

Abstract
Though fluorine has been shown to induce apoptosis in immune organs in vivo, there has no report on fluoride-induced apoptosis in the cultured lymphocytes. Therefore, this study was conducted with objective of investigating apoptosis induced by sodium fluoride (NaF) and the mechanism behind that in the cultured splenic lymphocytes by flow cytometry, western blot and Hoechst 33258 staining. The splenic lymphocytes were isolated from 3 weeks old male ICR mice and exposed to NaF (0, 100, 200, and 400 μmol/L) in vitro for 24 and 48 h. When compared to control group, flow cytometry assay and Hoechst 33258 staining showed that NaF induced lymphocytes apoptosis, which was promoted by decrease of mitochondria transmembrane potential, up-regulation of Bax, Bak, Fas, FasL, caspase 9, caspase 8, caspase 7, caspase 6 and caspase 3 protein expression (P < 0.05 or P <0.01), and down-regulation of Bcl-2 and Bcl-xL protein expression (P <0.05 or P <0.01). The above-mentioned data suggested that NaF-induced apoptosis in splenic lymphocytes could be mediated by mitochondrial and death receptor pathways.
Keywords: NaF, splenic lymphocytes, apoptosis, Bcl-2 family, caspases, Immunology and Microbiology Section, Immune response, Immunity
INTRODUCTION
Fluorine is one of the essential trace elements for human health [1]. However, excessive fluoride intake can cause serious tissue damage and lead to multiple organ dis-function [2, 3], which depends not only on the concentration and exposed duration [4, 5], but also on the absorption capacity, age, and nutritional status of the individual [6]. It has been demonstrated that fluoride can induce skeletal fluorosis and soft tissue damage [7-9]. Also, our previous studies have proved that fluorine can induce cytotoxicity, immunotoxicity, oxidative damage and pathological injury in the thymus, spleen, bursa of Fabricius, cecal tonsil, liver, kidney, peripheral blood and intestine of broiler chickens [10-36].
In recent years, fluoride has been shown to induce immunotoxicity in vivo and in vitro. Animal studies have showed that fluoride reduces T cell and B cell numbers [27, 37, 38], and increases G0/G1 thymocytes and splenocytes population in young broiler chickens [27, 28]. However, analysis of fluoride effects on cell cycle phases in cultured rat osteoblasts shows an increase number of cells at S phase and a decrease in cells at G2/M phase, while the cells in Go/G1 are not changed [39]. Refsnes et al. [40] have reported that fluoride can induce IL-6 and IL-8 production in human epithelial lung cells, while the concentration of IL-4, IL-6, TNF-α, and IFN-γ in the cecal tonsil and intestinal mucosa of broilers have been found to be decreased in the high dietary fluorine groups in vivo studies [29, 31]. Luo et al. and Liu et al. [13, 16] have reported that IgM and IgG contents in the cecal tonsil and intestines are decreased in high fluorine groups, which demonstrates that high dietary fluorine can impact the humoral immunity. In cecal tonsil of young chickens [30], the percentage of CD3+, CD3+ CD8+ and CD3+ CD4+ T lymphocytes are also decreased in high fluorine groups. Meanwhile, study in vitro has also reported [37] that fluorine ion can suppress T lymphocytes proliferation, IL-2 and IL-6 production, and decrease the CD4+ lymphocytes proportions.
Apoptosis, or programmed cell death, is an essential physiological process that plays a critical role in regulating cell growth and immune response via gene expression and/or protein activity [41, 42]. It has been reported that fluoride can induce cell apoptosis [43]. Our previous studies have found that fluorine can induce apoptosis by altering Bcl-2 and Bax expression in the splenic lymphocytes of broiler chickens [19, 25]. However, the underlying mechanisms of the fluoride-induced apoptosis of lymphocytes are still unknown.
Therefore, in order to reveal possible pathway involved in sodium fluoride (NaF)-induced apoptosis in the immune system, this in vitro study was conducted to investigate the apoptosis and apoptosis-related proteins in the cultured splenic lymphocytes from mice. The results would provide new experimental evidences for understanding the effect mechanism of NaF on splenic immune function.
RESULTS
NaF reduced viability of the splenic lymphocytes
CCK-8 assay was performed to test the cytotoxic effect of NaF on splenic lymphocyte viability. As shown in Figure 1, the viability of splenic lymphocytes was significantly decreased (P < 0.05) at 100-800 μmol/L NaF exposure for 12 h-72 h. And the IC50 value of NaF-decreased cell ability was estimated to be 400 μmol/L NaF for 48 h. Approximately 70% of cells were survived at 400 μmol/L NaF exposure for 24 h. Based on these findings, treatment of 100 (low-dose group, LG), 200 (medial -dose group, MG) and 400 (high-dose group, HG) μmol/L NaF for 24 h and 48 h was selected for the following studies.
Figure 1. Cell viabilities of cultured splenic lymphocytes.
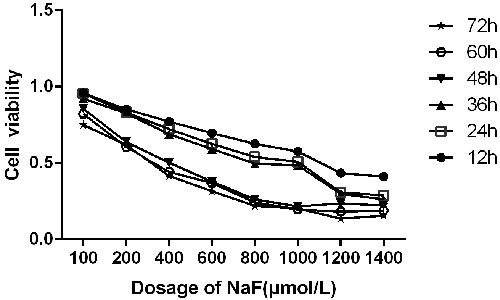
The cell viability was determined using a CCK8 assay as described in the methodology section. The splenic lymphocytes were incubated with different NaF concentrations at different times. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Dispensable of ROS activity in NaF induced cell apoptosis
To determine whether ROS is involved in NaF-induced cell death of splenic lymphocytes, the level of ROS production was measured by using DCFH-DA. Figure 2 and 3 showed that the concentration of NaF did not affect the ROS level in splenic lymphocytes. Overall, NaF induced apoptotic cell death in splenic lymphocytes was not through ROS pathway.
Figure 2. Effect of NaF on ROS generation at 24 h.
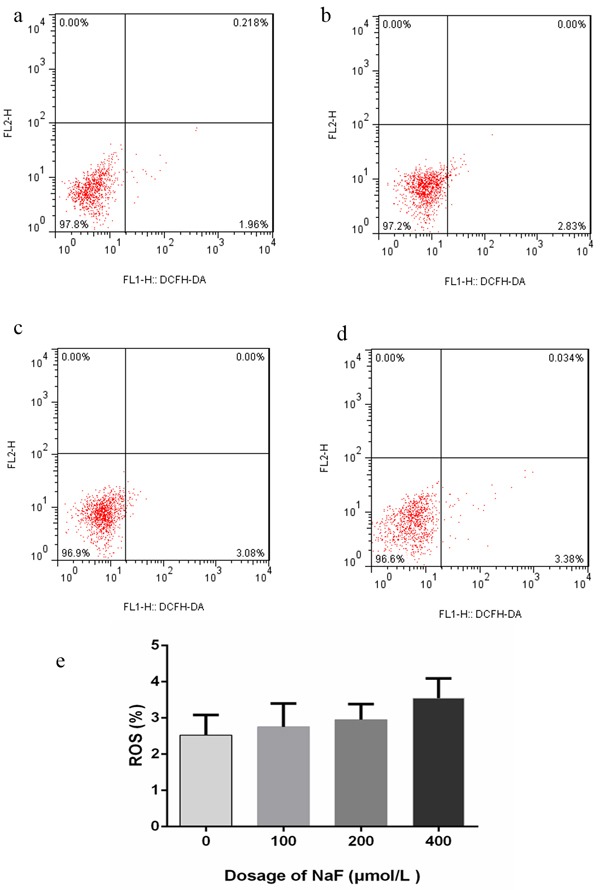
a.-d. Two-dimension scatter plots depicting distribution of cells positively stained for DCFH-DA. (a) CG, (b) LG, (c) MG and (d) HG. e. Quantitative analysis of ROS generation. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 3. Effect of NaF on ROS generation at 48 h.
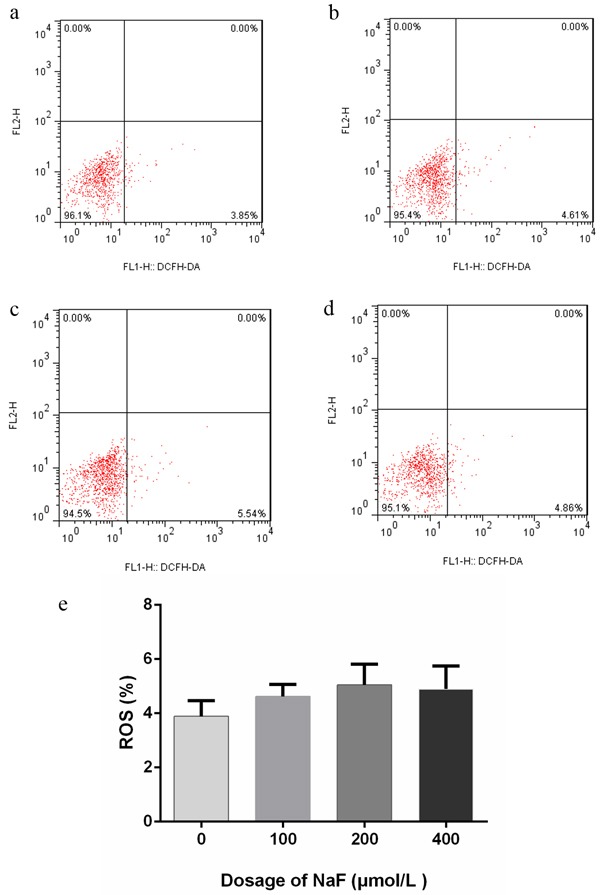
a-d. Two-dimension scatter plots depicting distribution of cells positively stained for DCFH-DA. (a) CG, (b) LG, (c) MG and (d) HG. e. Quantitative analysis of ROS generation. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Effects of NaF on mitochondria transmembrane potential (MMP) in splenic lymphocytes
It was well established that the induction of apoptosis was associated with the perturbation of mitochondrial functions. Here, we examined the changes of MMP by using the fluorescent dyes JC-1. Cells in right upper quadrant correspond to red fluorescence signal intensity, and cells in right lower quadrant correspond to green fluorescence signal intensity. As shown in Figure 4 and 5, the percentages of lymphocytes depolarized with collapse of the Δψm were significantly increased (p < 0.01) in the MG and HG in the presence of NaF for 24 h. Moreover, NaF treatment for 48 h decreased (p < 0.01) the percentage of lymphocytes depolarized with collapse of the Δψm in the LG, MG and HG when compared to CG. Those results suggested that NaF caused the MMP change, which could lead to the apoptosis of splenic lymhoocytes.
Figure 4. Effect of NaF on the MMP Δψm of splenic lymphocytes at 24 h.
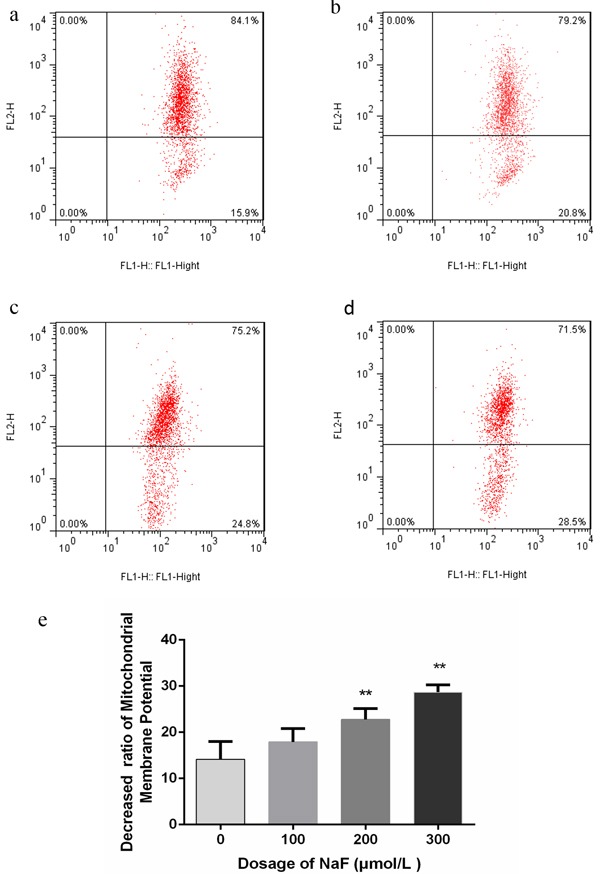
a.-d. Assessment of mitochondrial membrane potential of lymphocytes with JC-1 staining by flow cytometry method. (a) CG, (b) LG, (c) MG and (d) HG. e. Quantitative analysis of the percentage of lymphocytes with green fluorescence. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 5. Effect of NaF on the MMP Δψm of splenic lymphocytes at 48 h.
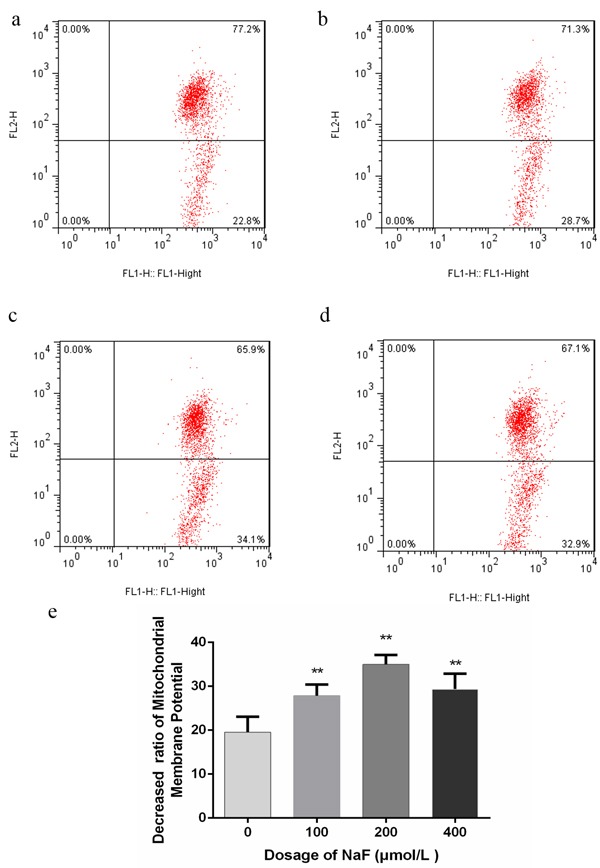
a.-d. Assessment of mitochondrial membrane potential of lymphocytes with JC-1 staining by flow cytometry method. (a) CG, (b) LG, (c) MG and (d) HG. e. Quantitative analysis of the percentage of lymphocytes with green fluorescence. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
NaF caused apoptosis in splenic lymphocytes
The flow cytometry assay showed that apoptotic lymphocytes were significantly increased (p < 0.01) in the MG and HG when compared with those in the LG and CG after NaF treatment for 24 h. And there was no significantly difference between LG and CG. However, with NaF treatment for 48 h, apoptotic lymphocytes were dramatically increased (p < 0.01) among NaF-treated groups and control group, and the apoptosis peaked at MG. The results were shown in Figure 6 and 7.
Figure 6. Effect of NaF treatment on apoptosis of cultured splenic lymphocytes at 24 h.
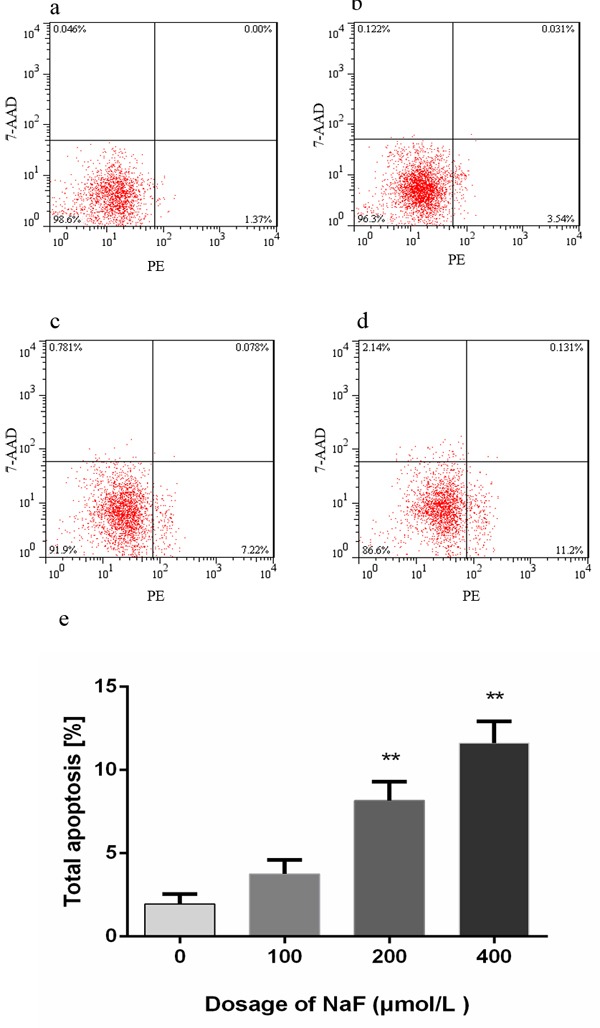
a.-d. Two-dimension scatter plots depicting distribution of cells positively stained for Annexin V-PE/7-AAD. (a) CG, (b) LG, (c) MG and (d) HG. Cells in lower left quadrant of each picture correspond to normal cells. Cells in right lower quadrant correspond to early apoptotic cells. Cells in right upper quadrant correspond to late apoptotic. Cells in left upper quadrant correspond to dead cells. e. Quantitative analysis of total apoptotic lymphocytes. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 7. Effect of NaF treatment on apoptosis of cultured splenic lymphocytes at 48 h.
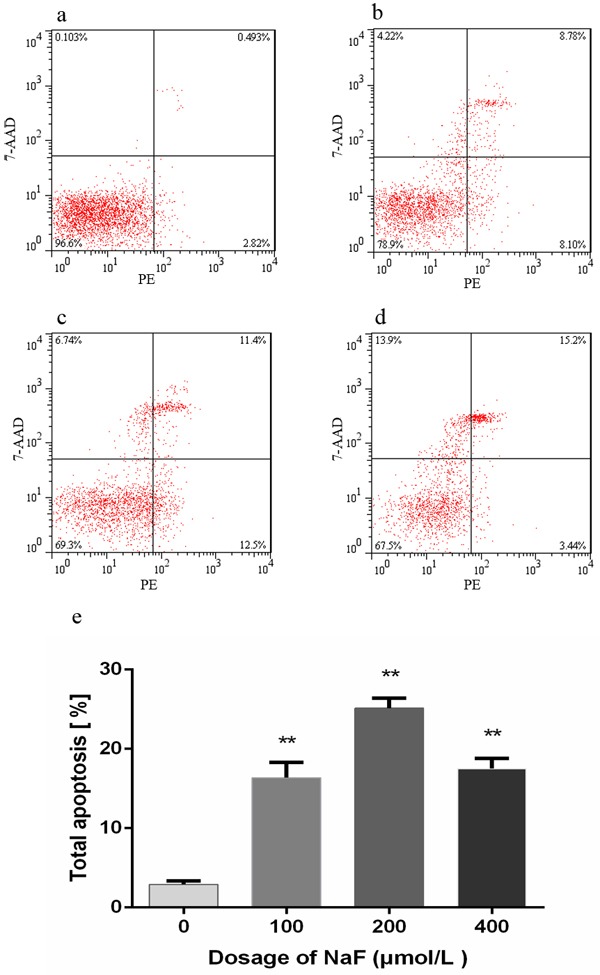
a.-d. Two-dimension scatter plots depicting distribution of cells positively stained for Annexin V-PE/7-AAD. (a) CG, (b) LG, (c) MG and (d) HG. Cells in lower left quadrant of each picture correspond to normal cells. Cells in right lower quadrant correspond to early apoptotic cells. Cells in right upper quadrant correspond to late apoptotic. Cells in left upper quadrant correspond to dead cells. e. Quantitative analysis of total apoptotic lymphocytes. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
NaF caused morphologic changes of apoptosis in splenic lymphocytes
As shown in Figure 8 and 9, cells treated with NaF for 24 h and 48 h showed the characteristic features of apoptosis under a light microscope, including nuclear shrinkage and chromatin fragmentation. Furthermore, the survival of the cells was significantly reduced. The number of apoptotic lymphocytes were significantly higher (p < 0.01) in the MG and HG than the LG and CG treated with NaF for 24 h. Also, exposed to NaF for 48 h, apoptotic lymphocytes were dramatically increased (p < 0.01) among NaF-treated groups and control group. Therefore, the short time treatment of NaF caused the typical apoptosis featured morphologic changes of splenic lymphocytes.
Figure 8. Effect of NaF treatment on morphologic changes of apoptosis in cultured splenic lymphocytes at 24 h.
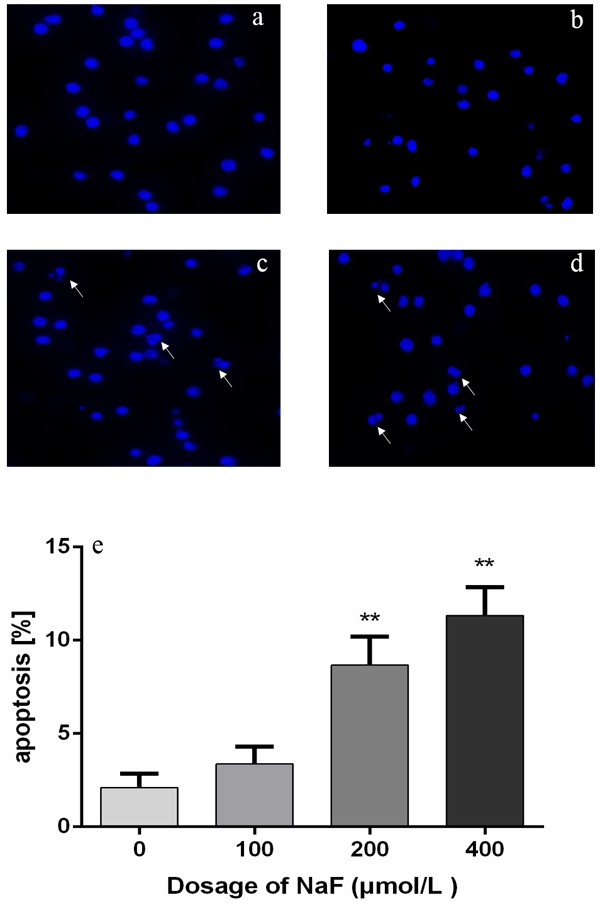
a.-d. the nuclear morphology of lymphocytes stained with Hoechst 33258. (a) CG, (b) LG, (c) MG and (d) HG. Cells are observed under fuorescence microscope. × 1000. e. Quantitative analysis of apoptotic lymphocytes. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 9. Effect of NaF treatment on morphologic changes of apoptosis in cultured splenic lymphocytes at 48 h.
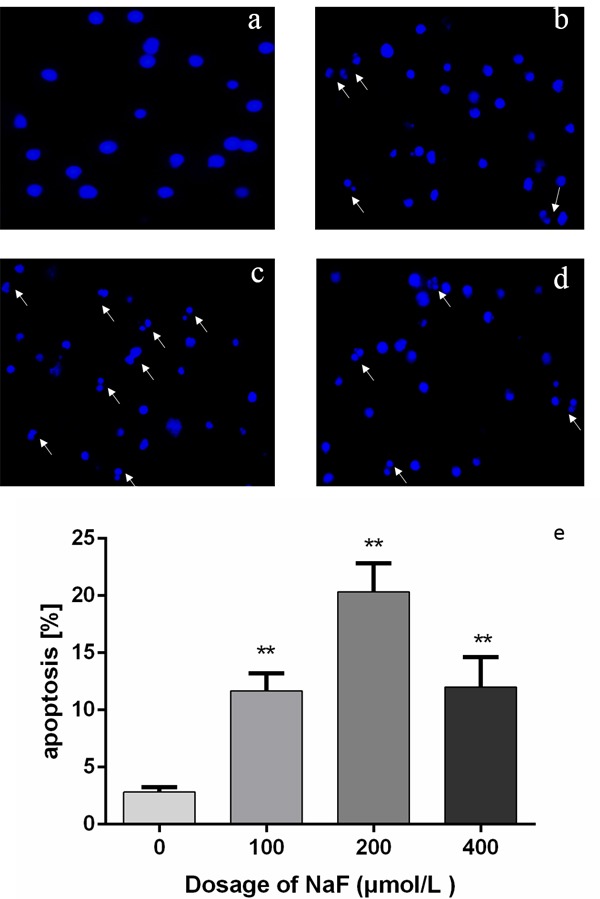
a.-d. the nuclear morphology of lymphocytes stained with Hoechst 33258. (a) CG, (b) LG, (c) MG and (d) HG. Cells are observed under fuorescence microscope. × 1000. e. Quantitative analysis of apoptotic lymphocytes. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Effects of NaF on caspase-9, -8, -7, -6 and -3 protein expression in splenic lymphocytes
To further investigate the potential mechanisms involved in splenic lymphocytes apoptosis induced by NaF, protein expression levels of caspase-9, -8, -7, -6, -3 and -3 were measured by western blot. After NaF treatment for 24 h, the caspase-9, -8, -7, -6 and -3 protein expression levels were significantly increased (p < 0.01) in the MG and HG than those in the CG. After NaF treatment for 48 h, caspase-9, -8, -7, -6, -3 protein expression levels (p < 0.01 or p < 0.05) were significantly increased in the LG, MG and HG, and peaked in the MG when compared with those in the CG, which was consistent with the results of flow cytometry and Hoechst 33258 staining assay. The results were shown in Figure 10 and 11. Collectively, those data suggested that NaF-induced apoptosis could mediated by up-regulation of the above caspase family proteins.
Figure 10. Effect of NaF treatment on protein expression levels of caspase-9, -8, -7, -6 and -3 in cultured splenic lymphocytes at 24 h.
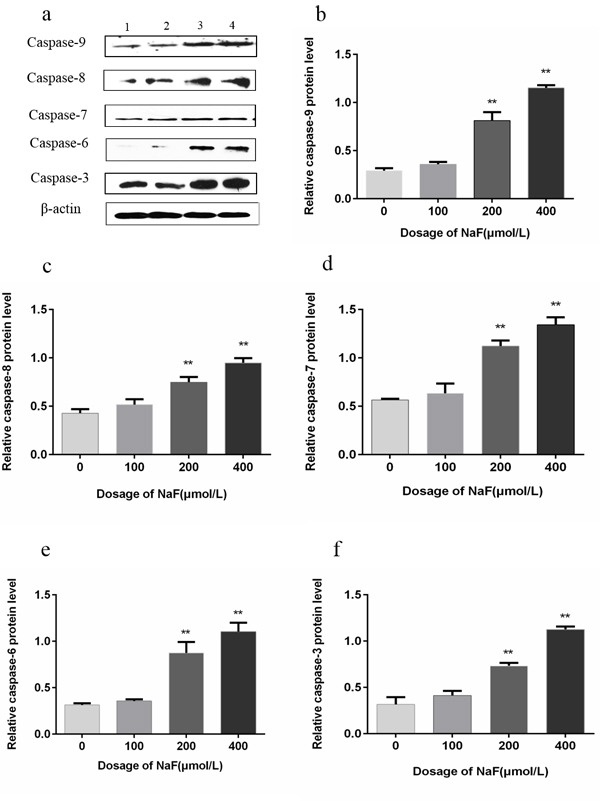
a. The western blot assay. (1) CG, (2) LG, (3) LG and (4) HG. b.-f. Quantitative analysis of caspase-related proteins expression. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 11. Effect of NaF treatment on protein expression levels of caspase-9, -8, -7, -6 and -3 in cultured splenic lymphocytes at 48 h.
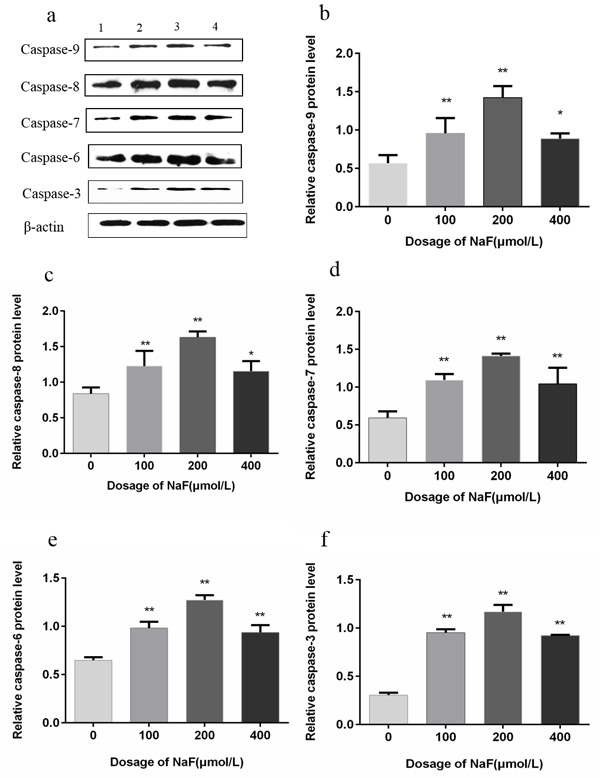
a. The western blot assay. (1) CG, (2) LG, (3) MG and (4) HG. b.-f. Quantitative analysis of caspase-related proteins expression. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
The effects of NaF on Bcl-2, Bcl-xL, Bax and Bak protein expression in splenic lymphocytes
The Bcl-2 family members play an important role in cell apoptosis and survival. As shown in Figure 12, Bax and Bak protein expression levels were significantly increased (p < 0.01) in the MG and HG in comparison with control group after NaF treatment for 24 h. Bcl-2 protein expression levels were significantly reduced (p < 0.01) in the MG and HG, and Bcl-xL protein levels were decreased (p < 0.01) in the HG. Also, Bax/Bcl-2 ratio was markedly increased (p < 0.01) in the MG and HG after NaF treatment for 24 h. Meanwhile, protein expression levels of Bax and Bak were higher (p < 0.01) in the LG, MG and HG than those in the CG after NaF treatment for 48 h. Bcl-2 and Bcl-xL protein expression levels were decreased (p < 0.01) in the LG, MG and HG (Figure 13). The Bax/Bcl-2 ratio was also increased (p < 0.01) in the NaF-treated groups after NaF treatment for 48 h.
Figure 12. Effect of NaF treatment on protein expression levels of Bcl-2, Bcl-xL, Bax and Bak in cultured splenic lymphocytes at 24h.
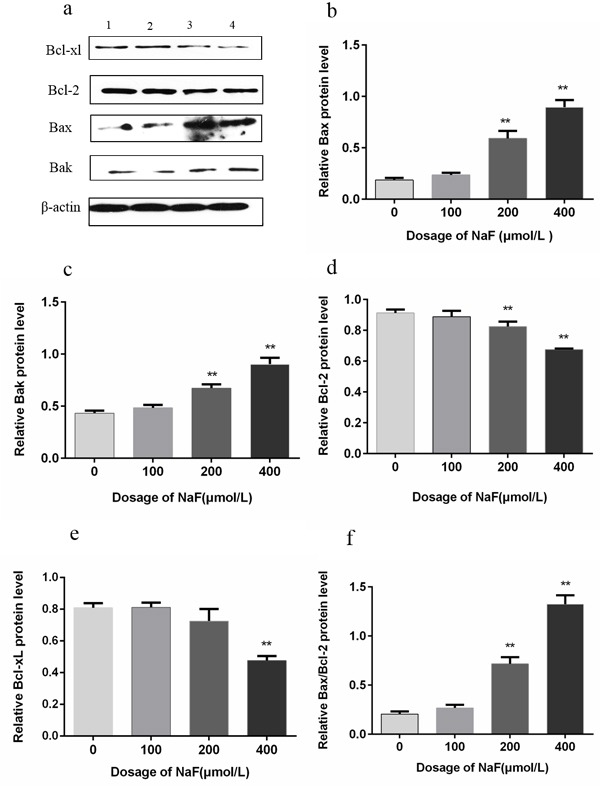
a. The western blot assay. (1) CG, (2) LG, (3) MG and (4) HG. b.-f. Quantitative analysis of caspase-related proteins expression. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Figure 13. Effect of NaF treatment on protein expression levels of Bcl-2, Bcl-xL, Bax and Bak in cultured splenic lymphocytes at 48 h.
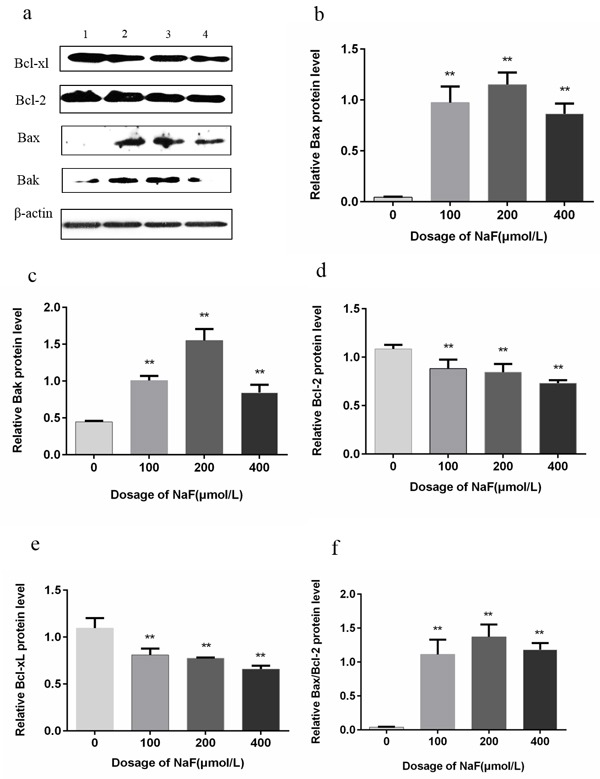
a. The western blot assay. (1) CG, (2) LG, (3) MG and (4) HG. b.-f. Quantitative analysis of caspase-related proteins expression. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
Effects of NaF on Fas and FasL protein expression in splenic lymphocytes
Fas and FasL are important molecules in the extrinsic apoptosis pathway, As shown in Figure 14a-14c, the protein expression levels of Fas and FasL were significantly higher (p < 0.01 or p < 0.05) in the MG and HG than those in the CG after NaF treatment for 24 h. After NaF treatment for 48 h, the Fas and FasL protein expression levels were significantly increased (p < 0.01) in the LG, MG and HG, and peaked in the MG when compared with those in the CG (Figure 14d-14f). Thus, the NaF caused the cell apoptosis through death receptor Fas/FasL pathway.
Figure 14. Effect of NaF treatment on protein expression levels of Fas and FasL in cultured splenic lymphocytes at 24 (a-c) and 48 h (d-f).
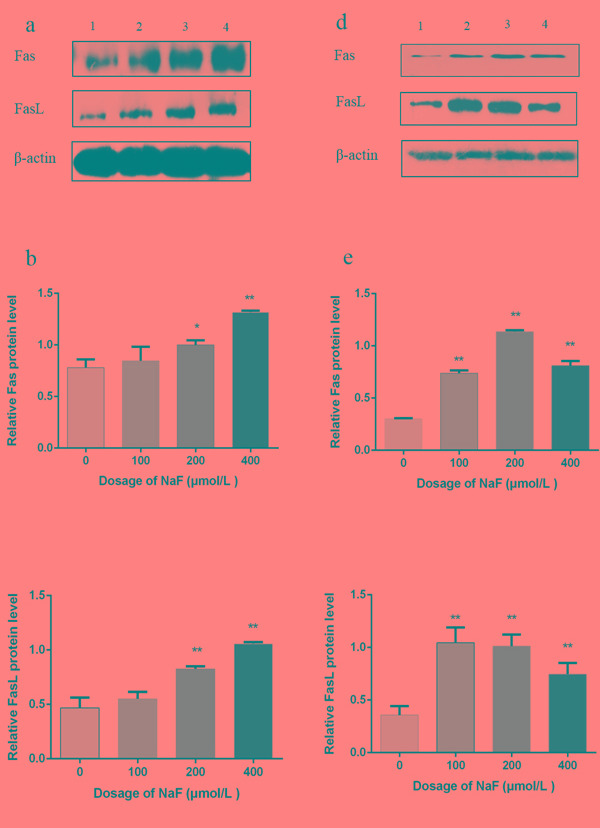
a., d. The western blot assay. (1) CG, (2) LG, (3) MG and (4) HG. b., c., e., f. Quantitative analysis of caspase-related proteins expression. Data are presented with the means ± standard deviation, * p < 0.05, ** p < 0.01, compared with the control group. Data were analyzed by the variance (ANOVA) test of the SPSS 19.0 software.
DISCUSSION
It is well known that fluoride have diverse effects on cells. In bone cells, fluorides elicited potentially beneficial effects by stimulating bone cell growth [6]. However, in other cultured cells, NaF can alter cellular metabolism [44], induce inflammatory cytokines production [40], inhibit protein secretion and synthesis [45], and influence signaling pathways involved in proliferation and apoptosis [46-48]. In this in vitro study, flow cytometry assay and Hoechst 33258 staining showed that NaF induced apoptosis in splenic lymphocytes from mice. It is well known that up-regulation of pro-apoptotic or down-regulation of anti-apoptotic proteins can lead to cell death. To reveal the pathway involved in NaF-induced apoptosis, we detected the ROS production, mitochondria transmembrane potential (MMP), protein expression of caspases, Fas, FasL and Bcl-2 family members.
Previous report has demonstrated that oxidative stress is induced in fluoride group in human blood lymphocytes and lead apoptosis [49]. However, our results showed that NaF did not change intracellular concentration of ROS, indicating that NaF-induced apoptosis of mouse splenic lymphocytes was independent of ROS production.
The mitochondrial pathway, which is controlled by Bcl-2 family members, is the most important intracellular apoptosis signaling cascade. Bcl-2 family, as a group of crucial regulatory factors in apoptosis, can indirectly regulate the activity of caspases in related apoptotic pathways [50]. Members of Bcl-2 family are classified into two groups: pro-apoptotic proteins and anti-apoptotic proteins. Pro-apoptotic proteins such as Bax and Bak induce apoptosis by causing loss of the mitochondrial membrane potential [51, 52]. Anti-apoptotic proteins such as Bcl-2 and Bcl-xL can maintain the normal permeability of mitochondrial permeability transition pore (MPTPs) and block the release of mitochondrial proapoptotic factor by binding selectively to the active conformation of Bax. Thus, the anti-apoptosis protein prevents cell apoptosis [52-54]. Yang et al. has reported that fluoride induce apoptosis in matured ameloblast-like LS8 cells by down-regulating Bcl-2 [55]. The similar picture has been observed in the NaF-treated renal tubules [56] and osteoblastic MC3T3-E1 cells [57], where the apoptosis induced by NaF is also associated with the down-regulation of Bcl-2 and up-regulation of Bax expression at both the mRNA and protein levels. In the present study, Bcl-2 and Bcl-xL was shown to be down-regulated whereas Bax and Bak were up-regulated in lymphocytes treated with NaF and an increase in the Bax/Bcl-2 ratio was also observed.
The mitochondrial-mediated apoptotic pathway is accompanied by ΔΨm depolarization, followed by cytochrome c release from mitochondria into the cytosol. After cytochrome c is released from mitochondria to the cytosol, caspase-9 is subsequently activated through binding to the CED-4 homolog Apaf-1 [58], and then caspase-3, -6 and -7 are activated [59]. Once executioner caspases such as caspase-3, -6 and -7 are activated, they subsequently cleave distinct cellular proteins to cause apoptosis [42, 60]. In the present paper, a decrease in ΔΨm was detected in the NaF-treated cells, together with the increased protein expression of caspase-3, -6, -7 and -9. Based on the above-mentioned data, mitochondria dependent apoptotic pathway may be involved in NaF-induced apoptosis in lymphocytes.
Most recent researches regarding apoptotic signaling cascades support a model in which two caspase generated pathways ensure cell death [61]. In the extrinsic apoptosis pathway, interaction of Fas and FasL lead to the activation of Caspase-8 [62]. Activated Caspase-8 can directly cleave and activate downstream effector proteases, such as Caspase-3, -6, -7, causing apoptosis [63]. In order to assess whether there is any role of the extrinsic apoptosis pathway in the activation of caspase-3, which is a key factor in apoptosis execution of both extrinsic and intrinsic pathways, we measured the protein levels of Fas, FasL and caspase-8. We found that they were also significantly up-regulated after NaF treatment. Therefore, the present study demonstrates that death receptor-mediated pathway may be also involved in NaF-induced apoptosis of lymphocytes. These results further suggest that NaF-induced apoptosis may occur via both mitochondria- and death receptor-dependent apoptosis pathways.
The spleen is a peripheral immune organ and plays an important role in maintaining immune homeostasis [64-66]. It is mainly composed of lymphocytes, which are involved in cellular immunity and humoral immunity. This in vitro study has proved that NaF-induced apoptosis in splenic lymphocytes, which impacts lymphocyte numbers and activities. Thus, the splenic immune function has finally been impaired due to the increased lymphocyte apoptosis and decreased lymphocyte numbers and activities.
MATERIALS AND METHODS
Chemicals and supplies
NaF (S6776) was purchased from Sigma Aldrich, UK. Lymphocyte separation medium (DKW33-R0100) were supplied by Dakewe Biotech Company, China. JC-1 mitochondrial membrane potential assay kit (C2006), ROS Assay kit (S0033), RIPA lysis buffer (P0013C), BCA Protein Assay Kit (P0012), Cell Counting Kit-8 (CCK-8) (C0038) and Hoechst 33258 (C1011) were obtained from Beyotime Biotechnology, China. RPMI 1640 (11875119) and fetal bovine serum (16000044) were supplied by Gibco, UK. The mouse Bc1-2 (3498T), Bcl-xl (2764T), Bax (14796S), Bak (12105T), caspase-9 (9508T), cleaved caspase-8 (8592T), cleaved caspase-7 (8438S), cleaved caspase-6 (9761T), cleaved caspase-3 (9664T) antibodies, rabbit IgG (7074P2) and mouse IgG (7076P2) were obtained from Cell Signaling Technology, UK. Fas (ab82419) and FasL (ab16104) were purchased from Abcam, UK. All other reagents used were analytical grade.
Lymphocyte isolation, culture and treatment
The ICR mice were obtained from the Experimental Animal Corporation of DOSSY Biological Technology Company. 3-week-old ICR male mice were anaesthetized and euthanized. After laparotomy, the spleen was separated from the mice and washed with cold phosphate buffered saline (PBS, pH 7.4). Then, the spleen was placed in a 200-mesh stain steel sieve over a culture dish containing 4-5 mL lymphocyte isolation separation medium and grounded into small pieces with the plunger of glass syringe. The liquid was transferred into a centrifuge tube, and then 200-500 μL RPMI-1640 medium was added and centrifuged at 800×g for 30 min at room temperature. Three layers were formed after centrifugation. The middle milky layer which contained lymphocytes was transferred into a test tube. The lymphocytes were washed twice with PBS and suspended in RPMI-1640 medium with 10 % fetal calf serum and then transferred into a culture bottle. All these processes were performed under sterile condition. The viability of the lymphocyte was estimated according to the trypan blue exclusion criteria, and the viability is over 95%.
To monitor various parameters (except the CCK-8 bioas-say), splenic lymphocytes were cultured in the RPMI-1640 medium (supplemented with 10% fetal calf serum, 100 U/mL penicillin, 100 μg/mL streptomycin) containing 0 (control group, CG), 100(low-dose group, LG), 200 (medial-dose group, MG), and 400 (high-dose group, HG) μmol/L NaF. Triplicates were performed in each treatment. All cells were maintained in a humidified incubator for 24 and 48 h at 37 °C with 5% CO2.
Our experiments involving the use of mice and all experimental procedures were approved by Animal Care and Use Committee, Sichuan Agricultural University.
Measurement of cell viability
Cell viability was assessed using the CCK-8 bioassay [67, 68]. Splenic lymphocytes (7×105/well) were seeded into 96-well flat-bottomed plates and were exposed to NaF (0-1400 μmol/L) for 12 h, 24 h, 36 h, 48 h and 72 h at 37 °C with 5% CO2. After NaF exposure, 10 μL CCK-8 solutions were added to each well, and incubated for 4 h. The optical density (OD) was measured at 450 nm using a microplate reader.
Detection of ROS production
The ROS production was evaluated by using an ROS detection Kit according to the manufacturer's protocol. Cells were collected by centrifugation and washed in PBS at 800 g for 5 min, and then resuspended in 100 μl PBS. Thereafter, cells stained with 10 μM DCFH-DA for 20 min at 37°C. Then, 400 μL PBS was added. The Rosup (a compound mixture) was used as a positive control. Data were then obtained by a FACSCalibur (Becton Dickinson, USA).
Measurement of mitochondria transmembrane potential (MMP)
JC-1 was used to determine mitochondrial membrane potential (Δψm). Cells were collected and transferred into 5 mL culture tube, and then centrifuged. Afterwards, 0.5 mL 1×JC-1 working solution was added and then incubated in the mixture for 20 min at 37 °C under 5% CO2 incubator. At the end of the incubations, cells were washed twice with 1× JC-1 Assay Buffer, and then resuspended each pellet in 450 μL 1× JC-1Assay Buffer. Finally, Δψm was assayed by FCM within 30 minutes [69].
Measurement of apoptosis
The percentage of apoptosis was evaluated by an Annexin V-PE/7-AAD staining detection Kit (BD Biosciences, San Jose, CA, USA) according to the manufacturer's protocol. Cells were cultured in the presence of 0, 100, 200 and 400 μmol/L NaF for 24 h and 48 h. Prior to toxicity detection, the cells were collected and washed two times with phosphate buffered saline (PBS, PH 7.4). Pellets were collected and resuspended in 100 μL PBS, and stained with PE Annexin V and 7-amino-actinomycin (7-AAD) for 15 minutes in the dark. Then, 400 μL binding buffer (BD Pharmingen) was added. Data were then obtained by a FACSCalibur (Becton Dickinson, USA).
Hoechst 33258 staining
Cells were collected in 1.5 ml centrifuge tubes, washed with PBS and fixed with 4% paraformaldehyde for 10 min. And then centrifuged at 2000×g for 5 min. Cells were then washed with PBS for two times again and stained with Hoechst 33258 (Beyotime, Jiangsu, China) at 10 μg/mL for 10 minutes in dark. After washed with PBS, the cells were added to the glass slide and then covered with a clean cover glass. Nuclear morphology was observed using fluorescence microscopy. Apoptotic cells were identified by the reduction in the volume, chromatin condensation and fragmentation of their nuclei [70-72]. The percentage of apoptotic cells was calculated from ten randomly selected microscopic fields (1000×magnification).
Western blot analysis
Cells were lysed and proteins were extracted with RIPA lysis buffer and then kept in laemmli buffer. Protein samples were separated by SDS-PAGE (10%-15% gels) and transferred to nitrocellulose filter membranes. The membranes were blocked in 5% nonfat dry milk for 1 h and then incubated with the primary antibodies overnight at 4°C. The primary antibodies were Bax, Bak, Bcl-2, Bcl-xL, Fas, FasL and caspase-3, -6, -7, -8, -9. The membranes were then washed with PBS-Tween (PBST) and incubated with the biotin-conjugated secondary antibodies for 1 h, and washed again with PBS-Tween (PBST). Blots were visualized by ECL™ (Bio-Rad) and X-ray film. When the band is thicker and darker, protein expression is higher, otherwise it is lower. Then, the statistical data of protein expression was done with ImageJ2x software
Statistical analysis
All the data were analyzed by SPSS 19.0. All the results were expressed as mean±SD. Data were analyzed by one way analysis of variance (ANOVA). p < 0.05 was considered as a significant difference.
Conclusions
In summary, 200 μmol/L and 400 μmol/L NaF induce the cultured splenic lymphocytes apoptosis, which is promoted by up-regulating of Bax, Bak, caspase-9, caspase-8, caspase-7, caspase-6 and caspase-3 protein expression, and down-regulating of mitochondria membrane potential, Bcl-2 and Bcl-xL protein expression. Also, the effect of NaF on cultured splenic lymphocytes apoptosis shows time and dose dependent.
Acknowledgments
The study was supported by the program for Changjiang scholars and innovative research team in university (IRT 0848) and the Shuangzhi project of Sichuan Agricultural University (03570327; 03571198).
Footnotes
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
REFERENCES
- 1.Guo HS, Rui LW, Zhao YC, Zhang B, Hai LW, Mao LL, Ji PG, Xiao YY. Toxic effects of sodium fluoride on cell proliferation and apoptosis of Leydig cells from young mice. J Physiol Biochem. 2014;70:761–768. doi: 10.1007/s13105-014-0344-1. [DOI] [PubMed] [Google Scholar]
- 2.Ersoy IH, Alanoglu EG, Koroglu BK, Varol S, Akcay S, Ugan Y, Ersoy S, Tamer MN. Effect of endemic fluorosis on hematological parameters. BiolTrace ElemRes. 2010;138:22–27. doi: 10.1007/s12011-009-8595-3. [DOI] [PubMed] [Google Scholar]
- 3.Baszczyk I, Birkner E, Gutowska I, Romuk E, Chlubek D. Influence of methionine and vitamin E on fluoride concentration in bones and teeth of rats exposed to sodium fluoride in drinking water. BiolTrace Elem Res. 2012;146:335–339. doi: 10.1007/s12011-011-9251-2. [DOI] [PubMed] [Google Scholar]
- 4.Qu WJ, Zhong DB, Wu PF, Wang JF, Han B. Sodium fluoride modulates caprine osteoblast proliferation and differentiation. J Bone Miner Metab. 2008;26:328–334. doi: 10.1007/s00774-007-0832-2. [DOI] [PubMed] [Google Scholar]
- 5.Wang SH, Shih YL, Ko WC, Wei YH, Shih CM. Cadmium-induced autophagy and apoptosis are mediated by a calcium signaling pathway. Cellular and molecular life sciences. 2008;65:3640–3652. doi: 10.1007/s00018-008-8383-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Caverzasio J, Palmer G, Bonjour JP. Fluoride: mode of action. Bone. 1998;22:585–589. doi: 10.1016/s8756-3282(98)00058-1. [DOI] [PubMed] [Google Scholar]
- 7.Ba Y, Zhu JY, Yang YJ, Yu B, Huang H, Wang G, Ren LJ, Cheng XM, Cui LX, Zhang YW. Serum calciotropic hormone levels, and dental fluorisis in children exposed to different concentrations of fluoride and iodine in drinking water. Chin Med J (Engl) 2010;123:675–679. [PubMed] [Google Scholar]
- 8.Everett E. Fluoride's effects on the formation of teeth and bones, and the influence of genetics. J Dent Res. 2011;90:552–560. doi: 10.1177/0022034510384626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gazzano E, Bergandi L, Riganti C, Aldieri E, Doublier S, Costamagna C, Bosia A, Ghigo D. Fluoride effects: the two faces of janus. Curr Med Chem. 2010;17:2431–2441. doi: 10.2174/092986710791698503. [DOI] [PubMed] [Google Scholar]
- 10.Luo Q, Cui H, Peng X, Fang J, Zuo Z, Liu J, Wu B, Deng Y. The association between cytokines and intestinal mucosal immunity among broilers fed on diets supplemented with fluorine. Biol Trace Elem Res. 2013;25:37–42. doi: 10.1007/s12011-013-9612-0. [DOI] [PubMed] [Google Scholar]
- 11.Chen T, Cui H, Cui Y, Bai C, Gong T, Peng X. Cell-cycle blockage associated with increased apoptotic cells in the thymus of chickens fed on diets high in fluorine. Hum Exp Toxicol. 2011;30:685–692. doi: 10.1177/0960327110379022. [DOI] [PubMed] [Google Scholar]
- 12.Luo Q, Cui H, Peng X, Fang J, Zuo Z, Deng J, Liu J, Deng Y. Suppressive effects of dietary high fluorine on the intestinal development in broilers. Biol Trace Elem Res. 2013;156:153–165. doi: 10.1007/s12011-013-9845-y. [DOI] [PubMed] [Google Scholar]
- 13.Luo Q, Cui H, Peng X, Fang J, Zuo Z, Deng J, Liu J, Deng Y. Intestinal IgA+ Cell Numbers as well as IgA, IgG, and IgM Contents Correlate with Mucosal Humoral Immunity of Broilers During Supplementation with High Fluorine in the Diets. Biol Trace Elem Res. 2013;154:62–72. doi: 10.1007/s12011-013-9713-9. [DOI] [PubMed] [Google Scholar]
- 14.Luo Q, Cui H, Peng X, Fang J, Zuo Z, Liu J, Wu B, Wang H, Deng Y, Huang J. Intestinal oxidative stress in broilers caused by high dietary fluorine. Fluoride. 2012;45:349–356. [Google Scholar]
- 15.Liu J, Cui H, Peng X, Fang J, Zuo Z. Changes Induced by High Dietary Fluorine in the Cecal TONSIL Cytokine Content of Broilers. Fluoride. 2012;45:94–99. [Google Scholar]
- 16.Liu J, Cui H, Peng X, Fang J, Zuo Z, Deng J, Wang H, Wu B, Deng Y, Wang K. Decreased IgA+ B cells population and IgA, IgG, IgM contents of the cecal tonsil induced by dietary high fluorine in broilers. Int J Environ Res Public Health. 2013;10:1775–1785. doi: 10.3390/ijerph10051775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen T, Cui Y, Bai C, Gong T, Peng X, Cui H. Decreased Percentages of the Peripheral Blood T-Cell Subsets and the Serum IL-2 Contents in Chickens Fed on Diets Excess in Fluorine. SocFor. 2009;87:2228–2230. doi: 10.1007/s12011-009-8373-2. [DOI] [PubMed] [Google Scholar]
- 18.Chen T, Cui H, Cui Y, Bai C, Gong T. Decreased antioxidase activities and oxidative stress in the spleen of chickens fed on high-fluorine diets. Hum Exp Toxicol. 2011;30:1282–1286. doi: 10.1177/0960327110388538. [DOI] [PubMed] [Google Scholar]
- 19.Liu J, Cui H, Peng X, Fang J, Zuo Z, Wang H, Wu B, Deng Y, Wang K. Dietary high fluorine induces apoptosis and alters Bcl-2, Bax, and caspase-3 protein expression in the cecal tonsil lymphocytes of broilers. Biol Trace Elem Res. 2013;152:25–30. doi: 10.1007/s12011-012-9595-2. [DOI] [PubMed] [Google Scholar]
- 20.Bai C, Chen T, Cui Y, Gong T, Peng X, Cui H. Effect of high fluorine on the cell cycle and apoptosis of renal cells in chickens. Biol Trace Elem Res. 2010;138:173–180. doi: 10.1007/s12011-009-8599-z. [DOI] [PubMed] [Google Scholar]
- 21.Deng Y, Cui H, Peng X, Fang J, Zuo Z, Deng J, Luo Q. Effects of High Dietary Fluoride on Serum Biochemistry and Oxidative Stress Parameters in Broiler Chickens. Health. 2014;06:1840–1848. [Google Scholar]
- 22.Deng Y, Cui H, Peng X, Fang J, Zuo Z, Deng J, Luo Q. Effects of high dietary fluorine on erythrocytes and erythrocyte immune adherence function in broiler chickens. Biol Trace Elem Res. 2013;155:247–252. doi: 10.1007/s12011-013-9793-6. [DOI] [PubMed] [Google Scholar]
- 23.Wang H, Cui H, Peng X, Fang J, Zuo Z, Liu J, Wu B, Deng Y, Wang K. Effects of high dietary fluorine on fatty acid composition in breast muscle of broilers. Fluoride. 2012;45:100–107. [Google Scholar]
- 24.Deng Y, Cui H, Peng X, Fang J, Zuo Z, Deng J, Luo Q. High dietary fluorine alteration of serum cytokine and immunoglobulin in broilers. Fluoride. 2013;46:118–127. [Google Scholar]
- 25.Chen T, Cui Y, Bai C, Gong T, Peng X, Cui H. Increased apoptotic lymphocyte population in the spleen of young chickens fed diets high in fluorine. Fluoride. 2009;42:94–100. [Google Scholar]
- 26.Chen T, Cui H, Cui Y, Bai C, Gong T, Peng X. Inhibition of splenocyte proliferation and spleen growth in young chickens fed high fluoride diets. Fluoride. 2009;42:203–209. [Google Scholar]
- 27.Chen T, Cui Y, Gong T, Bai C, Peng X, Cui HM. Inhibition of splenocyte proliferation and spleen growth in young chickens fed high fluoride diets. Fluoride. 2009;42:203–209. [Google Scholar]
- 28.Chen T, Cui H, Cui Y, Bai C, Gong T, Peng X. Cell-cycle blockage associated with increased apoptotic cells in the thymus of chickens fed on diets high in fluorine. Hum Exp Toxicol. 2011;30:685–692. doi: 10.1177/0960327110379022. [DOI] [PubMed] [Google Scholar]
- 29.Liu J, Cui H, Peng X, Fang J, Zuo Z, Wang H, Wu B, Deng Y, Wang K. Changes induced by high dietary fluorine in the cecal tonsil cytokine content of broilers. Fluoride. 2012;45:102–106. [Google Scholar]
- 30.Liu J, Cui H, Peng X, Fang J, Zuo Z, Wang H, Wu B, Deng Y, Wang K. Decreased percentages of T-cell subsets and IL-2 contents in the cecal tonsil of broilers fed diets high in fluorine. Fluoride. 2012;45:53–57. [Google Scholar]
- 31.Luo Q, Cui H, Peng X, Fang J, Zuo Z, Liu J, Wu B, Deng Y. The association between cytokines and intestinal mucosal immunity among broilers fed on diets supplemented with fluorine. Biol Trace Elem Res. 2013;152:212–218. doi: 10.1007/s12011-013-9612-0. [DOI] [PubMed] [Google Scholar]
- 32.Chen T, Gong T, Bai C, Peng X, Cui H. Effect of Dietary High Fluorine on the Morphologic Structure, Cell Cycle and Apoptosis of bursa of Fabricius in Broilers. Acta Vet Zootech Sinica. 2009;40:1235–1243. [Google Scholar]
- 33.Gong T, Chen T, Bai C, Peng X, Cui H. Effect of Dietary High Fluorine on the Cell Cycle and Apoptosis of Liver in Chickens. Acta Vet Zootech Sinica. 2009;40:1675–1680. [Google Scholar]
- 34.Gong T, Chen T, Bai C, Peng X, Cui H. Effect of High Fluorine on the Antioxidant Function and Ultrastructure of Liver in Chickens. Acta Vet Zootech Sinica. 2009;40:1389–1394. [Google Scholar]
- 35.Bai C, Peng X, Chen T, Gong T, Cui H. Effect of high fluorine on the antioxygen function of kidney in chickens. Chin J Vet Sci. 2010;30:825–828. [Google Scholar]
- 36.Bai C, Chen T, Gong T, Peng X, Cui H. Pathological effect of high fluorine on kidney and the related biochemical para. Chin J Vet Sci. 2010;30:505–509. [Google Scholar]
- 37.Zhou B, Wang H, Wang J, Zhang J, Yan X, Wang J. Effects of malnutrition and supplemented nutrition on nonspecific immune function changes induced by fluoride in rabbits. Fluoride. 2007;40:169–177. [Google Scholar]
- 38.Machalinska A, Wiszniewska B, Tarasiuk J, Machalinski B. Morphological effects of sodium fluoride on hematopoietic organs in mice. Fluoride. 2002;35:231–238. [Google Scholar]
- 39.Zhang Y, Sun G, Jin Y, Wang Y. Effects of fluoride on cell cycle and apoptosis in cultured osteoblasts of rats. Jhygiene Res. 2003;32:432–433. [PubMed] [Google Scholar]
- 40.Refsnes M, Becher R, Låg M, Skuland T, Schwarze P. Fluoride-induced interleukin-6 and interleukin-8 synthesis in human epithelial lung cells. HumExp Toxicol. 1999;18:645–652. doi: 10.1191/096032799678839572. [DOI] [PubMed] [Google Scholar]
- 41.Franco R, Cidlowski JA. Apoptosis and glutathione: beyond an antioxidant. Cell Death & Differentiation. 2009;16:1303–1314. doi: 10.1038/cdd.2009.107. [DOI] [PubMed] [Google Scholar]
- 42.Fan TJ, Han LH, Cong RS, Liang J. Caspase Family Proteases and Apoptosis. Acta Biochimica Et Biophysica Sinica. 2005;37:719–727. doi: 10.1111/j.1745-7270.2005.00108.x. [DOI] [PubMed] [Google Scholar]
- 43.Barbier O, Arreola-Mendoza L, Del Razo LM. Molecular mechanisms of fluoride toxicity. ChemBiolInt. 2010;188:319–333. doi: 10.1016/j.cbi.2010.07.011. [DOI] [PubMed] [Google Scholar]
- 44.Curnutte JT, Babior BM, Karnovsky ML. Fluoride-mediated activation of the respiratory burst in human neutrophils. A reversible process. JClin Invest. 1979;63:637–647. doi: 10.1172/JCI109346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Aardema MJ, Tsutsui T. Sodium fluoride-induced chromosome aberrations in different cell cycle stages. Mut Resfundamental Mol Mechanisms Mut. 1995;331:171–172. doi: 10.1016/0027-5107(95)00053-l. [DOI] [PubMed] [Google Scholar]
- 46.Hirano S, Ando M. Apoptotic cell death following exposure to fluoride in rat alveolar macrophages. ArchToxicol. 1996;70:249–251. doi: 10.1007/s002040050268. [DOI] [PubMed] [Google Scholar]
- 47.Karube H, Nishitai G, Inageda K, Kurosu H, Matsuoka M. NaF activates MAPKs and induces apoptosis in odontoblast-like cells. J Dent Res. 2009;88:461–465. doi: 10.1177/0022034509334771. [DOI] [PubMed] [Google Scholar]
- 48.Ming Z, Aiguo W, Tao X, Ping H. Effects of fluoride on DNA damage, S-phase cell-cycle arrest and the expression of NF-kappaB in primary cultured rat hippocampal neurons. ToxicolLett. 2008;179:1–5. doi: 10.1016/j.toxlet.2008.03.002. [DOI] [PubMed] [Google Scholar]
- 49.Jothiramajayam M, Sinha S, Ghosh M, Nag A, Jana A, Mukherjee A. Sodium fluoride promotes apoptosis by generation of reactive oxygen species in human lymphocytes. Journal of Toxicology & Environmental Health Part A. 2014;77:1269–1280. doi: 10.1080/15287394.2014.928658. [DOI] [PubMed] [Google Scholar]
- 50.Fan T, Xia L, Han YR. Mitochondrion and Apoptosis. Acta Bioch Bioph Sin. 2001;33:7–12. [PubMed] [Google Scholar]
- 51.Starkov AA, Polster BM, Gary F. Regulation of hydrogen peroxide production by brain mitochondria by calcium and Bax. J Neurochem. 2002;83:220–228. doi: 10.1046/j.1471-4159.2002.01153.x. [DOI] [PubMed] [Google Scholar]
- 52.Javorina M, Sandra H, Claudia H, Martin S. The DNA damage-induced decrease of Bcl-2 is secondary to the activation of apoptotic effector caspases. Oncogene. 2003;22:6852–6856. doi: 10.1038/sj.onc.1206716. [DOI] [PubMed] [Google Scholar]
- 53.Lü CX, Fan TJ, Hu GB, Cong RS. Apoptosis-inducing factor and apoptosis. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2003;35:881–885. [PubMed] [Google Scholar]
- 54.Fu YF, Fan TJ. Bcl-2 Family proteins and apoptosis. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2002;34:389–394. [PubMed] [Google Scholar]
- 55.Yang T, Zhang Y, Li Y, Hao Y, Zhou M, Dong N, Duan X. High amounts of fluoride induce apoptosis/cell death in matured ameloblast-like LS8 cells by downregulating Bcl-2. ArchOral biol. 2013;58:1165–1173. doi: 10.1016/j.archoralbio.2013.03.016. [DOI] [PubMed] [Google Scholar]
- 56.Xu H, Jin XQ, Jing L, Li GS. Effect of sodium fluoride on the expression of bcl-2 family and osteopontin in rat renal tubular cells. BiolTrace ElemRes. 2006;109:55–60. doi: 10.1385/BTER:109:1:055. [DOI] [PubMed] [Google Scholar]
- 57.Yang S, Wang Z, Farquharson C, Alkasir R, Zahra M, Ren G, Han B. Sodium fluoride induces apoptosis and alters bcl-2 family protein expression in MC3T3-E1 osteoblastic cells. Biochem Biophys Res Commun. 2011;410:910–915. doi: 10.1016/j.bbrc.2011.06.094. [DOI] [PubMed] [Google Scholar]
- 58.Cecconi F, Alvarez-Bolado G, Meyer BI, Roth KA, Gruss P. Apaf1 (CED-4 homolog) regulates programmed cell death in mammalian development. Cell. 1998;94:727–737. doi: 10.1016/s0092-8674(00)81732-8. [DOI] [PubMed] [Google Scholar]
- 59.Budihardjo I, Oliver H, Lutter M, Luo X, Wang X. Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell DevBi. 1999;15:269–290. doi: 10.1146/annurev.cellbio.15.1.269. [DOI] [PubMed] [Google Scholar]
- 60.Nan Z, Yali C, Ruixuan J, Erwei L, Xiuling C, Zhijun X, Yinglu G, Xingzhong L, Yuguang Z, Yongsheng C. PARP and RIP 1 are required for autophagy induced by 11′-deoxyverticillin A, which precedes caspase-dependent apoptosis. Autophagy. 2011;7:598–612. doi: 10.4161/auto.7.6.15103. [DOI] [PubMed] [Google Scholar]
- 61.Kumar S, Dorstyn L. Analysing caspase activation and caspase activity in apoptotic cells. Methods in Molecular Biology. 2009;559:3–17. doi: 10.1007/978-1-60327-017-5_1. [DOI] [PubMed] [Google Scholar]
- 62.Scott FL, Stec B, Pop C, Lee JEJ, Monosov E, Robinson H, Salvesen GS, Schwarzenbacher R, Riedl SJ. The Fas/FADD death domain complex structure unravels signaling by receptor clustering. Nature. 2009;457:1019–1022. doi: 10.1038/nature07606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lavrik IN, Krammer PH. Regulation of CD95/Fas signaling at the DISC. Cell Death & Differentiation. 2011;19:36–41. doi: 10.1038/cdd.2011.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Jianying H, Hengmin C, Xi P, Jing F, Zhicai Z, Junliang D, Bangyuan W. The association between splenocyte apoptosis and alterations of Bax, Bcl-2 and caspase-3 mRNA expression, and oxidative stress induced by dietary nickel chloride in broilers. Int J Environ Res Public Health. 2013;10:7310–7326. doi: 10.3390/ijerph10127310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Mebius RE, Georg K. Structure and function of the spleen. Nature Reviews Immunology. 2005;5:606–616. doi: 10.1038/nri1669. [DOI] [PubMed] [Google Scholar]
- 66.Cesta MF. Normal structure, function, and histology of the spleen. Toxicologic Pathology. 2006;34:455–465. doi: 10.1080/01926230600867743. [DOI] [PubMed] [Google Scholar]
- 67.Tominaga H, Ishiyama M, Ohseto F, Sasamoto K, Hamamoto T, Suzuki K, Watanabe M. A water-soluble tetrazolium salt useful for colorimetric cell viability assay. Analytical Communications. 1999;36:47–50. [Google Scholar]
- 68.Miyamoto T, Min W, Lillehoj HS. Lymphocyte proliferation response during Eimeria tenella infection assessed by a new, reliable, nonradioactive colorimetric assay. Avian Diseases. 2002;46:10–16. doi: 10.1637/0005-2086(2002)046[0010:LPRDET]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 69.Peng X, Yu Z, Liang N, Chi X, Li X, Jiang M, Fang J, Cui H, Lai W, Zhou Y. The mitochondrial and death receptor pathways involved in the thymocytes apoptosis induced by aflatoxin B1. Oncotarget. 2016;7:12222–12234. doi: 10.18632/oncotarget.7731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhang HB, Lu P, Guo QY, Zhang ZH, Meng XY. Baicalein induces apoptosis in esophageal squamous cell carcinoma cells through modulation of the PI3K/Akt pathway. Oncol Lett. 2013;5:722–728. doi: 10.3892/ol.2012.1069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Jia Jun Liu YZ, Guang Wei Bin, Yang Hong Zhi, Lin Dong Jun, Xiao Ruo Zhi. Ponicidin Inhibits Monocytic Leukemia Cell Growth by Induction of Apoptosis. Int J MolSci. 2008;9:2265–2277. doi: 10.3390/ijms9112265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Wen B., Li H., Hong S., D Y, Ou Y., Huan J., Jing F., Xian H. The immunosuppressive effect of gossypol in mice is mediated by inhibition of lymphocyte proliferation and by induction of cell apoptosis. Acta Pharmacol Sin. 2009;30:597–604. doi: 10.1038/aps.2009.35. [DOI] [PMC free article] [PubMed] [Google Scholar]