

Abstract
The live oral typhoid vaccine Ty21a elicits predominantly CD8+, as well as CD4+ T cells mediated immune responses. Clinical field studies showed that Ty21a is moderately effective against S. Typhi and S. Paratyphi B, but not S. Paratyphi A infections. In this study we describe the in depth characterization of S. Typhi, S. Paratyphi A and S. Paratyphi B cross-reactive CD4+ T cell responses elicited following immunization with Ty21a. PBMC samples were collected from 16 healthy volunteers before and 42/84 days after Ty21a immunization and stimulated ex-vivo with Salmonella-infected targets. Multiparametric flow cytometry was used to detect the vaccine elicited Salmonella-specific responses in T effector/memory (TEM) and CD45RA+ T effector/memory (TEMRA) CD4+ cell subsets, by measuring CD4+ multifunctional (MF) cells that concomitantly produced IFN-γ, TNF-α, IL-2, MIP-1β, IL-17A and/or expressed CD107a. Post-vaccination increases in S. Typhi-specific MF cells were observed in CD4+ TEM and TEMRA subsets which predominantly produced IFN-γ and/or TNF-α, while IL-2 was produced by a smaller cell subset. A small proportion of those MF cells also produced MIP-1β, IL-17A and expressed CD107a (a marker associated with cytotoxicity). Approximately one third of these specific MF cells have the potential to migrate to the gut mucosa, as evidenced by co-expression of the gut-homing molecule integrin α4β7. In contrast to our previous observations with CD8+T cells, MF CD4+ T cell responses to the different Salmonella serovars evaluated were similar in magnitude and characteristics. We conclude that although induction of cross-reactive CD4+ MF effector T cells suggest a possible role in Salmonella-immunity, these responses are unlikely to provide an immunological basis for the observed efficacy of Ty21a against S. Typhi and S. Paratyphi B, but not to S. Paratyphi A.
Keywords: Ty21a, Cross-reactive, CD4 T cells, Multifunctional, Salmonella, Human
1. Introduction
Enteric fevers are caused by infection with the human-host restricted organisms Salmonella enterica serovar Typhi (S. Typhi), S. Paratyphi A, S. Paratyphi B and more rarely, S. Paratyphi C [1]. Typhoid fever, caused by S. Typhi, has long been recognized as a major health problem and two moderately effective vaccines, i.e., live attenuated oral vaccine Ty21a (Ty21a) and parenteral polysaccharide Vi (Vi) vaccines have been used extensively in the US, as well as many other countries [2]. Recently, the incidence of paratyphoid A fever has been on the rise in South, Southeast and East Asia, as well as among US and European travelers returning from those areas [3–5]. However, in contrast to typhoid fever, no vaccine is available to prevent paratyphoid A (or B) fever.
Salmonella serovars causing enteric fever show a high degree of homology at the DNA level. However Salmonella virulence factor Vi polysaccharide, which has been purified and used as a Vi vaccine, is expressed by S. Typhi, but not by S. Paratyphi A or S. Paratyphi B [6]. Several field studies performed to investigate whether immunization with live oral vaccine Ty21a could protect against typhoid fever also offered an opportunity to assess cross-protection against S. Paratyphi B or S. Paratyphi A infections because those strains were also prevalent causes of enteric fever in the field trial sites [7, 8]. The Santiago, Chile study indicated that Ty21a conferred a moderate degree of cross-protection against S. Paratyphi B disease [9], while the Plaju, Indonesia trial suggested that Ty21a provided little protection against S. Paratyphi A disease. Thus, developing an effective vaccine against S. Paratyphi A has emerged as a public health priority [9, 10].
Successful development of a vaccine against S. Paratyphi A will be aided by a better understanding of the complex human host-immune responses mediating protection against Salmonella. Unfortunately, this endeavor is challenging due to the lack of a suitable animal model that can reliably mimic enteric fevers. The widely used mouse model with S. Typhimurium infection contributed significant insights into the Salmonella specific host innate and adaptive immune responses. However, less is known regarding the protective mechanism(s) against Salmonella infection in humans, which appear to involve both humoral and complex CMI responses [11–14]. Most of the available information regarding S.
Typhi immunity in humans was derived from clinical studies involving the immunization of healthy adults with various attenuated Salmonella vaccine strains and also from the limited body of work with natural infection and a handful of human challenge studies with wild-type S. Typhi [15–18]. Over the past two decades we and others have extensively studied S. Typhi specific humoral as well as cell-mediated immune (CMI) responses elicited in humans following the administration of live attenuated oral live typhoid vaccines [19–30]. Using advanced multi-parametric flow cytometric methods we demonstrated that immunization of healthy volunteers with attenuated live oral S. Typhi vaccines elicit both CD4+ and CD8+ T cell responses, including cytotoxic T cells (CTL) [21, 24–26, 28–31]. Furthermore, we revealed that these vaccine induced S. Typhi-specific CD8+ as well as CD4+ T cells, were mostly mediated by T effector/memory (TEM; CD45RA−CD62L) and CD45RA+TEM (TEMRA; CD45RA+CD62L−) subsets of T memory (TM) cells [21–26, 28–30]. Specific responses were also observed, albeit of lower magnitude, in T central/memory (TCM; CD45RA−CD62L+) cells. A significant portion of these S. Typhi-specific T cells also expressed the gut homing molecule integrin α4β7, suggesting their potential to migrate to the primary site of infection [24, 29, 30, 32].
The recent urgency in developing an effective vaccine against S. Paratyphi A and the observation in the field that Ty21a partially cross-protects against S. Paratyphi B but not S. Paratyphi A prompted us to evaluate cross-reactive immune responses elicited by Ty21a against the Salmonella serovars S. Paratyphi A and S. Paratyphi B) [7, 9]. We and others have studied Ty21a-induced cross-reactive humoral and CD8+ T cell mediated CMI responses against S. Typhi, S. Paratyphi A and B [30, 33–35]. We recently described that immunization with Ty21a elicited CD8+ T mediated multifunctional (MF) cross-reactive Salmonella specific responses against all three Salmonella strains and that S. Typhi specific responses were similar to those observed against S. Paratyphi B but not S. Paratyphi A [30]. In the present study we markedly extend these observations by describing, for the first time, that Ty21a also elicits CD4+ T cells that cross-react with S. Typhi, S. Paratyphi A and S. Paratyphi B and that these responses are mostly mediated by MF cells, capable of simultaneously producing more than one cytokine (i.e., IFN-γ, TNF-α, IL-2, MIP-1β, IL-17A) and/or expressing CD107a.
2. Materials and methods
2.1 Subjects, immunization and isolation of peripheral blood mononuclear cells (PBMC)
Sixteen healthy adults (median age 42 years, range 23 to 52 years) were immunized with the recommended four spaced doses of Ty21a vaccine (Vivotif enteric-coated capsules [Crucell] [29]. Blood samples were drawn pre-vaccination (day 0) and 42 (day 42) and/or 84 (day 84) days post-vaccination. PBMC were isolated immediately after blood draws by density gradient centrifugation and were cryopreserved in liquid nitrogen as previously described [31, 36]. The study was approved by University of Maryland Baltimore Institutional Review Board. All the volunteers gave informed consent prior to their recruitment into this study.
2.2 Target/stimulator cell preparation
Autologous Epstein Barr virus (EBV)-transformed B-LCL (EBV-B cells) were generated from PBMC obtained from Ty21a vaccinees as previously described [30]. Salmonella strains, i.e., wild-type S. Typhi strain (ISP-1820, Vi+, a clinical isolate from Chile), S. Paratyphi A (CV 223, ATCC# 9150), and S. Paratyphi B (CV 23, a clinical isolate from Chile) were obtained from the Center for Vaccine Development, University of Maryland, USA (CVD) reference stocks. EBV-B cells were infected with Salmonella strains, at an MOI of 10:1 (bacteria:cell) as previously described and following overnight resting, infected cells were gamma-irradiated (6,000 rad) before being used as “targets” for ex-vivo PBMC stimulation. To confirm the adequacy of the infection with S. Typhi, S. Paratyphi A or S. Paratyphi B, infected EBV-B cells were stained with anti-Salmonella common structural Ag (CSA-1)-FITC (Kierkegaard & Perry, Gaithersburg, MD) and analyzed by flow cytometry using a customized LSR-II instrument (BD, Franklin Lakes, NJ, USA) [30].
2.3 Ex-vivo PBMC stimulation
Thawed, overnight rested PBMC were stimulated with autologous S. Typhi-, S. Paratyphi A- or B- infected targets (section 2.2) at a ratio of 10:1 (PBMC:target). After 2 hours, the protein transport blockers Monensin (1 μg/ml, Sigma) and Brefeldin A (2 μg/ml; Sigma) were added to the PBMC cultures that were continued overnight at 37°C in 5% CO2. Media alone and uninfected autologous EBV-B cells were used as negative controls. Staphylococcal enterotoxin B (SEB) (10 μg/mL; Sigma) was used as a positive control.
2.4 Surface and intracellular staining
Surface and intracellular staining (ICS) was performed as described previously [30]. Briefly, ex-vivo stimulated PBMC were first stained for live/dead discrimination using LIVE/DEAD fixable violet dead cell stain kit (Invitrogen, Carlsbad, CA) and then surface stained with a panel of fluorochrome conjugated monoclonal antibodies (mAbs) that included CD14-Pacific Blue (TuK4, Invitrogen), CD19-Pacific Blue (SJ25-C1, Invitrogen), CD3-Qdot 655 (UCHT1, BD), CD4- PerCP-Cy5.5 (SK3, BD), CD8-Qdot 705 (HIT8A, Invitrogen), CD45RA-biotin (HI100, BD), CD62L-APC-EF780 (Dreg 56, Invitrogen), integrin α4β7-Alexa 488 (clone ACT-1; conjugated in house) and CD107a-A647(eBioH4A3, eBiosciences, San Diego, CA). The anti-CD107a mAb was added during the overnight ex-vivo stimulation to maximize its detection. The cells were then fixed and permeabilized with Fix & Perm cell buffers (Invitrogen) and ICS was performed with a panel of mAbs against IFN-γ-PE-Cy7 (B27, BD), TNF-α-Alexa 700 (MAb11, BD), IL-2-PE (5344.111, BD) and CD69-ECD, (TP1.55.3, Beckman Coulter, CA, USA). A modified panel of mAbs (14 colors) was used in some experiments to concomitantly detect two additional cytokines, i.e., MIP-1β and IL-17 following same surface and ICS staining method. This modified panel included, Live/DEAD fixable yellow dead-cell staining kit (Invitrogen), surface staining with mAbs against CD14-Brilliant violet (BV) 570 (TuK4, Invitrogen), CD19-BV570 (HIB19, Biolegend, San Diego, CA, USA), CD3-BV650 (OKT3, Biolegend), CD4-PE-Cy5 (RPA-T4, BD), CD8-PerCP-Cy5.5 (SK1, BD), CD45RA-biotin (HI100, BD)-secondary staining, with streptavidin Qdot 800 (Invitrogen), CD62L-APC-EF780 (Dreg 56, eBioscience), CD107a-FITC (H4A3, BD) and integrin α4β7-A647(ACT-1; conjugated in house) and ICS staining with mAbs against IFN-γ-PE-Cy7 (B27, BD), TNF-α-Alexa 700 (MAb11, BD), IL-2-BV605 (MQ1-17H12, Biolegend), IL-17A-BV421 (BL168, Biolegend), MIP-1β-PE (24006, R&D, Minneapolis, MN, USA) and CD69-ECD or -PE (TP1.55.3, eBioscience. )
Following staining cells were fixed in 1% paraformaldehyde and stored at 4°C until analyzed. Flow cytometry was performed using a customized LSRII flow cytometer (BD) and data were analyzed using WinList version 7 (Verity Software House, Topsham, ME, USA).
2.5 Gating protocol
CD4+ (live CD3+, CD8-CD4+) T cell responses in study subjects were evaluated in different T memory (TM) subsets defined by their expression of CD45RA and CD62L i.e., T central memory (TCM; CD62L+ CD45RA−), T effector memory (TEM; CD62L−CD45RA−) and T effector memory CD45RA+ (TEMRA; CD62LCD45RA+) and Naïve T cells (TN: CD62L+ CD45RA+) (Fig. S1). The FCOM analysis tool (WinList version 7) was used to classify events based on combinations of gates in multidimensional space, i.e., whether cells express single (single positives; S+) or multiple intracellular cytokines and/or CD107a alone or in all possible combinations) for the detection of multifunctional (MF) cells. Flow cytometric analyses were performed in at least 160,000 events in the live lymphocyte gate (Fig. S1A).
2.6 Statistical analyses
The statistical tests used to analyze each set of experiments are indicated in each Figure Legend. P values of <0.05 were considered significant.
3. Results
In this study, we used PBMC samples collected from volunteers before (day 0) and after (42/84 days) immunization with Ty21a as described in Materials and methods (section 2.1). Routine complete blood counts (CBC) were performed on every blood sample collected. Estimated absolute numbers of lymphocytes in each of the participating volunteers (calculated from the CBC) were not statistically different (p>0.3) when the results of pre-vaccination were compared to those recorded at days 42 or 84 following vaccination [30]
To measure Salmonella-specific responses PBMC were stimulated ex-vivo with S. Typhi-, S. Paratyphi A- and B-infected autologous EBV-B cells as described in Materials and Methods (section 2.3). The “net” Salmonella-specific responses at pre- and post-vaccination days were obtained by subtracting the respective non-specific background responses (i.e., those observed with the corresponding non-infected EBV-B cells). The Ty21a elicited post-vaccination increases (in S+ and each of the MF subsets), were then calculated from the “net” Salmonella-specific responses observed on post-vaccination days 42 or 84 after subtracting the corresponding day 0 levels.
Our results showed that activation of CD4+ T cells (expressing the lymphocyte activation marker CD69) following stimulation with all Salmonella-infected targets, were mostly observed in TEM, albeit lower percentages were also observed in TEMRA, TCM and TN subsets (Fig. S2B). However, activated Salmonella-specific CD4+CD69+ cells that produced cytokines (e.g., IFN-γ; Fig. S2C), TNF-α (Fig. S2D), IL-2 (Fig. S2E) or expressed the degranulation marker CD107a, a marker of cytotoxicity (Fig. S2F) resided almost exclusively in CD62L-TM cells, i.e., in TEM and TEMRA subsets of CD4+T cells. Based on these observations, subsequent analyses were focused in the CD4+ TEM and TEMRA cell subsets.
3.1 Evaluation of Salmonella-specific multifunctional CD4+ T cells
3.1.1 CD4+ TEM
The Salmonella-specific effector cells were characterized into single positives (S+) or those concomitantly producing of two or more cytokines (IFN-γ, TNF-α, IL-2) and/or expressing CD107a as multifunctional (MF) cells, using the FCOM analysis tool. Post-vaccination increases observed in Salmonella-specific IFN-γ+ (Fig. 1A), TNF-α+ (Fig. 1B) or IL-2+ (Fig. 1C) MF cells were significantly higher (p<0.001) compared to the corresponding single cytokine producing CD4+ TEM cells. Significantly increased expression of CD107a+ in MF over that in S+ cells (p<0.05) was observed in response to S. Paratyphi A–infected targets. Similar trends, albeit not significant, were also observed in response to S. Typhi (p=0.07) and S. Paratyphi B (p=0.11)-infected targets (Fig. 1D). The magnitude of these S. Typhi, S. Paratyphi A or S. Paratyphi B-specific MF responses (Fig. 1) showed no significant differences among them.
Figure 1. Induction of multifunctional CD4+ TEM cells following immunization with Ty21a.

PBMC collected from Ty21a vaccinees (n=16) were stimulated ex-vivo with S. Typhi- (ST), S. Paratyphi A- (PA) or S. Paratyphi B (PB)-infected targets. Data were obtained using FCOM analysis in TEM subsets of CD4+ T cells. Shown are the peak post-vaccination increases in Salmonella specific single positive (S+) and multifunctional (MF) IFN-γ+ (A), TNF-α+ (B), IL-2+ (C) and CD107a+ (D) cells. Post-vaccination peaks: peak of the responses at days 42 or 84 minus pre-vaccination (day 0) levels. Horizontal bars represent mean±SEM
TEM, T effector/memory; ***p<0.001 **p<0.01. *p<0.05 compared with the corresponding single-positive cells by Wilcoxon signed rank test, two tailed.
Overall, the MF subsets comprised almost two thirds (58–67%) of the vaccine induced Salmonella-specific CD4 TEM cell responses (Fig. 2). To characterize MF cells in further detail, we first, categorized these Ty21a induced MF cells those into double (2+), triple (3+) or quadruple (4+) positive subsets based on the number of cytokines (IFN-γ, TNF-α, IL-2) and/or CD107a simultaneously produced and/or expressed (Fig. 2). Post-vaccination increases in S. Typhi (Fig. 2A) and S. Paratyphi A (Fig. 2B)-specific MF cells were predominantly comprised of 2+ cells showing a hierarchy of 2+ >3+ >4+, while for S. Paratyphi B (Fig. 2C), we observed that was 2+ = 3+ >4+. We further characterized Salmonella-specific MF cells by investigating all possible subsets producing/expressing different combinations of cytokine production (IFN-γ, TNF-α, IL-2) and CD107a expression. In most of the volunteers only 5 out of all possible MF subsets were found to be “high frequency” subsets showing post-vaccination increases of >0.05% of CD4 TEM cells. Of note, when combined, these 5 subsets typically represented on average (mean±SE) 73.3±5.8%, 74.7±5.5% and 71.2±6.4% of S. Typhi-, S. Paratyphi A- and S. Paratyphi B- specific total MF cells, respectively (Fig. 3). Of interest, the MF cells producing IFN-γ and TNF-α were present in 4 out of these 5 “high frequency” subsets, while IL-2 and CD107a were each observed in 2 MF subsets each (Fig. 3). No statistically significant differences were observed among the percentages of these 5 Salmonella-specific dominant MF subsets of CD4+TEM specific to S. Typhi-, S. Paratyphi A or S. Paratyphi B infected cells (Fig. 3).
Figure 2. Ty21a elicited Salmonella-specific functional CD4+ TEM cells.

Post-vaccination peak increases in functional CD4 TEM (CD45RA−CD62L−) cells obtained from Ty21a immunized volunteers (n=16) that produced cytokines (i.e., IFN-γ, TNF-α, IL-2) or expressed CD107a in response to S. Typhi (A), S. Paratyphi A (B) and S. Paratyphi B (C)–infected targets were measured as described in the text. These functional cells were segregated into Single+ cells (S+; producing only one cytokine or expressing just CD107a [107a]) and multifunctional (MF; simultaneously producing two (2+), three (3+) or four (4+) cytokines and/or expressing CD107a. Bar graphs represent the mean+SE (error bar) of percentages of total functional cell for each of the subsets ( [subset]/[sum of S+ and MF cells] X100). Post-vaccination peak increases were calculated as peak levels at days 42 or 84 post-vaccination minus the corresponding pre-vaccination levels. Statistical analysis:
***: p<0.001, **: p<0.01, *: p<0.05, #: p=0.06 by Wilcoxon paired t-test, 2 tail
Figure 3. Post-vaccination increases in multifunctional (MF) CD4+T EM cells.

S. Typhi (A), S. Paratyphi A (B) and S. Paratyphi B (C) -infected target specific CD4+ TEM MF cells were subdivided into 11 different subsets based on the possible combinations of simultaneous production IFN-γ, TNF-α and/or IL-2- and/or expression of CD107a using the FCOM analysis tool. Shown are the post-vaccination peak increases in the 5/11 dominant subsets and sum of the remaining 6/11 (Others) subsets of specific MF cells. Data are presented as mean percentages of total MF cells for each subset and “Others” in Ty21a vaccinees (n=16).
Post vaccination increase: Peak level at days 42 or 84 post-vaccination minus the corresponding pre-vaccination levels. No statistical significant differences were observed among the various subsets with one way ANOVA (non parametric).
3.1.2 CD4+TEMRA
TEMRA constituted a relatively a small subpopulation of CD4+ T cells than TEM, TCM or TN subsets (Fig. S1). However, as mentioned earlier (Section 3) the induction of “functional cells” following stimulation with Salmonella-infected targets were mostly observed in TEM and TEMRA subsets (Fig. S2). Therefore, we performed a similar analysis to characterize the Ty21a-induced cross-reactive MF responses in CD4+ TEMRA subsets following the strategy described for TEM cells (Fig. 1–3). The magnitude of post-vaccination increases observed in Salmonella-specific IFN-γ+, TNF-α+ and IL-2+ CD4+ TEMRA MF cells, were significantly higher than the corresponding S+ cells (Fig. 4A, B, C). Significant increases in MF TEMRA CD4+ CD107a+ cells were also observed following stimulation with S. Typhi-infected targets (Fig. 4A).
Figure 4. Post-vaccination increases in multifunctional (MF) CD4+T EMRA cells.

PBMC collected from Ty21a vaccinees (n=16) were stimulated ex-vivo with Salmonella-infected targets. Peak post-vaccination increases in single (S) and multifunctional (MF) IFN-γ+, CD107a+, TNF-α+ and IL2+ cells specific for S. Typhi (A)-, S. Paratyphi A (B)- and S. Paratyphi B (C)-infected targets were measured in CD4+T EMRA subsets. Salmonella-infected targets specific CD4+ TEMRA MF cells were further subdivided into 11 different subsets based on the possible combinations of simultaneous production IFN-γ, TNF-α and/or IL-2- and/or expression of CD107a using the FCOM analysis tool. Shown are the post-vaccination peak increases in the 5/11 dominant subsets and sum of the remaining 6/11 (Others) subsets of specific MF cells. Data are presented as mean percentages of total MF cells for each subset and “Others” in Ty21a vaccinees (n=16).
T EMRA, CD45RA+ T effector/memory; Post vaccination increase: Peak level at days 42 or 84 post-vaccination minus the corresponding pre-vaccination levels
***p<0.001 **p<0.01. *p<0.05 compared with corresponding single-positive cells by Wilcoxon signed rank test, two tailed. (A–C).
No statistical significant differences were observed among the various subsets with nonparametric one-way ANOVA (D–F).
Ty21a immunization elicited Salmonella-specific MF cells in CD4+TEMRA subsets were mostly 2+ (~53%) as well as 3+ (~36%) and 4+ (~19%) cells. These proportions of MF CD+TEMRA subsets did not show any statistical differences when stimulated with S. Typhi-, S. Paratyphi A- or S. Paratyphi B-infected targets. Similar to our observations with CD4+ TEM cells (Fig. 3), Ty21a elicited increases in Salmonella specific cells were mostly observed in 5 subsets of CD4+ TEMRA subsets, together constituting almost two-thirds (60~63%) of total CD4 TEMRA Salmonella-specific MF cells (Fig. 4D–F). A similar representation of the 5 MF subsets (as percentage of total MF cells) was observed for TEMRA specific for S. Typhi- (Fig. 4D), S. Paratyphi A- (Fig. 4E) or S. Paratyphi B- (Fig. 4F) infected targets as that observed in the TEM subset.
3.2 Cross-reactive Salmonella-specific MIP-1β and IL-17A responses
We have previously reported that Ty21a elicited the appearance of Salmonella-specific MIP-1β+ and albeit very low magnitude also IL17A+ CD8+ T cells [30]. Therefore, it was of great interest to investigate whether MIP-1β and IL17A producing MF CD4+ T cells are also induced following stimulation of PBMC obtained from Ty21a vaccinees (n=8) with Salmonella-infected targets. To this end, we used an optimized 14-color flow cytometry panel (described in section 2.4.) that included mAbs against MIP-1β and IL-17A as well as those described above, i.e., IFN-γ, CD107a, TNF-α and IL-2. Similar responses were observed following immunization with Ty21a in the capacity of CD4+ TEM and TEMRA subsets to produce MIP-1β in response S. Typhi-, S. Paratyphi A-, and S. Paratyphi B-infected targets. Post-vaccination increases observed with all three Salmonella-specific MF MIP-1β+ cells were higher than the corresponding S+ cells (Fig. 5A). However, these differences following stimulation with S. Typhi- and S. Paratyphi B-infected targets reached statistical significance (p=<0.05), whilst only a trend (p=0.12) was observed for S. Paratyphi A-infected targets (Fig. 5A). In contrast, in CD4+TEMRA subsets, all three Salmonella- specific MF MIP-1β+ cells were elicited at significantly (p<0.05) higher magnitudes than the corresponding S+ cells(Fig. 5B).
Figure 5. Post-vaccination increases in MIP-1β producing CD4+ cells.

Post-vaccination peak increases in Single (S+) and multifunctional (MF) MIP-1β producing CD4+ T EM (A) and T EMRA (B) cells in Ty21a vaccinees (n=8) following stimulation with S. Typhi (ST)-, S. Paratyphi A (PA)- and S.
Paratyphi B (PB)-infected targets.
Error bars indicate mean±SEM.
*p<0.05, MF compared to the corresponding single positive cells. Wilcoxon signed rank test, 2-tail.
Interleukin-17A-producing CD4+ T cells are the core component of the Th17 helper subset which is thought to mediate protection against several organisms, including intracellular bacteria [12, 13, 37]. Therefore, we next investigated whether immunization with Ty21a induces S. Typhi-specific IL-17A producing cells, as well as those that cross-react with S. Paratyphi A or S. Paratyphi B. Post-vaccination increases (>0.05%) in S. Typhi-specific total IL-17A producing CD4+ TEM cells were observed in 75% (6 out of 8) of the Ty21a vaccinated volunteers, while 38% (3 out of 8) and 63% (5 out of 8) showed similar increases against S. Paratyphi A- and S. Paratyphi B-infected targets, respectively (Fig. S3A). Although, the magnitude of post-vaccination increases with S. Typhi-specific total IL-17A+ CD4+ TEM cells were somewhat higher than those specific for S. Paratyphi A- or S. Paratyphi B-infected targets, these differences did not reach statistical significance (Fig. 6A). We also characterized IL-17A+ cells, by dividing those into S+ and MF cells, as described for other cytokines. Our results showed that the post-vaccination increases in S. Typhi-and S. Paratyphi B-specific Salmonella specific IL-17A producing CD4+ TEM MF cells showed a strong statistical trend to be higher (p=0.06) than the increases observed in corresponding IL-17A S+ subset (Fig. 6B, and Fig. S3). Unfortunately, due to the low number of events, it was not possible to further characterize IL-17A+ MF cell subsets as described for other cytokines (Figs. 2 and 3). Interleukin-17 secreting cells were also observed in the TEMRA subset, although the responses were lower in magnitude and fewer cells were IL-17A+ than those observed in TEM cells.
Figure 6. Post-vaccination increases in IL-17A producing CD4+ T EM cells.

Total IL-17A producing CD4+T T EM cells responses (A) were measured in Ty21a vaccinees (n=8) following stimulation with S. Typhi (ST)-, S. Paratyphi A (PA)- and S. Paratyphi B (PB)-infected targets. Post-vaccination increases in Salmonella-specific-IL17A+ producing CD4+ T EM cells were further categorized into Single positive (S+) and multifunctional (MF) cells (sum of all subsets except S+) (B). Error bars represents Mean+SEM.
The peak-post vaccination increases (days 42 or 84) were calculated by subtracting the corresponding pre-vaccination (day 0) levels.
p values shown were determined by Wilcoxon signed rank test, 2-tail.
3.3 Characterization of the gut-homing potential of Salmonella–specific MF CD4+ TEM cells
T cells that have the potential to migrate to the gut mucosa can be measured by evaluating the expression of integrin α4β7 [38]. Because Ty21a immunization elicited a predominantly MF Salmonella-specific CD4+ TEM cell response (Fig. 1), we further investigated the expression of integrin α4β7 by Salmonella-specific MF cells in PBMC obtained from Ty21a-immunized volunteers (n=12). Although post vaccination increases in Salmonella-specific MF cells consisted mostly of integrin α4β7 negative cells (Fig. 7), a considerable proportion of those S. Typhi-, S. Paratyphi A-, and S. Paratyphi B–specific cells (mean±SE: 29.7±6.7, 28.6±6.3 and 29.2±7.5%, respectively) also expressed integrin α4β7. Similar proportions of integrin α4β7+ cells were also observed in Salmonella-specific CD4+ TEMRA MF cells.
Figure 7. Concomitant expression of the gut homing molecule integrin α4β7 by MF cells.
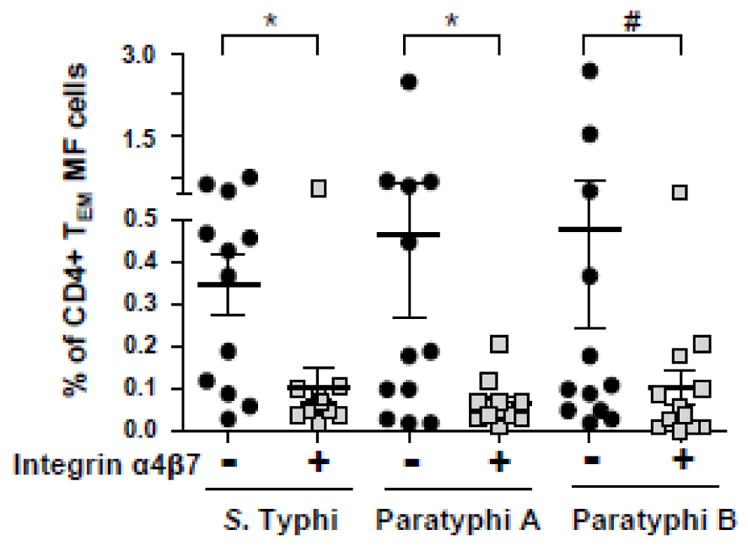
PBMC collected from Ty21a vaccinees (n=12) were stimulated with S. Typhi-, S. Paratyphi A-, and S. Paratyphi B-infected targets and the sum of all CD4+ T EM MF subsets that co-expressed integrin α4β7 (α4β7+) or not (α4β7-) were calculated using the FCOM analysis tool.
Horizontal bars represents mean±SEM of the post-vaccination increases calculated as peak responses at days 42 or 84 minus pre-vaccination [day 0] levels.
*p<0.05, # p=0.06, Wilcoxon signed rank test, 2-tail.
Due to limited numbers of events, it was not possible to further subclassify integrin α4β7+ MF cells into different subsets producing 2–5 cytokines or expressing CD107a.
4. Discussion
Typhoid and paratyphoid fevers due to the distinct Salmonella serovars S. Typhi and S. Paratyphi A and B, respectively, cause indistinguishable clinical syndromes [1]. Ty21a, the only FDA licensed live oral attenuated S. Typhi vaccine elicits both serological and CMI responses mediated by B and T effector/memory cells in humans. An effective CMI recall response appears to be essential in mediating the long-term protective efficacy of the Ty21a vaccine [11, 14, 39]. Evidence from the field indicates that Ty21a also confers moderate protection against S. Paratyphi B but apparently not against S. Paratyphi A [7]. To understand the possible immunological basis for these observations from field studies, in the past few years we have systemically studied the cross-reactivity between S. Typhi, S. Paratyphi A and S. Paratyphi B responses following Ty21a immunization. We have recently reported our results regarding humoral (the induction and functionality of the antibody responses) as well as MF CD8+ T cell-mediated CMI responses [30, 33, 34]. In this study, we confirmed and extended our previous observations with oral typhoid vaccine strain-induced S. Typhi specific CD4+ T cell mediated CMI responses [26, 29] and contributed new insights regarding Ty21a-induced cross-reactive CD4+ T cell responses against S. Paratyphi A and S. Paratyphi B..
Studies in the mouse model of infection with virulent S. Typhimurium evidenced a rapid and efficient induction of effector CD4+ T cell mediated T helper (Th)1 responses, e.g., production of IFN-γ, TNF-α and/or IL-2, which were associated with clearance or resistance to Salmonella infection in susceptible and immunized mice, respectively [13, 40–42].
In humans, immunization with Ty21a, besides S. Typhi-specific strong responses mediated by CD8+ T cells, also elicits specific CD4+ T cell responses (e.g., IFN-γ production). These CD4+ T cell responses were more pronounced to purified antigens (i.e., flagella) than to infected targets [11, 26, 29]. In the present study, we demonstrate that Ty21a induced S. Typhi specific CD4+ TEM and TEMRA cells were the dominant multifunctional (MF) cells. However, because TEMRA constitute a significantly smaller proportion of T effector/memory CD4+ T cells in PBMC (compared with TEM or TCM cells), our current observations suggest that CD4+ S. Typhi-specific T cell responses were mostly mediated by MF TEM cells. Of note, following ex-vivo stimulation of PBMC with all three Salmonella-infected targets we also observed activation of the CD4+ TCM as defined by expression of CD69, but very few of those produced cytokines or expressed CD107a. It is possible that these lower levels of functional CD4+ TCM cells are due to activated TCM cells producing cytokine levels below the level of sensitivity of our assays. Alternatively, or in addition, the samples collected at late post-vaccination time points (days 42 or 84) may have been beyond the peak response observable in peripheral blood (e.g., most of these Salmonella–specific cells may have already homed to secondary lymphoid tissues such as lymph nodes or the gut).
The ability of both mouse and human memory T cells to produce multiple cytokines (MF cells) has been shown to correlate with superior functional ability and protective capacity compared to populations displaying a more restricted cytokine secretion pattern (i.e., Single positive cells) [43–46]. It is now recognized that efficient vaccine induced immunity can be better assessed by evaluating the overall quality of the T cell response, rather than just measuring single functions, e.g., the frequency of IFN-γ producing CD4+T cells [44, 47]. Of note, in this study we observed that Ty21a elicited S. Typhi-specific MF TEM as well as TEMRA subsets following the hierarchy 2+ > 3+ > 4+ MF cells. Previously, we have shown that, Ty21a induced S.Typhi specific CD8+ mediated CMI responses were predominantly comprised of MF cells producing IFN-γ and/or expressing CD107a, classical characteristics of CTL [30]. Moreover, few CD8+ T cells also produced IL-2. On the contrary, in this study, vaccine induced MF CD4+ T cells were mostly dominated by subsets that produced IFN-γ and/or TNF-α, although some cells co-produced IL-2, indicative of a predominant Th1 response. These results are in agreement with the notion that rapid acquisition of CD4+ Th1 effector functions following immunization that include IFN-γ, TNF-α and IL-2 production protect against Salmonella illness [12, 48].
Salmonella specific post-vaccination increases were also observed in a smaller but recognizable subset of CD4+ T cells that expressed CD107a, a marker associated with CTL activity [49]. Classically, CTL activity is mediated by CD8+ T cells. However, a subset of CD4+ cell T cells that exhibits MHC class II-mediated cytotoxic activity has been recently identified in chronic viral infections, autoimmune diseases and cancer; yet their presence and role in gut mucosal immunity remains undefined [50]. The observation of the induction of Salmonella MF CD4+ T cells expressing CD107a following immunization with Ty21a adds information regarding these intriguing cell subsets. However, their precise role in protection from typhoid and other enteric diseases requires further investigation.
Of importance, PBMC obtained from S. Typhi-infected convalescent patients showed an increase in the production of β-chemokines (i.e., RANTES, MIP-1α, MIP-1β) in response to S. Typhi antigens [51]. Recently, we have shown the induction of Salmonella-cross-reactive MF CD8+ MIP-1β+ cells following immunization with Ty21a, suggesting an important role of MIP-1β in host immunity to Salmonella [30]. In this study, we also observed that Ty21a elicited S. Typhi MIP-1β+ cells in CD4+ T cell subset, albeit of lower magnitude to those we observed previously in CD8+ T cells [30]. An appropriate cytokine milieu generated by CD4+ helper T-cells is critical for the generation of strong immune responses against bacterial pathogens, including antibody and CD8+ responses, leading to the elimination of S. Typhi-infected host cells. Although the precise role of Ty21a-induced MF CD4+ TEM and TEMRA cells in protection against typhoid fever remains unknown, the evidence presented in this manuscript supports the notion that Ty21a induced CD4+ MF cells may play an important role in modulating CMI responses which are essential for protection against typhoid fever [11, 14].
Most studies on the role CD4+ T cells in protection against Salmonella have demonstrated their ability to induce potent Th1 or Th2 responses. Recently IL-17-producing CD4+ T helper cells (Th-17) cells have been described as a distinct cell lineage that contributes to host defense against various microbes including Salmonella [37, 52]. In addition to Th1 cells, IL-17A and IL-22 producing cells play a critical role in preventing intestinal dissemination of Salmonella in animal models of infections [37]; in humans, production of IL-17 by CD4+ cells were reported among convalescent but not acute stage of typhoid fever patients [51]. We also previously reported that immunization with Ty21a elicited only small increases in IL-17+ CD8+ T cells [30]. However, in this study we observed sizable post-vaccination increases in specific CD4+ IL-17A+ cells in post-vaccination samples, albeit these subsets was smaller than other Th1 cytokine producing cells (i.e., those producing IFN-γ, TNF-α). These Salmonella specific MF IL-17A+ cells were largely observed in CD4+ TEM subsets. These data are in agreement with previous observations that among the various cell types (i.e., CD4+, CD8+ or γδT cells), CD4+ cells are the major producers of IL-17A [52]. Here, we report for the first time that Ty21a immunization elicited S. Typhi specific CD4+ Th17 cells and that these responses cross-react to S. Paratyphi A or S. Paratyphi B. These data showing that Ty21a induced MF IL-17A+ cells suggest that IL-17A may contribute in protective immunity against Salmonella infection in humans. However, many questions remained to be investigated including the kinetics of induction of IL-17A producing cells, their concomitant expression of IL-23, retinoic acid receptor-related orphan nuclear receptor (ROR)-t transcription factor and CCR6, as well as cytokines associated with IL-17 (e.g., TNF-α, IL-22, IL-12, IL-6, IL-1β) [21, 52].
Salmonella enterica serovars causing enteric fevers in humans enter the host via the gut, causing both local and systemic pathology [1]. Therefore, effector immune responses at the gut level are presumed to be a critical component of vaccine induced protective immunity against these organisms. In past reports we have demonstrated that significant components of the immunity elicited by oral Salmonella vaccines included both CD4+ and CD8+ S. Typhi-specific TM cells, comprised of effector cells with the potential to migrate to gut (integrin α4β7+) [24, 29, 30]. Moreover, we described that higher vaccine induced responses were observed among the integrin α4β7 expressing cells than those that did not express integrin α4β7 [29]. In the present study we extended these observations by demonstrating that S. Typhi, S. Paratyphi A and S. Paratyphi B cross-reactive CD4+ MF T cells elicited following immunization with Ty21a consisted of cells expressing, or not, integrin α4β7. Similar to our previous observations with CD8+ T cells, we detected that a lower percentage of Salmonella-specific CD4+ MF integrin α4β7+ cells in circulation compared to integrin α4β7 negative cells. This observation could be due to the fact that a sizable fraction of the T cells in circulation do not express integrin α4β7 due to the fact that most of Salmonella-specific integrin α4β7+ cells have already migrated and reside in the gut. Regarding cross-reactivity to other Salmonella species responsible for enteric fevers, we observed that similar responses in terms of magnitude and characteristics of these CD4+ MF T cells were observed against S. Paratyphi A and S. Paratyphi B. It is possible that these responses are mediated through Salmonella common antigens (e.g., epitopes contained in flagellar subunit protein FliC) that are presented by Salmonella-infected autologous targets. However, since we did not see differences in the induction of these CD4+ MF cells that cross-react to S. Typhi, S. Paratyphi A and S. Paratyphi B, it is unlikely that these cells are responsible for the observed cross-protection of Ty21a vaccination to S. Paratyphi B infection but not to S. Paratyphi A [7, 8].
This study was partly limited by the unavailability of a larger sample size, due to inherent properties of such clinical studies. Moreover, we could utilize PBMC obtained at only two post-vaccination time points, which did not allow us to evaluate the precise kinetics of the induction and persistence of CD4+ CMI responses mediated by multiple cytokines or chemokines. Further in-depth studies with additional subjects and time points after vaccination will be required to fully understand the role of MF CD4+ T cells in Ty21a induced protective immunity against Salmonella.
5. Conclusions
In this study we show that live oral typhoid vaccines induce multifunctional S. Typhi-specific CD4+ T cell responses that cross-react with S. Paratyphi A and S. Paratyphi B. Our results further showed, for the first time in enteric bacterial diseases, that a small subset of vaccine-induced Salmonella-specific MF CD4+ T cells have cytotoxic potential, as assessed by expression of CD107a. In addition, we also demonstrate that immunization with Ty21a resulted in MF CD4+ cells that produce the pro-inflammatory cytokines MIP-1β and IL-17A, the latter a critical cytokine indicative of the induction of Th17 cells, and that these responses cross-react with S. Paratyphi A and S. Paratyphi B. Finally, we also show that a significant proportion of Salmonella-specific CD4+ T cells expressed the gut homing marker integrin α4β7. These results with CD4+ T cells, in contrast to our previous observations with Ty21a elicited CD8+T cells responses, did not show significant differences among S. Typhi, S. Paratyphi A or S. Paratyphi B. Overall, these data in conjunction with previous reports on humoral and CD8+ T cell mediated cross-reactive responses, leads us to hypothesize that, while both humoral and a CD4+ T cell responses might play an important role, it is an efficient and dominant CD8+ T cell response that may be the key CMI response responsible for the observed long-protection conferred by Ty21a against S. Typhi or S. Paratyphi B, but not S. Paratyphi A. However, the ultimate implications of these immunological responses in vaccine-induced protection against Salmonella will need to be evaluated further in human wild-type challenge models. In addition, the results of this study may also prove useful in exploring the use of attenuated S. Typhi oral vaccine strains to deliver foreign vaccine antigens for which CD4-mediated responses are considered to be of importance [53, 54].
Supplementary Material
Highlights.
Ty21a elicits Multifunctional (MF) Salmonella-specific CD4+ T cells in humans
MF cells predominantly produce IFN-γ, TNF-α and, to a lesser extent, IL-2
Some Ty21a elicited specific-MF CD4+ cells exhibit cytotoxic and Th-17 phenotypes
A third of CD4+ specific MF cells expressed the gut homing molecule integrin-α4β7
Similar MF CD4+ T cell responses were elicited against all Salmonella strains
Acknowledgments
We thankfully acknowledge Regina Harley, Catherine Storrer, Haiyan Chen and Shah Zafar for excellent technical assistance. We also thank the participating volunteers, Robin Barnes and the staff from the Recruiting Section of Center for Vaccine Development for the blood specimens. This study was funded by NIAID, NIH, DHHS grants R01-AI036525 (to M.B.S.), U19 AI082655 (Cooperative Center for Translational Research in Human Immunology and Biodefense [CCHI], to M.B.S.), and U54-AI057168 (Regional Center for Excellence for Biodefense and Emerging Infectious Diseases Research Mid-Atlantic Region [MARCE] and U19-AI109776 (Center of Excellence for Translational Research [CETR], to M.M.L. and M.B.S).
Abbreviations
- CMI
Cell-mediated immune
- S+
Single positive cells
- TEM
T effector/memory
- TEMRA
CD45RA+ T effector/memory
- MF
Multifunctional
Footnotes
Disclosures. The authors declare no conflict of interest. The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases or the National Institutes of Health.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Parry CM, Hien TT, Dougan G, White NJ, Farrar JJ. Typhoid fever. N Engl J Med. 2002;347:1770–1782. doi: 10.1056/NEJMra020201. [DOI] [PubMed] [Google Scholar]
- 2.Crump JA, Mintz ED. Global trends in typhoid and paratyphoid Fever. Clin Infect Dis. 2010;50:241–246. doi: 10.1086/649541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Arndt MB, Mosites EM, Tian M, Forouzanfar MH, Mokhdad AH, Meller M, Ochiai RL, Walson JL. Estimating the burden of paratyphoid a in Asia and Africa. PLoS Negl Trop Dis. 2014;8:e2925. doi: 10.1371/journal.pntd.0002925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Meltzer E, Stienlauf S, Leshem E, Sidi Y, Schwartz E. A Large Outbreak of Salmonella Paratyphi A Infection Among Israeli Travelers To Nepal. Clinical Infectious Diseases. 2014;58:359–364. doi: 10.1093/cid/cit723. [DOI] [PubMed] [Google Scholar]
- 5.Teh CS, Chua KH, Thong KL. Paratyphoid fever: splicing the global analyses. Int J Med Sci. 2014;11:732–741. doi: 10.7150/ijms.7768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McClelland M, Sanderson KE, Clifton SW, Latreille P, Porwollik S, Sabo A, Meyer R, Bieri T, Ozersky P, McLellan M, Harkins CR, Wang C, Nguyen C, Berghoff A, Elliott G, Kohlberg S, Strong C, Du F, Carter J, Kremizki C, Layman D, Leonard S, Sun H, Fulton L, Nash W, Miner T, Minx P, Delehaunty K, Fronick C, Magrini V, Nhan M, Warren W, Florea L, Spieth J, Wilson RK. Comparison of genome degradation in Paratyphi A and Typhi, human-restricted serovars of Salmonella enterica that cause typhoid. Nat Genet. 2004;36:1268–1274. doi: 10.1038/ng1470. [DOI] [PubMed] [Google Scholar]
- 7.Levine MM, Ferreccio C, Black RE, Lagos R, San MO, Blackwelder WC. Ty21a live oral typhoid vaccine and prevention of paratyphoid fever caused by Salmonella enterica Serovar Paratyphi B. Clin Infect Dis. 2007;45(Suppl 1):S24–S28. doi: 10.1086/518141. [DOI] [PubMed] [Google Scholar]
- 8.Simanjuntak CH, Paleologo FP, Punjabi NH, Darmowigoto R, Soeprawoto, Totosudirjo H, Haryanto P, Suprijanto E, Witham ND, Hoffman SL. Oral immunisation against typhoid fever in Indonesia with Ty21a vaccine. Lancet. 1991;338:1055–1059. doi: 10.1016/0140-6736(91)91910-m. [DOI] [PubMed] [Google Scholar]
- 9.Fangtham M, Wilde H. Emergence of Salmonella paratyphi A as a major cause of enteric fever: need for early detection, preventive measures, and effective vaccines. J Travel Med. 2008;15:344–350. doi: 10.1111/j.1708-8305.2008.00237.x. [DOI] [PubMed] [Google Scholar]
- 10.Gupta SK, Medalla F, Omondi MW, Whichard JM, Fields PI, Gerner-Smidt P, Patel NJ, Cooper KL, Chiller TM, Mintz ED. Laboratory-based surveillance of paratyphoid fever in the United States: travel and antimicrobial resistance. Clin Infect Dis. 2008;46:1656–1663. doi: 10.1086/587894. [DOI] [PubMed] [Google Scholar]
- 11.Sztein MB. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica Serovar Typhi strains used as live oral vaccines in humans. Clin Infect Dis. 2007;45(Suppl 1):S15–S19. doi: 10.1086/518140. [DOI] [PubMed] [Google Scholar]
- 12.Griffin AJ, McSorley SJ. Development of protective immunity to Salmonella, a mucosal pathogen with a systemic agenda. Mucosal Immunol. 2011;4:371–382. doi: 10.1038/mi.2011.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dougan G, John V, Palmer S, Mastroeni P. Immunity to salmonellosis. Immunol Rev. 2011;240:196–210. doi: 10.1111/j.1600-065X.2010.00999.x. [DOI] [PubMed] [Google Scholar]
- 14.Sztein MB, Salerno-Goncalves R, McArthur MA. Complex adaptive immunity to enteric fevers in humans: lessons learned and the path forward. Front Immunol. 2014;5:516. doi: 10.3389/fimmu.2014.00516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McArthur MA, Fresnay S, Magder LS, Darton TC, Jones C, Waddington CS, Blohmke CJ, Dougan G, Angus B, Levine MM, Pollard AJ, Sztein MB. Activation of Salmonella Typhi-specific regulatory T cells in typhoid disease in a wild-type S. Typhi challenge model. PLoS Pathog. 2015;11:e1004914. doi: 10.1371/journal.ppat.1004914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Toapanta FR, Bernal PJ, Fresnay S, Darton TC, Jones C, Waddington CS, Blohmke CJ, Dougan G, Angus B, Levine MM, Pollard AJ, Sztein MB. Oral Wild-Type Salmonella Typhi Challenge Induces Activation of Circulating Monocytes and Dendritic Cells in Individuals Who Develop Typhoid Disease. PLoS Negl Trop Dis. 2015;9:e0003837. doi: 10.1371/journal.pntd.0003837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Waddington CS, Darton TC, Jones C, Haworth K, Peters A, John T, Thompson BAV, Kerridge SA, Kingsley RA, Zhou L, Holt KE, Yu L-M, Lockhart S, Farrar JJ, Sztein MB, Dougan G, Angus B, Levine MM, Pollard AJ. An Outpatient, Ambulant-Design, Controlled Human Infection Model Using Escalating Doses of Salmonella Typhi Challenge Delivered in Sodium Bicarbonate Solution. Clinical Infectious Diseases. 2014;58:1230–1240. doi: 10.1093/cid/ciu078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fresnay S, McArthur MA, Magder L, Darton TC, Jones C, Waddington CS, Blohmke CJ, Angus B, Levine MM, Pollard AJ, Sztein MB. Salmonella Typhi-specific multifunctional CD8+ T cells play a dominant role in protection from typhoid fever in humans. J Transl Med. 2016;14:62. doi: 10.1186/s12967-016-0819-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kirkpatrick BD, McKenzie R, O’Neill JP, Larsson CJ, Bourgeois AL, Shimko J, Bentley M, Makin J, Chatfield S, Hindle Z, Fidler C, Robinson BE, Ventrone CH, Bansal N, Carpenter CM, Kutzko D, Hamlet S, LaPointe C, Taylor DN. Evaluation of Salmonella enterica serovar Typhi (Ty2 aroC-ssaV-) M01ZH09, with a defined mutation in the Salmonella pathogenicity island 2, as a live, oral typhoid vaccine in human volunteers. Vaccine. 2006;24:116–123. doi: 10.1016/j.vaccine.2005.08.008. [DOI] [PubMed] [Google Scholar]
- 20.Lindow JC, Fimlaid KA, Bunn JY, Kirkpatrick BD. Antibodies in action: role of human opsonins in killing Salmonella enterica serovar Typhi. Infect Immun. 2011;79:3188–3194. doi: 10.1128/IAI.05081-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.McArthur MA, Sztein MB. Heterogeneity of multifunctional IL-17A producing S. Typhi-specific CD8+ T cells in volunteers following Ty21a typhoid immunization. PLoS One. 2012;7:e38408. doi: 10.1371/journal.pone.0038408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Salerno-Goncalves R, Fernandez-Vina M, Lewinsohn DM, Sztein MB. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J Immunol. 2004;173:5852–5862. doi: 10.4049/jimmunol.173.9.5852. [DOI] [PubMed] [Google Scholar]
- 23.Salerno-Goncalves R, Pasetti MF, Sztein MB. Characterization of CD8(+) effector T cell responses in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J Immunol. 2002;169:2196–2203. doi: 10.4049/jimmunol.169.4.2196. [DOI] [PubMed] [Google Scholar]
- 24.Salerno-Goncalves R, Wahid R, Sztein MB. Immunization of volunteers with Salmonella enterica serovar Typhi strain Ty21a elicits the oligoclonal expansion of CD8+ T cells with predominant Vbeta repertoires. Infect Immun. 2005;73:3521–3530. doi: 10.1128/IAI.73.6.3521-3530.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Salerno-Goncalves R, Wahid R, Sztein MB. Ex Vivo kinetics of early and long-term multifunctional human leukocyte antigen E-specific CD8+ cells in volunteers immunized with the Ty21a typhoid vaccine. Clin Vaccine Immunol. 2010;17:1305–1314. doi: 10.1128/CVI.00234-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Salerno-Goncalves R, Wyant TL, Pasetti MF, Fernandez-Vina M, Tacket CO, Levine MM, Sztein MB. Concomitant induction of CD4+ and CD8+ T cell responses in volunteers immunized with Salmonella enterica serovar typhi strain CVD 908-htrA. J Immunol. 2003;170:2734–2741. doi: 10.4049/jimmunol.170.5.2734. [DOI] [PubMed] [Google Scholar]
- 27.Wahid R, Pasetti MF, Maciel M, Jr, Simon JK, Tacket CO, Levine MM, Sztein MB. Oral priming with Salmonella Typhi vaccine strain CVD 909 followed by parenteral boost with the S. Typhi Vi capsular polysaccharide vaccine induces CD27+IgD-S. Typhi-specific IgA and IgG B memory cells in humans. Clin Immunol. 2011;138:187–200. doi: 10.1016/j.clim.2010.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wahid R, Salerno-Goncalves R, Tacket CO, Levine MM, Sztein MB. Cell-mediated immune responses in humans after immunization with one or two doses of oral live attenuated typhoid vaccine CVD 909. Vaccine. 2007;25:1416–1425. doi: 10.1016/j.vaccine.2006.10.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wahid R, Salerno-Goncalves R, Tacket CO, Levine MM, Sztein MB. Generation of specific effector and memory T cells with gut- and secondary lymphoid tissue- homing potential by oral attenuated CVD 909 typhoid vaccine in humans. Mucosal Immunol. 2008;1:389–398. doi: 10.1038/mi.2008.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wahid R, Fresnay S, Levine MM, Sztein MB. Immunization with Ty21a live oral typhoid vaccine elicits crossreactive multifunctional CD8+ T-cell responses against Salmonella enterica serovar Typhi, S. Paratyphi A, and S. Paratyphi B in humans. Mucosal Immunol. 2015;8:1349–1359. doi: 10.1038/mi.2015.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sztein MB, Tanner MK, Polotsky Y, Orenstein JM, Levine MM. Cytotoxic T lymphocytes after oral immunization with attenuated vaccine strains of Salmonella typhi in humans. J Immunol. 1995;155:3987–3993. [PubMed] [Google Scholar]
- 32.Lundin BS, Johansson C, Svennerholm AM. Oral Immunization with a Salmonella enterica Serovar Typhi Vaccine Induces Specific Circulating Mucosa-Homing CD4+ and CD8+ T Cells in Humans. Infection and Immunity. 2002;70:5622–5627. doi: 10.1128/IAI.70.10.5622-5627.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wahid R, Simon R, Zafar SJ, Levine MM, Sztein MB. Live oral typhoid vaccine Ty21a induces cross-reactive humoral immune responses against Salmonella enterica serovar Paratyphi A and S. Paratyphi B in humans. Clin Vaccine Immunol. 2012;19:825–834. doi: 10.1128/CVI.00058-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wahid R, Zafar SJ, McArthur MA, Pasetti MF, Levine MM, Sztein MB. Live Oral Salmonella enterica Serovar Typhi Vaccines Ty21a and CVD 909 Induce Opsonophagocytic Functional Antibodies in Humans That Cross-React with S. Paratyphi A and S. Paratyphi B. Clinical and Vaccine Immunology. 2014;21:427–434. doi: 10.1128/CVI.00786-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pakkanen SH, Kantele JM, Kantele A. Cross-reactive gut-directed immune response against Salmonella enterica serovar Paratyphi A and B in typhoid fever and after oral Ty21a typhoid vaccination. Vaccine. 2012;30:6047–6053. doi: 10.1016/j.vaccine.2012.07.051. [DOI] [PubMed] [Google Scholar]
- 36.Sztein MB, Wasserman SS, Tacket CO, Edelman R, Hone D, Lindberg AA, Levine MM. Cytokine production patterns and lymphoproliferative responses in volunteers orally immunized with attenuated vaccine strains of Salmonella typhi. J Infect Dis. 1994;170:1508–1517. doi: 10.1093/infdis/170.6.1508. [DOI] [PubMed] [Google Scholar]
- 37.Pham OH, McSorley SJ. Protective host immune responses to Salmonella infection. Future Microbiol. 2015;10:101–110. doi: 10.2217/fmb.14.98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Johansson-Lindbom B, Agace WW. Generation of gut-homing T cells and their localization to the small intestinal mucosa. Immunol Rev. 2007;215:226–242. doi: 10.1111/j.1600-065X.2006.00482.x. [DOI] [PubMed] [Google Scholar]
- 39.Levine MM, Ferreccio C, Abrego P, Martin OS, Ortiz E, Cryz S. Duration of efficacy of Ty21a, attenuated Salmonella typhi live oral vaccine. Vaccine. 1999;17(Suppl 2):S22–S27. doi: 10.1016/s0264-410x(99)00231-5. [DOI] [PubMed] [Google Scholar]
- 40.Mastroeni P, Villarreal-Ramos B, Hormaeche CE. Role of T cells, TNF alpha and IFN gamma in recall of immunity to oral challenge with virulent salmonellae in mice vaccinated with live attenuated aro- Salmonella vaccines. Microb Pathog. 1992;13:477–491. doi: 10.1016/0882-4010(92)90014-f. [DOI] [PubMed] [Google Scholar]
- 41.Hess J, Ladel C, Miko D, Kaufmann SH. Salmonella typhimurium aroA- infection in gene-targeted immunodeficient mice: major role of CD4+ TCR-alpha beta cells and IFN-gamma in bacterial clearance independent of intracellular location. J Immunol. 1996;156:3321–3326. [PubMed] [Google Scholar]
- 42.Pasetti MF, Simon JK, Sztein MB, Levine MM. Immunology of gut mucosal vaccines. Immunological Reviews. 2011;239:125–148. doi: 10.1111/j.1600-065X.2010.00970.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Maybeno M, Redeker A, Welten SP, Peters B, Loughhead SM, Schoenberger SP, Sette A, Arens R. Polyfunctional CD4+ T cell responses to immunodominant epitopes correlate with disease activity of virulent Salmonella. PLoS One. 2012;7:e43481. doi: 10.1371/journal.pone.0043481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Seder RA, Darrah PA, Roederer M. T-cell quality in memory and protection: implications for vaccine design. Nat Rev Immunol. 2008;8:247–258. doi: 10.1038/nri2274. [DOI] [PubMed] [Google Scholar]
- 45.Stanley DA, Honko AN, Asiedu C, Trefry JC, Lau-Kilby AW, Johnson JC, Hensley L, Ammendola V, Abbate A, Grazioli F, Foulds KE, Cheng C, Wang L, Donaldson MM, Colloca S, Folgori A, Roederer M, Nabel GJ, Mascola J, Nicosia A, Cortese R, Koup RA, Sullivan NJ. Chimpanzee adenovirus vaccine generates acute and durable protective immunity against ebolavirus challenge. Nature medicine. 2014;20:1126–1129. doi: 10.1038/nm.3702. [DOI] [PubMed] [Google Scholar]
- 46.Thakur A, Pedersen LE, Jungersen G. Immune markers and correlates of protection for vaccine induced immune responses. Vaccine. 2012;30:4907–4920. doi: 10.1016/j.vaccine.2012.05.049. [DOI] [PubMed] [Google Scholar]
- 47.Darrah PA, Patel DT, De Luca PM, Lindsay RWB, Davey DF, Flynn BJ, Hoff ST, Andersen P, Reed SG, Morris SL, Roederer M, Seder RA. Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat Med. 2007;13:843–850. doi: 10.1038/nm1592. [DOI] [PubMed] [Google Scholar]
- 48.Srinivasan A, Foley J, McSorley SJ. Massive number of antigen-specific CD4 T cells during vaccination with live attenuated Salmonella causes interclonal competition. J Immunol. 2004;172:6884–6893. doi: 10.4049/jimmunol.172.11.6884. [DOI] [PubMed] [Google Scholar]
- 49.Betts MR, Brenchley JM, Price DA, De Rosa SC, Douek DC, Roederer M, Koup RA. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. Journal of Immunological Methods. 2003;281:65–78. doi: 10.1016/s0022-1759(03)00265-5. [DOI] [PubMed] [Google Scholar]
- 50.Cheroutre H, Husain MM. CD4 CTL: living up to the challenge. Semin Immunol. 2013;25:273–281. doi: 10.1016/j.smim.2013.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bhuiyan S, Sayeed A, Khanam F, Leung DT, Rahman Bhuiyan T, Sheikh A, Salma U, LaRocque RC, Harris JB, Pacek M, Calderwood SB, LaBaer J, Ryan ET, Qadri F, Charles RC. Cellular and Cytokine Responses to Salmonella enterica Serotype Typhi Proteins in Patients with Typhoid Fever in Bangladesh. The American Journal of Tropical Medicine and Hygiene. 2014;90:1024–1030. doi: 10.4269/ajtmh.13-0261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.McGeachy MJ, McSorley SJ. Microbial-induced Th17: Superhero or Supervillain? Journal of immunology (Baltimore, Md : 1950) 2012;189:3285–3291. doi: 10.4049/jimmunol.1201834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pasetti MF, Anderson RJ, Noriega FR, Levine MM, Sztein MB. Attenuated deltaguaBA Salmonella typhi vaccine strain CVD 915 as a live vector utilizing prokaryotic or eukaryotic expression systems to deliver foreign antigens and elicit immune responses. Clin Immunol. 1999;92:76–89. doi: 10.1006/clim.1999.4733. [DOI] [PubMed] [Google Scholar]
- 54.Tacket CO, Galen J, Sztein MB, Losonsky G, Wyant TL, Nataro J, Wasserman SS, Edelman R, Chatfield S, Dougan G, Levine MM. Safety and immune responses to attenuated Salmonella enterica serovar typhi oral live vector vaccines expressing tetanus toxin fragment C. 97. 2000. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.