

Abstract
Three studies examined the hypothesis that bilinguals can more rapidly disengage attention from irrelevant information than monolinguals by investigating the impact of previous trial congruency on performance in a simple flanker task. In Study 1, monolingual and bilingual young adults completed two versions of a flanker task. There were no differences between language groups on mean reaction time using standard analyses for congruent or incongruent trials or the size of the flanker effect. Sequential congruency effects (SCEs) however, which account for previous trial congruency, were smaller for bilinguals than for monolinguals. This finding was strongest at the shortest response-to-stimulus interval (RSI). Study 2 replicated this effect using a slightly different flanker task and a shorter RSI than study 1. Study 3 showed that at long RSIs, where behavioral SCE differences between groups disappear because of sufficient time to recover from the previous trial, event-related potentials demonstrated a continued influence of previous trial congruency for monolinguals but not bilinguals at both the N2 and the P3, replicating the reaction time effects in Studies 1 and 2. Together, these studies demonstrate that bilinguals experience less influence from previous trial congruency and have greater ability to disengage attention from the previous trial in order to focus attention on the current trial than is found for monolinguals.
Keywords: Conflict adaptation, Sequential congruency effects, Bilingualism, Disengagement, Executive control, Flanker task
1. Introduction
It is well established that the two languages of bilinguals are constantly active to some degree, creating a situation in which bilinguals must continually manage attention to the target language to avoid interference from the other language (review in Kroll Dussias, Bogulski, & Valdes Kroff, 2012). The suggestion has been that this linguistic conflict recruits the domain-general executive control system, thereby enhancing executive control for other tasks, including nonverbal ones (reviews in Bialystok, in press; Bialystok, Craik, Green, & Gollan, 2009). Neuroimaging support for this position comes from studies showing overlapping brain networks for language control and cognitive control in bilinguals (De Baene, Duyck, Brass, & Carreiras, 2015; Luk, Green, Abutalebi, & Grady, 2012). Behavioral evidence for the enhancement of domain-general executive control in bilinguals has accrued for infants (Kovács & Mehler, 2009; Pons, Bosch, & Lewkowicz, 2015; Singh et al., 2015), children (meta-analysis in Adesope et al., 2010; review in Barac, Bialystok, Castro, & Sanchez, 2014), and older adults (Bialystok, Craik, Klein, & Viswanathan, 2004; Gold et al., 2013). However, studies with young adults often show no language group differences in performing these tasks (Bialystok, 2006; Bialystok, Martin & Viswanathan, 2005; Paap & Greenberg, 2013; Gathercole et al., 2014; Kalia, Wilbourne, & Ghio, 2014), leading to a debate about the validity of these effects (Paap, Johnson, & Sawi, 2015). Logically, it would be surprising that a processing effect found in childhood and older age disappeared in young adulthood. The present study addresses the possibility that the standard statistical approach used in this literature lacks the sensitivity required to detect the processing differences that discriminate between groups of young adults performing these tasks. Evidence for this hypothesis will contribute to both a more detailed understanding of executive control and the controversy surrounding the cognitive effects of bilingualism.
Several factors can lead to null results when investigating group differences in performance between monolinguals and bilinguals on simple conflict tasks. First, the distinction between monolinguals and bilinguals may not be clearly demarcated, blurring the difference between groups (Bialystok, 2016; Luk & Bialystok, 2013). Second, behavioral responses may lack the variance needed for reliable group differences to emerge in high-performing young adults (Marian, Chabal, Bartolotti, Bradley, & Hernandez, 2014), particularly on simple tasks where performance is effectively at ceiling. The average reaction time for the tasks used in much of this research is about 500 ms, so group differences need to be large for statistically significant differences to emerge. In contrast, the average reaction time for children or older adults performing such tasks is often twice as long, allowing more room for experience to push performance in a particular direction. Bilingual young adults outperform monolinguals when task demands are increased (Bialystok, 2006; Friesen, Latman, Calvo, & Bialystok, 2015). For example, Costa et al. (2009) showed that when the number of congruent and incongruent trials was equivalent, bilinguals were faster than monolinguals, despite equivalent accuracy. In this condition, there was always a possibility that the next trial involved conflict, making responses less predictable. However, when the majority of the trials was either congruent or incongruent so the next trial was more predictable, monolinguals and bilinguals performed equivalently. Their interpretation was that in situations requiring higher levels of monitoring, bilinguals are better able than monolinguals to efficiently resolve conflict.
Third, task domain is important because language tasks are typically more effortful for bilinguals. Bilinguals identify pictures more slowly (Gollan et al., 2007; Ivanova & Costa, 2008), generate fewer words in verbal fluency tasks (Bialystok, Craik & Luk, 2008) and have smaller receptive vocabularies (Bialystok & Luk, 2012) than their monolingual peers. Because bilinguals divide their time between two languages, they have less experience in each language than monolinguals (Gollan, Montoya, Cera, & Sandoval, 2008; Gollan, Montoya, Fennema-Notestine, & Morris, 2005; Gollan, Sandoval, & Salmon, 2011). Thus, verbal tasks often disadvantage bilinguals and may mask group differences in executive control.
One way of detecting processing differences that may be concealed by analyses of overall RT is to examine the influence of the previous trial on performance. The ability to disengage from previous information is central to executive control (Elsabbagh et al., 2013; Landry & Bryson, 2004) but is not considered in analyses that compare overall RT on congruent (C) and incongruent (I) trials, or the difference between them (I-C; congruency effect). For example, in the flanker task in which one responds to a central target among flanking distractors, the congruency effect is the difference between I (< < > < <) and C trials (< < < < <). Studies examining differences between monolinguals and bilinguals have largely ignored the influence of the previous trial. Yet with practice, individuals not only become faster, but also rely less on previous trial congruency to achieve this speed (Mayr & Awe, 2009; van Steenbergen, 2015). The hypothesis is that young adult bilinguals will be better able to disengage attention from the previous trial in responding to the current trial, signaling better executive control in the absence of overall differences in RT.
The notion that bilinguals might be better at disengaging attention fits well with existing evidence. A prevailing view of how bilinguals manage attention to the target language in the face of joint activation is that the non-target language is inhibited (Green, 1998), a view consistent with the executive function model proposed by Mikaye et al. (2000) that assigns a prominent role to inhibition. One problem with the inhibition view, however, is the finding that bilingual preverbal infants with no practice in using language also outperform their monolingual peers on executive function tasks (Kovács & Mehler, 2009; Pons, Bosch, & Lewkowicz, 2015; Singh et al., 2015). Infants in the first year of life have representations for both languages (Weikum et al., 2007) and can attend to each selectively. Therefore, bilingual infants have more experience than monolingual infants in controlling attention to one of two language representations. This experience of bilingual infants that requires them to pay attention to multiple sources of input within various linguistic contexts makes it adaptive for them to rapidly disengage attention from stimuli once they are processed so that attention can be re-engaged to currently relevant stimuli. Such early experience in attending to and disengaging from linguistic cues may shape these attentional processes in later development.
Studies with young adults that show faster performance on a flanker task by bilinguals than monolinguals generally find the difference in both congruent and incongruent trials (Costa et al., 2008, 2009; Emmorey et al., 2008; see Hilchey & Klein, 2011 for a review). This pattern, too, is contrary to the prediction from the inhibition view in which language group differences would only be expected on I trials. The mechanism responsible for the effects of bilingualism, therefore, needs to affect both C and I trials. Disengagement of attention might be such a mechanism given that some amount of disengagement is required on all trial types to avoid devoting all resources to elaborative processing of the no longer relevant (previous) stimulus.
The sequential congruency effect (SCE)1 is the index of online reactive adjustment in performance in response to the congruency of the previous trial (Gratton, Coles, & Donchin, 1992) and has been shown to be important for understanding the role of executive control in interference tasks (Botvinick et al., 2001; Wang et al., 2015; Weissman et al., 2015; review in Egner, 2014). In this sense, the SCE reflects the speed of disengagement of attention. Accounting for previous trial congruency produces four trial types: congruent trials in which the previous trial was congruent (cC), congruent trials in which the previous trial was incongruent (iC), incongruent trials in which the previous trial was incongruent (iI), and incongruent trials in which the previous trial was congruent (cI). RT is facilitated when trial type is repeated (iI and cC) and slowed when trial type is changed (cI and iC). This leads to two flanker effects: I-C difference following congruent trials (c-flanker effect) and I-C difference following incongruent trials (i-flanker effect). The difference between these two flanker effects is the SCE (c-flanker – i-flanker). Larger SCEs indicate greater influence of previous trials on performance for both congruent and incongruent trials, and thus, slower disengagement of attention from those trials. Generally, the i-flanker effect is smaller than the c-flanker effect (see Figure 1).
Figure 1.
Depiction of the sequential congruency effect showing facilitation of RT with repeated trial type and slower RT with changed trial type. c-flanker effect is the congruency effect following congruent stimuli (cI – cC) and i-flanker effect is the congruency effect following incongruent stimuli (iI – iC). The sequential congruency effect (SCE) is the difference between these and represents the influence of previous trial congruency on the flanker effect.
Interpretations of SCEs include cognitive accounts based on top-down control (Egner, Ely, & Grinband, 2010) and associative accounts based on bottom-up retrieval (Mayr, Awh, & Laurey, 2003; Schmidt, 2013), both of which require some amount of disengagement before processing the current trial. One cognitive account, the conflict monitoring theory (Botvinick et al. 2001; Kerns, 2004) posits that the anterior cingulate cortex (ACC) detects conflict on incongruent trials and signals the dorsolateral prefrontal cortex to focus on task-relevant features on subsequent trials. Thus, incongruent trials facilitate performance on subsequent trials by refocusing attention to task-relevant features, making later incongruent trials less interfering. In contrast, encountering a congruent trial broadens the focus of attention for subsequent trials and leads to facilitation on subsequent congruent trials but interference on subsequent incongruent trials because distractors are also incorporated in the broadening of attention. In the repetition expectancy account (Gratton et al., 1992), the detection of congruency biases the individual to expect the same type of stimulus on the next trial. Thus, cognitive accounts assume that SCEs are the result of top-down attentional control applied from trial to trial. Note that in both of these accounts, the previous stimulus biases processing of the current stimulus, but some form of disengagement is required in order to avoid devoting all resources to elaborative processing of the no longer relevant stimulus. This interpretation is supported by EEG studies that show an initial increase in cortical activity immediately following a trial (reflecting adjustments in control), followed by a decrease in activity (reflecting disengagement of resources), and finally an increase in cortical activity immediately preceding the next stimulus (reflecting engagement; Compton et al., 2012).
Associative accounts such as feature-integration (Hommel, Proctor, & Vu, 2004) explain SCEs in terms of retrieval of partial or complete overlap of stimulus and response features from trial to trial. On any given trial, stimulus and response features are bound together as an “event file” and stored as a memory representation, and when those features are encountered on subsequent trials, they act as cues for past event files. Interference or facilitation occurs depending on the degree of overlap between the current event-file and the retrieved event-file(s). For example, in the flanker task, if a trial includes left pointing flankers (< < - < <) and a right pointing central target (- - > - -), and require a right button press, these features (i.e., left flanking arrows, right central target, right button press) are bound together as one event file. Subsequently encountering another right-pointing central arrow with right flanking arrows that requires a right button press cues retrieval of the prior event file. To the extent that the current and previous event files share all features, the current trial is facilitated. However, if there is only partial overlap as in the example of the right-pointing central arrow with right flanking arrows, the retrieved file needs to be “unbound” (i.e., disengaged) to execute the correct response (Egner, 2014). Thus, performance depends not on the value of the current trial as C or I but rather on the overlap between consecutive trials; that is, iI and cC trials are complete overlap trials and result in facilitation and iC and cI are partial overlap trials and result in interference. An intermediate position proposed by Egner (2014) argues that bottom-up associative and top-down cognitive accounts are not mutually exclusive but rather differ in the level of abstraction of the mechanism. He argues that stimulus and response features are bound together on a trial-by-trial basis like the associative accounts suggest, but that more complex top-down control states are also bound in time to these event files. Thus, both top-down and bottom-up processes contribute to the size of the SCE when event-files are retrieved from memory.
What is common to these views is their reliance on disengagement of attention from previous trial information. In cognitive accounts, disengagement from the previous trial precedes the implementation of control to complete the current trial; otherwise one would be devoting all resources to elaborative processing of the no longer relevant trial. In associative and Egner’s multi-level perspectives, the previous event file must be “unbound” (i.e., disengaged) in order to respond to the current stimulus (Egner, 2014). Therefore, disengagement of attention from the previous stimulus from a cognitive perspective occurs prior to presentation of the current stimulus, whereas disengagement of attention from the previous stimulus from an associative perspective occurs while the current stimulus is presented. In both accounts, degree and speed of disengagement from the previous trial contribute to the size of the SCE, with faster disengagement leading to smaller SCEs, a point supported in recent EEG studies showing disengagement between trials revealed by alpha power analyses (Compton et al., 2011, 2012).
The ACC is believed to be responsible for SCE adjustments (Botvinick et al., 2001). The most recent proposals for the ACC that incorporate both bottom-up and top-down perspectives in relation to SCE effects include the idea that the ACC is involved in predictions of upcoming cognitive load (Alexander & Brown, 2011; Holroyd, & Yeung, 2012; Shenhav, Botvinick, & Cohen, 2013; Shenhav, Cohen, & Botvinick, 2016; Sheth et al., 2012). These perspectives hinge on the idea that the ACC is responsible for recording and evaluating current and recent control requirements. The ACC then provides predictions of upcoming cognitive load/value or cost of reward, and these predictions are used by other centers to implement appropriate control. For example, Sheth et al. (2012) demonstrated using single-cell recordings that ACC neurons were activated in response to current conflict trials as well as trials following conflict. Furthermore, they showed that lesions in the ACC did not disrupt how people responded to current conflict trials compared to non-conflict trials, but that ACC lesions abolished the history-dependant behavioural adjustments; in other words, ACC lesions eliminated SCE effects. Because ACC activation was seen on both conflict trials and trials following conflict despite divergent behavioural adjustments, the authors proposed that the role of the ACC was to maintain a continuously updated account of predicted cognitive demand, rather than one of conflict detection per se. In a sense, this perspective is an extension of the conflict monitoring theory (Shenhav, Botvinick, & Cohen, 2013). Therefore, if bilinguals disengage attention more rapidly than monolinguals, then smaller predictions of upcoming cognitive load would be recorded by the ACC because control requirements induced by the previous stimulus do not last as long, leading to smaller behavioral SCE effects. This account is consistent with evidence showing less ACC activity during conflict resolution for bilinguals than monolinguals (Abutalebi et al., 2012).
There is some existing evidence that bilinguals disengage from previous distracting information more rapidly than monolinguals (Grundy & Keyvani Chahi, 2017; Mishra, Hilchey, Singh, & Klein, 2012). Task-switching studies that examine the cost of switching from one task to another versus repeating the same task have shown that bilinguals show smaller switch costs than monolinguals (Prior & Gollan, 2011; Prior & MacWhinney, 2010). Rapid disengagement is one of the mechanisms believed to be responsible for reduced switch costs (Allport & Wylie, 2000), although the finding of reduced switch costs for bilinguals is not always replicated (Hernández, Martin, Barceló, & Costa, 2013). Using an inhibition of return paradigm, Mishra and colleagues (2012) showed that high proficiency bilinguals showed the inhibition of return effect at earlier stimulus-onset-asynchronies than low proficiency bilinguals, a pattern consistent with more rapid disengagement from the preceding irrelevant distractor (Klein, 2000). Thus, higher proficiency in a second language is associated with more rapid disengagement.
More direct evidence for the possibility that bilinguals are better able to disengage attention than monolinguals comes from a study by Costa and colleagues (Costa, Hernández, & Sebastián-Gallés, 2008). The researchers examined switching costs for congruent and incongruent trials separately during an attention network flanker task (ANT; Fan et al., 2002) and examined the four critical trial types (cC, cI, iC, iI) involved in the SCE calculation. Congruent switch trials are similar to iC trials, congruent repeat trials are similar to cC trials, incongruent switch trials are similar to cI trials, and incongruent repeat trials are similar to iI trials. The authors found that monolinguals showed larger switch costs than bilinguals for congruent trials (iC – cC) and showed numerically (but not statistically) larger switch costs for monolinguals on incongruent trials (iI – cI). These results provide preliminary evidence for the idea that monolinguals are more influenced by previous trial congruency than bilinguals, at least for current congruent trials. However, several caveats need to be considered. First, the critical interaction needed to justify the breakdown of congruent and incongruent trials by previous trial type was never reported. Second, SCE scores were not calculated; SCE scores incorporate all four trial types into a single index of performance to provide a measure of how previous congruency is affecting current trial congruency. Group differences for switch costs on incongruent trials in Costa et al.’s (2008) study were in the direction of showing less influence of previous trial congruency for bilinguals than monolinguals in that bilinguals showed smaller switch costs, but this was not statistically significant. Incorporating both congruent and incongruent trials in the same index of performance strengthens the ability to find these group differences and is essential for an interpretation of SCEs. Third, the ANT presents cues before the critical flanker trials, which means that disengagement of attention would occur not only for the critical trials but also for the cues. To isolate disengagement from the critical trials rather than to the cues, a simple flanker task is more appropriate. The present studies addressed these issues by using a simple flanker task with no cues prior to stimulus onset, reporting the critical interaction, and calculating the SCE scores for both monolinguals and bilinguals. If bilinguals show smaller SCE scores in this case, the disengagement hypothesis will be supported.
An implication of the disengagement hypothesis is that time between trials will affect ease of disengagement and consequently the size of the SCE. For this reason, smaller SCEs have been associated with longer response-to-stimulus intervals (RSI; Egner, Ely, & Grinband, 2010) where more time is available to disengage from the previous trial. Thus, bilinguals might show smaller SCEs than monolinguals at short, but not long RSIs, where there is adequate time for all participants to disengage attention. However, neural processes should continue to reflect more rapid disengagement of attention for bilinguals. Specifically, the N2 and P3 event-related potential (ERP) components are sensitive to trial modulations involved in SCEs (Clayson & Larson, 2011a, 2011b). The N2 is especially sensitive to conflict (Donkers, & van Boxtel, 2004; Nieuwenhuis et al. 2003) and the P3 to stimulus categorization (Polich, 2012). Monolinguals should show larger and longer lasting modulations of these components than bilinguals because of the previous stimulus, reflecting the longer lasting influence of conflict at the N2 and more resources devoted to stimulus categorization at the P3.
In sum, the present set of studies investigated the hypothesis that bilinguals will show smaller SCEs than monolinguals, even when overall RT is comparable between groups. The smaller SCE reflects more rapid disengagement of attention and greater ability to refocus on the current trial. Although components of executive control, these processes are not captured by overall comparison of mean RT between groups. The purpose is to identify processing differences between groups that may help to explain the larger pattern of group differences found across the lifespan.
2. Study 1
Study 1 presented a flanker task to monolinguals and bilinguals using two RSI conditions to assess the effect of length of time before the next trial on performance. Two versions of the flanker task were included, verbal and nonverbal, to examine the effect of verbal stimuli on performance for bilinguals. The hypotheses were that bilinguals would show smaller SCEs than monolinguals, with larger differences between groups for nonverbal stimuli and for short RSIs.
2.1 Method
2.1.1. Participants and Procedures
Thirty English-speaking monolingual and 34 bilingual young adults participated in exchange for course credit. Five participants were excluded because of failure to understand the instructions or not completing the experiment. The final sample consisted of 28 monolinguals and 31 bilinguals. Twenty-one bilinguals reported English as their first or dominant language (L1). Non-English second languages (L2) included Arabic, Bisaya, Farsi, Gujarati, Italian, Japanese, Korean, Lithuanian, Russian, Somali, Spanish, Tagalog, Tamil, Urdu, and Yoruba. Those who reported a non-English L1 included Arabic, Cantonese, Ilonggo, Portuguese, Punjabi, Russian, Spanish, Twi, and Urdu. All bilinguals were early acquisition bilinguals, with a mean age of learning L2 at 1.7 years (SD = 2.9, range 0–10 years). Participants were assessed for language experience and proficiency on scales from 0 to 100 using the Language and Social Background Questionnaire (LSBQ; Luk & Bialystok, 2013). L2 proficiency in speaking, understanding, reading, and writing were assessed by asking participants to estimate their proficiency in each on this scale from 0 (no proficiency) to 100 (native). Frequency of use of the L2 and the L1 were assessed using a 5-point scale that asked participants the extent to which they used each of the languages to speak and listen throughout the day. These were then converted to relative percentages. Background information is reported in Table 1.
Table 1.
Mean scores (standard deviation) for background measures by language group in Study 1.
| Monolingual | Bilingual | |
|---|---|---|
|
| ||
| N | 28 | 31 |
| Age (years) | 19.1 (1.5) | 19.3 (1.9) |
| Shipley Vocabulary | 102.4 (9.1) | 100.6 (12.9) |
| Shipley Reasoning | 102.8 (11.7) | 103.8 (10.7) |
| SES (parents’ education) | 3.2 (1.1) | 3.3 (1.3) |
| L2 proficiency (%) | ||
| Speaking* | 2.8 (6.3) | 81.8 (16.9) |
| Understanding* | 5.1 (11.7) | 89.2 (12.3) |
| Reading* | 0.6 (2.8) | 54.7 (33.8) |
| Writing* | 0.6 (2.8) | 43.1 (32.7) |
| Frequency of use (%) | ||
| L1* | 95 (10.1) | 57 (11.7) |
| L2* | 5 (10.1) | 43 (11.7) |
| Control trial RTs (ms) | ||
| Nonverbal | 481 (69) | 482 (67) |
| Verbal | 532 (80) | 523 (72) |
Asterisks indicate significant differences between groups.
Participants were also tested on English receptive vocabulary and non-verbal intelligence through the Shipley-2 Institute of Living Scale Verbal and Blocks (Shipley, Gruber, Martin, & Klein, 2009). Shipley measures were converted to standardized scores (μ = 100, SD = 15).
Procedures complied with all relevant policies on ethics and were approved by the University Ethics Board. After obtaining informed consent, participants completed the three background measures followed by the flanker tasks, with order of the flanker tasks counterbalanced across participants. The experiment lasted approximately 50 minutes.
2.1.2 Flanker tasks
Stimuli were presented using E-prime 2.0 software (Psychology Software Tools, Inc., version 2.0.10.353) on a 19-inch computer at a distance of approximately 50 cm from participants. Refresh rate was set to 75 Hz. Stimuli were presented in the center of the screen subtending a vertical visual angle of approximately 1.2° and horizontal visual angle of 6.9°. For the nonverbal task, chevrons (< >) were presented in 18-point Verdana font; for the verbal task, the words “right” and “left” were presented in 14-point Verdana font. Stimuli remained on the screen until participants responded, or 2000 ms had elapsed. The RSI was set to 500 or 1000 ms, randomized within block. Flanker blocks that contained randomly intermixed congruent and incongruent trials were preceded and followed by control blocks consisting of target stimuli surrounded by neutral flankers (- -) as a measure of response speed. Participants were instructed to respond to the central stimulus while ignoring the flankers (i.e., right button press for nonverbal [< < > < <] or verbal [left right left] displays). There were six blocks in total; the two flanker blocks each contained 320 (160 congruent, 160 incongruent) trials and the four control blocks each contained 40 trials.
2.2 Results
Groups were equivalent on verbal and non-verbal intelligence, Fs < 1, and differed only in second-language use, ps < 0.001 (see Table 1). Mean RTs for control trials in both flanker tasks were also equivalent, Fs < 1. Accuracy was at ceiling (> 97% correct) and not analyzed further.
Mean RTs for the nonverbal flanker are shown in Figure 2A. A 2-way group × congruency mixed-measures ANOVA revealed a significant main effect of congruency, F(1, 57) = 302.48, p < 0.0001, ηp2 = 0.84, with no effect of group F< 1, or group by congruency interaction, F(1, 57) = 3.26, p = 0.08, ηp2 = 0.05.
Figure 2.
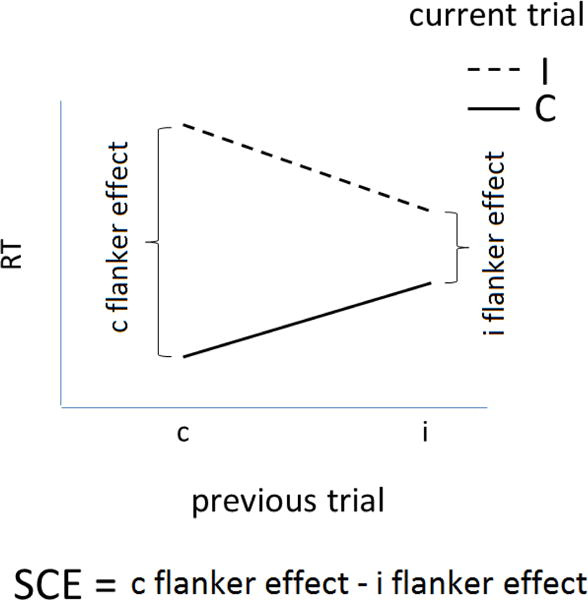
A) Mean RT (standard error) by group for current congruent and incongruent trials on the non-verbal arrow flanker task in Study 1. B) Mean RT as a function of current and previous congruency for each response-to-stimulus interval. C) SCE for each response-to-stimulus interval. * p < 0.05.
Figure 2 also shows mean RT as a function of current and previous trial congruency (Figure 2B) and SCEs (Figure 2C). A 4-way ANOVA indicated a significant effect of RSI, F(1, 57) = 9.89, p = 0.003, ηp2 = 0.15, current trial congruency, F(1, 57) = 308.88, p < 0.001, ηp2 = 0.84, and an interaction of previous trial congruency and current trial congruency, F(1, 57) = 36.10, p < 0.0001, ηp2 = 0.39. This interaction confirms that the flanker effect was larger following congruent (c-flanker effect = 97 ms) than incongruent (i-flanker effect = 61 ms) trials. These effects were qualified by a 3-way interaction between previous trial congruency, current trial congruency, and group, F(1, 57) = 5.74, p = 0.02, ηp2 = 0.09. One-way ANOVAs revealed that the SCE was larger for monolinguals than bilinguals, F(1, 57) = 5.74, p = 0.01, ηp2 = 0.09 (Figure 2C). More detailed analyses examining the interaction effects are reported in the Appendix. No other effects reached significance, Fs < 2.8.
Mean RTs for the verbal task are shown in Figure 3A. Again, the standard analysis revealed a significant main effect of congruency, F(1, 57) = 217.72, p < 0.001, ηp2 = 0.80, with no effect of group or group by congruency interaction, Fs < 1.
Figure 3.
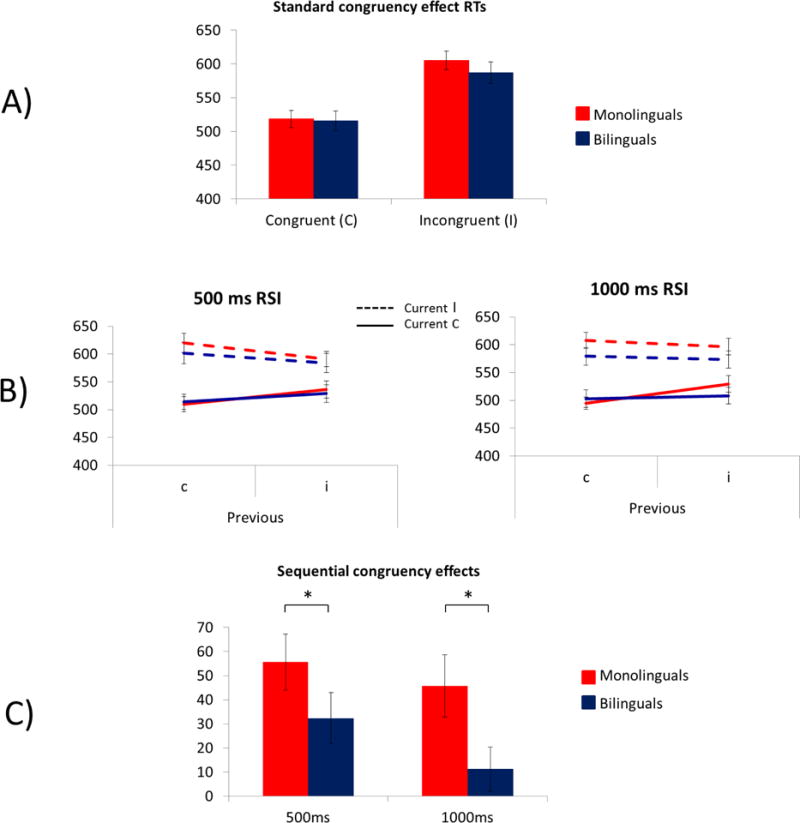
A) Mean RT to current congruent and incongruent trials by language group on the word flanker task in Study 1. B) Mean RT as a function of current and previous congruency for each response-to-stimulus interval. C). SCE for each response-to-stimulus interval. * p < 0.05.
Mean RT as a function of current and previous trial congruency and SCEs are depicted in Figure 3B and 3C, respectively. The 4-way ANOVA revealed a significant main effect of current trial congruency, F(1, 57) = 222.34, p < 0.001, ηp2 = 0.80, a two-way interaction between RSI and current trial congruency, F(1, 57) = 4.10, p = 0.05, ηp2 = 0.07, a two-way interaction between previous trial congruency and current trial congruency, F(1, 57) = 60.52, p < 0.001, ηp2 = 0.52, and a 4-way interaction between previous trial congruency, current trial congruency, RSI, and group, F(1, 57) = 4.41, p = 0.04, ηp2 = 0.07. The interaction between previous trial congruency and current trial congruency is the SCE. The 4-way interaction was explored by conducting a 3-way ANOVA for each RSI. Follow-up one-way ANOVAS showed that bilinguals had smaller SCE scores than monolinguals at the short RSI, F(1, 57) = 4.53, p = 0.04, ηp2 = 0.06, but not at the long RSI, F < 1.
2.3 Discussion
As in some previous research (e.g., Paap & Greenberg, 2013), there were no group differences in mean RT to congruent and incongruent stimuli. However, taking into account previous trial congruency revealed larger SCEs for monolinguals than bilinguals at both RSIs in the nonverbal flanker task and at the short RSI in the verbal flanker task. Thus, simple analyses that focus only on current trial congruency are insensitive to processing differences that exist between monolingual and bilingual young adults. Furthermore, group differences in SCE are more likely to appear in non-verbal tasks than in verbal ones. This difference in task domain might reflect the general finding that bilinguals must manage attention to ongoing competition between languages. As such, they become efficient and fast so that the conflict has less enduring impact on their processing. At long RSIs, monolinguals are able to “catch up” with bilinguals in terms of their ability to disengage attention from the previous stimulus. This leads to larger SCEs for bilinguals than monolinguals only at the long, but not short, RSI.
Study 1 provides initial support for the idea that bilinguals are better able to disengage attention from previous information in that they were less influenced by previous trial congruency. The next two studies attempted to replicate this pattern with unpublished flanker task data from our lab. There were two criteria for selecting these data sets: 1) sufficient numbers of trials (at least 15 observations per trial condition) to calculate the four critical trials required for SCE analyses, and 2) sufficiently large sample size to address power issues.
3. Study 2
The data for Study 2 were obtained from a study in which participants completed a block of trials using a similar version of the nonverbal flanker task as presented in Study 1. The main difference was that the RSI was shorter than that used in Study 1. There were also more participants in this study so power was increased.
3.1 Method
3.1.1. Participants and Procedure
Fifty-three monolinguals and 58 bilinguals participated in exchange for course credit. As in Study 1, classification into language groups was determined by responses to the LSBQ (Luk & Bialystok, 2013). Thirty-three bilinguals reported English as their L1, with non-English L1s including Arabic, Cantonese, Dari, Farsi, French, Gujarati, Hebrew, Korean, Persian, Portuguese, Punjabi, Russian, Somali, Tamil, Twi, Urdu, Yoruba. Non-English L2 included Arabic, Armenian, Farsi, French, Greek, Gujarati, Hebrew, Hindi, Kiswahili, Korean, Lingala, Macedonian, Panjabi, Pashto, Farsi, Polish, Portuguese, Punjabi, Romanian, Russian, Somalian, Spanish, Swahili, Tamil, Telugu, Twi, and Urdu. Bilinguals reported a mean age of L2 acquisition of 5.2 years (SD = 3.6, range 0–19 years).
English receptive vocabulary and non-verbal fluid intelligence were measured by the Peabody Picture Vocabulary Test (PPVT-III; Dunn & Dunn, 1997) and the Cattell Cultural Fair Test (Cattell, 1957), respectively. In both cases, raw scores were converted to standard scores based on a population mean of 100 and a SD of 15. Background data are reported in Table 2.
Table 2.
Mean scores (and standard deviations) for background measures by language group in Study 2.
| Monolingual | Bilingual | |
|---|---|---|
|
| ||
| N | 53 | 58 |
| Age (years) | 21.8 (3.3) | 22.0 (4.1) |
| PPVT-III* | 101.3 (8.97) | 92.2 (13.42) |
| Cattell | 104.9 (12.0) | 103.7 (12.6) |
| SES (parents’ education) | 2.9 (1.3) | 3.4 (1.6) |
| L2 proficiency (%) | ||
| Speaking* | 19.7 (17.8) | 90.9 (13.5) |
| Understanding* | 33.5 (22.4) | 96.1 (8.0) |
| Reading* | 22.0 (20.5) | 95.3 (9.8) |
| Writing* | 15.9 (17.7) | 92.8 (12.5) |
| Frequency of use (%) | ||
| L1* | 89.3 (14.6) | 52.9 (4.2) |
| L2* | 10.7 (14.6) | 47.1 (4.2) |
Asterisks indicate significant differences between groups.
After obtaining informed consent, participants completed the LSBQ. The flanker task was administered next, followed by the PPVT and the Cattell Cultural Fair Test. Procedures complied with all relevant policies on ethics and were approved by the University Ethics Board.
3.1.2 Flanker task
The materials and apparatus were similar to those used in the non-verbal flanker task in Study 1 with the following differences: 1) the RSI was set to a fixed 250 ms, and 2) there were 36 congruent and 36 incongruent trials in the critical mixed block.
3.2 Results
Groups were equivalent on non-verbal intelligence (Cattell), F < 1. Monolinguals obtained higher scores than bilinguals on the PPVT, F(1, 109) = 16.65, p < 0.001, ηp2 = 0.13, consistent with past research (Bialystok & Luk, 2012). Monolinguals and bilinguals differed on second-language use and proficiency measures, all ps < 0.001 (see Table 2). Accuracy on the flanker task was at ceiling (> 96% correct) and not analyzed further.
Mean RTs for the flanker task are shown in Figure 4A. A 2-way group × congruency mixed-measures ANOVA revealed a significant main effect of congruency, F(1, 109) = 112.33, p < 0.0001, ηp2 = 0.51, with no effect of group, or group by congruency interaction, Fs < 1.7.
Figure 4.
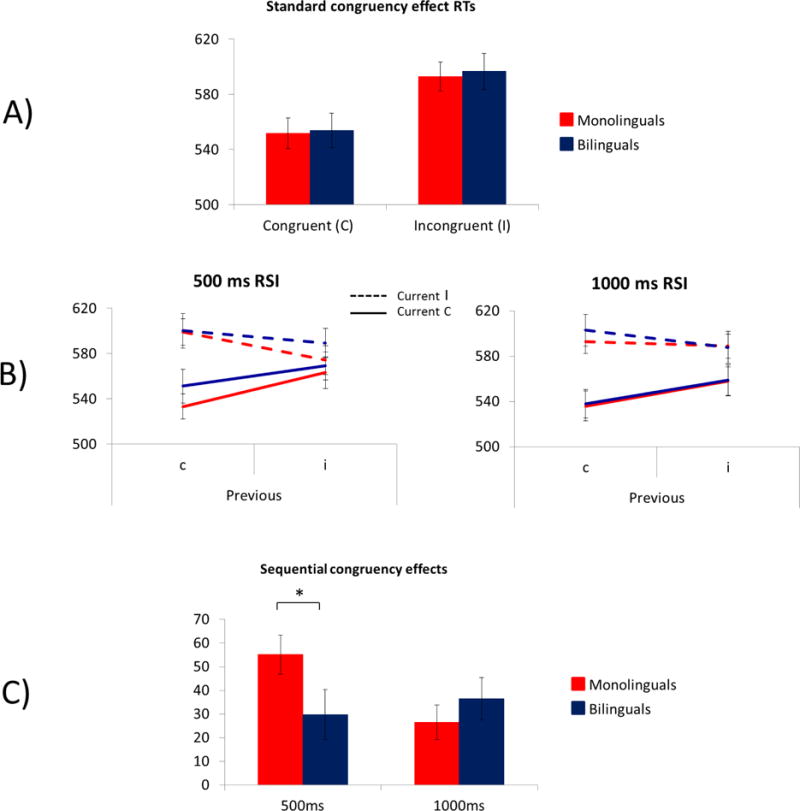
A) Mean RT to current congruent and incongruent trials by language group in Study 2. B) Mean RT as a function of current and previous trial congruency C) SCE by language group in Study 2. The response-to-stimulus intervals was 250 ms. *p < 0.05.
Mean RT as a function of current and previous trial congruency (Figure 4B) and SCEs (Figure 4C) are included in Figure 4. A 3-way ANOVA indicated a significant effect of current trial congruency, F(1, 109) = 112.33, p < 0.0001, ηp2 = 0.51, and an interaction of previous trial congruency and current trial congruency, F(1, 109) = 32.20, p < 0.0001, ηp2 = 0.23. This interaction confirms that the flanker effect was larger following congruent (c-flanker effect = 36 ms) than incongruent (i-flanker effect = 17 ms) trials. These effects were qualified by a 3-way interaction between previous trial congruency, current trial congruency, and group, F(1, 109) = 5.37, p = 0.02, ηp2 = 0.05. Follow-up one-way ANOVAs revealed that the SCE was larger for monolinguals than bilinguals, F(1, 109) = 5.37, p = 0.02, ηp2 = 0.05 (Figure 4). No other effects reached significance, Fs < 1.7.
3.3 Discussion
Study 2 replicated the findings from Study 1 using a different sample of participants and a shorter RSI. Bilinguals and monolinguals did not differ in terms of overall RT on congruent and incongruent trials, but monolinguals were more influenced by previous trial congruency than were bilinguals. These results again are consistent with the interpretation of better disengagement of attention from the previous trial for bilinguals than monolinguals.
4. Study 3
Study 3 applied the SCE analysis to a data set that included measures of EEG while participants performed a flanker task. Therefore, the study explored the electrophysiological correlates of SCEs between monolinguals and bilinguals during a flanker task. EEG requires that long, jittered RSIs are used during ERP recordings in order to avoid overlapping electrophysiological signals between time epochs from trial to trial (Woldorff, 1993). At these long RSIs, all participants are given sufficient time to disengage attention from the previous trial, possibly eliminating behavioral group differences, as found in Study 1. However, EEG signals are sensitive to changes in attention, so if bilinguals are disengaging attention from the previous stimulus more than monolinguals, then this should be reflected in the electrophysiological responses even in the absence of RT differences at long RSIs. Specifically, bilinguals should show less influence than monolinguals from the previous trial on the N2 and P3 ERP components of the current trial.
The N2 is a negative deflection that typically peaks around 250–350 ms after stimulus onset and has a scalp distribution at fronto-central electrode sites (Folstein & Van Petten, 2008; Nieuwenhuis, Yeung, Van Den Wildenberg, & Ridderinkhof, 2003; Yeung, Botvinick, & Cohen, 2004; Yeung & Cohen, 2006). The N2 is sensitive to conflict monitoring (Donkers, & van Boxtel, 2004; Nieuwenhuis et al. 2003), novelty detection (Suwazono, Machado, & Knight, 2000), and error processing (Folstein & Van Petten, 2008). Source localization studies place the generator of the N2 at the ACC (van Veen & Carter, 2002a, 2002b), which is involved in the modulations observed for SCEs (Sheth et al., 2012; Shenhav, Botvinick, & Cohen, 2013). More importantly, recent studies have found that the N2 is also sensitive to SCEs during the flanker task (Clayson & Larson, 2011a, 2011b).
The P3 is an ERP component that occurs approximately 350–500 ms after stimulus onset and has been implicated in stimulus categorization (Mecklinger & Ullsperger, 1993; Nasman & Rosenfeld, 1990), novelty detection (Friedman, Cycowicz, & Gaeta, 2001), and working memory updating before response execution (Polich, 2012); the P3 is also sensitive to SCEs during the flanker task (Clayson & Larson, 2011a, 2011b). The prediction was that monolinguals will show greater modulation of the N2 and P3 components reflecting more influence from the previous trial than bilinguals, even in the absence of behavioral group differences.
4.1 Method
4.1.1 Participants and Procedures
Fifty-five monolinguals and 56 bilinguals participated in exchange for course credit. Participants were assessed for language experience and proficiency using the LSBQ (Luk & Bialystok, 2013). Thirty-three bilinguals reported English as their L1, with non-English L1s including Arabic, Cantonese, Creole, Farsi, French, Hebrew, Korean, Russian, Spanish, Tamil, Vietnamese, and Punjabi. Non-English L2s included Albanian, Bengali, Creole, Farsi, French, Gujrati, Kashmiri, Korean, Nepali, Polish, Punjabi, Russian, Serbian, Spanish, Telugu, Ukranian, and Urdu. The mean age of L2 learning for bilinguals was 4.4 years (SD = 4.0, range 0–12 years). All participants were right-handed, had normal or corrected-to-normal vision, and no history of head injuries or neurological disorders. Background measures for participants are presented in Table 3.
Table 3.
Mean scores (and standard deviations) for background measures by language group in Study 3.
| Monolingual | Bilingual | |
|---|---|---|
|
| ||
| N | 55 | 56 |
| Age (years) | 22.7 (3.1) | 20.6 (2.7) |
| PPVT-III* | 105.3 (11.8) | 100.2 (12.3) |
| SES (parents’ education) | 3.2 (1.1) | 3.4 (1.0) |
| L2 proficiency (%) | ||
| Speaking | – | 83.0 (16.7) |
| Understanding | – | 88.0 (13.3) |
| Reading | – | 68.3 (34.3) |
| Writing | – | 58.9 (34.3) |
| Frequency of use (%) | ||
| L1* | 98.3 (3.3) | 59.3 (26.9) |
| L2* | 1.7 (3.3) | 40.7 (26.9) |
Asterisks indicate significant differences between groups.
As in Study 2, English receptive vocabulary was measured by the PPVT. Participants were first administered the LSBQ and PPVT, and then set up for EEG acquisition. ERPs were recorded while participants completed the flanker task.
4.1.2 Flanker task
The materials and apparatus were similar to Study 2 with the following exceptions: 1) RSI was randomly varied between 1000 ms, 1250 ms, and 1500 ms, 2) there were 100 congruent (C) and 100 incongruent (I) trials randomly intermixed within the experimental block, and 3) responses to C and I trials did not have a time limit.
4.1.3 EEG Recordings
Using the BioSemi Acquisition System, EEG was continuously recorded at a sampling rate of 512 Hz with an online band-pass filter of .01–80 Hz from 64 Ag/AgCl active electrodes that were placed according to the international 10–20 system. Six additional flat type electrodes were applied for offline re-referencing (one electrode on each mastoid), eye blinks (one electrode 1 cm below each eye), and horizontal eye-movements (one electrode placed 1 cm to the left and right of the outer-canthi of each eye). During recording, electrodes were referenced to the common mode sense electrodes and impedances were maintained below 20 kOhm at all electrode sites.
Off-line processing was performed using the EEGLAB v11.0.2.1b toolbox under MATLAB v7.14 (2012, Mathworks, Natick, MA). The electroencephalogram was re-referenced to the averaged mastoids. Stimulus-locked epochs from −200 to 800 ms after stimulus presentation were computed with a 200 ms pre-stimulus baseline. Electrode sites with high frequency noise were interpolated. Trials indicative of muscle tension or drift were removed prior to conducting the eye artifact detection procedure using a simple voltage threshold of 400 microvolts. Horizontal eye-movements and eye blinks were detected and corrected using the Independent Components Analysis (Makeig, Bell, Jung, & Sejnowski, 1996) procedure. The percentage of trials removed was 2.86% and 2.71% for monolinguals and bilinguals, respectively. Grand average event-related potentials were obtained for each participant by electrode site and condition.
4.2 Results
4.2.1 Background Measures
Bilinguals and monolinguals differed in terms of frequency of use of both their L1 and L2, both ps < 0.001. Because an older version of the LSBQ had been used, speaking, understanding, reading, and writing scores were not obtained for monolinguals. Consistent with previous literature, bilinguals had a lower PPVT score than monolinguals, F(1, 110) = 4.83, p = 0.03 (cf. Bialystok & Luk, 2012).
4.2.2 Behavioral Results
Incorrect responses and RTs above 2000 ms were removed from the analyses. This was done in order to be consistent with Studies 1 and 2 that both had a 2000 ms response time cutoff. Accuracy rates for all trial types exceeded 95% and were not analyzed further.
Mean RTs for the flanker task are shown in Figure 5A. A 2-way group × congruency mixed-measures ANOVA revealed a significant main effect of congruency, F(1, 109) = 79.62, p < 0.001, ηp2 = 0.42, with no effect of group, or group by congruency interaction, Fs < 1.
Figure 5.
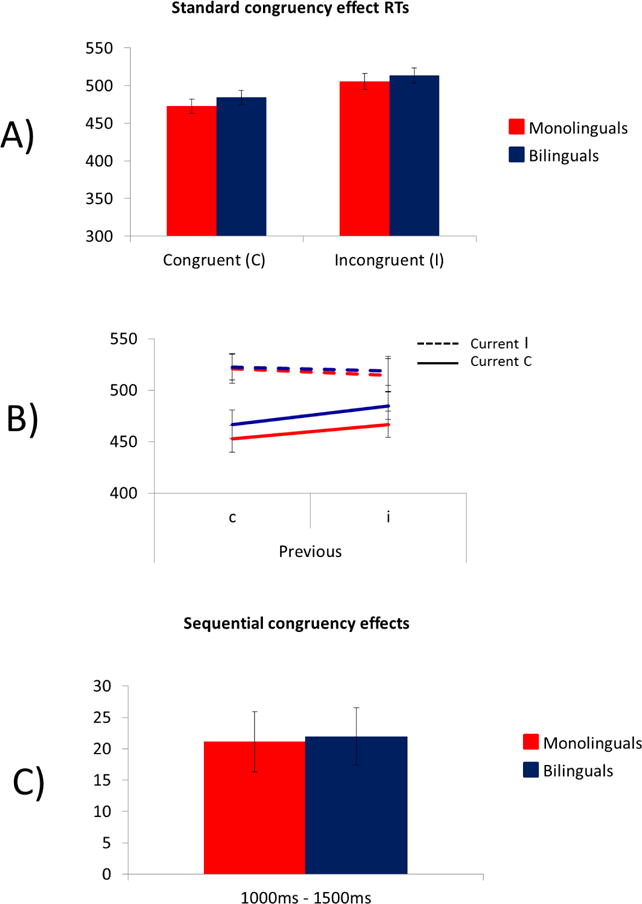
A) Mean RT to current congruent and incongruent trials by language group in Study 3. B) Mean RT as a function of current and previous trial congruency C) SCE by language group in Study 3. Response-to-stimulus intervals varied randomly between 1000–1500 ms.
Figure 5B displays mean RT as a function of current and previous trial congruency and Figure 5C shows SCEs. There was a significant main effect of previous trial congruency, F(1, 109) = 69.01, p < 0.001, ηp2 = 0.39, with faster RT when the previous trial was congruent (mean = 481 ms) than incongruent (mean = 507 ms). A main effect of current trial congruency, F(1, 109) = 79.62, p < 0.001, ηp2 = 0.42, showed faster RTs for congruent trials (mean = 478 ms) than incongruent trials (mean = 509 ms). The previous trial congruency by current trial congruency interaction was significant, F(1,109) = 42.10, p < 0.001, ηp2 = 0.28, showing again that the c-flanker effect (41 ms) was larger than the i-flanker effect (20 ms). There were no main effects or interactions with language group, all Fs < 1.
4.2.3 Electrophysiological Results
Mean amplitude analyses were conducted on the N2 (230–330ms) and P3 (330–500ms) waveforms. For the N2, electrode sites consisted of frontal medial sites (i.e., AF3, AFz, AF4, F1, Fz, F2, FC1, FCz, FC2) as determined by visual inspection and evidence that the N2 flanker effect is focal over frontal medial sites (Folstein & Van Petten, 2008; Nieuwenhuis, Yeung, Van Den Wildenberg, & Ridderinkhof, 2003; Yeung, Botvinick, & Cohen, 2004; Yeung & Cohen, 2006). For the P3, evidence suggests that the P3 is largest at centro-parietal medial sites (Johnson, 1986; Polich, 2012). Thus, the following electrode sites were chosen for the P3: FC1, FCz, FC2, C1, Cz, C2, CP1, CPz, CP2, P1, Pz, P2. The ANOVA for the N2 consisted of 4 within-subject factors: current trial congruency, previous trial congruency, laterality (left, midline, right), and anterior-posterior (anterior-frontal [AF3, AFz, AF4], frontal [F1, Fz, F2], frontal-central [FC1, FCz, FC2]). The ANOVA for the P3 also consisted of 4 within-subject variables: current trial congruency, previous trial congruency, laterality (left, midline, right), and anterior-posterior (frontal-central [FC1, FCz, FC2], central [C1, Cz, C2], central-parietal [CP1, CPz, CP2], parietal [P1, Pz, P2]). Language group was the between-subjects factor for both ANOVAs. The Greenhouse-Geisser correction was applied to variables with more than one degree of freedom in the numerator. Only analyses that contained the critical congruency factors (previous trial congruency, current trial congruency) and/or the group factor are reported.
Analyses on the N2 mean amplitudes revealed that there was no main effect of language group, F < 1. A larger N2 for the current incongruent trials (μV = 0.74, SE = 0.38) than the current congruent trials (μV = 1.43, SE = 0.36), F(1,109) = 11.48, p = 0.001, ηp2 = 0.10, indicated a significant flanker effect. This effect was qualified by a significant interaction of current trial congruency and previous-trial congruency, F(1,109) = 4.14, p = 0.04, ηp2 = 0.04, such that the electrophysiological flanker effect was significant when preceded by an incongruent trial, (congruent: μV = 1.55, SE = 0.38; incongruent: μV = 0.49, SE = 0.39), p < 0.001, F(1,109) = 17.49, p <0.001, ηp2= 0.14, but not by a congruent trial, F(1, 109) =1.25, p = 0.27, (congruent: μV = 1.32, SE = 0.39; incongruent: μV = 0.99, SE = 0.40). Furthermore, language group impacted the size of the flanker effect based on the previous trial as evidenced by a significant interaction of laterality, current-trial congruency, previous-trial congruency and language group, F(2, 218) = 5.62, p = 0.009, ηp2 = 0.05. Simple main effect analyses for each group revealed that, consistent with the above results, the flanker effect was not significant when the previous trial was congruent, all ps > 0.22. However, for monolinguals, when the previous trial was incongruent, the flanker effect was significant across all three levels of laterality, all ps < 0.001. In contrast, for bilinguals, the flanker effect following incongruent trials was smaller and only reached significance on the right side of the midline (the average of AF4, F2, FC2), p = 0.038. These results are shown in Figure 6.
Figure 6.
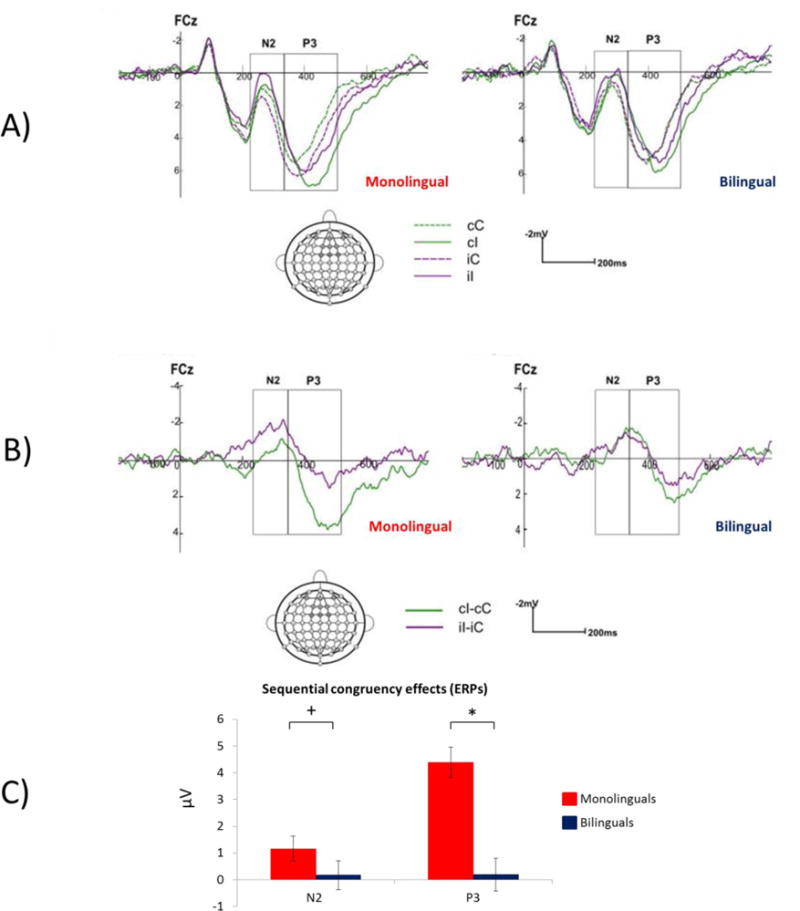
A). ERPs for the four critical trial types used to calculate SCEs. B) ERP difference waves representing the c-flanker effect (cI-cC; green) and the i-flanker effect (iI-iC; purple). Note that for monolinguals the N2 is more sensitive to the i-flanker effect (previous trial incongruent) whereas the P3 is more sensitive to the c-flanker effect (previous trial congruent). C) SCEs (c-flanker effect – i-flanker effect) at FCz. + p = 0.08, *p < 0.05.
Analyses on the P3 mean amplitudes revealed a main effect of current trial congruency, F(1,109) = 6.60, p = 0.012, ηp2 = .06, with larger P3 for incongruent trials (μV = 5.94, SE = 0.41) than congruent trials (μV = 5.34, SE = 0.37). There was no main effect of language group, F < 1. There was an interaction of current trial congruency by previous trial congruency, F(1, 109) = 4.01, p = 0.048, ηp2 = .04, such that the flanker effect was significant when the preceding trial was congruent (congruent: μV = 5.23, SE = 0.38; incongruent: μV = 6.23, SE = 0.43), F(1,109) = 10.24, p = 0.001, ηp2 = .09, but not when it was incongruent, F < 1 (congruent: μV = 5.44, SE = 0.41; incongruent: μV = 5.66, SE = 0.43), F < 1. There were also significant interactions of language group by anterior-posterior by previous trial congruency by current trial congruency, F(3, 327) = 6.30, p = 0.007, ηp2 = 0.06, and the five-way interaction of language group, anterior-posterior, laterality, previous-trial congruency and current-trial congruency, F(6, 678) = 3.30, p = 0.02, ηp2 = 0.03. Simple main effects analyses of the 5-way interaction revealed that for monolinguals, the flanker effect was significant after a congruent trial at all electrode sites with larger differences between the congruent and incongruent trial observed both at anterior sites and along the midline than other sites, all ps < 0.02. For bilinguals, no significant flanker effects were observed at any electrode sites, all ps > 0.122.
4.3 Discussion
Study 3 investigated whether bilinguals and monolinguals differed in their electrophysiological responses to the flanker task as a function of the congruency of the preceding trial. The results showed that monolinguals were more impacted by the preceding trial than were bilinguals.
Behaviorally, no group differences between monolinguals and bilinguals emerged in terms of RT or size of the SCE. This is not surprising given that the experiment required long RSIs during which all participants would have had sufficient time to disengage attention. However, the electrophysiological results showed larger SCEs for monolinguals than bilinguals, indicating more neural processes devoted to previous trial features and more effort disengaging attention.
At the N2, monolinguals showed a large SCE that was driven primarily by the influence of the preceding incongruent trial and this effect was distributed across all frontal-central electrode sites. Bilinguals did not show this pattern, with a much smaller influence of previous trial congruency and only appearing on the right side of the midline. At the P3, monolinguals again showed a large SCE effect, this time driven primarily by the influence of previous congruent trials and this was again distributed across all electrode sites included in the region of interest. Bilinguals showed no evidence of being influenced by previous trials at the P3.
These findings support the interpretation that bilinguals have disengaged attention from the previous stimulus while performing the current trial whereas monolinguals show a lag in their ability to redirect attention to the next trial. Monolinguals continue to be influenced by the previous stimulus at the P3, whereas bilinguals do not. Furthermore, more resources are required for monolinguals to disengage attention at the N2, as evidenced by larger amplitude differences and more distributed processes across electrode sites. In sum, bilinguals disengaged attention from previous information more efficiently and rapidly than monolinguals.
5. General Discussion
Across three experiments, monolinguals and bilinguals who performed equivalently on a flanker task when assessed by mean RT to congruent and incongruent trials showed systematically different patterns of attention as revealed by analyses of SCEs. There were four main findings. First, group differences in SCEs emerged in spite of no difference between groups in standard measures of reaction time. Second, the largest group differences were found for nonverbal tasks. Third, effects were more reliable for shorter RSIs than longer ones. Fourth, electrophysiological responses revealed less influence of previous trial congruency for bilinguals than monolinguals at long RSIs. Each of these findings will be discussed in turn.
Previous studies based on mean RT to congruent and incongruent trials frequently show no difference between language groups for young adults, leading to the possibility that performance does not differ between monolinguals and bilinguals, at least for this age (e.g., Paap & Greenberg, 2013). This pattern of null effects was replicated in all three experiments in the current study. However, calculating mean RT without regard for the previous trial averages performance across a potentially important factor, masking its effects. Incorporating previous trial information through the calculation of SCEs revealed that bilinguals were less influenced by the features of the previous trial than monolinguals. These findings provide insight into why some studies show bilingual advantages on executive control tasks and some do not. Specifically, reliance on previous trial information is beneficial when the trial type is repeated (cC, iI) but slows performance when the trial type is changed (cI, iC), but because bilinguals disengage attention from previous information more rapidly than monolinguals, these effects are diminished. Better disengagement results in more efficient performance over time, as evidenced by the finding that practice leads to less reliance on previous trial information and smaller SCEs (Mayr & Awe, 2009; van Steenbergen et al., 2015).
Study 1 also confirmed that the task domain influenced results by showing larger group differences in the nonverbal than the verbal task. In both tasks, monolinguals experienced larger facilitation effects for repeat trials and larger interference effects for change trials than bilinguals but the pattern was muted in the verbal task. In this case, there were no group differences in SCE for the long RSI. Possibly, faster language processing by monolinguals facilitated disengagement so that at the longer RSI, the monolinguals caught up with the bilinguals.
Smaller SCEs for bilinguals at short RSIs suggest that more rapid disengagement for bilinguals might occur very early, within 500 ms after response. Behavioral results from all three studies and electrophysiological responses from Study 3 support this conclusion. In Studies 1 and 2, the RSI ranged from 250 ms (Study 2) to 1000 ms (Study 1), with significant group differences at all intervals, but the largest difference occurring within 500 ms. In Study 3 where the RSI was jittered from 1000 ms to 1500 ms, there were no behavioral effects but electrophysiological group differences in SCEs occurred within 500 ms after stimulus onset. This early disengagement for bilinguals may explain why tasks with long RSIs do not produce group differences in SCEs; both groups have had enough time to disengage from the previous stimulus.
Electrophysiological responses at the N2 revealed larger SCEs for monolinguals than bilinguals. Monolinguals exhibited a larger flanker effect following incongruent stimuli than bilinguals, an effect that was distributed across frontal electrode sites. The appearance of this N2 effect following incongruent trials, but not congruent trials, is consistent with the idea that the N2 is sensitive to conflict and reflects the adjustment of responses to recently encountered conflict (Yeung, Botvinick, & Cohen, 2004). It is also consistent with the conflict monitoring theory (Botvinick et al., 2001), which suggests that incongruent trials result in refocused attention to task-relevant features, making the following incongruent trials the focus of greater monitoring and therefore modulating the size of the N2. One might have expected the N2 to be smaller for iI trials than cI trials given that greater N2 amplitudes have been associated with more focus on distractors, rather than task-relevant information (Yeung & Cohen, 2006; Yeung, Ralph, & Nieuwenhuis, 2007). However, larger N2 amplitudes are also associated with more conflict influencing performance at a global level (Grützmann et al., 2014); global processing of conflict could lead to greater N2 amplitudes for iI trials than cI trials. Importantly, this N2 modulation was substantially greater for monolinguals than bilinguals, suggesting a role for the bilingual experience of rapidly disengaging attention from multiple linguistic contexts.
P3 responses also revealed large language group differences, again indicating less influence of the previous trials for bilinguals than monolinguals. In this case, bilinguals showed no effect of previous trial congruency, presumably because they had already disengaged attention from the previous stimulus by this point. These results are again consistent with a conflict monitoring and resource allocation model of the P3 for monolinguals, in which previous trial information leads to subsequent changes in attentional focus (Clayson & Larson, 2011a). P3 amplitudes were greatest for cI trials and smallest for cC trials, suggesting that the previous congruent trials lead to a broadening of attention that results in the incorporation of more flanker information on subsequent trials. Therefore, cC trials benefited from the broadening of attention, so fewer attentional resources were required, resulting in smaller P3 amplitude. In contrast, more attention needed to be devoted to cI trials in order to parse out the irrelevant information because of the broadening of attention.
Although these results are consistent with top-down control interpretations, such as conflict monitoring, it is possible that the results of the N2 and P3 analyses reflect a combination of top-down control and bottom-up retrieval processes, consistent with Egner’s (2014) view that the two simultaneously contribute to the size of the SCE. For example, the sensitivity of the P3 to previous trial information for monolinguals could reflect greater interference following retrieval of past event files. In other words, monolinguals have a more difficult time disengaging from elaborative processing following retrieval of past event files when the current stimulus is presented. In this case, cI trials produce greater P3 amplitudes than cC trials because stimulus categorization and memory updating required greater effort to disengage from elaborative processing of the retrieved partial match trial than the complete overlap trial. By both top-down and bottom-up accounts, bilinguals are less influenced than monolinguals by previous trial information. The interpretation is that this ability is the result of extensive linguistic experience with disengagement of attention for bilinguals, leading to more efficient neural processing.
It is interesting to note that behavioral group differences in Studies 1 and 2 appear more driven by flanker effects following congruent (c-flanker effect) rather than incongruent (i-flanker effect) trials. Similarly, in Study 3, the P3 is sensitive to group differences in the c-flanker effect but not the i-flanker effect. It is possible that groups differ mainly in how congruent trials affect subsequent performance and that this is indexed by P3 modulations. However, such an interpretation is entirely speculative: We had no a priori reason to expect larger group differences for c-flanker than i-flanker effects and in Study 3, the N2 indicated group differences for the i-flanker and not the c-flanker effect. It is more likely that both previous congruent and previous incongruent trials are contributing to the overall SCE differences between groups. Future studies should examine these more nuanced details.
A theoretical implication of these results concerns the task-specificity of SCEs. Previous work has suggested that SCEs do not transfer across tasks that have different sources of conflict (e.g., feature vs. response-based conflict; Kim, Chung, & Kim, 2012; Lee, & Cho, 2013; for a review see Braem, Abrahamse, Duthoo, & Notebaert, 2014). For example, Egner, Delano and Hirsch (2007) had participants perform both a Simon and a Stroop task within the same block and found SCEs only when the previous trial was from the same task (Stroop to Stroop or Simon to Simon) but not when the task changed (Stroop to Simon or Simon to Stroop). The present results show that bilingual experience dealing with linguistic conflict may support more general processes that do transfer across tasks (i.e., linguistic to flanker). Thus, the results both endorse a theoretical interpretation of SCEs and help explain why executive control differences between monolinguals and bilinguals are sometimes not found.
These results are also consistent with an account in which predictions of upcoming cognitive load are based on current and recent history with conflict (Grundy & Shedden, 2014a, 2014b; Shenhav, Botvinick, & Cohen, 2013; Sheth et al., 2012). Neuroimaging studies examining SCEs have implicated the ACC in predicting changes in upcoming cognitive load (Shenhav, Botvinick, & Cohen, 2013; Sheth et al., 2012). Furthermore, the N2 has an ACC generator (van Veen & Carter, 2002a, 2002b), and the ACC is a cognitive control center that has been shown to be critical in dissociating processes between monolinguals and bilinguals on conflict tasks (Abutalebi et al., 2012; Gold et al., 2013). Sheth et al. (2012) provided evidence using SCEs that more conflict leads to increased predictions in upcoming cognitive load in the ACC. By this account, because bilinguals demonstrate smaller SCEs than monolinguals, bilinguals also make predictions of smaller upcoming cognitive load, possibly because of extensive practice with conflict in their environments. Therefore, large changes in upcoming cognitive load are not made, and this leads to more consistent performance. If bilinguals disengage attention from previous information more rapidly than monolinguals, predictions of upcoming cognitive load would be smaller because conflict trials do not influence subsequent performance to the same degree. Future neuroimaging work is needed to confirm that the ACC can detect these differences.
It is important to explain the present results in light of findings that SCE effects are not present in children but develop over time (Waxer & Morton, 2011). Thus it may seem contradictory that smaller SCEs would be considered more efficient; monolinguals might appear like more mature performers than bilinguals. However, it is important to note that even though bilinguals showed smaller SCE effects than monolinguals, they nonetheless showed an SCE effect. This means that, unlike children, bilinguals were influenced by previous trial congruency but to a lesser degree than monolinguals. Furthermore, SCE effects are reduced with practice for all individuals (Mayr & Awe, 2009; van Steenbergen, 2015), suggesting that less reliance on previous trial information is more efficient over time, at least for conflict resolution tasks.
As in some previous research (e.g., Paap & Greenberg, 2013), standard analyses of overall RT failed to detect differences between monolingual and bilingual young adults on a simple conflict task. The expectation that RT on congruent and incongruent trials or the difference between them discriminates performance across language groups follows from the conception of executive functions proposed by Miyake et al. (2000) based on three unique component processes, namely, inhibition, shifting and updating. The notion that bilinguals had better inhibition because of their experience in inhibiting the unwanted language had been the prevailing view in the field (e.g., Bialystok et al., 2009) and was congenial with the Miyake model. However, this inhibition view failed both as an explanation of bilingualism, in which it is widely accepted that both languages are constantly active and no inhibition is involved (e.g., Kroll et al, 2012) and as an account of the data in which bilinguals typically outperformed monolinguals not only on incongruent trials that require inhibition but also on congruent trials that do not (e.g., Hilchey & Klein, 2011). Instead, bilingual language processing is more compatible with a view in which constant monitoring and disengagement of attention from the non-target language are required to focus on the target language in the face of active competition. This attention and monitoring interpretation is consistent with the observation of smaller SCEs for bilinguals and furthermore explains why language group differences are often not found in tasks designed to test inhibition and switching in young adults.
Understanding SCEs as a mechanism of attention has proven to be a fruitful approach to investigating cognitive processes across the lifespan (review in Egner, 2014) and has been linked to ACC function (Botvinick et al., 2001), a center critical to conflict processing. The present studies provide evidence that SCEs can also be crucial to understanding how bilingual experience modifies cognitive processes. From early on, bilinguals learn to modulate attention to multiple sources of linguistic input, an experience that appears to have broad consequences. These consequences may impact general cognitive function across the lifespan.
Acknowledgments
The research reported in this paper was funded by grant R01HD052523 from the US National Institutes of Health and grant A2559 from the Natural Sciences and Engineering Research Council of Canada to EB. We thank Michelle Goodman & Jiyoon Oh for data collection in Experiment 3.
Appendix: Interaction Effects in Behavioral Results
Study 1
- Non-verbal arrow flanker ➔ 3-way interaction between previous trial congruency, current trial congruency, and group:
-
○Between-group comparisons for c-flanker and i-flanker effects
-
▪c-flanker effect was larger for monolinguals than bilinguals: F(1, 57) = 6.41, p = 0.014, ηp2 = 0.101
-
▪No difference was observed for the i-flanker effect, F(1, 57) = 0.01, p = 0.94, ηp2 = 0.00.
-
▪
-
○Within-group comparisons between iC-cC and iI-iI
-
▪Monolinguals:
- iC-cC ➔ 30.7, t(27) = 5.10, p <0.001
- cI-iI ➔ 20.0, t(27) = 3.81, p = 0.001
-
▪Bilinguals:
- iC-cC ➔ 9.9, t(30) =1.80, p =0.083
- cI-iI ➔ 11.9, t(30) =2.02, p = 0.052
-
▪
-
○
- Verbal word flanker ➔ 4-way interaction between RSI, previous trial congruency, current trial congruency, and group:
-
○500ms:
-
▪Between-group comparisons for c-flanker and i-flanker effects
- c-flanker effect was marginally larger for monolinguals than bilinguals: F(1, 57) =2.97, p =0.09, ηp2 =0.05
- No difference was observed for the i-flanker effect, F(1, 57) = 1.15, p = 0.29, ηp2 = 0.02
-
▪Within-group comparisons between iC-cC and iI-iI
- Monolinguals:
-
○iC-cC ➔30.1 ms, t(27) = 3.22, p = 0.003
-
○cI-iI ➔24.9 ms, t(27) = 2.98, p = 0.006
-
○
- Bilinguals:
-
○iC-cC ➔18.9 ms, t(30) =2.09, p = 0.045
-
○cI-iI ➔10.9 ms, t(30) =1.18, p = 0.246
-
○
-
▪
-
○1000ms:
-
▪No group difference was observed for the c-flanker effect, F(1, 57) = 0.58, p = 0.45, ηp2 = 0.01
-
▪No group difference was observed for the i-flanker effect, F(1, 57) = 0.04, p = 0.85, ηp2 = 0.001
-
▪Monolinguals:
- iC-cC ➔22.3 ms, t(27) = 2.71, p = 0.01
- cI-iI ➔4.2 ms, t(27) =0.82, p =0.42
-
▪Bilinguals:
- iC-cC ➔21.3 ms, t(30) = 3.67, p = 0.001
- cI-iI ➔15.1 ms, t(30) =2.26, p = 0.03
-
▪
-
○
Study 2
- Non-verbal arrow flanker ➔ 3-way interaction between previous trial congruency, current trial congruency, and group:
-
○Between-group comparisons for c-flanker and i-flanker effects
-
▪c-flanker effect was larger for monolinguals than bilinguals: F(1, 109) = 6.10, p = 0.02, ηp2 = 0.05
-
▪No difference was observed for the i-flanker effect, F(1, 109) = 0.21, p = 0.65, ηp2 = 0.002
-
▪
-
○Within-group comparisons between iC-cC and iI-iI
-
▪Monolinguals:
- iC-cC ➔17.1 ms, t(52) = 4.06, p < 0.001
- cI-iI ➔15.6 ms, t(52) =3.65, p = 0.001
-
▪Bilinguals:
- iC-cC ➔3.3 ms, t(57) = 0.81, p = 0.42
- cI-iI ➔10.3 ms, t(57) = 2.50, p = 0.02
-
▪
-
○
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
The sequential congruency effect is also commonly referred to as the “conflict adaptation effect” (e.g. Botvinick et al., 2001) or the “Gratton effect” (e.g. Schmidt & De Houwer, 2011). We prefer the term “sequential congruency effect” because of its descriptive value for the effect and theoretical neutrality with respect to mechanism.
Visual inspection of the waveforms suggested that the previous trial might influence the flanker effect for bilinguals within the 380–500 ms window. Statistics within this window revealed a significant flanker effect (p <0.05), but no influence of previous trial congruency or any interactions with previous trial congruency.
References
- Abutalebi J, Della Rosa PA, Green DW, Hernandez M, Scifo P, Keim R, Costa A. Bilingualism tunes the anterior cingulate cortex for conflict monitoring. Cerebral Cortex. 2012;22:2076–2086. doi: 10.1093/cercor/bhr287. [DOI] [PubMed] [Google Scholar]
- Adesope OO, Lavin T, Thompson T, Ungerleider C. A systematic review and meta-analysis of the cognitive correlates of bilingualism. Review of Educational Research. 2010;80:207–245. [Google Scholar]
- Alexander WH, Brown JW. Medial prefrontal cortex as an action-outcome predictor. Nature neuroscience. 2011;14(10):1338–1344. doi: 10.1038/nn.2921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allport A, Wylie G. Task switching, stimulus-response bindings, and negative priming. Control of cognitive processes: Attention and performance XVIII. 2000:35–70. [Google Scholar]
- Barac R, Bialystok E, Castro DC, Sanchez M. The cognitive development of young dual language learners: A critical review. Early Childhood Research Quarterly. 2014;29:699–714. doi: 10.1016/j.ecresq.2014.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bialystok E. The bilingual adaptation: How minds accommodate experience. Psychological Bulletin. doi: 10.1037/bul0000099. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bialystok E. The signal and the noise: Finding the pattern in human behavior. Linguistic Approaches to Bilingualism. 2016;6:517–534. [Google Scholar]
- Bialystok E. Effect of bilingualism and computer video game experience on the Simon task. Canadian Journal of Experimental Psychology. 2006;60:68–79. doi: 10.1037/cjep2006008. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Craik FI, Green DW, Gollan TH. Bilingual minds. Psychological Science in the Public Interest. 2009;10:89–129. doi: 10.1177/1529100610387084. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Craik FI, Klein R, Viswanathan M. Bilingualism, aging, and cognitive control: evidence from the Simon task. Psychology and Aging. 2004;19:290–303. doi: 10.1037/0882-7974.19.2.290. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Craik FI, Luk G. Lexical access in bilinguals: Effects of vocabulary size and executive control. Journal of Neurolinguistics. 2008;21:522–538. [Google Scholar]
- Bialystok E, Craik FI, Luk G. Bilingualism: consequences for mind and brain. Trends in Cognitive Sciences. 2012;16:240–250. doi: 10.1016/j.tics.2012.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bialystok E, Luk G. Receptive vocabulary differences in monolingual and bilingual adults. Bilingualism: Language and Cognition. 2012;15:397–401. doi: 10.1017/S1366728909990423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bialystok E, Martin MM, Viswanathan M. Bilingualism across the lifespan: The rise and fall of inhibitory control. International Journal of Bilingualism. 2005;9:103–119. [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychological Review. 2001;108:624–652. doi: 10.1037/0033-295x.108.3.624. [DOI] [PubMed] [Google Scholar]
- Braem S, Abrahamse EL, Duthoo W, Notebaert W. What determines the specificity of conflict adaptation? A review, critical analysis, and proposed synthesis. Frontiers in Psychology. 2014;5:1134. doi: 10.3389/fpsyg.2014.01134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cattell RB. Culture fair intelligence test, a measure of “g”. Savoy, IL: Western Psychological Services; 1957. [Google Scholar]
- Clayson PE, Larson MJ. Conflict adaptation and sequential trial effects: Support for the conflict monitoring theory. Neuropsychologia. 2011a;49:1953–1961. doi: 10.1016/j.neuropsychologia.2011.03.023. [DOI] [PubMed] [Google Scholar]
- Clayson PE, Larson MJ. Effects of repetition priming on electrophysiological and behavioral indices of conflict adaptation and cognitive control. Psychophysiology. 2011b;48:1621–1630. doi: 10.1111/j.1469-8986.2011.01265.x. [DOI] [PubMed] [Google Scholar]
- Compton RJ, Arnstein D, Freedman G, Dainer’Best J, Liss A. Cognitive control in the intertrial interval: evidence from EEG alpha power. Psychophysiology. 2011;48:583–590. doi: 10.1111/j.1469-8986.2010.01124.x. [DOI] [PubMed] [Google Scholar]
- Compton RJ, Huber E, Levinson AR, Zheutlin A. Is “conflict adaptation” driven by conflict? Behavioral and EEG evidence for the underappreciated role of congruent trials. Psychophysiology. 2012;49:583–589. doi: 10.1111/j.1469-8986.2012.01354.x. [DOI] [PubMed] [Google Scholar]
- Costa A, Hernández M, Costa-Faidella J, Sebastián-Gallés N. On the bilingual advantage in conflict processing: Now you see it, now you don’t. Cognition. 2009;113:135–149. doi: 10.1016/j.cognition.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Costa A, Hernández M, Sebastián-Gallés N. Bilingualism aids conflict resolution: Evidence from the ANT task. Cognition. 2008;106:59–86. doi: 10.1016/j.cognition.2006.12.013. [DOI] [PubMed] [Google Scholar]
- De Baene W, Duyck W, Brass M, Carreiras M. Brain circuit for cognitive control is shared by task and language switching. Journal of Cognitive Neuroscience. 2015;27:1752–1765. doi: 10.1162/jocn_a_00817. [DOI] [PubMed] [Google Scholar]
- Dunn LM, Dunn LM. Peabody Picture Vocabulary Test–III. Circle Pines, MN: American Guidance Service; 1997. [Google Scholar]
- Egner T. Creatures of habit (and control): a multi-level learning perspective on the modulation of congruency effects. Frontiers in Psychology. 2014;5:1247. doi: 10.3389/fpsyg.2014.01247. Article number. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egner T, Delano M, Hirsch J. Separate conflict-specific cognitive control mechanisms in the human brain. Neuroimage. 2007;35:940–948. doi: 10.1016/j.neuroimage.2006.11.061. [DOI] [PubMed] [Google Scholar]
- Egner T, Ely S, Grinband J. Going, going, gone: characterizing the time-course of congruency sequence effects. Frontiers in Psychology. 2010;1:154. doi: 10.3389/fpsyg.2010.00154. Article number. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emmorey K, Luk G, Pyers JE, Bialystok E. The source of enhanced cognitive control in bilinguals: Evidence from bimodal bilinguals. Psychological Science. 2008;19:1201–1206. doi: 10.1111/j.1467-9280.2008.02224.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folstein JR, Van Petten C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 2008;45:152–170. doi: 10.1111/j.1469-8986.2007.00602.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman D, Cycowicz YM, Gaeta H. The novelty P3: An event-related brain potential (ERP) sign of the brain’s evaluation of novelty. Neuroscience & Biobehavioral Reviews. 2001;25(4):355–373. doi: 10.1016/s0149-7634(01)00019-7. [DOI] [PubMed] [Google Scholar]
- Gathercole VCM, Thomas EM, Kennedy I, Prys C, Young N, Viñas Guasch N, Jones L. Does language dominance affect cognitive performance in bilinguals? Lifespan evidence from preschoolers through older adults on card sorting, Simon, and metalinguistic tasks. Frontiers in Psychology. 2014;5:11. doi: 10.3389/fpsyg.2014.00011. Article number. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gollan TH, Fennema-Nostestine C, Montoya RI, Jernigan TL. The bilingual effect on Boston naming test performance. Journal of the International Neuropsychological Society. 2007;13:197–208. doi: 10.1017/S1355617707070038. [DOI] [PubMed] [Google Scholar]
- Gollan TH, Montoya RI, Cera C, Sandoval TC. More use almost always means a smaller frequency effect: Aging, bilingualism, and the weaker links hypothesis. Journal of Memory and Language. 2008;58:787–814. doi: 10.1016/j.jml.2007.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gollan TH, Montoya RI, Fennema-Notestine C, Morris SK. Bilingualism affects picture naming but not picture classification. Memory & Cognition. 2005;33:1220–1234. doi: 10.3758/bf03193224. [DOI] [PubMed] [Google Scholar]
- Gollan TH, Sandoval T, Salmon DP. Cross-language intrusion errors in aging bilinguals reveal the link between executive control and language selection. Psychological Science. 2011;22:1155–1164. doi: 10.1177/0956797611417002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gold BT, Kim C, Johnson NF, Kryscio RJ, Smith CD. Lifelong bilingualism maintains neural efficiency for cognitive control in aging. The Journal of Neuroscience. 2013;33:387–396. doi: 10.1523/JNEUROSCI.3837-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gratton G, Coles MG, Donchin E. Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General. 1992;121:480–506. doi: 10.1037//0096-3445.121.4.480. [DOI] [PubMed] [Google Scholar]
- Grundy JG, Keyvani Chahi A. Post‐conflict slowing effects in monolingual and bilingual children. Developmental Science. 2017;20:e12488. doi: 10.1111/desc.12488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grundy JG, Shedden JM. A role for recency of response conflict in producing the bivalency effect. Psychological Research. 2014a;78:679–691. doi: 10.1007/s00426-013-0520-x. [DOI] [PubMed] [Google Scholar]
- Grundy JG, Shedden JM. Support for a history-dependent predictive model of dACC activity in producing the bivalency effect: An event-related potential study. Neuropsychologia. 2014b;57:166–178. doi: 10.1016/j.neuropsychologia.2014.03.008. [DOI] [PubMed] [Google Scholar]
- Hernández M, Martin CD, Barceló F, Costa A. Where is the bilingual advantage in task-switching? Journal of Memory and Language. 2013;69(3):257–276. [Google Scholar]
- Hilchey MD, Klein RM. Are there bilingual advantages on nonlinguistic interference tasks? Implications for the plasticity of executive control processes. Psychonomic Bulletin & Review. 2011;18:625–658. doi: 10.3758/s13423-011-0116-7. [DOI] [PubMed] [Google Scholar]
- Holroyd CB, Yeung N. Motivation of extended behaviors by anterior cingulate cortex. Trends in Cognitive Sciences. 2012;16(2):122–128. doi: 10.1016/j.tics.2011.12.008. [DOI] [PubMed] [Google Scholar]
- Ivanova I, Costa A. Does the bilingualism hamper lexical access in speech production? Acta Psychologica. 2008;127:277–288. doi: 10.1016/j.actpsy.2007.06.003. [DOI] [PubMed] [Google Scholar]
- Kalia V, Wilbourn MP, Ghio K. Better early or late? Examining the influence of age of exposure and language proficiency on executive function in early and late bilinguals. Journal of Cognitive Psychology. 2014;26:699–713. [Google Scholar]
- Kerns JG, Cohen JD, MacDonald AW, Cho RY, Stenger VA, Carter CS. Anterior cingulate conflict monitoring adjustments in control. Science. 2004;303(5660):1023–1026. doi: 10.1126/science.1089910. [DOI] [PubMed] [Google Scholar]
- Kim C, Chung C, Kim J. Conflict adjustment through domain-specific multiple cognitive control mechanisms. Brain Research. 2012;1444:55–64. doi: 10.1016/j.brainres.2012.01.023. [DOI] [PubMed] [Google Scholar]
- Klein RM. Inhibition of return. Trends in Cognitive Sciences. 2000;4:138–147. doi: 10.1016/s1364-6613(00)01452-2. [DOI] [PubMed] [Google Scholar]
- Kovács ÁM, Mehler J. Cognitive gains in 7-month-old bilingual infants. Proceedings of the National Academy of Sciences. 2009;106:6556–6560. doi: 10.1073/pnas.0811323106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroll JF, Dussias PE, Bogulski CA, Valdes Kroff JR. Juggling two languages in one mind: What bilinguals tell us about language processing and its consequences for cognition. In: Ross B, editor. The psychology of learning and motivation. Vol. 56. San Diego, CA: Academic Press; 2012. pp. 229–262. [Google Scholar]
- Landry R, Bryson SE. Impaired disengagement of attention in young children with autism. Journal of Child Psychology and Psychiatry. 2004;45:1115–1122. doi: 10.1111/j.1469-7610.2004.00304.x. [DOI] [PubMed] [Google Scholar]
- Luk G, Bialystok E, Craik FI, Grady CL. Lifelong bilingualism maintains white matter integrity in older adults. The Journal of Neuroscience. 2011;31:16808–16813. doi: 10.1523/JNEUROSCI.4563-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luk G, Bialystok E. Bilingualism is not a categorical variable: Interaction between language proficiency and usage. Journal of Cognitive Psychology. 2013;25:605–621. doi: 10.1080/20445911.2013.795574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luk G, Green DW, Abutalebi J, Grady C. Cognitive control for language switching in bilinguals: A quantitative meta-analysis of functional neuroimaging studies. Language and Cognitive Processes. 2012;27:1479–1488. doi: 10.1080/01690965.2011.613209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marian V, Chabal S, Bartolotti J, Bradley K, Hernandez AE. Differential recruitment of executive control regions during phonological competition in monolinguals and bilinguals. Brain and language. 2014;139:108–117. doi: 10.1016/j.bandl.2014.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayr U, Awh E. The elusive link between conflict and conflict adaptation. Psychological Research. 2009;73:794–802. doi: 10.1007/s00426-008-0191-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishra RK, Hilchey MD, Singh N, Klein RM. On the time course of exogenous cueing effects in bilinguals: higher proficiency in a second language is associated with more rapid endogenous disengagement. The Quarterly Journal of Experimental Psychology. 2012;65:1502–1510. doi: 10.1080/17470218.2012.657656. [DOI] [PubMed] [Google Scholar]
- Miyake A, Friedman NP, Emerson MJ, Witzki AH, Howerter A, Wager TD. The unity and diversity of executive functions and their contributions to complex “Frontal Lobe” tasks: a latent variable analysis. Cognitive Psychology. 2000;41:49–100. doi: 10.1006/cogp.1999.0734. [DOI] [PubMed] [Google Scholar]
- Nasman VT, Rosenfeld JP. Parietal P3 response as an indicator of stimulus categorization: increased P3 amplitude to categorically deviant target and nontarget stimuli. Psychophysiology. 1990;27:338–350. doi: 10.1111/j.1469-8986.1990.tb00393.x. [DOI] [PubMed] [Google Scholar]
- Nieuwenhuis S, Yeung N, Van Den Wildenberg W, Ridderinkhof KR. Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency. Cognitive, Affective and Behavioral Neuroscience. 2003;3:17–26. doi: 10.3758/cabn.3.1.17. [DOI] [PubMed] [Google Scholar]
- Paap KR, Greenberg ZI. There is no coherent evidence for a bilingual advantage in executive processing. Cognitive Psychology. 2013;66:232–258. doi: 10.1016/j.cogpsych.2012.12.002. [DOI] [PubMed] [Google Scholar]
- Paap KR, Johnson HA, Sawi O. Bilingual advantages in executive functioning either do not exist or are restricted to very specific and undetermined circumstances. Cortex. 2015;69:265–278. doi: 10.1016/j.cortex.2015.04.014. [DOI] [PubMed] [Google Scholar]
- Polich J. Neuropsychology of P300. Oxford handbook of event-related potential components. 2012:159–188. [Google Scholar]
- Pons F, Bosch L, Lewkowicz DJ. Bilingualism modulates infants’ selective attention to the mouth of a talking face. Psychological Science. 2015;26:490–498. doi: 10.1177/0956797614568320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prior A, Gollan TH. Good Language-Switchers are Good Task-Switchers: Evidence from Spanish–English and Mandarin–English Bilinguals. Journal of the International Neuropsychological Society. 2011;17:682–691. doi: 10.1017/S1355617711000580. [DOI] [PubMed] [Google Scholar]
- Prior A, MacWhinney B. A bilingual advantage in task switching. Bilingualism: Language and cognition. 2010;13(02):253–262. doi: 10.1017/S1366728909990526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt JR, De Houwer J. Now you see it, now you don’t: Controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychologica. 2011;138:176–186. doi: 10.1016/j.actpsy.2011.06.002. [DOI] [PubMed] [Google Scholar]
- Schmidt JR, Weissman DH. Congruency sequence effects without feature integration or contingency learning confounds. PloS One. 2014;9:e102337. doi: 10.1371/journal.pone.0102337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh L, Fu CS, Rahman AA, Hameed WB, Sanmugam S, Agarwal P, Rifkin‐Graboi A. Back to basics: A bilingual advantage in infant visual habituation. Child Development. 2015;86:294–302. doi: 10.1111/cdev.12271. [DOI] [PubMed] [Google Scholar]
- Shenhav A, Botvinick MM, Cohen JD. The expected value of control: An integrative theory of anterior cingulate cortex function. Neuron. 2013;79:217–240. doi: 10.1016/j.neuron.2013.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shenhav A, Cohen JD, Botvinick MM. Dorsal anterior cingulate cortex and the value of control. Nature Neuroscience. 2016;19(10):1286–1291. doi: 10.1038/nn.4384. [DOI] [PubMed] [Google Scholar]
- Sheth SA, Mian MK, Patel SR, Asaad WF, Williams ZM, Dougherty DD, Eskandar EN. Human dorsal anterior cingulate cortex neurons mediate ongoing behavioural adaptation. Nature. 2012;488:218–221. doi: 10.1038/nature11239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipley WC, Gruber CP, Martin TA, Klein AM. Shipley-2 Manual. Los Angeles, CA: Western Psychological Services; 2009. [Google Scholar]
- Spivey MJ, Marian V. Cross talk between native and second languages: Partial activation of an irrelevant lexicon. Psychological Science. 1999;10:281–284. [Google Scholar]
- Stroop JR. Studies of interference in serial verbal reactions. Journal of Experimental Psychology. 1935;18:643–662. [Google Scholar]
- van Steenbergen H, Haasnoot E, Bocanegra BR, Berretty EW, Hommel B. Practice explains abolished behavioural adaptation after human dorsal anterior cingulate cortex lesions. Scientific Reports. 2015;5:9721. doi: 10.1038/srep09721. Article number. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Veen V, Carter CS. The anterior cingulate as a conflict monitor: fMRI and ERP studies. Physiology & Behavior. 2002a;77:477–482. doi: 10.1016/s0031-9384(02)00930-7. [DOI] [PubMed] [Google Scholar]
- Van Veen V, Carter CS. The timing of action-monitoring processes in the anterior cingulate cortex. Journal of Cognitive Neuroscience. 2002b;14:593–602. doi: 10.1162/08989290260045837. [DOI] [PubMed] [Google Scholar]
- Wang X, Wang T, Chen Z, Hitchman G, Liu Y, Chen A. Functional connectivity patterns reflect individual differences in conflict adaptation. Neuropsychologia. 2015;70:177–184. doi: 10.1016/j.neuropsychologia.2015.02.031. [DOI] [PubMed] [Google Scholar]
- Waxer M, Morton JB. The development of future-oriented control: An electrophysiological investigation. NeuroImage. 2011;56(3):1648–1654. doi: 10.1016/j.neuroimage.2011.02.001. [DOI] [PubMed] [Google Scholar]
- Weikum WM, Vouloumanos A, Navarra J, Soto-Faraco S, Sebastian-Galles N, Werker JF. Visual language discrimination in infancy. Science. 2007;316:1159. doi: 10.1126/science.1137686. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Egner T, Hawks Z, Link J. The congruency sequence effect emerges when the distracter precedes the target. Acta Psychologica. 2015;156:8–21. doi: 10.1016/j.actpsy.2015.01.003. [DOI] [PubMed] [Google Scholar]
- Woldorff MG. Distortion of ERP averages due to overlap from temporally adjacent ERPs: Analysis and correction. Psychophysiology. 1993;30:98–119. doi: 10.1111/j.1469-8986.1993.tb03209.x. [DOI] [PubMed] [Google Scholar]
- Yeung N, Botvinick MM, Cohen JD. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 2004;111:931–959. doi: 10.1037/0033-295x.111.4.939. [DOI] [PubMed] [Google Scholar]
- Yeung N, Cohen JD. The impact of cognitive deficits on conflict monitoring: Predictable dissociations between the error-related negativity and N2. Psychological Science. 2006;17:164–171. doi: 10.1111/j.1467-9280.2006.01680.x. [DOI] [PubMed] [Google Scholar]