

ABSTRACT
Several bacteria use the plant hormone indole-3-acetic acid (IAA) as a sole carbon and energy source. A cluster of genes (named iac) encoding IAA degradation has been reported in Pseudomonas putida 1290, but the functions of these genes are not completely understood. The plant-growth-promoting rhizobacterium Paraburkholderia phytofirmans PsJN harbors iac gene homologues in its genome, but with a different gene organization and context than those of P. putida 1290. The iac gene functions enable P. phytofirmans to use IAA as a sole carbon and energy source. Employing a heterologous expression system approach, P. phytofirmans iac genes with previously undescribed functions were associated with specific biochemical steps. In addition, two uncharacterized genes, previously unreported in P. putida and found to be related to major facilitator and tautomerase superfamilies, are involved in removal of an IAA metabolite called dioxindole-3-acetate. Similar to the case in strain 1290, IAA degradation proceeds through catechol as intermediate, which is subsequently degraded by ortho-ring cleavage. A putative two-component regulatory system and a LysR-type regulator, which apparently respond to IAA and dioxindole-3-acetate, respectively, are involved in iac gene regulation in P. phytofirmans. These results provide new insights about unknown gene functions and complex regulatory mechanisms in IAA bacterial catabolism.
IMPORTANCE This study describes indole-3-acetic acid (auxin phytohormone) degradation in the well-known betaproteobacterium P. phytofirmans PsJN and comprises a complete description of genes, some of them with previously unreported functions, and the general basis of their gene regulation. This work contributes to the understanding of how beneficial bacteria interact with plants, helping them to grow and/or to resist environmental stresses, through a complex set of molecular signals, in this case through degradation of a highly relevant plant hormone.
KEYWORDS: indole-3-acetic acid catabolism, iac genes, Paraburkholderia phytofirmans, plant-growth-promoting rhizobacteria
INTRODUCTION
Indole-3-acetic acid (IAA) belongs to a class of plant hormones called auxins, which play a key role in plant growth and development, controlling cell division, elongation, differentiation, and tropism responses to gravity and light (1, 2). A crucial characteristic of auxin-mediated effects is related to the differential distribution of auxin in tissues (2); therefore, plants tightly control IAA levels through biosynthesis, conjugation, degradation, and intercellular transport (3). Aside from auxin being an essential molecule for plants, auxin production is widespread among microorganisms present in soil, water, and plant and animal hosts (4–6). Functions of bacterial IAA synthesis have primarily been associated with plant-microbe interactions, although it has been proposed that they also play a role in interaction with animal hosts and molecular signaling between microorganisms (4–6). Not only do bacteria have the ability to synthesize IAA, but a substantial diversity of microorganisms also possess the ability to transform or degrade IAA (7), raising questions about the ecological significance of auxin metabolism and turnover in microorganisms.
An anaerobic auxin degradation pathway has been proposed in Azoarcus evansii and Aromatoleum aromaticum (8), and complete aerobic degradation of this auxin has been reported in Bradyrhizobium (9, 10); Pseudomonas (11); Burkholderia, Rhodococcus, and Sphingomonas (12); and Acinetobacter (13) species. Although two routes for aerobic IAA degradation were proposed some time ago in Bradyrhizobium japonicum (9, 10), with anthranilic acid as a key intermediate, genetic and additional biochemical aspects of aerobic IAA degradation have only been recently addressed (12, 14). Leveau and Gerards (12) were the first to describe genes related to IAA degradation in Pseudomonas putida 1290, which carries the iacABCDEFGRHI gene cluster, encoding conversion of IAA to catechol as intermediate (Fig. 1). After initial functional analysis of some iac genes in P. putida 1290, Scott et al. (14) proposed two putative intermediates based on the fact that the iacA gene product would start IAA degradation, hydroxylating IAA in position 3 of the indole ring, thus generating 2-hydroxyindole-3-acetic acid by migration of the hydroxyl group from indole ring position 3 to position 2 (Fig. 1A). They also suggested that the iacE gene product would be involved in the next step, transforming 2-hydroxyindole-3-acetic acid to 3-hydroxy-2-oxindole-3-acetic acid (dioxindole-3-acetic acid [DOAA]), which would be the substrate for the iacC gene function (Fig. 1A), in a rate-limiting step that also requires the iacD gene function, proposed as a subunit of the iacC gene product, based on sequence similarity analysis (14). In addition, catechol was identified as the end product of this IAA degradation pathway (Fig. 1A) (12, 14). More recently, the iacR gene from Acinetobacter baumannii, encoding a putative MarR-type transcriptional regulator, was reported to control iac gene repression in the absence of IAA (15). The roles of other iac genes remain unknown.
FIG 1.

Indole-3-acetic acid (IAA) degradation pathways and gene clusters involved in IAA catabolism in Paraburkholderia phytofirmans PsJN and Pseudomonas putida 1290. (A) P. putida 1290 route channeling IAA to catechol (14), with iacA, iacE, and iacC genes encoding indicated catabolic steps. (Adapted from reference 14.) (B) iac genes involved in IAA catabolism. Black arrows indicate putative coding genes directly involved in IAA degradation, gray arrows indicate probable transporters of IAA metabolites, and dashed arrows indicate putative regulators of IAA degradation genes. Genes involved in catechol catabolism are shown in brackets. Thin arrows indicate putative promoter sequences from iac genes cloned to construct the corresponding lacZ transcriptional fusions.
iac gene homologues are present in several Proteobacteria and Actinobacteria but with different gene organizations (12). The well-known plant-growth-promoting rhizobacterium (PGPR) Paraburkholderia (formerly Burkholderia [16]) phytofirmans PsJN (17–19) possesses a cluster of iac genes related to those described in P. putida 1290 (Fig. 1B) (12) but with additional genes encoding putative regulatory, transport, and enzymatic functions. Remarkably, P. phytofirmans also synthesizes IAA (20) through at least two putative IAA biosynthesis routes (21), but additional studies are clearly required to understand IAA synthesis in this strain. Studies in P. phytofirmans PsJN indicate that production of the enzyme 1-aminocyclopropane-1-carboxylate deaminase, signaling via N-acylhomoserine lactones (quorum sensing), tryptophan-dependent IAA biosynthesis, and IAA degradation all play significant roles in plant colonization and plant growth promotion (19–22). Understanding the role of IAA in plant growth promotion in this and other bacteria is clearly precluded by the scarce knowledge on both the biochemical and genetic aspects of IAA degradation.
We reported here that P. phytofirmans PsJN mineralizes IAA using iac gene homologue-encoded functions. The role of some previously uncharacterized iac genes is also reported. IAA degradation by strain PsJN apparently uses some functionally redundant genes and other genes not previously reported in P. putida 1290, encoding transport and tautomerase superfamily enzymes. We also reported that, in contrast with P. putida and A. baumannii, where iac gene regulation is controlled by a MarR-type transcriptional repressor responding to the IAA molecule (14, 15), IAA degradation in P. phytofirmans is regulated by a two-component system and a LysR-type activator, which apparently respond to IAA in combination with DOAA, the previously proposed intermediate in strain 1290 (14).
RESULTS AND DISCUSSION
Genetic determinants involved in growth of P. phytofirmans PsJN on IAA.
To evaluate mineralization of IAA by strain PsJN, growth tests were carried out in minimal medium cultures containing exogenously supplied auxin. Results showed that strain PsJN uses IAA as a sole carbon and energy source (Fig. 2). P. phytofirmans also utilized IAA as a sole nitrogen source, since it grows in a minimal medium with no added nitrogen sources (data not shown). Strain PsJN presented maximum specific growth rates (μmax) (doubling times) of 0.21 ± 0.01 h−1 (3.24 ± 0.21 h) and 0.12 ± 0.01 h−1 (5.95 ± 0.16 h) with 2.5 and 5 mM IAA, respectively (Fig. 2). In contrast, strain PsJN exhibited μmax (doubling times) of 0.42 ± 0.15 h−1 (1.83 ± 0.67 h) and 0.38 ± 0.17 h−1 (2.09 ± 0.78 h) with 2.5 and 5 mM benzoate (Bz), respectively (Fig. 2). Growth yields were estimated in 1.15 ± 0.19 and 1.89 ± 0.42 mg of cells/mmol of added carbon, for IAA and Bz, respectively.
FIG 2.
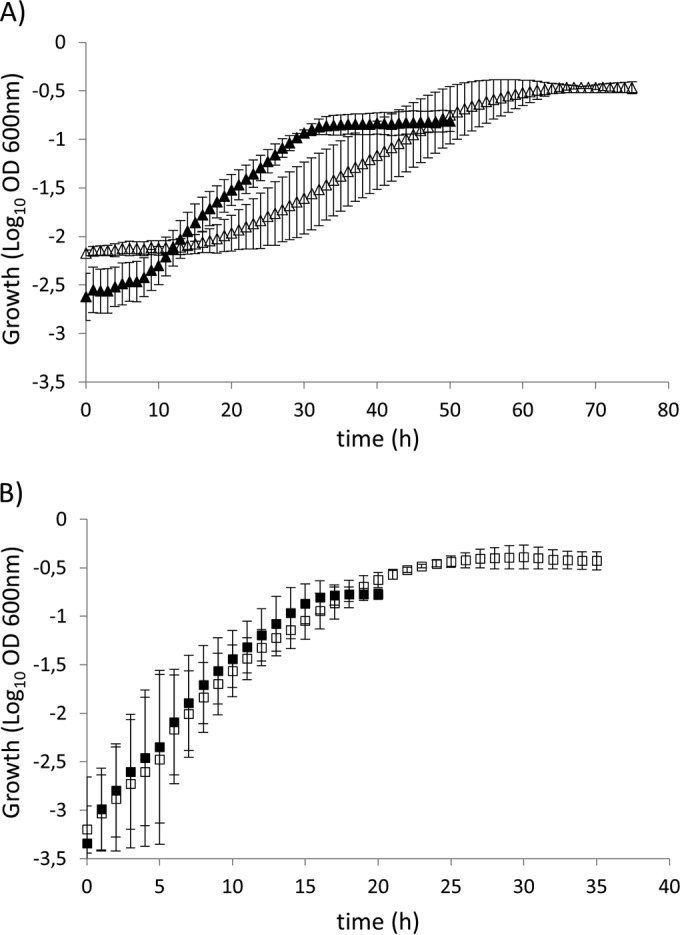
Growth of Paraburkholderia phytofirmans PsJN on indole-3-acetic acid (IAA) (A) or benzoate (Bz) (B) as the sole carbon and energy source. Symbols: closed squares, 2.5 mM Bz; open squares, 5 mM Bz; closed triangles, 2.5 mM IAA; open triangles, 5 mM IAA. Three biological replicates were performed for growth measurements. Error bars represent standard deviations of the means.
Lag-phase values for growth on IAA, calculated according to the method of Baty et al. (23), were 8.26 ± 0.18 h and 18.95 ± 5.54 h in 2.5 mM and 5 mM IAA, respectively. In contrast, growth on Bz exhibited lag phases of 3.61 ± 1.54 h (2.5 mM Bz) and 4.09 ± 2.81 h (5 mM Bz) (Fig. 2). The lag phase of strain PsJN for growth on IAA is also longer than that of P. putida 1290 (12), which may be explained by a tightly controlled iac gene induction to avoid formation of higher levels of toxic intermediates. In this context, accumulation of catechol, a toxic intermediate preventing bacterial growth (24–26), was reported by Scott et al. (14) and also detected in this study (see below). Since the same concentrations of IAA (a 10-carbon-atom molecule) and Bz (a seven-carbon molecule) produce similar biomass yields (Fig. 2), with the two compounds sharing the six-carbon-molecule catechol as a key intermediate, it is also possible that IAA degradation could generate some dead-end metabolites or CO2 as by-products, which are not utilized by cells.
Search of IAA degradation molecular determinants in the genome of strain PsJN allowed detection of P. putida 1290 iac gene homologues but with a different organization from that of strain 1290 (Fig. 1B) (12). Comparisons between iac homologues showed amino acid identities ranging from 38 to 62% (see Table S1 in the supplemental material). Only the iacR gene strain 1290 homologue, a transcriptional repressor belonging to the MarR family (14, 15), is absent in the strain PsJN genome. The strain PsJN iac gene sequences are adjacent to other genes, not found in P. putida 1290, possibly playing regulatory and transport roles in IAA degradation (Fig. 1B). Among these are a histidine kinase signal transducer-encoding gene (named iacS) and a LuxR family protein receptor (iacR1), forming a putative two-component system; a transcriptional regulator belonging to the LysR family (iacR2); a transporter belonging to the major facilitator superfamily (iacT1); and a protein related to the tautomerase superfamily (iacY) (see Table S2 in the supplemental material).
To test if these iac genes are involved in IAA degradation, the corresponding strain PsJN mutants were generated and growth tests were performed with IAA as a sole carbon and energy source. With the exceptions of the iacG gene, the putative regulatory iacS gene, the putative transporter iacT1 gene, and the iacY gene, all other iac genes (iacA, -B, -C, -D, -E, -F, -H, and -I) were required for growth of strain PsJN on IAA, as no growth of the corresponding mutant strains was detected, even if cells were cultured for 7 days (see Table S3 in the supplemental material). It was previously reported in P. putida 1290 that the iacH gene was not essential for IAA degradation (12), suggesting that insertional mutation (single crossover employing a suicide plasmid) could cause polar effects in strain PsJN over genes that would belong to the same transcriptional unit, i.e., iacABIHE and iacCDYT1 genes (Fig. 1B; see Fig. S1 in the supplemental material). In order to evaluate this possibility, the iacH mutant strain was complemented with the iacH gene or the contiguous downstream iacE gene. IAA growth was recovered only in the iacE-complemented strain (Table S3), strongly indicating that interruption of the iacH gene caused a polar effect over the iacE gene and supporting the idea that iacH was not essential for IAA degradation in strain PsJN, at least under the tested conditions.
To verify if other iac genes that may be organized as an operon (iacA, iacB, iacC, and iacI [Fig. 1B; Fig. S1]) are key in the IAA degradation pathway, mutant strains of these genes were supplemented with plasmid constructions containing all other iac genes (iacABIHECDGF), with or without the respective interrupted gene (plasmids in Table 1). Growth on IAA was recovered only in the presence of the corresponding interrupted genes, showing that iacA, iacB, iacC, and iacI genes are essential in the IAA degradation pathway (Table S3). In turn, growth on IAA of the iacG and iacS gene mutants was only partially diminished (Table S3), suggesting that their functions were in part replaced by other genes carried in the genome of strain PsJN. Remarkably, iacA, iacG, and iacT1 genes possess homologues in the P. phytofirmans genome, referred to as iacA2 (65% amino acid identity with iacA), iacG2 (63% amino acid identity with iacG), and iacT2 (51% amino acid identity with iacT1), which are clustered together (Fig. 1B). However, single mutants of these genes have no influence on growth of IAA, at least under the tested conditions (Table S3).
TABLE 1.
Bacterial strains and plasmids used in this study
| Strain or plasmid | Relevant phenotype and/or genotypea | Reference or source |
|---|---|---|
| Paraburkholderia phytofirmans strains | ||
| PsJN | IAA+ Bz+ fructose+ | 17 |
| PsJN::pCR2.1iacA | IAA− Kmr | This study |
| PsJN::pCR2.1iacB | IAA− Kmr | This study |
| PsJN::pCR2.1iacC | IAA− Kmr | This study |
| PsJN::pCR2.1iacD | IAA− Kmr | This study |
| PsJN::pCR2.1iacE | IAA− Kmr | This study |
| PsJN::pCR2.1iacF | IAA− Kmr | This study |
| PsJN::pCR2.1iacG | IAA+ Kmr | This study |
| PsJN::pCR2.1iacH | IAA− Kmr | This study |
| PsJN::pCR2.1iacI | IAA− Kmr | This study |
| PsJN::pCR2.1iacA2 | IAA+ Kmr | This study |
| PsJN::pCR2.1iacG2 | IAA+ Kmr | This study |
| PsJN::pCR2.1iacT1 | IAA+ Kmr | This study |
| PsJN::pCR2.1iacT2 | IAA+ Kmr | This study |
| PsJN::pCR2.1iacY | IAA+ Kmr | This study |
| PsJN::pCR2.1iacS | IAA+ Kmr | This study |
| PsJN::pCR2.1iacR1 | IAA− Kmr | This study |
| PsJN::pCR2.1iacR2 | IAA− Kmr | This study |
| PsJN::pCR2.1catA | IAA− Bz+ Kmr | This study |
| Other strains | ||
| Cupriavidus pinatubonensis | ||
| JMP134 | IAA− fructose+ | DSMZ (28) |
| JMP134iac1 | Strain expressing iacABIHECDGF genes driven by the heterologous PBAD promoter, Gm+ | This study |
| Escherichia coli Mach | ΔrecA1398 endA1 tonA ϕ80ΔlacM15 ΔlacX74 hsdR(rK− mK+) | Invitrogen, Carlsbad, CA, USA |
| Plasmids | ||
| pCR2.1-TOPO | Suicide vector in P. phytofirmans PsJN, Apr Kmr | Invitrogen, Carlsbad, CA, USA |
| pCR2.1iacb | pCR2.1 derivatives with internal fragment of iac genes, Apr Kmr | This study |
| pCR2.1catA | pCR2.1 derivative with internal fragment of catA gene, Apr Kmr | This study |
| pKGWP0 | BHR lacZ transcriptional fusion vector, Spr | 68 |
| pIacA-lacZ | PiacA pKGWP0 derivative Spr | This study |
| pIacC-lacZ | PiacC pKGWP0 derivative Spr | This study |
| pIacF-lacZ | PiacF pKGWP0 derivative Spr | This study |
| pIacG-lacZ | PiacG pKGWP0 derivative Spr | This study |
| pCatB-lacZ | PcatB pKGWP0 derivative Spr | This study |
| pBS1 | BHR gateway destination vector, araC-PBAD Gmr | 67 |
| pBS1iac1 | iacABIHECDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔA | iacBIHECDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔB | iacAIHECDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔCD | iacABIHEGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔE | iacABIHCDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔF | iacABIHECDG-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔG | iacABIHECDF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔH | iacABIECDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1iac1ΔI | iacABHECDGF-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacA | iacA-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacAG | iacAG-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacA2 | iacA2-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacA2G2 | iacA2G2-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacE | iacE-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacB | iacB-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacEB | iacEB-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacC | iacC-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacCD | iacCD-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacCDF | iacCDF-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacCDFI | iacCDFI-expressing pBS1 derivative, Gmr | This study |
| pBS1-iacH | iacH-expressing pBS1 derivative, Gmr | This study |
| p0-iacYT1 | iacYT1-expressing pKGWP0 derivative, Spr | This study |
IAA+, Bz+, and fructose+, ability to grow on indole-3-acetate, benzoate, and fructose, respectively; Apr, Gmr, Kmr, and Spr, resistance to ampicillin, gentamicin, kanamycin, and spectinomycin, respectively; BHR, broad host range.
Internal fragments of iacA, iacB, iacC, iacD, iacE, iacF, iacG, iacH, iacI, iacA2, iacG2, iacT1, iacT2, iacY, iacS, iacR1, and iacR2 cloned in pCR2.1 plasmid, to obtain their respective derivatives.
P. putida 1290 mutants for ortho-ring cleavage catechol degradation turn IAA growth test plates brown (12), and cell extracts of P. putida 1290 grown on IAA show elevated levels of catechol 1,2-dioxygenase activity (11), suggesting that IAA catabolism in strain 1290 produces catechol as intermediate, a possibility that was later confirmed by Scott et al. (14) through gas chromatography-mass spectroscopy analysis. The genome of P. phytofirmans PsJN has a putative ortho-ring cleavage catechol pathway gene cluster, encoding catechol 1,2-dioxygenase (catA), muconate cycloisomerase (catB), and muconolactone isomerase (catC), located adjacent to the iac gene cluster (Fig. 1B; Table S2), presumably involved in the degradation of catechol via cis,cis-muconic acid (27). A P. phytofirmans PsJN catA mutant was generated to evaluate if this putative catechol 1,2-dioxygenase effectively contributes to IAA degradation. The strain PsJN catA mutant turned fructose plates brown when supplemented with IAA (see Fig. S2 in the supplemental material), suggesting catechol accumulation and polymerization (24). This catA mutant was completely unable to grow on IAA (Table S3), strongly indicating its involvement in IAA degradation. A second putative catechol 1,2-dioxygenase-encoding gene (catA2) is also present in the strain PsJN genome, clustered with Bz 1,2-dioxygenase genes (Table S2) and therefore putatively involved in Bz catabolism. The catA2 gene seems to not be directly participating in IAA degradation because the catA gene mutant completely lost this ability. As expected, the catA mutant still grows on Bz (Table S3). When this catA mutant was exposed to 2.5 mM IAA plus 0.25 mM Bz (as an inducer of the additional CatA2 catechol 1,2-dioxygenase gene), this mutant recovered the ability to grow on IAA but turned the growth medium brown (data not shown), demonstrating that catechol accumulated by iac genes activity is partially metabolized by CatA2.
P. phytofirmans PsJN IAA degradation pathway.
Scott et al. (14) reported that the iacA, iacE, and iacC gene products are involved in consecutive steps of the IAA degradation pathway in P. putida 1290, whereas the roles of other iac genes remained unclear. To advance the elucidation of biochemical steps encoded by all strain PsJN iac genes in IAA degradation, several iac gene sets were cloned into the strain Cupriavidus pinatubonensis JMP134, which is heterologous but belongs to the same family (Burkholderiaceae). This strain, taxonomically and metabolically related to P. phytofirmans PsJN (28–30), lacks iac gene sequence homologues and is unable to use IAA as a sole carbon and energy source (Table S3), which allow its use as an appropriate heterologous host for iac genes. A strain JMP134 derivative carrying the P. phytofirmans iacABIHECDGF genes (named strain JMP134iac1) turned fructose growth test plates brown when supplemented with IAA, suggesting IAA removal and catechol accumulation comparable to those of the strain PsJN catA mutant (Fig. S2). As strain JMP134 has functional catA genes, most likely they were not induced by catechol produced from IAA under the tested conditions. Resting cell assays of strain JMP134iac1 demonstrated IAA consumption and transient catechol accumulation in high-performance liquid chromatography UV detection (HPLC-UV) profiles (see Fig. S3 in the supplemental material). Nevertheless, an additional intermediate was detected in these assays (initially designated compound 1 [Fig. S3]) that was not removed even after 43 h of incubation. In order to identify compound 1 as an intermediate during IAA degradation (see below), the accumulated metabolite was extracted from the JMP134iac1 supernatant organic phase at low pH using ethyl acetate, allowing purification of a single compound as detected by thin-layer chromatography (data not shown). Determination of the relative mass formula of the purified metabolite by mass spectrometry indicated that m/z of the molecular ion peak was 207.20, while infrared analysis showed that the molecule has -OH and CO- functional groups (infrared [IR] [KBr] cm−1, 3,313 [OH]; 1,702 [CO]). Analysis by 1H and 13C nuclear magnetic resonance (NMR) spectroscopy suggested that purified compound 1 is 2-(3-hydroxy-2-oxoindolin-3-yl)acetic acid (also named dioxindole-3-acetic acid) (see Table S4 in the supplemental material) comparable to that proposed by Scott et al. (14) (Fig. 1A). The molecular formula of this compound is C10H9NO4 with a calculated mass of 207.18 g/mol, which is quite close to the relative mass formula indicated above, strongly suggesting that the compound proposed by Scott et al. (14) is an IAA degradation product.
When strain PsJN IAA-grown cells were exposed to the supernatant containing compound 1, i.e., DOAA, fast and complete removal of this compound was observed (see Fig. S4 in the supplemental material), suggesting that strain JMP134iac1 lacks a function related to DOAA metabolism that is present in P. phytofirmans. A possible candidate for the missing function is the iacT1 gene (Fig. 1B; Table S2), encoding a putative transporter that could allow internalization of this intermediate to restart metabolism. Another possible candidate is the iacY gene (Table S2), located apparently in the same transcriptional unit next to the iacT1 gene (Fig. 1B; Fig. S1), encoding a protein belonging to a tautomerase superfamily. To test these possibilities, both the iacY and iacT1 gene sequences were cloned and introduced into the JMP134iac1 derivative accumulating DOAA. Results showed that only when the two genes were simultaneously present in the JMP134iac1 derivative was DOAA completely removed after 24 h of incubation (see Fig. S5 in the supplemental material). Remarkably, despite the fact that iacT1 and iacY mutant strains are able to use IAA as a sole carbon and energy source (Table S3), DOAA growth tests specifically performed with these mutants showed that they were unable to grow on DOAA, unlike the wild-type strain, indicating that the iacT1 and iacY genes are a functional part of the P. phytofirmans iac gene cluster, as their gene products play a role in DOAA removal in the biochemical route of IAA degradation. Database searches predicted that the iacT1 gene, and also the iacT2 gene found clustered with the iacA2 and iacG2 genes (Fig. 1B), belong to the major facilitator superfamily of transporters, which are single-polypeptide secondary carriers capable of transporting small solutes in response to chemiosmotic ion gradients (31), and specifically belong to the metabolite-H+ symporter family related to the shikimate transporter encoded by the shiA gene of Escherichia coli (∼40% amino acid identity) (32). These data suggest that the iacT1-encoded product is a DOAA transporter. In turn, the iacY gene would encode a protein belonging to a tautomerase superfamily characterized by catalytic promiscuity with a key catalytic amino-terminal proline (33). The iacY-encoded product is a long monomer related to the enzyme 4-oxalocrotonate tautomerase that catalyzes ketoenol tautomerization of 2-hydroxy-2,4-hexadienedioate (2-hydroxymuconate) to 2-keto-3-hexenedioate, which is part of the catabolic meta-cleavage pathway for aromatic compounds such as toluene or its derivatives (34). In the case of the iacY-encoded product, its role in DOAA metabolism remains elusive.
A series of additional incomplete JMP134iac1 derivatives were incubated with IAA, and the incubation supernatants were analyzed by HPLC-UV at different times. As expected, only the JMP134 strain carrying all iac genes except iacA (JMP134iac1ΔA) was unable to remove IAA (see Fig. S6 in the supplemental material), demonstrating that the iacA gene encodes the first step in the IAA degradation pathway of P. phytofirmans, in agreement with the findings reported by Lin et al. (13) and Scott et al. (14) in A. baumannii ATCC 19606 and P. putida 1290, respectively. Amino acid sequence analysis predicted that IacA would belong to the acyl coenzyme A (acyl-CoA) dehydrogenase flavoprotein family (13, 35). Conversion of indole into indigo (through indoxyl production) was demonstrated for the A. baumannii ATCC 19606 iacA gene product, whose reaction was absolutely dependent on NADH and flavin adenine dinucleotide (FAD) (13), supporting its classification as a flavoprotein. Moreover, Scott et al. (14) proposed that the IacA protein hydroxylates IAA to 2-hydroxyindole-3-acetic acid (Fig. 1A), indicating that IacA protein would have an IAA hydroxylase activity. Remarkably, the strain JMP134iac1ΔG derivative removed IAA significantly slower than strain JMP134iac1, as more than half of the initial IAA was still present after 10 h of incubation (Fig. S6). These results suggested that the iacA and the uncharacterized iacG gene products might work together and participate in the initial attack on IAA. To test this assumption, the iacA gene sequence was cloned in C. pinatubonensis JMP134, alone or in combination with the iacG gene, generating strain JMP134-iacA or -iacAG derivatives, respectively. Resting cells of strain JMP134-iacA exposed to IAA were unable to remove IAA (data not shown), whereas strain JMP134-iacAG cells quickly transformed IAA into different compounds (compounds X and Y [see Fig. S7A in the supplemental material]), supporting their role in the initial attack of IAA. Remarkably, the strain JMP134 derivative harboring redundant iacA2G2 genes (Fig. 1B), but not the JMP134 derivative carrying only the iacA2 gene, was also able to transform IAA to compounds X and Y (data not shown), suggesting that additional copies of iacAG are effectively functional. Cell extracts of derivatives JMP134-iacAG and -iacA2G2 confirmed consumption of NADH during IAA transformation, showing that enzymatic activity related to NADH conversion to NAD+ of the strain containing iacA2G2 was lower than that of the strain containing iacAG (data not shown). Interestingly, strain JMP134 derivatives carrying the iacAG genes but not the iacA2G2 genes have the capacity to transform indole into the blue insoluble compound named indigo (data not shown).
The role of the iacG gene does not seem to be essential for IAA degradation in P. phytofirmans because the iacG gene has at least two additional equivalents in the genome of strain PsJN (Table S1), one of them being functional (iacG2). Also, the IacG protein may have an accessory role, which is supported by the fact that the iacG gene sequence has the best hit (56% of amino acid identity) with the 4-hydroxyphenylacetate (4-HPA) 3-monooxygenase reductase component (hpaC gene) of E. coli (36), which is able to reduce FAD to FADH2, to dissociate from the enzyme, and later to be captured by the 4-HPA 3-monooxygenase (HpaB) to hydroxylate 4-HPA. Therefore, direct interaction between HpaB and HpaC is not critical, and thus, theoretically, any flavin reductase present in the host cell would replace HpaC (36). Correspondingly, IacG would provide reduced flavins to IacA in the first step of the IAA degradation pathway in P. phytofirmans in an NADH-dependent step (Fig. 3A).
FIG 3.

Putative roles of Paraburkholderia phytofirmans PsJN iac genes (A) and their regulation in auxin phytohormone indole-3-acetic acid (IAA) degradation (B). DOAA, dioxindole-3-acetic acid; in, intracellular space; out, extracellular space; ?, not known.
Additional experiments indicated that the strain JMP134iac1ΔE derivative completely removed IAA from the medium after 10 h, accumulating compounds X and Y (Fig. S6), similarly to the strain JMP134-iacAG derivative (Fig. S7A). Scott et al. (14) reported that the P. putida iacE gene product participates in the second step of the IAA pathway (Fig. 1A), encoding an enzyme involved in the formation of 3-hydroxy-2-oxindole-3-acetic acid (DOAA) from 2-hydroxyindole-3-acetic acid. Sequence analysis predicted that the iacE gene product of strain PsJN would be a classical member of the short-chain dehydrogenase/reductase (SDR) family, characterized by a Rossmann-fold scaffold, typically about 250 residues long, with specific cofactor (TGxxxGxG) and active site (YxxxK) sequence motifs and a reaction spectrum comprising the NAD(P)(H)-dependent oxidoreduction of hydroxy/keto groups (37). In turn, the strain JMP134iac1ΔB derivative exposed to IAA resulted in production of compounds X and Y but also detectable amounts of DOAA (Fig. S6), suggesting that IacE and IacB work together. To test this possibility, the ability of the strain JMP134-iacE, -iacB, or -iacEB derivatives to transform supernatants of strain JMP134-iacAG exposed to IAA, containing intermediate compounds X and Y (Fig. S7A), was determined. The strain JMP134-iacE and -iacEB derivatives transformed supernatants produced by the strain JMP134-iacAG derivative, generating DOAA, although the strain JMP134-iacEB derivative produced significantly larger amounts of DOAA (Fig. S7B), than the strain JMP134-iacE derivative (data not shown). These results strongly suggest that IacB may be an auxiliary or accessory protein of the IacE protein, as the strain JMP134-iacB derivative did not produce DOAA (data not shown). The P. phytofirmans iacB gene sequence analysis shows no conserved domains, and no function could be predicted from its primary structure, although a role of IacB as an IacE auxiliary or accessory protein cannot be dismissed, since production of DOAA would require a hydroxylation step (Fig. 1A) (19) not catalyzed by a classical dehydrogenase activity (37). Then, a second hydroxylation step in the IAA degradation pathway would be a result of the IacEB protein activity (Fig. 3A). However, a possible participation of iacAG gene products in a second hydroxylation on the IAA molecule cannot be discarded, since strain JMP134-iacAG exposed to IAA generated several compounds (Fig. S7A), which could correspond to interchangeable forms of the hydroxylated IAA, and therefore, IacEB may perform a subsequent hydroxy/keto reduction step to produce DOAA.
The strain JMP134iac1ΔCD, -iac1ΔF, and -iac1ΔI derivatives completely removed IAA and accumulated large amounts of DOAA (Fig. S6). When the strain JMP134-iacCDFI derivative was exposed to a supernatant containing only DOAA, this compound remained intact (data not shown), which clearly suggested that the iacT1 and iacY gene products (see above) are required before the participation of the iacCDFI gene products. To test this option, the strain JMP134-iacC, -iacCD, -iacCDF, or -iacCDFI derivatives additionally harboring iacY and iacT1 genes were exposed to supernatants containing DOAA. Results showed that strain JMP134-iacC and -iacCD derivatives were unable to transform DOAA (data not shown), whereas the strain JMP134-iacCDF derivative converted DOAA to catechol (Fig. S7C). It is worth noting that the strain JMP134-iacCDFI derivative transformed DOAA faster than the strain JMP134-iacCDF derivative, with no accumulation of catechol (data not shown). Sequence analysis showed that the iacC gene product of P. phytofirmans PsJN belongs to the Rieske nonheme iron oxygenase family (38, 39), whereas the iacD gene product belongs to the beta subunit of ring hydroxylating dioxygenases, which has a structure similar to that of NTF-2 and scytalone dehydratase (40, 41). Rieske nonheme iron oxygenase systems use nonheme Fe(II) to catalyze addition of hydroxyl groups to the aromatic ring, an initial step in oxidative degradation of aromatic compounds (38, 39), employing an electron transport chain to use NAD(P)H and activate molecular oxygen (38). Some oxygenase components contain a beta subunit, but with a purely structural function, although some reports suggest that they can influence substrate binding in some oxygenases (42). Then, alpha subunits are the catalytic components, carrying an N-terminal domain which binds a Rieske-like [2Fe-2S] cluster that accepts electrons from a reductase or ferredoxin and passes them to a C-terminal catalytic domain, which binds the nonheme Fe(II) for catalysis (38). Rieske nonheme iron oxygenase systems are constituted by additional components involved in transfer of an electron from NAD(P)H to O2: a reductase, a ferredoxin (only in three-component systems), and the previously indicated oxygenases (with oligomer α3 or α3β3) (38, 39). Sequence analysis of the iacF gene product revealed that it belongs to the ferredoxin-NADP reductase family (43), which contains an FAD or flavin mononucleotide (FMN) binding domain, an NADH binding domain, and a plant-type [2Fe-2S] cluster domain, catalyzing transfer of reducing equivalents between the NADP+/NADPH pair and the oxygenase component (38, 43). Consequently, the iacF gene product would be the reductase component of an aromatic ring hydroxylating dioxygenase, with the iacC and iacD genes as the oxygenase components (alpha and beta subunits, respectively). This assumption was supported by catechol production from DOAA in the strain JMP134 derivative carrying iacCDF genes plus iacY and iacT1 genes (Fig. S7C). Remarkably, the absence of the iacI gene in the strain JMP134iac1ΔI derivative produced an accumulation of DOAA (similar to the effect of the absence of the iacCD and iacF genes), and the strain PsJN iacI mutant was unable to use DOAA as sole carbon and energy source, suggesting a role in a biochemical step downstream from the reaction carried out by the iacY and iacT1 gene products. Amino acid sequence analysis of IacI showed similarity with members of the SnoaL-4 superfamily, which is a family of proteins that shares the SnoaL fold, mainly represented by polyketide cyclases catalyzing ring closure steps in polyketide antibiotic synthesis (44). The presence of the iacI gene in the strain JMP134 iacCDFYT1 gene derivative accelerated removal of DOAA. Thus, it is likely that IacI participates in the same or in a later IAA degradation step catalyzed by IacCDF (Fig. 3A).
Finally, the strain JMP134iac1ΔH derivative fully removed IAA and accumulated DOAA and catechol (data not shown), analogously to strain JMP134iac1, corroborating that the iacH gene-encoded product was not involved in IAA transformation to catechol under the tested conditions, similar to that reported in P. putida 1290 (12). Sequence analysis indicated that the iacH gene product of P. putida was related to amidases/amidohydrolases (12), analogous to that found for the strain PsJN homologue sequence. Auxin storage in plants includes conjugation to form IAA-amides, in which IAA is ligated to amino acids such as Ala, Asp, Phe, or Trp (45). Assays with a strain JMP134 derivative overexpressing the iacH gene dismissed the use of indole-3-acetamide or indole-3-acetonitrile (data not shown), although further research is needed to test a possible role of the iacH gene product in IAA-amide conjugate metabolism.
Analysis of iac and cat gene expression in P. phytofirmans PsJN.
To evaluate iac gene expression, a quantitative real-time PCR analysis of RNA extracted from mid-log-phase cells of P. phytofirmans PsJN growing on IAA, Bz, or fructose as a sole carbon and energy source was performed. Results showed that transcript levels of the iacA, iacC, iacF, and iacG genes in IAA-grown cells were at least 2 orders of magnitude higher than those in cells growing on fructose or Bz (Fig. 4). Transcript levels of the redundant iacA2 gene were also increased in IAA-grown cells, suggesting a functional role in IAA degradation. Remarkably, transcript levels of the iacS and iacR1 regulatory genes were also increased (approximately 50 times) in IAA-grown cells, whereas the iacR2 transcript levels increased only five times in comparison with cells growing without IAA (Fig. 4). Moreover, catA transcript levels in IAA-grown cells were about 500 times higher than those in fructose- or Bz-grown cells, indicating that this catechol 1,2-dioxygenase is strongly induced during growth on IAA. In contrast, catA2 transcript levels were not increased in IAA-grown cells but they were significantly induced in Bz-grown cells (Fig. 4), supporting the idea that the catA2 gene does not participate in auxin metabolism, as IAA is unable to induce this putative catechol 1,2-dioxygenase-encoding gene.
FIG 4.

Transcript levels of iac and cat genes from Paraburkholderia phytofirmans cells exposed to indole-3-acetic acid (IAA). Real-time PCR analysis of iac and cat genes in cells grown on fructose, benzoate, or IAA as a sole carbon and energy source. It should be noted that transcript levels of the ribosomal protein S7 (rpS7) and tryptophan dioxygenase (kynA) genes, used as expression controls, remained unchanged under these conditions. Transcript levels were normalized to the average value of the transcript levels in fructose treatment. 16S rRNA was used as a reference gene (internal control). Note different scales. All experiments were performed in three biological replicates. Error bars represent standard deviations. Different letters indicate statistically significant differences between treatments (one-way analysis of variance, P < 0.05; Tukey's test, P < 0.05).
Additional analysis of the iac-cat gene organization of P. phytofirmans PsJN showed the presence of putative iacABIHE, iacCDYT, iacF, iacG, and catBAC transcriptional units (Fig. 1B; Fig. S1) and made it interesting to analyze the promoter activity profile of IAA degradation genes. β-Galactosidase transcriptional fusion assays with putative promoters of these transcriptional units were performed. Results showed that all iac gene promoters tested, including the cat gene promoter, were more active in the presence of the mixture of IAA and supernatant containing DOAA or compounds X and Y as inducers than in the presence of IAA or the supernatant containing DOAA or compounds X and Y alone (Fig. 5A), suggesting a coordinated and presumably complex promoter regulation of iac/cat gene expression. Additional tests of the iac promoter activities were performed with P. phytofirmans regulatory mutants containing mutations in the iacS, iacR1, and iacR2 genes, exposed to mixtures of IAA and supernatant containing DOAA. Results showed that the iacC, iacF, and catB gene promoters were induced in the P. phytofirmans iacS or iacR1 gene mutants (Fig. 5B) and were not induced in the P. phytofirmans iacR2 gene mutant showing that iacC, iacF, and catB gene promoters are controlled by an iacR2-encoded regulator or another regulator controlled by IacR2 levels. Conversely, mixtures of IAA and supernatant containing DOAA did not induce the iacA and iacG gene promoters in the regulator mutant strains (Fig. 5B), indicating that these three iac regulators are involved in their control. Because iacR2-encoded regulators apparently handle all iac/cat promoters, quantitative real-time PCR analysis of RNA extracted from cells of the wild type and the iacR2 mutant growing on fructose and induced with IAA-DOAA mixtures was accomplished to find out if the iacR1 and iacS genes are controlled by IacR2. Results showed that cells with a background lacking the iacR2 gene were unable to induce iacR1 and iacS genes (see Fig. S8 in the supplemental material), demonstrating that transcription of both regulators is controlled by IacR2.
FIG 5.

Induction of iac and cat promoter activities from Paraburkholderia phytofirmans PsJN and derivative mutants by mixtures of indole-3-acetic acid (IAA) and dioxindole-3-acetic acid (DOAA) or compounds X and Y. β-Galactosidase activity levels obtained from transcriptional fusions of iacA, iacC, iacF, iacG, and catB genes exposed to IAA, DOAA, compounds X and Y or mixtures of IAA plus DOAA or compounds X and Y in the wild-type strain (A) and in the mutant derivatives defective in iacS, iacR1, or iacR2 (B). Compounds X and Y or DOAA was prepared using 4-fold-diluted supernatants of strain JMP134-iacAG or JMP134iac1 resting cells previously exposed to 1 mM IAA as the only carbon source for 2 h, respectively. Error bars indicate the standard deviations. WT, wild type; NI, noninduced. Different letters and asterisks indicate statistically significant differences between treatments (one-way analysis of variance, P < 0.05; Tukey's test, P < 0.05; Student's t test, P < 0.05).
Compared with P. putida 1290, where the iac gene cluster includes only a transcriptional repressor belonging to the MarR family, named iacR (14, 15), which is absent in the genome of P. phytofirmans PsJN, the regulation of IAA catabolism in strain PsJN is far more complex: a histidine kinase signal transducer and a LuxR family protein receptor (a putative two-component system), iacS and iacR1, respectively, and a LysR-type transcriptional regulator (LTTR), iacR2 (Fig. 1B). Only a few bacterial two-component systems involved in degradation of aromatic compounds have been characterized to date; they are related to toluene, biphenyl, and styrene degradation in Pseudomonas, Thauera, Azoarcus, and Rhodococcus strains (46–50). On the other hand, the LTTR family is a well-characterized group of transcriptional regulators, highly conserved and ubiquitous among bacteria (51), which have been involved in control of metabolism, cell division, quorum sensing, virulence, motility, and nitrogen fixation, among others (51–54). A larger subgroup of LTTRs is associated with degradation of aromatic compounds, in which regulators CatM and BenM of Bz catabolism are quite well studied (55, 56). It is reported here that the iacR1 and iacR2 genes are essential for P. phytofirmans IAA catabolism, whereas the lack of the iacS gene decreased catabolism of IAA. Based on their location in the iac gene cluster (Fig. 1B) and the promoter activities of the corresponding mutant strains reported here (Fig. 5B), it is quite possible that the iacS/iacR1 gene product pair is a two-component system, with the iacS gene product sensing added IAA, autophosphorylating, and then phosphorylating iacR1, which positively interacts with the iacA and iacG gene promoters (Fig. 5B). Interestingly, Leveau and Gerards (12) found that mutants in a gene with amino acid similarity to the kinase sensor of the two-component system CbrAB of Pseudomonas aeruginosa PAO1 are unable to use IAA. CbrAB-inactivated strain PAO1 derivatives are also unable to grow on several N-substrates, suggesting that CbrAB controls expression of catabolic pathways in response to changing intracellular C/N ratios (57). The results reported here allow us to propose that the iacR2 gene product is a classical LysR transcriptional regulator using DOAA generated by the iacEB-encoded products as a coinducer, thus inducing transcription of iacC, iacF, and catB genes (Fig. 4 and 5B). This system would also control transcription of iacR1 and iacS genes. The latter possibility is based on the fact that these genes showed an increased expression (∼50 times in comparison to the control) with IAA as an inducer, whereas the iacR2 gene exhibited a much lower induction level increase (Fig. 4), and that the P. phytofirmans iacR2 mutant is unable to induce any iac promoter and to produce iacS/iacR1 transcripts in the presence of the tested inducers (Fig. 5B; Fig. S8). In addition, IAA degradation cat genes in strain PsJN is associated with a CatR-like regulator (Fig. 1B), similar to CatM and BenM of Bz catabolism (55, 56). However, participation by this regulator remains unclear because strain PsJN cat genes are regulated by IacR2 (Fig. 5B). Additional research is required to find out if catR/iacR2-encoded regulators are able to interact.
Outlook.
The IAA degradation iac genes reported in P. putida 1290 are present in several bacteria belonging to Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, and Actinobacteria (12). The beneficial plant-related bacterium P. phytofirmans PsJN also harbors iac genes but with a different gene organization, including additional transport, enzymatic, and regulatory genes. Results reported here showed biochemical steps in which iac genes with unknown functions would be participating (Fig. 5A) and also showed two additional uncharacterized genes related to major facilitator and tautomerase superfamilies involved in removal of the DOAA intermediate (Fig. 1A and 3A). Additionally, iac genes encoding the initial attack on the IAA molecule would be redundant, as an iac-related second gene cluster is also present in P. phytofirmans. Apparent redundancy, indicated by results reported here for the iacA2 and iacG2 genes, as the iacA2 gene was induced in P. phytofirmans cells grown on IAA (Fig. 4), and indicated by the fact that the iacA2G2 genes expressed in combination enable the removal of IAA, may be utilized to extend the spectrum of plant-derived IAA-like molecules. Such a possibility would be supported by a different specificity of iacAG gene products, as the iacA2G2 gene products do not transform indole to indigo under the tested conditions, and the iacA2G2-encoded product has a lower enzymatic activity against IAA than does IacAG (data not shown). Furthermore, this apparent functional gene redundancy would provide a selective metabolic advantage associated with previously reported IAA toxicity over plant and animal membranes (58). For instance, two copies of (chloro)catechol 1,2-dioxygenase avoid accumulation of toxic (chloro)catechol intermediates (25, 26) or dead-end metabolite production, as in strains C. pinatubonensis JMP134 and Cupriavidus metallidurans CH34, which harbor two copies of phenol monooxygenases: one copy is associated with catechol 1,2-dioxygenase and the other with catechol 2,3-dioxygenase (30), depending on whether the provided substrate (e.g., phenol or methylphenols) is channeling through ortho (catechol 1,2-dioxygenase) or meta (catechol 2,3-dioxygenase) cleavage (59).
The iac gene regulation in P. phytofirmans PsJN is associated with an iacR2-encoded LysR-type regulator that would respond to a late metabolite (DOAA) of the IAA pathway, controlling transcription of iac genes related to later steps of IAA degradation and catechol ortho-ring cleavage (Fig. 3B) but also driving transcription of a putative two-component regulatory system, which apparently responds to the IAA molecule and regulates iac genes encoding early IAA degradation steps (Fig. 3B), thus suggesting complex regulatory mechanisms that would perfectly be operative in other iac gene-harboring bacteria (12) for controlling toxic intermediate accumulation and also IAA intermediate degradation.
Interestingly, it has been reported in Arabidopsis thaliana roots that 2-oxindole-3-acetic acid is a major primary IAA degradation metabolite (60), which is an isomer of the 2-hydroxyindole-3-acetic acid intermediate from the bacterial IAA degradation pathway (Fig. 1B), and DOAA derivatives which might regulate endogenous levels of IAA were detected in Citrus sinensis and Vicia faba (61, 62). Remarkably, A. thaliana plants exposed to IAA and to DOAA showed similar behavior (see Fig. S9 in the supplemental material), suggesting that this class of molecules is recognized by plants and could be present in plant root exudates and, therefore, available for plant-related bacteria carrying iac genes, including transporter- and tautomerase-related genes, participating in uptake and isomerization of DOAA-related molecules to be channeled through iac gene-encoded degradation.
The complex regulatory mechanisms in this PGPR could be related to its ecological importance and may be linked with its previously reported ability to synthesize IAA (19, 20, 22), raising questions about expression/coordination of IAA degradation and synthesis in natural environments.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.
Bacteria and plasmids used in this study are listed in Table 1. P. phytofirmans and its derivatives were grown at 30°C in mineral salts medium (63), supplemented with 2.5, 5, or 10 mM benzoate (Bz), IAA, or fructose, plus the appropriate antibiotics, kanamycin (Km; 50 μg ml−1), gentamicin (Gm; 30 μg ml−1), or spectinomycin (Sp; 100 μg ml−1). Cupriavidus pinatubonensis JMP134 (Table 1) and its derivatives were grown under the same conditions. Escherichia coli Mach (Invitrogen, Carlsbad, CA, USA) was grown at 37°C in Luria-Bertani (LB) medium. Growth was measured by optical density at 600 nm (OD600). For preparation of a nitrogen-free mineral salts medium, (NH4)2SO4 was removed and Ca(NO3)2·4H2O was replaced by CaSO4·2H2O (63). At least three biological replicates were performed for each growth measurement. Growth yield averages were estimated considering an OD600 of 0.6 equivalent to 0.1 mg of cells, based on the information that average bacterial weight is 10−12 g (64) and that an OD600 of 0.6 in strain PsJN corresponds to approximately 1 × 108 cells/ml.
Chromosomal disruption of gene sequences in P. phytofirmans PsJN.
Internal fragments of iac and associated genes (iacA, iacA2, iacB, iacC, iacD, iacE, iacF, iacG, iacG2, iacH, iacI, iacS, iacR1, iacR2, iacY, iacT1, iacT2, and catA) were amplified by PCR, using primer pairs listed in Table 2. The PCR products were cloned using the pCR2.1-TOPO system (Invitrogen, Carlsbad, CA, USA) to generate plasmids listed in Table 1. For gene inactivation, suicidal pCR2.1iac or pCR2.1catA plasmids (Table 1) were electroporated in P. phytofirmans PsJN to get a one-recombination-event disruption of the target gene, resulting in mutants (Table 1) which were selected on LB agar containing 50 μg ml−1 Km. Correct insertions in all mutants were confirmed by PCR using primer pairs located in genomic DNA and suicidal plasmid DNA and subsequent sequencing.
TABLE 2.
Primer pairs used in this study
| Purpose | Forward |
Reverse |
||
|---|---|---|---|---|
| Primer name | Sequence (5′→3′) | Primer name | Sequence (5′→3′) | |
| Inactivation of genes | Mut iacAFw | GCCCAGTTTCTCGACATGAT | Mut iacARv | GTCGTCTGCAACGAACTGGT |
| Mut iacBFw | CGAACAGATCGGCGAAGT | Mut iacBRv | CATAGCGCCACAGGTTGTATT | |
| Mut iacCFw | GGTCA ACGTCTTGCA GAACC | Mut iacCRv | GTTTCGTCGTCGATCGATTT | |
| Mut iacDFw | GACTATCGCGAGTGGCTCA | Mut iacDRv | CTTGCAAAACTGATGCGATG | |
| Mut iacEFw | AAGTCGTCTCCGTGATGGAC | Mut iacERv | GATCACCGCCTTCAGTTTGT | |
| Mut iacFFw | AACAGGAGCGCTTCGTATTG | Mut iacFRv | GCAAACCGTTCGAGATGAAT | |
| Mut iacGFw | TCGACGTCAAACAGCAATTC | Mut iacGRv | ACCGAAGCGTTCTTCCATC | |
| Mut iacHFw | ATCGGATTGAAGCCCACTTT | Mut iacHRv | CGAGGTCTCGACATTGATGA | |
| Mut iacIFw | GAAATTCTCGCGATGCTCAC | Mut iacIRv | GAAAGTGCGTGATGAACCAG | |
| Mut iacA2Fw | GTCGAGAACTGGAACGTGGT | Mut iacA2Rv | CAGTTGGCTTCTGCTTTCG | |
| Mut iacG2Fw | CCAGCTACGTGCATGACATC | Mut iacG2Rv | AATCTGCGCAAACATGACC | |
| Mut iacT1Fw | GGTGGAAAGCTTCATGTCGT | Mut iacT1Rv | GAGGAACACCAGCATTGGTT | |
| Mut iacT2Fw | AGCATTCTGCTGATTCACGA | Mut iacT2Rv | TTGAGAACGCGAAGAAGATG | |
| Mut iacYFw | ATTGATCGCGGCTCTCAC | Mut iacYRv | GCGATCAAGGCTTCTTTTTG | |
| Mut iacSFw | TCAGCGTGGAAGACACACTC | Mut iacSRv | GTTGTAACGTCCGGCGTATT | |
| Mut iacR1Fw | GTGCAAGCGGAGTTGAATC | Mut iacR1Rv | CTTCACGCTGATCGACAGAT | |
| Mut iacR2Fw | CGCTGGAACTGTCGACTATC | Mut iacR2Rv | GAAACGCCTTCAAACTCAGC | |
| Mut catAFw | TCTGGGTTTCGAGCATTTTC | Mut catARv | CGGAAGCTGTATTTGCCTTC | |
| RT-PCRa analysis | iacA intFw | GCCCAGTTTCTCGACATGAT | iacA intRv | AGATTTCCGCTTGTGTATGC |
| iacC intFw | GATGGTTATGAAGGCGTGGT | iacC intRv | GGCTCGATCTCCTGATTGAA | |
| iacF intFw | CTGATGCGATTCATCTCGAA | iacF intRv | ATGGTCGTGTCCTTGTCCAC | |
| iacG intFw | GCGACTTTCCAGAGCAACC | iacG intRv | ACCGAAGCGTTCTTCCATC | |
| iacA2 intFw | GTCGAGAACTGGAACGTGGT | iacA2 intRv | CAGTTGGCTTCTGCTTTCG | |
| iacR1intFw | GTGCAAGCGGAGTTGAATC | iacR1intRv | CTTCACGCTGATCGACAGAT | |
| iacR2intFw | CGCTGGAACTGTCGACTATC | iacR2intRv | GAAACGCCTTCAAACTCAGC | |
| iacSintFw | TTGTCCCTTCGTTTGTCCTT | iacSintRv | CCAGGTGGTGTTGAATCTGG | |
| catA intFw | ACGTACAACGGACCTTCGAT | catA intRv | TCTGGGTTTCGAGCATTTTC | |
| catA2 intFw | TTCAACAAGCTCGGACAGG | catA2 intRv | ACATACAGCGGACCTTCGAT | |
| kynA intFw | TGAAGCTCGCGTTGTATGAG | kynA intRv | TCCGATAAACCTCCAACCAG | |
| rpS7 intFw | CGAACAGATCCAGACCAAGG | rpS7 intRv | ACTTCTCGCTGCGCTTCTT | |
| 16S intFw | CGGGCTAGAGTATGGCAGAG | 16S intRv | CGTGCATGAGCGTCAGTATT | |
| Transcriptional fusions | PiacA Fw | GCGCGGTACCCAGGCGGTGAAAGTCTTCTT | PiacA Rv | TGTCCTCGAGGGTGAGGTATCTCCTTGATTCG |
| PiacC Fw | GCGCGGTACCGAATGAAAAGCGGGTCGTC | PiacC Rv | TCGCCTCGAGCTTGTGTAGGTGGGGGTGGT | |
| PiacF Fw | TAGAGGTACCGATTTGAATGTTCCGACTGGA | PiacF Rv | CGTTCTCGAGGGTTCTTTCTATGAGCTTGC | |
| PiacG Fw | TTATGGTACCCGAAGGCTGGGACATGGT | PiacG Rv | ATTTCTCGAGGATTCCTCGGGCAGGTGT | |
| PcatB Fw | GTTTGGTACCTTCCTCGATCTGCT GGATCT | PcatB Rv | ATTCCTCGAGCTCCTGAACTGCCTGACCTG | |
| iac gene expression | 1 (iacABIHE Fw) | TTGGGCTAGCGAATTCCTGCTTACGTTTCGATCCAACTTGC | 2 (iacABIHE Rv) | GTGGTCGATGAAGCGAAGTCGTCAGCGCATGAAGAGTCC |
| 3 (iacCD Fw) | GACTTCGCTTCATCGACCAC | 4 (iacCD Rv) | CAGGTGTCGAGTGGTTTGATTTCGAAAAGATCAAAGCAGGAA | |
| 5 (iacG Fw) | AATCAAACCACTCGACACCTG | 6 (iacG Rv) | GGTTGTGCGTCAAGGATAAGCCACTGTGCGTAATCAATGC | |
| 7 (iacF Fw) | CTTATCCTTGACGCACAACC | 8 (iacF Rv) | GTAATACGACTCACTATAGGAAGAAAAGGCGCTTGTGAAA | |
| 9 (pBS1 Fw) | CCTATAGTGAGTCGTATTAC | 10 (pBS1 Rv) | GCAGGAATTCGCTAGCCCAA | |
| 11 (iacB Fw) | TTGGGCTAGCGAATTCCTGCCTATCCGCACAACCAGGATT | 12 (iacA Rv) | AATCCTGGTTGTGCGGATAG | |
| 13 (iacI Fw) | CTATCCGCACAACCAGGATTCTTGAGCGTTTCGGAACAAC | 14 (iacH Rv) | GTGGTCGATGAAGCGAAGTCCCACTCCACTTCGAAATAACG | |
| 15 (iacE Rv) | CAGGTGTCGAGTGGTTTGATTATGCCGCTTTGGTGTTGT | 16 (iacG Rv2) | GTAATACGACTCACTATAGGCCACTGTGCGTAATCAATGC | |
| 17 (iacF Fw2) | TTCCTGCTTTGATCTTTTCGACTTATCCTTGACGCACAACC | 18 (iacI Rv) | CCTATCGACAGCGTCTGGA | |
| 19 (iacE Fw) | TCCAGACGCTGTCGATAGGGCCGTTATTTCGAAGTGGAG | 20 (iacB Rv) | TCCTGGTTCGCGTTCATT | |
| 21 (iacH Fw) | AATGAACGCGAACCAGGAAGGTCAACGCAAGGAAACC | 22 (iacA Rv2) | GTAATACGACTCACTATAGGAATCCTGGTTGTGCGGATA | |
| 23 (iacA Rv3) | CAGGTGTCGAGTGGTTTGATTAATCCTGGTTGTGCGGATAG | 24 (iacA2 Fw) | TTGGGCTAGCGAATTCCTGCAGCTTCTCCTGTACCGATCC | |
| 25 (iacA2 Rv) | GTAATACGACTCACTATAGGTTCCTTTTCCTTTTGCATGG | 26 (iacG2 Rv) | GTAATACGACTCACTATAGGGATGAAAGATCCGGAGAGCA | |
| 27 (iacE Fw2) | TTGGGCTAGCGAATTCCTGCGCCGTTATTTCGAAGTGGAG | 28 (iacE Rv2) | GTAATACGACTCACTATAGGATGCCGCTTTGGTGTTGT | |
| 29 (iacB Rv2) | GTAATACGACTCACTATAGGTCCTGGTTCGCGTTCATT | 30 (iacE Fw3) | AATGAACGCGAACCAGGAGCCGTTATTTCGAAGTGGAG | |
| 31 (iacC Fw) | TTGGGCTAGCGAATTCCTGCGACTTCGCTTCATCGACCAC | 32 (iacC Rv) | GTAATACGACTCACTATAGGAGCGTGTTTCTGTCGTCCAT | |
| 33 (iacD Rv) | GTAATACGACTCACTATAGGTCGAAAAGATCAAAGCAGGAA | 34 (iacF Rv1) | AAGAAAAGGCGCTTGTGAA | |
| 35 (iacI Fw2) | TTTCACAAGCGCCTTTTCTTCTTGAGCGTTTCGGAACAAC | 36 (iacI Rv2) | GTAATACGACTCACTATAGGCCTATCGACAGCGTCTGGA | |
| 37 (iacH Fw2) | TTGGGCTAGCGAATTCCTGCAGGTCAACGCAAGGAAACC | 38 (iacH Rv2) | GTAATACGACTCACTATAGGCCACTCCACTTCGAAATAACG | |
| 39 (iacY Fw) | ACAGGATGAGGATCGTTTCGATCGGCTTCCTGCTTTGAT | 40 (iacT1 Rv) | CAAGAAAGCTGGGTCGAATTGGCTCGAATGCAGCTTCTC | |
| 41 (pKan Fw) | CTAAGTAAGTAAGGAGAAGGGCGGGCTATCTGGACAAGGGAAA | 42 (pKan Rv) | CGAAACGATCCTCATCCTGT | |
| 43 (p0 Fw1) | AATTCGACCCAGCTTTCTTG | 44 (p0 Rv1) | CCTTTGAGTGAGCTGATACCG | |
| 45 (p0 Fw2) | CGGTATCAGCTCACTCAAAGG | 46 (p0 Rv2) | CGCCCTTCTCCTTACTTACTTAG | |
RT-PCR, reverse transcription-PCR.
Detection of transcripts by quantitative real-time PCR.
Cells of P. phytofirmans were grown on 2.5 mM fructose, Bz, or IAA as the sole carbon and energy source. Then, total RNA was obtained from 4 ml of mid-log-phase cells, using RNAprotect bacterial reagent and the RNeasy minikit (Qiagen, Chatsworth, CA, USA). The RNA was quantified using an Eon microplate spectrophotometer (BioTek, Winooski, VT, USA) and treated with the Turbo DNase kit (Ambion, Austin, TX, USA) to remove DNA contamination. The reverse transcription-PCR was performed using the ImProm-II reverse transcription system (Promega Corporation, Madison, WI, USA) with 1 μg of RNA in 20-μl reaction mixtures. Real-time PCR was performed using the Power SYBR green PCR master mix (Applied Biosystems, Warrington, UK) and Eco real-time PCR detection system (Illumina, San Diego, CA, USA). The PCR mixture (15 μl) contained 3.0 μl of template cDNA (diluted 1:10) and 0.2 μM (each) primer. Amplification was performed under the following conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 30 s, 60°C for 30 s, and 72°C for 40 s, and finishing with a melting cycle from 55 to 95°C. Relative gene expression values were calculated using the comparative cycle threshold (CT) method (also known as the 2−ΔΔCT method) (65). 16S rRNA gene sequence (Bphyt_R0071) was used as a reference gene (internal control) in these assays. Gene expression levels were normalized to the average value of the gene expression levels determined in the fructose treatment. Genes analyzed by quantitative real-time PCR were iacA (Bphyt_2161), iacC (Bphyt_2156), iacF (Bphyt_2150), iacG (Bphyt_2167), iacA2 (Bphyt_6911), catA (Bphyt_2152), catA2 (Bphyt_1590), iacR1 (Bphyt_2166), iacR2 (Bphyt_2155), iacS (Bphyt_2160), the 30S ribosomal protein S7 (rpS7) gene (Bphyt_3648), and kynA (Bphyt_3229). The latter two gene sequences were also used as gene expression controls. Experiments were done in three biological replicates.
Construction of plasmid derivatives expressing iac genes.
To obtain pBS1-iacABIHECDGF plasmid (pBS1-iac1), which contains the complete iac gene cluster under the control of an arabinose-inducible promoter, the Gibson et al. assembly method was used (66). In brief, PCR products comprising iacABIHE (primer pair 1-2), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes and pBS1 plasmid (primer pair 9-10) (67) were obtained using primer pairs listed in Table 2. These primers contain a 20-bp terminal sequence homologous to the terminus of the fragment to be linked, and the sequences were combined and ligated to generate a new DNA molecule in a one-step isothermal reaction (66). Also, PCR products lacking one or two of these iac genes were amplified using primer pairs listed in Table 2: iacBIHE (primer pair 2-11), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to obtain the iac1ΔA derivative; iacA (primer pair 1-12), iacIHE (primer pair 2-13), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to produce the iac1ΔB derivative; iacABIHE (primer pair 1-15), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to produce the iac1ΔCD derivative; iacABIH (primer pair 1-14), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to give the iac1ΔE derivative; iacABIHE (primer pair 1-2), iacCD (primer pair 3-4), and iacG (primer pair 5-16) genes were used to create the iac1ΔF derivative; iacABIHE (primer pair 1-2), iacCD (primer pair 3-4), and iacF (primer pair 8-17) genes were used to generate the iac1ΔG derivative; iacABI (primer pair 1-18), iacE (primer pair 2-19), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to make the iac1ΔH derivative; and iacAB (primer pair 1-20), iacHE (primer pair 2-21), iacCD (primer pair 3-4), iacG (primer pair 5-6), and iacF (primer pair 7-8) genes were used to yield the iac1ΔI derivative. In addition, expression plasmids containing single or combinations of some iac genes (Table 1) were generated, using primer pairs listed in Table 2: for iacA, primer pair 1-22; iacAG, primer pairs 1-23 and 5-16; iacA2, primer pair 24-25; iacA2G2, primer pair 24-26; iacE, primer pair 27-28; iacB, primer pair 11-29; iacEB, primer pairs 11-20 and 28-30; iacC, primer pair 31-32; iacCD, primer pair 31-33; iacCDF, primer pairs 4-31 and 8-17; iacCDFI, primer pairs 4-31, 17-34, and 35-36; iacH, primer pair 37-38; and iacYT1, primer pair 39-40. With the exception of iacYT1 genes, all fragments containing the 20-bp terminal sequence that overlapped each other were ligated to pBS1 plasmid (primer pair 9-10), using the Gibson et al. assembly method (66), to get plasmids that contain iac genes under the control of the AraBAD (PBAD) promoter in pBS1 (67). Additionally, the iacYT1 genes were ligated to pKGWP0 plasmid (68) lacking the lacZ gene (primer pairs 43-44 and 45-46), to which the constitutive promoter of kanamycin phosphotransferase of plasmid pCR2.1-TOPO (primer pair 41-42) was added to obtain constitutive expression of the iacYT1 genes, using the Gibson et al. assembly method (66). Plasmid pBS1 or pKGWP0 derivatives were electroporated in C. pinatubonensis JMP134 and selected in LB medium plus Gm (30 μg/ml) or Sp (100 μg/ml), respectively. For expression of iac genes driven by the heterologous PBAD promoter, these derivatives were exposed to 5 mM l-arabinose, which is not a carbon source for C. pinatubonensis JMP134. All plasmid constructions obtained by the Gibson et al. assembly method were confirmed by sequencing.
Construction of lacZ reporter fusions.
Putative promoter regions were fused to the lacZ gene of pKGWP0 (68). PCR products comprising approximately 200 bp of the upstream region contiguous to translational starts of the iacA, iacC, iacF, iacG, or catB genes were obtained, using primer pairs listed in Table 2. The amplified DNA fragments were cloned into the XhoI-KpnI restriction site of pKGWP0, forming pIacA-lacZ, pIacC-lacZ, pIacF-lacZ, pIacG-lacZ, and pCatB-lacZ plasmids; transferred into P. phytofirmans and its derivatives; and selected in minimal medium supplemented with 100 μg ml−1 Sp. To evaluate promoter induction profiles, bacteria carrying promoter constructions were grown overnight on 10 mM fructose, refreshed in the same medium to an OD600 of 0.2, and then induced during 6 h with 0.25 mM IAA and/or compounds X and Y or DOAA. Compounds X and Y and DOAA were prepared using 4-fold-diluted supernatants of strain JMP134-iacAG or JMP134iac1 resting cells previously exposed for 2 h to 1 mM IAA as the only carbon source, respectively. β-Galactosidase assays were performed according to standard protocols (69).
Analytical methods.
The presence of IAA and its intermediates was determined by high-performance liquid chromatography–UV detection (HPLC-UV) using cell-free supernatants from cells exposed to IAA. Samples (20 μl) were injected into a Hitachi LaChrom Elite high-performance liquid chromatograph (Tokyo, Japan) equipped with a Kromasil 100-3.5 C18 4.6-μm-diameter column. Methanol-H2O mixtures containing 0.1% (vol/vol) phosphoric acid were used as the solvent, at a flow rate of 1 ml min−1. The column effluent was monitored at 210 nm. Retention times with methanol-H2O (40:60) for IAA, catechol, and compound 1 (DOAA) were 6.75, 3.05, and 2.04 min, respectively, whereas retention times with methanol-H2O (80:20) for IAA and compound 1 were 3.18 and 1.03 min, respectively.
In order to identify compound 1 (DOAA), this intermediate was produced and accumulated in strain JMP134iac1 exposed to 2.5 mM IAA as the sole carbon source for 2 h. Then, cells were centrifuged at 9,000 rpm, supernatant containing compound 1 was filtered (filter unit of 0.22 μm), and its pH was lowered to 2.5 using hydrochloric acid. Then, compound 1 was extracted from the aqueous medium with 1 volume of ethyl acetate (three times). The pooled organic layer was dried over anhydrous Na2SO4 and filtered, and the solvent vacuum were removed. The crude product was subjected to spectroscopic analysis. NMR spectra were recorded on a Bruker Advance III HD 400 instrument at 400 MHz for 1H and 100 MHz for 13C, using the solvent signal of dimethyl sulfoxide (DMSO)-D6 as reference. The chemical shifts are expressed in parts per million (ppm) (δ scale) downfield from tetramethylsilane, and coupling constant values (J) are given in hertz. The IR spectrum was obtained on a Bruker Vector 22 spectrophotometer, using KBr discs. The mass spectra were determined in a TQ 4500 triple-quadrupole mass spectrometer coupled with electrospray ionization operated in the negative ion mode.
Bioinformatic tools.
The iac gene sequences were retrieved from the Integrated Microbial Genomes database (https://img.jgi.doe.gov/cgi-bin/m/main.cgi). Protein similarity and conserved domain searches were performed with the BLASTP program and the Conserved Domains Database from the NCBI website using default parameters (70, 71). Only proteins of P. phytofirmans PsJN displaying at least 30% amino acid identity with iac genes of P. putida 1290 were considered for analysis.
Chemicals.
IAA, indole, and catechol were obtained from Sigma-Aldrich (Steinheim, Germany). Bz, fructose, and l-arabinose were purchased from Merck (Darmstadt, Germany).
Supplementary Material
ACKNOWLEDGMENTS
The work was funded by FONDECYT grants 1110850 and 1151130, CONICYT grant FB 0002-2014, Millennium Nuclei in Plant Functional Genomics grant P10-062F, and Plant Systems and Synthetic Biology grant NC130030.
Footnotes
Supplemental material for this article may be found at https://doi.org/10.1128/AEM.01991-16.
REFERENCES
- 1.Schaller GE, Bishopp A, Kieber JJ. 2015. The yin-yang of hormones: cytokinin and auxin interactions in plant development. Plant Cell 27:44–63. doi: 10.1105/tpc.114.133595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vanneste S, Friml J. 2009. Auxin: a trigger for change in plant development. Cell 136:1005–1016. doi: 10.1016/j.cell.2009.03.001. [DOI] [PubMed] [Google Scholar]
- 3.Tromas A, Perrot-Rechenmann C. 2010. Recent progress in auxin biology. Crit Rev Biol 333:297–306. doi: 10.1016/j.crvi.2010.01.005. [DOI] [PubMed] [Google Scholar]
- 4.Duca D, Lorv J, Patten CL, Rose D, Glick BR. 2014. Indole-3-acetic acid in plant-microbe interactions. Antonie Van Leeuwenhoek 106:85–125. doi: 10.1007/s10482-013-0095-y. [DOI] [PubMed] [Google Scholar]
- 5.Patten CL, Blakney AJ, Coulson TJ. 2013. Activity, distribution and function of indole-3-acetic acid biosynthetic pathways in bacteria. Crit Rev Microbiol 39:395–415. doi: 10.3109/1040841X.2012.716819. [DOI] [PubMed] [Google Scholar]
- 6.Spaepen S, Vanderleyden J. 2011. Auxin and plant-microbe interactions. Cold Spring Harb Perspect Biol 3:a001438. doi: 10.1101/cshperspect.a001438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Arora PK, Sharma A, Bae H. 2015. Microbial degradation of indole and its derivatives. J Chem 2015:129159. doi: 10.1155/2015/129159. [DOI] [Google Scholar]
- 8.Ebenau-Jehle C, Thomas M, Scharf G, Kockelkorn D, Knapp B, Schühle K, Heider J, Fuchs G. 2012. Anaerobic metabolism of indoleacetate. J Bacteriol 194:2894–2903. doi: 10.1128/JB.00250-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Egebo LA, Nielsen SV, Jochimsen BU. 1991. Oxygen-dependent catabolism of indole-3-acetic acid in Bradyrhizobium japonicum. J Bacteriol 173:4897–4901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jensen JB, Egsgaard H, Van Onckelen H, Jochimsen BU. 1995. Catabolism of indole-3-acetic acid and 4- and 5-chloroindole-3-acetic acid in Bradyrhizobium japonicum. J Bacteriol 177:5762–5766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Leveau JH, Lindow SE. 2005. Utilization of the plant hormone indole-3-acetic acid for growth by Pseudomonas putida strain 1290. Appl Environ Microbiol 71:2365–2371. doi: 10.1128/AEM.71.5.2365-2371.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Leveau JH, Gerards S. 2008. Discovery of a bacterial gene cluster for catabolism of the plant hormone indole 3-acetic acid. FEMS Microbiol Ecol 65:238–250. doi: 10.1111/j.1574-6941.2008.00436.x. [DOI] [PubMed] [Google Scholar]
- 13.Lin GH, Chen HP, Huang JH, Liu TT, Lin TK, Wang SJ, Tseng CH, Shu HY. 2012. Identification and characterization of an indigo-producing oxygenase involved in indole 3-acetic acid utilization by Acinetobacter baumannii. Antonie Van Leeuwenhoek 101:881–890. doi: 10.1007/s10482-012-9704-4. [DOI] [PubMed] [Google Scholar]
- 14.Scott JC, Greenhut IV, Leveau JH. 2013. Functional characterization of the bacterial iac genes for degradation of the plant hormone indole-3-acetic acid. J Chem Ecol 39:942–951. doi: 10.1007/s10886-013-0324-x. [DOI] [PubMed] [Google Scholar]
- 15.Shu HY, Lin LC, Lin TK, Chen HP, Yang HH, Peng KC, Lin GH. 2015. Transcriptional regulation of the iac locus from Acinetobacter baumannii by the phytohormone indole-3-acetic acid. Antonie Van Leeuwenhoek 107:1237–1247. doi: 10.1007/s10482-015-0417-3. [DOI] [PubMed] [Google Scholar]
- 16.Sawana A, Adeolu M, Gupta RS. 2014. Molecular signatures and phylogenomic analysis of the genus Burkholderia: proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front Genet 5:429. doi: 10.3389/fgene.2014.00429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mitter B, Petric A, Shin MW, Chain PS, Hauberg-Lotte L, Reinhold-Hurek B, Nowak J, Sessitsch A. 2013. Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants. Front Plant Sci 4:120. doi: 10.3389/fpls.2013.00120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Poupin MJ, Timmermann T, Vega A, Zúñiga A, González B. 2013. Effects of the plant growth-promoting bacterium Burkholderia phytofirmans PsJN throughout the life cycle of Arabidopsis thaliana. PLoS One 8:e69435. doi: 10.1371/journal.pone.0069435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sun Y, Cheng Z, Glick BR. 2009. The presence of a 1-aminocyclopropane-1-carboxylate (ACC) deaminase deletion mutation alters the physiology of the endophytic plant growth-promoting bacterium Burkholderia phytofirmans PsJN. FEMS Microbiol Lett 296:131–136. doi: 10.1111/j.1574-6968.2009.01625.x. [DOI] [PubMed] [Google Scholar]
- 20.Zúñiga A, Poupin MJ, Donoso R, Ledger T, Guiliani N, Gutiérrez RA, González B. 2013. Quorum sensing and indole-3-acetic acid degradation play a role in colonization and plant growth promotion of Arabidopsis thaliana by Burkholderia phytofirmans PsJN. Mol Plant Microbe Interact 26:546–553. doi: 10.1094/MPMI-10-12-0241-R. [DOI] [PubMed] [Google Scholar]
- 21.Weilharter A, Mitter B, Shin MV, Chain PS, Nowak J, Sessitsch A. 2011. Complete genome sequence of the plant growth-promoting endophyte Burkholderia phytofirmans strain PsJN. J Bacteriol 193:3383–3384. doi: 10.1128/JB.05055-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Naveed M, Qureshi MA, Zahir ZA, Hussain MB, Sessitsch A, Mitter B. 2015. l-Tryptophan-dependent biosynthesis of indole-3-acetic acid (IAA) improves plant growth and colonization of maize by Burkholderia phytofirmans PsJN. Ann Microbiol 65:1381–1389. doi: 10.1007/s13213-014-0976-y. [DOI] [Google Scholar]
- 23.Baty F, Flandrois JP, Delignette-Muller ML. 2002. Modeling the lag time of Listeria monocytogenes from viable count enumeration and optical density data. Appl Environ Microbiol 68:5816–5825. doi: 10.1128/AEM.68.12.5816-5825.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schweigert N, Zehnder AJ, Eggen RI. 2001. Chemical properties of catechols and their molecular modes of toxic action in cells, from microorganisms to mammals. Environ Microbiol 3:81–91. doi: 10.1046/j.1462-2920.2001.00176.x. [DOI] [PubMed] [Google Scholar]
- 25.Ledger T, Pieper DH, González B. 2006. Chlorophenol hydroxylases encoded by plasmid pJP4 differentially contribute to chlorophenoxyacetic acid degradation. Appl Environ Microbiol 72:2783–2792. doi: 10.1128/AEM.72.4.2783-2792.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pérez-Pantoja D, Ledger T, Pieper DH, González B. 2003. Efficient turnover of chlorocatechols is essential for growth of Ralstonia eutropha JMP134(pJP4) in 3-chlorobenzoic acid. J Bacteriol 185:1534–1542. doi: 10.1128/JB.185.5.1534-1542.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Harwood CS, Parales RE. 1996. The beta-ketoadipate pathway and the biology of self-identity. Annu Rev Microbiol 50:553–590. doi: 10.1146/annurev.micro.50.1.553. [DOI] [PubMed] [Google Scholar]
- 28.Lykidis A, Pérez-Pantoja D, Ledger T, Mavromatis K, Anderson IJ, Ivanova NN, Hooper SD, Lapidus A, Lucas S, González B, Kyrpides NC. 2010. The complete multipartite genome sequence of Cupriavidus necator JMP134, a versatile pollutant degrader. PLoS One 5:e9729. doi: 10.1371/journal.pone.0009729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pérez-Pantoja D, De la Iglesia R, Pieper DH, González B. 2008. Metabolic reconstruction of aromatic compounds degradation from the genome of the amazing pollutant-degrading bacterium Cupriavidus necator JMP134. FEMS Microbiol Rev 32:736–794. doi: 10.1111/j.1574-6976.2008.00122.x. [DOI] [PubMed] [Google Scholar]
- 30.Pérez-Pantoja D, Donoso R, Agulló L, Córdova M, Seeger M, Pieper DH, González B. 2012. Genomic analysis of the potential for aromatic compounds biodegradation in Burkholderiales. Environ Microbiol 14:1091–1117. doi: 10.1111/j.1462-2920.2011.02613.x. [DOI] [PubMed] [Google Scholar]
- 31.Reddy VS, Shlykov MA, Castillo R, Sun EI, Saier MH Jr. 2012. The major facilitator superfamily (MFS) revisited. FEBS J 279:2022–2035. doi: 10.1111/j.1742-4658.2012.08588.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Whipp MJ, Camakaris H, Pittard AJ. 1998. Cloning and analysis of the shiA gene, which encodes the shikimate transport system of Escherichia coli K-12. Gene 209:185–192. doi: 10.1016/S0378-1119(98)00043-2. [DOI] [PubMed] [Google Scholar]
- 33.Huddleston JP, Burks EA, Whitman CP. 2014. Identification and characterization of new family members in the tautomerase superfamily: analysis and implications. Arch Biochem Biophys 564:189–196. doi: 10.1016/j.abb.2014.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Whitman CP, Aird BA, Gillespie WR, Stolowich NJ. 1991. Chemical and enzymatic ketonization of 2-hydroxymuconate, a conjugated enol. J Am Chem Soc 113:3154–3162. doi: 10.1021/ja00008a052. [DOI] [Google Scholar]
- 35.Ghisla S, Thorpe C. 2004. Acyl-CoA dehydrogenases. A mechanistic overview. Eur J Biochem 271:494–508. [DOI] [PubMed] [Google Scholar]
- 36.Galán B, Díaz E, Prieto MA, García JL. 2000. Functional analysis of the small component of the 4-hydroxyphenylacetate 3-monooxygenase of Escherichia coli W: a prototype of a new flavin:NAD(P)H reductase subfamily. J Bacteriol 182:627–636. doi: 10.1128/JB.182.3.627-636.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kavanagh KL, Jörnvall H, Persson B, Oppermann U. 2008. Medium- and short-chain dehydrogenase/reductase gene and protein families: the SDR superfamily: functional and structural diversity within a family of metabolic and regulatory enzymes. Cell Mol Life Sci 65:3895–3906. doi: 10.1007/s00018-008-8588-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ferraro DJ, Gakhar L, Ramaswamy S. 2005. Rieske business: structure-function of Rieske non-heme oxygenases. Biochem Biophys Res Commun 338:175–190. doi: 10.1016/j.bbrc.2005.08.222. [DOI] [PubMed] [Google Scholar]
- 39.Kweon O, Kim SJ, Baek S, Chae JC, Adjei MD, Baek DH, Kim YC, Cerniglia CE. 2008. A new classification system for bacterial Rieske non-heme iron aromatic ring-hydroxylating oxygenases. BMC Biochem 9:11. doi: 10.1186/1471-2091-9-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bullock TL, Clarkson WD, Kent HM, Stewart M. 1996. The 1.6 angstroms resolution crystal structure of nuclear transport factor 2 (NTF2). J Mol Biol 260:422–431. doi: 10.1006/jmbi.1996.0411. [DOI] [PubMed] [Google Scholar]
- 41.Lundqvist T, Rice J, Hodge CN, Basarab GS, Pierce J, Lindqvist Y. 1994. Crystal structure of scytalone dehydratase—a disease determinant of the rice pathogen, Magnaporthe grisea. Structure 2:937–944. doi: 10.1016/S0969-2126(94)00095-6. [DOI] [PubMed] [Google Scholar]
- 42.Hurtubise Y, Barriault D, Sylvestre M. 1998. Involvement of the terminal oxygenase beta subunit in the biphenyl dioxygenase reactivity pattern toward chlorobiphenyls. J Bacteriol 180:5828–5835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Aliverti A, Pandini V, Pennati A, de Rosa M, Zanetti G. 2008. Structural and functional diversity of ferredoxin-NADP(+) reductases. Arch Biochem Biophys 474:283–291. doi: 10.1016/j.abb.2008.02.014. [DOI] [PubMed] [Google Scholar]
- 44.Kallio P, Sultana A, Niemi J, Mäntsälä P, Schneider G. 2006. Crystal structure of the polyketide cyclase AknH with bound substrate and product analogue: implications for catalytic mechanism and product stereoselectivity. J Mol Biol 357:210–220. doi: 10.1016/j.jmb.2005.12.064. [DOI] [PubMed] [Google Scholar]
- 45.Staswick PE, Serban B, Rowe M, Tiryaki I, Maldonado MT, Maldonado MC, Suza W. 2005. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 17:616–627. doi: 10.1105/tpc.104.026690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Coschigano PW, Young LY. 1997. Identification and sequence analysis of two regulatory genes involved in anaerobic toluene metabolism by strain T1. Appl Environ Microbiol 63:652–660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lau PC, Wang Y, Patel A, Labbé D, Bergeron H, Brousseau R, Konishi Y, Rawlings M. 1997. A bacterial basic region leucine zipper histidine kinase regulating toluene degradation. Proc Natl Acad Sci U S A 94:1453–1458. doi: 10.1073/pnas.94.4.1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Achong GR, Rodriguez AM, Spormann AM. 2001. Benzylsuccinate synthase of Azoarcus sp. strain T: cloning, sequencing, transcriptional organization, and its role in anaerobic toluene and m-xylene mineralization. J Bacteriol 183:6763–6770. doi: 10.1128/JB.183.23.6763-6770.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Labbé D, Garnon J, Lau PC. 1997. Characterization of the genes encoding a receptor-like histidine kinase and a cognate response regulator from a biphenyl/polychlorobiphenyl-degrading bacterium, Rhodococcus sp. strain M5. J Bacteriol 179:2772–2776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.O'Leary ND, Mooney A, O'Mahony M, Dobson AD. 2014. Functional characterization of a StyS sensor kinase reveals distinct domains associated with intracellular and extracellular sensing of styrene in Pseudomonas putida CA-3. Bioengineering 5:114–122. doi: 10.4161/bioe.28354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Maddocks SE, Oyston PC. 2008. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 154:3609–3623. doi: 10.1099/mic.0.2008/022772-0. [DOI] [PubMed] [Google Scholar]
- 52.Cao H, Krishnan G, Goumnerov B, Tsongalis J, Tompkins R, Rahme LG. 2001. A quorum sensing-associated virulence gene of Pseudomonas aeruginosa encodes a LysR-like transcription regulator with a unique self-regulatory mechanism. Proc Natl Acad Sci U S A 98:14613–14618. doi: 10.1073/pnas.251465298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kim J, Kim JG, Kang Y, Jang JY, Jog GJ, Lim JY, Kim S, Suga H, Nagamatsu T, Hwang I. 2004. Quorum sensing and the LysR-type transcriptional activator ToxR regulate toxoflavin biosynthesis and transport in Burkholderia glumae. Mol Microbiol 54:921–934. doi: 10.1111/j.1365-2958.2004.04338.x. [DOI] [PubMed] [Google Scholar]
- 54.Lu Z, Takeuchi M, Sato T. 2007. The LysR-type transcriptional regulator YofA controls cell division through the regulation of expression of ftsW in Bacillus subtilis. J Bacteriol 189:5642–5651. doi: 10.1128/JB.00467-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Craven SH, Ezezika OC, Haddad S, Hall RA, Momany C, Neidle EL. 2009. Inducer responses of BenM, a LysR-type transcriptional regulator from Acinetobacter baylyi ADP1. Mol Microbiol 72:881–894. doi: 10.1111/j.1365-2958.2009.06686.x. [DOI] [PubMed] [Google Scholar]
- 56.Ezezika OC, Collier-Hyams LS, Dale HA, Burk AC, Neidle EL. 2006. CatM regulation of the benABCDE operon: functional divergence of two LysR-type paralogs in Acinetobacter baylyi ADP1. Appl Environ Microbiol 72:1749–1758. doi: 10.1128/AEM.72.3.1749-1758.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Nishijyo T, Haas D, Itoh Y. 2001. The CbrA-CbrB two component regulatory system controls the utilization of multiple carbon and nitrogen sources in Pseudomonas aeruginosa. Mol Microbiol 40:917–931. doi: 10.1046/j.1365-2958.2001.02435.x. [DOI] [PubMed] [Google Scholar]
- 58.Hąc-Wydro K, Flasiński M. 2015. The studies on the toxicity mechanism of environmentally hazardous natural (IAA) and synthetic (NAA) auxin. The experiments on model Arabidopsis thaliana and rat liver plasma membranes. Colloids Surf B Biointerfaces 130:53–60. doi: 10.1016/j.colsurfb.2015.03.064. [DOI] [PubMed] [Google Scholar]
- 59.Pieper DH, Engesser KH, Knackmuss HJ. 1989. Regulation of catabolic pathways of phenoxyacetic acids and phenols in Alcaligenes eutrophus JMP134. Arch Microbiol 151:365–371. doi: 10.1007/BF00406566. [DOI] [Google Scholar]
- 60.Pencík A, Simonovik B, Petersson SV, Henyková E, Simon S, Greenham K, Zhang Y, Kowalczyk M, Estelle M, Zazímalová E, Novák O, Sandberg G, Ljung K. 2013. Regulation of auxin homeostasis and gradients in Arabidopsis roots through the formation of the indole-3-acetic acid catabolite 2-oxindole-3-acetic acid. Plant Cell 25:3858–3570. doi: 10.1105/tpc.113.114421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chamarro J, Ostin A, Sandberg G. 2001. Metabolism of indole-3-acetic acid by orange (Citrus sinensis) flavedo tissue during fruit development. Phytochemistry 57:179–187. doi: 10.1016/S0031-9422(01)00023-1. [DOI] [PubMed] [Google Scholar]
- 62.Tsurumi S, Wada S. 1985. Identification of 3-(O-beta-glucosyl)-2-indolone-3-acetylaspartic acid as a new indole-3-acetic acid metabolite in Vicia seedlings. Plant Physiol 79:667–671. doi: 10.1104/pp.79.3.667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Dorn E, Hellwig M, Reineke W, Knackmuss HJ. 1974. Isolation and characterization of a 3-chlorobenzoate degrading pseudomonad. Arch Microbiol 99:61–70. doi: 10.1007/BF00696222. [DOI] [PubMed] [Google Scholar]
- 64.Scott TA, Mercer EI. 1996. Concise encyclopedia of biochemistry and molecular biology. Walter de Gruyter, Berlin, Germany. [Google Scholar]
- 65.Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative CT method. Nat Protoc 3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 66.Gibson DG, Young L, Chuang RY, Venter JC, Hutchison CA III, Smith HO. 2009. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods 6:343–345. doi: 10.1038/nmeth.1318. [DOI] [PubMed] [Google Scholar]
- 67.Bronstein PA, Marrichi M, Cartinhour S, Schneider DJ, DeLisa MP. 2005. Identification of a twin-arginine translocation system in Pseudomonas syringae pv. tomato DC3000 and its contribution to pathogenicity and fitness. J Bacteriol 187:8450–8461. doi: 10.1128/JB.187.24.8450-8461.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Marín M, Pérez-Pantoja D, Donoso RA, Wray V, González B, Pieper DH. 2010. Modified 3-oxoadipate pathway for the biodegradation of methylaromatics in Pseudomonas reinekei MT1. J Bacteriol 192:1543–1552. doi: 10.1128/JB.01208-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Miller JH. 1972. Experiments in molecular genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. [Google Scholar]
- 70.Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Marchler-Bauer A, Derbyshire MK, Gonzales NR, Lu S, Chitsaz F, Geer LY, Geer RC, He J, Gwadz M, Hurwitz DI, Lanczycki CJ, Lu F, Marchler GH, Song JS, Thanki N, Wang Z, Yamashita RA, Zhang D, Zheng C, Bryant SH. 2015. CDD: NCBI's conserved domain database. Nucleic Acids Res 43:D222–D226. doi: 10.1093/nar/gku1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.