptation [18]. Previous studies have reported that antibiotic monotherapy and combination therapy are the main causes of narrow- and broad-spectrum resistance, respectively [9]. The concentration of ciprofloxacin treated after the initial CIP exposure might be insufficient to suppress the development of antibiotic resistance [19]. The sequential antibiotic treatment, specifically 1/4CIP-CIP, can possibly cause a change in antibiotic resistance. This might be attributed to the mode of action of antibiotics, including time- and concentration-dependent activities. The time-dependent antibiotics include β-lactams and carbapenems, characterized by slow antibiotic activity according to exposure time above MIC [20, 21]. Ciprofloxacin is referred to as concentration-dependent antibiotic, showing antibiotic activity at high concentration [22]. Due to the bacterial adaptation, the sequential treatment of homogeneous antibiotic can reduce the susceptibility to same and different classes of antibiotics [19, 23]. Homogeneous treatment increases bacterial fitness and selection pressure, resulting in the development of antibiotic resistance [24, 25].
The strain of K. pneumoniae used in this study was intrinsically resistant to ampicillin (MIC > 256 μg/ml), piperazine (MIC > 16 μg/ml), cephalotin (MIC > 32 μg/ml), and cefoxitin (MIC > 16 μg/ml), resulted from the production of β-lactamases [26]. No significant change in β-lactamase activities was observed for 1/4CIP-CON when compared to the CON, while the β-lactamase activities were significantly decreased at the CON-CIP, CON-MER, 1/4CIP-CIP, and 1/4CIP-MER. The decrease in lactamase activity suggest that relatively fewer viable cells were observed at the treatments compared to the CON and the antibiotics used may act as a lactamase inhibitor [27]. K. pneumoniae cells exposed to CON-CIP showed the increased susceptibility to CTX (Fig. 3), which corresponds to the significant reduction in β-lactamase activity treated at CON-CIP (2.9 μmol/min/ml) (Fig. 4). The mechanisms of resistance to β-lactam antibiotics include not only the increase in β-lactamases-mediated hydrolysis but also the decrease in uptake through porin channels and the increase in efflux pump activity [6, 28]. The alteration in porin channel results in the decrease in membrane permeability to antibiotics such as β-lactams and quinolones, but not aminoglycosides that can enter the outer membrane through lipopolysaccharide (LPS) uptake system [19]. The results explain that the susceptibilities of K. pneumoniae cells grown at all treatments to KAN were constant regardless of the levels of β-lactamase activities (Fig. 3). Carbapenems such as imipenem and meropenem are stable to β-lactamases that can hydrolyze β-lactams [28–30]. The production of β-lactamases can contribute to the hydrolysis, resulting in the enhanced resistance to β-lactam antibiotics that cannot reach penicillin binding protein (PBP) [26]. This is in good agreement with the result of antibiotic susceptibility that K. pneumoniae possesses high intrinsic resistance to ampicillin (MIC > 256 μg/ml).
The efflux pump systems competitively expel substrates, which plays an important role in the development of multidrug resistance [2]. The specific substrates for efflux pumps in K. pneumoniae treated at 1/4CIP-CIP and 1/4CIP-CIP may include CHL, TET, NOR, NAL, and LEV, corresponding the enhanced multiple antibiotic resistance in disk-diffusion assay in Fig. 3. The intracellular concentrations of antibiotics substrates in bacteria are directly reduced by active efflux pump systems, leading to the decreased antibiotic susceptibility [31]. In other words, the inhibition of efflux pumps can restore antibiotic susceptibility in resistant bacteria, suggesting the EPI can be a possible control method against antibiotic-resistant bacteria [32–34]. The activation of efflux pumps is associated with the fast-acting antibiotic mechanism at the early stage of antibiotic resistance development [19]. The substrate-dependent efflux pump systems may contribute to cross-resistance to different classes of antibiotics [35].
Conclusions
This study highlights the influence of using sequential antibiotic treatment on the development of antibiotic resistance in K. pneumoniae. The difference in antibiotic activity against K. pneumoniae might be due to the time- and concentration-dependent modes of action of meropenem and ciprofloxacin, respectively. The antibiotic resistance mechanisms can occur within a short time in association with lactamase production, membrane permeability, and efflux pump activity. The efflux pump activities in K. pneumoniae cells treated at 1/4CIP-CON, 1/4CIP-CIP, and 1/4CIP-MER were still observed in the presence of PAβN, indicating that various types of substrate-dependent efflux pumps exist to reduce the intracellular concentrations of antibiotics. The induced efflux pump activity are responsible for the increased antibiotic resistance in K. pneumoniae. The most significant finding in this study was that K. pneumoniae treated with sequential antibiotics, specifically 1/4CIP-CIP, showed reduced susceptibility towards CHL, NOR, NAL, and LEV, leading to the multiple antibiotic resistance. Therefore, antibiotic therapies should take into account the history of pre-exposed antibiotics to prevent the development of antibiotic resistance. Further studies are needed to assess the risk of antibiotic resistance in sequential and combination antibiotic therapies, which is essential to design an effective strategy for controlling multiple antibiotic resistant bacteria.
Authors’ contributions
JK and AJ conducted all experiments and EC contributed to the writing and preparation of the manuscript. JA contributed to the experimental design, data interpretation, and manuscript writing. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The data supporting the conclusions are included within the manuscript.
Funding
This work supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (Grant number: HI15C-1798-000016).
Abbreviations
- ESBL
extended-spectrum β-lactamase
- MIC
minimum inhibitory concentration
- CIP
ciprofloxacin
- MER
meropenem
- CTX
cefotaxime
- CHL
chloramphenicol
- KAN
kanamycin
- LEV
levofloxacin
- NAL
nalidixic acid
- NOR
norfloxacin
- S/T
sulphamethoxazole/trimethoprim
- TET
tetracycline
- CFU
colony forming unit
- EtBr
ethidium bromide
- EPI
efflux pump inhibitor
- PAβN
phenylalanine-arginine-β-naphthylamide
Contributor Information
Jeongjin Kim, Email: kjj@kangwon.ac.kr.
Ara Jo, Email: jar1920@kangwon.ac.kr.
Ekachai Chukeatirote, Email: ekachai@mfu.ac.th.
Juhee Ahn, Phone: +82 33 250 6564, Email: juheeahn@kangwon.ac.kr.
References
- 1.Odonkor ST, Addo KK. Bacterial resistance to antibiotics: recent trends and challenges. Int J Bio Med Res. 2011;2(4):1204–1210. [Google Scholar]
- 2.HQ Zhong, Zhang S, Pan H, Cai T. Influence of induced ciprofloxacin resistance on efflux pump activity of Klebsiella pneumoniae. J Zhejiang Univ Sci B. 2013;14(9):837–843. doi: 10.1631/jzus.B1200221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brinkworth AJ, Hammer CH, Olano LR, Kobayashi SD, Chen L, Kreiswirth BN, et al. Identification of outer membrane and exoproteins of carbapenem-resistant multilocus sequence Type 258 Klebsiella pneumoniae. PLoS ONE. 2015;10(4):e0123219. doi: 10.1371/journal.pone.0123219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Latifpour M, Gholipour A, Damavandi MS. Prevalence of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae isolates in nosocomial and community-acquired urinary tract infections. Jundishapur J Microbiol. 2016;9(3):e31179. doi: 10.5812/jjm.31179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tenover FC. Mechanisms of antimicrobial resistance in bacteria. Am J Med. 2006;119(6A):S3–S10. doi: 10.1016/j.amjmed.2006.03.011. [DOI] [PubMed] [Google Scholar]
- 6.Pages J-M, Lavigne J-P, Leflon-Guibout V, Marcon E, Bert F, Noussair L, et al. Efflux pump, the masked side of β-lactam resistance in Klebsiella pneumoniae clinical isolates. PLoS ONE. 2009;4(3):e4817. doi: 10.1371/journal.pone.0004817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nichol D, Jeavons P, Fletcher AG, Bonomo RA, Maini PK, Paul JL, et al. Steering evolution with sequential therapy to prevent the emergence of bacterial antibiotic resistance. PLoS Comput Biol. 2015;11(9):e1004493. doi: 10.1371/journal.pcbi.1004493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Perron GG, Kryazhimskiy S, Rice DP, Buckling A. Multidrug therapy and evolution of antibiotic resistance: when order matters. Appl Environ Microbiol. 2012;78(17):6137–6142. doi: 10.1128/AEM.01078-12. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 9.Vestergaard M, Paulander W, Marvig RL, Clasen J, Jochumsen N, Molin S, et al. Antibiotic combination therapy can select for broad-spectrum multidrug resistance in Pseudomonas aeruginosa. Int J Antimicrob Agent. 2016;47(1):48–55. doi: 10.1016/j.ijantimicag.2015.09.014. [DOI] [PubMed] [Google Scholar]
- 10.Warren DK, Hill HA, Merz LR, Kollef MH, Hayden MK, Fraser VJ, et al. Cycling empirical antimicrobial agents to prevent emergence of antimicrobial-resistant Gram-negative bacteria among intensive care unit patients. Crit Care Med. 2004;32(12):2450–2456. doi: 10.1097/01.CCM.0000147685.79487.28. [DOI] [PubMed] [Google Scholar]
- 11.Gould IM, MacKenzie FM. Antibiotic exposure as a risk factor for emergence of resistance: the influence of concentration. J Appl Microbiol. 2002;92:78S–84S. doi: 10.1046/j.1365-2672.92.5s1.10.x. [DOI] [PubMed] [Google Scholar]
- 12.CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. Approved standard M07-A9, 9th edn. Wayne, PA: Clinical and Laboratory Standards Institute; 2012.
- 13.Matsumoto Y, Hayama K, Sakakihara S, Nishino K, Noji H, Iino R, et al. Evaluation of multidrug efflux pump inhibitors by a new method using microfluidic channels. PLoS ONE. 2011;6(4):e18547. doi: 10.1371/journal.pone.0018547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sharma S, Ramnani P, Virdi JS. Detection and assay of β-lactamases in clinical and non-clinical strains of Yersinia enterocolitica biovar 1A. J Antimicrob Chemother. 2004;54(2):401–405. doi: 10.1093/jac/dkh365. [DOI] [PubMed] [Google Scholar]
- 15.Costa SS, Falcão C, Viveiros M, Machado D, Martins M, Melo-Cristino J, et al. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Staphylococcus aureus. BMC Microbiol. 2011;11(1):1–12. doi: 10.1186/1471-2180-11-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Martins M, Viveiros M, Couto I, Costa SS, Pacheco T, Fanning S, et al. Identification of efflux pump-mediated multidrug-resistant bacteria by the ethidium bromide-agar cartwheel method. In Vivo. 2011;25(2):171–178. [PubMed] [Google Scholar]
- 17.Walsh F. Doripenem: a new carbapenem antibiotic a review of comparative antimicrobial and bactericidal activities. Ther Clin Risk Manag. 2007;3(5):789–794. [PMC free article] [PubMed] [Google Scholar]
- 18.var der Horst MA, Schuurmans JM, Smid MC, Koenders BB, ter Kuile BH. De vovo acquisition of resistance to three antibiotics by Escherichia coli. Microb Drug Resist. 2011;17(2):141–147. doi: 10.1089/mdr.2010.0101. [DOI] [PubMed] [Google Scholar]
- 19.Wu BM, Sabarinath SN, Rand K, Johnson J, Derendorf H. Suppression of ciprofloxacin-induced resistant Pseudomonas aeruginosa in a dynamic kill curve system. Int J Antimicrob Agent. 2011;37(6):519–524. doi: 10.1016/j.ijantimicag.2011.01.021. [DOI] [PubMed] [Google Scholar]
- 20.Williamson R, Tomasz A. Inhibition of cell wall synthesis and acylation of the penicillin binding proteins during prolonged exposure of growing Streptococcus pneumoniae to benzylpenicillin. Eur J Biochem. 1985;151(3):475–483. doi: 10.1111/j.1432-1033.1985.tb09126.x. [DOI] [PubMed] [Google Scholar]
- 21.Jacobs MR. Optimisation of antimicrobial therapy using pharmacokinetic and pharmacodynamic parameters. Clin Microbiol Infect. 2001;7(11):589–596. doi: 10.1046/j.1198-743x.2001.00295.x. [DOI] [PubMed] [Google Scholar]
- 22.Levison ME, Levison JH. Pharmacokinetics and pharmacodynamics of antibacterial agents. Infect Dis Clin North Am. 2009;23(4):791–815. doi: 10.1016/j.idc.2009.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bergstrom CT, Lo M, Lipsitch M. Ecological theory suggests that antimicrobial cycling will not reduce antimicrobial resistance in hospitals. Proc Nat Acad Sci. 2004;101(36):13285–13290. doi: 10.1073/pnas.0402298101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sandiumenge A, Diaz E, Rodriguez A, Vidaur L, Canadell L, Olona M, et al. Impact of diversity of antibiotic use on the development of antimicrobial resistance. J Antimicrob Chemother. 2006;57(6):1197–1204. doi: 10.1093/jac/dkl097. [DOI] [PubMed] [Google Scholar]
- 25.Bonhoeffer S, Lipsitch M, Levin BR. Evaluating treatment protocols to prevent antibiotic resistance. Proc Nat Acad Sci. 1997;94(22):12106–12111. doi: 10.1073/pnas.94.22.12106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zou L-K, Li L-W, Pan X, Tian G-B, Luo Y, Wu Q, et al. Molecular characterization of β-lactam-resistant Escherichia coli isolated from Fu River, China. World J Microbiol Biotechnol. 2012;28(5):1891–1899. doi: 10.1007/s11274-011-0987-9. [DOI] [PubMed] [Google Scholar]
- 27.Coleman K, Levasseur P, Girard A-M, Borgonovi M, Miossec C, Merdjan H, et al. Activities of ceftazidime and avibactam against β-lactamase-producing Enterobacteriaceae in a hollow-fiber pharmacodynamic model. Antimicrob Agent Chemother. 2014;58(6):3366–3372. doi: 10.1128/AAC.00080-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Poole K. Resistance to β-lactam antibiotics. Cell Mol Life Sci. 2004;61(17):2200–2223. doi: 10.1007/s00018-004-4060-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Franceschini N, Segatore B, Perilli M, Vessillier S, Franchino L, Amicosante G. Meropenem stability to β-lactamase hydrolysis and comparative in vitro activity against several β-lactamase-producing Gram-negative strains. J Antimicrob Chemother. 2002;49(2):395–398. doi: 10.1093/jac/49.2.395. [DOI] [PubMed] [Google Scholar]
- 30.Nicoletti G, Russo G, Bonfiglio G. Recent developments in carbapenems. Expert Opin Investig Drug. 2002;11(4):529–544. doi: 10.1517/13543784.11.4.529. [DOI] [PubMed] [Google Scholar]
- 31.Aathithan S, French GL. Prevalence and role of efflux pump activity in ciprofloxacin resistance in clinical isolates of Klebsiella pneumoniae. Eur J Clin Microbiol Infect Dis. 2011;30(6):745–752. doi: 10.1007/s10096-010-1147-0. [DOI] [PubMed] [Google Scholar]
- 32.Louw GE, Warren RM, Gey van Pittius NC, Leon R, Jimenez A, Hernandez-Pando R, et al. Rifampicin reduces susceptibility to ofloxacin in rifampicin-resistant Mycobacterium tuberculosis through efflux. Am J Respir Crit Care Med. 2011;184(2):269–276. doi: 10.1164/rccm.201011-1924OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Askoura M, Mottawea W, Abujamel T, Taher I. Efflux pump inhibitors (EPIs) as new antimicrobial agents against Pseudomonas aeruginosa. Libyan J Med. 2011;6:5870. doi: 10.3402/ljm.v6i0.5870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sun J, Deng Z, Yan A. Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochem Biophys Res Commun. 2014;453(2):254–267. doi: 10.1016/j.bbrc.2014.05.090. [DOI] [PubMed] [Google Scholar]
- 35.Blanco P, Hernando-Amado S, Reales-Calderon J, Corona F, Lira F, Alcalde-Rico M, et al. Bacterial multidrug efflux pumps: much more than antibiotic resistance determinants. Microorganisms. 2016;4(1):14. doi: 10.3390/microorganisms4010014. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data supporting the conclusions are included within the manuscript.