

Abstract
Cartilaginous fish are particularly vulnerable to anthropogenic stressors and environmental change because of their K-selected reproductive strategy. Accurate data from scientific surveys and landings are essential to assess conservation status and to develop robust protection and management plans. Currently available data are often incomplete or incorrect as a result of inaccurate species identifications, due to a high level of morphological stasis, especially among closely related taxa. Moreover, several diagnostic characters clearly visible in adult specimens are less evident in juveniles. Here we present results generated by the ELASMOMED Consortium, a regional network aiming to sample and DNA-barcode the Mediterranean Chondrichthyans with the ultimate goal to provide a comprehensive DNA barcode reference library. This library will support and improve the molecular taxonomy of this group and the effectiveness of management and conservation measures. We successfully barcoded 882 individuals belonging to 42 species (17 sharks, 24 batoids and one chimaera), including four endemic and several threatened ones. Morphological misidentifications were found across most orders, further confirming the need for a comprehensive DNA barcoding library as a valuable tool for the reliable identification of specimens in support of taxonomist who are reviewing current identification keys. Despite low intraspecific variation among their barcode sequences and reduced samples size, five species showed preliminary evidence of phylogeographic structure. Overall, the ELASMOMED initiative further emphasizes the key role accurate DNA barcoding libraries play in establishing reliable diagnostic species specific features in otherwise taxonomically problematic groups for biodiversity management and conservation actions.
Introduction
To assess and conserve biodiversity, it is critical to correctly identify species that occur in a given ecosystem in order to evaluate species richness and abundance. In recent years, methods of identification based on morphology were gradually integrated with methods based on DNA sequences, such as DNA barcoding thereby forming the so called molecular taxonomy. DNA barcoding seeks to advance both specimens identification and species discovery through the analysis of patterns of sequences divergence of a universal, standardized gene region. Many studies have shown the effectiveness of the mitochondrial cytochrome c oxidase subunit I (COI) gene as a universal barcode sequence for species identification in animal lineages [1,2]. Molecular taxonomy overcomes some problems posed by traditional morphological identification such as homoplasy [3] and phenotypic plasticity [4] of characters used for species identification. Secondly, because often the adopted morphological keys are effective only for a particular life stage, many individuals, especially in their juvenile phases, cannot be assigned to species [5]. Finally, traditional taxonomy doesn’t allow the identification of cryptic species [6]. DNA barcoding has been fundamental in case studies related to immature specimens’ identification (e.g. fish larvae, [5,7]; amphibians and reptiles, [8,9]). In many cases it has been successfully employed to resolve species boundaries between morphologically conserved taxa (e.g. tribe Bombini, [10]).
In the marine realm, the conservation of similar morphological traits appears quite common among sibling species [11–14] and it is often combined with the lack of visual communication in many taxa, in favour of chemical [15] or electrical signals [16–18], both intrinsic conditions to the definition of cryptic species. Among marine organisms, Chondrichthyans seem to have experienced frequent cryptic speciation events across different taxa: large lantern sharks [19], skates [20–22], blacktip sharks [23], hammerhead sharks [24] and guitarfish [25] might have undergone isolating mechanisms which precluded mating between co-occurring species. Despite controversies and criticisms [26,27], cryptic species discovery and, in general, species cataloguing are fundamental, as species represent the basic unit for the management, conservation, and legal protection of biodiversity and for the distribution of limited resources [28–33].
To maximize the potential of molecular taxonomy is necessary to have solid and comprehensive DNA barcodes reference libraries. The Fish Barcode of Life campaign (Fish-BOL, http://www.fishbol.org) is an initiative started in 2005 with the goal to barcode all fish species [34–36]. As of August 2016, 665 out of 1228 chondrichthyan species have been barcoded. Fish-BOL’s efforts comprise of numerous projects covering low-level taxonomic groups and several projects that have targeted specific regional chondrichthyan faunas [37–43]. In addition, recent studies demonstrated the effectiveness of DNA barcodes in describing phylogeographic patterns in this class [22,44,45].
One of those targeted Fish-BOL projects is the ELASMOMED Consortium, a regional network active since 2009 involving fish biologists, fishery scientists, and molecular zoologists from 15 research institutions, most of which also participate in the Mediterranean International Trawl Survey (MEDITS) scientific program[46]. The aims of this network are the sampling and barcoding Chondrichthyans of the Mediterranean Sea.
The Mediterranean Sea is appraised as a global marine biodiversity hotspot and it is home to 89 chondrichthyan species (sharks, skates, rays and chimaeras), which corresponds to about 7% of the global species diversity of the group [47–51]. In detail, the Mediterranean Chondrichthyans include one chimaera, 49 sharks belonging to 17 families and 27 genera, and 39 batoids consisting of nine families with 16 genera. Five batoids are considered endemic: Leucoraja melitensis, Raja asterias, Raja radula, Raja polystigma, and Mobula mobular, although the endemic status of the latter is uncertain as its separation from the widespread congener Mobula japonica has been recently questioned [52]. In parallel, a total of 86 Mediterranean Chondrichthyan species (S1 Table) was reviewed for the European Red List of Marine Fishes [53]. This list includes species threatened by extinction in European waters. Assessed taxa were categorized as Critically Endangered (15), Endangered (13), Vulnerable (11), Near Threatened (15), Least Concern (12) and Data Deficient (20).
Several exotic elasmobranchs have recently been recorded in the Mediterranean (S1 Table), however their particular status (vagrant, alien, or established) is still under debate [48,54–57]. The presence of six species (Carcharhinus melanopterus, Dipturus batis complex, Torpedo alexandrinsis, Leucoraja fullonica, Raja africana), however, has not been confirmed [50,58–61]. In contrast, the presence of Dipturus nidarosiensis in the Mediterranean, a species formerly reported only from the North-Eastern Atlantic, has been ascertained by recent studies [58,62].
Chondrichthyans’ reproductive strategy makes them particularly vulnerable to anthropogenic stressors, such as the use of different fishing gears and the direct and indirect effects of environmental changes and habitat fragmentation [63]. For example, neritic species such as Squatina spp. and Scyliorhinus stellaris are highly depleted as a consequence of the use of unselective fishing gear [64]. Available fisheries data for Chondrichthyes are incomplete and incorrect, because they are often recorded at higher taxonomic levels than species, with frequent misidentifications of individuals [51,65,66]. In 2012, the General Fisheries Commission for the Mediterranean (GFCM) issued Recommendation GFCM/36/2012/3 on fisheries management measures for conservation of sharks and rays in the GFCM area stating that “cartilaginous fish are kept on board, trans-shipped, landed and marketed at first sale in a way that species are recognizable and identifiable and catches, incidental takings and, whenever appropriate, releases by species can be monitored and recorded” [67]. However, morphological identification of cartilaginous fish remains difficult because of low levels of differentiation among species across multiple taxa and several diagnostic taxonomic characters are clearly exhibited by adult specimens but are less pronounced in juveniles [22,51,65,66]. This can lead to erroneous species attribution even among skate species that are not closely related, e.g. Leucoraja fullonica and L. circularis. Such taxonomic uncertainties often occur within a larger group constituted of Raja polystigma, R. montagui, R. asterias and R. brachyura [22,66]. For sharks, similar difficulties were reported for the congeneric species Squalus blainville and Squalus megalops [68,69] and for Mustelus mustelus and Mustelus punctulatus [70]. Finally, cryptic species (sensu Bickford et al. [6]) have frequently been reported in elasmobranchs as shown by molecular studies of the common skate Dipturus batis, once one of the most abundant skate resources in the North-Eastern Atlantic trawl fishery and today heavily depleted in most of its range [63]. D. batis actually comprises two cryptic species, Dipturus cf intermedia and Dipturus cf flossada [20,71,72]. A genetic analysis of Eastern Atlantic and Mediterranean species of the genus Raja revealed several recently diverged peripatric sibling species, such as Raja clavata and R. straeleni [73], R. polystigma and R. montagui [22,74], as well as the R. miraletus complex [75,76].
Here we report the establishment of DNA reference barcodes for 42 chondrichthyan species, mostly collected as part of the MEDITS program, as an integrative tool to improve the effectiveness of the above mentioned measures for conservation. This new library will: i) offer a valuable tool for reliable identification of specimens and clarify the taxonomic status of important cartilaginous fishes; ii) support taxonomists who are reviewing the current identification keys, and iii) provide a robust scientific baseline for management and conservation actions, especially in relation to endemic and endangered species. Finally, the comparison and integration of the ELASMOMED dataset with other public barcode datasets will also allow the preliminary identification of geographical population structure and help to determine candidate conservation units.
In general, the assembly of a comprehensive DNA barcode reference library for biological species would mean to compile a biodiversity inventory on different scales and dimensions, making molecular systematics a fundamental tool for the implementation of biodiversity monitoring programmes worldwide. Several of such programmes already exploit the potential of accurate DNA reference libraries by integrating environmental DNA analyses in their standard monitoring programmes [77–79].
Materials and Methods
Sampling
Specimens used in this study were collected from Mediterranean individuals caught during scientific research programs. No specific approval of this vertebrate work is required since the individuals sampled in this study were obtained from scientific and commercial fishing activities. A total of 998 individuals were collected from several locations within six Mediterranean FAO fishing divisions (http://www.fao.org/fishery/area/Area37/en). Starting in 2009, dedicated sampling was performed mainly in the framework of the MEDITS scientific surveys (http://www.sibm.it/SITO%20MEDITS/principaleprogramme.htm) or by contracted commercial fishermen on designated cruises (S2 Table). Additional samples were provided by each partner of the ELASMOMED Consortium, from each Institute’s collections. Specimen and collection data, as well as voucher digital images (when recorded) were uploaded to the “ELASMOMED Consortium” project (Project Code: ELAMO) accessible through the Barcode of Life Data system (BOLD, http://www.barcodinglife.org, [80]). Individual fin clips or skeletal muscle tissue samples were collected and preserved in 96% ethanol and kept at -20°C until laboratory analyses.
DNA extraction, amplification and sequencing
Laboratory work was jointly carried out by the Centre for Biodiversity Genomics (CBG) and at the University of Bologna (UNIBO). At CBG 650bp of the mitochondrial COI region were obtained by following standardized high-throughput protocols for DNA barcode amplification and sequencing [81]. At UNIBO the same COI fragment was amplified using the primer set FishF2 and FishR2 following the protocol described in Ward et al. [82]. Amplification products were checked on a 1.5% agarose gel. A commercial sequence service provider (Macrogen Europe, Amsterdam, Netherlands) performed sequencing employing the same primers used for the amplification. Trace files and sequence data were uploaded to BOLD and subsequently submitted to GenBank (Accession numbers are provided in S3 Table).
Specimens’ identification and spatial scale of barcode variation
Specimens were identified on board or in the lab using morphological taxonomic characters according to guidelines provided in [83]. A p-distance metric with pairwise deletion was used for sequence comparisons [84]. Genetic distances and Neighbour-joining (NJ) tree clustering [85] were obtained using MEGA version 6 [86]. Confidence in estimated relationships of NJ tree topologies was evaluated by a bootstrap analysis with 1,000 replicates [87].
The mean and maximum intraspecific genetic distances and the mean distance to the Nearest Neighbour (NN) were computed using the ‘Barcoding Gap Analysis’ tool on BOLD [80]. Maximum intraspecific distance was plotted against the mean distance to the NN for each species to infer the presence of a “barcode gap”, which is defined as a distinct gap between intraspecific and interspecific variability [84].
The Barcode Index Number (BIN) System clusters sequences using a Refined Single Linkage algorithm to produce operational taxonomic units that closely correspond to species. BINs were automatically assigned by BOLD and assessed using the ‘BIN Discordance Report’ analysis tool [88]. This tool labels a BIN as “concordant” when it comprises sequences attributed to the same species, and “discordant” when it comprises sequences of different species.
An arbitrary measure of taxonomic reliability was attributed to each barcoded taxon according to the criteria proposed by Costa et al. [89]. Representative barcode sequences for each species were queried using the BOLD Identification Engine with the Species Level option. Grades ranging from A (full concordance) to E (full discordance) were attributed according to the following criteria:
Grade A- External concordance: unambiguous species match with specimens from other BOLD projects or published sequences. Monophyletic species with a maximum of 2% (patristic) sequence divergence.
Grade B- Internal concordance: species congruent within our dataset, where at least 3 specimens of the same species are available, with a maximum of 2% (patristic) sequence divergence. No matching sequences found through the BOLD-IDS.
Grade C- Sub-optimal concordance (possible within species genetic structure): at least 3 specimens of the same species are available within the library and form a monophyletic cluster; however intraspecific distance is greater than 2%; and/or the BOLD-IDS indicates monophyletic nearest neighbour of the same species, with more than 2% patristic distance.
Grade D- Insufficient Data: low number of specimens analysed (1 or 2 individuals) and no matching sequence available in BOLD.
Grade E- Discordant species assignments: sequences for a given species in our dataset did not match with the same species in BOLD. The specimen may match with a different species or may display paraphyly or polyphyly.
Because of the several Mediterranean geographical areas covered by ELASMOMED, we tested for the presence of phylogeographic signal at regional level for species with barcode data from multiple FAO divisions. Species-specific haplotype networks were created using Haploviewer (http://www.cibiv.at/~greg/haploviewer). Parsimony trees required for Haploviewer were reconstructed with the dnapars of the PHYLIP package version 3.6 [90,91].
Results
DNA barcodes could be recovered for 884 of the 998 individuals. Stop codons were recorded only for two specimens: ELAMO028-15 (S. blainville) and ELAME1143-11 (Torpedo marmorata), which were excluded from further analyses. The newly generated Mediterranean barcode library ELASMOMED includes 42 species: 17 sharks, 24 skates/rays and one chimera, belonging to eight orders and 18 families (S2 Table). Overall nucleotide frequencies were 25.16% adenine (A), 26.12% cytosine (C), 16.78% guanine (G) and 31.94% thymine (T), with an average GC content of 42.90%.
A first Neighbour-Joining tree of 882 barcode sequences showed that 77 specimens (8.73%), representing 11 species, clustered with individuals of a closely related and morphologically similar species (Table 1), probably because of identification errors during field sampling. All these specimens were reassessed and subsequently renamed.
Table 1. Cases of specimens’ misidentification by FAO fishing division.
The number of misidentified individuals over the total number of barcoded individuals is given in parenthesis.
| Barcode ID | Morphological ID | FAO Division |
|---|---|---|
| Dasyatis centroura | Pteroplatytrygon violacea | 37.1.3—Sardina (1/1) |
| Pteroplatytrygon violacea | Dasyatis centroura | 37.1.1—Balearic (1/1) |
| Leucoraja circularis | Leucoraja fullonica | 37.1.3—Sardinia (2/8); 37.2.2—Ionian (1/5) |
| Raja polystigma | Raja montagui | 37.1.1—Balearic (2/5) |
| Torpedo marmorata | Torpedo nobiliana | 37.3.2—Levant (3/9) |
| Torpedo marmorata | Torpedo torpedo | 37.2.1—Adriatic (1/1) |
| Scyliorhinus canicula | Scyliorhinus stellaris | 37.2.1—Adriatic (4/20) |
| Mustelus punctulatus | Mustelus mustelus | 37.1.1—Balearic (5/5); 37.2.1—Adriatic (45/146) |
| Centrophorus granulosus | Centrophorus uyato | 37.1.1—Balearic (1/5) |
| Squalus blainville | Squalus acanthias | 37.1.1—Balearic (2/3); 37.3.2—Levant (5/5) |
| Squalus blainville | Squalus megalops | 37.2.2—Ionian (4/12) |
All further analyses were conducted using this curated and corrected dataset and the resulting Neighbour-Joining tree (Figs 1 and 2) indicated that most of the species formed cohesive units, concordant with the morphological identification. Species-specific clusters were supported by high bootstrap values (≥80), with the exception of the D. pastinaca cluster, which split into two fully supported sub-clusters (1 and 2; Fig 2, S1 Fig). The families Scyliorhinidae and Myliobatidae did not form a monophyletic clade, as did the genera Raja and Dasyatis (Figs 1 and 2).
Fig 1. Neighbour-Joining tree based on genetic p-distances of COI barcode sequences of 17 shark species.
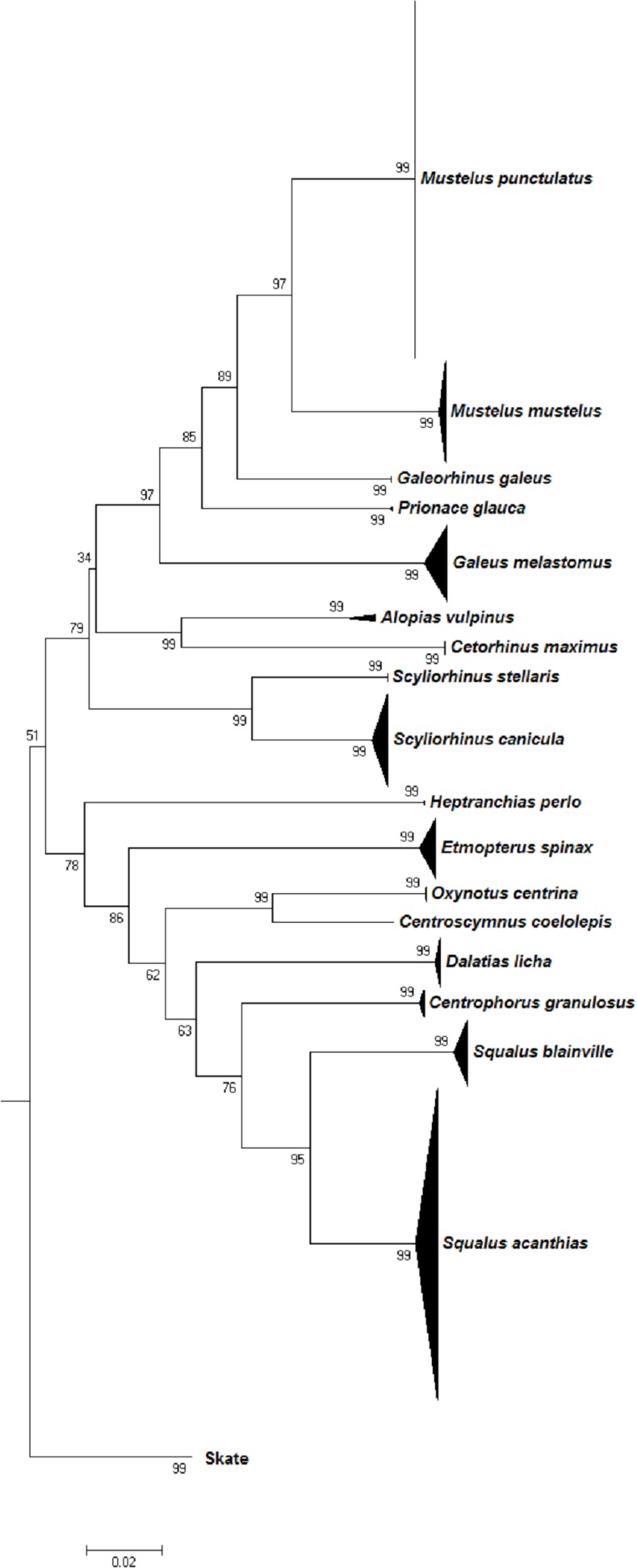
Numbers near nodes indicate bootstrap values (>50%). The distance scale bar is given. Each species' sample size and geographic origin are detailed in S2 Table.
Fig 2. Neighbour-Joining tree based on genetic p-distances of COI barcode sequences of 23 skate species.
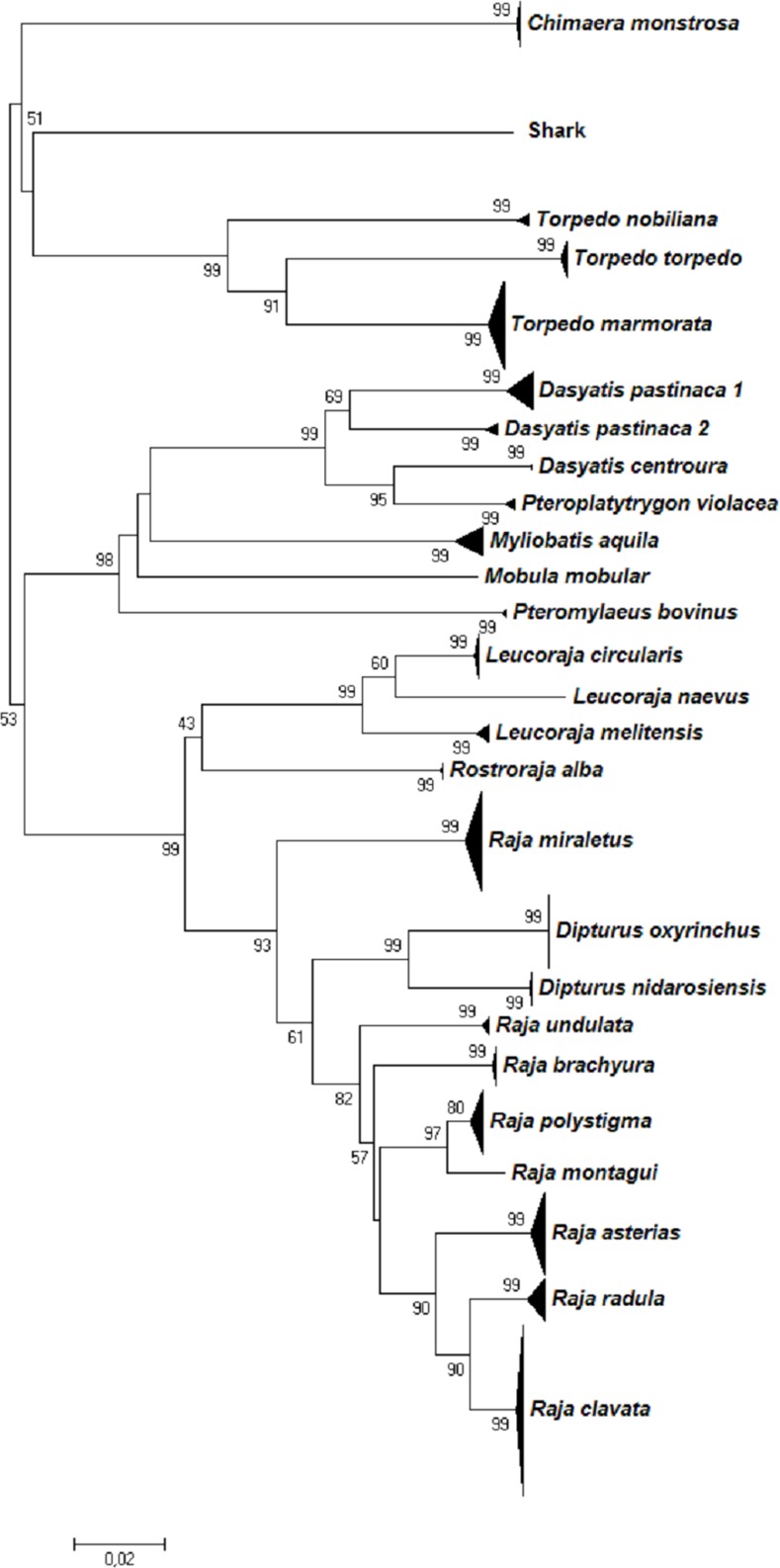
Numbers near nodes indicate bootstrap values (>50%). The distance scale bar is given. Each species' sample size and geographic origin are detailed in S2 Table.
The mean and maximum p-distance within species showed average values of 0.29% (range = 0–3.06%) and 0.93% (range = 0–8.72%), respectively (Table 2). The highest values for both distances occurred in Dasyatis pastinaca. Excluding this outlier taxon, highest mean and maximum p-distances were reduced to 0.43% (Raja asterias) and 1.55% (Galeus melastomus), respectively. The distances to the Nearest Neighbour (Table 2) varied from 1.71% (R. montagui vs. R. polystigma) to 12.83% (T. marmorata vs. Torpedo nobiliana) for congeneric taxa; while higher values were observed for comparisons at higher taxonomic levels (Table 2). Note that the maximum value of intraspecific p-distance was always lower than the distance to the Nearest Neighbour with the exception of D. pastinaca, highlighting the absence of a “barcode gap” for this species (Fig 3).
Table 2. Mean and maximum intraspecific distances and Nearest-Neighbour distance (p-distance values) of the 41 chondrichthyan species barcoded for the ELASMOMED library.
The Dasyatis pastinaca cluster is not separated.
| Species | Within-species distance | Nearest Neighbour | Distance to theNearest Neighbor | |
|---|---|---|---|---|
| Mean | Maximum | |||
| Chimaera monstrosa | 0.08 | 0.31 | Raja polystigma | 21.42 |
| Dasyatis centroura | 0 | 0 | Pteroplatytrygon violacea | 5.77 |
| Dasyatis pastinaca | 3.06 | 8.72 | Pteroplatytrygon violacea | 7.89 |
| Pteroplatytrygon violacea | 0.24 | 0.46 | Dasyatis centroura | 5.77 |
| Mobula mobular | N/A | N/A | Myliobatis aquila | 16.36 |
| Myliobatis aquila | 0.29 | 1.02 | Pteroplatytrygon violacea | 15.59 |
| Pteromylaeus bovinus | 0.11 | 0.17 | Myliobatis aquila | 16.79 |
| Dipturus nidarosiensis | 0.03 | 0.16 | Dipturus oxyrinchus | 5.96 |
| Dipturus oxyrinchus | 0 | 0 | Dipturus nidarosiensis | 5.96 |
| Leucoraja circularis | 0.03 | 0.16 | Leucoraja melitensis | 3.60 |
| Leucoraja melitensis | 0.34 | 0.63 | Leucoraja circularis | 3.60 |
| Leucoraja naevus | N/A | N/A | Leucoraja circularis | 5.02 |
| Raja asterias | 0.43 | 0.79 | Raja radula | 3.96 |
| Raja brachyura | 0.08 | 0.16 | Raja polystigma | 4.09 |
| Raja clavata | 0.06 | 0.47 | Raja radula | 2.51 |
| Raja miraletus | 0.30 | 1.28 | Raja polystigma | 6.79 |
| Raja montagui | N/A | N/A | Raja polystigma | 1.71 |
| Raja polystigma | 0.31 | 1.26 | Raja montagui | 1.71 |
| Raja radula | 0.35 | 1.11 | Raja clavata | 2.51 |
| Raja undulata | 0.31 | 0.64 | Raja polystigma | 3.68 |
| Rostroraja alba | 0.05 | 0.18 | Leucoraja melitensis | 12.24 |
| Torpedo marmorata | 0.42 | 1.30 | Torpedo torpedo | 10.77 |
| Torpedo nobiliana | 0.19 | 0.33 | Torpedo marmorata | 12.83 |
| Torpedo torpedo | 0.05 | 0.34 | Torpedo marmorata | 10.77 |
| Prionace glauca | 0.19 | 0.19 | Galeorhinus galeus | 10.63 |
| Galeus melastomus | 0.24 | 1.55 | Galeorhinus galeus | 13.63 |
| Scyliorhinus canicula | 0.35 | 1.40 | Scyliorhinus stellaris | 7.10 |
| Scyliorhinus stellaris | 0 | 0 | Scyliorhinus canicula | 7.10 |
| Galeorhinus galeus | 0 | 0 | Mustelus punctulatus | 9.25 |
| Mustelus mustelus | 0.07 | 0.64 | Mustelus punctulatus | 7.46 |
| Mustelus punctulatus | 0 | 0.18 | Mustelus mustelus | 7.46 |
| Heptranchias perlo | 0 | 0 | Centroscymnus coelolepis | 17.85 |
| Alopias vulpinus | 0.12 | 0.18 | Cetorhinus maximus | 13.72 |
| Cetorhinus maximus | 0 | 0 | Alopias vulpinus | 13.72 |
| Centrophorus granulosus | 0.08 | 0.52 | Squalus acanthias | 9.36 |
| Dalatias licha | 0.05 | 0.16 | Squalus acanthias | 12.40 |
| Etmopterus spinax | 0.28 | 1.15 | Centrophorus granulosus | 13.91 |
| Oxynotus centrina | 0 | 0 | Centroscymnus coelolepis | 7.82 |
| Centroscymnus coelolepis | N/A | N/A | Oxynotus centrina | 7.82 |
| Squalus acanthias | 0.11 | 1.10 | Squalus blainville | 6.87 |
| Squalus blainville | 0.33 | 1.30 | Squalus acanthias | 6.87 |
| Average | 0.29 | 0.93 | 8.79 | |
Fig 3. Maximum intraspecific distance plotted against Nearest Neighbour distance (p-distance values) for the COI barcode sequences of the 41 chondrichthyan species.

The black rhombus represents the value for Dasyatis pastinaca before its separation into two sub-clusters 1 and 2. The 1:1 equivalence (straight line) is indicated.
The BIN discordance analysis showed 17 discordant BINs out of 42 (40.47%). After reviewing the reasons for the observed discordance, 11 BINs could be reclassified as concordant (Table 3). This included cases such as the M. mustelus BIN with 72 records of M. mustelus and three sequences attributed to a provisional name, indicating that the discordance was caused by the use of an interim name. Other cases could be ascribed to erroneous morphological identifications of one or two records associated with the BIN, e.g. Raja clavata, where a few records were assigned differently. The five remaining discordant BINs contained several individuals belonging to more than one species, indicating that for these species either the barcode sequence or the BIN algorithm is not sufficient to discriminate them. However, the possibility of misidentification cannot be excluded either. Lastly, BIN AAD5036 associated with the D. pastinaca sub-cluster 2 consists of six individuals of Dasyatis tortonesei from the Muséum National d’Historie Naturelle, Paris, indicating a possible species misidentification of the four ELASMOMED specimens (Table 3).
Table 3. Species with discordant BINs found by the ‘BIN Discordance Report’ sequence analysis tool on BOLD.
Taxonomic ranks of conflict are reported including species of the same genus up to taxa belonging to different orders. The column BIN Taxon Variation shows the number of records for each taxon in parenthesis.
| Species | Discordant BIN | Rank of conflict | BIN Taxon Variation | After review |
|---|---|---|---|---|
| Dasyatis pastinaca 1 | BOLD:ACK8259 | Family | Dasyatidae[20], Rhinobatidae[1] | Concordant |
| Squalus blainville | BOLD:AAA1550 | Family | Squalidae[131], Centrophoridae[1] | Concordant |
| Prionace glauca | BOLD:AAA7096 | Genus | Prionace[68], Carcharhinus[1] | Concordant |
| Dasyatis centroura | BOLD:AAD5044 | Species | Dasyatis ushiei[7], Dasyatis centroura[7] | Discordant |
| Dasyatis pastinaca 2 | BOLD:AAD5036 | Species | Dasyatis tortonesei[6], Dasyatis pastinaca[4] | Discordant |
| Mobula mobular | BOLD:AAB8636 | Species | Mobula japanica[27], Mobula mobular[2], Mobula sp.[1], Mobula japonica[1] | Discordant |
| Dipturus oxyrinchus | BOLD:ABZ4263 | Species | Dipturus oxyrinchus sp1[56], Dipturus oxyrinchus[49] | Concordant |
| Raja brachyura | BOLD:AAA4358 | Species | Raja brachyura[36], Raja sp.[1] | Concordant |
| Raja clavata | BOLD:ACF2419 | Species | Raja clavata[107], Raja undulata[1] | Concordant |
| Raja polystigma | BOLD:ABY6158 | Species | Raja polystigma[24], Raja sp.[1] | Concordant |
| Torpedo nobiliana | BOLD:AAC6970 | Species | Torpedo nobiliana[9], Torpedo macneilli[5], Torpedo fairchildi[4], Torpedo tokionis[1], Torpedo sp. A[1] | Discordant |
| Mustelus mustelus | BOLD:AAA4345 | Species | Mustelus mustelus[72], Mustelus sp.[3] | Concordant |
| Mustelus punctulatus | BOLD:AAA4347 | Species | Mustelus punctulatus[164], Mustelus sp. zpl 00058[1] | Concordant |
| Centrophorus granulosus | BOLD:AAB4327 | Species | Centrophorus granulosus[59], Centrophorus zeehaani[28], Centrophorus lusitanicus[1] | Discordant |
| Oxynotus centrina | BOLD:AAF2493 | Species | Oxynotus centrina[8], Oxynotus paradoxus[1] | Concordant |
| Centroscymnus coelolepis | BOLD:AAB8284 | Species | Centroscymnus coelolepis[20], Centroscymnus sp.[1] | Concordant |
| Squalus acanthias | BOLD:AAA1547 | Species | Squalus acanthias[242], Squalus suckleyi[32] | Discordant |
We applied a ranking system analysis to all the barcoded species, treating the subclusters 1 and 2 of D. pastinaca as different species. The analysis ranked 36 species at Grade A (85.72%) and six species at Grade E (14.28%) (Table 4), with the last showing the post-revision BIN discordance. As all prior analyses confirmed the misidentification of specimens in the D. pastinaca sub-cluster 2, the four specimens belonging to this group were assigned to D. tortonesei, upgrading this cluster to Grade A. Both species are differentiated by 26 nucleotide substitutions (3.66%; S1 Fig). Individuals of D. tortonesei were found only in the Ionian division (FAO 37.2.2). All four samples were collected in the Strait of Sicily (South-Eastern part of the Ionian FAO division), while those of D. pastinaca were collected from the divisions Balearic (FAO 37.1.1), Sardinia (FAO 37.1.2) and Ionian (FAO 37.2.2).
Table 4. Attribution of grades from A (full concordance) to E (full discordance) to DNA barcodes of the 42 chondrichthyan species of the ELASMOMED reference library, following the ranking system proposed by Costa et al. (2012).
The Dasyatis pastinaca sub-clusters 1 and 2 are graded separately.
| Species | Grade | Species | Grade | Species | Grade |
|---|---|---|---|---|---|
| Chimaera monstrosa | A | Raja brachyura | A | Scyliorhinus stellaris | A |
| Dasyatis centroura | E | Raja clavata | A | Galeorhinus galeus | A |
| Dasyatis pastinaca 1 | A | Raja miraletus | A | Mustelus mustelus | A |
| Dasyatis pastinaca 2 | E | Raja montagui | A | Mustelus punctulatus | A |
| Pteroplatytrygon violacea | A | Raja polystigma | A | Heptranchias perlo | A |
| Mobula mobular | E | Raja radula | A | Alopias vulpinus | A |
| Myliobatis aquila | A | Raja undulata | A | Cetorhinus maximus | A |
| Pteromylaeus bovinus | A | Rostroraja alba | A | Centrophorus granulosus | E |
| Dipturus nidarosiensis | A | Torpedo marmorata | A | Dalatias licha | A |
| Dipturus oxyrinchus | A | Torpedo nobiliana | E | Etmopterus spinax | A |
| Leucoraja circularis | A | Torpedo torpedo | A | Oxynotus centrina | A |
| Leucoraja melitensis | A | Prionace glauca | A | Centroscymnus coelolepis | A |
| Leucoraja naevus | A | Galeus melastomus | A | Squalus acanthias | E |
| Raja asterias | A | Scyliorhinus canicula | A | Squalus blainville | A |
Among juvenile specimens, cases of species misidentification were encountered among smooth hound sharks, cat sharks and dogfish. Misidentification of skates and electric rays were recorded both for juveniles and adults (Table 1). Five juveniles of M. punctulatus from the Balearic division (FAO 37.1.1) as well as 45 juvenile specimens from the Adriatic division (FAO 37.2.1) were misidentified as M. mustelus, confirming that the characters used in the taxonomic keys are not adequate for immature individuals. The congeneric species Scyliorhinus canicula and S. stellaris collected in the Adriatic division (FAO 37.2.1) were confused in four cases. Moreover, four out of the 12 dogfishes collected in the Ionian (Strait of Sicily) division (FAO 37.2.2) were initially assigned morphologically to S. megalops, but were barcoded as S. blainville. A similar taxonomic confusion between Squalus acanthias and S. blainville occurred in both Balearic (FAO 37.1.1) and Levant (FAO 37.3.2) divisions.
Among rays and skates, T. marmorata individuals, one from Adriatic (FAO 37.2.1) and three from Levant (FAO 37.3.2) divisions were morphologically misidentified as T. nobiliana or T. torpedo because diagnostic spots are absent in juvenile specimens (Table 1). Two specimens of R. polystigma were misidentified as R. montagui. Both were collected in the Balearic division (FAO 37.1.1), a unique Mediterranean area where both species co-occur in sympatry. Other minor morphological misidentifications concern a few specimens of Centrophorus granulosus, Pteroplatytrygon violacea, D. centroura and L. circularis (Table 1).
Thirty-four out of the 42 barcoded species were assessed for spatial variation as multiple records were obtained from several FAO divisions (S2 Table). Among those, five batoids showed noticeable phylogeographic signals in the COI sequence variation, mainly represented by private haplotypes of some FAO divisions (Fig 4; the complete list of mutations and positions characterizing each haplotype is reported in S4 Table). Even if the present sample size is too limited to allow solid and definitive phylogeographic inferences, the data allow some preliminary insights from the patterns observed (Fig 4).
Fig 4. Haplotype parsimony networks of Raja miraletus, Raja asterias, Raja polystigma, Raja radula and Torpedo marmorata reconstructed using ELASMOMED COI barcode sequences.

Each circle represents one haplotype and its size is proportional to frequency. Colours indicate the origin of samples according to FAO fishing divisions. Mutations and positions characterizing each haplotype, as well as the size (bp), and the number of the sequences included in each species network, are provided in S4 Table.
Raja miraletus showed eight COI haplotypes, with a single private haplotype (Hrm8) comprising all samples from the Levant division. Raja asterias consisted of three slightly different haplotype groups: Hra4 and Hra5 for the Balearic division, Hra1-3 for Sardinia and Ionian divisions, and Hra6-9 from the Adriatic division. The latter group also included haplotype Hra10 that contained only a single specimen from the Strait of Sicily. Raja polystigma exhibited five haplotypes with the private haplotype Hrp5 of the Adriatic division, while all the others are shared among areas of the Western and Central Mediterranean. Two specimens of Raja radula from the Levant division formed private haplotype Hrr6, differentiated by three mutations from the Balearic haplotypes (Hrr1-5). Torpedo marmorata contained three haplotype groups: one formed by Htm1 and Htm2 found in the Balearic, Sardinia and Ionian divisions, the second formed by Htm3 and Htm4 found in individuals from Ionian and Adriatic divisions, and the third included the haplotypes Htm5 and Htm6 found only in the Levantine individuals (Fig 4).
Discussion
Towards completion of a DNA barcode reference library of the mediterranean chondrichthyes
The ELASMOMED initiative sampled and barcoded 42 out of the 89 chondrichthyan species occurring in the Mediterranean Sea, contributing ex novo barcodes for 24 species. In addition, it added several barcoded specimens from different geographical areas to those from previous initiatives. The integration of our results with those of other recent major fish barcoding initiatives carried out in the Mediterranean [39,92] raises the total count to 51 species with DNA barcodes (S1 Table), comprising the holocephalan Chimaera monstrosa, 26 sharks (53%), and 24 skates and rays (62%).
As the main source of specimens for the ELASMOMED initiative was the MEDITS program, demersal and bathydemersal species (N = 32) outnumbered pelagic and benthopelagic ones (N = 10; S1 Table). Regarding endemic species, barcode sequences of R. asterias, R. polystigma and M. mobular were newly obtained and the number of records for L. melitensis and R. radula greatly increased. As a symptom of the dangerously limited knowledge characterising the Mediterranean chondrichthyans faunas, only one sequence for each species was available until this study [92]. Multiple sequences from each species are needed to properly assess intra- vs inter-specific sequences variation and thus improve the robustness of the molecular assignment. Only one out of nine known alien species (C. altimus) was barcoded [39]. Of the 38 Mediterranean species without a barcode, 32 have been barcoded from other regions and only six have not been barcoded at all (Rhinoptera marginata, Glaucostegus halavi, Pristis pristis, Rhinobatos cemiculus, Torpedo alexandrinsis and Raja africana; S1 Table).
The ELASMOMED network allowed for an unprecedented large spatial coverage of data encompassing five FAO fishing divisions (S2 Table).Notably, ELASMOMED increased the number of barcoded species from North African countries, as 25 species collected from the Algerian waters were added to those of an earlier study [39]. About 43% of the Mediterranean Chondrichthyan diversity remains to be inventoried and ongoing efforts are needed to increase the number of species collected from both already targeted and remaining areas.
Despite a slower mutation rate of mtDNA genes in sharks [93], our results showed that COI barcoding is a reliable and efficient method for specimens identification, consistent with the findings of previous studies in marine fish [82,89,94,95]. The effectiveness of DNA barcoding was also demonstrated by the cohesive monophyletic clustering in our Neighbour-Joining analysis, the high number of concordant BINs and the majority of Grade A rankings (37 out of 42 species—88.1%) following the Costa el al. [89] criteria. Only in five species, categorized as Grade E, barcode sequences did not allow the discrimination of Mediterranean species from congeners inhabiting the Indo-Pacific Ocean, namely C. granulosus from Centrophorus zeehaani, D. centroura from Dasyatis ushiei, M. mobular from M. japanica, S. acanthias from Squalus suckley and T. nobiliana from Torpedo macneilli, Torpedo fairchildi and Torpedo tokionis. This barcode sharing could be attributed to various reasons, such as incomplete lineage sorting, past lineage introgression, taxonomic uncertainties, or matched sequences on BOLD were generated from misidentified specimens.
Although Neighbour-Joining trees based on COI sequence divergence alone are not useful as a phylogenetic tool, they allow limited insights into relationships at higher taxonomic levels [82]. All skate orders and two shark orders (Lamniformes, Squaliformes) were resolved as monophyletic groups with high bootstrap support (98–100)). Intermediate taxonomic levels such as family and genus were not well resolved. For example, the blackmouth catshark, Galeus melastomus (Scyliorhinidae) appears closer related to Prionace glauca (Carcharhinidae), M. mustelus, M. punctulatus and Galeorhinus galeus (Triakidae) than to other scyliorhinid sharks such as S. canicula and S. stellaris; Myliobatidae (Myliobatis aquila and Pteromylaeus bovinus) did not form a monophyletic clade, but rather cluster with the whiptail stingrays of the family Dasyatidae. Paraphyletic clusters were also observed for some Raja, Dasyatis and Dipturus species. These data emphasize the need for a classification revision and illustrate the limits of a single marker gene as phylogenetic tool [82]. However, some clusters might reflect insufficient taxonomy, e.g. the placement of D. centroura close to P. violacea as opposed to D. pastinaca and D. tortonesei, which supports the need for a taxonomic revision of the Mediterranean Dasyatidae. Such unresolved cases could be clarified by using a combined morphological and molecular phylogenetic analyses, as successfully performed for the Indo-Pacific stingrays [96]. The barcode-referenced library of voucher tissues held by ELASMOMED represents a unique opportunity for subsequent research intended to achieve taxonomic improvements.
From molecular taxonomy to conservation and management issues: the added value of DNA barcoding for chondrichthyans
Persistent problems regarding the morphological taxonomy of elasmobranchs are their pronounced morphological stasis and the fact that distinctive diagnostic characters are only exhibited in adults [51,65,66]. On-board identification of specimens during international trawl surveys usually involves several fisheries scientists from different research institutes or organizations, with varying levels of taxonomic skills and expertise. Field sorting and taxonomic classification are generally carried out using dichotomic keys and identification guides based on characters shown mainly in the adult stage [5]. This approach itself was shown to be misleading, because most individuals caught are either juveniles or sub-adults [83]. The approach adopted by the ELASMOMED Consortium has proven to be effective in detecting cases of misidentification among gulper sharks, smooth hound sharks, cat sharks, sting rays and electric rays. These instances are ascribable to a lack of clear traits and diagnostic characters, especially in immature individuals. The discrimination between Centrophorus granulosus and C. uyato is challenging and the taxonomy of the genus has been controversial and in need of revision[97–99]. The absence of characteristic dorsal black dots on the skin of juveniles of M. punctulatus (TL < 60cm) can cause confusion with M. mustelus [70]. Similarly, the characteristic dorsal coloration of mottled light on dark background and the knob-rimmed margins of the spiracles of Torpedo marmorata are not pronounced enough in small-size individuals which are often confused with the closely related species T. nobiliana. In cat sharks, the traits for discriminating S. canicula and S. stellaris are the distinct shape of the anterior nasal flap and the distribution pattern of coloured spots on the animal’s skin [39,100]. These diagnostic features were not clearly assessed in a few individuals collected in the Adriatic Sea. The genus Raja and, in particular, the sibling sister species R. montagui and R. polystigma, are prime examples of taxa with very similar morphological characters both in immature and mature individuals because of a very high level of stasis [22,73,101,102]. Such groups require very accurate, detailed, and time-consuming traditional taxonomic analyses of several individuals in order to assess species-specific characters. DNA barcoding can assign individuals to species even if this is not possible using morphological characters (e.g. in sibling and cryptic species) providing a basic yet very efficient tool to establish reliable diagnostic species-specific features in such problematic taxa. The prerequisite of such an important process is the development and implementation of large, almost complete and constantly updated barcode reference libraries with associated voucher specimens as started by ELASMOMED and other similar studies (e.g. [92]).
Such large repositories consisting of tissues and barcode sequences of dozens of samples per species collected across its entire known distribution area would also allow preliminary exploration of population structure. Intraspecific variation of barcode sequences is intrinsically low, but despite this and the rather low number of individuals and samples analysed we were able to detect signals of phylogeographic structure within some groups of the ELASMOMED Mediterranean dataset. Such preliminary evidence is insufficient and requires further inference of evolutionary patterns suitable for addressing management and conservation issues. The five batoid species R. miraletus, R. asterias, R. polystigma, R. radula and T. marmorata exhibited small but detectable phylogeographic structure among the Mediterranean samples, with divergent private haplotypes detected in the Levant area or, whenever the species do not occur that far to east (i.e. R. asterias and R. polystigma), in the Adriatic Sea. In addition, R. asterias showed also two private haplotypes in the Balearic area, likely highlighting a more marked geographical structure than the other four species. Geographical genetic breaks need to be accurately accounted for when delineating management and conservation units. Behavioural ecology and life-history traits of demersal and bathydemersal skates and rays [103–105] make this group particularly prone to genetic structuring and several cases have already been documented for the region [73,106,107]. Other studies used different mtDNA markers such as parts of the control region or cytochrome b (see for R. clavata [73,106]) to accurately unravel geographical structuring of populations. Recent studies in the Mediterranean used barcode data to assess genetic differentiation of the small-spotted catshark [44,45] and their findings describe phylogeographic signals comparable to those inferred by our ELASMOMED dataset.
Efforts toward the completion of the ELASMOMED database should be continued to expand taxonomic coverage and to improve the assessment of spatial and temporal patterns of species diversity of Chondrichthyes in the Mediterranean. The completion of the ELASMOMED barcode library will allow i) more accurate species-specific information from scientific survey and landing data essential to correctly assess stock status of Chondrichthyans [63,108]; ii) the detection of cryptic and invasive species and iii) the forensic traceability of cartilaginous fish products to fight illegal and unreported fisheries and support commercial trade [109].
DNA barcoding is also the starting point for other fast-evolving DNA-based techniques. The widespread use of environmental DNA allows for the extraction short sequences of multiple-species from complex matrices (e.g. seawater) in form of metabarcoding data [77,110–113].Because of the nature of this methodology, comprehensive barcoding libraries are even more essential in providing valid comparative data and strengthening biodiversity monitoring different geographical scales.
Supporting Information
Both the number of barcode records obtained from Mediterranean and non-Mediterranean specimens are reported. When the number of non-Mediterranean barcodes on BOLD was larger than 10, the exact number is not reported. Species author, Ecology (Bathydemersal, Demersal, Pelagic and Benthopelagic), IUCN Red List Category and Criteria (CR = Critically Endangered, EN = Endangered, VU = Vulnerable, NT = Near Threatened, LC = Least Concern, DD = Data Deficient, NE = Not Evaluated), Status and BIN available on BOLD are detailed for each species. Asterisks denote barcode sequences deposited in public repositories but not in BOLD.
(XLSX)
The number of collected and barcoded individuals are reported for each chondrichthyan species and each FAO division as well as in total.
(XLSX)
(XLSX)
For each species, the number of sequences used and the length of the alignment are reported.
(XLSX)
Each circle represents one haplotype and its size is proportional to frequency. Colours indicate the origin of samples according to FAO fishing divisions.
(TIF)
Acknowledgments
We thank Victoria Bertucci, Alessandro Velonà, Diego Galafassi, Nicola Di Francesco, Cecilia Pinto, Alessandra Nero, Lisa Capelli, Matteo Bertozzi, Paola Pasolini, Chiara Manfredi and Corrado Piccinetti (University of Bologna); Angelo Cau (University of Cagliari); Alvaro Abella, Roberto Bedini, Roberto Silvestro, Alessandro Voliani, Michela Ria and Enrico Cecchi (ARPAT Livorno); Giuseppe Scarcella (CNR-ISMAR Ancona); Kostas Efthimiadis (DFMR-MOA Republic of Cyprus) Nicola Ungaro (ARPA Puglia) for their contribution and support in sampling activities.
We would like to acknowledge Claudia Steinke and James Robertson (BOLD) for their expertise and help in managing the ELASMOMED Consortium Project.
This research work was supported by the University of Bologna, the Canadian Centre for DNA Barcoding, and the Biodiversity Institute of Ontario both supported by the government of Canada through Genome Canada and the Ontario Genomics Institute.
Data Availability
Sampling and biological data as well as digital images of dorsal/ventral sides (when recorded) are available in the “ELASMOMED Consortium” project (Project Code: ELAMO) accessible on the Barcode of Life Data system (BOLD, http://www.barcodinglife.org). GenBank accession numbers for all barcoded specimens are reported in S3 Table.
Funding Statement
This research work was supported by the University of Bologna, the Canadian Centre for DNA Barcoding, and the Biodiversity Institute of Ontario both supported by the government of Canada through Genome Canada and the Ontario Genomics Institute. Rita Cannas, Maria Cristina Follesa and Alessandro Cau were funded by the Autonomous Region of Sardinia (RAS) grant n° L.R.7 CRP-25321. Dirk Steinke was funded by the Alfred P Sloan Foundation. These latter funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Hebert PDN, Cywinska A, Ball SL, DeWaard JR. Biological identifications through DNA barcodes. Proc R Soc London B Biol Sci. 2003;270: 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Savolainen V, Cowan RS, Vogler AP, Roderick GK, Lane R. Towards writing the encyclopedia of life: an introduction to DNA barcoding. Philos Trans R Soc London B Biol Sci. 2005;360: 1805–1811. 10.1098/rstb.2005.1730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vences M, Thomas M, Bonett RM, Vieites DR. Deciphering amphibian diversity through DNA barcoding: chances and challenges. Philos Trans R Soc Lond B Biol Sci. 2005;360: 1859–1868. 10.1098/rstb.2005.1717 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Saunders GW. Applying DNA barcoding to red macroalgae: a preliminary appraisal holds promise for future applications. Philos Trans R Soc Lond B Biol Sci. 2005;360: 1879–1888. 10.1098/rstb.2005.1719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pegg GG, Sinclair B, Briskey L, Aspden WJ. MtDNA barcode identification of fish larvae in the southern Great Barrier Reef, Australia. Sci Mar. 2006;70S2: 7–12. [Google Scholar]
- 6.Bickford D, Lohman DJ, Sodhi NS, Ng PKL, Meier R, Winker K, et al. Cryptic species as a window on diversity and conservation. Trends Ecol Evol. 2007;22: 148–155. 10.1016/j.tree.2006.11.004 [DOI] [PubMed] [Google Scholar]
- 7.Puncher GN, Alemany F, Arrizabalaga H, Cariani A, Tinti F. Misidentification of bluefin tuna larvae: a call for caution and taxonomic reform. Rev Fish Biol Fish. 2015;25: 485–502. [Google Scholar]
- 8.Murphy RW, Crawford AJ, Bauer AM, Che J, Donnellan SC, Fritz U, et al. Cold Code: the global initiative to DNA barcode amphibians and nonavian reptiles. Mol Ecol Resour. 2013;13: 161–167. [Google Scholar]
- 9.Pfenninger M, Nowak C, Kley C, Steinke D, Streit B. Utility of DNA taxonomy and barcoding for the inference of larval community structure in morphologically cryptic Chironomus (Diptera) species. Mol Ecol. 2007;16: 1957–68. 10.1111/j.1365-294X.2006.03136.x [DOI] [PubMed] [Google Scholar]
- 10.Carolan JC, Murray TE, Fitzpatrick Ú, Crossley J, Schmidt H, Cederberg B, et al. Colour Patterns Do Not Diagnose Species: Quantitative Evaluation of a DNA Barcoded Cryptic Bumblebee Complex. PLoS One. Public Library of Science; 2012;7: 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Plass-Johnson JG, McQuaid CD, Hill JM. Morphologically similar, coexisting hard corals (Porites lobata and P. solida) display similar trophic isotopic ratios across reefs and depths. Mar Freshw Res. 2016;67: 671–676. [Google Scholar]
- 12.Andrews KR, Williams AJ, Fernandez-Silva I, Newman SJ, Copus JM, Wakefield CB, et al. Phylogeny of deepwater snappers (Genus Etelis) reveals a cryptic species pair in the Indo-Pacific and Pleistocene invasion of the Atlantic. Mol Phylogenet Evol. Elsevier Inc.; 2016;100: 361–371. 10.1016/j.ympev.2016.04.004 [DOI] [PubMed] [Google Scholar]
- 13.Ashton TJ, Kayoueche-Reeve M, Blight AJ, Moore J, Paterson DM. Duplex DNA barcoding allows accurate species determination of morphologically similar limpets (Patella spp.) from non-destructive sampling. J Mar Biol Assoc United Kingdom. 2016; 1–4. [Google Scholar]
- 14.Knowlton N. Sibling Species in the Sea. Annu Rev Ecol Syst. 1993;24: 189–216. [Google Scholar]
- 15.Zimmer RK, Ferrier GA, Kim SJ, Kaddis CS, Zimmer CA, Loo JA. A multifunctional chemical cue drives opposing demographic processes and structures ecological communities. Ecology. 2016;97: 2232–2239. 10.1002/ecy.1455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Camperi M, Tricas TC, Brown BR. From morphology to neural information: The electric sense of the skate. PLoS Comput Biol. 2007;3: 1083–1096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sisneros JA, Tricas TC. Neuroethology and life history adaptations of the elasmobranch electric sense. J Physiol Paris. 2002;96: 379–389. 10.1016/S0928-4257(03)00016-0 [DOI] [PubMed] [Google Scholar]
- 18.Klimley AP. The biology of sharks and rays University of Chicago Press; 2013. [Google Scholar]
- 19.Straube N, Kriwet J, Schliewen UK. Cryptic diversity and species assignment of large lantern sharks of the Etmopterus spinax clade from the Southern Hemisphere (Squaliformes, Etmopteridae). Zool Scr. 2011;40: 61–75. [Google Scholar]
- 20.Griffiths AM, Sims DW, Cotterell SP, El Nagar A, Ellis JR, Lynghammar A, et al. Molecular markers reveal spatially segregated cryptic species in a critically endangered fish, the common skate (Dipturus batis). Proc R Soc London B Biol Sci. 2010;277: 1497–1503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Iglésias SP, Toulhoat L, Sellos DY. Taxonomic confusion and market mislabelling of threatened skates: important consequences for their conservation status. Aquat Conserv Mar Freshw Ecosyst. Wiley Online Library; 2010;20: 319–333. [Google Scholar]
- 22.Frodella N, Cannas R, Velonà A, Carbonara P, Farrell ED, Fiorentino F, et al. Population connectivity and phylogeography of the Mediterranean endemic skate Raja polystigma and evidence of its hybridization with the parapatric sibling R. montagui. Mar Ecol Prog Ser. 2016;554: 99–113. [Google Scholar]
- 23.Ovenden JR, Morgan JAT, Kashiwagi T, Broderick D, Salini J. Towards better management of Australias shark fishery: Genetic analyses reveal unexpected ratios of cryptic blacktip species Carcharhinus tilstoni and C. limbatus. Mar Freshw Res. 2010;61: 253–262. [Google Scholar]
- 24.Quattro JM, Stoner DS, Driggers WB, Anderson CA, Priede KA, Hoppmann EC, et al. Genetic evidence of cryptic speciation within hammerhead sharks (Genus Sphyrna). Mar Biol. 2006;148: 1143–1155. [Google Scholar]
- 25.Sandoval-Castillo J, Rocha-Olivares A, Villavicencio-Garayzar C, Balart E. Cryptic isolation of Gulf of California shovelnose guitarfish evidenced by mitochondrial DNA. Mar Biol. 2004;145: 983–988. [Google Scholar]
- 26.Trontelj P, Fiser C. Cryptic species diversity should not be trivialised. Syst Biodivers. 2009;7: 1–3. [Google Scholar]
- 27.Meier R, Zhang G, Ali F. The use of mean instead of smallest interspecific distances exaggerates the size of the “barcoding gap” and leads to misidentification. Syst Biol. 2008;57: 809–813. 10.1080/10635150802406343 [DOI] [PubMed] [Google Scholar]
- 28.Lindenmayer D, Burgman M. Practical conservation biology. Csiro Publishing; 2005. [Google Scholar]
- 29.King M. Fisheries biology, assessment and management John Wiley & Sons; 2013. [Google Scholar]
- 30.Daugherty CH, Patterson GB, Hitchmough R a. Taxonomic and conservation review of the New Zealand herpetofauna. New Zeal J Zool. 1994;21: 317–323. [Google Scholar]
- 31.Daugherty CH, Cree A, Hay JM, Thompson M. Neglected taxonomy and continuing extinctions of tuatara(Sphenodon). Nature. 1990;374: 177–179. [Google Scholar]
- 32.Agapow P, Bininda-Emonds ORP, Crandall KA, Gittleman JL, Mace GM, Marshall JC, et al. The Impact of Species Concept on Biodiversity Studies. Q Rev Biol. 2004;79: 161–179. [DOI] [PubMed] [Google Scholar]
- 33.Crozier RH, Dunnett LJ, Agapow PM. Phylogenetic biodiversity assessment based on systematic nomenclature. Evol Bioinform Online. 2005;1: 11–36. [PMC free article] [PubMed] [Google Scholar]
- 34.Becker S, Hanner R, Steinke D. Five years of FISH-BOL: Brief status report. Mitochondrial DNA. 2011;22: 3–9. [DOI] [PubMed] [Google Scholar]
- 35.Ward RD, Hanner R, Hebert PDN. The campaign to DNA barcode all fishes, FISH-BOL. J Fish Biol. 2009;74: 329–356. 10.1111/j.1095-8649.2008.02080.x [DOI] [PubMed] [Google Scholar]
- 36.Ward RD. FISH-BOL, A Case Study for DNA Barcodes. DNA barcodes:Methods and Protocols. 2012. pp. 423–439. [DOI] [PubMed]
- 37.Coulson MW, Denti D, Van Guelpen L, Miri C, Kenchington E, Bentzen P. DNA barcoding of Canada’s skates. Mol Ecol Resour. 2011;11: 968–978. 10.1111/j.1755-0998.2011.03034.x [DOI] [PubMed] [Google Scholar]
- 38.Lynghammar A, Christiansen JS, Griffiths AM, Fevolden SE, Hop H, Bakken T. DNA barcoding of the northern Northeast Atlantic skates (Chondrichthyes, Rajiformes), with remarks on the widely distributed starry ray. Zool Scr. 2014;43: 485–495. [Google Scholar]
- 39.Moftah M, Abdel Aziz SH, Elramah S, Favereaux A. Classification of Sharks in the Egyptian Mediterranean Waters Using Morphological and DNA Barcoding Approaches. PLoS One. 2011;6: e27001 10.1371/journal.pone.0027001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Serra-Pereira B, Moura T, Griffiths AM, Serrano Gordo L, Figueiredo I. Molecular barcoding of skates (Chondrichthyes: Rajidae) from the southern Northeast Atlantic. Zool Scr. 2011;40: 76–84. [Google Scholar]
- 41.Smith PJ, Steinke D, Mcveagh SM, Stewart AL, Struthers CD, Roberts CD. Molecular analysis of Southern Ocean skates (Bathyraja) reveals a new species of Antarctic skate. J Fish Biol. 2008;73: 1170–1182. [Google Scholar]
- 42.Spies IB, Gaichas S, Stevenson DE, Orr JW, Canino MF. DNA-based identification of Alaska skates (Amblyraja, Bathyraja and Raja: Rajidae) using cytochrome c oxidase subunit I (coI) variation. J Fish Biol. 2006;69: 283–292. [Google Scholar]
- 43.Ward RD, Holmes BH, White WT, Last PR. DNA barcoding Australasian chondrichthyans: Results and potential uses in conservation. Mar Freshw Res. 2008;59: 57–71. [Google Scholar]
- 44.Barbieri M, Maltagliati F, Roldán MI, Castelli A. Molecular contribution to stock identification in the small-spotted catshark, Scyliorhinus canicula (Chondrichthyes, Scyliorhinidae). Fish Res. 2014;154: 11–16. [Google Scholar]
- 45.Kousteni V, Kasapidis P, Kotoulas G, Megalofonou P. Strong population genetic structure and contrasting demographic histories for the small-spotted catshark (Scyliorhinus canicula) in the Mediterranean Sea. Heredity (Edinb). 2014;114: 333–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bertrand JA, Gil de Sola L, Papaconstantinou C, Relini G, Souplet A. The general specifications of the MEDITS surveys. Sci Mar. 2002;66: 9–17. [Google Scholar]
- 47.Ben Souissi J, Golani D, Méjri H, Ben Salem M, Capapé C. First confirmed record of the Halave’s Guitarfish, Rhinobatos halavi (Forsskål, 1775) (Chondrichthyes: Rhinobatidae) in the Mediterranean Sea with a description of a case of albinism in elasmobranchs. Cah Biol March 2007;48: 67–75. [Google Scholar]
- 48.Bradai MN, Saidi B, Enajjar S. Elasmobranchs of the Mediterranean and Black Sea: Status, Ecology and Biology. Bibliographic Analysis. FAO; 2012.
- 49.Compagno LJV. Sharks of the world, an annotated and illustrated catalogue of shark species known to date—bullhead, mackerel & carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). Report No. 1, vol. 2.FAO; 2001.
- 50.Psomadakis PN, Giustino S, Vacchi M. Mediterranean fish biodiversity: an undated inventory with focus on the Ligurian and Tyrrhenian seas. Zootaxa. 2012;3263: 1–46. [Google Scholar]
- 51.Serena F. Field identification Guide to the sharks and rays of the Mediterranean and Black Sea. FAO Species Identification Guide for Fishery Purposes. FAO 97 pp; 2005.
- 52.Poortvliet M, Olsen JL, Croll DA, Bernardi G, Newton K, Kollias S, et al. A dated molecular phylogeny of manta and devil rays (Mobulidae) based on mitogenome and nuclear sequences. Mol Phylogenet Evol. 2015;83: 72–85. 10.1016/j.ympev.2014.10.012 [DOI] [PubMed] [Google Scholar]
- 53.Nieto A, Ralph GM, Comeros-Raynal MT, Kemp J, García Criado M, Allen DJ, et al. European Red List of Marine Fishes. European Commission; 2015. [Google Scholar]
- 54.CIESM. CIESM Atlas of Exotic Species in the Mediterranean- Vol. 1. Fishes by D. Golani, L. Orsi-Relini, E. Massutí and J.P. Quignard [Internet]. 2002. Available: http://www.ciesm.org/online/atlas/intro.htm
- 55.Zenetos A, Gofas S, Verlaque M, Inar ME, Garci JE, Bianchi CN, et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr Mar Sci. 2011;12/2: 509–514. [Google Scholar]
- 56.Zenetos A, Gofas S, Morri C, Rosso A, Violanti D, García Raso JE, et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr Mar Sci. 2012;13: 328–352. [Google Scholar]
- 57.Tobuni I, Ben Abdallah A, Serena F, Shakman E. First documented presence of Galeocerdo cuvier (Péron & Lesueur, 1822) (ELASMOBRANCHII, CARCHARHINIDAE) in the Mediterranean basin (Libyan waters). Mar Biodivers Rec. [Google Scholar]
- 58.Cannas R, Follesa MC, Cabiddu S, Porcu C, Salvadori S, Iglésias SP, et al. Molecular and morphological evidence of the occurrence of the Norwegian skate Dipturus nidarosiensis (Storm, 1881) in the Mediterranean Sea. Mar Biol Res. 2010;6: 341–350. [Google Scholar]
- 59.Serena F. Lo status degli elasmobranchi dei mari italiani (Elasmostat). I Programma Nazionale triennale della pesca e dell’acquacoltura 2007–2009 (prorogato a tutto il 2012). Progetto di ricerca: “7 –Tematica A3”. Rapporto finale, 28 febbraio 2014. 321 pp. 2014.
- 60.Eschmeyer WN (ed). Catalog of fishes: Genera, species, references [Internet]. 2015 [cited 1 Jan 2015]. Available: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp Electronic version accessed 05 June 2015
- 61.Froese R, Pauly D. FishBase. World Wide Web electronic publication. In: Editors. 2015. FishBase [Internet]. 2015 [cited 1 Jan 2015]. Available: www.fishbase.org, version (02/2015).
- 62.Follesa MC, Cannas R, Cabiddu S, Cau A l., Mulas A, Porcu C, et al. Preliminary observations of the reproductive biology and diet for the Norwegian skate Dipturus nidarosiensis (Rajidae) from the Central Western Mediterranean Sea. Cybium. 2012;36: 473–477. [Google Scholar]
- 63.Dulvy NK, Fowler SL, Musick JA, Cavanagh RD, Kyne PM, Harrison LR, et al. Extinction risk and conservation of the world’s sharks and rays [Internet]. Baldwin IT, editor. 2014. [DOI] [PMC free article] [PubMed]
- 64.Ragonese S, Vitale S, Dimech M, Mazzola S. Abundances of Demersal Sharks and Chimaera from 1994–2009 Scientific Surveys in the Central Mediterranean Sea. PLoS One. 2013;8: e74865 10.1371/journal.pone.0074865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Tinti F, Ungaro N, Pasolini P, De Panfilis M, Garoia F, Guarniero I, et al. Development of molecular and morphological markers to improve species-specific monitoring and systematics of Northeast Atlantic and Mediterranean skates (Rajiformes). J Exp Mar Bio Ecol. 2003;288: 149–165. [Google Scholar]
- 66.Serena F, Mancusi C, Barone M. Field identification guide to the skates (Rajidae) of the Mediterranean Sea. Guidelines for data collection and analysis. Biologia Marina Mediterranea, 17 (Sppl. 2); 2010. [Google Scholar]
- 67.GFCM—General Fisheries Commission for the Mediterranean. Recommendation GFCM/36/2012/3 on fisheries management measures for conservation of sharks and rays in the GFCM area. 2012.
- 68.Bonello J, Bonnici L, Ferrari A, Cariani A, Schembri PJ. Not all that clear cut: intraspecific morphological variability in Squalus blainville (Risso, 1827) and implications for identification of the species. J Mar Biol Assoc United Kingdom. 2015; 1–12. [Google Scholar]
- 69.Marouani S, Chaâba R, Kadri H, Saidi B, Bouain A, Maltagliati F, et al. Taxonomic research on Squalus megalops (Macleay, 1881) and Squalus blainvillei (Risso, 1827) (Chondrichthyes: Squalidae) in Tunisian waters (central Mediterranean Sea). Sci Mar. 2012;76: 97–109. [Google Scholar]
- 70.Marino IAM, Riginella E, Cariani A, Tinti F, Farrell ED, Mazzoldi C, et al. New molecular tools for the identification of 2 endangered Smooth-Hound Sharks, Mustelus mustelus and Mustelus punctulatus. J Hered. 2015;106: 123–130. 10.1093/jhered/esu064 [DOI] [PubMed] [Google Scholar]
- 71.Dulvy NK, Reynolds JD. Biodiversity: Skates on thin ice. Nature. 2009;462: 417–417. 10.1038/462417a [DOI] [PubMed] [Google Scholar]
- 72.Iglésias SP, Toulhoat L, Sellos DY. Taxonomic confusion and market mislabelling of threatened skates: Important consequences for their conservation status. Aquat Conserv Mar Freshw Ecosyst. 2010;20: 319–333. [Google Scholar]
- 73.Pasolini P, Ragazzini C, Zaccaro Z, Cariani A, Ferrara G, Gonzalez EG, et al. Quaternary geographical sibling speciation and population structuring in the Eastern Atlantic skates (suborder Rajoidea) Raja clavata and R. straeleni. Mar Biol. 2011;158: 2173–2186. [Google Scholar]
- 74.Cannas R, Pasolini P, Mancusi C, Follesa MC, Cabiddu S, Hemida F, et al. Distribution, molecular systematics and phylogeography of Raja polystigma and Raja montagui in the Mediterranean. Biol Mar Mediterr. 2008;15: 188–191. [Google Scholar]
- 75.McEachran JD, Seret B, Miyake T. Morphological variation within Raja miraletus and status of R. ocellifera (Chondrichthyes, Rajoidei). Copeia. 1989;3: 629–641. [Google Scholar]
- 76.Cariani A, Carlesi L, Tosarelli I, Seret B, Tinti F. Cryptic speciation and evolutionary history of the Raja miraletus species complex. Communication at 14th EEA Scientific Conference 10-13th November 2010, Galway, Ireland.; 2010.
- 77.Kelly RP, Port JA, Yamahara KM, Crowder LB. Using environmental DNA to census marine fishes in a large mesocosm. PLoS One. 2014;9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kelly RP, Port J a., Yamahara KM, Martone RG, Lowell N, Thomsen PF, et al. Harnessing DNA to improve environmental management. Science (80-). 2014;344: 1455–1456. [DOI] [PubMed] [Google Scholar]
- 79.Thomsen PF, Møller PR, Sigsgaard EE, Knudsen SW, Jørgensen OA, Willerslev E. Environmental DNA from Seawater Samples Correlate with Trawl Catches of Subarctic, Deepwater Fishes. PLoS One. 2016;11: e0165252 10.1371/journal.pone.0165252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ratnasingham S, Hebert PDN. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol Ecol Notes. 2007;7: 355–364. 10.1111/j.1471-8286.2007.01678.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Steinke D, Connell AD, Hebert PDN. Linking adults and immatures of South African marine fishes Genome. NRC Research Press; 2016; [DOI] [PubMed] [Google Scholar]
- 82.Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PDN. DNA barcoding Australia’s fish species. Philos Trans R Soc Lond B Biol Sci. 2005;360: 1847–1857. 10.1098/rstb.2005.1716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Serena F, Baino R, Rey J, Papaconstantinou C, Relini G. Catch composition and abundance of deep-sea elasmobranchs based on the MEDITS trawl surveys. Deep Sea 2003: Conference poster papers and workshop papers Vol 2 FAO. 2005. pp. 395–408.
- 84.Collins R a., Cruickshank RH. The seven deadly sins of DNA barcoding. Mol Ecol Resour. 2013;13: 969–975. 10.1111/1755-0998.12046 [DOI] [PubMed] [Google Scholar]
- 85.Saitou N, Nei M. The Neighbor-joining method: a new method for reconstructing phylogenetic trees. Biol Sci. 1987;4: 406–425. [DOI] [PubMed] [Google Scholar]
- 86.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30: 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution (N Y). 1985;39: 783–791. [DOI] [PubMed] [Google Scholar]
- 88.Ratnasingham S, Hebert PDN. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS One. 2013;8: e66213 10.1371/journal.pone.0066213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Costa FO, Landi M, Martins R, Costa MH, Costa ME, Carneiro M, et al. A ranking system for reference libraries of DNA barcodes: application to marine fish species from Portugal. PLoS One. 2012;7: e35858 10.1371/journal.pone.0035858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Felsenstein J. PHYLIP—Phylogeny Inference Package (Version 3.2). Cladistic. 1989;5: 164–166. [Google Scholar]
- 91.Felsenstein J. PHYLIP (Phylogeny Inference Package) version 3.6. Distributed by the author. Department of Genome Sciences, University of Washington, Seattle; 2005.
- 92.Landi M, Dimech M, Arculeo M, Biondo G, Martins R, Carneiro M, et al. DNA Barcoding for Species Assignment: The Case of Mediterranean Marine Fishes. PLoS One. 2014;9: e106135 10.1371/journal.pone.0106135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Martin AP, Palumbi SR. Body size, metabolic rate, generation time, and the molecular clock. Proc Natl Acad Sci U S A. 1993;90: 4087–4091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Steinke D, Zemlak TS, Boutillier JA, Hebert PDN. DNA barcoding of Pacific Canada’s fishes. Mar Biol. 2009;156: 2641–2647. [Google Scholar]
- 95.Zhang J, Hanner R. Molecular approach to the identification of fish in the South China Sea. PLoS One. 2012;7: e30621 10.1371/journal.pone.0030621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lim KC, Lim P-E, Chong VC, Loh K-H. Molecular and morphological analyses reveal phylogenetic relationships of stingrays focusing on the family Dasyatidae (Myliobatiformes). PLoS One. 2015;10: e0120518 10.1371/journal.pone.0120518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Wienerroither RM, Bjelland O, Bachmann L, Junge C. Northernmost record of the little gulper shark Centrophorus uyato in the north-eastern Atlantic Ocean, with taxonomical notes on Centrophorus zeehaani. J Fish Biol. 2015;86: 834–844. [DOI] [PubMed] [Google Scholar]
- 98.Veríssimo A, Cotton CF, Buch RH, Guallart J, Burgess GH. Species diversity of the deep-water gulper sharks (Squaliformes: Centrophoridae: Centrophorus) in North Atlantic waters—current status and taxonomic issues. Zool J Linn Soc. 2014; n/a-n/a. [Google Scholar]
- 99.White WT, Ebert DA, Naylor GJP, Ho HC, Clerkin P, Veríssimo A, et al. Revision of the genus Centrophorus (Squaliformes: Centrophoridae): Part 1—Redescription of Centrophorus granulosus (Bloch & Schneider), a senior synonym of C. acus Garman and C. niaukang Teng. Zootaxa. 2013;3752: 35–72. [DOI] [PubMed] [Google Scholar]
- 100.Ebert DA, Stehmann MFW. Sharks, Batoids and Chimeras of the North Atlantic. FAO Species Catalogue for Fishery Purposes No. 7. FAO; 2013.
- 101.Ebert DA, Compagno LJ V. Biodiversity and systematics of skates (Chondrichthyes: Rajiformes: Rajoidei). Environ Biol Fishes. 2007;80: 111–124. [Google Scholar]
- 102.Stehmann MFW, Bürkel DL. Rajidae Fishes of the North-Eastern Atlantic and the Mediterranean. UNESCO, Paris; 1984. pp. 163–196. [Google Scholar]
- 103.Compagno LJV. Systematics and body form. Sharks, Skates and Rays Johns Hopkins University Press, Baltimore; 1999. pp. 1–42. [Google Scholar]
- 104.Hamlett WC. Sharks, skates, and rays The biology of elasmobranch fishes. Baltimore, Johns Hopkins University Press; 1999. [Google Scholar]
- 105.McEachran JD, Miyake T. Zoogeography and bathymetry of skates (Chondrichthyes, Rajoidei). Elasmobranchs and Living Resources: Advances as the Biology, Ecology, Systematics and the Status of the Fisheries. NOAA Tech Rept 90, pp 285–304 (eds H.L.Pratt, S.H.Gruber and T.Taniuchi). Technical report National Marine Fisheries Service, no. 90, National Oceanic & Atmospheric Administration, Washington, D. C., 305–326 pp; 1990.
- 106.Chevolot M, Hoarau G, Rijnsdorp AD, Stam WT, Olsen JL. Phylogeography and population structure of thornback rays (Raja clavata L., Rajidae). Mol Ecol. 2006;15: 3693–3705. 10.1111/j.1365-294X.2006.03043.x [DOI] [PubMed] [Google Scholar]
- 107.Griffiths AM, Sims DW, Johnson A, Lynghammar A, McHugh M, Bakken T, et al. Levels of connectivity between longnose skate (Dipturus oxyrinchus) in the Mediterranean Sea and the north-eastern Atlantic Ocean. Conserv Genet. 2011;12: 577–582. [Google Scholar]
- 108.Dulvy NK, Metcalfe JD, Glanville J, Pawson MG, Reynolds JD. Fishery stability, local extinctions, and shifts in community structure in skates. Conserv Biol. 2000;14: 283–293. [Google Scholar]
- 109.FDA—Food & Drug Administration [Internet]. 2015. Available: http://www.fda.gov/Food/FoodScienceResearch
- 110.Valentini A, Taberlet P, Miaud C, Civade R, Herder J, Thomsen PF, et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol Ecol. 2016;25: 929–942. 10.1111/mec.13428 [DOI] [PubMed] [Google Scholar]
- 111.Simpfendorfer CA, Kyne PM, Noble TH, Goldsbury J, Basiita RK, Lindsay R, et al. Environmental DNA detects Critically Endangered largetooth sawfish in the wild. Endanger Species Res. 2016;30: 109–116. [Google Scholar]
- 112.Mahon AR, Jerde CL. Using Environmental DNA for Invasive Species Surveillance and Monitoring In: Bourlat SJ, editor. Marine Genomics: Methods and Protocols. New York, NY: Springer New York; 2016. pp. 131–142. [DOI] [PubMed] [Google Scholar]
- 113.Ficetola GFGF, Miaud C, Pompanon F, Taberlet P. Species detection using environmental DNA from water samples. Biol Lett. 2008;4: 423–425. 10.1098/rsbl.2008.0118 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Both the number of barcode records obtained from Mediterranean and non-Mediterranean specimens are reported. When the number of non-Mediterranean barcodes on BOLD was larger than 10, the exact number is not reported. Species author, Ecology (Bathydemersal, Demersal, Pelagic and Benthopelagic), IUCN Red List Category and Criteria (CR = Critically Endangered, EN = Endangered, VU = Vulnerable, NT = Near Threatened, LC = Least Concern, DD = Data Deficient, NE = Not Evaluated), Status and BIN available on BOLD are detailed for each species. Asterisks denote barcode sequences deposited in public repositories but not in BOLD.
(XLSX)
The number of collected and barcoded individuals are reported for each chondrichthyan species and each FAO division as well as in total.
(XLSX)
(XLSX)
For each species, the number of sequences used and the length of the alignment are reported.
(XLSX)
Each circle represents one haplotype and its size is proportional to frequency. Colours indicate the origin of samples according to FAO fishing divisions.
(TIF)
Data Availability Statement
Sampling and biological data as well as digital images of dorsal/ventral sides (when recorded) are available in the “ELASMOMED Consortium” project (Project Code: ELAMO) accessible on the Barcode of Life Data system (BOLD, http://www.barcodinglife.org). GenBank accession numbers for all barcoded specimens are reported in S3 Table.