

Abstract
Background
With the increasing capacity of present-day next-generation sequencers the field of mitogenomics is rapidly changing. Enrichment of the mitochondrial fraction, is no longer necessary for obtaining mitogenomic data. Despite the benefits, shotgun sequencing approaches also have disadvantages. They do not guarantee obtaining the complete mitogenome, generally require larger amounts of input DNA and coverage is low compared to sequencing with enrichment strategies. If the mitogenome could be amplified in a single amplification, additional time and costs for sample preparation might outweigh these disadvantages.
Results
A sequence of the complete mitochondrial genome of the pupilloid landsnail Orcula dolium is presented. The mitogenome was amplified in a single long-range (LR) PCR and sequenced on an Ion Torrent PGM (Life Technologies). The length is 14,063 nt and the average depth of coverage is 1112 X. This is the first published mitogenome for a member of the family Orculidae. It has the typical metazoan makeup of 13 protein coding genes (PCGs), 2 ribosomal RNAs (12S and 16S) and 22 transfer RNAs (tRNAs). Orcula is positioned between Pupilla and the Vertiginidae as the sister-group of Gastrocopta and Vertigo, together. An ancestral gene order reconstruction shows that Orthurethra in contrast to other Stylommatophora, have tRNA-H before tRNA-G and that the gene order in the ‘non-achatinoid’ clade is identical to that of closely related non-stylommatophoran taxa.
Conclusions
We show it is feasible to ultra-deep sequence a mitogenome from a single LR-PCR. This approach is particularly relevant to studies that have low concentrations of input DNA. It results in a more efficient use of NGS capacity (only the targeted fraction is sequenced) and is an effective selection against nuclear mitochondrial inserts (NUMTS). In contrast to previous studies based in particular on 28S, our results indicate that phylogeny reconstructions based on complete mitogenomes might be more suitable to resolve deep relationships within Stylommatophora. Ancestral gene order reconstructions reveal rearrangements that characterize systematic groups.
Electronic supplementary material
The online version of this article (doi:10.1186/s41065-017-0028-2) contains supplementary material, which is available to authorized users.
Keywords: Stylommatophora, Orthurethra, Mitochondrial genome, Gene arrangement
Background
A recent increase in the number of sequenced mitogenomes allows for a better understanding of gastropod evolution [1–3]. Of the more than sixty mitogenomes that are currently available for Gastropoda, less than twenty belong to the Eupulmonata sensu [4], of which the clade Stylommatophora represents the majority of the terrestrial snails. Although the first stylommatophoran mitogenome was sequenced nearly two decades ago [5], it took more than 15 years before new mitogenomes were added to this group on about a yearly basis, as is the case at present. Complete mitogenomes have been obtained for 18 species of Stylommatophora now (excluding Euhadra and Orcula; accessed 2016-10-17) (Table 1). These include representatives of the superfamilies Achatinelloidea, Clausilloidea, Helicoidea, Orthalicoidea, Pupilloidea, Succinoidea and Urocoptoidea, or more inclusively the subclades Elasmognatha, Orthurethra, the ‘Limacoid clade’, the informal group Sigmurethra sensu [6] and the ‘achatinoid clade’ sensu [7]. Here we report the mitochondrial genome of a fifth orthurethran species, the first one for the family Orculidae, viz. Orcula dolium (Drapernaud, 1801). It is the type species of the genus Orcula Held 1837, which comprises 13 species featuring ovate–cylindrical shells of 5 to 10 mm height. Of these, O. dolium shows the widest distribution and is ecologically most tolerant. Orcula is common in limestone areas of the Central European Alps and the Western Carpathians and is usually associated with mountainous forest habitats and rocky landscapes. Its altitudinal distribution covers a range from 200 m to 2160 m above sea level [8]. Loess sediments of the Pannonian Basin (Hungary, Republic of Croatia and Republic of Serbia) [9–11] and the periphery of the Western and Eastern Alps [12, 13] show that O. dolium was also widely distributed throughout glacial periods of the Late Pleistocene.
Table 1.
List of included mitogenomes (accessed 2016-10-17)
| Species | Length (nt) | GenBank | Reference |
|---|---|---|---|
| Stylommatophora | |||
| Orthurethra | |||
| Achatinella mustelina | 16,323 | KU525108 | [41] |
| Gastrocopta cristata | 14,060 | KC185403 | [42] |
| Orcula dolium | 14,063 | KJ867421 | This study |
| Pupilla muscorum | 14,149 | KC185404 | [42] |
| Vertigo pusilla | 14,078 | KC185405 | [42] |
| Sigmurethra | |||
| Achatina fulica | 15,057 | KJ744205 | [59] |
| Aegista aubryana | 14,238 | NC_029419 | [60] |
| Aegista diversifamilia | 14,039 | KR002567 | [61] |
| Albinaria caerulea (Deshayes, 1835) | 14,130 | NC_001761 | [5] |
| Camaena cicatricosa | 13,843 | KM365408 | [48] |
| Cernuella virgata | 14,147 | KR736333 | [62] |
| Cerion incanum | 15,177 | NC_025645 | [57] |
| Cepaea nemoralis Linnaeus, 1758 | 14,100 | NC_001816 | [63] |
| Cornu aspersum (Müller, 1774) | 14,050 | NC_021747 | [35] |
| Cylindrus obtusus (Draparnaud, 1805) | 14,610 | NC_017872 | [47] |
| Dolicheulota formosensis | 14,237 | KR338956 | [61] |
| Euhadra herklotsi | ? | Z71693- Z71701 | [64] |
| Mastigeulota kiangsinensis | 14,029 | KM083123 | [65] |
| Naesiotus nux | 15,197 | KT821554 | [24] |
| Elasmognatha | |||
| Succinea putris Linnaeus, 1758 | 14,092 | NC_016190 | [3] |
| Basommatophora | |||
| Hygrophila | |||
| Biomphalaria glabrata | 13,670 | NC_005439 | [66] |
| Biomphalaria tenagophila | 13,722 | NC_010220 | [67] |
| Galba pervia | 13,768 | NC_018536 | [68] |
| Physella acuta | 14,490 | JQ390525 | [69] |
| Archaepulmonata | |||
| Ellobioidea | |||
| Myosotella myosotis | 14,246 | NC_012434 | [70] |
| Pedipes pedipes | 16,708 | NC_016179 | [3] |
| Systellomatophora | |||
| Rhopalocaulis grandidieri | 14,523 | NC_016183 | [3] |
Methods
This study was carried out on a specimen of Orcula dolium dolium (RMNH 114169) collected in 2009 in SW Berchtesgaden (Bayern, Germany). DNA was extracted with a Qiagen DNA tissue kit. Total yield of DNA extracted was ~13 ug (DNA conc. 66.7 ng/ul with an elution volume of 200 ul). Using universal barcoding primers [14] a partial sequence (655 nt) of Cytochrome Oxidase subunit I (COI) was obtained using the procedure described in [15]. This sequence was used to design specimen specific primers (Orcula_529_COI_F 5′-CTAAGACTATTTGTGTGGTCGATCTTA-3′ and Orcula_336_COI_R 5′-TCTAGACCTAATCAAAAGAACAAATGAAG-3′) to amlify the complete mitogenome of O. dolium. An amplicon with a length of 13,871 nt was obtained with GoTaq long PCR Master Mix (Promega) using the manufacturers protocol. Thermocycling profile was 2 m. at 94 °C, followed by 40 cycles of 30 s. at 94 °C, 15 m. at 65 °C, 10 m. at 72 °C. The PCR product was gel purified with the Wizard SV gel and PCR cleanup-system (Promega) and subsequently checked on a BioAnalyzer 2100 using a DNA 12000 chip (Agilent). The Orcula library was part of a combined run in which different samples were pooled. The purified amplicon was enzymatically digested and individual samples were ligated with a unique Ion Express Barcode Adapter (Life Technologies) using the NEBNext Fast DNA & Library Prep Set for Ion Torrent (New England Biolabs), following the manufacturers instructions. After ligation samples were quantified on the Bioanalyser 2100 using a DNA High sensitivity chip (Agilent). An equimolair pool was prepared of the highest possible concentration. This equimolair pool was diluted according to the calculated template dilution factor to target 10–30% of all positive Ion Sphere Particles. Template preparation and enrichment was carried out with the Ion Touch 2 system, using the OT2 400 kit (Life Technologies), according to the manufactures protocol (7218RevA0). The enriched Ion Sphere Particles were prepared for sequencing on a Personal Genome Machine (PGM) with the Ion PGM 400 Sequencing kit as described in the protocol using a 316v2 chip.
Quality check and assembly
Reads from the Orcula library were separated from the pool based on their unique barcode tag by the Ion Torrent Server. Quality was checked using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Reads shorter than 35 nt or with a phred score below 28 were removed with Fastx_trimmer (http://hannonlab.cshl.edu/fastx_toolkit/). A ‘de novo’ assembly was carried out with Spades v.2.5.1. [16] and reads were mapped against available orthurethran sequences (KC185403-05, KU525108) using Geneious v.7.1.7 [17]. Finally ‘de novo’ contigs larger than 1 kb (for gene order assessment) and mapping assemblies were merged (see Additional file 1: Figure S1) and used as reference for iterative mapping (medium sensitivity, no fine tuning, not trimmed before mapping; other parameters left at default) of the quality checked reads (again using Geneious v.7.1.7). The reason for this two-step approach was that the initial ‘de novo’ assembly did not result in a contig of expected length. To confirm the final assembly (and enclosed gene order) a selection of matching reads (~ten fold downsampling) was analysed with another assembler, MITObim v. 1.8 [18] (using the 655 nt COI sequence as seed bait) and with Spades 2.5.1. [17] again also.
Annotation
The position of the protein coding genes (PCGs) was determined by alignment with available stylommatophoran mitogenomes (Table 1) and by locating start and stop codons. The identification of the ribosomal RNAs was done with BLAST searches. Arwen and Mitos [19] were used to locate the tRNAs and the secondary structures were all generated with Arwen [20]. The contig sequence was annotated in Geneious [17].
Phylogenetic analyses
A prerequisite for understanding (mitochondrial) gene rearrangements is a robust, well rooted phylogeny. Representatives of the clades Hygrophila and Eupulmonata were selected as outgroup because these taxa are supposed to be closely related to, but not part of the ingroup [3, 4, 21]. For each of the 13 PCGs alignments were made with TranslatorX [22]; a program that aligns nucleotide sequences based on their corresponding amino acid translations. Translator X was run with the MAFFT alignment module [23] and the invertebrate mitochondrial genetic code; other settings were left at default. A supposed ‘copy’ of ND4L [24] for Naesiotus nux (GenBank accession number NC_028553) was excluded because there are no stylommatophoran homologs known to align it with. Ribosomal genes were individually aligned in Geneious, again using MAFFT [23] and conserved datablocks were selected with Gblocks [25] using default settings. The PCG and ribosomal alignments were concatenated in Geneious and exported as a phylip formatted matrix (27 taxa, 13,419 nucleotides). With 41 datablocks specified (each codon position and the ribosomal RNAs), PartitionFinder v.1.1.1 [26] estimated the best partitioning scheme (33 partitions) and nucleotide substitution models. A Bayesian analysis (two simultaneous MCMC runs 10 M generations each) was conducted with MrBayes 3.2.3 hosted on the CIPRES Science Gateway [27]. Inspection of the parameter files with Tracer v.1.5 [28] showed proper mixing of the MCMC (effective sampling size values > 200). The first 2500 trees (25% of each tree-file) were discarded as burnin. Majority rule consensus trees were visualised and edited in FigTree v.1.4.0 [29]. The procedure was repeated with the amino acid alignments (13 datablocks, 4190 amino acids) from TranslatorX [22]. The estimated partitioning scheme now consisted of six partitions and the analysis was carried out with 2.5 M generations to keep computation time tenable. Nucleotide and amino acid datamatrices were analysed separately to see if both would yield phylogenies with similar topologies. Abbreviations: BPP = Bayesian Posterior Probability.
Mitochondrial gene arrangements
An ancestral gene order reconstruction [30] was done using the Maximum Likelihood Gene Order analysis (MLGO) web server (http://www.geneorder.org/server.php) using the phylogeny obtained in the previous step (outgroup reduced to Biomphalaria) as a fixed tree (ie. Small Parsimony Problem, using SPP option).
Taxonomic implications
For the classification and the nomenclature of the various taxa, the review by Bouchet and Rocroi [6] served as the primary starting point. Historic classifications such as the Pilsbry-Baker system [31, 32] are not exhaustively discussed. The more recent literature is dealt with only when it contains additional data. Ever since the rise of phylogenetic systematics, the increase of more detailed anatomical analyses and in particular the quickly growing quantity of molecular data, the classification of the gastropods has been in a confusing transitional state. In the monograph of Zilch [33] for example, published before the rise of phylogenetic systematics had started, the classis Gastropoda is subdivided into the subclasses Prosobranchia and Euthyneura. The latter subclassis contains the ordines Basommatophora and Stylommatophora, whereas the ordo Stylommatophora is further split into the subordines Orthurethra, Heterurethra and Sigmurethra. Many of these names are still in use, but often not for exactly the same group of species. Next to the classical taxonomical categories, new nominal taxa names were more recently introduced, like ‘clade’ and ‘informal group’. Bouchet et al. [6] for example, in a recent nomenclatorial monograph on gastropod classification and nomenclature, use ‘Informal Group Pulmonata’ as a partial synonym of Zilch’s Euthyneura. The Orthurethra are accepted by [6] as a ‘Subclade’, which is identical with Zilch’s subordo of that name. There are many more discrepancies, however. For a more recent contribution to this subject, see [4]. Here we do not aim at a summary of the changing and sometimes conflicting views regarding gastropod phylogeny.
Results
The Ion-Torrent run resulted in 118,854 reads of which 115,412 were retained after trimming with Fastx_trimmer. The retained reads were in the length range 35–313 nt and had an average length of 143.8 nt (sd. 42.8). Of these 107,569 could be mapped against the merged reference sequence, resulting in an average coverage of 1112X (min. 208X, max. 2483X; see Fig. 1). The mitogenome of O. dolium has a length of 14,063 nucleotides, which is within the known range for Stylommatophora (13.843 and 16.323 nt for Camaena and Achatinella, respectively) and very similar to that reported for other Pupilloidea (14,060 and 14,078 nt for Gastrocopta and Vertigo respectively). Both MITObim v.1.8 [18] and Spades v.2.5.1 [17] yielded the same length and nucleotide sequence using the downsampled dataset. The AT content is 66% and skews for AT and GC are −0.084 and 0.043. All 37 common metazoan genes (13 protein coding genes, two rRNAs and 22 tRNAs) were recovered. The gene order of PCGs and ribosomal RNAs was identical to those of most Stylommatophora (Achatinelloidea and certain Helicoidea excepted). The distribution of tRNAs was, with one mutational step, most similar to that of Vertigo and Albinaria. All tRNAs showed standard cloverleaf secondary structures except for tRNA-K and tRNA-W which had missing T-arms and tRNA-S1 which had a D-arm missing (see Additional file 2: Figure S2). The annotated mitogenome of O. dolium has been deposited in GenBank (accession number KJ867421).
Fig. 1.
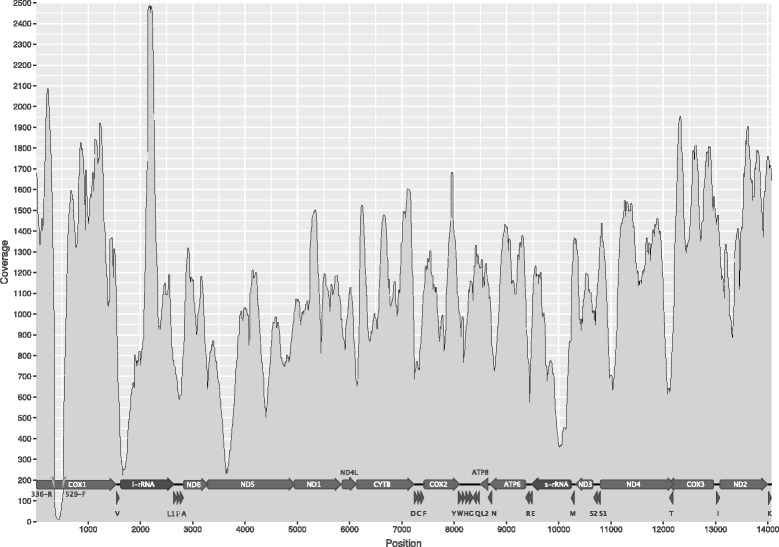
Coverage plot of the Orcula dolium mitochondrial genome (14,063 nt) and corresponding gene annotation
Discussion
Selection of outgroup
Recent studies have shown that the traditionally accepted nominal taxa Pulmonata and Opisthobranchia are not monophyletic, what necessitated a new classification of the Euthyneura [4]. Within that taxon these authors accept Panpulmonata and Eupulmonata, with the Stylommatophora belonging to the Eupulmonata. The Informal Group Basommatophora is assigned to a paraphyletic group Panpulmonata. Next to the selection of markers and taxa, outgroup selection strongly affects our concept of Euthyneuran phylogeny, as is evident from the literature [1, 3, 4, 21, 34–36]. Despite the differences, all these studies show that the clades Hygrophila (Lymnaeoidea & Planorboidea), Systellommatophora (Veronicelloidea) and Eupulmonata (Ellobioidea) are closely related to, but separate from the Stylommatophora (i.e. these are eligible outgroup taxa).
Gene order
Changes in mitochondrial gene order are common in gastropods [1, 3, 21]. Among the Stylommatophora however, the order is rather conserved (Fig. 2). Here most shifts occur in the arrangement of tRNAs. In the Stylommatophora PCG rearrangements have only been recorded in Helicidae (COIII), Aegista (Bradybaenidae; ND3) and Achatinella (Achatinellidae; COII). Most transpositions have been observed in the Helicoidea, but there is a strong bias in available data for that group.
Fig. 2.

Mitogenomic gene arrangement of Stylommatophora and Euthyneuran outgroup taxa
Polarisation
The monophyly of the Stylommatophora has been shown repeatedly [4, 7, 34, 37] (Fig. 2), but a subdivision into lower taxa remained problematic. Inclusion of additional genes or investigation of rare genomic changes (RGCs), such as changes in mitochondrial gene order [38] has been suggested [39] to solve this problem. Mitochondrial gene order data (Fig. 2) together with the phylogeny (Fig. 3) allowed for a reconstruction of the ancestral gene order (Fig. 4). Rearrangements that characterize established systematic groups are discussed below.
Fig. 3.
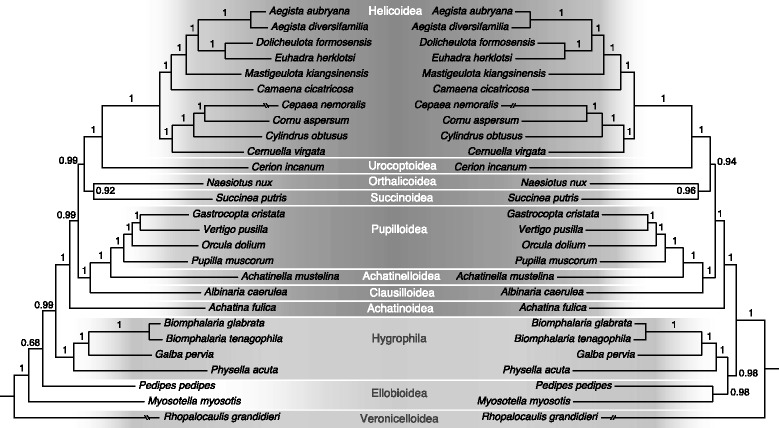
Bayesian mitogenomic phylogeny reconstructions of Stylommatophora based on nucleotide (left) and amino acid data (right)
Fig. 4.
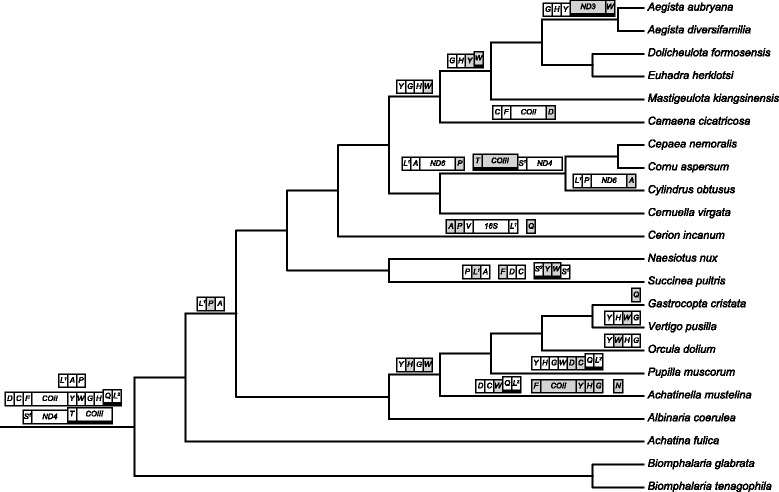
Ancestral (mt) gene order reconstruction for Stylommatophora
Comparison with existing classifications
The number of Stylommatophora with known mitogenomes is still limited, but representatives of most of the major groups are available now. This allows for a preliminary comparison.
Orthurethra [31]
Initially, Orthurethra was considered a ‘primitive’ group of Stylommatophora [31, 40]. Molecular analyses showed however, that the Orthurethra are a more derived taxon [7, 37]. Bouchet and Rocroi [6] accept five orthurethran superfamilies, one of which is the Pupilloidea. Complete mitogenomic sequences are available only for Achatinelloidea [41] and for Pupilloidea, i.e. for Pupillidae and Vertiginidae [42] and now additionally for Orculidae.
The Orthurethra and the Pupilloidea are both monophyletic. The Vertiginidae, represented by Gastrocopta and Vertigo are the sister-group of the Orculidae (Fig. 2). Recently, Gastrocopta (Gastrocoptinae) was excluded from the ‘vertiginid’-clade (Vertiginidae), mainly based on 28S sequence data; that phylogeny [43] showed two clades: one consisting of Pupilla and Vertigo and another with Orcula and Gastrocopta, albeit poorly supported. These sister-group relationships, as well as the exclusion of Gastrocoptinae from Vertiginidae are rejected by our data (Fig. 2).
The Clausilioidea, classified in an Informal Group Sigmurethra [6] and the Orthurethra are sister-groups (BPP = 1.0; Fig. 2).
In the Orthurethra (and Stylommatophora in general) most mitochondrial gene rearrangements occurred in the region between CytB and ATP8. Except for that region, the gene order of Pupilloidea is identical to that in Albinaria (Fig. 2). In the Achatinelloidea a number of rearrangements was found that are not observed in Pupilloidea (most prominent is the transposition of the region tRNA-F_COII_tRNA-YHG; Fig. 2). Both orthurethran superfamilies have the gene order tRNA-HG (or at least tRNA-H before tRNA-G) in which they deviate from other Stylommatophora (which have tRNA-GH). We hypothesize that the arrangement tRNA-HG is an apomorphic character state for the Orthurethra.
The MLGO result predicted tRNA-YHWG as the ancestral gene arrangement for Orthurethra (requiring additional steps for all orthurethrans except Vertigo). Alternatively tRNA-YHGW would have been equally parsimonious (also nine steps) and is more likely, given that tRNA-YHG is observed in Achatinella, Pupilla and Gastrocopta. Therefore the latter scenario was adopted in Fig. 4. In either reconstruction the shift from tRNA-GH to tRNA-H_before_G must have taken place early in orthurethran history. Additionaly, for the Pupilloidea, tRNA-W was transposed independently in Orcula and Vertigo, tRNA-DC in Pupilla, and tRNA-Q from L-strand to H-strand in Gastrocopta (Figs. 2 and 4).
The mitogenome of Achatinella mustelina is said to be “similar to those of other pulmonates” [41]. We assume that statement refers to gene composition, not gene order, because one of the largest transpositions recorded for Stylommatophora is seen in Achatinella. The latter taxon has transposed tRNA-F_COII_tRNA-YHG (assuming it was not the larger fragement tRNA-WQL2_ATP8-ATP6_tRNA-RE_12S), tRNA-W and tRNA-N (Figs. 2 and 4).
Heterurethra [31]
The Heterurethra sensu Pilsbry [31] or the subclade Elasmognatha according to [6] when Athoracophoroidea are included next to Succineoidea, were classified in a clade with Acavoidea (Leucotaenius) and Orthalicoidea (Placostylus) [7, 37], albeit poorly supported. Our analyses support (BPP = 0.92/0.97; Fig. 3) a sister-group relation between Elasmognatha (Succinea: Succineoidea) and Orthalicoidea (Naesiotus). In agreement with previous studies [7, 37] our data indicate that Succineoidea and Orthalicoidea together are the sister-group of the combined Urocoptoidea and Helicoidea (Fig. 3). Of the three gene rearrangements within Succinea (Figs. 2 and 4) none is shared with Naesiotus.
A peculiar feature of Naesiotus nux (genbank access.nr. KT821554) is a ‘duplication’ of ND4L (positioned between tRNA-L and tRNA-P; [24]). Since this ‘copy’ (sequence divergence > 60%) was not present in other Stylommatophora it could not be included in our analyses. It might partly explain the increased size of the mitogenome of N. nux compared to that of other Stylommatophora (Table 1).
Mesurethra [32]
Of this nominal taxon the Cerionidae and Clausiliidae, which were included by [44], are represented. Earlier studies showed the polyphyly of this group [7, 37]. Our phylogeny reconstructions (Fig. 3) also reject the hypothesis of a close relation between Cerionidae and Clausiliidae.
Recently, Urocoptidae and Cerionidae were shown to constitute a monophyletic group (based on 28S sequence data) for which the superfamily Urocoptoidea was introduced [45]. Urocoptoidea and Helicoidea (= Sigmurethra) are sister-groups (BPP = 1.0; Fig. 3). Vaught [46] classified only the Clausiloidea as Mesurethra.
Sigmurethra [31]
The Sigmurethra sensu Pilsbry [31] was already shown to be paraphyletic [7, 37]. Our data reconfirm that Sigmurethra is not monophyletic (Fig. 3). Bouchet and Rocroi [6] refer to the this nominal taxon as an ‘Informal Group’, which may remain in use as long as no preferential alternative has been advocated.
‘Achatinoid clade’ [7, 37]
In our analyses, Achatina fulica is the sister-group of the ‘non-Achatinoid’ clade. Stylommatophora (‘Achatinoid’ and ‘non-Achatinoid’ clades) is a monophyletic group (BPP = 1.0; Fig. 3). The mitogenomic gene arrangement of Achatina differs from that of the other Stylommatophora (Fig. 2), but it is identical to that of the outgroup genera Biomphalaria and Planorbarius, which are classified with the Planorboidea of the clade Hygrophila. Unlike the representatives of the ‘non-Achatinoid clade’, Achatina has the gene order tRNA-LAP (between 16S and ND6), which also occurs in more distantly related outgroups (Pedipes, Myosotella and Rhopalocaulis; Fig. 2). Therefore, using the principle of outgroup comparison, we hypothesize that the ancestral mitochondrial gene order of the Stylommatophora has been identical to the arrangement currently found in Achatina and in some taxa of the clade Hygrophila.
‘Non-Achatinoid clade’ [7, 37]
Our data show a strongly supported (BPP = 1.0; Fig. 3) basal split in the ‘non-Achatinoid clade’ with on the one hand Orthurethra (Achatinelloidea, Pupilloidea) + Clausilioidea and on the other hand Elasmognatha (= Heterurethra) + Orthalicoidea + Urocoptoidea + Helicoidea. In the latter clade Elasmognatha + Orthalicoidea is the sister-group of Urocoptoidea + Helicoidea.
Rearrangements of PCGs are thus far only observed in the Helicoidea and Achatinelloidea. Separation of ND6 and ND5 by a tRNA from the L-P-A region and transposition of tRNA-T + COIII might indeed be an apomorphy for the Helicidae, as suggested by [47]; the recently added mitogenome of Cornu aspersum [35] confirms this. We hypothesize that the gene arrangement tRNA-GH before tRNA-YW is an apomorphic character state characterizing Bradybaenidae. The transposition of ND3 is not observed in the other bradybaenids and at the moment being, is unique to Aegista. The mitogenomic gene arrangement of Camaenidae might be in between that of an helicoid ancestor and that of Bradybaenidae; tRNA-Y is still at the ancestral position, whereas tRNA-W is already transposed to the ‘bradybaenid’ location (albeit still on the heavy strand). Wang et al. concluded that the mitogenomic gene order of Camaena cicatricosa differs from that in other stylommatophores in the position of COII and the tRNA’s C, F, D, G, H and W [48]. The single transposition of tRNA-D is a more parsimonious explanation for the first four ‘differences’. The latter three could subsequently be explained by a single transposition of tRNA-W after tRNA-GH or the transposition of tRNA-GH (as a single unit) before tRNA-W. Movement of two consecutive tRNA’s as a single unit is not uncommon, as is exemplified by tRNA-YW in Succinea and tRNA-AP in Cerion (Fig. 2).
Camaenidae and Bradybaenidae have been considered confamilial by Scott [49], whereas both nominal taxa are paraphyletic according to others [7, 50]. According to our analyses, Camaenidae and Bradybaenidae form a clade within the Helicoidea (Fig. 3), leaving no place for a separate superfamily Camaenoidea, which was accepted by [51, 52].
The mitochondrial gene arrangements of Albinaria (Clausilioidea) and Naesiotus (Orthalicoidea) are identical, despite the fact that both taxa belong to different basal groups within the ‘non-Achatinoid’ clade. We hypothesize that this is the ancestral mitogenomic gene order of the ‘non-Achatinoid clade’ and that a rearrangement from tRNA-LAP to tRNA-LPA characterized the most recent common ancestor of this clade.
On the chosen sequence strategy
Amplification of the complete mitochondrial genome in a single LR-PCR with target specific primers for the snail species tested here was straightforward. Nevertheless, other studies show that full mitogenomic amplifications can be stochastic [53] and the method will not work with degraded material. Alternatively the mitochondrial fraction can be enriched by physical isolation (e.g. centrifugation in CsCl-gradient; requires significant amounts of starting material) or by target capture approaches, in which mitochondrial sequences are isolated by hybridisation to biotinylated probes [54]. Some studies abandon mitochondrial enrichment steps entirely and shotgun sequence (pools of) DNA extracts instead [41, 47, 55–58]. Each of these methods have their own strengths and weaknesses. In studies where multiple extracts are pooled and that do not make use of indexing tags, additional controls are necessary to check against ‘chimeric’ assemblies (especially when closely related taxa are involved). To this end (and for linking the obtained mitogenomes to species or vouchers) multiple ‘bait’ sequences have to be determined a priori [55, 56, 58]. A mitogenome assembled with pooled DNA from a large number of individuals of the same species [41], is inevitably artificial. Enrichment of the mitochondrial fraction by LR-PCR (and labeling using indexing tag sequences) is expensive and potentially time-consuming. With the capacity of current generation sequencers mitochondrial enrichment strategies might seem outdated. Advantages of the here demonstrated method are that it ensures obtainment of the complete mitochondrial locus from even minute amounts of starting material, there are no negative costs associated with increasing genome sizes (no mitochondrial to nuclear ratio effect) and it diminishes the risk of erroneoulsy sequencing NUMTs.
Conclusion
This study shows that a complete mitogenome can be amplified and sequenced with high coverage from a single LR-PCR. The approach might be especially relevant in situations where only small amounts of starting material are available. Phylogeny reconstructions based on entire mitogenomes are promising to resolve deep level relationships within Stylommatophora, that could not be resolved using only 28S sequence data. Well supported groups from previous studies based on ten-fold less sequence data [7, 37] were reconfirmed. The region between COII and ATP8 is apparently a hot spot for rearrangements in stylommatophoran mitogenomes. The ‘Achatinoid’ clade is a basal branch in Stylommatophora and has the same mitochondrial gene arrangement as closely related non-Stylommatophoran taxa (especially from the clade Hygrophila). Rearrangements in mitochondrial gene order can characterize different stylommatophoran ranks, e.g. (non-) ‘Achatinoid’ clade, Orthurethra, Helicidae and Bradybaenidae.
Additional files
Assembly strategy. (A) Regions where reads could not be mapped to the reference sequence (B) Regions that broke up the ‘de novo’ assembly (C) Mapping and de novo contigs merged. (PDF 15 kb)
Secondary structures of the inferred tRNAs of Orcula dolium. (PDF 107 kb)
Acknowledgements
We are grateful to M. Kolet for assisting with the fieldwork for this project.
Funding
The molecular work was funded in part by Fonds Economische Structuurversterking (FES).
Availability of data and materials
The annotated mitogenome of Orcula dolium is available from GenBank under accesssion number KJ867421.
Authors’ contributions
Experimental concept and design: DSJG. Labwork: ED, DSJG. Data analyses: DSJG. Manuscript writing: DSJG, EG, JH, ED. Figures: DSJG. All authors contributed to the analysis of data and writing of the manuscript and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
D. S. J. Groenenberg, Email: dick.groenenberg@naturalis.nl
J. Harl, Email: harl_josef@hotmail.com
E. Duijm, Email: elza.duijm@naturalis.nl
E. Gittenberger, Email: edi.gittenberger@naturalis.nl
References
- 1.Grande C, Templado J, Zardoya R. Evolution of gastropod mitochondrial genome arrangements. BMC Evol Biol. 2008;8:61. doi: 10.1186/1471-2148-8-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Medina M, Lal S, Valles Y, Takaoka TL, Dayrat BA, Boore JL, Gosliner T. Crawling through time: Transition of snails to slugs dating back to the Paleozoic, based on mitochondrial phylogenomics. Mar Genomics. 2011;4(1):51–9. doi: 10.1016/j.margen.2010.12.006. [DOI] [PubMed] [Google Scholar]
- 3.White TR, Conrad MM, Tseng R, Balayan S, Golding R, de Frias Martins AM, Dayrat BA. Ten new complete mitochondrial genomes of pulmonates (Mollusca: Gastropoda) and their impact on phylogenetic relationships. BMC Evol Biol. 2011;11(1):295. doi: 10.1186/1471-2148-11-295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Jörger KM, Stoger I, Kano Y, Fukuda H, Knebelsberger T, Schrodl M. On the origin of Acochlidia and other enigmatic euthyneuran gastropods, with implications for the systematics of Heterobranchia. BMC Evol Biol. 2010;10:323. doi: 10.1186/1471-2148-10-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hatzoglou E, Rodakis GC, Lecanidou R. Complete sequence and gene organization of the mitochondrial genome of the land snail Albinaria coerulea. Genetics. 1995;140(4):1353–66. doi: 10.1093/genetics/140.4.1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bouchet P, Rocroi J-P. Classification and nomenclator of gastropod families. Malacologia: International Journal of Malacology. Hackenheim: ConchBooks; 2005;47(1-2).
- 7.Wade CM, Mordan PB, Naggs F. Evolutionary relationships among the Pulmonate land snails and slugs (Pulmonata, Stylommatophora) Biol J Linn Soc. 2006;87(4):593–610. doi: 10.1111/j.1095-8312.2006.00596.x. [DOI] [Google Scholar]
- 8.Harl J, Duda M, Kruckenhauser L, Sattmann H, Haring E. In search of glacial refuges of the land snail Orcula dolium (Pulmonata, Orculidae)--an integrative approach using DNA sequence and fossil data. PLoS ONE. 2014;9(5):e96012. doi: 10.1371/journal.pone.0096012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fûköh L, Krolopp E, Sümegi P. Quaternary malacostratigraphy in Hungary. Malacological Newsletter. Gyöngyös: Mátra Múzeum; 1995;1:1–219.
- 10.Mitrović B. Pleistocene malacofauna of the Požarevac Danube area (NE Serbia) Geološki anali Balkanskoga poluostrva. 2007;68:81–9. doi: 10.2298/GABP0701081M. [DOI] [Google Scholar]
- 11.Molnár D, Hupuczi J, Galović L, Sümegi P. Preliminary malacological investigation on the loess profile at Zmajevac, Croatia. Cen Eur J Geogr. 2010;2(1):52–6. [Google Scholar]
- 12.Frank C. Plio-pleistozäne und holozäne Mollusken Österreichs. 2006. [Google Scholar]
- 13.Moine O, Rousseau D-D, Antoine P. Terrestrial molluscan records of Weichselian Lower to Middle Pleniglacial climatic changes from the Nussloch loess series (Rhine Valley, Germany): the impact of local factors. Boreas. 2005;34(3):363–80. doi: 10.1080/03009480510013060. [DOI] [Google Scholar]
- 14.Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3(5):294–9. [PubMed] [Google Scholar]
- 15.Groenenberg DSJ, Neubert E, Gittenberger E. Reappraisal of the “Molecular phylogeny of Western Palaearctic Helicidae s.l. (Gastropoda: Stylommatophora)”: when poor science meets GenBank. Mol Phylogenet Evol. 2011;61(3):914–23. doi: 10.1016/j.ympev.2011.08.024. [DOI] [PubMed] [Google Scholar]
- 16.Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19(5):455–77. doi: 10.1089/cmb.2012.0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28(12):1647–9. doi: 10.1093/bioinformatics/bts199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hahn C, Bachmann L, Chevreux B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads—a baiting and iterative mapping approach. Nucleic Acids Res. 2013;41(13):e129. [DOI] [PMC free article] [PubMed]
- 19.Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, Fritzsch G, Pütz J, Middendorf M, Stadler PF. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 2013;69(2):313–9. doi: 10.1016/j.ympev.2012.08.023. [DOI] [PubMed] [Google Scholar]
- 20.Laslett D, Canback B. ARWEN: a program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics. 2008;24(2):172–5. doi: 10.1093/bioinformatics/btm573. [DOI] [PubMed] [Google Scholar]
- 21.Williams ST, Foster PG, Littlewood DT. The complete mitochondrial genome of a turbinid vetigastropod from MiSeq Illumina sequencing of genomic DNA and steps towards a resolved gastropod phylogeny. Gene. 2014;533(1):38–47. doi: 10.1016/j.gene.2013.10.005. [DOI] [PubMed] [Google Scholar]
- 22.Abascal F, Zardoya R, Telford MJ. TranslatorX: multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010;38(Web Server issue):W7–13. doi: 10.1093/nar/gkq291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Katoh K, Toh H. Improved accuracy of multiple ncRNA alignment by incorporating structural information into a MAFFT-based framework. BMC Bioinf. 2008;9:212. doi: 10.1186/1471-2105-9-212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hunter SS, Settles ML, New DD, Parent CE, Gerritsen AT. Mitochondrial Genome Sequence of the Galápagos Endemic Land Snail Naesiotus nux. Genome Announc. 2016;4(1):e01362–01315. doi: 10.1128/genomeA.01362-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. 2000;17(4):540–52. doi: 10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- 26.Lanfear R, Calcott B, Ho SYW, Guindon S. PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol Biol Evol. 2012;29(6):1695–701. doi: 10.1093/molbev/mss020. [DOI] [PubMed] [Google Scholar]
- 27.Miller MA, Pfeiffer W, Schwartz T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Gateway Computing Environments Workshop (GCE), 2010: 2010: IEEE; 2010: 1–8.
- 28.Rambaut A, Drummond AJ. Tracer. v. 1.5. 2007. [Google Scholar]
- 29.Rambaut A. FigTree version 1.4. 0. 2012. [Google Scholar]
- 30.Hu F, Zhou L, Tang J. Reconstructing ancestral genomic orders using binary encoding and probabilistic models. In: Bioinformatics Research and Applications. Berlin: Springer; 2013. p. 17–27.
- 31.Pilsbry HA. Zoölogical Position of Partula and Achatinella. 1900. pp. 561–7. [Google Scholar]
- 32.Baker HB. Heterurethrous and aulacopod. Nautilus. 1955;68(4):109–12. [Google Scholar]
- 33.Zilch A. Euthyneura. Handbuch der Paläozoologie. 1960;2(6):401–835. [Google Scholar]
- 34.Dayrat B, Conrad M, Balayan S, White TR, Albrecht C, Golding R, Gomes SR, Harasewych MG, de Frias Martins AM. Phylogenetic relationships and evolution of pulmonate gastropods (Mollusca): new insights from increased taxon sampling. Mol Phylogenet Evol. 2011;59(2):425–37. doi: 10.1016/j.ympev.2011.02.014. [DOI] [PubMed] [Google Scholar]
- 35.Gaitán-Espitia JD, Nespolo RF, Opazo JC. The complete mitochondrial genome of the land snail Cornu aspersum (Helicidae: Mollusca): intra-specific divergence of protein-coding genes and phylogenetic considerations within Euthyneura. PLoS ONE. 2013;8(6):e67299. doi: 10.1371/journal.pone.0067299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kocot KM, Halanych KM, Krug PJ. Phylogenomics supports Panpulmonata: opisthobranch paraphyly and key evolutionary steps in a major radiation of gastropod molluscs. Mol Phylogenet Evol. 2013;69(3):764–71. doi: 10.1016/j.ympev.2013.07.001. [DOI] [PubMed] [Google Scholar]
- 37.Wade CM, Mordan PB, Clarke B. A phylogeny of the land snails (Gastropoda: Pulmonata) Proc Biol Sci/R Soc. 2001;268(1465):413–22. doi: 10.1098/rspb.2000.1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rokas A, Holland PWH. Rare genomic changes as a tool for phylogenetics. Trends Ecol Evol. 2000;15(11):454–9. doi: 10.1016/S0169-5347(00)01967-4. [DOI] [PubMed] [Google Scholar]
- 39.Mordan P, Wade C. Heterobranchia II. Berkeley: Phylogeny and evolution of the Mollusca University of California Press; 2008. pp. 409–26. [Google Scholar]
- 40.Nordsieck H. The system of the Stylommatophora (Gastropoda), with special regard to the systematic position of the Clausiliidae. I: Importance of the excretory and genital systems. Archiv für Molluskenkunde der Senckenbergischen Naturforschenden Gesellschaft. 1985;116(1–3):1–24. [Google Scholar]
- 41.Price MR, Forsman ZH, Knapp I, Hadfield MG, Toonen RJ. The complete mitochondrial genome of Achatinella mustelina (Gastropoda: Pulmonata: Stylommatophora) Mitochondrial DNA Part B. 2016;1(1):183–5. doi: 10.1080/23802359.2016.1149787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Marquardt JD. Mitochondrial Genome Evolution in Pupillid Land Snails. 2013. [Google Scholar]
- 43.Nekola JC, Coles BF. Supraspecific taxonomy in the Vertiginidae (Gastropoda: Stylommatophora). J Moll Stud. 2016;82(1):208–12.
- 44.Baker HB. Puerto Rican pupillids and clausilioids. Nautilus. 1961;75(1):33–6. [Google Scholar]
- 45.Uit de Weerd DR. Delimitation and phylogenetics of the diverse land-snail family Urocoptidae (Gastropoda: Pulmonata) based on 28S rRNA sequence data: a reunion with Cerion. J Molluscan Stud. 2008;74(4):317–29. doi: 10.1093/mollus/eyn023. [DOI] [Google Scholar]
- 46.Vaught KC. A classification of the living Mollusca. Melbourne: Amer Malacologists Inc.; 1989;1–195.
- 47.Groenenberg DSJ, Pirovano W, Gittenberger E, Schilthuizen M. The complete mitogenome of Cylindrus obtusus (Helicidae, Ariantinae) using Illumina next generation sequencing. BMC Genomics. 2012;13(1):114. doi: 10.1186/1471-2164-13-114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang P, Yang HF, Zhou WC, Hwang CC, Zhang WH, Qian ZX. The mitochondrial genome of the land snail Camaenacicatricosa (Muller, 1774) (Stylommatophora, Camaenidae): the first complete sequence in the family Camaenidae. Zookeys. 2014;451:33–48. doi: 10.3897/zookeys.451.8537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Scott B. Phylogenetic relationships of the Camaenidae (Pulmonata: Stylommatophora: Helicoidea) J Molluscan Stud. 1996;62(1):65–73. doi: 10.1093/mollus/62.1.65. [DOI] [Google Scholar]
- 50.Wade CM, Hudelot C, Davison A, Naggs F, Mordan PB. Molecular phylogeny of the helicoid land snails (Pulmonata: Stylommatophora: Helicoidea), with special emphasis on the Camaenidae. J Molluscan Stud. 2007;73(4):411–5. doi: 10.1093/mollus/eym030. [DOI] [Google Scholar]
- 51.Solem A. Classification of the land Mollusca. Pulmonates. 1978;2:49–97. [Google Scholar]
- 52.Nordsieck H. The system of the Stylommatophora (Gastropoda), with special regard to the systematic position of the Clausiliidae, II: Importance of the Shell and distribution. Archiv für Molluskenkunde der Senckenbergischen Naturforschenden Gesellschaft. 1986;117(1–3):93–116. [Google Scholar]
- 53.Briscoe AG, Goodacre S, Masta SE, Taylor MI, Arnedo MA, Penney D, Kenny J, Creer S. Can long-range PCR be used to amplify genetically divergent mitochondrial genomes for comparative phylogenetics? A case study within spiders (Arthropoda: Araneae) PLoS ONE. 2013;8(5):e62404. doi: 10.1371/journal.pone.0062404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gillett CPDT, Crampton-Platt A, Timmermans MJTN, Jordal B, Emerson BC, Vogler AP. Bulk de novo mitogenome assembly from pooled total DNA elucidates the phylogeny of weevils (Coleoptera: Curculionoidea). Mol Biol Evol. 2014;31(8):2223–37. [DOI] [PMC free article] [PubMed]
- 55.Timmermans MJ, Lees DC, Simonsen TJ. Towards a mitogenomic phylogeny of Lepidoptera. Mol Phylogenet Evol. 2014;79:169–78. doi: 10.1016/j.ympev.2014.05.031. [DOI] [PubMed] [Google Scholar]
- 56.Crampton-Platt A, Timmermans MJTN, Gimmel ML, Kutty SN, Cockerill TD, Khen CV, Vogler AP. Soup to tree: the phylogeny of beetles inferred by mitochondrial metagenomics of a Bornean rainforest sample. Mol Biol Evol. 2015;32(9):2302–16. doi: 10.1093/molbev/msv111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.González VL, Kayal E, Halloran M, Shrestha Y, Harasewych MG. The complete mitochondrial genome of the land snail Cerion incanum (Gastropoda: Stylommatophora) and the phylogenetic relationships of Cerionidae within Panpulmonata. J Moll Stud. 2016;82(4):525–33.
- 58.Breeschoten T, Doorenweerd C, Tarasov S, Vogler AP. Phylogenetics and biogeography of the dung beetle genus Onthophagus inferred from mitochondrial genomes. Mol Phylogenet Evol. 2016;105:86–95. doi: 10.1016/j.ympev.2016.08.016. [DOI] [PubMed] [Google Scholar]
- 59.He ZP, Dai XB, Zhang S, Zhi TT, Lun ZR, Wu ZD, Yang TB. Complete mitochondrial genome of the giant African snail, Achatina fulica (Mollusca: Achatinidae): a novel location of putative control regions (CR) in the mitogenome within Pulmonate species. Mitochondrial DNA Part A. 2014;27(2):1084–85. [DOI] [PubMed]
- 60.Yang X, Xie G-L, Wu X-P, Ouyang S. The complete mitochondrial genome of Chinese land snail Aegista aubryana (Gastropoda: Pulmonata: Bradybaenidae) Mitochondrial DNA Part A. 2016;27(5):3538–9. doi: 10.3109/19401736.2015.1074207. [DOI] [PubMed] [Google Scholar]
- 61.Huang C-W, Lin S-M, Wu W-L. Mitochondrial genome sequences of landsnails Aegista diversifamilia and Dolicheulota formosensis (Gastropoda: Pulmonata: Stylommatophora). Mitochondrial DNA. 2015(ahead-of-print):1–3. [DOI] [PubMed]
- 62.Lin J-H, Zhou W, Ding H-L, Wang P, Ai H-M. The mitochondrial genome of the land snail Cernuella virgata (Da Costa, 1778): the first complete sequence in the family Hygromiidae (Pulmonata, Stylommatophora) ZooKeys. 2016;589:55. doi: 10.3897/zookeys.589.7637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Terrett JA, Miles S, Thomas RH. Complete DNA sequence of the mitochondrial genome of Cepaea nemoralis (Gastropoda: Pulmonata) J Mol Evol. 1996;42(2):160–8. doi: 10.1007/BF02198842. [DOI] [PubMed] [Google Scholar]
- 64.Yamazaki N, Ueshima R, Terrett JA, Yokobori S-i, Kaifu M, Segawa R, Kobayashi T, Numachi K-i, Ueda T, Nishikawa K. Evolution of pulmonate gastropod mitochondrial genomes: comparisons of gene organizations of Euhadra, Cepaea and Albinaria and implications of unusual tRNA secondary structures. Genetics. 1997;145(3):749–58. doi: 10.1093/genetics/145.3.749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Deng PJ, Wang WM, Huang XC, Wu XP, Xie GL, Ouyang S. The complete mitochondrial genome of Chinese land snail Mastigeulota kiangsinensis (Gastropoda: Pulmonata: Bradybaenidae). Mitochondrial DNA Part A. 2014;27(2):1441–42. [DOI] [PubMed]
- 66.DeJong RJ, Emery AM, Adema CM. The mitochondrial genome of Biomphalaria glabrata (Gastropoda: Basommatophora), intermediate host of Schistosoma mansoni. J Parasitol. 2004;90(5):991–7. doi: 10.1645/GE-284R. [DOI] [PubMed] [Google Scholar]
- 67.Jannotti-Passos LK, Ruiz JC, Caldeira RL, Murta SM, Coelho PM, Carvalho OS. Phylogenetic analysis of Biomphalaria tenagophila (Orbigny, 1835) (Mollusca: Gastropoda) Mem Inst Oswaldo Cruz. 2010;105(4):504–11. doi: 10.1590/S0074-02762010000400027. [DOI] [PubMed] [Google Scholar]
- 68.Liu GH, Wang SY, Huang WY, Zhao GH, Wei SJ, Song HQ, Xu MJ, Lin RQ, Zhou DH, Zhu XQ. The complete mitochondrial genome of Galba pervia (Gastropoda: Mollusca), an intermediate host snail of Fasciola spp. PLoS ONE. 2012;7(7):e42172. doi: 10.1371/journal.pone.0042172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nolan JR, Bergthorsson U, Adema CM. Physella acuta: atypical mitochondrial gene order among panpulmonates (Gastropoda) J Moll Stud. 2014;80(4):388–99. doi: 10.1093/mollus/eyu025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Grande C, Templado J, Cervera JL, Zardoya R. Phylogenetic relationships among Opisthobranchia (Mollusca: Gastropoda) based on mitochondrial cox 1, trnV, and rrnL genes. Mol Phylogenet Evol. 2004;33(2):378–88. doi: 10.1016/j.ympev.2004.06.008. [DOI] [PubMed] [Google Scholar]
- 71.Feldmeyer B, Hoffmeier K, Pfenninger M. The complete mitochondrial genome of Radix balthica (Pulmonata, Basommatophora), obtained by low coverage shot gun next generation sequencing. Mol Phylogenet Evol. 2010;57(3):1329–33. doi: 10.1016/j.ympev.2010.09.012. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Assembly strategy. (A) Regions where reads could not be mapped to the reference sequence (B) Regions that broke up the ‘de novo’ assembly (C) Mapping and de novo contigs merged. (PDF 15 kb)
Secondary structures of the inferred tRNAs of Orcula dolium. (PDF 107 kb)
Data Availability Statement
The annotated mitogenome of Orcula dolium is available from GenBank under accesssion number KJ867421.