

Abstract
Background
Motivated by the first mathematical model for schistosomiasis proposed by Macdonald and Barbour’s classical schistosomiasis model tracking the dynamics of infected human population and infected snail hosts in a community, in our previous study, we incorporated seasonal fluctuations into Barbour’s model, but ignored the effect of bovine reservoir host in the transmission of schistosomiasis. Inspired by the findings from our previous work, the model was further improved by integrating two definitive hosts (human and bovine) and seasonal fluctuations, so as to understand the transmission dynamics of schistosomiasis japonica and evaluate the ongoing control measures in Liaonan village, Xingzi County, Jiangxi Province.
Methods
The basic reproductive ratio R 0 and its computation formulae were derived by using the operator theory in functional analysis and the monodromy matrix theory. The mathematical methods for global dynamics of periodic systems were used in order to show that R 0 serves as a threshold value that determines whether there was disease outbreak or not. The parameter fitting and the ratio calculation were performed with surveillance data obtained from the village of Liaonan using numerical simulation. Sensitivity analysis was carried out in order to understand the impact of R 0 on seasonal fluctuations and snail host control. The modified basic reproductive ratios were compared with known results to illustrate the infection risk.
Results
The Barbour’s two-host model with seasonal fluctuations was proposed. The implicit expression of R 0 for the model was given by the spectral radius of next infection operator. The R 0 s for the model ranged between 1.030 and 1.097 from 2003 to 2010 in the village of Liaonan, Xingzi County, China, with 1.097 recorded as the maximum value in 2005 but declined dramatically afterwards. In addition, we proved that the disease goes into extinction when R 0 is less than one and persists when R 0 is greater than one. Comparisons of the different improved models were also made.
Conclusions
Based on the mechanism and characteristics of schistosomiasis transmission, Barbour’s model was improved by considering seasonality. The implicit formula of R 0 for the model and its calculation were given. Theoretical results showed that R 0 gave a sharp threshold that determines whether the disease dies out or not. Simulations concluded that: (i) ignoring seasonality would overestimate the transmission risk of schistosomiasis, and (ii) mollusiciding is an effective control measure to curtail schistosomiasis transmission in Xingzi County when the removal rate of infected snails is small.
Electronic supplementary material
The online version of this article (doi:10.1186/s13071-017-1983-1) contains supplementary material, which is available to authorized users.
Keywords: Schistosomiasis japonica, Mathematical model, Seasonal fluctuation, Basic reproductive ratio, Parameter estimation, Barbour’s two-host model
Background
Schistosomiasis japonica, a parasitic disease caused by Schistosoma japonicum, has been in existence in the People’s Republic of China (P. R. China) for over 2000 years with considerable public-health and economic significance [1, 2]. A large-scale national schistosomiasis control programme was initiated in the mid-1950s [2, 3], when China’s population was approximately 600 million. An estimated 11.8 million people were infected with S. japonicum [4]. Sustained control efforts have contributed significantly to the dramatic reduction of both transmission intensity and schistosomiasis distribution in China in the past six decades [5–9]. Recent data have shown that progress toward schistosomiasis elimination encountered difficulties and setback due to high re-infection rates, particularly in lake region of the endemic areas [10]. For instance, schistosomiasis re-emerged shortly after the termination of The World Bank Loan Project (WBLP) at the end of 2001 [1, 5, 11]. In order to achieve the goal of schistosomiasis elimination in P. R. China, Chinese government strengthened the national schistosomiasis control programme in 2004. This made schistosomiasis control top priority along with the list of other communicable diseases such as HIV/AIDS, tuberculosis, and hepatitis B in China [4, 12]. Moreover, a revised strategy to effectively control schistosomiasis by using integrated measures in the national control programme has been implemented since 2005 [11].
Over 40 different species of wild and domestic animals have been identified as definitive hosts of S. japonicum [13]. Bovines are the major reservoirs for S. japonicum in the lake and marshland regions of southern China [14, 15]. A large number of schistosome-infected bovines are distributed in these regions and they excrete large quantities of S. japonicum eggs, the majority of which are deposited near or in the lake [14]. The daily faecal output from a water buffalo (~25 kg) has been estimated to be at least 100 times more than that (~250 g) from an individual human [16, 17]. It was reported that the overall prevalence of S. japonicum was 9.6 and 7.2% in water buffalo and cattle, respectively, in 1995 [18]. High prevalence of S. japonicum infection in bovine reservoir hosts is believed to be the major factor maintaining active transmission in certain areas. A cluster randomized intervention trial which was designed to compare the control (human treatment) and intervention (human and bovine treatment) in some villages concluded that the incidence of human S. japonicum infection is reduced with a decline in the infection rates of water buffaloes [19]. In order to remove bovines as a source of infection, several effective measures, including replacing cattle with farm machinery, isolating marshland, and prohibiting grazing in susceptible areas, have been implemented.
Mathematical modelling is a powerful tool to study the transmission dynamics of schistosomiasis [20]. It was first proposed by Macdonald [21]. Following this pioneering work, many mathematical models have been developed, all of which show great potential in aiding our understanding of the interplay of biology, transmission dynamics and control of schistosomiasis [22–27]. In 1996, Macdonald’s model was improved by Barbour [28]. It tracks dynamics of both infected human and snails in a community. Barbour’s model has played an important role in evaluating possible control strategies [29]. Traditionally, schistosomiasis models assume that all parameters are constant. A real world environment is obviously non-stationary, and should include seasonal variations in snail population. Infection rates vary seasonally due to natural factors (i.e. changes in moisture and temperature) and social factors (i.e. changes in contact rates). Therefore, it is more realistic to assume that the infection rates are periodic rather than constant. In our previous study [30], we constructed the Barbour’s single-host model with seasonal fluctuations (BSHSF model) and calculated the basic reproductive ratio to assess the effect of integrated control measures against schistosomiasis in Liaonan village, Xingzi County, Jiangxi Province. However, the impact of the bovine reservoir host on the transmission of schistosomiasis should not be ignored.
This study aims to further modify the Barbour’s two-host model with seasonal fluctuations (BTHSF model) and give an implicit expression of the basic reproductive ratio of schistosomiasis and computation method. Also, we will further establish some indexes that predict prevalence variation characteristics of schistosomiasis, and evaluate the prevention and control strategies for schistosomiasis.
Methods
Mathematical formulation
The Barbour’s single-host model with seasonal fluctuations (BSHSF model) in Gao et al. [30] was given by the following equations:
| 1 |
All variables of Eq. (1) are described in Table 1. The infection rate a(t) represents the rate at which a single definitive host becomes infected at unit density of infected snails at time t. The infection rate b(t) is the rate at which snails become infected at time t. The two infection rates can be calculated as follows (see [28]):
Table 1.
Interpretation of the model (1)
| Parameter | Interpretation |
|---|---|
| a(t) | the rate of incidence for a single definitive host at unit density of infected snails at time t |
| b(t) | the rate at which an infected definitive host causes snail infections at time t |
| g | the recovery rate for definitive host infections |
| Δ | the density of snails |
| Σ | the density of definitive hosts |
| μ | per capita removal rate of infected snails |
Abbreviations: P, the prevalence of infection in the definitive host population; y, the proportion of infected snails
Here, α1 denotes the rate at which a host has contact with contaminated water per unit time, α2 denotes the density of cercariae, α3 denotes the probability that an encounter with cercaria leads to the establishment of an adult parasite, β 1 is the rate of egg-laying, β 2 is the probability of an egg developing into a miracidium, β 3 is the probability that a miracidium penetrates a snail, β 4 is the probability that miracidial penetration into an uninfected snail develops into infection.
Seasonality was taken into consideration by assuming that the infection rates a and b in model (1) were time-varying and periodic. a(t), b(t) were taken as the absolute value of sine functions with 365 days period, while the average values of functions a(t) and b(t) per year (365 days) were assumed to be equal to the values of parameters a and b in [31], respectively.
Furthermore, we considered the impacts of bovine reservoir host on the transmission of schistosomiasis. The BSHSF model (1) was modified and used to create the BTHSF model. The dynamics of the model are governed by the following differential equations:
| 2 |
All variables of Eq. (2) are described in Table 2, where a 1(t) denotes the rate of incidence for one human host at unit density infected by one infectious snail per unit time (one day) at time t, a 2(t) denotes the rate of incidence for one bovine host at unit density infected by one infectious snail per unit time (1 day) at time t, b 1(t) is the rate of incidence for one susceptible snail at unit density infected by one infected human per unit time (1 day) at time t, b 2(t) is the rate of incidence for one susceptible snail at unit density infected by one infected bovine per unit time (1 day) at time t. In this study, we assume that b 1 is proportional to b 1, i.e. b 2 = kb 1, where k is the coefficient ratio. According to Zhao et al. [32]:
where A i (i = 1,2) denote faeces output per human and bovine, B i (i = 1,2) denote the epg of human and bovine, respectively, here epg is the number of eggs per gram of faeces. The number of eggs per bovine per day is more than 105 [33], and the number of eggs per human per day is about ~2800–3000 [34]. Furthermore, many effective measures, such as improved sanitation and health education are implemented to control the spread of schistosomiasis among human beings. In this paper, we choose k =50. We also take the infection rates as:
Table 2.
Interpretation of the model (2)
| Parameter | Interpretation |
|---|---|
| a 1(t) | the infection rate from snail to human at time t |
| a 2(t) | the infection rate from snail to bovine at time t |
| b 1(t) | the infection rate from human to snail at time t |
| b 2(t) | the infection rate from bovine to snail at time t |
| g 1 | the recovery rate for human host infections |
| g 2 | the recovery rate for bovine host infections |
| Δ | the density of snails |
| Σ1 | the density of human hosts |
| Σ2 | the density of bovine hosts |
| μ | per capita removal rate of infected snails |
Abbreviations: P 1, prevalence of infection in the human host population; P 2, the prevalence of infection in the bovine host population; y, the proportion of infected snails
Since infection rates a i and b i (i = 1,2) denote the average values of a i(t) and b i(t) per year, respectively, we have
Thus we can derive and
The basic reproductive ratio (R0)
For epidemiological models, the basic reproductive ratio, which is often introduced as a threshold parameter, is defined as the expected number of secondary infections produced by a single infective individual in a completely susceptible population during its entire infectious period [35]. R 0 has often been used to predict the trend of the disease transmission and also to assess the effects of control measures. Here, we give the implicit formula R 0 of model (2). It is expected that the infection will persist if R 0 is greater than one. As R 0 increases, the spreading rate of the disease also increases. This implies that more intervention efforts or improved control measures should be implemented. We have also shown that R 0 is a sharp threshold value which determines whether the disease dies out or not in Additional file 1: Appendix B. That is, if R 0 < 1, then the disease will die out, whereas if R 0 > 1, the disease will be endemic.
With reference to Bacaër [36], a biologically meaningful threshold value R 0 of BTHSF model (2) was derived by using operator theory in functional analysis and the monodromy matrix of linear periodic system theory. The numerical computation of the basic reproductive ratio was carried out using the mathematical programming language MATLAB 7.1. Also, sensitivity analysis was carried out in order to assess the impact of seasonal fluctuations on R 0
Study area and estimation of model parameters
Liaonan village, Xingzi County, Jiangxi Province was selected as the study area. The basic reproductive ratio was calculated based on the parameters in model (2). Determination of accurate model parameters was the priority when this threshold was applied. The details of the technique used in estimating the parameters have been described in our previous study [30]. In this paper, part of the parameters in model (2) were determined according to their biological significance. In addition, the remaining parameters were estimated based on the annual report surveillance data of Xingzi County from 2003 to 2010. The meanings of parameters g i, Σ i(i = 1,2), μ, ω are consistent with those reported in [30] (see Tables 1 and 2).
The parameters are stated as follows; g1 = 0.00093 per day, μ = 0.0055 per day, and g2 = 0.00183 per day (the data are given by Wu [31]). The density of snails, Δ(/m 2), and the density of definitive hosts, Σ1, Σ2(/m2), were estimated from annual report surveillance data (see Table 3).
Table 3.
The data for Δ, Σ1, Σ2, , and from annual report data in the village of Liaonan, Xingzi County, Jiangxi Province, China
| Parameter | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|
| Δ | 27.997 | 33.9230 | 91.4602 | 133.5238 | 41.0615 | 17.4960 | 11.3121 | 2.4183 |
| Σ1 | 0.0300 | 0.0199 | 0.0475 | 0.0167 | 0.0309 | 0.0178 | 0.0309 | 0.0176 |
| Σ2 | 0.0013 | 0.0007 | 0.0020 | 0.0006 | 0.0010 | 0.0006 | 0.0013 | 0.0008 |
| 0.0671 | 0.0533 | 0.0631 | 0.0454 | 0.0454 | 0.0454 | 0.04530 | 0.0457 | |
| 0.0283 | 0.1379 | 0.1756 | 0.1144 | 0.1084 | 0.0676 | 0.0639 | 0.0466 | |
| 0.0002 | 0.00005 | 0.0001 | 0.0001 | 0.0002 | 0.0005 | 0.0008 | 0.0015 |
It is assumed that the coefficients of model (2) are constant. By setting the right-hand side of equations in model (2) to zero, the steady-state (equilibrium) values for the prevalence of infection in humans, bovines and snails were obtained [28]
| 3 |
| 4 |
where t (i)MS = b i∑i/(μΔ) and t (i)SM = a i Δ/g i(i = 1, 2) are the transmission factors for definitive host i. By solving (3) and (4), we obtain the prevalence of infection in humans, bovines and snails from 2003 to 2010.
Results
Calculation of R0
Following the results of Bacaër [36] and Wang & Zhao [37], we show that R 0 is equal to the value of λ, where λ is the positive root of the equation
| 5 |
Here
and ρ(W(365, 0, λ)) is the spectral radius of matrix W(365, 0, λ).
The mathematical details that were used to derive the expression for the basic reproductive ratio can be found in Additional file 1: Appendices A, B (Derivation of R 0 and proof of main results).
Numerical simulation
Based on the annual report data for Xingzi County from 2003 to 2010, the equilibrium values (prevalence of infection in human host, bovine reservoir host and snail) are shown in Table 3. In view of the parameters’ values given above (see Table 3), the infection rates a 1, a 2, b 1 and b 2 of BTH model was derived from Eqs. (3) and (4), and are shown in Table 4.
Table 4.
The calculated values of composite parameters a 1, a 2, b 1 and b 2
| Parameters | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|
| a 1 | 0.0116 | 0.0309 | 0.0051 | 0.0024 | 0.0061 | 0.0050 | 0.0051 | 0.0120 |
| a 2 | 0.0093 | 0.1726 | 0.0320 | 0.0128 | 0.0305 | 0.0151 | 0.0144 | 0.0241 |
| b 1 | 0.0082 | 0.0015 | 0.0032 | 0.0260 | 0.0057 | 0.0171 | 0.0085 | 0.0075 |
| b 2 | 0.4084 | 0.0754 | 0.1606 | 1.2996 | 0.2871 | 0.8565 | 0.4268 | 0.3725 |
In Additional file 1: Appendix A, we illustrate that R 0 is the positive root of the equation ρ(W(365, 0, λ)) = 1. Using the data in Tables 3 and 4, the values of R 0 for BTHSF model were calculated and shown in Table 5. Let R ′0 be the basic reproductive ratio for BSHSF model. It was calculated using established methods (see Table 6).
Table 5.
The result of the basic reproductive ratio from annual report data in the village of Liaonan, Xingzi County, Jiangxi Province, China
| The basic reproductive ratios | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|
| R 0 (BTHSF model) | 1.031 | 1.074 | 1.097 | 1.060 | 1.057 | 1.038 | 1.037 | 1.030 |
| ℜ 0 (BTH model) | 1.052 | 1.142 | 1.192 | 1.114 | 1.107 | 1.066 | 1.064 | 1.050 |
Table 6.
The result of the basic reproductive ratio from annual report data in the village of Liaonan, Xingzi County, Jiangxi Province, China
| The basic reproductive ratios | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 |
|---|---|---|---|---|---|---|---|---|
| R 0 (BTHSF model) | 1.031 | 1.074 | 1.097 | 1.060 | 1.057 | 1.038 | 1.037 | 1.030 |
| R '0 (BSHSF model) | 1.043 | 1.035 | 1.040 | 1.031 | 1.031 | 1.031 | 1.031 | 1.032 |
Barbour [28], proposed the following model without seasonality (BTH model):
and obtained the basic reproductive ratio
Using the data in Tables 3 and 4, we obtain the values of R 0 in the village of Liaonan from 2003 to 2010.
R 0 for BTHSF model is shown in Table 3, and it clearly shows that the values range between 1.030 and 1.097 from 2003 to 2010 in the village of Liaonan. Moreover, R 0 for BTH model is always greater than that of BTHSF model at the same time. Furthermore, the variation tendency of R 0 for BTHSF model is shown in Fig. 1.
Fig. 1.
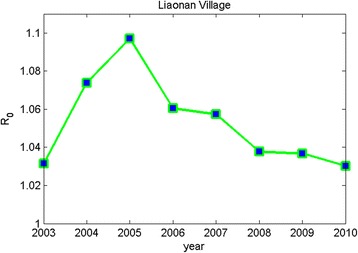
Changes in the basic reproductive ratio in Liaonan village from 2003 to 2010
The prevalence of two hosts and vector host has strong effect on R 0 for BTHSF model. Except in 2003 and 2010, the values of R 0 for BTHSF model are greater than those for BSHSF model in the other years (see Table 4). The prevalence of infected bovines in 2003 and 2010 are less than the other years (see Table 2). This phenomenon illustrates that if the prevalence ratio of infected bovine hosts and infected human hosts is small, R 0 for BTHSF model may be smaller than for BSHSF model. Moreover, we note that R 0 and the prevalence of infection in bovines for BTHSF model are higher in 2004 and 2005 among the years under study as shown in (Tables 1 and 3).
Sensitivity analysis
In model (2), infection rates a 1(t) and b 1(t) (i = 1,2) were taken as sine functions of 365 days period, with the form and . Thus, πa i/2 and πb i/2 reflect the amplitude of infection rates a i(t) and b i(t), respectively. We fix g1 = 0.00093, g2 = 0.00183, Δ = 91.4602, Σ1 = 0.0475, g1 = 0.00093, Σ2 = 0.0020, μ = 0.0055, a 2 = 0.0426, b 2 = 0.1224, and vary a 1 and b 1 in [0,0.04] in (5), with other parameters unchanged as above, by numerical simulations we obtain the curve of R 0 with respect to a 1 and b 1 (see Fig. 2). It shows that the larger a 1 and b 1 are the higher R 0 is. When a 1 and b 1 are near to 0, R 0 is less than 1, which indicates that the control strategy is very effective and disease transmission will be interrupted. This implies that R 0 is very sensitive to the changes in a 1 and b 1 when their values are higher than 0.03. Next, if we fix a 1 = 0.0068, b 1 = 0.0024, and let a 2 and b 2 vary. A graph indicating the relationship between R 0 and a 2 and b 2 was obtained (Fig. 3). This graph shows that R 0 increases with an increase in the amplitude of a 2 and b 2. Finally, as μ varies in [0,0.012] with other parameters unchanged as above, numerical simulations provide the relationship between R 0 and μ (Fig. 4). Figure 4 shows that R 0 is more sensitive to changes in μ when μ is less than 0.002.
Fig. 2.
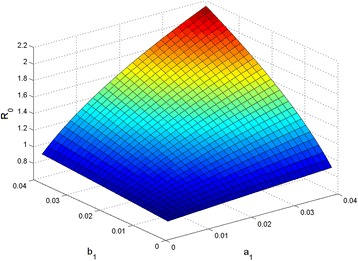
The relationship between a 1, b 1 and R 0. The graph demonstrates the sensitivity of the basic reproductive ratio to the changes of composite parameters a 1 and b 1
Fig. 3.
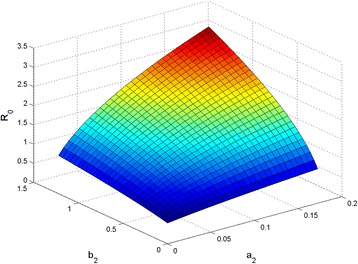
The relationship between a 2, b 2 and R 0. The graph demonstrates the sensitivity of the basic reproductive ratio to the changes of composite parameters a 2 and b 2
Fig. 4.
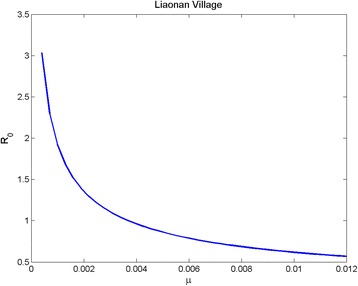
The relationship between μ and R 0. The graph demonstrates the sensitivity of the basic reproductive ratio to the changes of parameter μ
Discussion
This paper proposed Barbour’s two-host model with seasonality (BTHSF model), which is an improvement on the models BTH and BSHSF. Following the idea of Wang & Zhao [37], the implicit formula of R 0 for BTHSF was given. By computation, the annual changing trend of R 0 s in Liaonan village from 2003 to 2010 was given (see Fig. 1). It showed that R 0 peaked in 2005 at 1.09 and declined dramatically until 2010. There are three possible factors responsible for this decline. First, in 2001, the World Bank Loan Project (WBLP) for schistosomiasis control (1992–2001) was terminated. This led to the resurgence of schistosomiasis transmission after 2001 [38, 39]. Secondly, the implementation of integrated control strategies with emphasis on infection source control commenced in 2005. Finally, the high prevalence of S. japonicum in bovines contributes to the high value of R 0.
By numerical simulation, we realize that the basic reproductive number for Barbour’s model with constant infection rates is always higher than that of periodic infection rates. This illustrates that ignoring seasonal fluctuations would lead to overestimating schistosomiasis transmission risk.
Chemotherapy, health education, provision of water, sanitation and hygiene, bovine control and snail host control are the main measures for schistosomiasis control. We know that the implementation of chemotherapy strategy is beneficial to the recovery and well-being of patients and cattle. This would result in increasing the parameters g 1 and g 2 (recovery rates) in model (2). Accordingly, the implementation of control strategies, such as health education, provision of water, sanitation and hygiene, bovine control would lead to decrease in; the host’s contact rate with contaminated water, the probability that an encounter with cercariae results in the development of an adult parasite and the rate at which schistosome eggs are laid. Therefore, the infection rates (a 1(t), a 2(t), b 1(t), b 2(t)) in model (2) would decrease. More so, the implementation of snail control strategy will increase the removal rate of the snail (μ). Due to the implementation of preventive chemotherapy in Xingzi County, we assessed the sensitivity analysis of the average infection rates and removal rate. Numerical results have shown that R 0 is a monotone increasing function of the four infection rates and a monotone decreasing function of the removal rate. This implies that as the infection rates increases or the removal rate decreases, R 0 increases. Our results also indicate that mollusiciding is an effective means of controlling schistosomiasis transmission in Xingzi County when the removal rate of snails is small.
Finally, in line with the theoretical concept of this paper, we introduce several indexes that were explored extensively. Similar to the methods established in [28], R 0 s are estimated from prevalence data and assumed to be at a steady equilibrium state, so they are always more than 1. In reality, R 0 may be less than 1. However, we can study the transmission intensity by using the values of R 0. For example, if we define the relative basic reproductive ratio as:
where denotes the basic reproductive ratio of year k. If R ji > 1, the transmission status of schistosomiasis in year j is more serious than year i and vice versa.
Next, we define the relative mean basic reproductive ratio as
Here, shows that the basic reproductive ratio in year m is greater than the average ratio from year i to j, and vice versa. Taking Liaonan village as the example, the relative basic reproductive ratio in 2005 and 2006 is defined as:
This illustrates that the epidemic trend in 2005 is relatively strong, compared to 2006. In addition,
and
This indicates that the epidemic trends in 2010 are slower than the average level from 2003 to 2010.
Similarly, it is also able to measure the disease transmission situation between two different regions. Let denote the basic reproductive ratios of two different regions in year i, respectively. The index of the basic reproductive ratio is defined as:
γ i can be used to compare the effect of control and intervention between two regions at the same time. However, many factors such as heterogeneity of definitive host, drug resistance and optimal control method are yet to be considered in this mathematical model. These issues need to be addressed in the future studies.
Conclusions
We modified and improved Barbour’s model to investigate the role of seasonal fluctuations in the spread of schistosomiasis. The formula for the basic reproductive number R 0 with a single host was generalized with that of two hosts. Theoretical results showed that it is the basic reproductive ratio which is the threshold value that determines whether schistosomiasis should persist or not. The values of R 0 in Liaonan village from 2003 to 2010 were calculated using the mathematical programming language MATLAB 7.1. The results from this study have shown that ignoring seasonality would overestimate the transmission risk of schistosomiasis, and mollusiciding would be the most effective control measure to curtail schistosomiasis transmission in Xingzi County. Also, the indexes established in this paper will lead to the re-evaluation of control strategies between different areas.
Acknowledgments
This study was supported by Dr Bo Tao and his team from the Schistosomiasis Control Station of Xinzhi County, Jiangxi province, People’s Republic of China, on the field work and data collection. Thank also to Dr. Eniola M. Abe for his assistance on language corrections.
Funding
This project was supported by the Natural Science Foundation of China (grant no. 11561004) and the National Natural Science Foundation of China (grant no. 11261004) through SJG, and by the National Natural Science Foundation of China (grant no. 81573261) through GJY, as well as by the National Science and Technology Major Program (grant no. 2012ZX10004 220) and Shanghai Science and Technology Committee (grant no. 11XD1405400) through XNZ.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and its additional file.
Authors’ contributions
SJG and XNZ conceived and designed the study; YJL contributed with numerical simulation and performed the drawing; all authors contributed to the analysis of the data and to writing the manuscript, and gave final approval for publication. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
This study was approved by the Institutional Ethics Review Committee (ERC) of the National Institute of Parasitic Diseases, China CDC. All the data are from previous databases or yearbooks, and did not include individual data. Therefore, the informed consent was not necessary in this study.
Abbreviations
- BSH model
Barbour’s single-host model
- BSHSF model
Barbour’s single-host model with seasonal fluctuations
- BTH model
Barbour’s two-host model
- BTHSF model
Barbour’s two-host model with seasonal fluctuations
- epg
Eggs per gram
- ERC
Institutional Ethics Review Committee
- R0
The basic reproductive ratio
- WBLP
The World Bank Loan Project
Additional file
Appendix A. Derivarion of R0. Appendix B. Proof of the main results. (PDF 67 kb)
Contributor Information
Shu-Jing Gao, Email: gaosjmath@126.com.
Hua-Hua Cao, Email: 870577346@qq.com.
Yu-Ying He, Email: heyuyinghyy@126.com.
Xiang-Yu Zhang, Email: xyzhang5@163.com.
Guo-Jing Yang, Email: guojingyang@hotmail.com.
Xiao-Nong Zhou, Email: zhouxn1@chinacdc.cn.
References
- 1.Zhou XN, Wang LY, Chen MG, Wu XH, Jiang QW, Chen XY, et al. The public health significance and control of schistosomiasis in China - then and now. Acta Trop. 2005;96:97–105. doi: 10.1016/j.actatropica.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 2.Zhang Z, Clark AB, Bivand R, Chen Y, Carpenter T, Peng W, et al. Nonparametric spatial analysis to detect high-risk regions for schistosomiasis in Guichi, China. Trans R Soc Trop Med Hyg. 2009;103:1045–1052. doi: 10.1016/j.trstmh.2008.11.012. [DOI] [PubMed] [Google Scholar]
- 3.Chen X, Wang L, Cai J, Zhou XN, Zheng J, Guo J, et al. Schistosomiasis control in China: the impact of a 10-year World Bank Loan Project (1992–2001) Bull World Health Organ. 2005;83:43–48. [PMC free article] [PubMed] [Google Scholar]
- 4.Report Inter-country meeting on strategies to eliminate schistosomiasis from the Eastern Mediterranean Region. Muscat, Oman. 2007.
- 5.Zhou XN, Guo JG, Wu XH, Jiang QW, Zheng J, Dang H, et al. Epidemiology of schistosomiasis in the People’s Republic of China, 2004. Emerg Infect Dis. 2007;13:1470–1476. doi: 10.3201/eid1310.061423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shrivastava J, Qian BZ, Mcvean G, Webster JP. An insight into the genetic variation of Schistosoma japonicum in mainland China using DNA microsatellite markers. Mol Ecol. 2005;14:839–849. doi: 10.1111/j.1365-294X.2005.02443.x. [DOI] [PubMed] [Google Scholar]
- 7.Engels D, Chitsulo L, Montresor A, Savioli L. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Trop. 2002;82:139–146. doi: 10.1016/S0001-706X(02)00045-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bergquist R, Tanner M. Controlling schistosomiasis in Southeast Asia: a tale of two countries. Adv Parasitol. 2010;72:109–144. doi: 10.1016/S0065-308X(10)72005-4. [DOI] [PubMed] [Google Scholar]
- 9.Zhao QP, Jiang MS, Dong HF, Nie P. Diversification of Schistosoma japonicum in mainland China revealed by mitochondrial DNA. PLoS Neglect Trop Dis. 2012;6:e1503. doi: 10.1371/journal.pntd.0001503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Seto EYW, Remais JV, Carlton EJ, Wang S, Liang S, Brindley PJ, Qiu D, et al. Toward sustainable and comprehensive control of schistosomiasis in China: lessons from Sichuan. PLoS Neglect Trop Dis. 2011;5:e1372. doi: 10.1371/journal.pntd.0001372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li SZ, Luz A, Wang XH, Xu LL, Wang Q, Qian YJ, et al. Schistosomiasis in China: acute infections during 2005–2008. Chinese Med J Eng. 2009;122:1009–1014. [PubMed] [Google Scholar]
- 12.Engels D, Wang LY, Palmer KL. Control of schistosomiasis in China. Acta Trop. 2005;96:67–68. doi: 10.1016/j.actatropica.2005.07.004. [DOI] [PubMed] [Google Scholar]
- 13.Chen MG, Zheng F. Schistosomiasis control in China. Parasitol Int. 1999;48:11–19. doi: 10.1016/S1383-5769(99)00004-5. [DOI] [PubMed] [Google Scholar]
- 14.Guo JG, Li YS, Gray D, Ning A. A drug-based intervention study on the importance of buffaloes for human Schistosoma japonicum infection around Poyang Lake, People’s Republic of China. Am J Trop Med Hyg. 2006;74:335–341. [PubMed] [Google Scholar]
- 15.Yang G, Ross AGP, Lin DD, Williams GM, Chen HG, Li Y, et al. A baseline study on the importance of bovines for human Schistosoma japonicum infection around Poyang Lake, China. Am J Trop Med Hyg. 2001;65:272–278. doi: 10.4269/ajtmh.2001.65.272. [DOI] [PubMed] [Google Scholar]
- 16.Ross AGP, Sleigh AC, Li Y, Davis GM, Williams GW, Jiang Z, et al. Schistosomiasis in the People’s Republic of China: prospects and challenges for the 21st century. Clin Microbiol Rev. 2001;14:270–295. doi: 10.1128/CMR.14.2.270-295.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.He YX, Salafsky B, Ramaswamy K. Host-parasite relationships of Schistosoma japonicum in mammalian hosts. Trends Parasitol. 2001;17:320–324. doi: 10.1016/S1471-4922(01)01904-3. [DOI] [PubMed] [Google Scholar]
- 18.Dakui E. Epidemic status of schistosomiasis in China, a nation-wide sampling survey in 1995. Naning: Nanjing University Press; 1998. [Google Scholar]
- 19.Gray DJ, Williams GM, Li Y, Chen H, Forsyth S, Li RS, et al. A Cluster-randomised intervention trial against Schistosoma japonicum in the Peoples’ Republic of China: Bovine and human transmission. PLoS One. 2009;4:e5900. doi: 10.1371/journal.pone.0005900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Webb GF, Browne CJ. A model of the Ebola epidemics in West Africa incorporating age of infection. J Biol Dynam. 2016;10:18–30. doi: 10.1080/17513758.2015.1090632. [DOI] [PubMed] [Google Scholar]
- 21.MacDonald G. The dynamics of helminth infections, with special reference to schistosomes. Trans R Soc Trop Med Hyg. 1965;59:489–506. doi: 10.1016/0035-9203(65)90152-5. [DOI] [PubMed] [Google Scholar]
- 22.Williams GM, Sleigh AC, Li Y, Feng Z, Davis GM, Chen H, et al. Mathematical modelling of schistosomiasis japonica: comparison of control strategies in the People’s Republic of China. Acta Trop. 2002;82:253–262. doi: 10.1016/S0001-706X(02)00017-7. [DOI] [PubMed] [Google Scholar]
- 23.Feng Z, Li CC, Milner FA. Schistosomiasis models with two migrating human groups. Math Comput Model. 2005;41:1213–1230. doi: 10.1016/j.mcm.2004.10.023. [DOI] [Google Scholar]
- 24.Chen ZM, Zou L, Shen DW, Zhang WN, Ruan SG. Mathematical modelling and control of schistosomiasis in Hubei Province, China. Acta Trop. 2010;115:119–125. doi: 10.1016/j.actatropica.2010.02.012. [DOI] [PubMed] [Google Scholar]
- 25.Zhang P, Feng Z, Milner F. A schistosomiasis model with an age-structure in human hosts and its application to treatment strategies. Math Biosci. 2007;205:83–107. doi: 10.1016/j.mbs.2006.06.006. [DOI] [PubMed] [Google Scholar]
- 26.Das P, Mukheriee D, Sarkar AK. A study of schistosome transmission dynamics and its control. J Biol Syst. 2006;14:295–302. doi: 10.1142/S0218339006001799. [DOI] [Google Scholar]
- 27.Gray DJ, Williams GM, Li YS, McManus DP. Transmission dynamics of Schistosoma japonicum in the lakes and marshlands of China. PLoS One. 2008;3:e4058. doi: 10.1371/journal.pone.0004058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Barbour AD. Modeling the transmission of schistosomiasis: an introductory view. Am J Trop Med Hyg. 1996;55(Suppl):135–143. doi: 10.4269/ajtmh.1996.55.135. [DOI] [PubMed] [Google Scholar]
- 29.Yang Y, Xiao D. A mathematical model with delays for schistosomiasis japonicum transmission. Chinese Ann Math. 2010;31B:433–446. doi: 10.1007/s11401-010-0596-1. [DOI] [Google Scholar]
- 30.Gao SJ, He YY, Liu YJ, Yang GJ, Zhou XN. Field transmission intensity of Schistosoma japonicum measured by basic reproduction ratio from modified Barbour’s model. Parasit Vectors. 2013;6:141. doi: 10.1186/1756-3305-6-141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wu KC. Mathematical model and transmission dynamics of schistosomiasis and its application. Chinese Trop Med. 2005;5:837–844. [Google Scholar]
- 32.Zhao J, Ouyang T, Ding G. Numerical evaluation of new control method for schistosomiasis. Chinese J Biomed Eng. 2011;28:725–731. [PubMed] [Google Scholar]
- 33.Chen XQ, Xu MG. Analysis of the infection in cattle schistosomiasis in Hubei. Hubei J Anim Vet Sci. 1993;1:23–24. [Google Scholar]
- 34.Mao SB. Schistosome biology and control of schistosomiasis. Being: People’s Health Publication House; 1990. [Google Scholar]
- 35.Bacaër N, Guernaoui S. The epidemic threshold of vector-borne diseases with seasonality. J Math Biol. 2006;53:421–436. doi: 10.1007/s00285-006-0015-0. [DOI] [PubMed] [Google Scholar]
- 36.Bacaër N. Approximation of the basic reproduction number R0 for vector-borne diseases with a periodic vector population. Bull Math Biol. 2007;69:1067–1091. doi: 10.1007/s11538-006-9166-9. [DOI] [PubMed] [Google Scholar]
- 37.Wang WD, Zhao XQ. Threshold dynamics for compartmental epidemic models in periodic environments. J Dyn Differ Equ. 2008;20:699–717. doi: 10.1007/s10884-008-9111-8. [DOI] [Google Scholar]
- 38.Zhou XN, Bergquist R, Leonardo L, Yang GJ, Yang K, Sudomo M, Olveda R. Schistosomiasis japonica control and research needs. Adv Parasitol. 2010;72:145–78. [DOI] [PubMed]
- 39.Liang S, Yang C, Zhong B, Qiu D. Re-emerging schistosomiasis in hilly and mountainous areas of Sichuan, China. Bull World Health Organ. 2006;84:139–144. doi: 10.2471/BLT.05.025031. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article and its additional file.