

Abstract
Sequence divergence of orthologous proteins enables adaptation to environmental stresses and promotes evolution of novel functions. Limits on evolution imposed by constraints on sequence and structure were explored using a model TIM barrel protein, indole-3-glycerol phosphate synthase (IGPS). Fitness effects of point mutations in three phylogenetically divergent IGPS proteins during adaptation to temperature stress were probed by auxotrophic complementation of yeast with prokaryotic, thermophilic IGPS. Analysis of beneficial mutations pointed to an unexpected, long-range allosteric pathway towards the active site of the protein. Significant correlations between the fitness landscapes of distant orthologues implicate both sequence and structure as primary forces in defining the TIM barrel fitness landscape and suggest that fitness landscapes can be translocated in sequence space. Exploration of fitness landscapes in the context of a protein fold provides a strategy for elucidating the sequence-structure-fitness relationships in other common motifs.
The TIM barrel fold is an evolutionarily conserved motif found in proteins with a variety of enzymatic functions. Here the authors explore the fitness landscape of the TIM barrel protein IGPS and uncover evolutionary constraints on both sequence and structure, accompanied by long range allosteric interactions.
Proteins carry out diverse and essential functions in all living organisms. Notable among these functions is the catalysis of a host of complex chemical reactions under a broad range of environmental conditions. Based on enzyme classification, proteins catalyse over 5,700 unique biochemical reactions by employing ∼1,400 unique folds1. This structural redundancy shows that enzymes capitalize on robust structural platforms to introduce novel chemistries through sequence diversification. The TIM barrel fold is one of the oldest and most common motifs in biology2. The polarized structure has a well-ordered scaffold that is fused to an autonomous domain for substrate binding, catalysis, and product releases; the spatial segregation of the active site from the stabilizing protein core is an elegant solution for evolving new functions of TIM barrels3. The TIM barrel superfamily contains 57 distinct families and encompasses five of the six enzyme commission functional categories catalysing at least 34 unique functions4.
As sequences and functions of TIM barrel fold proteins emerged through a divergent evolutionary process, a quantitative description of the TIM barrel fitness landscape would be a crucial step in our understanding of protein evolutionary dynamics. Recent deep mutational scanning experiments provided significant insight into the fitness landscapes of individual proteins5,6,7,8,9,10,11,12,13 or very closely related homologues14. It was found that thermodynamic effects of mutations are not very sensitive to sequence background, prompting biophysical models of sequence evolution10,15,16,17,18,19. Furthermore, mutational scans showed that site-specific amino-acid preferences and, presumably, fitness landscapes are nearly identical in homologues of influenza virus nucleoprotein with 94% identity14. To date, it remains unclear whether the fitness landscapes remain similar in homologous proteins with a strongly divergent evolutionary history.
Here we perform a mutational scan of three orthologous TIM barrel fold proteins, indole-3-glycerol phosphate synthase (IGPS), to experimentally determine the fitness landscapes of proteins sharing the same fold and function, but with ancient divergences and low sequence identity. IGPS proteins were mutagenized in eight 10-residue segments, following the fold symmetry and covering the β-barrel core and adjacent elements of the αβ- and βα-loops. A tryptophan auxotrophic S. cerevisiae yeast strain, created by deletion of the endogenous IGPS gene, was transformed to prototrophy with each of the three orthologous genes. Relative fitness of the mutants, which depends on IGPS activity, was determined by the abundance of the mutant DNA sequence relative to wildtype (WT) over time20.
A comprehensive analysis of 5,040 mutations in the three orthologues demonstrated that the fitness landscapes of IGPS orthologues are statistically significantly correlated to each other, despite the sequence identity of approximately 30–40%. Surprisingly, we found that fitness can be dramatically enhanced by mutations distal from the active site in all three orthologues. Fold geometry and structural elements of the TIM barrel fold, as well as sequence conservation and amino-acid biochemistry, all impose measurable constraints on the fitness landscape. Principal component analyses (PCA) detected commonality between sources of fitness variance in the three IGPS TIM barrel orthologues, while statistical coupling analysis (SCA) revealed evolutionary correlations between the active site and positions of the distal beneficial mutations. These results have significant implications for the design of TIM barrel enzymes with novel functions not found in nature and insights into the unanticipated allostery for the TIM barrel motif.
Results
Fitness landscape of individual IGPS TIM barrel proteins
We applied the EMPIRIC deep mutational scan approach, developed by the Bolon group20, to explore the fitness landscape of a monofunctional enzyme of the TIM barrel fold. In the canonical TIM barrel structure, eight β-strands and α-helices alternate in sequence, and the β-strands assemble sequentially into a cylindrical core around which the α-helices form a helical shell (Fig. 1a). The TIM barrel scaffold is highly symmetrical, displaying a four-fold βαβα symmetry at the level of the smallest independent folding unit, the βαβ module (Fig. 1b)21. We chose three phylogenetically divergent IGPS orthologues from a thermophilic archeaon, S. solfataricus (SsIGPS), and two thermophilic bacteria, T. maritima (TmIGPS) and T. thermophilus (TtIGPS), Table 1. For the three proteins, we carried out selection experiments in yeast to determine the fitness of all possible mutations in libraries of 10 positions spanning the 8 αβ-loops, β-strands, and initial portion of the βα-loops (Fig. 2). Mutations were introduced by restriction of the gene followed by its ligation with a cassette containing all 64 codons for a given position, as opposed to transcribing the gene using an error-prone polymerase9,20. This approach generates a well-defined, rather than random, sequence diversity in the mutagenized region, reducing experimental noise.
Figure 1. Orthologous IGPS proteins fold into highly similar canonical TIM barrel structures.

(a) Ribbon diagrams of structurally aligned SsIGPS (PDB: 2C3Z), TmIGPS (PDB: 1I4N), and TtIGPS (PDB: 1VC4). EMPIRIC mutagenesis library positions are highlighted in orange. The parallel β-strands vary in length from 4–6 residues and the β-barrel structure forms four layers of side chains from alternating β-strands that protrude into the protein core (Supplementary Fig. 1). Preceding the N terminus of the β-strands, short αβ-loops, generally 3–4 residues in length, are proposed to play a role in stability74. At the C-terminus of the β-strands, long-βα loops, between 5 and 13 residues, link the C termini of the β-strands to the N termini of the subsequent α-helices and invariably form the active site of the enzyme. (b) The four-fold symmetric βαβα repeat unit holds the minimal independently folding unit, the βαβ module, highlighted in orange, wheat, and orange, respectively. The even helix shaded in grey does not contribute significantly to stability of the four-fold unit. Within the βαβ module, a canonical βα hairpin clamp formed between a main chain amide H-bond donor in +1 position of the odd β-strand and the side chain acceptor at the −1 position in the following even αβ-loop is highlighted in blue.
Table 1. Pairwise sequence and structure similarity of the three IGPS orthologues and mutagenized libraries.
| SsIGPS | TmIGPS | TtIGPS | |
|---|---|---|---|
| SsIGPS | — | Library sequence:Identical: 51%Similar: 71% | Library sequence:Identical: 49%Similar: 68% |
| TmIGPS | Full-length sequence:Alignment length: 267Identical: 30%Similar: 49% | — | Library sequence:Identical: 54%Similar: 73% |
| Structure:Alignment length: 216r.m.s.d.: 1.58 Å | |||
| TtIGPS | Full-length sequence:Alignment length: 271Identical: 35%Similar: 49% | Full-length sequence:Alignment length: 277Identical: 27%Similar: 43% | — |
| Structure:Alignment length: 214r.m.s.d.: 1.24 Å | Structure:Alignment length: 241r.m.s.d.: 1.72 Å |
Figure 2. Fitness landscapes of the three IGPS orthologues.

Values of the selection coefficient are colour-coded on a continuous scale from 1 to −1.5 indicated by the colorbar. WT residues and positions are labelled at the bottom of each panel. Mutant amino acids are indicated on the vertical axis. Within the heatmap, WT residues are indicated by the black outline. Low-quality data filtered from analysis are indicated by the red checkered boxes. Canonical secondary structures are drawn at the top of the panels. Active sites are indicated by the red stars at the top of the position columns. βα hairpin clamps are indicated by the black brackets at the bottom of the position columns. Fitness gains were observed with several mutations of the βα hairpin clamps. For example, more than half the mutations in the three β3α3 hairpin clamps SsIGPS I107 and D128, TmIGPS I105 and D126, TtIGPS I109 and S130 are beneficial (s>0). Two non-canonical interactions analogous to the βα hairpin clamp in SsIGPS are indicated by the red brackets. An ionic interaction is observed between E155 and R175. A hydrophobic stacking interaction is observed between I45, V78, and F40 (not included in library). All mutations of SsIGPS I45, except to stop codons, resulted in fitness advantage over WT.
Fitness was quantified as the selection coefficient s, that is, the slope of the relative abundance of mutant to WT IGPS over time (see Methods and Supplementary Fig. 2). Mutants with selection coefficient of 0 have doubling time equal to WT, while those with a selection coefficient of −1 are lethal. Over 55% of the mutations were deleterious with s<−0.75. The β-strands showed a particularly low tolerance to mutation. Presumably, mutations in the core destabilize the native state, resulting in a lower population of functional IGPS enzymes, or distort the active site, reducing catalytic power. Six catalytically important residues form the active site and are located at or near the C termini of β1, β3, β5, β6 and β7. Mutations at these highly conserved residues were strictly not tolerated. Bimodal distributions are often observed with fitness landscapes, describing a thermodynamic ‘cliff' at which proteins unfold and the mutations are lethal17,19,22. While we observed the expected bimodal distribution, the high fitness mode showed a bias towards beneficial rather than neutral fitness (Supplementary Fig. 3). Positive selection coefficients were associated with several mutations in the active site βα-loops at positions that do not directly support enzyme chemistry. Remarkably, fitness can be increased by mutations far from the active site in the αβ-loops, most notably at the positions of βα hairpin clamps (Fig. 2). βα hairpin clamps are long-range hydrogen bonds between the even numbered αβ-loops and the preceding odd numbered β-strands, enforcing β-strand alignment and providing stability (Fig. 1b)23. Based on the repeating fitness patterns in the minimally independent folding βαβ modules contained within the four-fold βαβα symmetrical units for each of the three orthologues, we analysed the selection coefficients in groups of two consecutive libraries, that is, odd (n) and even (n+1) strands (Fig. 1b).
αβ-loops
Within the canonical TIM barrel structure, there is a bias towards the glycine-x-aspartic acid motif in the αβ-loops leading up to the even number β-strands2,23. In the even numbered αβ-loops, mutations away from small hydrophobic residues at −2 position (X in the motif) with respect to the β-strand, prevent the tight turn required to dock the preceding helix. We found that this distortion of the βαβα module results in low organismal fitness, likely reflecting destabilization of the native state from poor packing. In contrast, the odd numbered αβ-loops link adjacent modules and are less constrained in their residue choice. Many mutations at the −3 and −2 positions of the odd strands were beneficial. At the −1 position, the higher fitness in the even versus the odd strand reflects their different accessible surface area (ASA). The average selection coefficient of these positions is correlated with the ASA, where mutations at the more buried even loops resulted in lower fitness than mutations in the more solvent exposed odd loops (Supplementary Fig. 4). Similar results were found using the relative surface area (RSA)24 instead of ASA.
βα hairpin clamp
Loss of the βα hairpin clamp between the canonical aspartic acid at the −1 position of the even strand and the main chain amide hydrogen beneath a large hydrophobic side chain at the N terminus of the preceding odd numbered β-strand elicited a fitness gain relative to the WT protein (Figs 1b,2). This enhancement is the most dramatic at the β3α3 hairpin clamp in all three orthologues. Significantly, mutation of I107 in the SsIGPS β3α3 hairpin clamp results in high fitness, while just one position away in the β-barrel, L108, mutations are mostly deleterious. The distinctly different responses highlight the different roles of the αβ-loop and the β-strand in protein stability and enzymatic function, each driving fitness of the host organism (Fig. 2). The most beneficial mutation in SsIGPS was I45Q, with selection coefficient of +0.89. Position I45 is involved in a triple stack of hydrophobic side chains where F40 (α0) is sandwiched between I45 (αβ0-loop) and V78 (αβ1-loop). This bridge between strands β1 and β2 is an alternative solution to the βα hairpin clamp typically found to stabilize odd and even β-strands. Mutations of the I45 to any other amino acid, except stop codons, are uniformly beneficial (Supplementary Fig. 5).
β-strands
The largely deleterious mutations in the β-strands indicate that minimal changes are tolerated within the protein core. Similar to the αβ-loops, the residues in each β-strand can be further stratified by side chain orientation into or out of the β-barrel and by participation in the four layer levels within the β-barrel (Supplementary Fig. 1). The evolved structure maximizes buried surface area of side chains, while minimizing steric clashes, by alternating side chain orientation between odd and even strands within a layer and between layers within β-strands25,26. Side chains that point inward form a highly stable hydrophobic core, while side chains pointing outward provide docking surfaces for the concentric α-helices by hydrophobic interactions. For side chains pointing outward in layers 2–4, a significant fraction of mutations is beneficial, in contrast to the almost complete lethality of their inward facing counterparts (Fig. 3). Mutations of β-strand residues pointing out of the barrel would be expected to perturb the α-helix and β-strand interface as well as the intervening βα-loop. Interestingly, inward facing side chains in layer 1 can also increase the fitness for a significant fraction of the mutations. For mutations in the fourth layer, the poor fitness may reflect the known sensitivity of the βα-loops to the interface between the stability core and the active site loops27.
Figure 3. Distribution of fitness values displayed as violin plots stratified by side chain orientation ‘in' (left) and ‘out' (right) and by layers (L1 to L4).

Guide lines are drawn at s=0 in green and s=−1 in blue. The mean and s.d. are indicated by the red diamond and error bars. Sample numbers are indicated beneath each layer. Permutation tests (n=10,000) were used to assess statistical significant differences between distributions based on ratio of beneficial (s>0) to total mutations (**P<0.02). See text and Supplementary Fig. 1 for details.
βα-loops and the active site
The active site spans multiple β-strands and subsequent βα-loops (Fig. 2). Although mutations at catalytically important residues were uniformly deleterious, some nearby mutations showed improved fitness over WT. Across the three orthologues, beneficial mutations were observed at position 52 (SsIGPS numbering) between the glutamic acid (E51) and lysine (K53) active site residues bridging the β1-strand and βα1-loop interface. Coordination of the substrate to its reactive conformation is mediated by electrostatic interactions between K53 in the βα1-loop and K110 in the βα3-loop28. However, molecular dynamics simulations of thermophilic SsIGPS under mesophilic temperatures suggest that strong electrostatic interactions between K53 and the substrate result in a nonproductive substrate geometry29. Our fitness results suggest that disrupting the ionic interactions between E51 and the two lysines, K53 and K110, may favour the reactive conformation of the substrate. Other beneficial mutations include the phosphate binding residue, S234, and its adjacent residue, S235. These residues, located in the eighth ‘βα-loop' form a 310-helix, and mutation of either may distort the phosphate binding pocket so as to improve substrate binding and/or product release.
Correlation of fitness landscapes between IGPS orthologues
A major question emerging from this screen is whether protein fitness landscapes are conserved across sequence and/or structure space. We compared the fitness landscapes of the orthologues by calculating the Pearson correlation coefficient R between fitness values of the 20 mutant amino acids at a pair of positions in two orthologues. The probability distribution of R for a specific set of positions was then used to assess the similarity of fitness landscapes. We considered the following four sets of positions: (a) identical WT amino acids in a pair of orthologues irrespective of structural alignment; (b) all structurally aligned positions irrespective of WT amino acid; (c) all structurally aligned position with non-identical WT amino acids; and (d) positions aligned by their four-fold symmetry in the TIM barrel, irrespective of WT amino acid, Fig. 4.
Figure 4. Fitness landscapes of orthologous proteins are correlated despite their low sequence identity.

Distributions of Pearson correlation coefficients between fitness landscapes of (a) identical WT amino acids in a pair of orthologues irrespective of structural alignment, (b) all structurally aligned positions irrespective of WT amino acid; (c) all structurally aligned position with non-identical WT amino acids, and (d) positions aligned by their four-fold symmetry in the TIM barrel, irrespective of WT amino acid. Fitness landscapes are statistically significantly correlated despite the low sequence identity and ancient divergence of IGPS orthologues (**P<0.02). Statistical significance is detailed in Supplementary Table 1.
In all cases, the fitness landscapes of these groups were significantly different from the null model (pairwise correlations between all positions) according to Kolmogorov–Smirnov test (P<10−4, Supplementary Table 1). The correlations in all three pairs of orthologues were similar in magnitude, and in most cases their distributions were statistically indistinguishable (KS test, P>0.05, Supplementary Table 2). The modes of these specific distributions corresponded to the Pearson correlation between ∼0.5 and 0.7. On the other hand, distributions of correlations for different sets of positions (for example, (a) versus (b) for all orthologues) were different in most cases (KS test, P<10−4, Supplementary Table 3), suggesting that structure and sequence features define the fitness landscape. These results are statistically significant compared to variation between experimental replicates (see Methods and Supplementary Figs 9 and 10).
Despite ancient divergence and bacterial versus archaeal origins, we found that fitness landscapes of three IGPS proteins are significantly correlated. Remarkably, for positions exhibiting as low as 40% sequence conservation in the multiple sequence alignment of IGPS proteins, experimentally determined fitness landscapes of the three orthologues remained correlated to each other, R≥0.4 (Fig. 5a), complementing earlier observations using sequence alignments and mutational scans14,30.
Figure 5. Sequence conservation and epistasis affect fitness landscapes.

(a) Fitness landscapes of IGPS orthologues are significantly correlated (R>0.4) for IGPS positions ('open circle' odd numbered libraries, 'filled circle' even numbered libraries) displaying 40 to 100% conservation in the multiple sequence alignment (red box): the fitness landscapes of highly conserved positions are strongly correlated. At the same time, the wide range of correlations before the ‘cliff' of 40% conservation suggests that sequence conservation is not the only determinant of fitness. (b) Mutations that ‘transform' one IGPS orthologue into another primarily have neutral or beneficial effects (pink histogram) compared to all mutations (grey). Epistatic interactions are responsible for the existence of transformative detrimental mutations (minor peak on the pink histogram around s=−1). The distribution of selection coefficients for the transformative mutations is significantly different that than of the null distribution (**P<0.02).
Correlation of fitness landscapes and epistasis
The correlations between fitness landscapes of orthologous proteins are intricately connected to the epistatic interactions in these proteins. If epistasis was absent, we would expect a near-perfect correlation between landscapes, as corresponding sites would have the same amino-acid preferences. Conversely, if epistasis was very strong, the fitness effects of every mutation would be entirely determined by the protein as a whole, and one would expect the fitness landscapes to become uncorrelated. Loosely, correlation of fitness landscapes is inversely proportional to the degree of epistasis. To unravel potential epistatic interactions in IGPS, for each orthologue, we selected ‘transformative' mutations that correspond to WT residues in another orthologue, ‘transforming' one protein into another. The distribution of selection coefficients of transformative mutations compared to the distributions of all selection coefficients in the three orthologues is significantly different (Fig. 5b). Transformative mutations are significantly enriched in neutral and beneficial mutations; deleterious mutations are depleted. As widely expected, an amino acid that has naturally evolved in one of the orthologues is often tolerated in another orthologue. However, the existence of detrimental transformative mutations suggests epistatic interactions, as their fitness is strongly affected by interactions with the rest of the protein. We found that about 31% of the transformative mutations were detrimental (s<−0.5) in IGPS. For comparison, a previous study with isopropymalate dehydrogenase from E. coli and P. aeruginosa found that 38% of 168 transformative mutations were detrimental in vitro31.
Sources of variance in experimental fitness landscapes of IGPS
To determine if the major sources of fitness variance are equivalent across the three orthologues, we applied PCA to the 80 × 20 matrices of selection coefficients of all mutations at each site surveyed, Supplementary Fig. 6. For each orthologue, the first two principal components accounted for almost 70% of observed fitness effects. The first principal component (PC1) explains ∼51% of the fitness variance and, by construction, is proportional to the average fitness of the position (Fig. 6a). We did not detect a significant linear relationship of PCA components to features such as ASA, RSA, b-factor, and WT amino-acid sequence conservation, if all three orthologues are considered together (Fig. 6b). Still, a periodicity observed in the PC1 scores along the library positions prompted a closer look at the relationship between PC1 and the four-fold symmetric structure (Fig. 6c). This scatterplot of PC1 showed differences in the deviation of the principal component scores along the coarse-grained ten library positions. The highest scores, representing the highest average fitness, are found mainly at the first three positions of the odd strand libraries and the last position of the even strand libraries, associated with the αβ-loops and βα-loops, respectively. The lowest scores, representing the lowest average fitness, were found at positions six and eight, associated with the β-strands whose residues point into the β-barrel. The average four-fold PC1 varied linearly with average four-fold IGPS conservation (R=0.84) (Fig. 6d). Thus, fitness must relate to structural conservation of the four-fold symmetry. The second principal component (PC2) explains ∼17% of the variance in our fitness data set and is correlated best with the hydrophobicity of the WT residues (REMPIRIC PC2 to Hydrophobicity∼0.67). Hydrophobicity of the WT residues is a major factor affecting the fitness outcome, independent of the residue position in the structure.
Figure 6. The first principal component of fitness landscape is related to average four-fold conservation.

(a) By construction, the first principal component (PC1) of the EMPIRIC PCA is linearly related to average fitness (R=0.996, 'open circle' odd numbered libraries, 'filled circle' even numbered libraries). (b) A linear relationship between EMPIRIC PC1 and IGPS conservation of varying strength was observed for each orthologue. A linear correlation of R=0.408 was observed if all three orthologues were considered together. (c) The values of EMPIRIC PC1 vary with four-fold aligned positions, implicating structure in fitness determination. The canonical secondary structures are indicated above the plot. The green line indicates the average score. Highest scores were observed at both ends of the library positions, associated with the αβ and βα-loops (red boxes). Lowest average scores were observed in the intermediate positions, associated with the β-strands (blue boxes). (d) Average EMPIRIC PC1 scores based on the four-fold alignment correlate linearly with average four-fold IGPS conservation (R=0.842).
Experimental data versus evolved IGPS and TIM sequences
To separate the factors that are important for function from those important for stability in our fitness assay, we performed separate PCA analyses of representative sequences of IGPS and TIM barrel proteins, using amino-acid frequencies at each site as a proxy for their evolutionary fitness (see Methods). Since all IGPS enzymes are TIM barrel proteins, we expected that the major drivers of sequence variance will reflect stability and structure for the TIM PCA and stability, structure, and function for the IGPS PCA. By selecting the 80 aligned positions examined in our fitness assay, we directly compare and contrast the experimental EMPIRIC IGPS fitness landscape to the naturally evolved amino-acid preferences in IGPS and TIM barrels.
The first principal component for both the IGPS PCA and TIM PCAs accounted for ∼30% of the variance found in their amino-acid preferences and was correlated with WT amino-acid hydrophobicity (RIGPS PC1 to hydrophobicity∼0.70). The two components correlated with each other (RIGPS PC1 to TIM PC1∼0.71) and with the second component of the EMPIRIC PCA (REMPIRIC PC2 to IGPS PC1∼0.69, REMPIRIC PC2 to TIM PC1∼0.59), which correlated with hydrophobicity. Thus, stability appears to be the main driver for the residue selection within IGPS and TIM barrels, in general, for the segments examined.
The second principal component accounts for ∼20% of the variance observed in IGPS and TIM barrels and is collinear with information content of the sequence alignment, indicating conservation. Similar to PC1, the two PC2 of the IGPS PCA and TIM PCA were correlated (RIGPS PC2 to TIM PC2∼0.68). At the same time, no correlation was observed between TIM conservation and EMPIRIC fitness in the IGPS orthologues. Presumably, conservation in a sample of 71 representative TIM sequences only reflects very general fold patterns, and has little predictive value for the fitness of mutants in the specific family such as IGPS.
Statistical coupling analysis
While active site residues are highly conserved to preserve enzymatic function, other sites indirectly supporting enzyme activity or those supporting protein stability can also show position-specific sequence constraints and correlated substitutions between positions within a protein family32,33,34. SCA was used to characterize functionally important co-evolving residues in SsIGPS that may inform our fitness results32. Two significant sectors were identified. Sector one (46 positions) involved mainly amino acids at the interface between the β-barrel and α-helical shell (Supplementary Fig. 7a). The distribution of fitness effects for the 17 positions with known fitness was unimodally deleterious. Over 37% of the residues were branched aliphatic residues and 28% were charged residues, highlighting the importance of hydrophobic and electrostatic interactions in stabilizing the β-barrel/helical shell interface throughout evolution. In contrast, sector two (45 positions) described both stability and functional properties. The distribution of fitness effects for the 25 positions with known selection coefficients was bimodal, smodes∼−0.9 and −0.3, reflecting highly conserved sites that are intolerant of mutations and sites of high adaptation potential. Active site residues, including the substrate-binding site, made up ∼20% of the sector (Supplementary Fig. 7b). Of particular interest, two non-active site positions found to improve SsIGPS fitness with mutation were identified in sector two, residues I45 and D128. Corresponding mutations of the β3α3 hairpin clamp in the other two orthologues yielded similar fitness gains. Serving as a putative conduit between the active site and these two residues are select α-helical and β-strand residues in the SCA that span the two ends of the protein (Supplementary Fig. 8). The distributed nature of covariation, or SCA sectors, over the IGPS structure is consistent with statistical observations of long-range effects of mutations35.
Discussion
Systematic exploration of fitness landscapes has become possible using EMPIRIC and other high throughout mutagenesis approaches9,10,11,12,13,14,20,36. We have explored the fitness landscapes of three orthologous TIM barrel proteins from archaea and bacteria to answer two important questions at the intersection of biophysics and evolutionary biology: (1) what are the salient sequence and structural correlates of a TIM barrel fitness landscape, and (2) are the fitness landscapes of phylogenetically divergent orthologous proteins correlated with each other and, if so, what is the basis of the correlation?
Protein fitness landscapes depend on stability and enzymatic function, which are both affected by sequence and structure3,13,18. Thermophilic proteins employ multiple mechanisms to maintain their structure at high temperatures, including a higher content of salt bridges and H-bonds relative to their mesophilic counterparts37,38. Increased stability is certainly required at high temperature, but it can be a liability at lower temperatures39. The native basin of proteins and enzymes contains a dynamic ensemble of rapidly interconverting states, some of which may be more favourable for particular functions, for example, substrate binding or the catalysis of chemical reactions40. For IGPS, ring closure of the substrate, 1-(o-carboxyphenylamino)-1-deoxyribulose 5-phosphate (CdRP), requires a minimal distance and favourable orientation between C1 and C2' for the reaction to proceed28,29. This conformation is mediated by electrostatic interactions between the substrate and the active site lysine residues29. At lower temperatures, the conserved lysine residues show decreased flexibility and restricted structural orientation leading to a greater population of CdRP in an extended, unproductive conformation. At higher temperatures, electrostatic interactions between IGPS and CdRP favour a reactive substrate conformation29. Access to these productive higher energy states in the native basin would increase if a mutation leads to a destabilized unproductive native state conformation without a concomitant destabilization of the productive higher energy state (Fig. 7a,b). We hypothesize that fitness gains are driven by increased local flexibility and/or dynamics accompanied by a population shift toward the productive catalytic conformation(s). Conversely, deleterious effects may be caused by a decrease in stability and the concomitant decrease in the concentration of the enzyme or by distortion of the active site and ensuing reduction of catalytic properties (Fig. 7c). In fact, largely aliphatic residues in the barrel interior (Supplementary Table 4) are essential for structural integrity and incur a high proportion of deleterious mutations (Fig. 3). Native state hydrogen exchange protection patterns in the HisF TIM barrel from T. maritima revealed that layers 2 and 3 were strongly protected from solvent exchange, implying a major role in stabilizing the native state41. The side chains in these two layers participate in large clusters of densely packed ILV residues, known to form cores of stability in globular proteins42. We argue that the core of the barrel is critical for the stability of both the unproductive and productive conformations of TIM barrels.
Figure 7. Putative effects of mutations on the energy landscape of IGPS.

Free energy diagrams showing three possible scenarios for the effect of a single mutation on the folding free energy surface (ΔG°, Gibbs free energy of folding; U, unfolded state; N, native state ensemble). An ensemble of rapidly interconverting productive (Nprod.) and nonproductive (Nunprod.) conformations resides in the native basin. At mesophilic temperatures, thermophilic IGPS access the productive conformation to a lesser extent than at their native thermophilic temperatures. (a) Beneficial mutations may destabilize the unproductive native state without a concomitant destabilization of the higher energy, productive conformation, resulting in a shift in the population from the unproductive to the productive conformation. Increased activity improves fitness. (b) WT-like mutations may destabilize both the unproductive and productive states, resulting in no net change in the population. No change in fitness is observed. (c) Deleterious mutations may greatly destabilize the native state, resulting in a population shift to inactive, partially or fully unfolded, states that are susceptible to proteolysis. Poor fitness is associated with loss of enzymatic capabilities.
Increased activity of mutant thermophilic proteins, including IGPS, at a lower temperature has been previously reported, and typically involves modifications to the active site or substrate-binding site43. In contrast, many positions where mutations increased the fitness in our assay were distal to the active site in the βα-loops. For example, deletion of the canonical β3α3-hairpin clamp, such as SsIGPS I107 and D128, and several other hairpin clamps within the four-fold symmetrical βαβ modules consistently shortened doubling time relative to the WT. Similarly, for the hydrophobic stack between SsIGPS F40, I45 and V78, all mutations at I45, except to stop codons, displayed increased fitness. These long-range effects on fitness revealed an unexpected allostery between the αβ-loops at one end of the TIM barrel and the active site at the opposite end. The biological relevance of this allostery is supported by known cross-activating communication between HisF synthase TIM barrel and HisH glutaminase in the histidine biosynthetic pathway44. Activities of these two proteins are coupled through a physical interaction between the two enzymes, where the αβ-loops of the HisF TIM barrel dock onto the oxyanion hole of the HisH glutaminase active site44. Signal transduction between the two active sites spans the entire length of the TIM barrel and is mediated by correlated motions of several networks of residues within the β-barrel and α-helices45.
Although the IGPS fitness landscape observed in our experiments in yeast may differ from that experienced during evolution of these thermophilic proteins, we believe that the basic biophysical and structural constraints we observed hold true in the natural environment. Moreover, thermophilic orthologues functioning at mesophilic temperatures provide targets of opportunity to detect and map pathways of allostery leading to beneficial fitness under stress conditions. Bernhardt speculated long ago that consecutive enzymes in metabolic pathways might preferentially associate with each other to enhance the throughput of substrates to products46. While the assembly status of the three IGPS in our study is monomeric, various combinations of bifunctional and multifunctional enzymes from the tryptophan biosynthetic pathway occur in nature47. Thus, the inherent ability of a TIM barrel to stimulate or enhance the activity of another protein in an enzymatic pathway similar to the HisF/HisH pair through allosteric interactions would be advantageous.
Our fitness data for IGPS suggest that mutations in the αβ-loops induce changes in the distal active site βα-loops to favour the catalytically active conformation (Fig. 8). They may do so by mimicking the physiological stimulation by partner proteins or by inducing the conformation naturally preferred at their respective optimal growth temperatures. The greater fitness observed for mutations in the outward facing side chain positions in the β-barrel (Fig. 3) compared to mutations with inward facing side chains, implicates perturbations in the α-helical shell as the conduit for the allostery. This conjecture is supported by the SCA results that point to a possible pathway of signal transduction between the active site and the αβ-loops in SsIGPS, mediated by intervening helical elements (Supplementary Fig. 8).
Figure 8. The active site of IGPS coordinates the ring closure event converting substrate CdRP to product IGP.

The active site orients the C1 and C2' atoms of the substrate to a specific distance and geometry, favoring conversion of CdRP to IGP. Correlated motions of grouped amino acids dispersed throughout the protein influence conformation of the active site. This access to the productive enzyme conformation dictates enzyme activity. Under mesophilic conditions, thermophilic IGPS enzymes are less dynamic, less flexible and less active than at their native thermophilic temperatures. (a) During yeast growth at 30 °C, the active site of thermophilic IGPS favours an unproductive conformation, where CdRP is oriented in an extended, non-reactive conformation. Reduced activity results in reduced fitness of the WT thermophilic IGPS compared to the WT mesophilic IGPS. (b) An allosteric beneficial effect on fitness was observed with certain mutations of the αβ-loop, resulting in greater fitness of the mutants compared to the thermophilic WT IGPS proteins at mesophilic temperatures. These mutations transmit their allostery via correlated protein breathing motions to favour the productive conformation of the active site, where CdRP is oriented optimally in a reactive conformation. Improved fitness observed for β-strand residues whose side chain point to the β-strand/α-helices interface implicates the α-helices (Fig. 3) as the conduit for the allostery between the active site βα-loops and the αβ-loop.
Experimental mapping of fitness landscapes has progressed from sampling of some or all combinations of diallelic loci48,49 towards large-scale mutational scans9,10,11,12,13,14,20,36. Although there is evidence about the roughness of fitness landscapes for small deviations from the WT sequence6,50,51, our finding of correlations between the fitness landscapes of divergent IGPS orthologues suggests that the common fold and function act together to smoothen the landscape. In fact, low throughput experiments have shown that the effects of mutations on biophysical properties such as Tm and ΔΔG° are largely conserved across homologues of influenza nucleoprotein52, and modern versus ancestral thioredoxins and β-lactamases53. These studies suggest that amino-acid preferences at a given position in the structure are mostly conserved during evolution, contrary to a modelling prediction of Pollock et al.54 However, these works focused on specific biophysical properties of the proteins, rather their full functional consequences. This limitation was lifted in a deep mutational scan of two influenza virus nucleoproteins (94% sequence identity) in their full physiological context14. Again, the amino-acid preferences were found to be conserved at most sites, suggesting strongly correlated fitness landscapes of aligned positions. In agreement, we found significantly higher correlation of fitness landscapes of aligned positions even when the WT amino acids differed between orthologues pairs (Fig. 4c,d).
Beyond constraints imposed by the structure, ancestral sequence reconstruction of ribonuclease H1 suggests that various molecular mechanisms are employed to stabilize proteins during evolution in response to environmental stress55. Diverse distributions of ionic, H-bond, and hydrophobic interactions are observed in our three orthologues56,57,58, indicating that each protein sampled different sequence space and evolutionary paths as part of its ‘thermodynamic system drift'55. At the same time, high correlation of fitness landscapes between positions with identical WT amino acids irrespective of structural alignment (Fig. 4a) highlight the biochemical and physical constraints required for properly maintaining the protein fold.
In summary, we found a very strong correlation of organism-level fitness landscapes of three extant proteins at 30–40% sequence identity, and of bacterial versus archaeal origin. Two related interpretations of this result appear possible. First, conservation of amino-acid preferences does persist across the two phylogenic domains and low sequence identity. Second, in a complementary way, we interpret this conservation as translocation of fitness landscapes in sequence space: fitness landscapes of single point-mutants can be successfully translocated to a different starting point in sequence space. We propose to visualize translocation by presenting the fitness landscapes of single point-mutants as pinwheels in sequence space (Fig. 9). All mutants are one change away from the WT, with the colour and height of each mutant representing its fitness. In this analogy, translocation means that pinwheels for orthologues centered far apart in sequence space still maintain a similar shape and colour profile.
Figure 9. Translocation of fitness landscapes in the sequence space of orthologous TIM barrels.

(a) Over evolutionary time, orthologous proteins adapt to changing environment for optimal fitness. Steps in sequence space select for protein stability and activity under their native conditions, all while retaining the TIM barrel fold and IGPS function. (b) Experimentally derived fitness landscapes mapped from point mutations represent single steps from WT sequence. Despite significant divergence of WT in sequence space, the fitness landscapes of IGPS orthologues remain correlated (dashed lines). Rather than traditional two-dimensional heatmaps, fitness values are displayed on a three-dimensional pinwheel, highlighting the wide range of possible fitness effects of a single sequence step. The profiles of the pinwheels are similar, indicating the correlation of fitness landscapes, even if WT sequences (centers of the wheels) are only ∼40% identical and widely separated. PCA demonstrates a correlation between experimental fitness landscapes and amino-acid preferences in evolved sequences.
Understandably, deviations from the perfect correlation between the landscapes of orthologues result from epistatic effects within the protein. As mentioned previously, the magnitude of epistasis is inversely related with the correlation between the fitness landscapes. For proteins of relatively small divergence, biophysical models achieve a significant power to predict the effects of mutations59,60, especially if informed by high-throughput mutational scan data10,15. Our findings suggest that such approaches may be extended to proteins with a greater degree of sequence divergence. Further development of the models and metrics for comparing the fitness landscapes61,62 will produce appropriate tools for quantifying the landscapes at various degrees of divergence between sequences.
Translocation of fitness landscape is facilitated by plasticity of the TIM barrel fold, both in natural sequences and in vitro, as stable TIM barrels can be created by fusions of natural half barrels, (βα)4 (refs 63, 64). Indeed, it is well established that the inverse protein folding problem, designing sequences for a given template structure, has multiple solutions65. Recently, the first successful de novo TIM barrel design employed a completely symmetric four-fold repeat25,66. Observations of the four-fold symmetry in both our experimental fitness landscape and in silico design serve to validate both approaches in understanding the fundamental properties of TIM protein architecture. From an experimental perspective, fitness studies on other common proteins platforms have the potential to reveal unanticipated sequence-structure-fitness relationships and provide new strategies for de novo protein design25,67.
Methods
Strains and culture conditions
S. cerevisiae strain BY4742 ΔIGPS::KanMX was produced using the same PCR-generated deletion strategy described by the Saccharomyces Genome Deletion Project68. The last 810 bp of the TRP3 gene encoding IGPS were replaced with the KanMX gene. Deletion of IGPS with the KanMX gene was confirmed by Sanger sequencing.
The pRS416 vector carrying the auxotrophic URA3 marker was a gift from Daniel Bolon's lab. A lower expressing Tma19 promoter, also provided by the Bolon laboratory, was used to increase the sensitivity of the fitness assay. Three silent mutations were introduced into the plasmid at the URA3 marker, the ampicillin resistance marker, and the Tma19 promoter to disrupt BSAI recognition sites. The BSAI enzyme was used to create the saturating mutagenesis libraries.
IGPS genes were purchased from Genscript. An N-terminal 6 × His tag and Tev protease recognition site were added to each construct in anticipation of future studies requiring protein abundance measurements and protein purification. In anticipation for in vitro folding studies, the non-canonical N-terminal α00 of each gene was deleted to reduce aggregation during refolding of purified proteins (SsIGPS Δ1–26, TmIGPS Δ1–31, TtIGPS Δ1–34). To prevent non-specific cleavage when using Tev protease, position 18 was mutated from arginine to serine in SsIGPS. To prevent disulfide bonds and oxidation by molecular oxygen, position 102 was mutated from a cysteine to a serine in TmIGPS. IGPS genes were cloned into the pRS416 vector using restriction sites, SpeI and BamHI.
For each of the orthologues, we created 8 plasmid libraries where a 10 amino-acid region was mutagenized using the EMPIRIC method, see Hietpas et al.20 for a detailed protocol. Briefly, inverted BsaI restriction sites are introduced by PCR within the region of interest. BsaI digestion is followed by directional sticky-end ligation of oligonucleotide cassettes containing a single randomized site containing all 64 codons. Thus, each library contained 640 sequence variants corresponding to all possible amino-acid (and codon) mutations at each position. The sequence variance was deterministic, as no error-prone polymerases were used.
Yeast transformation was performed using the LiAc/SS carrier DNA/PEG method as described by Gietz and Schiestl69. Yeast cells were grown in rich media with G418 to select for IGPS knockout yeast. Transformed cells were selected on synthetic minimal media lacking uracil. Selection for IGPS activity was achieved through growth of transformed yeast (one plasmid library per culture) in synthetic drop-out medium lacking tryptophan. All growth experiments were performed at 30 °C. Liquid cultures were maintained in log phase throughout the fitness assay by periodic dilution. G418 selection was maintained throughout the growth. The oligonucleotide sequences for mutant library generation and primer sequences used for creating the plasmid libraries and processing the deep-sequencing samples are listed in Supplementary Data 1.
EMPIRIC data processing
Illumina deep sequencing (36 base single reads) was performed by Elim Biopharmaceuticals. In-house analysis of the deep-sequencing results was performed using custom software. Reads were stringently filtered based on several criteria: Phred score>20 across all 36 bases (error probability of 0.01), valid barcode match, single reference sequence match for anticipated single codon mutations, and absence of MmeI recognition site. MmeI enzyme was used to create overhangs for ligation of time-stamping barcodes. Sequence counts were tracked on the nucleotide and amino-acid level. A total of 5,040 (10 AA/library × 8 libraries/orthologue × 3 orthologues × 21 mutation types) amino-acid mutations were created. Raw fitness was calculated as the slope of the log2 relative abundance of the mutant to WT versus time over 6 to 10 time points spread over 4 doubling periods of the yeast,
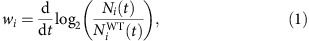 |
where Ni(t) is the abundance (count) of mutant i at time t, and NiWT(t) is the count of WT amino acid at the corresponding position. Slope was determined using the linear regression, Supplementary Fig. 2. From the raw fitness w, selection coefficients were determined according to
 |
where wSTOP is the average raw fitness of all mutations to a stop codon within a 10 amino-acid region, corresponding to an individual selection experiment, 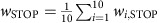 . This normalization ensures that on average, stop codons have a selection coefficient of −1. Fitness calculations were performed on all values except 79 mutations leading to MmeI recognition sites. Seven mutations had poor coverage in our mutagenesis libraries and were also removed from analysis: SsIGPS A174W, TmIGPS I131M, TmIGPS I178C, TmIGPS I178W, TtIGPS G78M, TtIGPS L153C and TtIGPS L153Y. After filtering, a total of 4,954 mutations were analysed.
. This normalization ensures that on average, stop codons have a selection coefficient of −1. Fitness calculations were performed on all values except 79 mutations leading to MmeI recognition sites. Seven mutations had poor coverage in our mutagenesis libraries and were also removed from analysis: SsIGPS A174W, TmIGPS I131M, TmIGPS I178C, TmIGPS I178W, TtIGPS G78M, TtIGPS L153C and TtIGPS L153Y. After filtering, a total of 4,954 mutations were analysed.
To assess the reproducibility of our fitness results, we compared selection coefficients of two full biological replicates of regions β3, β4 of SsIGPS comprising 399 mutations (R=0.947, Supplementary Fig. 9a).
Sequence and structural analyses
Pairwise sequence alignments of the three orthologues were performed using Clustal Omega provided by EMBL-EBI70. Sequence identities and similarities were calculated using Ident and Sim Program in the Sequence Manipulation Suite provided by bioinformatics.org. PDB accession codes used for structural analyses were 2C3Z for SsIGPS56, 1I4N for TmIGPS57 and 1VC4for TtIGPS58. RMSDs of structural alignments were performed using SPalign71. Structural figures were generated using PyMOL Molecular Graphics System v1.8.2.0. Accessible surface area (ASA) was calculated using EBI PISA tool. RSA was calculated from ASA using an empirical scale24.
Comparison of fitness distributions
For structural analyses, specific groups of residues were compared to identify differences in their distributions of fitness effects. The fraction of beneficial mutations was used to describe the shape of fitness distribution. A permutation test was used to determine if two fitness distributions differed significantly. For each subset, the original fitness measurements for 19 amino-acid substitutions at a given position were reassigned to a random position and the corresponding fraction of beneficial mutations was determined. Statistical significance (P value) of the observed fraction of beneficial mutations was calculated from a distribution (N=10,000) of the randomized values. Plots were generated in R version 3.2.0.
Correlations of fitness landscapes
The Pearson correlation coefficient was used to assess the similarity in response to the set of 19 amino-acid substitutions for a given pair of residues. To assess the statistical significance, we compared the fitness landscapes of two full biological replicates of regions β3, β4 of SsIGPS comprising 20 positions (Supplementary Fig. 9b). The average correlation between fitness landscapes of the replicates was R=0.89, much higher than all the correlations between the orthologues (Supplementary Fig. 10). The difference between the distributions of correlation coefficients of orthologues and that of the replicates was highly statistically significant (KS test, P<10−4).
Principal component analysis
PCA was used to identify major sources of variance in our data set. PCA analyses were performed using custom scripts written in Python version 2.7.6. For the EMPIRIC PCA, fitness values were normalized for every position to have the same mean and s.d. PCA was performed separately for each of the orthologues. For the IGPS PCA, the multiple sequence alignment (MSA), 1,744 representative sequences was obtained from PFam (accession code PF00218). Two positions in the fifth library of SsIGPS had no structural match with positions of the TmIGPS and TtIGPS libraries. Therefore, three separate alignments were created using positions corresponding to all 80 library positions in each of the orthologues. The minimal pairwise overlap between the three alignments is >60 positions. Amino-acid frequencies corresponding to our library positions were extracted and the resulting three matrices of size 80 × 20 were log-transformed and normalized for PCA application. No significant differences in the IGPS-PCA results were observed between the three sequence alignments. For the TIM PCA, structural alignment of 71 non-redundant TIM-barrel proteins72 was constructed using SPalign71. Pairwise structural alignment was performed for each of template structures SsIGPS, TmIGPS, and TtIGPS to the 71 representative TIM barrel structures. As with the IGPS-PCA, we extracted the corresponding 80 library positions from the pairwise alignment to obtain three multiple structure sequence alignments (MSSA) for each of the templates. No significant differences in the TIM-PCA results were observed between the three structural alignments.
Statistical coupling analysis
SCA was used to identify groups of co-evolving residues that have functional roles in protein activity and stability. A modified IGPS-MSA was generated for the SCA in order to minimize large gaps in the alignment. The full-length sequence of SsIGPS was used as a BLAST search seed in the NCBI non-redundant database. COBALT73 was used to align 1,000 protein sequences with length between 200 and 300, and redundancy filter reduced number of sequence down to 537 (35 to 95% sequence identity). Aligned positions with >80% gaps were removed from the alignment. This modified alignment was passed into SCA v5.0 MATLAB toolbox32 and analysed using the default parameters. Two eigenvalues from the SCA positional correlation matrix exceeded the significance threshold. Therefore, the expected number of sectors was set to kmax=2. Spectral analysis of SCA sequence correlation matrix revealed no phylogenetic and/or sequence sampling biases, permitting functional interpretation of the two identified protein sectors.
Data availability
All data, primer sequences, alignments, and scripts for data analyses are available at https://github.com/yvehchan/TIM_EMPIRIC or from the corresponding author upon request.
Additional information
How to cite this article: Chan, Y. H. et al. Correlation of fitness landscapes from three orthologous TIM barrels originates from sequence and structure constraints. Nat. Commun. 8, 14614 doi: 10.1038/ncomms14614 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Supplementary Figures and Supplementary Tables
Oligonucleotide sequences for mutant library generation. Primer sequences used for creating the plasmid libraries and processing the deep sequencing samples are available for download.
Acknowledgments
This research was supported by NIH GM23303 to C.R.M. We thank Daniel Bolon for providing valued expertise, material, and comments on the manuscript. We thank current/past members of the Bolon lab: Ryan Hietpas, Benjamin Roscoe, Parul Mishra and Li Jiang, and Sagar Kathuria from the Matthews lab for insightful discussions during our experimental setup. We thank Troy Whitfield for insightful discussions during our data analysis.
Footnotes
The authors declare no competing financial interests.
Author contributions C.R.M. and Y.H.C. conceived and designed the studies. Y.H.C. performed the experiments. C.R.M. and K.B.Z. guided the data analyses. K.B.Z., Y.H.C. and S.V.V. analysed the data. Y.H.C. wrote the manuscript with contributions from all authors.
References
- Berman H. M. The protein data bank. Nucleic Acids Res. 28, 235–242 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagano N., Orengo C. A. A. & Thornton J. M. M. One fold with many functions: The evolutionary relationships between TIM barrel families based on their sequences, structures and functions. J. Mol. Biol. 321, 741–765 (2002). [DOI] [PubMed] [Google Scholar]
- Tóth-Petróczy Á. & Tawfik D. S. The robustness and innovability of protein folds. Curr. Opin. Struct. Biol. 26, 131–138 (2014). [DOI] [PubMed] [Google Scholar]
- Goldman A. D., Beatty J. T. & Landweber L. F. The TIM Barrel architecture facilitated the early evolution of protein-mediated metabolism. J. Mol. Evol. 82, 17–26 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hietpas R. T., Jensen J. D. & Bolon D. N. A. Experimental illumination of a fitness landscape. Proc. Natl Acad. Sci. USA 108, 7896–7901 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bank C., Hietpas R. T., Jensen J. D. & Bolon D. N. A. A Systematic Survey of an Intragenic Epistatic Landscape. Mol. Biol. Evol. 32, 229–238 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiffler M. A., Hekstra D. R. & Ranganathan R. Evolvability as a function of purifying selection in TEM-1 β-Lactamase. Cell 160, 882–892 (2015). [DOI] [PubMed] [Google Scholar]
- Starita L. M. et al. Massively parallel functional analysis of BRCA1 RING domain variants. Genetics 200, 413–422 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doud M. B. & Bloom J. D. Accurate measurement of the effects of all amino-acid mutations on influenza hemagglutinin. Viruses 8, 155 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom J. D. An experimentally determined evolutionary model dramatically improves phylogenetic fit. Mol. Biol. Evol. 31, 1956–1978 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roscoe B. P., Thayer K. M., Zeldovich K. B., Fushman D. & Bolon D. N. A. Analyses of the effects of all ubiquitin point mutants on yeast growth rate. J. Mol. Biol. 425, 1363–1377 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melamed D., Young D. L., Gamble C. E., Miller C. R. & Fields S. Deep mutational scanning of an RRM domain of the Saccharomyces cerevisiae poly(A)-binding protein. RNA 19, 1537–1551 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkisyan K. S. et al. Local fitness landscape of the green fluorescent protein. Nature 533, 397–401 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doud M. B., Ashenberg O. & Bloom J. D. Site-specific amino acid preferences are mostly conserved in two closely related protein homologs. Mol. Biol. Evol. 32, 2944–2960 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom J. D. An experimentally informed evolutionary model improves phylogenetic fit to divergent lactamase homologs. Mol. Biol. Evol. 31, 2753–2769 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom J. D., Wilke C. O., Arnold F. H. & Adami C. Stability and the Evolvability of Function in a Model Protein. Biophys. J. 86, 2758–2764 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeldovich K. B., Chen P. & Shakhnovich E. I. Protein stability imposes limits on organism complexity and speed of molecular evolution. Proc. Natl Acad. Sci. USA 104, 16152–16157 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokuriki N. & Tawfik D. S. Stability effects of mutations and protein evolvability. Curr. Opin. Struct. Biol. 19, 596–604 (2009). [DOI] [PubMed] [Google Scholar]
- Wylie C. S. & Shakhnovich E. I. A biophysical protein folding model accounts for most mutational fitness effects in viruses. Proc. Natl Acad. Sci. USA 108, 9916–9921 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hietpas R., Roscoe B., Jiang L. & Bolon D. N. A. Fitness analyses of all possible point mutations for regions of genes in yeast. Nat. Protoc. 7, 1382–1396 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zitzewitz J. A., Gualfetti P. J., Perkons I. A., Wasta S. A. & Matthews C. R. Identifying the structural boundaries of independent folding domains in the alpha subunit of tryptophan synthase, a βα barrel protein. Protein Sci. 8, 1200–1209 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L., Mishra P., Hietpas R. T., Zeldovich K. B. & Bolon D. N. A. Latent effects of hsp90 mutants revealed at reduced expression levels. PLoS Genet. 9, e1003600 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X., Kathuria S. V., Vadrevu R. & Matthews C. R. βα-Hairpin clamps brace βαβ modules and can make substantive contributions to the stability of TIM barrel proteins. PLoS ONE 4, e7179 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tien M. Z., Meyer A. G., Sydykova D. K., Spielman S. J. & Wilke C. O. Maximum allowed solvent accessibilites of residues in proteins. PLoS ONE 8, e80635 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang P.-S. et al. De novo design of a four-fold symmetric TIM-barrel protein with atomic-level accuracy. Nat. Chem. Biol. 12, 29–34 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murzin A. G., Lesk A. M. & Chothia C. Principles determining the structure of β-sheet barrels in proteins I. A theoretical analysis. J. Mol. Biol. 236, 1369–1381 (1994). [DOI] [PubMed] [Google Scholar]
- Ochoa-Leyva A. et al. Protein Design through Systematic Catalytic Loop Exchange in the (β/α)8 Fold. J. Mol. Biol. 387, 949–964 (2009). [DOI] [PubMed] [Google Scholar]
- Hennig M., Darimont B. D., Jansonius J. N. & Kirschner K. The catalytic mechanism of indole-3-glycerol phosphate synthase: crystal structures of complexes of the enzyme from Sulfolobus solfataricus with substrate analogue, substrate, and product. J. Mol. Biol. 319, 757–766 (2002). [DOI] [PubMed] [Google Scholar]
- Mazumder-Shivakumar D., Kahn K. & Bruice T. C. Computational study of the ground state of thermophilic indole glycerol phosphate synthase: structural alterations at the active site with temperature. J. Am. Chem. Soc. 126, 5936–5937 (2004). [DOI] [PubMed] [Google Scholar]
- Meyer A. G., Dawson E. T. & Wilke C. O. Cross-species comparison of site-specific evolutionary-rate variation in influenza haemagglutinin. Philos. Trans. R. Soc. Lond. B. Biol. Sci 368, 20120334 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lunzer M., Brian Golding G. & Dean A. M. Pervasive cryptic epistasis in molecular evolution. PLoS Genet. 6, 1–10 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halabi N., Rivoire O., Leibler S. & Ranganathan R. Protein sectors: evolutionary units of three-dimensional structure. Cell 138, 774–786 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vijayabaskar M. S. & Vishveshwara S. Insights into the fold organization of TIM barrel from interaction energy based structure networks. PLoS Comput. Biol. 8, e1002505 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ovchinnikov S. et al. Large-scale determination of previously unsolved protein structures using evolutionary information. Elife 4, e09248 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jack B. R., Meyer A. G., Echave J. & Wilke C. O. Functional sites induce long-range evolutionary constraints in enzymes. PLoS Biol. 14, e1002452 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fowler D. M. et al. High-resolution mapping of protein sequence-function relationships. Nat. Methods 7, 741–746 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S., Tsai C.-J. & Nussinov R. Factors enhancing protein thermostability. Protein Eng. 13, 179–191 (2000). [DOI] [PubMed] [Google Scholar]
- Szilágyi A. & Závodszky P. Structural differences between mesophilic, moderately thermophilic and extremely thermophilic protein subunits: Results of a comprehensive survey. Structure 8, 493–504 (2000). [DOI] [PubMed] [Google Scholar]
- Závodszky P., Kardos J., Svingor Á. & Petsko G. A. Adjustment of conformational flexibility is a key event in the thermal adaptation of proteins. Proc. Natl Acad. Sci. USA 95, 7406–7411 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalbitzer H. R., Spoerner M., Ganser P., Hozsa C. & Kremer W. Fundamental link between folding states and functional states of proteins. J. Am. Chem. Soc. 131, 16714–16719 (2009). [DOI] [PubMed] [Google Scholar]
- Gangadhara B. N., Laine J. M., Kathuria S. V., Massi F. & Matthews C. R. Clusters of branched aliphatic side chains serve as cores of stability in the native state of the HisF TIM barrel protein. J. Mol. Biol. 425, 1065–1081 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kathuria S. V., Chan Y. H., Nobrega R. P., Özen A. & Matthews C. R. Clusters of isoleucine, leucine, and valine side chains define cores of stability in high-energy states of globular proteins: Sequence determinants of structure and stability. Protein Sci. 25, 662–675 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki T., Yasugi M., Arisaka F., Yamagishi A. & Oshima T. Adaptation of a thermophilic enzyme, 3-isopropylmalate dehydrogenase, to low temperatures. Protein Eng. 14, 85–91 (2001). [DOI] [PubMed] [Google Scholar]
- Douangamath A. et al. Structural evidence for ammonia tunneling across the (β/α)8 barrel of the imidazole glycerol phosphate synthase bienzyme complex. Structure 10, 185–193 (2002). [DOI] [PubMed] [Google Scholar]
- Rivalta I. et al. Allosteric pathways in imidazole glycerol phosphate synthase. Proc. Natl Acad. Sci. USA 109, E1428–E1436 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava D. K. & Bernhard S. A. Biophysical chemistry of metabolic reaction sequences in concentrated enzyme solution and in the cell. Annu. Rev. Biophys. Biophys. Chem. 16, 175–204 (1987). [DOI] [PubMed] [Google Scholar]
- Farber G. K. & Petsko G. A. The evolution of α/β barrel enzymes. Trends Biochem. Sci. 15, 228–234 (1990). [DOI] [PubMed] [Google Scholar]
- Weinreich D. M., Delaney N. F., DePristo M. A. & Hartl D. L. Darwinian evolution can follow only very few mutational paths to fitter proteins. Science 312, 111–114 (2006). [DOI] [PubMed] [Google Scholar]
- Olson C. A. A., Wu N. C. C. & Sun R. A comprehensive biophysical description of pairwise epistasis throughout an entire protein domain. Curr. Biol. 24, 2643–2651 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinkley T. et al. A systems analysis of mutational effects in HIV-1 protease and reverse transcriptase. Nat. Genet. 43, 487–489 (2011). [DOI] [PubMed] [Google Scholar]
- Kondrashov D. A. & Kondrashov F. A. Topological features of rugged fitness landscapes in sequence space. Trends Genet. 31, 24–33 (2015). [DOI] [PubMed] [Google Scholar]
- Ashenberg O., Gong L. I. & Bloom J. D. Mutational effects on stability are largely conserved during protein evolution. Proc. Natl Acad. Sci. USA 110, 21071–21076 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risso V. A. et al. Mutational studies on resurrected ancestral proteins reveal conservation of site-specific amino acid preferences throughout evolutionary history. Mol. Biol. Evol. 32, 440–455 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollock D. D., Thiltgen G. & Goldstein R. A. Amino acid coevolution induces an evolutionary Stokes shift. Proc. Natl Acad. Sci. USA 109, E1352–E1359 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hart K. M. et al. Thermodynamic system drift in protein evolution. PLoS Biol. 12, e1001994 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider B. et al. Role of the N-terminal extension of the (βα)8 -barrel enzyme indole-3-glycerol phosphate synthase for its fold, stability, and catalytic activity. Biochemistry 44, 16405–16412 (2005). [DOI] [PubMed] [Google Scholar]
- Knochel T., Pappenberger A., Jansonius J. N. J. & Kirschner K. The crystal structure of indoleglycerol-phosphate synthase from Thermotoga maritima. Kinetic stabilization by salt bridges. J. Biol. Chem. 277, 8626–8634 (2002). [DOI] [PubMed] [Google Scholar]
- Bagautdinov B. & Yutani K. Structure of indole-3-glycerol phosphate synthase from Thermus thermophilus HB8: Implications for thermal stability. Acta Crystallogr. D Biol. Crystallogr. 67, 1054–1064 (2011). [DOI] [PubMed] [Google Scholar]
- Bershtein S. et al. Protein homeostasis imposes a barrier on functional integration of horizontally transferred genes in bacteria. PLoS Genet. 11, e1005612 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigues J. V. et al. Biophysical principles predict fitness landscapes of drug resistance. Proc. Natl Acad. Sci. USA 113, E1470–E1478 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Visser J. A. G. M. & Krug J. Empirical fitness landscapes and the predictability of evolution. Nat. Rev. Genet. 15, 480–490 (2014). [DOI] [PubMed] [Google Scholar]
- Ferretti L. et al. Measuring epistasis in fitness landscapes: The correlation of fitness effects of mutations. J. Theor. Biol. 396, 132–143 (2016). [DOI] [PubMed] [Google Scholar]
- Sterner R. & Höcker B. Catalytic versatility, stability, and evolution of the (βα)8-barrel enzyme fold. Chem. Rev. 105, 4038–4055 (2005). [DOI] [PubMed] [Google Scholar]
- Gerlt J. A. & Babbitt P. C. Barrels in pieces? Nat. Struct. Mol. Biol. 8, 5–7 (2001). [DOI] [PubMed] [Google Scholar]
- Yue K. & Dill K. A. Inverse protein folding problem: designing polymer sequences. Proc. Natl Acad. Sci. USA 89, 4163–4167 (1992). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagarajan D., Deka G. & Rao M. Design of symmetric TIM barrel proteins from first principles. BMC Biochem. 16, 18 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolfson D. N. et al. De novo protein design: How do we expand into the universe of possible protein structures? Curr. Opin. Struct. Biol. 33, 16–26 (2015). [DOI] [PubMed] [Google Scholar]
- Winzeler E. A. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 285, 901–906 (1999). [DOI] [PubMed] [Google Scholar]
- Gietz R. D. & Schiestl R. H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2, 31–34 (2007). [DOI] [PubMed] [Google Scholar]
- Sievers F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7, 539 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y., Zhan J., Zhao H. & Zhou Y. A new size-independent score for pairwise protein structure alignment and its application to structure classification and nucleic-acid binding prediction. Proteins 80, 2080–2088 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gromiha M. M., Pujadas G., Magyar C., Selvaraj S. & Simon I. Locating the stabilizing residues in (β/α)8 barrel proteins based on hydrophobicity, long-range interactions, and sequence conservation. Proteins 55, 316–329 (2004). [DOI] [PubMed] [Google Scholar]
- Papadopoulos J. S. & Agarwala R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 23, 1073–1079 (2007). [DOI] [PubMed] [Google Scholar]
- Schneider B. et al. Role of the N-terminal extension of the (βα)-barrel enzyme indole-3-glycerol phosphate synthase for its fold, stability, and catalytic activity. Biochemistry 44, 16405–16412 (2005). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figures and Supplementary Tables
Oligonucleotide sequences for mutant library generation. Primer sequences used for creating the plasmid libraries and processing the deep sequencing samples are available for download.
Data Availability Statement
All data, primer sequences, alignments, and scripts for data analyses are available at https://github.com/yvehchan/TIM_EMPIRIC or from the corresponding author upon request.