

Abstract
The role of transforming growth factor beta-type-1 (TGFβ1) in pancreatic ductal adenocarcinoma (PDAC) progression is stage-dependent. We hypothesized that TGFβ1 expression is associated with survival and proliferation markers in patients with early-stage PDAC. We acquired clinicopathologic, treatment, and mRNA expression data from The Cancer Genome Atlas data set for 106 patients identified with stage I/II PDAC who underwent pancreaticoduodenectomy. Patients were categorized as high expression when mRNA expression was ≥75th percentile for each gene. Average log2 mRNA expression of TGFβ1 in patients with high expression was 11.6 ± 0.2 and 10.5 ± 0.6 in patients with low expression (P<0.001). Low TGFβ1 expression is associated with shorter median survival compared with high TGFβ1 expression (17 versus at least 60 months; P=0.005). Patients with tumors demonstrating high MKI67 (the gene encoding Ki-67) expression have shorter median survival versus those with lowerMKI67 expression (16 versus 20 months; P=0.026). TGFβ1 and MKI67 are inversely associated (P=0.009). On multivariate analysis, improved survival is associated with TGFβ1 overexpression (P=0.017), adjuvant chemotherapy (P=0.001), and adjuvant radiotherapy (P=0.017), whereas positive surgical margins are associated with worse survival (P=0.002). In patients who undergo pancreaticoduodenectomy for PDAC, high TGFβ1 expression may counteract the worse survival associated with high proliferation.
Keywords: pancreatic carcinoma, TGF-β, Ki-67, MKI67
INTRODUCTION
The precise role of transforming growth factor beta type 1 (TGFβ1) in the progression of pancreatic ductal adenocarcinoma (PDAC) is unknown, and its relation to survival is context dependent [1, 2]. This is referred to as the TGFβ paradox, where TGFβ expression in early tumors (AJCC stage I and II) is a favorable characteristic but a detrimental characteristic in advanced-stage III or IV disease [3].
TGFβ is a complex signaling molecule that exists in 3 isoforms and as part of large family of polypeptides [4]. Although it is critical for growth arrest, it is also a growth factor responsible for differentiation in embryonic progenitor cells [5]. Among other signaling cascades, the canonical pathway for activated TGFβ receptors leads to downstream serine phosphorylation of SMAD transcription factors [6]. Important in PDAC, TGFβ signaling induces peritumoral fibrosis by myofibroblast differentiation when exposed to radiation [7] or by increased collagen production in the setting of chronic pancreatitis [8].
Although the role of TGFβ has been better delineated in late-stage PDAC, its role in early-stage PDAC is unclear. In late-stage PDAC, targeted therapeutics against the TGFβ pathway are undergoing clinical trials as data are more mature in this area [9] and suggest that angiogenesis is critical to PDAC tumorigenesis [3, 10]. There are conflicting data in the literature regarding the role of TGFβ in patients with early-stage PDAC. Some evidence has suggested that early-stage tumors are similar to late-stage tumors in terms of a proangiogenic environment [11]. However, other data have suggested that TGFβ blocks tumor cell proliferation [12]. Due to these conflicting data, we investigated the association between TGFB1 and RNA expression of the proliferation marker Ki-67 in PDAC.
Because of its complex signaling cascade, it has been suggested that TGFβ is both a proto-oncogene and tumor suppressor gene, depending on the context [6, 13]. Work by Zhang et al. have demonstrated that TGFβ1 overexpression inhibits cell growth via SMAD3 phosphorylation, hence behaving as a tumor suppressor [3]. Likewise, that group also provided limited data that TGFβ1 overexpression results in SMAD7 activation, which leads to β-catenin/VEGF-A signaling and tumor neo-angiogenesis. Others have demonstrated similar oncogenic behavior of TGFβ, especially in the context of proliferation and inflammation [2, 10].
A potential mechanism for TGFβ1-mediated carcinogenesis is an overwhelming, proliferative peritumoral desmoplastic response [14]. In a non-malignant environment, collagen production by fibroblasts is mediated by TGFβ1 signaling [7, 15, 16]. In the malignant setting, pre-clinical and in vivo data have suggested that collagen deposition by pancreatic stellate cells and pancreatic carcinoma cells both lead to a pro-carcinogenic desmoplastic response [17–19]. In addition, increased proliferation as measured by Ki-67 expression in PDAC cancer cells is associated with therapeutic changes in animal models and early recurrence in retrospective human studies [20–22]. Interestingly, an mRNA transcript leading to type I collagen production, COL1A2, has been associated with proliferative events and markers of aggressive tumor biology, including epithelial to mesenchymal transition [8]. Another group recently demonstrated that the same specific mRNA transcript (COL1A2) may be a serum biomarker for pancreatic carcinoma [19].
We hypothesized that survival in patients with resected early-stage PDAC is associated with TGFβ1 and MKI67 expression (the gene encoding the proliferation marker Ki-67). To control for unknown features of cancer biology, we utilized a strict inclusion/exclusion criteria to identify a relatively homogenous patient population from The Cancer Genome Atlas (TCGA) with stage I and II PDAC who underwent pancreaticoduodenectomy.
RESULTS
Early pancreatic ductal adenocarcinoma in the TCGA data set
Our TCGA data set consisted of 175 tumor samples with pancreatic malignancies. Five of these patients demonstrated histologies not consistent with “typical” PDAC to include neuroendocrine and mucinous colloid carcinomas. Likewise, a minority of patients underwent distal pancreatectomy, total pancreatectomy, or other pancreatic resection (n = 38) and 10 patients had stage III or IV disease.
After applying the inclusion and exclusion criteria, 106 patients were identified for analyses. Of these patients, the average age was 65 ± 11 years, 58% of patients were male, and 91% were white. The median follow-up time was 15 months (range, 0.3-75 months). Seven patients (7%) had a documented family history of PDAC. Only 1 patient received neoadjuvant therapy. In our patient group, 13%, 58%, and 28% of tumors were histologically grade I, II, and III/IV, respectively. The mean number of lymph nodes examined was 18 ± 8 per patient with an average of 3 ± 3 positive lymph nodes per patient. Overall, 60 patients (57%) received a margin-negative (R0) resection, while 56 patients received margin-positive resections (R1 or R2).
TGFβ1 expression in early PDAC from the TCGA
The average expression of TGFβ1 in the top quartile (high expression group) was 11.6 ± 0.2, whereas it was 10.5 ± 0.6 in patients with low expression (P < 0.001, Figure 1A). mRNA expression of MKI67 was 11.2 ± 0.4 in the high expression group and 9.8 ± 0.8 in the low expression group (Figure 1B, P<0.001). Finally, log2 mRNA expression of COL1A2 was 17.5 ± 0.5 in the high expression group and 15.7 ± 1.1 in the low expression group (P<0.001, Figure 1C).
Figure 1. Graphical representation of the log2 mRNA levels of TGFβ1 A. MKI67 B. COL1A2 C.

High TGFβ1 expression is associated with improved overall survival
The clinical and treatment characteristics for patients with tumors expressing high or low levels of TGFβ1 were similar between the two groups, with the exception of radiotherapy (Table 1). Low TGFβ1 expression was associated with median survival of 17 months, whereas median survival was not reached for the high TGFβ1 expression group (at least 60 months, Figure 2; P = 0.007). All long-term survivors (> 40 months) were in the high TGFβ1 expression group. The univaraiate Cox proportion hazard ratio (HR) for high TGFβ1 group was 0.35 (P = 0.009; Table 2). Finally, the mean log2 expression level of TGFβ1 in patients with a family history of PDAC was lower than in patients without a family history of PDAC (10.18 versus 10.84; P = 0.02).
Table 1. Patient and treatment characteristics.
| Characteristic | TGFβ1 Low | TGFβ1 High | P Value |
|---|---|---|---|
| n = 78 | n = 28 | ||
| Age, mean ± SD | 65 ± 11 years | 66 ± 10 years | 0.61 |
| Male, n (%) | 46 (58%) | 16 (59%) | 0.92 |
| AJCC Stage | 0.96 | ||
| T1-2 | 8 (10%) | 5 (18%) | |
| T3 | 70 (90%) | 23 (82%) | |
| N1 | 63 (80%) | 21 (78%) | |
| Family history of pancreatic cancer, n (%) | 7 (9%) | 0 (0%) | 0.11 |
| High-grade tumor, n (%) | 22 (28%) | 8 (29%) | 0.97 |
| Positive surgical margin, n (%) | 34 (44%) | 12 (43%) | 0.9 |
| Radiotherapy, n (%) | 17 (22%) | 15 (57%) | 0.003 |
| Chemotherapy, n (%) | 55 (71%) | 21 (75%) | 0.65 |
Figure 2. Patients with tumors demonstrating TGFβ1 expression ≥75th percentile (high TGFβ1) have longer median survival than other patients (> 60 vs 17 months, P = 0.005).
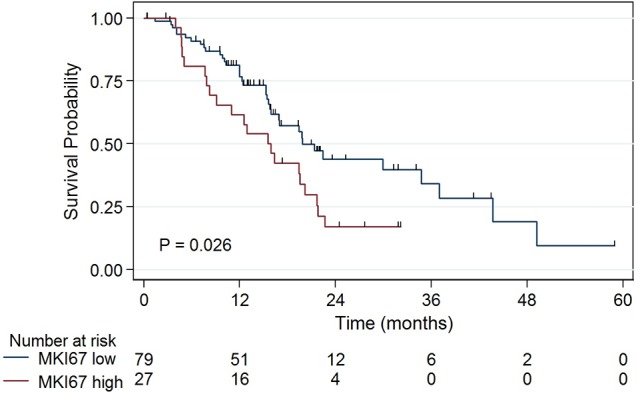
Table 2. Characteristics associated with overall survival.
| Variable | Univariate Cox HR | P Value | Multivariate Cox HR | P Value |
|---|---|---|---|---|
| Age, per year | 1.03 | 0.03 | 1.01 | 0.428 |
| Male gender | 0.67 | 0.13 | ||
| Family history | 2.26 | 0.06 | 3.73 | 0.02 |
| Positive lymph nodes | 2.61 | 0.019 | 3.68 | 0.013 |
| Positive surgical margin | 2.3 | 0.003 | 3.37 | 0.002 |
| High-grade tumor | 1.67 | 0.07 | 1.62 | 0.179 |
| Chemotherapy | 0.39 | 0.001 | 0.30 | 0.001 |
| Radiotherapy | 0.36 | 0.003 | 0.38 | 0.017 |
| High TGFβ1 expression | 0.35 | 0.009 | 0.29 | 0.017 |
| High MKI67 expression | 1.85 | 0.028 | 1.37 | 0.373 |
| High COL1A2 expression | 0.45 | 0.02 | 0.77 | 0.567 |
Interestingly, high mRNA expression of SMAD3, SMAD4, and TGFβR1 were not associated with survival (P=0.87, 0.53, and 0.14, respectively). Likewise, there was no association between the mRNA expressions of these genes with expression of TGFβ1 (each P > 0.1).
Increased proliferation is associated with worse survival
Because Ki-67 is an established marker for cell proliferation in cancer, we investigated its mRNA expression in PDAC. Patients with tumors demonstrating MKI67 expression ≥75th percentile have shorter median survival versus patients with tumors demonstrating lower MKI67 expression (16 months versus 20 months; P = 0.026, Figure 3). Interestingly, patients in the high MKI67 expression group showed lower levels of TGFβ1 expression compared with patients in the low MKI67 expression group (0.07 versus 0.33; P = 0.009). Furthermore, 32% of patients with low tumor TGFβ1 expression have high MKI67 expression, whereas only 7% of patients in the high TGFβ1 expression group have high MKI67 expression (P = 0.009).
Figure 3. Patients with tumors demonstrating MKI67 expression ≥75th percentile have shorter median survival than patients with tumors demonstrating lower MKI67 expression (16 vs 20 months, P = 0.026).
Because there was correlation between TGFβ1 and MKI67 expression, we investigated survival in patients by gene expression status (Table 2). The median survival in the low TGFβ1/low MKI67 expression group was 17 months, whereas patients in the high/high group demonstrated a median survival of 4 months. In the low TGFβ1/high MKI67 expression group, the median survival was 16 months, whereas it was greater than 60 months in the high TGFβ1/low MKI67 expression group (overall P = 0.015). All long-term survivors were in the high/low group.
On multivariate Cox regression analyses, high TGFβ1 expression remained associated with longer survival (Cox HR = 0.29; P = 0.017). Neither MKI67 expression nor COL1A2 expression was independently associated with survival on multivariate Cox regression (Table 2). However, a positive surgical margin remained not only statistically associated with worse survival on multivariate analysis, but it had one of the largest associations with overall survival except for high TGFβ1 expression and a family history of PDAC (Table 2). Importantly, whereas high TGFβ1 expression is associated with radiotherapy (Table 1) and radiotherapy was associated with survival on univariate analysis (Table 2), on multivariate analysis TGFβ1 expression remained statistically significantly associated with longer survival and radiotherapy showed less of an overall effect on survival than high TGFβ1 expression (Cox HR = 0.38, P = 0.017).
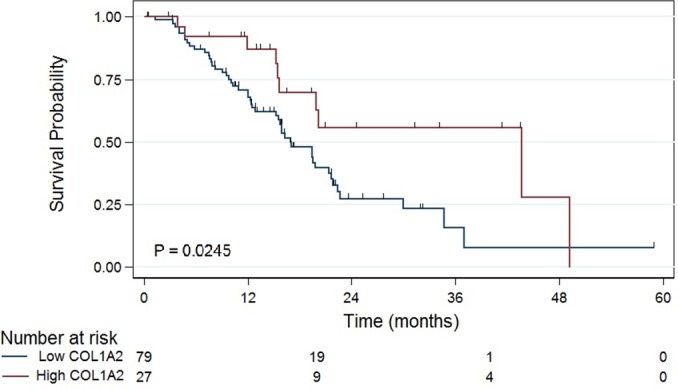
Overexpression of COL1A2, a component of collagen production, is associated with high TGFβ1 expression. We found that 51% of tumors in the high TGFβ1 expression group had high COL1A2 expression, whereas only 24% of tumors in the low TGFβ1 expression group had high COL1A2 expression (P < 0.001). Furthermore, median survival was significantly lower in the low COL1A2 expression group (Figure 4; 17 versus 44 months; P = 0.02). In the low TGFβ1 expression group, high COL1A2 expression was associated with improved survival (Cox HR = 0.39; P = 0.03). There was no association between MKI67 and COL1A2 expression (data not shown; P = 0.3).
Figure 4. High expression of COL1A2 is associated with prolonged survival in patients with resected early-stage pancreatic carcinoma (P = 0.0245).

Of the 106 patients, the overall Kaplan-Meier median survival was 19 months. On univariate analyses, age, positive surgical margins, chemotherapy, radiotherapy, high TGFβ1 expression, high MKI67 expression, high COL1A2 expression, and positive lymph node spread were all statistically associated with survival (Table 2). Radiotherapy was significantly associated with longer survival in margin-positive (R1 or R2) resections (Cox HR = 0.3; P = 0.01), whereas radiotherapy was not significantly associated with longer survival in margin-negative (R0) resection (Cox HR = 0.4; P = 0.07).
DISCUSSION
The TGFβ1 signaling pathway activates transcription factors intimately involved in many normal and benign functions that, when commandeered by PDAC, permit the cancer to be refractory to standard therapy. For example, fibrosis, a critical response to wound healing, also provides PDAC with a mechanical barrier to chemotherapeutic delivery [23, 24]. Adding to the complexity (and perhaps the paradox), work by Jones et al identified over 30 distinct mutations in the TGFβ1 signaling pathway in 24 pancreatic carcinoma tumors, with 100% of tumors containing at least 1 mutation [25]. While this suggests that the TGFβ1 signaling pathway may be extremely important in PDAC carcinogenesis, it also demonstrates that the TGFβ1 signaling pathway may be far more complex than other common mutated pathways.
In this study, our data suggest that the TGFβ1 paradox may be related to proliferation in patients who undergo pancreaticoduodenectomy for PDAC. High TGFβ1 expression may be able to ameliorate the decreased survival associated with highly proliferative tumors that have high levels of Ki-67 mRNA. Alternatively, high levels of Ki-67 may overwhelm tumor suppressor effects of TGFβ1. Importantly, in the multivariate analysis, MKI67 expression was no longer significant when analyzed in conjunction with the other variables in the model, specifically TGFβ1 and radiotherapy, a treatment that targets proliferating cells. High TGFβ1 expression remained statistically significant and has the greatest association with survival except for family history of PDAC and positive lymph node disease (Table 2). While PDAC family history is a non-modifiable factor, positive lymph node disease is the definition of aggressive tumor biology on the macroscopic level. Further investigations are needed to delineate the signaling interactions between other proliferation markers and the TGFβ pathway, which may help further unravel the paradox. Finally, the role of Ki-67 in predicting radiosensitivity needs to be elucidated in PDAC.
We investigated the role of COL1A2 expression by PDAC cells since collagen deposition is often a key component of the desmoplastic response, a component of epithelial to mesenchymal transition, and this specific mRNA transcript may serve as a biomarker for PDAC [8, 19, 26]. Although it has been demonstrated that the peritumoral desmoplasia is created by pancreatic stellate cells [17, 24], we did not identify any evidence in the literature of a relationship between COL1A2 and survival in patients with PDAC. The data here demonstrated a survival advantage associated with high COL1A2 expression in patients with PDAC, but, importantly, this was not significant on multivariate analysis where other patients and tumor characteristics dominated the survival model. Most patients in the high TGFβ1 expression group had high COL1A2 expression and low MKI67 expression, which was not seen in the low TGFβ1 expression group, suggesting the possibility of other interactions not currently investigated. In the low TGFβ1 expression group, high COL1A2 expression was statistically associated with improved survival. It is unclear whether PDAC cells are actually producing collagen and creating a desmoplastic response or whether this is an effect (autocrine or otherwise) to aberrant TGFβ1 signaling pathways.
While analyzing a large data set such as the TCGA is quite useful to investigate the interaction of gene expression and clinical data, there are some significant limitations. The major limitation of this analysis is the inability to interrogate tissue specimens or use patient-derived samples to test hypotheses. Specifically, correlating TGFβ1 serum levels, TGFβ1 tissue levels, and clinical outcomes is a future research target to help exact the relationship between MKI67 expression and TGFβ1 levels. However, these data can serve to outline future research targets, specifically the interaction between cellular proliferation and TGFβ1 pathways. Furthermore, we attempted to limit the effects of the TGFβ1 paradox by using strict inclusion/exclusion criteria to identify patients with “early” disease. However, there is no method to objectively define this patient population other than the self-fulfilling prophecy of investigating patients with only stage I and II PDAC who went to resection. This is selection bias is difficult to quantify.
In summary, our study demonstrates the potential role of proliferation to help understand the TGFβ1 paradox. Patients with early-stage PDAC have longer median survival when TGFβ1 expression is high, which may be due to limited cellular proliferation. Further work is needed to delineate the mechanism of this interaction, investigate it in patients with advanced PDAC, and identify opportunities to modulate the intra- and extracellular TGFβ1 signaling cascade to improve survival and decrease PDAC proliferation.
PATIENTS AND METHODS
Patients and data acquisition
Data from TCGA were downloaded on February 5, 2016. Clinical data, tumor DNA sequence, and mRNA expression data were acquired from the TCGA data set from the data access committee. Except for a single patient, this represents an untreated patient population with PDAC who went directly to surgery for resection. After resection, tumor specimens were dissected from surrounding stroma. mRNA was sequenced with next generation sequencing, usually on the Illuminia platform, as part of the TCGA network with quality controls in place at multiple steps. The TCGA project has been well-described elsewhere [27], but briefly it is a consortium of numerous institutions collaborating to collect, analyze, and integrate data at the genomic, RNA, proteomic, and clinical level for a number of human malignancies.
Inclusion and exclusion criteria
Inclusion criteria were complete data available, pancreatic adenocarcinoma or ductal carcinoma histology, American Joint Committee on Cancer (AJCC) stage I or II, and underwent pancreaticoduodenectomy. Patients with stage III or IV disease based on the 7th edition of the AJCC staging system were excluded.
Expression and gene analyses
Patient clinical data (Level 2.0.46.0) and RNASeqV2 normalized gene expression data (Level 3.1.8.0) were downloaded from the TCGA data portal (https://tcga-data.nci.nih.gov/) open access HTTP directory. Gene expression data were further normalized against the median sample using IRON v2.1.6 (iron_generic --rnaseq --unlog) to correct for the remaining minor differences in dynamic range between samples [28]. High gene expression was based on ≥ 75th percentile log2 mRNA expression value for each gene. Low gene expression was based on values < 75th percentile log2 mRNA expression value for each gene. We specifically investigated the relationship among TGFβ1, MKI67, and COL1A2 and survival in patients with resected PDAC in the head of the pancreas and staged as I or II.
Statistical analyses
Data analyses were performed with STATA 13 (StataCorp LP, College Station, TX). Significance was set to α = 0.05. Differences in variables between the two groups were investigated with Student t test or χ2 as appropriate. Overall survival was investigated with the Kaplan-Meier method utilizing the log-rank test. Censoring occurred for days to last follow-up with the number of patients at risk listed for each time point below the graphs. Variables associated with survival were investigated with univariate and multivariate Cox regression analyses with the proportional hazard ratio (HR). The multivariate survival model was based on clinical and gene expression data as described below utilizing a cut-off of univariate P = 0.1 to be included in the model.
ACKNOWLEDGMENTS AND FUNDING SUPPORT
The authors would like to thank Rasa Hamilton and Kim Murphy for administrative support. The results shown here are based on data generated by the TCGA Research Network: http://cancergenome.nih.gov/. Our study received assistance from the Cancer Informatics Core Facility at the H. Lee Moffitt Cancer Center & Research Institute, an NCI-designated Comprehensive Cancer Center, supported under NIH grant P30-CA76292. Part of this work was presented at the AACR Special Conference on Pancreatic Cancer (May 12-15, 2016, Orlando, FL).
Footnotes
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
- 1.Massague J. TGF beta signalling in context. Nat Rev Mol Cell Biol. 2012;13:616–630. doi: 10.1038/nrm3434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Akhurst RJ, Hata A. Targeting the TGF beta signalling pathway in disease. Nat Rev Drug Discov. 2012;11:790–811. doi: 10.1038/nrd3810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang H, Liu C, Kong Y, Huang H, Wang C, Zhang H. TGFbeta signaling in pancreatic ductal adenocarcinoma. Tumour Biol. 2015;36:1613–1618. doi: 10.1007/s13277-014-2757-4. [DOI] [PubMed] [Google Scholar]
- 4.Heldin CH, Landstrom M, Moustakas A. Mechanism of TGF-beta signaling to growth arrest, apoptosis, and epithelial-mesenchymal transition. Curr Opin Cell Biol. 2009;21:166–176. doi: 10.1016/j.ceb.2009.01.021. [DOI] [PubMed] [Google Scholar]
- 5.Moustakas A, Heldin CH. The regulation of TGFbeta signal transduction. Development. 2009;136:3699–3714. doi: 10.1242/dev.030338. [DOI] [PubMed] [Google Scholar]
- 6.Kubiczkova L, Sedlarikova L, Hajek R, Sevcikova S. TGF-beta - an excellent servant but a bad master. J Transl Med. 2012;10:183. doi: 10.1186/1479-5876-10-183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Judge JL, Owens KM, Pollock SJ, Woeller CF, Thatcher TH, Williams JP, Phipps RP, Sime PJ, Kottmann RM. Ionizing radiation induces myofibroblast differentiation via lactate dehydrogenase. Am J Physiol Lung Cell Mol Physiol. 2015;309:L879–887. doi: 10.1152/ajplung.00153.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shields MA, Ebine K, Sahai V, Kumar K, Siddiqui K, Hwang RF, Grippo PJ, Munshi HG. Snail cooperates with KrasG12D to promote pancreatic fibrosis. Mol Cancer Res. 2013;11:1078–1087. doi: 10.1158/1541-7786.MCR-12-0637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Akinleye A, Iragavarapu C, Furqan M, Cang S, Liu D. Novel agents for advanced pancreatic cancer. Oncotarget. 2015;6:39521–39537. doi: 10.18632/oncotarget.3999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Craven KE, Gore J, Wilson JL, Korc M. Angiogenic gene signature in human pancreatic cancer correlates with TGF-beta and inflammatory transcriptomes. Oncotarget. 2016;7:323–341. doi: 10.18632/oncotarget.6345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gore J, Craven KE, Wilson JL, Cote GA, Cheng M, Nguyen HV, Cramer HM, Sherman S, Korc M. TCGA data and patient-derived orthotopic xenografts highlight pancreatic cancer-associated angiogenesis. Oncotarget. 2015;6:7504–7521. doi: 10.18632/oncotarget.3233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hezel AF, Deshpande V, Zimmerman SM, Contino G, Alagesan B, O'Dell MR, Rivera LB, Harper J, Lonning S, Brekken RA, Bardeesy N. TGF-beta and alphavbeta6 integrin act in a common pathway to suppress pancreatic cancer progression. Cancer Res. 2012;72:4840–4845. doi: 10.1158/0008-5472.CAN-12-0634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Candido J, Hagemann T. Cancer-Related Inflammation. J Clin Immunol. 2013;33:S79–S84. doi: 10.1007/s10875-012-9847-0. [DOI] [PubMed] [Google Scholar]
- 14.Pickup M, Novitskiy S, Moses HL. The roles of TGF beta in the tumour microenvironment. Nat Rev Cancer. 2013;13:788–799. doi: 10.1038/nrc3603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Medina A, Ghahary A. Fibrocytes can be reprogrammed to promote tissue remodeling capacity of dermal fibroblasts. Mol Cell Biochem. 2010;344:11–21. doi: 10.1007/s11010-010-0524-4. [DOI] [PubMed] [Google Scholar]
- 16.Yen TW, Aardal NP, Bronner MP, Thorning DR, Savard CE, Lee SP, Bell RH., Jr Myofibroblasts are responsible for the desmoplastic reaction surrounding human pancreatic carcinomas. Surgery. 2002;131:129–134. doi: 10.1067/msy.2002.119192. [DOI] [PubMed] [Google Scholar]
- 17.Incio J, Suboj P, Chin SM, Vardam-Kaur T, Liu H, Hato T, Babykutty S, Chen I, Deshpande V, Jain RK, Fukumura D. Metformin reduces desmoplasia in pancreatic cancer by reprogramming stellate cells and tumor-associated macrophages. PLoS One. 2015;10:e0141392. doi: 10.1371/journal.pone.0141392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuninty PR, Bojmar L, Tjomsland V, Larsson M, Storm G, Ostman A, Sandstrom P, Prakash J. MicroRNA-199a and -214 as potential therapeutic targets in pancreatic stellate cells in pancreatic tumor. Oncotarget. 2016;7:16396–408. doi: 10.18632/oncotarget.7651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang X, Tong P, Chen J, Pei Z, Zhang X, Chen W, Xu J, Wang J. Identification of novel biomarkers for pancreatic cancer using integrated transcriptomics with functional pathways analysis. J Cell Physiol. 2016 doi: 10.1002/jcp.26066. [DOI] [PubMed] [Google Scholar]
- 20.Kim H, Park CY, Lee JH, Kim JC, Cho CK, Kim HJ. Ki-67 and p53 expression as a predictive marker for early postoperative recurrence in pancreatic head cancer. Ann Surg Treat Res. 2015;88:200–207. doi: 10.4174/astr.2015.88.4.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jamieson NB, Carter CR, McKay CJ, Oien KA. Tissue biomarkers for prognosis in pancreatic ductal adenocarcinoma: a systematic review and meta-analysis. Clin Cancer Res. 2011;17:3316–3331. doi: 10.1158/1078-0432.CCR-10-3284. [DOI] [PubMed] [Google Scholar]
- 22.Glazer ES, Kaluarachchi WD, Massey KL, Zhu C, Curley SA. Bioengineered arginase I increases caspase-3 expression of hepatocellular and pancreatic carcinoma cells despite induction of argininosuccinate synthetase-1. Surgery. 2010;148:310–318. doi: 10.1016/j.surg.2010.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Koerdt S, Rohleder NH, Rommel N, Nobis C, Stoeckelhuber M, Pigorsch S, Duma MN, Wolff KD, Kesting MR. An expression analysis of markers of radiation-induced skin fibrosis and angiogenesis in wound healing disorders of the head and neck. Radiat Oncol. 2015;10:202. doi: 10.1186/s13014-015-0508-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moir JA, Mann J, White SA. The role of pancreatic stellate cells in pancreatic cancer. Surg Oncol. 2015;24:232–238. doi: 10.1016/j.suronc.2015.05.002. [DOI] [PubMed] [Google Scholar]
- 25.Jones S, Zhang X, Parsons DW, Lin JC, Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, Hong SM, Fu B, Lin MT, Calhoun ES, Kamiyama M, Walter K, et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science. 2008;321:1801–1806. doi: 10.1126/science.1164368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kim H, Choi JA, Kim JH. Ras promotes transforming growth factor-beta (TGF-beta)-induced epithelial-mesenchymal transition via a leukotriene B4 receptor-2-linked cascade in mammary epithelial cells. J Biol Chem. 2014;289:22151–22160. doi: 10.1074/jbc.M114.556126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cancer Genome Atlas Research N, Weinstein JN, Collisson EA, Mills GB, Shaw KR, Ozenberger BA, Ellrott K, Shmulevich I, Sander C, Stuart JM. The Cancer Genome Atlas Pan-Cancer analysis project. Nat Genet. 2013;45:1113–1120. doi: 10.1038/ng.2764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Welsh EA, Eschrich SA, Berglund AE, Fenstermacher DA. Iterative rank-order normalization of gene expression microarray data. BMC Bioinformatics. 2013;14:153. doi: 10.1186/1471-2105-14-153. [DOI] [PMC free article] [PubMed] [Google Scholar]