

Supplemental Digital Content is available in the text
Keywords: developing world, HIV, enteric, microbiome, systemic inflammation, virome
Abstract
Purpose of review
Despite HIV therapy advances, average life expectancy in HIV-infected individuals on effective treatment is significantly decreased relative to uninfected persons, largely because of increased incidence of inflammation-related diseases, such as cardiovascular disease and renal dysfunction. The enteric microbial community could potentially cause this inflammation, as HIV-driven destruction of gastrointestinal CD4+ T cells may disturb the microbiota–mucosal immune system balance, disrupting the stable gut microbiome and leading to further deleterious host outcomes.
Recent findings
Varied enteric microbiome changes have been reported during HIV infection, but unifying patterns have emerged. Community diversity is decreased, similar to pathologies such as inflammatory bowel disease, obesity, and Clostridium difficile infection. Many taxa frequently enriched in HIV-infected individuals, such as Enterobacteriaceae and Erysipelotrichaceae, have pathogenic potential, whereas depleted taxa, such as Bacteroidaceae and Ruminococcaceae, are more linked with anti-inflammatory properties and maintenance of gut homeostasis. The gut viral community in HIV has been found to contain a greater abundance of pathogenesis-associated Adenoviridae and Anelloviridae. These bacterial and viral changes correlate with increased systemic inflammatory markers, such as serum sCD14, sCD163, and IL-6.
Summary
Enteric microbial community changes may contribute to chronic HIV pathogenesis, but more investigation is necessary, especially in the developing world population with the greatest HIV burden (Video, Supplemental Digital Content 1, which includes the authors’ summary of the importance of the work).
INTRODUCTION
The enteric ‘microbiome’ consists of a diverse collection of trillions of Bacteria, Archaea, Eukarya, and viruses [1–5], with a large aggregate genome, referred to as the ‘metagenome’, that contributes to normal immune development [6] and a number of pathological processes [7–9]. The host immune system acts as an essential curator for this luminal enteric microbial community, serving to shape and control the structure and function of this diverse collection of organisms [10–12]. HIV infection leads to the widespread destruction of host immune function [13,14], including the rapid and profound depletion of CD4+ T cells within gut-associated lymphoid tissue [15,16]. As could be predicted from the loss of mucosal immune cells, a wide range of changes in the enteric microbial community have been reported during HIV infection [17,18▪▪,19,20▪,21▪▪,22▪,23▪▪,24▪–27▪,28▪▪–32▪▪,33▪] (Fig. 1 and Table S1). In light of the role of the enteric microbiome in many inflammation-associated pathologies such as diabetes, obesity, and inflammatory bowel disease (IBD) [7–9], it has been proposed that HIV-associated microbiome shifts could contribute to inflammation-related noncommunicable diseases (NCD) that are responsible for a large share of the increased mortality observed during chronic HIV infection [34–39]. Investigation of microbial changes associated with HIV infection has the potential to aid in the development of therapeutic interventions that could improve many of the pathologic consequences of chronic HIV infection.
FIGURE 1.

Alterations in the gut microbiota reported in HIV microbiome studies. Black and gray boxes, respectively, indicate reported increases and decreases in relative abundance. Bracketed letter indicates taxonomic level of classification (e.g., [p] indicates phylum), parenthetical taxa indicate subtaxa that commonly drive significance of parent taxa. aDysbiosis observed in HIV-positive subjects with less than 200 CD4+ T cells/μL. Black dashed box outlines the only study to date that investigated the relationship between gut microbiota and HIV in a sub-Saharan African cohort. bFecal microbiome data only obtained for HIV seronegative unexposed and exposed infants. cProteobacteria class indicated by corresponding Greek letter (e.g., β for Betaproteobacteria). Enterobacteriaceae are Gammaproteobacteria [c] within Proteobacteria [p]. ART, antiretroviral therapy.
Box 1.

no caption available
CHRONIC SYSTEMIC IMMUNE ACTIVATION IN HIV INFECTION
Despite advancements in HIV antiretroviral therapy (ART), average life expectancy in HIV-infected individuals on effective treatment is 14% less than that of uninfected persons [35]. The ART Cohort Collaboration Study found that deaths were largely because of inflammation-related clinical diseases, such as stroke, long-bone fractures, cardiovascular disease (CVD), and renal dysfunction [34,35,37,38]. CVD, which constitutes a large proportion of HIV-associated NCD [40–42], is increased in HIV infection [43–45] and associated with systemic immune activation as measured by markers such as serum IL-6, sCD163, and C-reactive protein [46–51]. In HIV-uninfected individuals, microbiota-induced inflammation has been shown to drive CVD pathogenesis [52–55], suggesting that this disorder could occur in HIV as well.
Further supporting the connection between systemic inflammation and chronic pathogenesis, individuals with the highest degree of persistent elevated immune activation while on suppressive ART experience higher overall mortality, even with CD4+ T-cell reconstitution more than 500 cells/μl [56,57]. Although systemic immune activation declines after initiation of ART, it remains persistently elevated in the majority of study participants even after years of therapy [58–60] and has also been observed in individuals with undetectable viral loads [61]. In a cohort of HIV-infected Ugandans, a 1.6-fold increased hazard of death was associated with each 10% increase in CD8+ T-cell activation following initial viral suppression with ART [62].
POTENTIAL ROLE OF HIV-ASSOCIATED GUT MICROBIAL CHANGES IN HIV DISEASE PROGRESSION
HIV-associated changes to the enteric microbiome may lead to systemic inflammation by disrupting the balance of metabolic functions performed by the microbiota, such as short-chain fatty acid or bile acid metabolism [63], or causing increased translocation of bacterial products into the systemic circulation [64]. Elevated plasma kynurenine, a tryptophan metabolite, has been found to be associated with CD8+ T-cell activation [65] and mortality [66] in HIV-infected individuals and HIV infection was associated with the presence of a gut microbial community with both the genetic capacity to metabolize tryptophan into kynurenine and demonstrable kynurenine production in vitro[21▪▪]. In a similar paradigm, independent of HIV, the enteric microbiome can drive CVD pathogenesis by transforming dietary constituents such as phosphatidylcholine and bile acids into reactive intermediates such as trimethylamine N-oxide that can lead to macrophage and platelet activation, thrombosis, and arterial plaque formation [67–71].
Early in HIV infection the intestinal mucosa is a critical reservoir of viral replication and substantial CD4+ T-cell depletion (80–90%) [72–80]. This depletion is attributed to the widespread surface expression of CCR5 on gastrointestinal T cells, which serves as the coreceptor for HIV entry early on in infection [13,72]. Because certain lymphocyte subsets are important to the maintenance of enteric epithelial integrity (e.g., IL-22-producing ILC3 and Th17 cells [12,81]), it is believed that such a loss of immune surveillance may result in impaired intestinal epithelial barrier function, increased gut permeability, and the translocation of gut bacterial products into systemic circulation [16,64]. These products could trigger persistent systemic immune activation and drive turnover of CD4+ and CD8+ T cells leading to clonal exhaustion and progressive impairment of T-cell function [82]. HIV-associated chronic systemic immune activation, secondary to a loss of gut epithelial homeostasis, may then lead to disease progression in both treated and untreated HIV infection. This model is supported by data showing that circulating levels of lipopolysaccharide (LPS), a bacterial product known to elicit an innate immune response by binding host Toll-like receptors [83], increase significantly as HIV infection progresses [84], though initiation of ART lowers circulating LPS levels [64]. Individuals with both treated and untreated HIV infection, however, exhibit elevated serum levels of bacterial LPS and systemic inflammation relative to uninfected controls [64,85,86].
BACTERIAL MICROBIOME DIFFERENCES IN HIV INFECTION
Changes in the composition of the enteric microbiome that cause it to deviate from what is considered a healthy baseline state are often collectively referred to as ‘dysbiosis’. Dysbiotic states are thought to underlie many of the pathologies linked with the microbiota [7–9]. Comparative studies between HIV-infected and uninfected individuals have differed in methods and patient populations studied (Fig. 2 and Table S1) and found a wide spectrum of differences associated with HIV infection (Figs. 1 and S1). However, several overarching themes have emerged. HIV-infected individuals often have reduced diversity of their enteric microbiome, which is also observed in a number of pathologies such as IBD [87–89], type 1 diabetes mellitus [90], obesity [91], and Clostridium difficile infection [92,93]. HIV infection also frequently correlates with increased tissue and circulating measures of inflammation (e.g., sCD14, IL-6, CD38+HLA-DR+CD8+ T cells) [64,85,86], as well as increases in traditionally pathogenic bacteria such as Enterobacteriaceae [20▪,21▪▪,24▪,29▪▪,31▪▪,32▪▪,33▪] and decreases in commensals such as Lactobacillaceae, Lachnospiraceae, and Ruminococcaceae [19,20▪,21▪▪,22▪,23▪▪,25▪,26▪,28▪▪–30▪▪,32▪▪,94,95].
FIGURE 2.
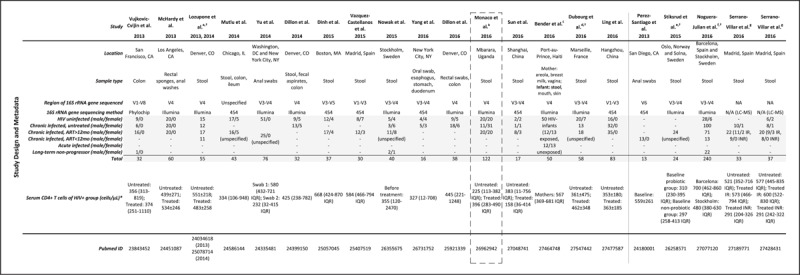
Study design and patient metadata in HIV microbiome studies. ∗CD4 cell counts (cells/μL) displayed as either median [range (not notated in table)] median (IQR), or average ± SD. †Sex composition of clinical study participant groups not specified. aThe same patient cohort was used for both Lozupone et al. 2013 and 2014. Displayed is patient information from Lozupone et al. 2014, which included 17 additional subjects. Proportion of males versus females in each clinical group is only described in Lozupone et al. 2013 (male/female: 8/5 seronegative; 11/0 HIV+ untreated; 7/1 HIV+ treated). bThe black dashed box outlines the only study to date that investigated the relationship between gut microbiota and HIV in a sub-Saharan African cohort. cFecal microbiome data only obtained for HIV seronegative unexposed and exposed infants. dThe HIV-infected group has a total of 31 individuals, including 22 males and 9 females, specific clinical group not specified. eThirty study participants (22 males, 8 females) originally enrolled but only 24 study participants completed the study, specific clinical group not specified. fThe HIV-infected groups of the Barcelona and Stockholm cohorts, respectively, included 101 males and 28 females, and 46 males and 31 females, specific clinical group not specified. gImmunological responders (IR) and nonresponders (INR) respectively defined as having at least 350 and less than 350 CD4+ T-cell counts/μl after more than 2 years of ART. ART, antiretroviral therapy.
Taxa from four bacterial phyla – Proteobacteria, Fusobacteria, Bacteroidetes, and Firmicutes – have been reported in multiple studies to differ between HIV-infected and HIV-uninfected individuals. Proteobacteria were more abundant in HIV-infected individuals in 10 of the 16 published studies whereas others reported no change [17,18▪▪,19,20▪,21▪▪,22▪,23▪▪,24▪,27▪,30▪▪,32▪▪,33▪]. The phylum Proteobacteria includes numerous pathogens, such as Shigella, Salmonella, and Helicobacter. Many specific Proteobacteria genera have been shown to be enriched in HIV-infected individuals and are capable of potentially pathogenic activities in this context [96]: Pseudomonas[21▪▪,95] is an opportunistic pathogen that is capable of impairing host mucus production [97,98]; Desulfovirbrio[17] can produce hydrogen sulfide compounds and inflame the host epithelium [99]; Acinetobacter[23▪▪] can produce LPS [100] and in vitro is able to induce IL-8 production and neutrophil recruitment that could lead to inflammatory tissue damage [101,102]; and Campylobacter[20▪] produces multiple toxins [103] that can induce mucosal inflammation [104]. The Proteobacterial family Enterobacteriaceae is associated with inflammation [105,106] and was enriched in HIV-infected individuals in seven of the 16 studies surveyed [20▪,21▪▪,24▪,29▪▪,31▪▪,32▪▪,33▪] (most frequently driven by its constituent genus Escherichia, which is capable of pathogenic bile acid transformations and degradation of host mucus [98,107]). The phylum Fusobacteria, which is of particular interest because of its previous associations with intestinal inflammation and colorectal cancer [108–110], was enriched (most frequently driven by its constituent genus Fusobacterium) in HIV-infected individuals in four of the 16 previously published studies [19,20▪,22▪,32▪▪].
Taxa in the phylum Bacteroidetes, including the families Prevotellaceae, Porphyromonadaceae, Bacteroidaceae, and Rikenellaceae, exhibited a more heterogeneous pattern of changes in HIV-infected individuals. The family Prevotellaceae (most frequently driven by its constituent genus Prevotella) was enriched in HIV-infected individuals in seven of 16 studies [17,18▪▪,20▪,23▪▪,26▪,27▪,28▪▪,33▪] and depleted in one study [31▪▪]. Taxa within the family Prevotellaceae have been associated with inflammation (particularly in the context of autoinflammatory disease [111,112]) and activation of gut dendritic cells [28▪▪], but a greater abundance of Prevotellaceae is also characteristic of the baseline enteric microbial community of healthy individuals in developing world countries such as Burkina Faso, Venezuela, Malawi, or Papua New Guinea [17,113–117]. Taxa from the family Porphyromonadaceae (most frequently driven by the genera Barnesiella or Odoribacter) were generally depleted in HIV-infected individuals, exhibiting a decrease in nine of 16 studies [17,18▪▪,20▪,21▪▪,22▪,23▪▪,30▪▪–32▪▪,33▪] and an increase in two studies [19,24▪]. Independent of HIV, Porphyromonadaceae exhibit a diverse and complex array of functions, with both positive [110,118,119] and negative [119] associations with colorectal cancer, and negative associations with C. difficile[120–122], Salmonella[123], vancomycin-resistant Enterococcus[124], and Citrobacter rodentium[125,126] infection that imply a putative protective role. The family Bacteroidaceae (mostly driven by the abundance of the genus Bacteroides) is overall depleted in HIV infection, showing a reduced HIV-associated abundance in 10 of 16 studies [17,18▪▪,21▪▪,23▪▪,26▪,28▪▪–31▪▪,33▪]. This family is generally considered to play an anti-inflammatory role [127–131], with the species Bacteroides fragilis promoting regulatory T-cell differentiation and IL-10 production via secreted bacterial products [127–129]. The family Rikenellaceae (mostly driven by the abundance of the genus Alistipes) is also depleted in HIV infection, with decreased abundance in seven studies [17,18▪▪,19,20▪,21▪▪,23▪▪,24▪,28▪▪] and this bile-tolerant family [132] displays protective properties against C. difficile infection [133] and a negative association with obesity [134] as well as positive associations with both type 1 [135] and type 2 [136] diabetes mellitus.
Similar to the phylum Bacteroidetes, bacterial families within the phylum Firmicutes were in general reduced in abundance in HIV-infected individuals, though this pattern did not hold true for every family within this phylum. The phylum overall behaved in this manner, with decreased abundance in five of 16 studies [19,20▪,22▪,23▪▪,27▪] and increased abundance in two studies [30▪▪,33▪]. The Firmicutes phylum is quite diverse, but broadly can be characterized as associated with developed world individuals [113] as well as obesity and increased energy harvest from diet [137,138]. Within the Firmicutes, the class Clostridia, which was overall depleted in HIV infection with decreased abundance in eight of 16 studies [19,20▪,21▪▪,22▪,23▪▪,30▪▪,32▪▪,33▪], is characterized by taxa that often function in anti-inflammatory roles by producing butyrate and other short-chain fatty acids (SCFA) [107,139] and shifting T-cell differentiation toward regulatory T cells [140–143]. Within the class Clostridia, the family Lachnospiraceae, which was decreased in abundance in HIV-infected individuals in six of 16 studies [19,20▪,21▪▪,23▪▪,25▪,30▪▪], includes members that are commonly found to be uniquely effective metabolizers of complex polysaccharides [144,145] and characterized by the production of SCFA such as butyrate [146] and acetate [147] that are thought to be anti-inflammatory. Also within the class Clostridia, the family Peptostreptococcaceae varied in its HIV-associated shifts, with two of 16 studies [20▪,32▪▪] showing a relative decrease in abundance in HIV-infected individuals and one study [19] showing a relative increase. Peptostreptococcaceae have been found to function in mostly a proinflammatory role, with positive associations with C. difficile infection [121,148], viral diarrhea [149], intestinal inflammation [150], and the mucosal [151] and fecal [152] communities of individuals with colorectal cancer. In contrast to the family Peptostreptococcaceae, the Clostridia family Ruminococcaceae was in general decreased in HIV infection, with 10 of 16 studies [19,20▪,21▪▪,22▪,23▪▪,26▪,28▪▪–30▪▪,32▪▪] reporting decreased abundance in HIV-infected individuals and only one study [17,18▪▪] reporting increased abundance. Ruminococcaceae have been associated with both protective and disruptive roles within the gut microbial community, such as the production of anti-inflammatory SCFA [144] or the degradation of host mucus and potential proinflammatory role in IBD [153], and functional effects within this family have been found to be highly species dependent [132,154–156]. The bacterial family Erysipelotrichaceae, which is contained within the separate class Erysipelotrichia, was on the whole found to be increased in association with HIV infection with a greater abundance demonstrated in six of 16 studies [17,18▪▪,20▪,21▪▪,24▪,26▪,33▪] and decreased abundance in only one study [31▪▪]. Erysipelotrichaceae are described as adhesive and potentially pathogenic [157], and have been found to be positively associated with obesity [158,159] and luminal microbial communities in colorectal cancer [151,160]. Interestingly, this family is also found to be enriched in the enteric communities of the Hadza hunter-gatherers of Tanzania [157].
MICROBIOME DIFFERENCES AND SAMPLING SITE
Some of the variation in the findings among these studies may be attributed to the differences in body site sampled and sampling methodology. The predominant collection method was stool sampling [17,18▪▪,20▪,23▪▪,24▪–26▪,28▪▪–32▪▪,33▪,161▪–164▪] which is most representative of the luminal microbial community, though anal swabs and washings [19,22▪,165] or mucosal biopsies [20▪,21▪▪,23▪▪,27▪,28▪▪] were also used. Independent of HIV, mucosal and luminal microbial communities have been shown to differ to varying degrees [166–171], though in some contexts, the mucosal and luminal communities correlate strongly and are representative of one another [170]. With regard to functional differences, facultative anaerobes have been found to be more abundant in mucosal-associated environments, whereas obligate anaerobes are more prevalent in the gut lumen [170]. Studies of the HIV-associated microbiota that used both mucosal and luminal-targeted sampling techniques [20▪,23▪▪,28▪▪] found a variety of differences between these techniques. In the most concordant finding, the two sampling sites produced similar conclusions, with most taxa showing the same patterns of HIV-associated enrichment or depletion at both sampling sites (Fig. 1 and Table S1) [20▪]. However, the HIV-associated reduction in diversity was more pronounced in mucosa than stool samples. Other studies found greater variation in the HIV-associated differences by body site. For instance, Dillon et al.[28▪▪] detected an HIV-associated increase in abundance of Prevotellaceae at both sampling sites, but only observed HIV-associated decreases in taxa in the phyla Bacteroidetes and Firmicutes and increases in Proteobacteria in the mucosal samples. The study with the greatest discordance between mucosal and luminal findings observed many more HIV-associated differences in mucosal-associated communities than luminal communities [23▪▪]. In addition, mucosal community composition changes were more closely associated with mucosal cellular immune activation. Mucosal findings may be more sensitive or representative of the causative community, as these microorganisms are in the closest contact with epithelial cells and immune cells. These advantages, however, must be weighed against the relative difficulty of obtaining mucosal samples.
GEOGRAPHICAL CONTEXT FOR BACTERIAL MICROBIOTA DIFFERENCES
Most current work investigating HIV and the enteric microbiome has focused on populations in the developed world (Fig. 3), as opposed to the developing world where HIV burden is greatest [172]. As the burden of NCD in chronic HIV is growing rapidly in the HIV-infected population in sub-Saharan Africa [173–176], this region could potentially benefit from the deployment of microbiota-directed therapeutics. However, geographic differences in the gut microbiome may make it difficult to translate data derived from developed world subjects to target populations in the developing world.
FIGURE 3.
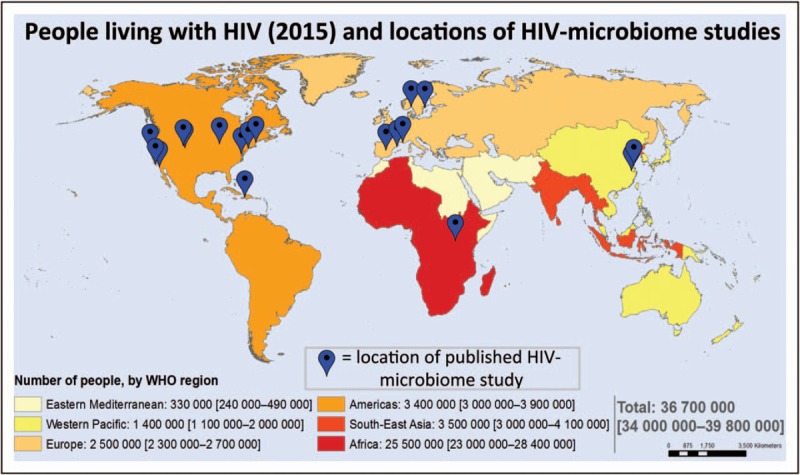
People living with HIV and locations of HIV-microbiome studies. Locations of HIV-microbiome cohorts overlaid on world map of HIV prevalence. The vast majority of HIV-microbiome studies have been conducted in developed world settings. Map source: Adults and children estimated to be living with HIV 2015, WHO/UNAIDS/UNICEF.
The microbiota differences observed between HIV-infected and uninfected individuals as a whole [17,18▪▪] mirror some differences seen at baseline between populations in the developing and developed world [113,115,117]. It is currently thought that the differences between developed and developing world microbiota are primarily a result of the corresponding dietary differences, with greater consumption of fat and simple carbohydrates in the developed world and greater consumption of more complex carbohydrates in the developing world [114,132,177]. The observations that a Prevotellaceae-rich community is frequently observed in healthy individuals in developing nations such as Burkina Faso, Venezuela, Malawi, or Papua New Guinea [17,113–117] argue against the simple conclusion that defined HIV-associated taxa changes in the gut microbiota (e.g., an increase in Prevotellaceae and a decrease in Firmicutes) alone are responsible for chronic inflammation and disorder in HIV-infected individuals. Rather, these potentially conflicting observations suggest a more complex relationship wherein a mismatch between the extant taxa and their host context causes inflammation [18▪▪]. One possible example of this mismatch might be a microbial community that is inappropriate for the host diet and as a result produces inflammatory metabolites from these dietary constituents. This ‘context-dependent’ model of microbial-driven systemic inflammation would further heighten the need to study HIV-associated microbial differences in appropriate populations, as therapeutic interventions constructed from data originating from one population may be poorly suited for application in a population with a different microbial and environmental milieu.
VIROME DIFFERENCES IN HIV INFECTION
The diverse human enteric virome includes a wide array of eukaryotic viruses, bacteriophages, and endogenous retroviruses [178–181] and is largely unexplored, with some estimates that only 1% of the virome has been sequenced [182]. To date, only a handful of studies have examined alterations in the virome during lentiviral infection [29▪▪,183–186]. New sequencing technologies offer a unique opportunity for unbiased examination of the virome including discovery of potentially novel unculturable viruses [187]. Enteric eukaryotic viruses can drive host pathology that directly causes gastroenteritis, enteritis, or colitis. Bacteriophages, which are the most abundant enteric viruses, can initiate changes in the bacterial community that influence gut health and may also be able to directly interact with the host immune system [178–190]. In IBD, diversity of the bacteriophage virome increases whereas diversity of the bacterial microbiome decreases, suggesting a potential competitive relationship between bacteria and bacteriophages during enteric inflammation [191]. In light of the emerging recognition of the potential role of the enteric virome in gastrointestinal pathology, there is now a growing focus on characterizing the gut virome associated with HIV infection.
Nonhuman primate simian immunodeficiency virus models of HIV infection have shown expansion of the enteric virome, with significant increases in Picornaviridae, Adenoviridae, and Parvoviridae abundance in pathologic simian immunodeficiency virus infection of rhesus macaques [183,184]. In this model, Adenoviridae were associated with development of severe gastrointestinal disease [183,184] and enteric viruses spread to other tissue compartments, with Parvoviridae detected in the serum and Adenoviridae found in the enteric mucosa [184].
HIV-associated differences in the virome have also been observed in HIV-infected individuals. In a Ugandan cohort, CD4+ T-cell counts less than 200 cells/μl were associated with a significant increase in enteric Adenoviridae and Anelloviridae[29▪▪]. Anelloviridae are small, nonenveloped viruses with circular negative-sense ssDNA genomes often found in human serum [192]. Anelloviridae have not yet been identified as causative disease agents, but have been shown to be increased in the serum of immunosuppressed solid organ transplant patients [193] and (not using modern sequencing methods) HIV-infected study participants [194,195]. Similar alterations to the plasma virome were observed in HIV-infected individuals in the United States and Uganda, with an increase in Anelloviridae[185]. In the United States but not the Ugandan cohort, there was also a significant increase in total plasma viral sequences, mainly attributed to bacteriophages, in HIV-infected individuals with low (< 20 cells/μl) CD4+ T-cell counts as compared with individuals with high (> 700 cells/μl) CD4+ T-cell counts [185].
CONCLUSION
The role of enteric microbial changes in HIV disease progression has been the focus of increasing investigation. Models regarding the connections between gut microbiome changes and chronic HIV pathogenesis hypothesize a role for gut epithelial damage and systemic immune activation as an intermediate mechanism. Investigations into HIV-associated differences in the gut microbial community have found varied changes, but a few overall consistent patterns have emerged. In general, HIV infection is associated with decreases in many bacterial families within the phyla Bacteroidetes and Firmicutes as well as increases in the family Prevotellaceae and families within the phyla Proteobacteria and Fusobacteria. This community shift implies an overall pathogenic or proinflammatory outcome based on the functions of the differentially abundant microorganisms, but this causative relationship has not been conclusively shown and bacterial behavior can vary widely based on context. The viral component of the gut microbial community has received relatively little attention, but findings have generally shown an increase in the family Anelloviridae, which is frequently found in immunocompromised hosts, and in potentially pathogenic viruses such as the family Adenoviridae.
The heterogeneity of the conclusions drawn to date within this field may be in part because of the variation in study populations and methods employed (Fig. 2 and Table S1). Greater coordination of methodologies would allow more robust analysis of multiple studies and potentially reconcile conflicting conclusions. There is also a need to more finely examine the taxa differentially abundant in HIV infection, as meaningful functional variation can often occur at the species or strain level rather than the family or genera level [196–198]. 16S rRNA gene sequencing cannot provide this taxonomic resolution, creating a role for techniques such as shotgun metagenomic sequencing, which could detect strain or gene content variation. In light of the current findings of HIV-associated Prevotellaceae enrichment and the great abundance of this taxa in developing world populations that constitute the majority of HIV-infected individuals, there is a dire need for further examination of HIV-associated gut microbial differences in developing world populations. It is possible that therapeutic strategies that consider Prevotellaceae enrichment a pathogenic state would be ill suited for HIV-infected individuals in the developing world. In addition, microbiota-targeted probiotic interventions have produced mixed results, with some therapeutics reducing inflammatory markers and others lacking efficacy [162▪,199–208]. Further understanding of the enteric microbial changes associated with HIV infection, especially among developing world populations that bear the greatest burden of HIV infection, is therefore necessary to design therapeutic strategies that could alleviate the sequelae of systemic inflammation and NCD in chronic HIV infection.
Acknowledgements
The authors would like to acknowledge Dr Herbert W. Virgin, Dr Scott Handley, and Dr Guoyan Zhao for informative discussions regarding the content of this review.
Financial support and sponsorship
Funding received from NIH (NIDDK and NIAID), and the Burroughs Wellcome Fund.
Conflicts of interest
There are no conflicts of interest.
REFERENCES AND RECOMMENDED READING
Papers of particular interest, published within the annual period of review, have been highlighted as:
▪ of special interest
▪▪ of outstanding interest
Supplementary Material
Supplementary Material
REFERENCES
- 1.Delwart E. A roadmap to the human virome. PLoS Pathog 2013; 9:e1003146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ley RE, Peterson DA, Gordon JI. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006; 124:837–848. [DOI] [PubMed] [Google Scholar]
- 3.Virgin HW, Todd JA. Metagenomics and personalized medicine. Cell 2011; 147:44–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Virgin HW, Wherry EJ, Ahmed R. Redefining chronic viral infection. Cell 2009; 138:30–50. [DOI] [PubMed] [Google Scholar]
- 5.Wylie KM, Weinstock GM, Storch GA. Emerging view of the human virome. Transl Res 2012; 160:283–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Honda K, Littman DR. The microbiome in infectious disease and inflammation. Annu Rev Immunol 2012; 30:759–795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cho I, Blaser MJ. The human microbiome: at the interface of health and disease. Nat Rev Genet 2012; 13:260–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Friedrich MJ. Genomes of microbes inhabiting the body offer clues to human health and disease. JAMA 2013; 309:1447–1449. [DOI] [PubMed] [Google Scholar]
- 9.Littman DR, Pamer EG. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011; 10:311–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science 2012; 336:1268–1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang H, Luo XM. Control of commensal microbiota by the adaptive immune system. Gut Microbes 2015; 6:156–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thaiss CA, Zmora N, Levy M, Elinav E. The microbiome and innate immunity. Nature 2016; 535:65–74. [DOI] [PubMed] [Google Scholar]
- 13.Okoye AA, Picker LJ. CD4(+) T-cell depletion in HIV infection: mechanisms of immunological failure. Immunol Rev 2013; 254:54–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Andrews CA, Koup RA. The immunopathology of HIV infection. J Antimicrob Chemother 1996; 37 Suppl B:13–25. [DOI] [PubMed] [Google Scholar]
- 15.Douek DC, Roederer M, Koup RA. Emerging concepts in the immunopathogenesis of AIDS. Annu Rev Med 2009; 60:471–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sandler NG, Douek DC. Microbial translocation in HIV infection: causes, consequences and treatment opportunities. Nat Rev Microbiol 2012; 10:655–666. [DOI] [PubMed] [Google Scholar]
- 17.Lozupone CA, Li M, Campbell TB, et al. Alterations in the gut microbiota associated with HIV-1 infection. Cell Host Microbe 2013; 14:329–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18▪▪.Lozupone CA, Rhodes ME, Neff CP, et al. HIV-induced alteration in gut microbiota: driving factors, consequences, and effects of antiretroviral therapy. Gut Microbes 2014; 5:562–570. [DOI] [PubMed] [Google Scholar]; The study observed increased alpha diversity and Prevotella/Bacteroides ratio in HIV. This study showed that alterations to the gut microbiota in HIV persist with ART and the microbiota of individuals on ART are more similar to that of untreated HIV-infected than uninfected individuals.
- 19.McHardy IH, Li X, Tong M, et al. HIV Infection is associated with compositional and functional shifts in the rectal mucosal microbiota. Microbiome 2013; 1:26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20▪.Mutlu EA, Keshavarzian A, Losurdo J, et al. A compositional look at the human gastrointestinal microbiome and immune activation parameters in HIV infected subjects. PLoS Pathog 2014; 10:e1003829. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study was the first to show that dysbiosis and reduced alpha diversity occur in the terminal ileum and colon in HIV infection. This study observed reduced alpha diversity and increased ratio of Prevotella/Bacteroides in the HIV-infected group.
- 21▪▪.Vujkovic-Cvijin I, Dunham RM, Iwai S, et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci Transl Med 2013; 5: 193ra91. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study revealed an increased ratio of Proteobacteria/Bacteroides in viremic, untreated HIV-infected study participants and an intermediate dysbiosis in the ART group. This study found that the degree of dysbiosis was strongly correlated with IP-10, the kynurenine pathway of tryptophan metabolism, and T-cell activation in both the colon and peripheral blood.
- 22▪.Yu G, Fadrosh D, Ma B, et al. Anal microbiota profiles in HIV-positive and HIV-negative MSM. AIDS 2014; 28:753–760. [DOI] [PubMed] [Google Scholar]; Longitudinal microbiome analyses in this study showed that alpha diversity decreases with advanced HIV infection.
- 23▪▪.Dillon SM, Lee EJ, Kotter CV, et al. An altered intestinal mucosal microbiome in HIV-1 infection is associated with mucosal and systemic immune activation and endotoxemia. Mucosal Immunol 2014; 7:983–994. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study demonstrated an increased ratio of Prevotella/Bacteroides in HIV-infected group, whereas other features of dysbiosis were inconsistent across sample types, such as mucosa-specific increases in Proteobacteria and alpha diversity. In this study, the degree of dysbiosis correlated with plasma LPS levels, peripheral blood CD4 and CD8 activation, and colonic T cell and myeloid dendritic cell activation.
- 24▪.Dinh DM, Volpe GE, Duffalo C, et al. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J Infect Dis 2015; 211:19–27. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this study, Gammaproteobacteria, Enterobacteriales, and Enterobacteriaceae abundance positively correlated with levels of markers of systemic immune activation such as sCD14, IFNγ, and IL-1β.
- 25▪.Nowak P, Troseid M, Avershina E, et al. Gut microbiota diversity predicts immune status in HIV-1 infection. AIDS (London, England) 2015; 29:2409–2418. [DOI] [PubMed] [Google Scholar]; The study suggested that Prevotella abundance and alpha diversity are further reduced following ART, and that alpha diversity inversely correlates with CD4 cell count and positively correlated with plasma LPS, LBP, sCD14, and sCD163. In this study, elite controllers were comparable to HIV-uninfected study participants at the phylum level, and had a higher abundance of Bacteroidetes than viremic study participants.
- 26▪.Vázquez-Castellanos JF, Serrano-Villar S, Latorre A, et al. Altered metabolism of gut microbiota contributes to chronic immune activation in HIV-infected individuals. Mucosal Immunol 2015; 8:760–772. [DOI] [PubMed] [Google Scholar]; The study revealed an increased ratio of Prevotella/Bacteroides and depletion of Coprococcus in the HIV-infected group. Shotgun metagenomic data in this study demonstrated functional differences in the microbiota of HIV-infected study participants, including enrichment in pathogenic processes, LPS biosynthesis, bacterial translocation, and other inflammatory pathways, as well as the depletion of amino acid metabolism and energy processes.
- 27▪.Yang L, Poles MA, Fisch GS, et al. HIV-induced immunosuppression is associated with colonization of the proximal gut by environmental bacteria. AIDS 2016; 30:19–29. [DOI] [PMC free article] [PubMed] [Google Scholar]; This was the first study to suggest that HIV impairs resistance of the duodenum to colonization of environmental bacteria such as Burkholderia fungorum and Bradyrhizobium pachyrhizi, whose abundance was inversely correlated with CD4 cell count but not viral load. The study also observed significant clustering of microbiota based on HIV phenotype in the duodenum but not in other compartments of the proximal gut, characterized by an enrichment of Proteobacteria and depletion of Firmicutes and Lactobacillus.
- 28▪▪.Dillon SM, Lee EJ, Kotter CV, et al. Gut dendritic cell activation links an altered colonic microbiome to mucosal and systemic T-cell activation in untreated HIV-1 infection. Mucosal Immunol 2016; 9:24–37. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study found that Prevotella positively correlated with colonic myeloid dendritic cell activation in vivo and induced proinflammatory cytokine production by colonic mDCs in vitro. These findings led them to propose a model describing myeloid dendritic cells and the gut microbiota as drivers of mucosal immune activation and chronic inflammation in HIV.
- 29▪▪.Monaco CL, Gootenberg DB, Zhao G, et al. Altered virome and bacterial icrobiome in human immunodeficiency virus-associated Acquired Immunodeficiency Syndrome. Cell Host Microbe 2016; 19:311–322. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this first study investigating the gut microbiota in HIV in sub-Saharan Africa, low CD4 T-cell levels correlated with alterations in gut bacterial communities independent of ART treatment, including reduced phylogenetic diversity and enrichment in Enterobacteriaceae. This was also the first study to examine the enteric virome in HIV disease progression, demonstrating an expansion of enteric adenovirus sequences in HIV-infected study participants with low CD4 cell counts.
- 30▪▪.Sun Y, Ma Y, Lin P, et al. Fecal bacterial microbiome diversity in chronic HIV-infected patients in China. Emerg Microbes Infect 2016; 5:e31. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this first study investigating the gut microbiota in HIV in an Asian cohort, infected individuals had decreased alpha diversity, were enriched for Firmicutes and Proteobacteria at the phylum level, and were depleted in the families Ruminococcaceae and Lachnospiraceae and the class Clostridia.
- 31▪▪.Bender JM, Li F, Martelly S, et al. Maternal HIV infection influences the microbiome of HIV-uninfected infants. Sci Transl Med 2016; 8: 349ra100. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study demonstrated that human breast milk oligosaccharides correlated with the abundance of specific gut bacteria in high-risk infants (increases in HIV-infected mothers’ N-fucopentaose-1 in breast milk correlated with increases in Bifidobacteriaceae in infant's stool). In this study, HIV exposed, uninfected infants had less diverse microbiota with greater abundances of Prevotella and Psuedomonas. Maternal CD4 cell count positively correlated with the diversity of infant gut microbiota. Trending but insignificant proportion of HIV exposed, uninfected infant stool microbiota resembled HIV-infected mother's breast millk and areolar skin, suggesting direct seeding of infant gut microbiome via breasfeeding in HIV-infected mothers. HIV-exposed infants had ‘younger/immature’ gut microbiota.
- 32▪▪.Dubourg G, Lagier JC, Hue S, et al. Gut microbiota associated with HIV infection is significantly enriched in bacteria tolerant to oxygen. BMJ Open Gastroenterol 2016; 3:e000080. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study showed that the microbiota in HIV-positive study participants are enrichment in aerotolerant taxa such as Enterobacteriaceae and Enterococcaceae, and depleted in anaerobic taxa such as Ruminococcus. Reduced diversity in HIV individuals not restored by ART. The increased ratio of aerotolerant to anaerobic taxa in HIV-infected study participants was inversely correlated with inflammation and immune activation markers (IP10, CD14, CD163).
- 33▪.Ling Z, Jin C, Xie T, et al. Alterations in the fecal microbiota of patients with HIV-1 infection: an observational study in a Chinese population. Sci Rep 2016; 6:30673. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this study, the ratio of Firmicutes/Bacteroidetes and Prevotella abundance increased in the HIV-infected group. Despite reduced viral loads, ART did not restore the diversity or composition of the gut microbiota.
- 34. Centers for Disease Control and Prevention: HIV Surveillance Report; 2008. [Google Scholar]
- 35.Egger M, May M, Chene G, et al. Prognosis of HIV-1-infected patients starting highly active antiretroviral therapy: a collaborative analysis of prospective studies. Lancet 2002; 360:119–129. [DOI] [PubMed] [Google Scholar]
- 36.Guaraldi G, Orlando G, Zona S, et al. Premature age-related comorbidities among HIV-infected persons compared with the general population. Clin Infect Dis 2011; 53:1120–1126. [DOI] [PubMed] [Google Scholar]
- 37.Lewden C, Chene G, Morlat P, et al. HIV-infected adults with a CD4 cell count greater than 500 cells/mm3 on long-term combination antiretroviral therapy reach same mortality rates as the general population. J Acquir Immune Defic Syndr 2007; 46:72–77. [DOI] [PubMed] [Google Scholar]
- 38.Lohse N, Hansen A-BE, Pedersen G, et al. Survival of persons with and without HIV infection in Denmark, 1995–2005. Ann Intern Med 2007; 146:87–95. [DOI] [PubMed] [Google Scholar]
- 39.Reid MJA, Mosepele M, Tsima BM, Gross R. Addressing the challenge of the emerging NCD epidemic: lessons learned from Botswana's response to the HIV epidemic. Public Health Action 2012; 2:47–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Muyanja D, Muzoora C, Muyingo A, et al. High prevalence of metabolic syndrome and cardiovascular disease risk among people with HIV on stable ART in Southwestern Uganda. AIDS Patient Care STDS 2015; 30:4–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Okello S, Kanyesigye M, Muyindike WR, et al. Incidence and predictors of hypertension in adults with HIV-initiating antiretroviral therapy in south-western Uganda. J Hypertens 2015; 33:2039–2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Siedner MJ, Kim J-H, Nakku RS, et al. Persistent immune activation and carotid atherosclerosis in HIV-infected Ugandans receiving antiretroviral therapy. J Infect Dis 2016; 213:370–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chow FC, Regan S, Feske S, et al. Comparison of ischemic stroke incidence in HIV-infected and non-HIV-infected patients in a US healthcare system. J Acquir Immune Defic Syndr 2012; 60:351–358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Freiberg MS, Chang C-CH, Kuller LH, et al. HIV infection and the risk of acute myocardial infarction. JAMA Intern Med 2013; 173:614–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Marcus JL, Leyden WA, Chao CR, et al. HIV infection and incidence of ischemic stroke. AIDS 2014; 28:1911–1919. [DOI] [PubMed] [Google Scholar]
- 46.Duprez DA, Neuhaus J, Kuller LH, et al. Inflammation, coagulation and cardiovascular disease in HIV-infected individuals. PLoS One 2012; 7:e44454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McKibben RA, Margolick JB, Grinspoon S, et al. Elevated levels of monocyte activation markers are associated with subclinical atherosclerosis in men with and those without HIV infection. J Infect Dis 2015; 211:1219–1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ross AC, Rizk N, O’Riordan MA, et al. McComsey Ga: Relationship between inflammatory markers, endothelial activation markers, and carotid intima-media thickness in HIV-infected patients receiving antiretroviral therapy. Clin Infect Dis 2009; 49:1119–1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Subramanian S, Tawakol A, Burdo TH, et al. Arterial inflammation in patients with HIV. JAMA 2012; 308:379–386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Triant VA, Grinspoon SK. Immune dysregulation and vascular risk in HIV-infected patients: implications for clinical care. J Infect Dis 2011; 203:439–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Triant VA, Meigs JB, Grinspoon SK. Association of C-reactive protein and HIV infection with acute myocardial infarction. J Acquir Immune Defic Syndr 2009; 51:268–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fisher EA, Feig JE, Hewing B, et al. High-density lipoprotein function, dysfunction, and reverse cholesterol transport. Arterioscler Thromb Vasc Biol 2012; 32:2813–2820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Khovidhunkit W, Kim M-S, Memon RA, et al. Effects of infection and inflammation on lipid and lipoprotein metabolism: mechanisms and consequences to the host. J Lipid Res 2004; 45:1169–1196. [DOI] [PubMed] [Google Scholar]
- 54.Neves AL, Coelho J, Couto L, et al. Metabolic endotoxemia: a molecular link between obesity and cardiovascular risk. J Mol Endocrinol 2013; 51:R51–R64. [DOI] [PubMed] [Google Scholar]
- 55.Yuan H, Zelkha S, Zelka S, et al. Pivotal role of NOD2 in inflammatory processes affecting atherosclerosis and periodontal bone loss. Proc Natl Acad Sci U S A 2013; 110:E5059–E5068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hunt PW. HIV and inflammation: mechanisms and consequences. Curr HIV/AIDS Rep 2012; 9:139–147. [DOI] [PubMed] [Google Scholar]
- 57.Tien PC, Choi AI, Zolopa AR, et al. Inflammation and mortality in HIV-infected adults: analysis of the FRAM study cohort. J Acquir Immune Defic Syndr 2010; 55:316–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Gandhi RT, Spritzler J, Chan E, et al. Effect of baseline- and treatment-related factors on immunologic recovery after initiation of antiretroviral therapy in HIV-1-positive subjects: results from ACTG 384. J Acquir Immune Defic Syndr 2006; 42:426–434. [DOI] [PubMed] [Google Scholar]
- 59.Goicoechea M, Smith DM, Liu L, et al. Determinants of CD4+ T cell recovery during suppressive antiretroviral therapy: association of immune activation, T cell maturation markers, and cellular HIV-1 DNA. J Infect Dis 2006; 194:29–37. [DOI] [PubMed] [Google Scholar]
- 60.Hunt PW, Martin JN, Sinclair E, et al. T cell activation is associated with lower CD4+ T cell gains in human immunodeficiency virus-infected patients with sustained viral suppression during antiretroviral therapy. J Infect Dis 2003; 187:1534–1543. [DOI] [PubMed] [Google Scholar]
- 61.Hunt Peter W, Brenchley J, Sinclair E, et al. Relationship between T cell activation and CD4 + T cell count in HIV-seropositive individuals with undetectable plasma HIV RNA levels in the absence of therapy. J Infect Dis 2008; 197:126–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hunt PW, Cao HL, Muzoora C, et al. Impact of CD8+ T-cell activation on CD4+ T-cell recovery and mortality in HIV-infected Ugandans initiating antiretroviral therapy. AIDS 2011; 25:2123–2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol 2012; 30:149–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Brenchley JM, Price Da, Schacker TW, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med 2006; 12:1365–1371. [DOI] [PubMed] [Google Scholar]
- 65.Jenabian MA, El-Far M, Vyboh K, et al. Immunosuppressive tryptophan catabolism and gut mucosal dysfunction following early HIV infection. J Infect Dis 2015; 212:355–366. [DOI] [PubMed] [Google Scholar]
- 66.Hunt PW, Sinclair E, Rodriguez B, et al. Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J Infect Dis 2014; 210:1228–1238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Brown JM, Hazen SL. The gut microbial endocrine organ: bacterially derived signals driving cardiometabolic diseases. Annu Rev Med 2015; 66:343–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Koeth RA, Levison BS, Culley MK, et al. γ-butyrobetaine is a proatherogenic intermediate in gut microbial metabolism of L-carnitine to TMAO. Cell Metab 2014; 20:799–812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Koeth RA, Wang Z, Levison BS, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 2013; 19:576–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011; 472:57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zhu W, Gregory JC, Org E, et al. Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell 2016; 165:111–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Brenchley JM, Schacker TW, Ruff LE, et al. CD4+ T cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J Exp Med 2004; 200:749–759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Clayton F, Snow G, Reka S, Kotler DP. Selective depletion of rectal lamina propria rather than lymphoid aggregate CD4 lymphocytes in HIV infection. Clin Exp Immunol 1997; 107:288–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Heise C, Miller CJ, Lackner A, Dandekar S. Primary acute simian immunodeficiency virus infection of intestinal lymphoid tissue is associated with gastrointestinal dysfunction. J Infect Dis 1994; 169:1116–1120. [DOI] [PubMed] [Google Scholar]
- 75.Li Q, Duan L, Estes JD, et al. Peak SIV replication in resting memory CD4+ T cells depletes gut lamina propria CD4+ T cells. Nature 2005; 434:1148–1152. [DOI] [PubMed] [Google Scholar]
- 76.Mattapallil JJ, Douek DC, Hill B, et al. Massive infection and loss of memory CD4+ T cells in multiple tissues during acute SIV infection. Nature 2005; 434:1093–1097. [DOI] [PubMed] [Google Scholar]
- 77.Mehandru S, Poles MA, Tenner-Racz K, et al. Primary HIV-1 infection is associated with preferential depletion of CD4+ T lymphocytes from effector sites in the gastrointestinal tract. J Exp Med 2004; 200:761–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Schneider T, Ullrich R, Zeitz M. The immunologic aspects of human immunodeficiency virus infection in the gastrointestinal tract. Semin Gastrointest Dis 1996; 7:19–29. [PubMed] [Google Scholar]
- 79.Veazey RS, DeMaria M, Chalifoux LV, et al. Gastrointestinal tract as a major site of CD4+ T cell depletion and viral replication in SIV infection. Science 1998; 280:427–431. [DOI] [PubMed] [Google Scholar]
- 80.Schuetz A, Deleage C, Sereti I, et al. Initiation of ART during early acute HIV infection preserves mucosal Th17 function and reverses HIV-related immune activation. PLoS Pathog 2014; 10:e1004543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Maloy KJ, Kullberg MC. IL-23 and Th17 cytokines in intestinal homeostasis. Mucosal Immunol 2008; 1:339–349. [DOI] [PubMed] [Google Scholar]
- 82.Torres B, Guardo AC, Leal L, et al. Protease inhibitor monotherapy is associated with a higher level of monocyte activation, bacterial translocation and inflammation. J Int AIDS Soc 2014; 17:19246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007; 56:1761–1772. [DOI] [PubMed] [Google Scholar]
- 84.Marchetti G, Tincati C, Silvestri G. Microbial translocation in the pathogenesis of HIV infection and AIDS. Clin Microbiol Rev 2013; 26:2–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Cassol E, Malfeld S, Mahasha P, et al. Persistent microbial translocation and immune activation in HIV-1-infected South Africans receiving combination antiretroviral therapy. J Infect Dis 2010; 202:723–733. [DOI] [PubMed] [Google Scholar]
- 86.Sandler NG, Wand H, Roque A, et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis 2011; 203:780–790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kostic AD, Xavier RJ, Gevers D. The microbiome in inflammatory bowel disease: current status and the future ahead. Gastroenterology 2014; 146:1489–1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Willing BP, Dicksved J, Halfvarson J, et al. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 2010; 139:1844–1854. e1841. [DOI] [PubMed] [Google Scholar]
- 89.Scher JU, Ubeda C, Artacho A, et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol 2015; 67:128–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kostic AD, Gevers D, Siljander H, et al. The dynamics of the human infant gut microbiome in development and in progression toward type 1 diabetes. Cell Host Microbe 2015; 17:260–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Sze MA, Schloss PD. Looking for a signal in the noise: revisiting obesity and the microbiome. MBio 2016; 7:e01018-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Weingarden A, Gonzalez A, Vazquez-Baeza Y, et al. Dynamic changes in short- and long-term bacterial composition following fecal microbiota transplantation for recurrent Clostridium difficile infection. Microbiome 2015; 3:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Chang JY, Antonopoulos DA, Kalra A, et al. Decreased diversity of the fecal Microbiome in recurrent Clostridium difficile-associated diarrhea. J Infect Dis 2008; 197:435–438. [DOI] [PubMed] [Google Scholar]
- 94.Ellis CL, Ma ZM, Mann SK, et al. Molecular characterization of stool microbiota in HIV-infected subjects by panbacterial and order-level 16S ribosomal DNA (rDNA) quantification and correlations with immune activation. J Acquir Immune Defic Syndr 2011; 57:363–370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Gori A, Tincati C, Rizzardini G, et al. Early impairment of gut function and gut flora supporting a role for alteration of gastrointestinal mucosa in human immunodeficiency virus pathogenesis. J Clin Microbiol 2008; 46:757–758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Zevin AS, McKinnon L, Burgener A, Klatt NR. Microbial translocation and microbiome dysbiosis in HIV-associated immune activation. Curr Opin HIV AIDS 2016; 11:182–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.de Bentzmann S, Plesiat P. The Pseudomonas aeruginosa opportunistic pathogen and human infections. Environ Microbiol 2011; 13:1655–1665. [DOI] [PubMed] [Google Scholar]
- 98.McGuckin MA, Linden SK, Sutton P, Florin TH. Mucin dynamics and enteric pathogens. Nat Rev Microbiol 2011; 9:265–278. [DOI] [PubMed] [Google Scholar]
- 99.Rey FE, Gonzalez MD, Cheng J, et al. Metabolic niche of a prominent sulfate-reducing human gut bacterium. Proc Natl Acad Sci U S A 2013; 110:13582–13587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Peleg AY, de Breij A, Adams MD, et al. The success of acinetobacter species; genetic, metabolic and virulence attributes. PLoS One 2012; 7:e46984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.March C, Regueiro V, Llobet E, et al. Dissection of host cell signal transduction during Acinetobacter baumannii-triggered inflammatory response. PLoS One 2010; 5:e10033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Knapp S, Wieland CW, Florquin S, et al. Differential roles of CD14 and toll-like receptors 4 and 2 in murine Acinetobacter pneumonia. Am J Respir Crit Care Med 2006; 173:122–129. [DOI] [PubMed] [Google Scholar]
- 103.Walker RI, Caldwell MB, Lee EC, et al. Pathophysiology of Campylobacter enteritis. Microbiol Rev 1986; 50:81–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Bereswill S, Fischer A, Plickert R, et al. Novel murine infection models provide deep insights into the ‘menage a trois’ of Campylobacter jejuni, microbiota and host innate immunity. PLoS One 2011; 6:e20953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Guentzel MN: Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacter, and Proteus. In Medical Microbiology. 4th ed Edited by Baron S; 1996. [PubMed] [Google Scholar]
- 106.Lupp C, Robertson ML, Wickham ME, et al. Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2007; 2:119–129. [DOI] [PubMed] [Google Scholar]
- 107.Nicholson JK, Holmes E, Kinross J, et al. Host-gut microbiota metabolic interactions. Science 2012; 336:1262–1267. [DOI] [PubMed] [Google Scholar]
- 108.Kostic AD, Chun E, Robertson L, et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013; 14:207–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Shaw KA, Bertha M, Hofmekler T, et al. Dysbiosis, inflammation, and response to treatment: a longitudinal study of pediatric subjects with newly diagnosed inflammatory bowel disease. Genome Med 2016; 8:75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Sinha R, Ahn J, Sampson JN, et al. Fecal microbiota, fecal metabolome, and colorectal cancer interrelations. PLoS One 2016; 11:e0152126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lukens JR, Gurung P, Vogel P, et al. Dietary modulation of the microbiome affects autoinflammatory disease. Nature 2014; 516:246–249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Scher JU, Sczesnak A, Longman RS, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife 2013; 2:e01202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.De Filippo C, Cavalieri D, Di Paola M, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A 2010; 107:14691–14696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Wu GD, Chen J, Hoffmann C, et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011; 334:105–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Yatsunenko T, Rey FE, Manary MJ, et al. Human gut microbiome viewed across age and geography. Nature 2012; 486:222–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Martinez I, Stegen JC, Maldonado-Gomez MX, et al. The gut microbiota of rural papua new guineans: composition, diversity patterns, and ecological processes. Cell Rep 2015; 11:527–538. [DOI] [PubMed] [Google Scholar]
- 117.Gorvitovskaia A, Holmes SP, Huse SM. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016; 4:15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Vogtmann E, Hua X, Zeller G, et al. Colorectal cancer and the human gut microbiome: reproducibility with whole-genome shotgun sequencing. PLoS One 2016; 11:e0155362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zackular JP, Baxter NT, Iverson KD, et al. The gut microbiome modulates colon tumorigenesis. MBio 2013; 4:e00692–e00613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Schubert AM, Sinani H, Schloss PD. Antibiotic-induced alterations of the murine gut microbiota and subsequent effects on colonization resistance against Clostridium difficile. MBio 2015; 6:e00974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Milani C, Ticinesi A, Gerritsen J, et al. Gut microbiota composition and Clostridium difficile infection in hospitalized elderly individuals: a metagenomic study. Sci Rep 2016; 6:25945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Schubert AM, Rogers MA, Ring C, et al. Microbiome data distinguish patients with Clostridium difficile infection and non-C. difficile-associated diarrhea from healthy controls. MBio 2014; 5: e01021–e01014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Ferreira RB, Gill N, Willing BP, et al. The intestinal microbiota plays a role in Salmonella-induced colitis independent of pathogen colonization. PLoS One 2011; 6:e20338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Ubeda C, Bucci V, Caballero S, et al. Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect Immun 2013; 81:965–973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Bailey MT, Dowd SE, Parry NM, et al. Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium. Infect Immun 2010; 78:1509–1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wlodarska M, Willing B, Keeney KM, et al. Antibiotic treatment alters the colonic mucus layer and predisposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect Immun 2011; 79:1536–1545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mazmanian SK, Round JL, Kasper DL. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008; 453:620–625. [DOI] [PubMed] [Google Scholar]
- 128.Troy EB, Kasper DL. Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front Biosci (Landmark Ed) 2010; 15:25–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci U S A 2010; 107:12204–12209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Gauffin Cano P, Santacruz A, Moya A, Sanz Y. Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. PLoS One 2012; 7:e41079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Vatanen T, Kostic AD, d’Hennezel E, et al. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell 2016; 165:842–853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.David LA, Maurice CF, Carmody RN, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014; 505:559–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Khanna S, Montassier E, Schmidt B, et al. Gut microbiome predictors of treatment response and recurrence in primary Clostridium difficile infection. Aliment Pharmacol Ther 2016; 44:715–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Harris K, Kassis A, Major G, Chou CJ. Is the gut microbiota a new factor contributing to obesity and its metabolic disorders? J Obes 2012; 2012:879151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Candon S, Perez-Arroyo A, Marquet C, et al. Antibiotics in early life alter the gut microbiome and increase disease incidence in a spontaneous mouse model of autoimmune insulin-dependent diabetes. PLoS One 2015; 10:e0125448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Qin J, Li Y, Cai Z, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012; 490:55–60. [DOI] [PubMed] [Google Scholar]
- 137.Ley R, Turnbaugh P, Klein S, Gordon J. Microbial ecology: human gut microbes associated with obesity. Nature 2006; 444:1022–1023. [DOI] [PubMed] [Google Scholar]
- 138.Turnbaugh PJ, Ley RE, Mahowald MA, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006; 444:1027–1031. [DOI] [PubMed] [Google Scholar]
- 139.Louis P, Flint HJ. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol Lett 2009; 294:1–8. [DOI] [PubMed] [Google Scholar]
- 140.Furusawa Y, Obata Y, Fukuda S, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013; 504:446–450. [DOI] [PubMed] [Google Scholar]
- 141.Atarashi K, Tanoue T, Oshima K, et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013; 500:232–236. [DOI] [PubMed] [Google Scholar]
- 142.Atarashi K, Tanoue T, Shima T, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011; 331:337–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Kunisawa J, Kiyono H. Peaceful mutualism in the gut: revealing key commensal bacteria for the creation and maintenance of immunological homeostasis. Cell Host Microbe 2011; 9:83–84. [DOI] [PubMed] [Google Scholar]
- 144.Flint HJ, Bayer EA, Rincon MT, et al. Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat Rev Microbiol 2008; 6:121–131. [DOI] [PubMed] [Google Scholar]
- 145.Biddle A, Stewart L, Blanchard J, Leschine S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013; 5:627. [Google Scholar]
- 146.Meehan CJ, Beiko RG. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol Evol 2014; 6:703–713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Rey FE, Faith JJ, Bain J, et al. Dissecting the in vivo metabolic potential of two human gut acetogens. J Biol Chem 2010; 285:22082–22090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Sangster W, Hegarty JP, Schieffer KM, et al. Bacterial and fungal microbiota changes distinguish C. difficile infection from other forms of diarrhea: results of a prospective inpatient study. Front Microbiol 2016; 7:789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ma C, Wu X, Nawaz M, et al. Molecular characterization of fecal microbiota in patients with viral diarrhea. Curr Microbiol 2011; 63:259–266. [DOI] [PubMed] [Google Scholar]
- 150.Tannock GW, Lawley B, Munro K, et al. Comprehensive analysis of the bacterial content of stool from patients with chronic pouchitis, normal pouches, or familial adenomatous polyposis pouches. Inflamm Bowel Dis 2012; 18:925–934. [DOI] [PubMed] [Google Scholar]
- 151.Chen W, Liu F, Ling Z, et al. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PloS one 2012; 7:e39743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Wang T, Cai G, Qiu Y, et al. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J 2012; 6:320–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Png CW, Lindén SK, Gilshenan KS, et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am J Gastroenterol 2010; 105:2420–2428. [DOI] [PubMed] [Google Scholar]
- 154.Ze X, Duncan SH, Louis P, Flint HJ. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J 2012; 6:1535–1543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Crost EH, Tailford LE, Le Gall G, et al. Utilisation of mucin glycans by the human gut symbiont Ruminococcus gnavus is strain-dependent. PLoS One 2013; 8:e76341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Hsiao A, Ahmed AM, Subramanian S, et al. Members of the human gut microbiota involved in recovery from Vibrio cholerae infection. Nature 2014; 515:423–426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Turroni S, Rampelli S, Centanni M, et al. Enterocyte-associated microbiome of the Hadza Hunter-Gatherers. Front Microbiol 2016; 7:865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Zietak M, Kovatcheva-Datchary P, Markiewicz LH, et al. Altered microbiota contributes to reduced diet-induced obesity upon cold exposure. Cell Metab 2016; 23:1216–1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Greiner T, Backhed F. Effects of the gut microbiota on obesity and glucose homeostasis. Trends Endocrinol Metab 2011; 22:117–123. [DOI] [PubMed] [Google Scholar]
- 160.Kaakoush NO. Insights into the role of Erysipelotrichaceae in the human host. Front Cell Infect Microbiol 2015; 5:84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161▪.Noguera-Julian M, Rocafort M, Guillén Y, et al. Gut microbiota linked to sexual preference and HIV infection. EBioMedicine 2016. 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this study, dysbiosis was observed only when grouping based on sexual behavior, suggesting that future HIV gut microbiome studies should control for HIV risk factors such as MSM. The MSM group was more diverse and enriched for Prevotella, whereas the non-MSM group was enriched for Bacteroides.
- 162▪.Stiksrud B, Nowak P, Nwosu FC, et al. Reduced levels of D-dimer and changes in gut microbiota composition after probiotic intervention in HIV-infected individuals on stable ART. J Acquir Immune Defic Syndr 2015; 70:329–337. [DOI] [PubMed] [Google Scholar]; The study performed an interventional probiotic trial and demonstrated that the HIV-infected individuals receiving the probiotic had an increased ratio of Lactobacilli and Bifidobacteria/Bacteroides, which correlated with lower levels of plasma LPS. Reductions in D-dimer, IL-6, and C-reactive protein after 8 weeks of probiotic intervention in HIV patients on ART. Probiotic intervention had no impact on microbial translocation or T-cell activation.
- 163▪.Serrano-Villar S, Rojo D, Martinez-Martinez M, et al. Gut bacteria metabolism impacts immune recovery in HIV-infected individuals. EBioMedicine 2016; 8:203–216. [DOI] [PMC free article] [PubMed] [Google Scholar]; The study demonstrated that gut bacteria metabolomic profiles cluster based on CD4 cell count, but that HIV-infected study participants with high CD4 T-cell concentrations did not cluster with HIV-uninfected study participants. In this study, proteomic phylogenetic analyses revealed specific proteins characteristic of bacterial taxa that may serve as active biomarkers of different clinical HIV phenotypes. Enrichment in proteins from Acidaminococcaceae family were only active biomarkers of HIV infection.
- 164▪.Serrano-Villar S, Rojo D, Martinez-Martinez M, et al. HIV infection results in metabolic alterations in the gut microbiota different from those induced by other diseases. Sci Rep 2016; 6:26192. [DOI] [PMC free article] [PubMed] [Google Scholar]; Using microbial metabolomics, this study showed that the gut microbiota in HIV has an impaired ability to synthesize Pro, Phe, and Lys, a finding that may partially explain the nutritional deficiency of wasting syndrome seen in advanced HIV. This study also showed that 3-hydroxyanthranilate, a product of the kynurenine pathway during tryptophan catabolism, is increased in the gut microbiota of HIV patients.
- 165.Pérez-Santiago J, Gianella S, Massanella M, et al. Gut Lactobacillales are associated with higher CD4 and less microbial translocation during HIV infection. AIDS 2013; 27:1921–1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Bajaj JS, Hylemon PB, Ridlon JM, et al. Colonic mucosal microbiome differs from stool microbiome in cirrhosis and hepatic encephalopathy and is linked to cognition and inflammation. Am J Physiol Gastrointest Liver Physiol 2012; 303:G675–G685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Eckburg PB, Bik EM, Bernstein CN, et al. Diversity of the human intestinal microbial flora. Science 2005; 308:1635–1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Morgan XC, Tickle TL, Sokol H, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol 2012; 13:R79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Stearns JC, Lynch MD, Senadheera DB, et al. Bacterial biogeography of the human digestive tract. Sci Rep 2011; 1:170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Yasuda K, Oh K, Ren B, et al. Biogeography of the intestinal mucosal and lumenal microbiome in the rhesus macaque. Cell Host Microbe 2015; 17:385–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Zoetendal EG, von Wright A, Vilpponen-Salmela T, et al. Mucosa-associated bacteria in the human gastrointestinal tract are uniformly distributed along the colon and differ from the community recovered from feces. Appl Environ Microbiol 2002; 68:3401–3407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. UNAIDS. GLOBAL REPORT: UNAIDS report on the global AIDS epidemic 2013. UNAIDS; 2013. [Google Scholar]
- 173.Hirschhorn LR, Kaaya SF, Garrity PS, et al. Cancer and the ’other’ noncommunicable chronic diseases in older people living with HIV/AIDS in resource-limited settings: a challenge to success. AIDS 2012; 26 Suppl 1:S65–S75. [DOI] [PubMed] [Google Scholar]
- 174.Hontelez JAC, de Vlas SJ, Baltussen R, et al. The impact of antiretroviral treatment on the age composition of the HIV epidemic in sub-Saharan Africa. AIDS 2012; 26 Suppl 1:S19–S30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Justice AC, Braithwaite RS. Lessons learned from the first wave of aging with HIV. AIDS 2012; 26 Suppl 1:S11–S18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Mills EJ, Bärnighausen T, Negin J. HIV and aging: preparing for the challenges ahead. N Engl J Med 2012; 366:1270–1273. [DOI] [PubMed] [Google Scholar]
- 177.Kovatcheva-Datchary P, Nilsson A, Akrami R, et al. Dietary fiber-induced improvement in glucose metabolism is associated with increased abundance of Prevotella. Cell Metab 2015; 22:971–982. [DOI] [PubMed] [Google Scholar]
- 178.Virgin HW. The virome in mammalian physiology and disease. Cell 2014; 157:142–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Cadwell K. Expanding the role of the virome: commensalism in the gut. J Virol 2015; 89:1951–1953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Norman JM, Handley SA, Virgin HW. Kingdom-agnostic metagenomics and the importance of complete characterization of enteric microbial communities. Gastroenterology 2014; 146:1459–1469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Ogilvie LA, Jones BV. The human gut virome: a multifaceted majority. Front Microbiol 2015; 6:918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Mokili JL, Rohwer F, Dutilh BE. Metagenomics and future perspectives in virus discovery. Curr Opin Virol 2012; 2:63–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Handley SA, Desai C, Zhao G, et al. SIV infection-mediated changes in gastrointestinal bacterial microbiome and virome are associated with immunodeficiency and prevented by vaccination. Cell Host Microbe 2016; 19:323–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Handley SA, Thackray LB, Zhao G, et al. Pathogenic simian immunodeficiency virus infection is associated with expansion of the enteric virome. Cell 2012; 151:253–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Li L, Deng X, Linsuwanon P, et al. AIDS alters the commensal plasma virome. J Virol 2013; 87:10912–10915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Li S-K, Leung RK-K, Guo H-X, et al. Detection and identification of plasma bacterial and viral elements in HIV/AIDS patients in comparison to healthy adults. Clin Microbiol Infect 2012; 18:1126–1133. [DOI] [PubMed] [Google Scholar]
- 187.Zhao G, Krishnamurthy S, Cai Z, et al. Identification of novel viruses using VirusHunter: an automated data analysis pipeline. PloS One 2013; 8:e78470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Duerkop BA, Hooper LV. Resident viruses and their interactions with the immune system. Nat Immunol 2013; 14:654–659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Focà A, Liberto MC, Quirino A, et al. Gut inflammation and immunity: what is the role of the human gut virome? Mediators Inflamm 2015; 2015:326032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Reyes A, Wu M, McNulty NP, et al. Gnotobiotic mouse model of phage-bacterial host dynamics in the human gut. Proc Natl Acad Sci U S A 2013; 110:20236–20241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Norman JM, Handley SA, Baldridge MT. Disease-specific alterations in the enteric virome in inflammatory bowel disease. Cell 2015; 160:447–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Bendinelli M, Maggi F. Tidona C, Darai G. Anellovirus. The Springer Index of Viruses. New York: Springer; 2011. 1963–1973. [Google Scholar]
- 193.De Vlaminck I, Khush KK, Strehl C, et al. Temporal response of the human virome to immunosuppression and antiviral therapy. Cell 2013; 155:1178–1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Christensen JK, Eugen-Olsen J, SŁrensen M, et al. Prevalence and prognostic significance of infection with TT virus in patients infected with human immunodeficiency virus. J Infect Dis 2000; 181:1796–1799. [DOI] [PubMed] [Google Scholar]
- 195.Debiaggi M, Zara F, Sacchi P, et al. Transfusion-transmitted virus infection in HIV-1-seropositive patients. Clin Microbiol Infect 2000; 6:246–250. [DOI] [PubMed] [Google Scholar]
- 196.Ley RE. Gut microbiota in 2015: Prevotella in the gut: choose carefully. Nat Rev Gastroenterol Hepatol 2016; 13:69–70. [DOI] [PubMed] [Google Scholar]
- 197.Ahern PP, Faith JJ, Gordon JI. Mining the human gut microbiota for effector strains that shape the immune system. Immunity 2014; 40:815–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Faith JJ, Colombel JF, Gordon JI. Identifying strains that contribute to complex diseases through the study of microbial inheritance. Proc Natl Acad Sci U S A 2015; 112:633–640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Cunningham-Rundles S, Ahrne S, Johann-Liang R, et al. Effect of probiotic bacteria on microbial host defense, growth, and immune function in human immunodeficiency virus type-1 infection. Nutrients 2011; 3:1042–1070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.d’Ettorre G, Ceccarelli G, Giustini N, et al. Probiotics reduce inflammation in antiretroviral treated, HIV-infected individuals: results of the ‘Probio-HIV’ Clinical Trial. PLoS One 2015; 10:e0137200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.González-Hernández LA, Jave-Suarez LF, Fafutis-Morris M, et al. Synbiotic therapy decreases microbial translocation and inflammation and improves immunological status in HIV-infected patients: a double-blind randomized controlled pilot trial. Nutr J 2012; 11:90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Hummelen R, Hemsworth J, Changalucha J, et al. Effect of micronutrient and probiotic fortified yogurt on immune-function of antiretroviral therapy naive HIV patients. Nutrients 2011; 3:897–909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Irvine SL, Hummelen R, Hekmat S, et al. Probiotic yogurt consumption is associated with an increase of CD4 count among people living with HIV/AIDS. J Clin Gastroenterol 2010; 44:e201–e205. [DOI] [PubMed] [Google Scholar]
- 204.Miller H, Ferris R, Phelps BR. The effect of probiotics on CD4 counts among people living with HIV: a systematic review. Benef Microbes 2016; 7:345–351. [DOI] [PubMed] [Google Scholar]
- 205.Monachese M, Cunningham-Rundles S, Diaz MA, et al. Probiotics and prebiotics to combat enteric infections and HIV in the developing world: a consensus report. Gut Microbes 2011; 2:198–207. [DOI] [PubMed] [Google Scholar]
- 206.Reid G. The potential role for probiotic yogurt for people living with HIV/AIDS. Gut Microbes 2010; 1:411–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Schunter M, Chu H, Hayes TL, et al. Randomized pilot trial of a synbiotic dietary supplement in chronic HIV-1 infection. BMC Complement Altern Med 2012; 12:84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Villar-Garcia J, Hernandez JJ, Guerri-Fernandez R, et al. Effect of probiotics (Saccharomyces boulardii) on microbial translocation and inflammation in HIV-treated patients: a double-blind, randomized, placebo-controlled trial. J Acquir Immune Defic Syndr 2015; 68:256–263. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.