

Abstract
Background
Formic acid is an inhibitory compound present in lignocellulosic hydrolysates. Understanding the complex molecular mechanisms underlying Saccharomyces cerevisiae tolerance to this weak acid at the system level is instrumental to guide synthetic pathway engineering for robustness improvement of industrial strains envisaging their use in lignocellulosic biorefineries.
Results
This study was performed to identify, at a genome-wide scale, genes whose expression confers protection or susceptibility to formic acid, based on the screening of a haploid deletion mutant collection to search for these phenotypes in the presence of 60, 70 and 80 mM of this acid, at pH 4.5. This chemogenomic analysis allowed the identification of 172 determinants of tolerance and 41 determinants of susceptibility to formic acid. Clustering of genes required for maximal tolerance to this weak acid, based on their biological function, indicates an enrichment of those involved in intracellular trafficking and protein synthesis, cell wall and cytoskeleton organization, carbohydrate metabolism, lipid, amino acid and vitamin metabolism, response to stress, chromatin remodelling, transcription and internal pH homeostasis. Among these genes is HAA1 encoding the main transcriptional regulator of yeast transcriptome reprograming in response to acetic acid and genes of the Haa1-regulon; all demonstrated determinants of acetic acid tolerance. Among the genes that when deleted lead to increased tolerance to formic acid, TRK1, encoding the high-affinity potassium transporter and a determinant of resistance to acetic acid, was surprisingly found. Consistently, genes encoding positive regulators of Trk1 activity were also identified as formic acid susceptibility determinants, while a negative regulator confers protection. At a saturating K+ concentration of 20 mM, the deletion mutant trk1Δ was found to exhibit a much higher tolerance compared with the parental strain. Given that trk1Δ accumulates lower levels of radiolabelled formic acid, compared to the parental strain, it is hypothesized that Trk1 facilitates formic acid uptake into the yeast cell.
Conclusions
The list of genes resulting from this study shows a few marked differences from the list of genes conferring protection to acetic acid and provides potentially valuable information to guide improvement programmes for the development of more robust strains against formic acid.
Electronic supplementary material
The online version of this article (doi:10.1186/s13068-017-0781-5) contains supplementary material, which is available to authorized users.
Keywords: Formic acid tolerance, Formic acid toxicity, Chemogenomic analysis, Trk1, Haa1, Lignocellulosic hydrolysates, Yeast robustness
Background
Formic acid (pK a = 3.74; octanol–water partition coefficient, LogP = −0.54 [1]) is the simplest monocarboxylic acid in nature. The worldwide production of this acid is in large part (35%) used to prevent fungal and bacterial growth in silage [2]. Formic acid is also among the inhibitors found in lignocellulosic hydrolysates, typically at concentrations around 30 mM although they are variable depending on the type of treatment used to prepare the hydrolysates [3–6]. A comprehensive understanding of the mechanisms underlying tolerance to formic acid is therefore important to guide the design of more robust industrial yeast strains able to more efficiently use lignocellulosic hydrolysates as growth medium in biorefineries [7].
The straightforward exploitation of Omic approaches combined with metabolic engineering and synthetic biology strategies in Saccharomyces cerevisiae is considered instrumental to guide the improvement of yeast strains for production of biofuels and added-value chemicals [8–14]. The application of Omic analyses to elucidate the genes and the pathways involved in yeast adaptive response and tolerance to stress induced by weak acids, in particular acetic acid, has been essential to identify their molecular targets and guide the design of more robust strains for efficient production of organic acids or to cope with their presence in the lignocellulosic hydrolysates and to their increased concentration during fermentation [8, 11, 15–19]. However, equivalent studies to unravel the molecular determinants of toxicity and tolerance to formic acid, especially at a genome-wide scale, are scarce [20].
As described for other weak acids, it is believed that the undissociated form of formic acid may enter the yeast cells through simple diffusion and dissociate in the cytosol, leading to the disruption of the electrochemical gradient across the plasma membrane, to the decrease of internal pH and the triggering the production of reactive oxygen species (ROS) [20–22]. Adaptation to weak acids involves the increase of activity of plasma membrane H+-ATPase and vacuolar H+-ATPase to counteract the acidification of cytosolic pH and the dissipation of plasma membrane H+ gradient [23, 24]. Several other mechanisms of toxicity and tolerance to weak acids have been reported [8, 9, 15, 16, 18]. Recently, genes involved in lipid metabolism emerged as determinants of yeast response and tolerance to acetic acid stress [15], the Tor complex 2-Ypk1 signalling pathway being involved in this response [25].
Key regulatory pathways involved in yeast global response and tolerance to weak acids have been unveiled exploring genome-wide approaches [15, 16, 18, 26]. Among them is the signalling pathway regulated by the transcription factor Haa1. Yeast response to short-chain weak acids, as it is the case of acetic, propionic and lactic acids [18, 27], is regulated by Haa1, which is considered the main orchestrator of the reprograming of yeast transcriptome in response to acetic acid [28]. Indeed, Haa1 regulates a large regulon including 80% of the acetic acid-stress responsive genes, including those involved in transcription (e.g., MSN4, MCM1, FKH2 and COM2), multidrug resistance (MDR) transport (e.g., TPO2, TPO3 and AQR1), cell wall remodelling (e.g., YGP1 and SPI1), nucleic acid processing (e.g., SAP30) and in lipid (e.g., YCP1 and SUR2) and carbohydrate metabolism (HRK1) [15, 28]. Considering the central role of Haa1 in yeast adaptive response and tolerance to acetic acid and other low-chain fatty acids, the manipulation of the Haa1-signalling pathway either by increasing the expression of HAA1, based on the expression of extra copies of the gene and/or the use of more efficient promoters [29–33], or by the insertion of beneficial mutations in the coding sequence [33, 34], was successfully attempted.
Formic acid was found to be co-utilized with glucose by S. cerevisiae in glucose-limited cultures (7.5 g L−1) under aerobic conditions to generate NADH molecules for respiratory dissimilation [35, 36]. This weak acid is rapidly converted into CO2 and water by formate dehydrogenases (FDH) encoded by FDH1 and FDH2 genes with NADH molecules being produced in the process, and this activity was proposed to contribute to formic acid detoxification [36–38]. In mammalian and plant cells, formate is believed to bind cytochrome c oxidase (complex VI of the electron transport chain), which catalyses the reduction of molecular oxygen to water using cytochrome c as the electron donor [39, 40]. The inhibition of this last step of the electron transport chain hampers proton gradient maintenance at physiological values and ultimately ATP synthesis [41]. In mammalian cells, this process was found to be accompanied by an increase of ROS production (i.e., superoxide anions and hydroxyl radicals) in the mitochondria, leading to the oxidative damage of proteins, lipids and DNA [42–45]. With the exception of a proteomic analysis [20], little has been done to understand the cytotoxic effects of formic acid and the response mechanisms in S. cerevisiae. This study is a first attempt to identify, in a comprehensive manner, genes involved in yeast tolerance and susceptibility to formic acid at a genome-wide scale. Among the selected determinants identified through the chemogenomic analysis performed is the Haa1 regulon, for the first time demonstrated to be required for tolerance to formic acid. Quite unexpectedly, the high-affinity potassium transporter Trk1 [46] and the positive regulators of its activity emerged as susceptibility determinants to this acid. Trk1 activity is positively regulated by the Serine/threonine protein kinases Sat4 and Hal5 [47], and negatively regulated by the Serine/threonine protein phosphatase Ppz1 [48, 49]. The activity of the Trk1 transporter is also positively regulated by the activity of Hal3, a negative regulatory subunit of the protein phosphatase Ppz1 [48]. Contrasting with the results from this study, TRK1 was found to confer protection against acetic acid in yeast [15]. Moreover, yeast susceptibility to acetic acid was found to be attenuated by growth medium supplementation with K+ ions [15]. Considering the results obtained in this study, the unexpected role of Trk1 in S. cerevisiae tolerance to formic acid was examined.
Methods
Strains and growth media
The haploid parental strain S. cerevisiae BY4741 (MATa, his3Δ1, leu2Δ0, met15Δ0, ura3Δ0) and the derived collection of single deletion mutants, in which each non-essential gene was individually deleted, were obtained from EUROSCARF (Frankfurt, Germany). Yeast cells were batch cultured in YPD medium containing 2% (w/v) glucose (Merck), 2% (w/v) yeast extract (VWR International) and 1% (w/v) peptone (VWR International), or in MM4 medium containing 1.7 g L−1 Yeast Nitrogen Base (YNB) w/o amino acids or ammonium (Difco), 20 g L−1 glucose, 2.65 g L−1 (NH4)2SO4 (Scharlau), supplemented with 20 mg L−1 methionine, 30 mg L−1 lysine, 60 mg L−1 leucine, 40 mg L−1 tryptophan, 20 mg L−1 histidine and 20 mg L−1 uracil (all from Sigma-Aldrich). Cell cultures were cultivated at 30 °C with orbital agitation (250 rev min−1). To test the growth of wild-type BY4741 and trk1Δ- or trk2Δ-derived strains under potassium limitation, an ammonium phosphate-derived medium was used, containing 0.492 g L−1 MgSO4·7H2O (Merck), 0.02 g L−1 CaCl2 (Sigma-Aldrich), 1.056 g L−1 (NH4)2HPO4 (Merck), 3.96 g L−1 (NH4)2SO4, 20 g L−1 glucose (Merck), 2 mg L−1 niacin, 2 mg L−1 pyridoxine, 2 mg L−1 thiamine, 2 mg L−1 pantothenate, 0.02 mg L−1 biotin, 20 mg L−1 methionine, 60 mg L−1 leucine, 20 mg L−1 histidine and 20 mg L−1 uracil supplemented with the desired concentration of KCl (all from Sigma). A 3 M formic acid stock solution (Sigma-Aldrich) was prepared in water and the pH of the solution adjusted to 4.0 or to 4.5 with NaOH depending on the growth medium supplementation. Solid media were prepared by addition of 20 g L−1 agar (IberAgar) to the liquid media.
High-throughput screening of the deletion mutant collection
To screen the EUROSCARF deletion mutant collection for susceptibility or tolerance to formic acid, strains were grown for 16 h in MM4 medium in 96-well plates as described before [15, 16]. Using a 96-pin replica platter, the cell suspensions were spotted onto the surface of MM4 solid medium acidified with HCl to pH 4.5 and supplemented, or not, with formic acid to a final concentration of 60, 70 or 80 mM. Depending on the severity of growth inhibition, plates were incubated at 30 °C for 2 or 3 days. Two datasets containing genes identified as determinants of resistance (Additional file 1, Fig. 1a) or of susceptibility (Additional file 2, Fig. 1b) to formic acid were obtained. Both datasets were clustered according to biological process Gene Ontology (GO) assignments using the MIPS functional catalogue (http://mips.helmholtz-muenchen.de/funcatDB; over-representation of functional categories was considered for a p value ≤0.01) and this analysis was complemented using the information available in Saccharomyces Genome Database (SGD) (http://www.yeastgenome.org). The simultaneous occurrence of genes from the same dataset in a shared metabolic pathway or signalling pathway was determined using the Kyoto Encyclopedia for Genes and Genomes (KEGG) Mapper Search pathway tool (http://www.genome.jp/kegg/tool/map_pathway1.html).
Fig. 1.
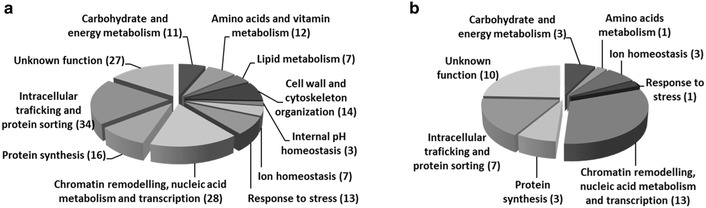
Biological functions found to be enriched in the datasets of genes found to confer tolerance or susceptibility to formic acid. a Genes found to confer tolerance to formic acid, listed in Additional file 1, were clustered according their biological process GO assignments using the MIPS functional catalogue (http://mips.helmholtz-muenchen.de/proj/funcatDB), and functional categories were considered to be over-represented if p value ≤0.01. b Genes found to confer susceptibility to formic acid were manually clustered according to the information available in SGD (http://www.yeastgenome.org) into the indicated categories. The number of genes identified within each category is indicated in brackets
Susceptibility assays to formic acid of selected strains
The susceptibility of selected S. cerevisiae BY4741-derived deletion mutants was compared with the parental strain in shake flasks or petri dishes in MM4 medium (pH 4.0) or in solid MM4 medium (pH 4.5). To test the strain susceptibility to formic acid (30 mM) in liquid MM4 medium, mid-exponential cells pre-grown in MM4 medium (pH 4.0) to an optical density at 600 nm (OD600) of 0.8 ± 0.05 were used to inoculate the same basal medium, either or not, supplemented with 30 mM formic acid, with an initial OD600 of 0.05 ± 0.005. Growth curves were followed based on the increase of the OD600 and by determining the number of colony-forming units (CFU) per ml of cell suspension in solid YPD medium. To test the susceptibility of selected mutants to formic acid in solid MM4 medium (pH 4.5), mid-exponential cells grown in MM4 liquid medium (pH 4.5) to a final OD600 of 0.8 ± 0.05 were diluted to an OD600 of 0.05 ± 0.005 and spotted onto solid medium supplemented or not with formic acid (50 and 80 mM). Plates were incubated for 3 days at 30 °C.
The susceptibility of wild-type BY4741 and trk1Δ and trk2Δ mutants to formic acid was also compared by spot assays performed in the presence of increasing concentrations of potassium. The solid ammonium phosphate-derived medium (at pH 4.5) was supplemented with KCl (stock solution of 2 M) to final concentrations of 0.5, 2 and 20 mM and supplemented or not with formic acid at the indicated concentrations. The cells used to inoculate the agar plates were grown until an OD600 of 0.4 in liquid ammonium phosphate-derived medium (at pH 4.5) supplemented with 20 mM of KCl. Cells were diluted in sterile H2O to an OD600 of 0.05, and this solution was used to prepare 1:3 and 1:15 diluted suspensions. Four microliters of each cell suspension were spotted onto solid media and the plates incubated at 30 °C for 5 days, depending on the severity of growth inhibition.
Effect of formic acid in the transcriptional level from Haa1-regulated genes
Real time RT-PCR was used to compare mRNA levels from a number of Haa1-target genes (TPO2, TPO3, COM2, HRK1, SAP30 and SUR2) in BY4741 and BY4741haa1Δ genetic backgrounds, cultivated in MM4 medium (pH 4.0) supplemented or not with 30 mM of formic acid, as described above for formic acid susceptibility assays, except that the initial OD600 of the growth curves which was set at 0.4 ± 0.05. Cells incubated for 30 min in presence or absence of 30 mM of formic acid were harvested by centrifugation at 5000g, 4 °C, for 3 min and the pellet immediately frozen in liquid nitrogen and stored at −80 °C until used. Total RNA was extracted from these pellets using the hot phenol method [50], treated with DNAaseI (Invitrogen) according to the manufacturer instructions, and 1 μg of the treated RNA was used in the reverse transcription step (Taqman® reverse transcription reagents). Primers used for the PCR amplification of the selected genes (Additional file 3) were designed using the Primer Express Software (Applied Biosystems), and 62.5 ng of the synthesized cDNA were used as template for the amplification step. ACT1 gene was used as the endogenous control. Values of mRNA determined for BY4741 cells cultivated in the absence of the acid were used as normalization factor, being the mRNA levels from the different genes determined for the same RNA sample set as 1.
Cellular accumulation assays for [14C]formic acid
BY4741 and BY4741trk1Δ cells, pre-grown in MM4 medium (pH 4.0), were used to inoculate fresh medium with an initial OD of 0.1 and cultivated at 30 °C with orbital agitation (250 rev min−1) until an OD600 of 0.3–0.5 was reached. An appropriate volume of cell suspension was filtered and cells resuspended in 5 ml of fresh MM4 medium (pH 4.0) with an OD600 of 0.5 ± 0.05 and incubated for 10 min at 30 °C (150 rev min−1) before the accumulation experiment was performed. The experiment started after the addition of cold formic acid (final concentration of 30 mM) and 2.5 μL of [14C]formic acid ([14C]sodium formate, 1 mCi ml−1, ARC0163A, American Radiolabeled Chemicals). At the indicated time points, 200 μL of the cell suspension were filtered through glass microfiber filters (Whatman GF/C) and washed twice with ice-cold TM buffer containing 100 mM MES and 41 mM Tris buffers (both from Sigma, pH of 4.0 adjusted with HCl). The filters containing the adsorbed cells were placed into scintillation vials containing 5 ml of scintillation cocktail (Ultima Gold MV, PerkinElmer) and the radioactivity of the samples measured in a Beckman LS 5000TD scintillation counter. The radioactivity of 100 μL of the culture supernatant was also measured at each time point. The accumulation ratio of intracellular/extracellular [14C]formic acid at the indicated time points was calculated considering that the internal volume (V i) of cells of both strains was equal to 2.5 μL per mg of dry weight [23]. The difference between the means determined for wild-type and trk1Δ cells were considered statistically significant for p values <0.05 using a one-way ANOVA.
Results
Genome-wide identification of S. cerevisiae genes that confer tolerance to formic acid
The EUROSCARF collection, containing approximately 5000 single deletion mutant strains devoid of all non-essential S. cerevisiae genes, was screened to identify genes contributing to tolerance to formic acid-induced stress, based on the identification of those mutants exhibiting increased susceptibility to 60, 70 or 80 mM of formic acid at pH 4.5, when compared to the parental strain (Additional file 4). This chemogenomic analysis allowed the identification of a total of 172 tolerance genes (Additional file 1). Two levels of susceptibility were considered, depending on the severity of growth inhibition of the deletion mutants when compared to the wild-type strain, as illustrated in Additional file 4. All genes that were found to be determinants of tolerance to formic acid (despite the level of susceptibility conferred by the deletion of the gene) are indicated in Additional file 1. These genes were grouped using the MIPS functional catalogue (http://mips.helmholtz-muenchen.de/proj/funcatDB). The different functional categories identified as being over-represented (p value ≤0.01) in the dataset, when compared to the genome and the list of genes and the number of genes in each category, are depicted in Fig. 1a and in Additional file 1.
Haa1 and the Haa1-regulon are involved in response and tolerance to formic acid
Among the tolerance genes identified based on the deletion mutant collection screening, those encoding the transcription factor Haa1 and genes of the Haa1-regulon (direct or indirect target genes for Haa1 regulation under acetic acid stress) were selected for further studies. In fact, their prominent role in yeast adaptive response to acetic acid and other short-chain fatty acids has been proposed. To confirm the putative role of Haa1 and of genes of the Haa1-regulon in yeast response and tolerance to formic acid, the susceptibility to this weak acid of the parental strain and of several individual deletion mutants with genes of the Haa1-regulon deleted, in particular, tpo2Δ, tpo3Δ, com2Δ, sur2Δ, hrk1Δ and sap30Δ, was compared based on formic acid-induced increase of the duration of the latency period for these mutant strains when cultivated in MM4 medium (pH 4.0) supplemented with 30 mM of formic acid (Fig. 2b). With the exception of TPO2 gene (data not shown), all the Haa1-regulated genes tested were shown to contribute significantly to yeast tolerance to formic acid, exerting a significant effect by decreasing the duration of formic acid-induced lag-phase (Fig. 2b). As described for acetic acid [15], a particularly marked effect of HRK1 and SAP30 deletion in the increase of formic acid susceptibility was observed, being even higher than the phenotype registered for cells with the HAA1 deleted (Fig. 2b).
Fig. 2.
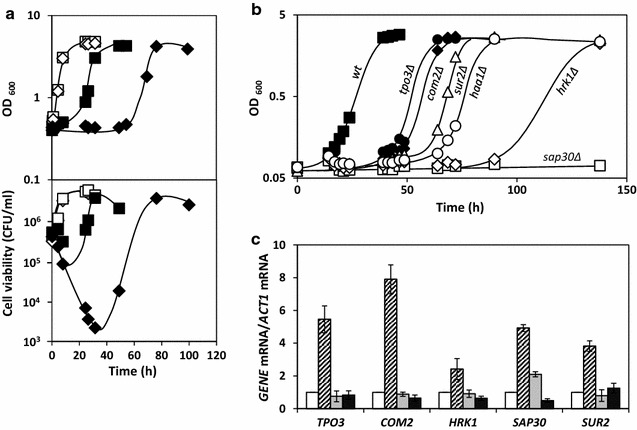
The expression of HAA1 and of Haa1-regulated genes is required for S. cerevisiae adaptation and tolerance to formic acid. a Cells of the parental strain (squares) and deletion mutant haa1Δ (diamons) were grown in liquid MMB medium (pH 4.0) supplemented (closed symbols), or not (open symbols), with 30 mM of formic acid (upper panel) and growth was followed based on culture optical density at 600 nm (OD600). Cell viability (CFU ml−1) was also determined during cultivation (lower panel). b Growth curves of the parental BY4741 strain and of strains with the Haa1-target genes deleted were incubated in liquid MMB medium (pH 4.0) supplemented with 30 mM of formic acid. The growth curves indicated in panels (a) and (b) are representative of at least three independent experiments. c Comparison of the mRNA levels from HAA1 and selected Haa1-regulated genes assessed in wild-type cells incubated in basal medium (white bars), or exposed to formic acid (30 mM) for 30 min (diagonal stripes bars) and in haa1Δ cells in cultivated in those same conditions (grey and black bars, respectively). The bars represent the average values of at least three independent experiments, and the error bars the associated standard deviation
The transcriptional levels from the Haa1-target genes TPO3, COM2, SUR2, HRK1 and SAP30 were also compared in parental strain and haa1Δ mutant during early response to formic acid (Fig. 2c) to test this weak acid effect in the hypothesized increase of mRNA levels from Haa1-target genes and its dependence on Haa1. To compare the mRNA levels from the above referred genes, the parental and haa1Δ cells were cultivated in MM4 medium at pH 4.0 and collected after 30 min of incubation in medium supplemented, or not, with 30 mM of formic acid (Fig. 2c). Transcription levels from selected Haa1-target genes under formic acid stress were found to be significantly higher compared with those in unstressed parental cells, while this formic acid-induced transcriptional activation of the target genes was drastically reduced in cells with the HAA1 gene deleted, supporting the idea that Haa1 is an activator of these genes increased transcription in response to formic acid stress (Fig. 2c). The mRNA levels from the SAP30 gene in unstressed haa1Δ cells were found to be higher than those registered in the parental strain but no likely justification can be put forward for this result at this time.
Genome-wide identification of S. cerevisiae genes whose expression increases susceptibility to formic acid
The chemogenomic analysis carried out also led to the identification of 41 genes whose individual deletion results in increased tolerance to formic acid of each single mutant when compared to the parental strain, as illustrated in Additional file 4. According to the MIPS functional catalogue, no significant enrichment in GO terms (p value ≤0.01) could be identified, probably due to the high variety of functions associated to this small gene dataset. For this reason, the identified determinants of susceptibility to formic acid were manually grouped according to their description in SGD (www.yeastgenome.org) into the following categories: “carbohydrate and energy metabolism”, “amino acids metabolism”, “ion homeostasis”, “response to stress”, “protein synthesis”, “intracellular trafficking”, “protein sorting” and “chromatin remodelling, nucleic acid metabolism and transcription” (Additional file 2 and Fig. 1b). Among the genes found to contribute to increased yeast susceptibility to formic acid, two genes are highlighted: the gene CYC3, encoding a cytochrome c heme lyase, and the ARN2 gene, encoding a transporter of siderophore–iron complexes involved in iron uptake.
From the EUROSCARF collection screening, genes SAT4 and HAL5, two positive regulators of Trk1 activity, were found to confer susceptibility to yeast. Based on this result, it was considered of interest to systematically compare the susceptibility to formic acid of an individual mutant with the TRK1 gene deleted and of mutants with the positive (HAL3, SAT4 or HAL5) and negative (PPZ1) regulators [47, 51] of its activity individually deleted (Fig. 3a, b). Surprisingly, results indicate that the TRK1 gene, encoding the high-affinity potassium transporter [46], as well as genes SAT4 and HAL5 are determinants of susceptibility to formic acid (Additional file 2, Fig. 3a), suggesting a detrimental effect of the biological activity of Trk1 in yeast protection to formic acid. Consistent with this hypothesis, the mutant with the gene encoding the negative regulator of Trk1, Ppz1, deleted is more susceptible to formic acid than the parental strain, whereas the mutant deleted for the gene encoding the negative regulator of Ppz1, Hal3, is more tolerant to the acid (Fig. 3a, b).
Fig. 3.
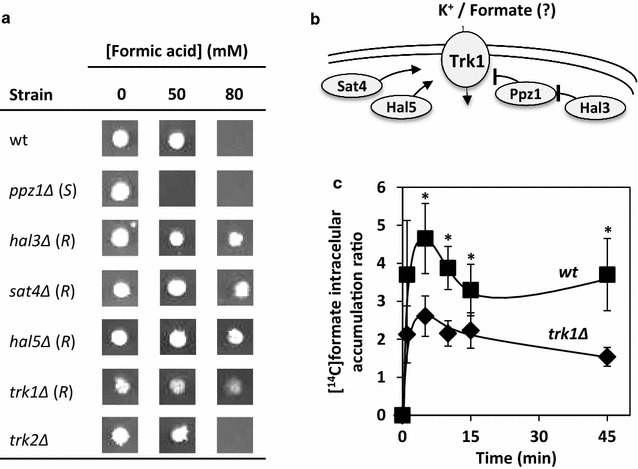
TRK1 gene and genes encoding positive/negative regulators of Trk1 activity are determinants of susceptibility/tolerance to formic acid. a Wild-type (wt), trk1Δ and deletion mutants where the genes coding for regulators of Trk1 activity (HAL3, SAT4, HAL5 and PPZ1) were deleted were tested for their susceptibility to formic acid in solid MMB medium (pH 4.5) supplemented with the acid at the indicated concentrations. Resistance (R) and susceptibility (S) phenotypes. b Schematic representation of the positive and negative regulators of Trk1 activity. c Time-course accumulation of [14C]formic acid in non-adapted wild-type cells and trk1Δ cells cultivated in MM4 growth medium (at pH 4.0) after exposure to 30 mM of formic acid during a 45-min period. Values represent the average of at least three independent experiments and the error bars represent the associated standard deviation. The asterisks indicate a significant different (p value <0.05) between the accumulation of [14C]formic acid in wild-type and trk1Δ cells at that time point
However, the deletion of TRK2, coding for the low-affinity potassium transporter, had no detectable effect on yeast susceptibility to formic acid (Fig. 3a). Since the screening of the deletion mutant phenotypes was performed at a non-limiting K+ concentration (8 mM), to understand the conjugated effect of potassium availability and the presence of Trk1 or Trk2 in yeast tolerance to formic acid, the susceptibility to formic acid of wild-type BY4147 cells and TRK1 or TRK2 deleted mutants was compared by spot assays at K+ concentrations ranging from 0.5 to 20 mM of KCl (Fig. 4). At the higher concentration of 20 mM of KCl, ectopic uptake of K+ is sufficient to fulfil cellular requirements [52]. Results show that the deletion of the TRK1 gene increases, at the highest K+ concentration tested, the tolerance of yeast cells to high concentrations of formic acid, allowing growth at 40 and 50 mM under the experimental conditions used (Fig. 4). However, the effect of TRK2 gene deletion was not detected at any of the K+ concentrations tested (Fig. 4). Based on these results, it was hypothesized that the high-affinity K+ transporter, Trk1, besides its main biological function, may also facilitate the uptake of formic acid/formate into the yeast cell. This hypothesis is consistent with the higher accumulation of radiolabeled [14C]formic acid in the parental strain compared with trk1Δ cells (Fig. 3c).
Fig. 4.
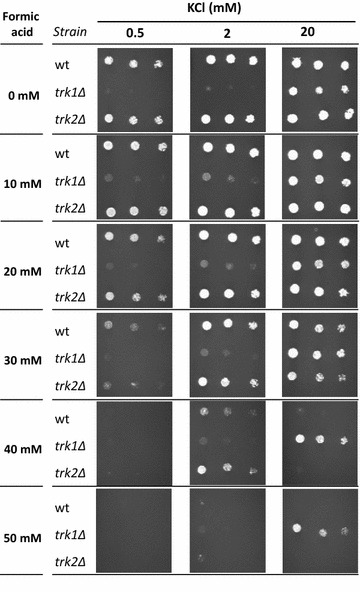
Effect of potassium concentration in tolerance to formic acid of yeast cells with different capacities of K+ uptake. Comparison of the susceptibility of wild-type (wt) BY4741, trk1Δ and trk2Δ deletion mutants to different concentrations of formic acid (0, 10, 20, 30, 40 and 50 mM) in ammonium phosphate medium supplemented with increasing concentrations of KCl (0.5, 2 and 20 mM). Serial dilutions (1:0, 1:3 and 1:15) of a water-diluted cellular suspension with an OD600 of 0.05 were spotted onto agar plates and incubated during 5 days at 30 °C
Discussion
From the chemogenomic analysis performed in this study, several genes belonging to the functional categories “intracellular trafficking and protein sorting”, “chromatin remodelling, nucleic acid metabolism and transcription” and “intracellular pH homeostasis” were found to be over-represented in the dataset of the genes required for yeast adaptation and tolerance to formic acid. These categories were also found to be enriched among the genes required for yeast maximal tolerance to acetic and propionic acids [15, 16]. Vacuolar protein sorting-associated genes have been implicated in multidrug resistance (MDR) [53], and acetic acid-triggered intracellular acidification was shown to lead to trafficking defects, hampering vesicle shuffling from endosome to vacuole [54]. Since formic acid is believed to share part of the cytotoxic effects associated to acetic and propionic acids, in particular the induction of intracellular acidification, the protective effect conferred by the genes grouped in those categories against formic acid was not surprising.
Genes involved in oxidative stress response were also found to confer tolerance to formic acid, consistent with recent evidences indicating that formic acid induces yeast apoptosis, accompanied by the increase of ROS production [20]. Also, the expression of the ARN2 gene, encoding a transporter for siderophore-iron complexes involved in iron uptake, was found to increase yeast susceptibility to formic acid possibly due to iron ability to catalyse ROS production via Fenton reaction. Collectively, these results indicate that formic acid induces oxidative stress and cell damage.
The deletion of CYC3, encoding the enzyme that catalyses the insertion of an heme group into the apo-form of cytochrome c [55], was found to confer susceptibility to formic acid. In yeast, CYC3 deletion leads to increased H2O2 levels and consequently to increased oxidative stress [56]. However, our results suggest that the deletion of the CYC3 gene is beneficial for yeast tolerance to formic acid. Since formate is believed to bind cytochrome c oxidase (complex VI of the electron transport chain), specifically inhibiting cytochrome c oxidase activity [39, 40, 45], this effect may affect the re-oxidation of cytochrome c with consequences for the performance of CYC3-expressing cells.
Two genes (GPD2 and GPP2) involved in glycerol biosynthesis were also identified as determinants of tolerance to formic acid, as well as FPS1, encoding a aquaglyceroporin involved in the control of glycerol homeostasis [54, 57]. These results are in line with those reported for non-S. cerevisiae yeast strains exhibiting higher intracellular levels of glycerol, presumably acting in osmoprotection [58]. The production of glycerol by glycerol 3-phosphate dehydrogenase (GPD2) contributes to reduce the NADH pool which is important since formate dehydrogenase activity generates NADH and is believed to be required for formic acid detoxification [36]. For the same reason, the role of GPP2, involved in the catalysis of the last step of glycerol synthesis, is also a determinant of formic acid tolerance.
Genes involved in ergosterol and long-fatty acid metabolism and in cell wall biosynthesis and integrity were also identified as determinants of tolerance to formic acid. Changes in the saturation degree of fatty acid acyl chains, phospholipids and plasma membrane ergosterol content are believed to affect cell permeability to weak acids [15, 16, 59]. The increased saturation degree of fatty acids acyl chains leads to a more packed membrane structure that is more impermeable to weak acids [60]. Changes in the ergosterol content of plasma membrane influence its permeability and fluidity, and can even affect the activity of membrane transporters including those involved in multidrug resistance [59, 61].
Several genes involved in amino acid biosynthesis were also found to confer increased tolerance to formic acid, suggesting that this acid may cause a depletion of their pool, as reported for acetic acid [26, 62]. On the other hand, it is important to mention that weak acids inhibit amino acid uptake, by affecting the activity of the amino acids permeases, thus exacerbating the influence of the auxotrophic marks present in the parental strain in the susceptibility phenotypes [62, 63]. Since S. cerevisiae BY4741 and the derived deletion mutant collection have several auxotrophic markers, it is possible that the susceptibility phenotypes observed with strains where genes encoding proteins involved in amino acids homeostasis and synthesis are deleted are an artefact related with those auxotrophic markers.
Among the determinants of resistance identified in this chemogenomic analysis is the VAM6 gene, encoding a guanine nucleotide exchange factor that is an activator of the Tor Complex 1 (TORC1) [64] and two genes encoding targets of TORC1 (SCH9 and RTG2). The TORC1 pathway is activated in response to carbon and amino acid starvation and hyperosmotic and redox stresses (reviewed in [65]) and also regulates apoptosis in acetic acid-challenged cells [26]. Our results suggest that the TORC1 pathway may also mediate yeast response to formic acid, possibly in response to amino acid starvation or osmotic or oxidative stress-induced stimuli [21, 26].
The HAA1 gene, encoding the main orchestrator of S. cerevisiae global response to acetic acid [15], was demonstrated to be a determinant of tolerance to formic acid, as well as the deletion of Haa1-target genes TPO3, YGP1, SAP30 and HRK1. Moreover, the transcriptional levels from the above mentioned Haa1-target genes increase in response to formic acid and this transcriptional activation is dependent on Haa1, suggesting that the Haa1-regulon is also activated in response to formic acid stress.
A concentration of 30 mM of formic acid (at pH 4.0) was found to cause a latency period identical to the latency period induced by a higher concentration of acetic acid (60 mM) (see Additional file 5). Considering the pK a (3.74) and LogP (−0.54) constants for formic acid (at pH 4.0), the higher toxicity of formic acid compared to acetic acid strongly suggests that there are mechanisms underlying formic acid toxicity that are specific to this weak acid. Among these hypothesized mechanisms specific to formic acid (C1) toxicity or tolerance that emerged from this work is a mechanism that has not been observed before for weak acids in general, not even for acetic acid (C2): the hypothesized uptake of formic acid through the high-affinity potassium transporter, Trk1. This unexpected proposed mechanism is supported by a number of evidences gathered during this study. Among them is the increased accumulation of radiolabeled formic acid in cells devoid of TRK1. The involvement of the Trk1 transporter in facilitating formic acid uptake is consistent with the role of genes encoding the positive (SAT4, HAL3 and HAL5) and negative (PPZ1) regulators of Trk1 biological activity, in decreased or increased tolerance to formic acid, respectively. The lack of detectable effect of TRK2 deletion in yeast tolerance to formic acid is also consistent with this hypothesized role of Trk1 as a facilitator of formic acid uptake into the cell given that Trk1 is a low-affinity K+ transporter, whereas Trk1 is a high-affinity K+ transporter. Remarkably, S. cerevisiae TRK-potassium transporters have been described as mediators of currents of different anions, including Cl− ≫ formate > gluconate > acetate ≫ phosphate (order of selectivity for several slightly permeant anions at pH0 = 5.5) [66]. Our results also indicate that when the growth media has saturating concentrations of K+ (20 mM), the deletion of TRK1 can be explored to increase yeast robustness against formic acid stress.
Conclusion
This study provides the first genome-wide identification of determinants and mechanisms of formic acid toxicity and tolerance in yeast. Among the relevant insights obtained, the role of the Haa1 transcription factor and the Haa1-regulon in the adaptive response and tolerance to formic acid and the biological activity of Trk1 and of its positive regulators in increasing yeast susceptibility to formic acid are highlighted. Collectively, the indications obtained are considered useful to guide the design of fermentation media (increasing K+ concentration) and the genetic manipulation of yeast cell to obtain more robust strains (engineering the Haa1 regulon) for second-generation bio-ethanol production in lignocellulosic biorefineries. The deletion of the TRK1 gene and of genes encoding Trk1 positive regulators is also suggested for maximal yeast tolerance to formic acid at concentrations of K+ for which ectopic uptake of K+ is sufficient to fulfil cellular requirements.
Additional files
Additional file 1: Table S1.List of genes whose expression increases S. cerevisiae tolerance to formic acid based on the screening of the EUROSCARF deletion mutant collection (the elimination of the indicated genes increases yeast susceptibility to formic acid). The severity of growth inhibition is indicated by “+” or “++” and the level was attributed according to the criteria described in Additional file 3.
Additional file 2: Table S2. List of genes whose expression increases S. cerevisiae susceptibility to formic acid based on the screening of the EUROSCARF deletion mutant collection (the elimination of the indicated genes increases yeast tolerance to formic acid).
Additional file 3: Table S3. Primers used in this study.
Additional file 4: Figure S1. Description of the criteria used to define the different levels of susceptibility to formic acid of the deletion mutant strains. Wild-type and deletion mutant strains were spotted onto solid MMB medium (pH 4.5) supplemented with increasing concentrations of formic acid (60, 70 and 80 mM). Strains that did not grow in the presence of 70 mM of formic acid were considered susceptible strains (S); two levels of susceptibility where defined when mutant growth was reduced (+), or completely abolished (++), when cultivated in the presence of 60 mM of formic acid. Strains that grew in medium supplemented with 80 mM of formic acid were considered resistant strains (R).
Additional file 5: Table S4. Comparison of lag-phase duration of the parental strain BY4741 cells cultivated in the presence of equitoxic concentrations of short-chain monocarboxylic acids that cause a latency phase of approximately 17 h. The duration of the lag-phases was determined in cultures of non-adapted BY4741 cells cultivated in MM4 medium (pH 4.0) after sudden exposure to equitoxic concentrations of the listed weak acids (CT, in bold). [HA] concentration of the weak acid protonated at pH 4.0; LogP, logarithm of the partition coefficient of the weak acid between octanol and water.
Acknowledgments
Authors’ contributions
SFH, NPM and ISC conceived and designed the experiments. SFH carried out the experimental work, prepared the figures and contributed to the writing of the manuscript under the scientific supervision of ISC, who coordinated the study. All authors read and approved the final manuscript.
Acknowledgements
Not applicable
Competing interests
The authors declare that they have no competing interests.
Funding
This work was financially supported by “Fundação para a Ciência e a Tecnologia” (FCT) (Project contract PTDC/BBB-BEP/0385/2014 and the PhD fellowship SFRH/BD/78058/2011 to SFH). The financial support received by iBB—Institute for Bioengineering and Biosciences from FCT (UID/BIO/04565/2013) and Programa Operacional Regional de Lisboa 2020 (No. 007317) is acknowledged.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ROS
Reactive oxygen species
- FDH
Formate dehydrogenase
- SGD
Saccharomyces genome database
- KEGG
Kyoto encyclopedia for genes and genomes
- GO
Gene ontology
- MDR
MultiDrug resistance
- MAP
Mitogen-activated protein
- LogP
Coefficient of octanol–water partition
- TORC1
Tor complex 1
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s13068-017-0781-5) contains supplementary material, which is available to authorized users.
Contributor Information
Sílvia F. Henriques, Email: silvia.henriques@tecnico.ulisboa.pt
Nuno P. Mira, Email: nuno.mira@tecnico.ulisboa.pt
Isabel Sá-Correia, Phone: +351-218417682, Email: isacorreia@tecnico.ulisboa.pt.
References
- 1.Sangster J. Octanol–water partition-coefficients of simple organic-compounds. J Phys Chem Ref Data. 1989;18(3):1111–1229. doi: 10.1063/1.555833. [DOI] [Google Scholar]
- 2.Reutemann W, Kieczka H. Formic acid. In: Ullmann’s encyclopedia of industrial chemistry. Wiley-VCH Verlag GmbH & Co. KGaA; 2000.
- 3.Rasmussen H, Sorensen HR, Meyer AS. Formation of degradation compounds from lignocellulosic biomass in the biorefinery: sugar reaction mechanisms. Carbohydr Res. 2014;385:45–57. doi: 10.1016/j.carres.2013.08.029. [DOI] [PubMed] [Google Scholar]
- 4.Greetham D, Wimalasena T, Kerruish DW, Brindley S, Ibbett RN, Linforth RL, Tucker G, Phister TG, Smart KA. Development of a phenotypic assay for characterisation of ethanologenic yeast strain sensitivity to inhibitors released from lignocellulosic feedstocks. J Ind Microbiol Biotechnol. 2014;41(6):931–945. doi: 10.1007/s10295-014-1431-6. [DOI] [PubMed] [Google Scholar]
- 5.Tomas-Pejo E, Oliva JM, Ballesteros M, Olsson L. Comparison of SHF and SSF processes from steam-exploded wheat straw for ethanol production by xylose-fermenting and robust glucose-fermenting Saccharomyces cerevisiae strains. Biotechnol Bioeng. 2008;100(6):1122–1131. doi: 10.1002/bit.21849. [DOI] [PubMed] [Google Scholar]
- 6.Jonsson LJ, Martin C. Pretreatment of lignocellulose: formation of inhibitory by-products and strategies for minimizing their effects. Bioresour Technol. 2016;199:103–112. doi: 10.1016/j.biortech.2015.10.009. [DOI] [PubMed] [Google Scholar]
- 7.dos Santos SC, Sa-Correia I. Yeast toxicogenomics: lessons from a eukaryotic cell model and cell factory. Curr Opin Biotechnol. 2015;33:183–191. doi: 10.1016/j.copbio.2015.03.001. [DOI] [PubMed] [Google Scholar]
- 8.Mira NP, Teixeira MC, Sa-Correia I. Adaptive response and tolerance to weak acids in Saccharomyces cerevisiae: a genome-wide view. OMICS. 2010;14(5):525–540. doi: 10.1089/omi.2010.0072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Teixeira MC, Mira NP, Sa-Correia I. A genome-wide perspective on the response and tolerance to food-relevant stresses in Saccharomyces cerevisiae. Curr Opin Biotechnol. 2011;22(2):150–156. doi: 10.1016/j.copbio.2010.10.011. [DOI] [PubMed] [Google Scholar]
- 10.de Jong B, Siewers V, Nielsen J. Systems biology of yeast: enabling technology for development of cell factories for production of advanced biofuels. Curr Opin Biotechnol. 2012;23(4):624–630. doi: 10.1016/j.copbio.2011.11.021. [DOI] [PubMed] [Google Scholar]
- 11.Chen Y, Sheng J, Jiang T, Stevens J, Feng X, Wei N. Transcriptional profiling reveals molecular basis and novel genetic targets for improved resistance to multiple fermentation inhibitors in Saccharomyces cerevisiae. Biotechnol Biofuels. 2016;9:9. doi: 10.1186/s13068-015-0418-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Caspeta L, Chen Y, Ghiaci P, Feizi A, Buskov S, Hallstrom BM, Petranovic D, Nielsen J. Altered sterol composition renders yeast thermotolerant. Biofuels. 2014;346(6205):75–78. doi: 10.1126/science.1258137. [DOI] [PubMed] [Google Scholar]
- 13.Li M, Borodina I. Application of synthetic biology for production of chemicals in yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2015;15(1):1–12. doi: 10.1093/femsyr/fou003. [DOI] [PubMed] [Google Scholar]
- 14.Lindahl L, Santos AX, Olsson H, Olsson L, Bettiga M. Membrane engineering of S. cerevisiae targeting sphingolipid metabolism. Sci Rep. 2017;7:41868. doi: 10.1038/srep41868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mira NP, Palma M, Guerreiro JF, Sa-Correia I. Genome-wide identification of Saccharomyces cerevisiae genes required for tolerance to acetic acid. Microb Cell Fact. 2010;9:79. doi: 10.1186/1475-2859-9-79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mira NP, Lourenco AB, Fernandes AR, Becker JD, Sa-Correia I. The RIM101 pathway has a role in Saccharomyces cerevisiae adaptive response and resistance to propionic acid and other weak acids. FEMS Yeast Res. 2009;9(2):202–216. doi: 10.1111/j.1567-1364.2008.00473.x. [DOI] [PubMed] [Google Scholar]
- 17.Hasunuma T, Sanda T, Yamada R, Yoshimura K, Ishii J, Kondo A. Metabolic pathway engineering based on metabolomics confers acetic and formic acid tolerance to a recombinant xylose-fermenting strain of Saccharomyces cerevisiae. Microb Cell Fact. 2011;10(1):2. doi: 10.1186/1475-2859-10-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Abbott DA, Suir E, van Maris AJ, Pronk JT. Physiological and transcriptional responses to high concentrations of lactic acid in anaerobic chemostat cultures of Saccharomyces cerevisiae. Appl Environ Microbiol. 2008;74(18):5759–5768. doi: 10.1128/AEM.01030-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chen Y, Stabryla L, Wei N. Improved acetic acid resistance in Saccharomyces cerevisiae by overexpression of the WHI2 gene identified through inverse metabolic engineering. Appl Environ Microbiol. 2016;82(7):2156–2166. doi: 10.1128/AEM.03718-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lee SE, Park BS, Yoon JJ. Proteomic evaluation of cellular responses of Saccharomyces cerevisiae to formic acid stress. Mycobiology. 2010;38(4):302–309. doi: 10.4489/MYCO.2010.38.4.302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Du L, Su Y, Sun D, Zhu W, Wang J, Zhuang X, Zhou S, Lu Y. Formic acid induces Yca1p-independent apoptosis-like cell death in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2008;8(4):531–539. doi: 10.1111/j.1567-1364.2008.00375.x. [DOI] [PubMed] [Google Scholar]
- 22.Piper PW. Yeast superoxide dismutase mutants reveal a pro-oxidant action of weak organic acid food preservatives. Free Radic Biol Med. 1999;27(11–12):1219–1227. doi: 10.1016/S0891-5849(99)00147-1. [DOI] [PubMed] [Google Scholar]
- 23.Carmelo V, Santos H, Sa-Correia I. Effect of extracellular acidification on the activity of plasma membrane ATPase and on the cytosolic and vacuolar pH of Saccharomyces cerevisiae. Biochim Biophys Acta. 1997;1325(1):63–70. doi: 10.1016/S0005-2736(96)00245-3. [DOI] [PubMed] [Google Scholar]
- 24.Makrantoni V, Dennison P, Stark MJ, Coote PJ. A novel role for the yeast protein kinase Dbf2p in vacuolar H+-ATPase function and sorbic acid stress tolerance. Microbiology. 2007;153(12):4016–4026. doi: 10.1099/mic.0.2007/010298-0. [DOI] [PubMed] [Google Scholar]
- 25.Guerreiro JF, Muir A, Ramachandran S, Thorner J, Sa-Correia I. Sphingolipid biosynthesis upregulation by TOR complex 2-Ypk1 signaling during yeast adaptive response to acetic acid stress. Biochem J. 2016;473(23):4311–4325. doi: 10.1042/BCJ20160565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Almeida B, Ohlmeier S, Almeida AJ, Madeo F, Leao C, Rodrigues F, Ludovico P. Yeast protein expression profile during acetic acid-induced apoptosis indicates causal involvement of the TOR pathway. Proteomics. 2009;9(3):720–732. doi: 10.1002/pmic.200700816. [DOI] [PubMed] [Google Scholar]
- 27.Fernandes AR, Mira NP, Vargas RC, Canelhas I, Sa-Correia I. Saccharomyces cerevisiae adaptation to weak acids involves the transcription factor Haa1p and Haa1p-regulated genes. Biochem Biophys Res Commun. 2005;337(1):95–103. doi: 10.1016/j.bbrc.2005.09.010. [DOI] [PubMed] [Google Scholar]
- 28.Mira NP, Henriques SF, Keller G, Teixeira MC, Matos RG, Arraiano CM, Winge DR, Sa-Correia I. Identification of a DNA-binding site for the transcription factor Haa1, required for Saccharomyces cerevisiae response to acetic acid stress. Nucleic Acids Res. 2011;39(16):6896–6907. doi: 10.1093/nar/gkr228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Inaba T, Watanabe D, Yoshiyama Y, Tanaka K, Ogawa J, Takagi H, Shimoi H, Shima J. An organic acid-tolerant HAA1-overexpression mutant of an industrial bioethanol strain of Saccharomyces cerevisiae and its application to the production of bioethanol from sugarcane molasses. AMB Express. 2013;3(1):74. doi: 10.1186/2191-0855-3-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sakihama Y, Hasunuma T, Kondo A. Improved ethanol production from xylose in the presence of acetic acid by the overexpression of the HAA1 gene in Saccharomyces cerevisiae. J Biosci Bioeng. 2015;119(3):297–302. doi: 10.1016/j.jbiosc.2014.09.004. [DOI] [PubMed] [Google Scholar]
- 31.Tanaka K, Ishii Y, Ogawa J, Shima J. Enhancement of acetic acid tolerance in Saccharomyces cerevisiae by overexpression of the HAA1 gene, encoding a transcriptional activator. Appl Environ Microbiol. 2012;78(22):8161–8163. doi: 10.1128/AEM.02356-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kondo A, Hasunuma T, Sakihama Y. Method For Producing Ethanol From Biomass. 2015. US20150218592.
- 33.Swinnen S, Henriques SF, Shrestha R, Ho PW, Sa-Correia I, Nevoigt E. Improvement of yeast tolerance to acetic acid through Haa1 transcription factor engineering: towards the underlying mechanisms. Microb Cell Fact. 2017;16(1):7. doi: 10.1186/s12934-016-0621-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zahn K, Jacobson S. Acetate resistance in yeast based on introduction of a mutant haa1 allele. 2014. WO/2014/018450.
- 35.Babel W, Brinkmann U, Muller RH. The auxiliary substrate concept—an approach for overcoming limits of microbial performances. Acta Biotechnol. 1993;13(3):211–242. doi: 10.1002/abio.370130302. [DOI] [Google Scholar]
- 36.Overkamp KM, Kotter P, van der Hoek R, Schoondermark-Stolk S, Luttik MA, van Dijken JP, Pronk JT. Functional analysis of structural genes for NAD(+)-dependent formate dehydrogenase in Saccharomyces cerevisiae. Yeast. 2002;19(6):509–520. doi: 10.1002/yea.856. [DOI] [PubMed] [Google Scholar]
- 37.Hazeu W, Donker RA. A continuous culture study of methanol and formate utilization by the yeast Pichia pastoris. Biotechnol Lett. 1983;5(6):399–404. doi: 10.1007/BF00131280. [DOI] [Google Scholar]
- 38.Hasunuma T, Sung KM, Sanda T, Yoshimura K, Matsuda F, Kondo A. Efficient fermentation of xylose to ethanol at high formic acid concentrations by metabolically engineered Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2011;90(3):997–1004. doi: 10.1007/s00253-011-3085-x. [DOI] [PubMed] [Google Scholar]
- 39.Iwaki M, Rich PR. Direct detection of formate ligation in cytochrome c oxidase by ATR-FTIR spectroscopy. J Am Chem Soc. 2004;126(8):2386–2389. doi: 10.1021/ja039320j. [DOI] [PubMed] [Google Scholar]
- 40.Haritos VS, Dojchinov G. Cytochrome c oxidase inhibition in the rice weevil Sitophilus oryzae (L.) by formate, the toxic metabolite of volatile alkyl formates. Comp Biochem Physiol C: Toxicol Pharmacol. 2003;136(2):135–143. doi: 10.1016/s1532-0456(03)00173-x. [DOI] [PubMed] [Google Scholar]
- 41.Nicholls P. Formate as an inhibitor of cytochrome c oxidase. Biochem Biophys Res Commun. 1975;67(2):610–616. doi: 10.1016/0006-291X(75)90856-6. [DOI] [PubMed] [Google Scholar]
- 42.Chacon E, Acosta D. Mitochondrial regulation of superoxide by Ca2+: an alternate mechanism for the cardiotoxicity of doxorubicin. Toxicol Appl Pharmacol. 1991;107(1):117–128. doi: 10.1016/0041-008X(91)90336-D. [DOI] [PubMed] [Google Scholar]
- 43.Bralet J, Bouvier C, Schreiber L, Boquillon M. Effect of acidosis on lipid peroxidation in brain slices. Brain Res. 1991;539(1):175–177. doi: 10.1016/0006-8993(91)90703-X. [DOI] [PubMed] [Google Scholar]
- 44.Dikalova AE, Kadiiska MB, Mason RP. An in vivo ESR spin-trapping study: free radical generation in rats from formate intoxication—role of the Fenton reaction. Proc Natl Acad Sci USA. 2001;98(24):13549–13553. doi: 10.1073/pnas.251091098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Srinivasan S, Avadhani NG. Cytochrome c oxidase dysfunction in oxidative stress. Free Radic Biol Med. 2012;53(6):1252–1263. doi: 10.1016/j.freeradbiomed.2012.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gaber RF, Styles CA, Fink GR. TRK1 encodes a plasma membrane protein required for high-affinity potassium transport in Saccharomyces cerevisiae. Mol Cell Biol. 1988;8(7):2848–2859. doi: 10.1128/MCB.8.7.2848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mulet JM, Leube MP, Kron SJ, Rios G, Fink GR, Serrano R. A novel mechanism of ion homeostasis and salt tolerance in yeast: the Hal4 and Hal5 protein kinases modulate the Trk1–Trk2 potassium transporter. Mol Cell Biol. 1999;19(5):3328–3337. doi: 10.1128/MCB.19.5.3328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yenush L, Mulet JM, Arino J, Serrano R. The Ppz protein phosphatases are key regulators of K+ and pH homeostasis: implications for salt tolerance, cell wall integrity and cell cycle progression. EMBO J. 2002;21(5):920–929. doi: 10.1093/emboj/21.5.920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yenush L, Merchan S, Holmes J, Serrano R. pH-Responsive, posttranslational regulation of the Trk1 potassium transporter by the type 1-related Ppz1 phosphatase. Mol Cell Biol. 2005;25(19):8683–8692. doi: 10.1128/MCB.25.19.8683-8692.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kohrer K, Domdey H. Preparation of high molecular weight RNA. Methods Enzymol. 1991;194:398–405. doi: 10.1016/0076-6879(91)94030-G. [DOI] [PubMed] [Google Scholar]
- 51.Ferrando A, Kron SJ, Rios G, Fink GR, Serrano R. Regulation of cation transport in Saccharomyces cerevisiae by the salt tolerance gene HAL3. Mol Cell Biol. 1995;15(10):5470–5481. doi: 10.1128/MCB.15.10.5470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Madrid R, Gomez MJ, Ramos J, Rodriguez-Navarro A. Ectopic potassium uptake in trk1 trk2 mutants of Saccharomyces cerevisiae correlates with a highly hyperpolarized membrane potential. J Biol Chem. 1998;273(24):14838–14844. doi: 10.1074/jbc.273.24.14838. [DOI] [PubMed] [Google Scholar]
- 53.Hillenmeyer ME, Fung E, Wildenhain J, Pierce SE, Hoon S, Lee W, Proctor M, St Onge RP, Tyers M, Koller D, et al. The chemical genomic portrait of yeast: uncovering a phenotype for all genes. Science. 2008;320(5874):362–365. doi: 10.1126/science.1150021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Duskova M, Borovikova D, Herynkova P, Rapoport A, Sychrova H. The role of glycerol transporters in yeast cells in various physiological and stress conditions. FEMS Microbiol Lett. 2015;362(3):1–8. doi: 10.1093/femsle/fnu041. [DOI] [PubMed] [Google Scholar]
- 55.Dumont ME, Cardillo TS, Hayes MK, Sherman F. Role of cytochrome c heme lyase in mitochondrial import and accumulation of cytochrome c in Saccharomyces cerevisiae. Mol Cell Biol. 1991;11(11):5487–5496. doi: 10.1128/MCB.11.11.5487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Barros MH, Netto LE, Kowaltowski AJ. H(2)O(2) generation in Saccharomyces cerevisiae respiratory pet mutants: effect of cytochrome c. Free Radic Biol Med. 2003;35(2):179–188. doi: 10.1016/S0891-5849(03)00307-1. [DOI] [PubMed] [Google Scholar]
- 57.Tamas MJ, Luyten K, Sutherland FC, Hernandez A, Albertyn J, Valadi H, Li H, Prior BA, Kilian SG, Ramos J, et al. Fps1p controls the accumulation and release of the compatible solute glycerol in yeast osmoregulation. Mol Microbiol. 1999;31(4):1087–1104. doi: 10.1046/j.1365-2958.1999.01248.x. [DOI] [PubMed] [Google Scholar]
- 58.Oshoma CE, Greetham D, Louis EJ, Smart KA, Phister TG, Powell C, Du C. Screening of non-Saccharomyces cerevisiae strains for tolerance to formic acid in bioethanol fermentation. PLoS One. 2015;10(8):e0135626. doi: 10.1371/journal.pone.0135626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kodedova M, Sychrova H. Changes in the sterol composition of the plasma membrane affect membrane potential, salt tolerance and the activity of multidrug resistance pumps in Saccharomyces cerevisiae. PLoS One. 2015;10(9):e0139306. doi: 10.1371/journal.pone.0139306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lindberg L, Santos AX, Riezman H, Olsson L, Bettiga M. Lipidomic profiling of Saccharomyces cerevisiae and Zygosaccharomyces bailii reveals critical changes in lipid composition in response to acetic acid stress. PLoS One. 2013;8(9):e73936. doi: 10.1371/journal.pone.0073936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Daum G, Lees ND, Bard M, Dickson R. Biochemistry, cell biology and molecular biology of lipids of Saccharomyces cerevisiae. Yeast. 1998;14(16):1471–1510. doi: 10.1002/(SICI)1097-0061(199812)14:16<1471::AID-YEA353>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 62.Bauer BE, Rossington D, Mollapour M, Mamnun Y, Kuchler K, Piper PW. Weak organic acid stress inhibits aromatic amino acid uptake by yeast, causing a strong influence of amino acid auxotrophies on the phenotypes of membrane transporter mutants. Eur J Biochem. 2003;270(15):3189–3195. doi: 10.1046/j.1432-1033.2003.03701.x. [DOI] [PubMed] [Google Scholar]
- 63.Hueso G, Aparicio-Sanchis R, Montesinos C, Lorenz S, Murguia JR, Serrano R. A novel role for protein kinase Gcn2 in yeast tolerance to intracellular acid stress. Biochem J. 2012;441(1):255–264. doi: 10.1042/BJ20111264. [DOI] [PubMed] [Google Scholar]
- 64.Binda M, Peli-Gulli MP, Bonfils G, Panchaud N, Urban J, Sturgill TW, Loewith R, De Virgilio C. The Vam6 GEF controls TORC1 by activating the EGO complex. Mol Cell. 2009;35(5):563–573. doi: 10.1016/j.molcel.2009.06.033. [DOI] [PubMed] [Google Scholar]
- 65.Loewith R, Hall MN. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics. 2011;189(4):1177–1201. doi: 10.1534/genetics.111.133363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rivetta A, Kuroda T, Slayman C. Anion currents in yeast K+ transporters (TRK) characterize a structural homologue of ligand-gated ion channels. Pflugers Arch. 2011;462(2):315–330. doi: 10.1007/s00424-011-0959-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1.List of genes whose expression increases S. cerevisiae tolerance to formic acid based on the screening of the EUROSCARF deletion mutant collection (the elimination of the indicated genes increases yeast susceptibility to formic acid). The severity of growth inhibition is indicated by “+” or “++” and the level was attributed according to the criteria described in Additional file 3.
Additional file 2: Table S2. List of genes whose expression increases S. cerevisiae susceptibility to formic acid based on the screening of the EUROSCARF deletion mutant collection (the elimination of the indicated genes increases yeast tolerance to formic acid).
Additional file 3: Table S3. Primers used in this study.
Additional file 4: Figure S1. Description of the criteria used to define the different levels of susceptibility to formic acid of the deletion mutant strains. Wild-type and deletion mutant strains were spotted onto solid MMB medium (pH 4.5) supplemented with increasing concentrations of formic acid (60, 70 and 80 mM). Strains that did not grow in the presence of 70 mM of formic acid were considered susceptible strains (S); two levels of susceptibility where defined when mutant growth was reduced (+), or completely abolished (++), when cultivated in the presence of 60 mM of formic acid. Strains that grew in medium supplemented with 80 mM of formic acid were considered resistant strains (R).
Additional file 5: Table S4. Comparison of lag-phase duration of the parental strain BY4741 cells cultivated in the presence of equitoxic concentrations of short-chain monocarboxylic acids that cause a latency phase of approximately 17 h. The duration of the lag-phases was determined in cultures of non-adapted BY4741 cells cultivated in MM4 medium (pH 4.0) after sudden exposure to equitoxic concentrations of the listed weak acids (CT, in bold). [HA] concentration of the weak acid protonated at pH 4.0; LogP, logarithm of the partition coefficient of the weak acid between octanol and water.