

Abstract
The mammalian hippocampus shows a remarkable capacity for continued neurogenesis throughout life. Newborn neurons, generated by the radial neural stem cells (NSCs), are important for learning and memory as well as mood control. During aging, the number and responses of NSCs to neurogenic stimuli diminish, leading to decreased neurogenesis and age-associated cognitive decline and psychiatric disorders. Thus, adult hippocampal neurogenesis has garnered significant interest because targeting it could be a novel potential therapeutic strategy for these disorders. However, if we are to use neurogenesis to halt or reverse hippocampal-related pathology, we need to understand better the core molecular machinery that governs NSC and their progeny. In this review, we summarize a wide variety of mouse models used in adult neurogenesis field, present their advantages and disadvantages based on specificity and efficiency of labeling of different cell types, and review their contribution to our understanding of the biology and the heterogeneity of different cell types found in adult neurogenic niches.
Keywords: adult neurogenesis, mouse models, neural stem cells, neuroprogenitors, lineage tracing
In the adult mammalian brain, two regions, the subgranular zone of the dentate gyrus of hippocampus (SGZ) and the subventricular zone (SVZ), harbor stem/progenitor cells that give rise to newborn neurons and astrocytes throughout life (Altman, 1962; Eriksson et al., 1998; Ming and Song, 2011; Morshead et al., 1994; Palmer et al., 1997; Spalding et al., 2013). The neuroprogenitor cells from the SVZ give rise to GABAergic interneurons of the olfactory bulb, responsible for odor discrimination and mating behavior (Doetsch et al., 1997, 1999; Gheusi et al., 2000; Mak et al., 2007; Mouret et al., 2009; Sakamoto et al., 2014). The neuroprogenitor cells from the SGZ give rise to glutamatergic granule cells, associated with hippocampal-dependent spatial learning and memory (Clelland et al., 2009; Deng et al., 2009; Farioli-Vecchioli et al., 2008; Imayoshi et al., 2008; Kitamura et al., 2009; Saxe et al., 2006; Shors et al., 2002; Zhang et al., 2008), mood control (Encinas et al., 2006; Jacobs et al., 2000; Malberg et al., 2000; Santarelli et al., 2003; Snyder et al., 2011), pattern separation (Aimone et al., 2011; Pan et al., 2012; Sahay and Hen, 2008; Sahay et al., 2011; Wojtowicz et al., 2008; Zhao et al., 2008), and stress response (Heine et al., 2005; Joels et al., 2007; Lucassen et al., 2015; Snyder et al., 2011; Surget et al., 2011). Adult neurogenesis is very-well established in the human hippocampus as well (Djuric et al., 2008; Eriksson et al., 1998; Manganas et al., 2007; Pereira et al., 2007; Quinones-Hinojosa et al., 2006; Sierra et al., 2011; Spalding et al., 2013), thus sparking interests with regards to the potential of utilizing adult neurogenesis for regenerative medicine and treatment of neuropsychiatric disorders (David et al., 2010; Mainen et al., 1999; Manganas and Maletic-Savatic, 2005; Abiega et al, 2016). If adult neurogenesis is to be used as a therapeutic modality for brain repair, it is very important to understand the unique properties of individual cell types that are responsible for this phenomenon. However, the ever-proliferating and differentiating spectrum of cells in both adult neurogenic niches poses a great challenge to study adult neurogenesis one cell type at a time.
Classification of different cell types in both adult neurogenic niches, from primary stem cells to their end-progeny, is hard. More and more it has become evident that these cells are heterogeneous populations with different lifespans, lineage plasticity and regenerative potential, that all depend on age (Alvarez-Buylla et al., 2008; Bonaguidi et al., 2012; Bond et al., 2015; Chojnacki et al., 2009; Giachino and Taylor, 2014). Criteria used to classify different cell types in both adult neurogenic niches depend not only on marker combinations but also on morphology of the cells. Although there is a consensus for establishing the main cell types of the adult neurogenic niches (Figures 1, 2), consensus for defining the neural stem/progenitor subtypes, lineages and some of their properties (self-renewal, cell cycle, quiescence) is still far away. Although experimental differences (e.g. timing and dose of tamoxifen, different cell cycle indicators, etc.) may account for some of the discrepancy, transgenic mouse lines are the primary suspect accounting for most of the observed differences: lineage tracing mouse models generated based on the same gene but utilizing different regulatory regions produce different experimental outcomes. A striking example is Nestin, the main powerhouse for generation of transgenic mouse lines. Nine different NestinCreERT2 lines have been designed using Nestin promoter; however, only a subset of them are capable of targeting neurogenic regions of the adult brain with significant differences in efficiency (Sun et al., 2014). As many transgenic mouse models have been developed for labeling and tracing of different cell types in the adult neurogenic niches (Table 1, Figures 1, 2), it is important to understand their advantages and disadvantages. Here, we review mouse models most widely used for studies of adult neural stem/progenitor cells and their lineage.
Figure 1. Summary of immunohistochemical markers, reporter and lineage tracing mouse models used to identify different cell types of the adult hippocampal neurogenic niche.
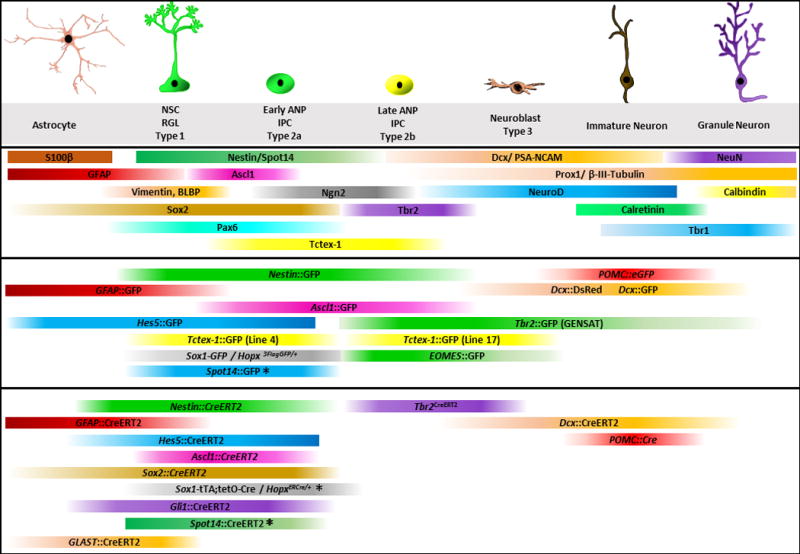
NSC, neural stem cells; RGL, radial glia like cells; ANP, amplifying neuroprogenitors; IPC, intermediate progenitor cells. Upper part: Markers, Middle part: Reporter lines; Lower part: Lineage tracing lines. * denotes the mouse models that target mostly quiescent neural stem cells
Figure 2. Summary of immunohistochemical markers, reporter and lineage tracing mouse models used to identify different cell types of the adult subventricular zone neurogenic niche.
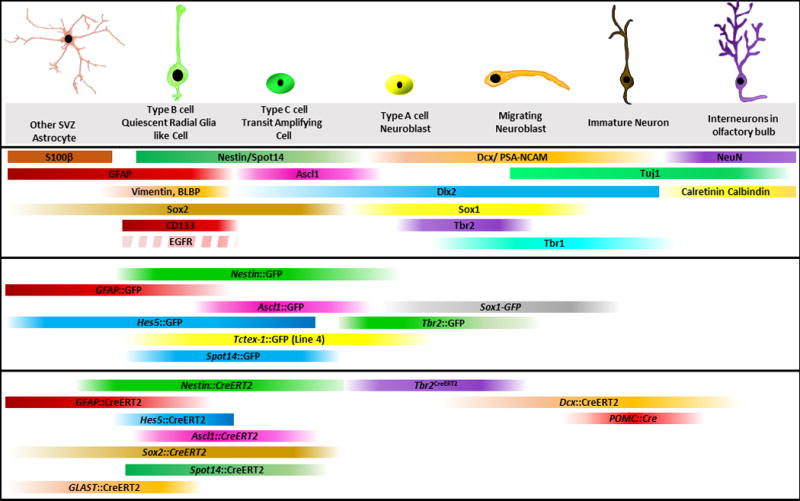
Broken strip (EGFR) indicates expression in only subset of Type B cells. EGFR+ population is actively cycling whereas EGFR− population consists of quiescent SVZ neural stem cells. SVZ=subventricular zone; RMS=rostral migratory stream; OB=olfactory bulb. Upper part: Markers, Middle part: Reporter lines; Lower part: Lineage tracing lines.
Table 1. List of reporter mice used in adult neurogenesis research.
NSC=neural stem cells, ANP = amplifying neuroprogenitors, NB=neuroblasts, IN=immature neurons, GZ=granule cells, SGZ=subgranular zone, SVZ=subventricular zone. When not specified, the mouse model is used for studies of both SGZ and SVZ neurogenesis.
| Line | Use | Reference: | |
|---|---|---|---|
| Nestin | Nestin∷lacZ | Characterization of regulatory regions of Nestin | (Johansson et al., 2002; Zimmerman et al., 1994) |
| Nestin∷CFPnuc | Quantification of NSCs and progenitor cells | (Encinas et al., 2006) | |
| Nestin∷GFP, Nestin∷eGFP | Population studies of NSCs and progenitor cells | (Kawaguchi et al., 2001; Mignone et al., 2004; Yamaguchi et al., 2000) | |
|
Nestin∷tTA, Nestin-rtTA, Nestin-rtTA-M2-eGFP Nestin-Cre, Nestin-CreER; Nestin-CreERT2 |
Lineage tracing | (Beech et al., 2004; Mitsuhashi et al., 2001; Yu et al., 2005) (Betz et al., 1996; Kuo et al., 2006); (Dranovsky et al., 2011; Imayoshi et al., 2006; Lagace et al., 2007) |
|
| GFAP |
GFAP∷GFP, GFAP∷eGFP GFAP∷AmCyan1, AsRed2, mRFP1 |
Population studies of astrocytes and NSCs | (Nolte et al., 2001; Platel et al., 2009; Zhuo et al., 1997); (Hirrlinger et al., 2005) |
| GFAP∷TK | Specific ablation of NSCs | (Garcia et al., 2004) | |
|
GFAP-Cre, GFAP-CreERT2 |
Lineage tracing of NSCs | (Casper and McCarthy, 2006; Ganat et al., 2006; Marino et al., 2000) | |
| Sox1 | Sox1-GFP | Population studies of a subset of NSCs and ANPs | (Aubert et al., 2003) |
| Sox1-tTA;tetO-Cre | Lineage tracing of NSCs and ANPs | (Venere et al., 2012) | |
| Sox2 | Sox2β-Geo | Strong activity in SVZ but weak in SGZ | (Zappone et al., 2000) |
|
Sox2∷GFP, Sox2-eGFP |
Population studies of NSCs, ANPs, and astrocytes | (D’Amour and Gage, 2003; Ellis et al., 2004) | |
| Sox2∷Cre, Sox2∷CreER, Sox2∷CreERT2 | Lineage tracing | (Favaro et al., 2009; Hayashi et al., 2007; Kang and Hebert, 2012) | |
| Ascl1 | Ascl1-GFPnuc | Strong expression in ANPs but weak in NSCs | (Leung et al., 2007) |
| Ascl1-CreERT2 | Lineage tracing in SGZ | (Kim et al., 2011) | |
| Glast |
Glast∷eGFP Glast∷dsRed |
Population studies of astrocytes and NSCs | (Gong et al., 2003) (Glowatzki et al., 2006; Regan et al., 2007) |
|
Glast-CreERT2; Glast∷CreERT2 |
Lineage tracing | (Mori et al., 2006); (Slezak et al., 2007) | |
| Blbp |
Blbp∷GFP, BLBP∷eGFP Blbp∷eYFP, Blbp∷dsRed2 Blbp∷mCherry |
Population studies of astrocytes and NSCs | (Anthony et al., 2004; Gong et al., 2003; Schmid et al., 2006); (Giachino et al., 2014) |
| Blbp∷Cre | Lineage tracing | (Anthony et al., 2004; Hegedus et al., 2007) | |
| Hes5 | Hes5∷GFP | Population studies of a subset of NSCs and ANPs | (Basak and Taylor, 2007) |
| Hes5∷CreERT2 | Lineage tracing | (Lugert et al., 2012) | |
| Tbr2 | EOMES∷GFP | Labeling of late ANPs and early NBs | (Kwon and Hadjantonakis, 2007) |
| Tbr2∷GFP | Labeling of late ANPs, NBs, INs and some GCs | (Gong et al., 2003) | |
|
EOMES-CreERT2 Tbr2∷CreERT2 |
Lineage tracing of late ANPs and NBs | (Pimeisl et al., 2013) (Berg et al., 2015) |
|
| Spot14 | Spot14∷GFP | Labeling of subset of NSCs and ANPs | (Knobloch et al., 2013; Knobloch et al., 2014) |
| Spot14∷CreERT2 | Lineage tracing | (Knobloch et al., 2013) | |
| Gli1 | Gli1∷CreERT2 | Lineage tracing of NSCs in SGZ | (Ahn and Joyner, 2004) |
| Prss56 | Prss56-CreERT2 | Lineage tracing of NSCs in SGZ | (Jourdon et al., 2015) |
| Hopx | Hopx-3FlagGFP/+ | Labeling of NSCs of SGZ | (Li et al., 2015) |
| Hopx-ERCre/+ | Lineage tracing of NSCs in SGZ | (Li et al., 2015) | |
| Tctex-1 |
Tctex-1∷GFP, Tctex-1∷DsRed |
Labeling of NSCs, ANPs and NBs of SGZ | (Tseng et al., 2010) |
| Pomc | Pomc∷eGFP | Labeling of NBs/INs | (Cowley et al., 2001) |
| Pomc-Cre | Lineage tracing of NBs/INs | (Balthasar et al., 2004) | |
| Dcx |
Dcx-eGFP Dcx-DsRed |
Labeling of NBs/Ins Labeling of NBs/Ins |
(Couillard-Despres et al., 2006; Walker et al., 2007); (Couillard-Despres et al., 2006; Wang et al., 2007) |
Nestin
Arguably the most popular mouse models to study adult neurogenesis use Nestin regulatory elements as drivers of reporter gene expression. Nestin was cloned by finding the target protein of Rat401 antibody (Hockfield and McKay, 1985), namely the antigen enriched in the proliferative zone of the neural tube. It was subsequently named based on its activation in neuroepithelial stem cells (Lendahl et al., 1990). Nestin is an intermediate filament protein expressed in many tissues in addition to neural tissue, including muscle (Day et al., 2007), retina (Mayer et al., 2003), testis (Lobo et al., 2004), hair follicle (Li et al., 2003), and others (Wiese et al., 2004). The first study that characterized the regulatory regions of the Nestin revealed the challenges of using these regulatory regions for transgenic approaches. Initially, two enhancer regions localized to the first and second intron were characterized (Zimmerman et al., 1994). Subsequently, it was discovered that the enhancer element in the second intron of the human Nestin contains TR (Thyroid hormone receptor), RXR (Retinoid X receptor), RAR (all-trans retinoic acid receptor), and COUP-TF (Chicken ovalbumin upstream promoter-transcription factor) binding motifs that control Nestin activation in the embryonic brain (Lothian et al., 1999). In addition, enhancers located in the second intron of the rat Nestin seemed to be active in a region-specific manner: one was active throughout the developing brain, while the other was specific for the ventral midbrain (Yaworsky and Kappen, 1999). Interestingly, in these early years of the Nestin characterization, no study reported the expression of nestin in the adult neurogenic regions (Lothian et al., 1999; Yaworsky and Kappen, 1999; Zimmerman et al., 1994). In 2000, Yamaguchi and colleagues generated the first Nestin∷GFP mouse and reported the expression of GFP in adult dentate gyrus, SVZ and rostral migratory stream (RMS) (Yamaguchi et al., 2000). Later, this mouse was used to demonstrate that neural stem cells (NSCs) in the SGZ shared a lot of features with astrocytes, such as the expression of glial fibrillary acidic protein (GFAP) and passive and non-inactivating currents with a linear current-voltage relationship (Filippov et al., 2003). The same group also thoroughly characterized the morphological properties of those NSCs, including the characteristic tree-like branching endings in the molecular layer of the dentate gyrus (Filippov et al., 2003).
The second mouse model that used Nestin as a driver of the fluorescent reporters was generated by Mignone and collaborators (Mignone et al., 2004). In their transgenic Nestin∷GFP mouse, Mignone and colleagues used the enhancer located in the second intron in addition to Nestin promoter. The GFP accurately marked both neurogenic regions of the adult brain and GFP+ cells efficiently formed neurospheres after sorting. In SVZ and RMS, Nestin∷GFP expression was localized to Type A, B, and C cells (Mignone et al., 2004), whereas within the dentate gyrus, it marked NSCs, amplifying neuroprogenitors (ANPs), as well as other non-progenitor cell types including pericytes, endothelial cells, and oligodendrocyte progenitors (Encinas et al., 2011). Although cytoplasmic GFP is good for visualization of the morphological features of the cells where it is expressed, it is not ideal for quantification because of the abundance of expressing cells, particularly in young animals. Thus, another mouse model was generated, using the cyan fluorescent protein (CFP) fused with the nuclear localization signal (Nestin∷CFPnuc transgenic mouse). In this mouse, it was easy to visualize the neurogenic areas as dotted patterns, which allowed the unambiguous enumeration of the nuclei of Nestin-expressing cells (Encinas et al., 2006). Thus, for quantitative studies of the SGZ neuroprogenitors, the Nestin∷CFPnuc is a valuable mouse model.
Soon after Nestin∷GFP mouse models were published, the first inducible Nestin mouse line was generated (Beech et al., 2004). It expressed tetracycline transactivator (tTA) under the control of a wide genomic region covering the 5.8kb upstream and 5.4 kb downstream of Nestin, and was then crossed with the TetOP-mCREB-FLAG mouse (Chen et al., 1998) to visualize the Nestin expressing cells and their progeny in the absence of doxycycline (tet-off). Using this mouse, it was demonstrated for the first time that Nestin expressing cells in the SVZ were capable of giving rise to periglomerular cells in the olfactory bulb (Beech et al., 2004). Surprisingly, granule cells in the olfactory bulb, another type of interneurons known to be sourced from the SVZ (Lois and Alvarez-Buylla, 1994; Luskin, 1993), were not labeled in this mouse model. This study was thus important because it pointed to two observations that the field still continues to tease out: 1) NSC population in the SVZ is heterogeneous; and 2) correct labeling of the stem/progenitor cell population is not enough to ensure that the lineage tracing will proceed as expected.
Several other transgenic mice using tetracycline responsive element were generated. Yu and collaborators developed a line with eGFP sequence upstream of the rtTA element (Nestin-rtTA-M2-eGFP) and crossed it with TetOP-Cre (Perl et al., 2002) and Rosa26-LacZ (Soriano, 1999) to demonstrate the temporal induction of Cre in neurogenic regions (Yu et al., 2005). Although eGFP+ cells have been observed in the SGZ and SVZ, no lineage tracing data in the adult mice have been reported using this mouse model. Dupret and collaborators developed a tet-on model using the second intron of rat Nestin (Dupret et al., 2008). In this mouse, GFP was seen only in the SGZ but not SVZ, suggesting that Nestin dependent transgenic reporter and inducible lines have considerable variations in their targets, depending on the genomic element used to drive the expression of transgene. Another Nestin∷CreERT2 line, expressing CreERT2 under the control of neural specific Nestin enhancer (Zimmerman et al., 1994), was used to show that Smoothened mediated Hedgehog signaling is important for SVZ neurogenesis (Balordi and Fishell, 2007). Later, the same inducible line was used in two other studies that reached controversial conclusions. Encinas and collaborators performed a comprehensive double labeling study with two BrdU analogs, CldU and IdU, and developed an algorithm that estimated that SGZ NSCs, after entering into cell cycle, repeatedly divide three times and then terminally differentiate into astrocytes, suggesting that these NSCs are disposable (Encinas et al., 2011). On the other hand, Bonaguidi and collaborators used an induction protocol to activate Cre in sparse, individual NSCs and traced their respective lineages. They concluded that SGZ NSCs have the ability to both self-renew and give rise to multilineage progeny (Bonaguidi et al., 2011). These two disparate findings then led Dranovsky and collaborators to generate yet another Nestin∷CreERT2 mouse, based on the pNerv-SXN vector, where brain-specific enhancer of Nestin located in the second intron (Josephson et al., 1998) drives the expression of CreERT2 (Dranovsky et al., 2011). They showed that NSCs have the ability to produce expanding and persisting populations of both neurons and stem cells, but that this effect depends mostly on the external experiences of the animal. Social isolation and X-ray irradiation seemed to facilitate accumulation of NSCs while enriched environment shifted the balance in the favor of newborn neuron production (Dranovsky et al., 2011).
Nestin-dependent inducible lines have thus been invaluable for deciphering mechanisms underlying the complex nature of adult neurogenic niches, at the same time raising controversial questions. This is not surprising, as recent report suggests (Sun et al., 2014). Namely, all these lines show variable stem cell labeling efficiency and sometimes suffer from leakage to other cell types, which might all contribute to different findings despite the use of the “same” transgenic line. This is important to remember when authenticating each line, as strain, age, promoter, induction protocol used and other factors may affect the outcome of the experiment.
GFAP
One of the indelible markers commonly used to identify adult NSCs of the SGZ is glial fibrillary acidic protein (GFAP). GFAP labels radial processes that originate from the apical surface of the NSCs and span throughout the granular cell layer. It is not expressed in amplifying neuroprogenitors (ANPs) and this distinct expression pattern has been invaluable to differentiate between these two types of proliferating cells.
GFAP expression in the NSCs turned out to be critical for their characterization and naming. Cameron and colleagues showed for the first time that [3H]thymidine+ cells in the adult rat SGZ contained GFAP+ cells resembling radial glia, with triangular cell bodies and a thin radial process (Cameron et al., 1993). These were SGZ primary NSCs, and “radial glia-like” cells is still in use to name these primary SGZ stem cells. Despite the fact that GFAP+ cells in the SGZ were shown to proliferate, the authors of the first GFAP∷GFP mouse (Zhuo et al., 1997) did not report on the GFP expression in the dentate gyrus. It took several years to begin to utilize this line for studies of NSCs, but eventually it found the application particularly for studies of the SVZ neurogenic niche: it was used to describe its vascular nature (Shen et al., 2008; Tavazoie et al., 2008) and to isolate and study both activated (Pastrana et al., 2009) and quiescent progenitors (Codega et al., 2014). Another GFAP reporter line, based on the human promoter, has been generated (Nolte et al., 2001) and although it has been extensively used in the astrocyte field, it has not been very popular for studies of the adult neurogenesis (Beckervordersandforth et al., 2014; Sultan et al., 2013).
First direct proof of “astrocytes” giving rise to neurons came in 2001. With the help of BrdU labeling and retroviral lineage tracing, Seri and colleagues found that GFAP+ “astrocytes” in the SGZ were capable of proliferating in vivo and generating neurons (Seri et al., 2001). Using GFAP∷GFP adenovirus, the same group defined the morphology of NSCs (Seri et al., 2004), and later that same year, a new line, GFAP∷TK (thymidine kinase), was generated to transgenically ablate the dividing GFAP+ progenitors (Garcia et al., 2004). This seminal study demonstrated that GFAP+ progenitors are the principal source of adult neurons in both SVZ and SGZ. To further prove this observation, an inducible Cre recombinase (CreERT2) controlled by the human GFAP promoter was used for lineage tracing of GFAP+ cells and confirmed that those cells can give rise to neuronal precursors and neurons (Ganat et al., 2006). The same line was subsequently used to conditionally ablate Notch1 in NSCs and their progeny, revealing the role of Notch signaling in regulation of the cell fate and dendrite morphology of newborn neurons (Breunig et al., 2007).
Although human GFAP promoter has been useful in the generation of mouse models that allowed the studies of NSCs, abundant GFAP expression in astrocytes has generated a major problem for purification and lineage tracing of NSCs. To overcome this problem, a split-Cre approach, in which two lentiviral constructs (one containing the C-terminal of the Cre driven by Prominin1 p2 promoter and one containing the N-terminal of the Cre driven by human GFAP promoter) have been used to target specifically the SGZ NSCs (Beckervordersandforth et al., 2014). Immunofluorescence against h-GFAP∷GFP and Prominin1 showed that the combinatorial approach labeled NSCs with radial morphology; however these represented only 30% of the labeled cells.
Despite being expressed in astrocytes, abundantly present in both adult neurogenic niches, GFAP based models have been useful for adult neurogenic studies. For isolation of SVZ NSCs, lack of specificity of GFAP∷GFP has been compensated by EGFR and/or CD133 as additional markers for NSC purification (Codega et al., 2014). For lineage tracing, in comparison to Nestin based models, GFAP based models have the advantage of avoiding the initial activation of Cre in ANPs. Thus, this enables lineage tracing directly from the origin, the NSC. However, these studies suffer from estimating the contribution of astrocytes to the final lineage outcome, and consequently, data interpretation and particularly quantitative studies of potential transformation of NSCs into astrocytes are jeopardized.
Sox proteins
Sox (sex determining region Y box family) proteins that bind to DNA via their high-mobility group (HMG) domains are conserved transcription factors that take role in cell fate specification and differentiation of many tissues (Abraham et al., 2013a; Abraham et al., 2013b; Kamachi and Kondoh, 2013). The most well-known member of this family, Sox2, has been characterized as one of the Yamanaka factors required for generation of induced pluripotent stem cells (Takahashi and Yamanaka, 2006). Sox2 is required for embryonic stem cell pluripotency and thus, it is one of the earliest known genes to be activated in the vertebrate neural tube (Uwanogho et al., 1995). In addition, it is also essential for maintenance of many adult tissue resident stem cells (Arnold et al., 2011), including both SVZ and SGZ NSCs (Ferri et al., 2004). The first studies of adult neurogenic niches using Sox2 expression came from Sox2β-Geo knock-in mice (Zappone et al., 2000). Ferri and colleagues showed that X-gal staining was localized to mainly SGZ of the dentate gyrus and the lateral ventricle walls, as well as that Sox2 was expressed in BrdU+ GFAP+ NSCs of both SGZ and SVZ niches. Moreover, in Sox2ΔENH mice, in which neural specific enhancer of Sox2 is deleted, there was a dramatic reduction in the number of proliferating cells in both niches, indicating the specificity of Sox2 expression in neural stem and progenitor cells (Ferri et al., 2004). Further characterization of Sox2 regulatory regions revealed that another enhancer element, Sox2 regulatory region2 (SRR2) that spans +3300 to +4124bp respective to the translation initiation codon, is able to discriminate between SVZ and SGZ. Sox2∷β-geo transgenic line, which utilized SRR2 element to drive β-geo reporter, showed strong expression pattern in the SVZ and rostral migratory stream, but showed a weak labeling of cells in the SGZ (Miyagi et al., 2006). This report indicates that the regulatory regions governing Sox2 expression are very important to consider when choosing the best mouse model for a given experiment.
First Sox2∷GFP transgenic mice were generated in 2003 (D’Amour and Gage, 2003) and have been used to characterize the Sox2+ cells in the hippocampal niche (Suh et al., 2007). Three types of cells were expressing Sox2∷GFP in the dentate gyrus, NSCs, ANPs, and astrocytes. The same group also generated Sox2∷Cre-GFP retro and lenti viruses to show that Sox2+ cells can give rise to neurons, astrocytes and other cells that retain Sox2 expression. Thus, this study was the first to hint at the self-renewal ability of Sox2+ cells in the SGZ (Suh et al., 2007). Finally, conditional removal of Sox2 was achieved in Sox2∷CreERT2 line (Favaro et al., 2009), which utilized the telencephalic specific enhancer-promoter region (Zappone et al., 2000). In the embryonic brain, this removal resulted in aberrant dentate gyrus (Favaro et al., 2009). In the adult brain, it led to significant loss of hippocampal NSCs. These effects were attributed to Shh pathway, as Sox2 is required for transcription of Shh (Favaro et al., 2009).
Sox1, another well-known member of Sox family, is one of the earliest genes to be expressed in the ectodermal cells that are committed to neural fate (Collignon, 1992). Its expression correlates with the dividing neural precursors of the embryonic central nervous system but not with the differentiation along the dorso-ventral axis (Pevny et al., 1998). First Sox1GFP mouse, a knock-in construct that replaced Sox1 locus with GFP, was used to identify the neural genes from embryonic neuroepithelial cells. Interestingly, GFP was also localized in SGZ cells that resembled NSCs (Aubert et al., 2003). Characterization of these Sox1GFP+ cells in SGZ revealed that 35% of the population had GFAP+ radial processes (and thus were most likely NSCs), another 30% were GFAP− (most likely ANPs) and the remaining 35% were uncharacterized cell type(s). Despite labeling quite a bit of NSCs and ANPs, Sox1GFP is expressed in only 40% the total population of NSCs and ANPs, suggesting that it labels only a subset of them (Venere et al., 2012). By generating a Sox1 driven tet-on dependent lineage tracing line (Sox1-tTA;tetO-Cre), Venere and colleagues were able to show that Sox1 expressing cells were able to generate granule neurons and hilar astrocytes. In SVZ however, Sox1GFP+ cells mostly co-localized with PSA-NCAM+ late progenitors/neuroblasts and not with early progenitors (Venere et al., 2012), arguing that the role of this gene depends on the neurogenic niche.
ASCL1 (Mash1)
Achaete-Scute Family BHLH Transcription Factor 1 (Ascl1) or Mash1 (mammalian Ascl1) is a member of the basic helix loop helix family of transcription factors. It is one of the key members of the proneural transcription factors, and it is necessary and sufficient to promote neurogenesis (Bertrand et al., 2002). Ascl1 sequentially activates target genes by increasing the accessibility of target genes both in proliferating and differentiating neuroprogenitors (Raposo et al., 2015). Ascl1 has also been extensively used to reprogram somatic cells to induce formation of neuroprogenitors and neurons (Berninger et al., 2007; Karow et al., 2012; Pollak et al., 2013; Vierbuchen et al., 2010). Interestingly, retrovirus-mediated overexpression of Ascl1 in the adult SGZ NSCs resulted in generation of oligodendrocytic lineage at the expense of newborn granule neurons, suggesting the complex nature of differentiation cascade governed by Ascl1 (Jessberger et al., 2008).
The first GFP reporter mouse model, Ascl1GFPnuc knock-in, replaced one allele of Ascl1 coding sequence with the nuclear localized GFP (Leung et al., 2007). Two seminal studies were published using this mouse model: Leung and collaborators demonstrated that the horizontal basal cells are the stem cell population of olfactory neuroepithelium (Leung et al., 2007), while Kim and collaborators showed that Ascl1 expression was limited to transient ANPs in both SVZ and SGZ (Kim et al., 2007). Later, weak expression pattern of the Ascl1 was reported in the subset of the SGZ NSCs, using an inducible Ascl1CreERT2 knock-in mouse (Kim et al., 2011). Namely, after a short term induction (1 week), labeled cells included Type 1 cells (NSCs) in the SGZ and a subset of GFAP+ progenitors in the SVZ (Kim et al., 2011). Lineage tracing indicated that Ascl1 expressing cells were able to generate only newborn neurons and not astrocytes, supporting the observation that Ascl1 labeled mostly ANPs. Compared to other lineage tracing lines, this particular transgenic mouse model provided superbly clean labeling of neuronal progeny, demonstrating almost complete repopulation of the granule cell layer three months following the induction (Yang et al., 2015). Thus, this mouse model may become invaluable for quantitative studies of intermediate and late stages of neurogenesis as well as of newborn neuron integration.
GLAST
GLAST1 (Glial High affinity glutamate transporter) or SLC1A3 (solute carrier family 1, member3) gene encodes an astrocyte specific high affinity glutamate transporter, which buffers excess of extracellular glutamate from the tissue. Early in situ hybridization data showed that GLAST mRNA was localized in the ventricular zone of the fetal brain (Shibata et al., 1996; Sutherland et al., 1996) and radial glia cells in the developing spinal cord (Shibata et al., 1997). Characterization of radial glia throughout embryonic development suggested that not all radial glia in the embryonic brain express GLAST, but that GLAST+ cells consist of functionally distinct subsets of precursor cells with different cell cycle characteristics (Hartfuss et al., 2001). In addition, neurospheres generated from the subependymal zone of adult mice contained GLAST+ progenitor cells (Hartfuss et al., 2001). Moreover, GLAST was expressed in the human fetal telencephalon radial glia cells too (Zecevic, 2004).
Early postnatal data on the expression of GLAST in BrdU+ and Ki67+ cells of dentate gyrus showed that majority of the proliferating cells were GLAST+ and S100β+ (Namba et al., 2005). This suggested that GLAST expression is not only specific for embryonic radial glia but also present in the adult neurogenic regions, which held in many species including mouse, rat, cat, marmoset, and pig (Williams et al., 2005).
In 2006, first inducible line for GLAST (Glast-CreERT2) targets many of the astrocytes and NSCs of both dentate gyrus and SVZ that give rise to granule neurons in olfactory bulb (Mori et al., 2006). Later, this inducible line was used to detail the contribution of GLAST and Nestin lineages to adult hippocampal neurogenesis under different conditions (DeCarolis et al., 2013). In this study, authors showed that although both Nestin and GLAST lineages gave rise to granule neurons, contribution of the latter seemed to continuously increase over time while contribution of the former reached a plateau. The difference between the contributions of two lineages to neurogenesis became more apparent when animals were exposed to experimental manipulations, either ablation (by anti-mitotic drug, AraC) or stimulation (running) of neuroprogenitors. After either experimental condition, labeled NSCs in Glast-CreERT2 line appeared to contribute to neurogenesis, while those labeled in Nestin-CreERT2 did not (DeCarolis et al., 2013). Further characterization of Nestin∷GFP mice showed that, although GLAST+ GFAP+ radial cells were mostly Nestin∷GFP+, some GLAST+ GFAP+ radial cells were Nestin∷GFP−, suggesting that not all cells with radial morphology express Nestin∷GFP. These data emphasized that careful reevaluation of the conventional morphology/marker based classification of the cell types, especially for NSCs, in adult neurogenic niches is a necessity. GLASTCreERT2 line thus provides undoubtedly critical tool for understanding the heterogeneity of the NSC population.
Hes5
Notch signaling is an evolutionarily conserved pathway (Andersson et al., 2011) that plays a key role in development, through diverse effects on survival, differentiation and proliferation (Alunni et al., 2013; Breunig et al., 2007; Giachino and Taylor, 2014), that depend on signal strength (Chapouton et al., 2010; Gama-Norton et al., 2015; Ninov et al., 2012; Shimojo et al., 2008) and cellular context (Basak et al., 2012; Farnsworth et al., 2015; Lugert et al., 2010). Hairy and enhancer of split (Hes) genes are downstream targets of Notch pathway. They encode transcriptional repressors that predominantly control the proneural basic helix-loop-helix genes and thus regulate the maintenance of undifferentiated cells (Kageyama and Ohtsuka, 1999; Ohtsuka et al., 2001).
Hes5 starts to be expressed at E8.5 in the midbrain/hindbrain mouse region and continues to be expressed exclusively in the developing nervous system (Akazawa et al., 1992; Hatakeyama et al., 2004; Ohtsuka et al., 2001). First reporter mouse, Hes5∷GFP, has been developed by Basak and colleagues and replicated in situ data (Basak and Taylor, 2007). Hes5∷GFP expression was completely abolished in Notch1 deficient E9.5 embryos, suggesting that Hes5∷GFP can also be used to report Notch1 activity in vivo (Basak and Taylor, 2007). Further characterization of this mouse in SGZ revealed that Hes5∷GFP is expressed in cells that have both radial (54%) and horizontal (46%) morphologies. However, only 60% of Hes5∷GFP+ cells overlapped with Sox2, BLBP, and GFAP, indicating that Hes5∷GFP labels only a subset of the NSC population (Lugert et al., 2010). This is a very important finding, which suggests that either only some NSCs have active Notch signaling or that dynamics of Hes5∷GFP expression reflects potential oscillations in Notch signaling, labeling only the NSCs with high Notch activity. Given that Hes5∷GFP+ cells consists of both quiescent and actively dividing cells that respond selectively to physiological (running) and pathological (kainic acid induced seizures) stimuli, different levels of Notch in these cells is a plausible explanation. Generation of Hes5∷CreERT2 line enabled further characterization of Hes5+ stem/progenitor cell population and their lineage (Lugert et al., 2012). The Hes5∷CreERT2 line provided new data on the proliferation dynamics of different ANP subtypes. It has been accepted that Ascl1high Type2a cells are the driving force of proliferation in the SGZ niche. In contrary, Lugert and colleagues argued that Tbr2+ Type2b cells that originated from Ascl1high Type2a cells are highly mitotic and responsible for the expansion of the precursor pool before further differentiating into post-mitotic neuroblasts (Lugert et al., 2012). Collectively, these data suggested that Notch signaling might be one of the candidate factors that contributes to the heterogeneity of NSCs. Thus, utilization of Hes5∷CreERT2 and Hes5∷GFP mice with different conditional alleles of Notch pathway elements will be useful for understanding the Notch dependent and potentially Notch independent nature of NSCs.
Tbr2 (EOMES)
Eomesodermin (Eomes, Tbr2) is a member of T-box family and crucial for embryonic development. It is expressed in mouse oocytes and also detected in the pre-implantation embryos by in situ hybridization (McConnell et al., 2005). This observation was verified in EOMES∷GFP mouse, where it was also shown that eomesodermin was expressed in trophectoderm, primitive streak and telencephalon (Kwon and Hadjantonakis, 2007). In the developing neocortex, Tbr2 expression is associated with downregulation of Pax6 and transitioning of radial glia to intermediate progenitor cell stage (Englund et al., 2005). Similarly, in the adult hippocampus, Tbr2 is specific for intermediate progenitors in the SGZ (Hodge et al., 2008). There, Tbr2+ cells represent a highly heterogeneous population: they colocalize with PSA-NCAM, Dcx, Sox2 (weak overlap), Ascl1 (weak overlap), Pax6, NeuroD1, and proliferation marker PCNA, but not GFAP. This expression pattern thus indicates that Tbr2 is present in a wide spectrum of the differentiating cells, from ANPs to neuroblasts (Hodge et al., 2008).
On the other hand, another transgenic reporter mouse, Tbr2∷GFP, generated by GENSAT (Gong et al., 2003) suffered from substantial leakage to other cell types (Hodge et al., 2008). Although all Tbr2+ cells were Tbr2∷GFP+, they only constituted 30% of the Tbr2∷GFP+ cells. GFP was expressed in immature neurons, colocalizing with calretinin, and in granule cell, colocalizing with NeuN (Hodge et al., 2008). The widely different expression of GFP in two transgenic mice, EOMES∷GFP and Tbr2∷GFP, indicates the variability most likely due to the use of BAC clones and prompts careful authentication of such lines to avoid misinterpretation of data. In the SVZ, Tbr2+ cells were detected in the dorsal subregion of subependymal zone and in the rostral migratory stream. Short term lineage tracing revealed that Tbr2+ cells gave rise to glutamergic olfactory bulb interneurons (Brill et al., 2009). In the SGZ, the neurogenic potential of Tbr2+ cells was demonstrated in the first inducible knock-in line, EOMESCreER, in which Tbr2+ cells gave rise to Dcx+ immature neurons (Pimeisl et al., 2013). Although neuronal lineage of Tbr2+ cells is a well-accepted finding in the neurogenesis field, proliferation potential of these cells is still debated (Berg et al., 2015; Lugert et al., 2012).
Another Tbr2 dependent lineage tracing mouse line, Tbr2∷CreERT2, was used to show that Tbr2+ cells do not undergo significant amplification, but rather differentiate and generate 1.2 immature neurons per cell by 15 days following induction (Berg et al., 2015). As Cre is expressed mostly in homogenous population of unipotent Type2b cells that develop into immature neurons, but not into astrocytes or other cell types, Tbr2∷CreERT2 is a unique mouse model that can find applications for studies of newborn neurons with similar birthdays.
Spot14 (Thrsp)
SPOT14, or Thyroid hormone responsive protein (Thrsp), is involved in de novo lipogenesis and mainly expressed in the lipogenic tissues such as liver, fat, and lactating mammary gland (Jump and Oppenheimer, 1985). It regulates the activity of fatty acid synthase (Fasn), the key enzyme in biosynthesis of fatty acids, by reducing the availability of its substrate, malonyl-CoA. Lipid metabolism and signaling has been highly implicated in the function of stem cells, including neural stem cells (Allen and Maletic-Savatic, 2011; Allen et al., 2013; Arnold et al., 2015; Maletic-Savatic et al., 2008; Manganas et al., 2007; Walker et al., 2016). In 2013, Knobloch and colleagues showed that Fasn is highly expressed in the adult neural stem/progenitor cells of both SGZ and SVZ but downregulated after differentiation (Knobloch et al., 2013). SPOT-14∷GFP reporter mouse showed that Spot14+ cells have radial (43.1%) and non-radial morphology (56.9%), and that GFP expression overlapped with Sox2, GFAP and Nestin. Only 1.7% of the radial population and 7.8% of the non-radial population were Ki67+, indicating that Spot14+ cells are mostly quiescent (Knobloch et al., 2013). Long term (1 and 3 months) lineage analysis in Spot14∷CreERT2 mouse confirmed that Spot14+ cells constitute neurogenic stem/progenitor cells in the adult brain (Knobloch et al., 2013). Further characterization of Spot14+ population revealed that their numbers decline with aging, and that they responded to both positive (running) and negative (temozolomide) neurogenic regulators (Knobloch et al., 2014). Since Spot14 has been shown to be responsible for regulating the quiescence of stem cells via limiting lipid synthesis (Knobloch et al., 2013), transgenic lines that depend on the regulatory regions of Spot14 would be useful for future studies aimed at investigations of the metabolic aspects of the stem cell quiescence.
Gli1
Sonic hedgehog (Shh) pathway is important for adult neurogenesis (Lai et al., 2003; Machold et al., 2003). Gli1 (GLI-Krupperl family member 1) has been used as a sensitive readout of the Shh activity. Gli1 promoter was thus used to drive the expression of CreERT2 in the Gli1CreERT2 line (Ahn and Joyner, 2004), in order to study the role of Shh in SGZ NSCs. Indeed, Shh-responsive NSCs had the ability to self-renew as they expanded in number and generated neurons (Ahn and Joyner, 2005). Encinas and collaborators crossed Gli1CreERT2 with the RCE (Rosa26/CMV-loxP-stop-loxP-GFP) reporter line (Balordi and Fishell, 2007) to drive the expression of GFP exclusively in the SGZ NSCs (Encinas et al., 2011).
Other mouse models
Due to limitations of aforementioned mouse models either for specificity or efficiency of labeling of neural stem/progenitor cells, groups all over the world took different approaches to generate other valuable mouse models. Among those, recently developed Prss56CreERT2 line stands out, as it provides labeling of NSCs and respective lineages not only in the adult SGZ and SVZ, but also in the hypothalamus ventricular zone (Jourdon et al., 2015). Further, Hopx, atypical homeodomain only protein, previously reported to be expressed in the stem cell populations of intestine (Takeda et al., 2011) and hair follicle (Takeda et al., 2013), is also a potential marker of SGZ but not SVZ stem/progenitor cells (Li et al., 2015). In Hopx3FlagGFP/+ line, GFP expression localized to a subset of Sox2+ cells of the SGZ as well as GFAP+, Nestin+ and BrdU retaining cells. HopxERCre/+ lineage tracing showed that Hopx expressing cells consist of almost exclusively quiescent NSCs (Shin et al., 2015), which have the ability to give rise to neuronal lineage in the dentate gyrus but not in the olfactory bulb (Li et al., 2015). Thu, this unique mouse model may provide key knowledge on the biology of quiescence. Tctex-1, dynein light chain, is expressed in the neurogenic regions of the adult brain (Chuang et al., 2001; Dedesma et al., 2006). Further characterization of Tctex-1 expressing cells showed that they are mostly ANPs and neuroblasts but not NSCs (Tseng et al., 2010). On the contrary, two different Tctex-1∷GFP lines generated in the same study showed conflicting outcomes: in one line (Line 4), GFP was expressed in NSCs and early ANPs while in the other line (Line 17), it was expressed in late ANPs and neuroblasts. This variable expression patterns even in transgenic models that use the same constructs again prompts full authentication of the mouse models, prior to start of any experimentation.
Albeit we focused here mainly on the mouse models used to study neural stem and progenitor cells, various other models have been developed to study newborn neurons (Figures 1, 2). Among those, Pomc-based (Pomc∷eGFP (Cowley et al., 2001), Pomc-Cre (Balthasar et al., 2004)) and Dcx-based (two Dcx-eGFP (Couillard-Despres et al., 2006; Walker et al., 2007), and two Dcx-DsRed (Couillard-Despres et al., 2006; Wang et al., 2007)) lines stand out. Those models have been instrumental for our understanding of the differentiation and integration of neuroblasts and granule neurons (Hunt et al., 2012; McHugh et al., 2007; Nakashiba et al., 2012) as well as for distinguishing the NSC population and their progeny from each other (Bracko et al., 2012). However, complete review of these mouse models is beyond the scope of this review.
CONCLUSION
Despite its early discovery (Altman, 1962), adult mammalian neurogenesis has gained recognition only in the early 1990s, but since then, there has been a substantial increase in the number of reporter and inducible mouse lines to study this phenomenon. Finding the right genes and their regulatory regions to drive the expression of fluorescent reporters or the Cre variants with optimum specificity and efficiency has been the major challenge. Thus, characterization of enhancer elements is very important to increase the specificity and efficiency of the current models. Finding new markers for different cell types is also important for not only developing new tools but also for understanding the stem/progenitor cell heterogeneity. Single-cell RNA sequencing studies offer great opportunities for finding new markers for different cell types. A recent study has demonstrated the utility of the single cell RNA sequencing for delineating the transcriptional profiles of NSCs along their developmental trajectory (Shin et al., 2015).
Most of the transgenic models that we summarized here use GFP or eGFP as their reporters. Although stability and signal intensity of GFP is an advantage, sometimes it poses a challenge. Tbr2∷GFP is a good example: while Tbr2 is expressed in ANPs, GFP overlaps with calretinin and even with NeuN, and is thus found in neuroblasts and neurons in the dentate gyrus (Hodge et al., 2008). Using destabilized GFP protein fused with the PEST sequence, with a half-life of 2hr (Li et al., 1998) might be an alternative strategy to limit the leakage of the reporter gene to progeny. Developing additional reporter mouse lines that utilize other parts of the spectrum, such as Nestin∷CFPnuc (Encinas et al., 2006), and BLBP∷mCherry (Giachino et al., 2014) would be extremely useful for the validation of our current marker paradigm and also for understanding the cellular heterogeneity of stem/progenitor cell population.
Acknowledgments
We thank the members of Maletic-Savatic lab for comments and critical reading of the paper. This project was supported by the Dana Foundation, the McKnight Endowment for Science, the CPRIT grant (RP130573CPRIT), and the Nancy Chang Award for Research Excellence (M.M.S.). The research was also supported in part by the Baylor College of Medicine Microscopy Core (P30HD024064 Intellectual and Developmental Disabilities Research Grant from the Eunice Kennedy Shriver National Institute of Child Health and Human Development). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
Conflict of interest
Fatih Semerci and Mirjana Maletić-Savatić declare no conflicts of interest.
Human and animal rights, and informed consent
This manuscript is a review article and does not involve a research protocol requiring approval by the relevant institutional review board or ethics committee.
References
- Abiega O, Beccari S, Diaz-Aparicio I, Nadjar A, Layé S, Leyrolle Q, Gómez-Nicola D, Domercq M, Pérez-Samartín A, Sánchez-Zafra V, Paris I, Valero J, Savage JC, Hui CW, Tremblay MÈ, Deudero JJ, Brewster AL, Anderson AE, Zaldumbide L, Galbarriatu L, Marinas A, Vivanco Md, Matute C, Maletic-Savatic M, Encinas JM, Sierra A. Neuronal Hyperactivity Disturbs ATP Microgradients, Impairs Microglial Motility, and Reduces Phagocytic Receptor Expression Triggering Apoptosis/Microglial Phagocytosis Uncoupling. PLoS Biol. 2016 May 26;14(5) doi: 10.1371/journal.pbio.1002466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abraham AB, Bronstein R, Chen EI, Koller A, Ronfani L, Maletic-Savatic M, Tsirka SE. Members of the high mobility group B protein family are dynamically expressed in embryonic neural stem cells. Proteome Sci. 2013a;11:18. doi: 10.1186/1477-5956-11-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abraham AB, Bronstein R, Reddy AS, Maletic-Savatic M, Aguirre A, Tsirka SE. Aberrant Neural Stem Cell Proliferation and Increased Adult Neurogenesis in Mice Lacking Chromatin Protein HMGB2. PLoS One. 2013b;8:e84838. doi: 10.1371/journal.pone.0084838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahn S, Joyner AL. Dynamic changes in the response of cells to positive hedgehog signaling during mouse limb patterning. Cell. 2004;118:505–516. doi: 10.1016/j.cell.2004.07.023. [DOI] [PubMed] [Google Scholar]
- Ahn S, Joyner AL. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature. 2005;437:894–897. doi: 10.1038/nature03994. [DOI] [PubMed] [Google Scholar]
- Aimone JB, Deng W, Gage FH. Resolving new memories: a critical look at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron. 2011;70:589–596. doi: 10.1016/j.neuron.2011.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akazawa C, Sasai Y, Nakanishi S, Kageyama R. Molecular characterization of a rat negative regulator with a basic helix-loop-helix structure predominantly expressed in the developing nervous system. J Biol Chem. 1992;267:21879–21885. [PubMed] [Google Scholar]
- Allen GI, Maletic-Savatic M. Sparse non-negative generalized PCA with applications to metabolomics. Bioinformatics. 2011;27:3029–3035. doi: 10.1093/bioinformatics/btr522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen GI, Peterson C, Vannucci M, Maletic-Savatic M. Regularized Partial Least Squares with an Application to NMR Spectroscopy. Stat Anal Data Min. 2013;6:302–314. doi: 10.1002/sam.11169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altman J. Are new neurons formed in the brains of adult mammals? Science. 1962;135:1127–1128. doi: 10.1126/science.135.3509.1127. [DOI] [PubMed] [Google Scholar]
- Alunni A, Krecsmarik M, Bosco A, Galant S, Pan L, Moens CB, Bally-Cuif L. Notch3 signaling gates cell cycle entry and limits neural stem cell amplification in the adult pallium. Development. 2013;140:3335–3347. doi: 10.1242/dev.095018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez-Buylla A, Kohwi M, Nguyen TM, Merkle FT. The heterogeneity of adult neural stem cells and the emerging complexity of their niche. Cold Spring Harb Symp Quant Biol. 2008;73:357–365. doi: 10.1101/sqb.2008.73.019. [DOI] [PubMed] [Google Scholar]
- Andersson ER, Sandberg R, Lendahl U. Notch signaling: simplicity in design, versatility in function. Development. 2011;138:3593–3612. doi: 10.1242/dev.063610. [DOI] [PubMed] [Google Scholar]
- Anthony TE, Klein C, Fishell G, Heintz N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 2004;41:881–890. doi: 10.1016/s0896-6273(04)00140-0. [DOI] [PubMed] [Google Scholar]
- Arnold JM, Choi WT, Sreekumar A, Maletic-Savatic M. Analytical strategies for studying stem cell metabolism. Front Biol (Beijing) 2015;10:141–153. doi: 10.1007/s11515-015-1357-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold K, Sarkar A, Yram MA, Polo JM, Bronson R, Sengupta S, Seandel M, Geijsen N, Hochedlinger K. Sox2(+) adult stem and progenitor cells are important for tissue regeneration and survival of mice. Cell Stem Cell. 2011;9:317–329. doi: 10.1016/j.stem.2011.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubert J, Stavridis MP, Tweedie S, O’Reilly M, Vierlinger K, Li M, Ghazal P, Pratt T, Mason JO, Roy D, et al. Screening for mammalian neural genes via fluorescence-activated cell sorter purification of neural precursors from Sox1-gfp knock-in mice. Proc Natl Acad Sci U S A. 2003;100(Suppl 1):11836–11841. doi: 10.1073/pnas.1734197100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balordi F, Fishell G. Mosaic removal of hedgehog signaling in the adult SVZ reveals that the residual wild-type stem cells have a limited capacity for self-renewal. J Neurosci. 2007;27:14248–14259. doi: 10.1523/JNEUROSCI.4531-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balthasar N, Coppari R, McMinn J, Liu SM, Lee CE, Tang V, Kenny CD, McGovern RA, Chua SC, Jr, Elmquist JK, et al. Leptin receptor signaling in POMC neurons is required for normal body weight homeostasis. Neuron. 2004;42:983–991. doi: 10.1016/j.neuron.2004.06.004. [DOI] [PubMed] [Google Scholar]
- Basak O, Giachino C, Fiorini E, Macdonald HR, Taylor V. Neurogenic subventricular zone stem/progenitor cells are Notch1-dependent in their active but not quiescent state. J Neurosci. 2012;32:5654–5666. doi: 10.1523/JNEUROSCI.0455-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basak O, Taylor V. Identification of self-replicating multipotent progenitors in the embryonic nervous system by high Notch activity and Hes5 expression. Eur J Neurosci. 2007;25:1006–1022. doi: 10.1111/j.1460-9568.2007.05370.x. [DOI] [PubMed] [Google Scholar]
- Beckervordersandforth R, Deshpande A, Schaffner I, Huttner HB, Lepier A, Lie DC, Gotz M. In vivo targeting of adult neural stem cells in the dentate gyrus by a split-cre approach. Stem Cell Reports. 2014;2:153–162. doi: 10.1016/j.stemcr.2014.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beech RD, Cleary MA, Treloar HB, Eisch AJ, Harrist AV, Zhong W, Greer CA, Duman RS, Picciotto MR. Nestin promoter/enhancer directs transgene expression to precursors of adult generated periglomerular neurons. J Comp Neurol. 2004;475:128–141. doi: 10.1002/cne.20179. [DOI] [PubMed] [Google Scholar]
- Berg DA, Yoon KJ, Will B, Xiao AY, Kim NS, Christian KM, Song H, Ming G-L. Tbr2-expressing intermediate progenitor cells in the adult mouse hippocampus are unipotent neuronal precursors with limited amplification capacity under homeostasis. Frontiers in Biology. 2015;10:262–271. [Google Scholar]
- Berninger B, Costa MR, Koch U, Schroeder T, Sutor B, Grothe B, Gotz M. Functional properties of neurons derived from in vitro reprogrammed postnatal astroglia. J Neurosci. 2007;27:8654–8664. doi: 10.1523/JNEUROSCI.1615-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertrand N, Castro DS, Guillemot F. Proneural genes and the specification of neural cell types. Nat Rev Neurosci. 2002;3:517–530. doi: 10.1038/nrn874. [DOI] [PubMed] [Google Scholar]
- Betz UA, Vosshenrich CA, Rajewsky K, Muller W. Bypass of lethality with mosaic mice generated by Cre-loxP-mediated recombination. Curr Biol. 1996;6:1307–1316. doi: 10.1016/s0960-9822(02)70717-3. [DOI] [PubMed] [Google Scholar]
- Bonaguidi MA, Song J, Ming GL, Song H. A unifying hypothesis on mammalian neural stem cell properties in the adult hippocampus. Curr Opin Neurobiol. 2012;22:754–761. doi: 10.1016/j.conb.2012.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonaguidi MA, Wheeler MA, Shapiro JS, Stadel RP, Sun GJ, Ming GL, Song H. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell. 2011;145:1142–1155. doi: 10.1016/j.cell.2011.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bond AM, Ming GL, Song H. Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later. Cell Stem Cell. 2015;17:385–395. doi: 10.1016/j.stem.2015.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bracko O, Singer T, Aigner S, Knobloch M, Winner B, Ray J, Clemenson GD, Jr, Suh H, Couillard-Despres S, Aigner L, et al. Gene expression profiling of neural stem cells and their neuronal progeny reveals IGF2 as a regulator of adult hippocampal neurogenesis. J Neurosci. 2012;32:3376–3387. doi: 10.1523/JNEUROSCI.4248-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breunig JJ, Silbereis J, Vaccarino FM, Sestan N, Rakic P. Notch regulates cell fate and dendrite morphology of newborn neurons in the postnatal dentate gyrus. Proc Natl Acad Sci U S A. 2007;104:20558–20563. doi: 10.1073/pnas.0710156104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brill MS, Ninkovic J, Winpenny E, Hodge RD, Ozen I, Yang R, Lepier A, Gascon S, Erdelyi F, Szabo G, et al. Adult generation of glutamatergic olfactory bulb interneurons. Nat Neurosci. 2009;12:1524–1533. doi: 10.1038/nn.2416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron HA, Woolley CS, McEwen BS, Gould E. Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat. Neuroscience. 1993;56:337–344. doi: 10.1016/0306-4522(93)90335-d. [DOI] [PubMed] [Google Scholar]
- Casper KB, McCarthy KD. GFAP-positive progenitor cells produce neurons and oligodendrocytes throughout the CNS. Mol Cell Neurosci. 2006;31:676–684. doi: 10.1016/j.mcn.2005.12.006. [DOI] [PubMed] [Google Scholar]
- Chapouton P, Skupien P, Hesl B, Coolen M, Moore JC, Madelaine R, Kremmer E, Faus-Kessler T, Blader P, Lawson ND, et al. Notch activity levels control the balance between quiescence and recruitment of adult neural stem cells. J Neurosci. 2010;30:7961–7974. doi: 10.1523/JNEUROSCI.6170-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ. Transgenic animals with inducible, targeted gene expression in brain. Mol Pharmacol. 1998;54:495–503. doi: 10.1124/mol.54.3.495. [DOI] [PubMed] [Google Scholar]
- Chojnacki AK, Mak GK, Weiss S. Identity crisis for adult periventricular neural stem cells: subventricular zone astrocytes, ependymal cells or both? Nat Rev Neurosci. 2009;10:153–163. doi: 10.1038/nrn2571. [DOI] [PubMed] [Google Scholar]
- Chuang JZ, Milner TA, Sung CH. Subunit heterogeneity of cytoplasmic dynein: Differential expression of 14 kDa dynein light chains in rat hippocampus. J Neurosci. 2001;21:5501–5512. doi: 10.1523/JNEUROSCI.21-15-05501.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clelland CD, Choi M, Romberg C, Clemenson GD, Jr, Fragniere A, Tyers P, Jessberger S, Saksida LM, Barker RA, Gage FH, et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science. 2009;325:210–213. doi: 10.1126/science.1173215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Codega P, Silva-Vargas V, Paul A, Maldonado-Soto AR, Deleo AM, Pastrana E, Doetsch F. Prospective identification and purification of quiescent adult neural stem cells from their in vivo niche. Neuron. 2014;82:545–559. doi: 10.1016/j.neuron.2014.02.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collignon J. Phd Thesis. CNAA; London: 1992. Study of a new family of genes related to the mammalian testis determining gene. [Google Scholar]
- Couillard-Despres S, Winner B, Karl C, Lindemann G, Schmid P, Aigner R, Laemke J, Bogdahn U, Winkler J, Bischofberger J, et al. Targeted transgene expression in neuronal precursors: watching young neurons in the old brain. Eur J Neurosci. 2006;24:1535–1545. doi: 10.1111/j.1460-9568.2006.05039.x. [DOI] [PubMed] [Google Scholar]
- Cowley MA, Smart JL, Rubinstein M, Cerdan MG, Diano S, Horvath TL, Cone RD, Low MJ. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature. 2001;411:480–484. doi: 10.1038/35078085. [DOI] [PubMed] [Google Scholar]
- D’Amour KA, Gage FH. Genetic and functional differences between multipotent neural and pluripotent embryonic stem cells. Proc Natl Acad Sci U S A. 2003;100(Suppl 1):11866–11872. doi: 10.1073/pnas.1834200100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- David DJ, Wang J, Samuels BA, Rainer Q, David I, Gardier AM, Hen R. Implications of the functional integration of adult-born hippocampal neurons in anxiety-depression disorders. Neuroscientist. 2010;16:578–591. doi: 10.1177/1073858409360281. [DOI] [PubMed] [Google Scholar]
- Day K, Shefer G, Richardson JB, Enikolopov G, Yablonka-Reuveni Z. Nestin-GFP reporter expression defines the quiescent state of skeletal muscle satellite cells. Dev Biol. 2007;304:246–259. doi: 10.1016/j.ydbio.2006.12.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeCarolis NA, Mechanic M, Petrik D, Carlton A, Ables JL, Malhotra S, Bachoo R, Gotz M, Lagace DC, Eisch AJ. In vivo contribution of nestin- and GLAST-lineage cells to adult hippocampal neurogenesis. Hippocampus. 2013;23:708–719. doi: 10.1002/hipo.22130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dedesma C, Chuang JZ, Alfinito PD, Sung CH. Dynein light chain Tctex-1 identifies neural progenitors in adult brain. J Comp Neurol. 2006;496:773–786. doi: 10.1002/cne.20958. [DOI] [PubMed] [Google Scholar]
- Deng W, Saxe MD, Gallina IS, Gage FH. Adult-born hippocampal dentate granule cells undergoing maturation modulate learning and memory in the brain. J Neurosci. 2009;29:13532–13542. doi: 10.1523/JNEUROSCI.3362-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Djuric PM, Wagshul ME, Henn FB, H EG, Maletic-Savatic M. Singular Value Decomposition algorithm for detection of neural progenitor cells in the live human brain. Science. 2008;321:640. doi: 10.1126/science.1156889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci. 1997;17:5046–5061. doi: 10.1523/JNEUROSCI.17-13-05046.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Regeneration of a germinal layer in the adult mammalian brain. Proc Natl Acad Sci U S A. 1999;96:11619–11624. doi: 10.1073/pnas.96.20.11619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dranovsky A, Picchini AM, Moadel T, Sisti AC, Yamada A, Kimura S, Leonardo ED, Hen R. Experience dictates stem cell fate in the adult hippocampus. Neuron. 2011;70:908–923. doi: 10.1016/j.neuron.2011.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupret D, Revest JM, Koehl M, Ichas F, De Giorgi F, Costet P, Abrous DN, Piazza PV. Spatial relational memory requires hippocampal adult neurogenesis. PLoS One. 2008;3:e1959. doi: 10.1371/journal.pone.0001959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis P, Fagan BM, Magness ST, Hutton S, Taranova O, Hayashi S, McMahon A, Rao M, Pevny L. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev Neurosci. 2004;26:148–165. doi: 10.1159/000082134. [DOI] [PubMed] [Google Scholar]
- Encinas JM, Michurina TV, Peunova N, Park JH, Tordo J, Peterson DA, Fishell G, Koulakov A, Enikolopov G. Division-coupled astrocytic differentiation and age-related depletion of neural stem cells in the adult hippocampus. Cell Stem Cell. 2011;8:566–579. doi: 10.1016/j.stem.2011.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Encinas JM, Vaahtokari A, Enikolopov G. Fluoxetine targets early progenitor cells in the adult brain. Proc Natl Acad Sci U S A. 2006;103:8233–8238. doi: 10.1073/pnas.0601992103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Englund C, Fink A, Lau C, Pham D, Daza RA, Bulfone A, Kowalczyk T, Hevner RF. Pax6, Tbr2, and Tbr1 are expressed sequentially by radial glia, intermediate progenitor cells, and postmitotic neurons in developing neocortex. J Neurosci. 2005;25:247–251. doi: 10.1523/JNEUROSCI.2899-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- Farioli-Vecchioli S, Saraulli D, Costanzi M, Pacioni S, Cina I, Aceti M, Micheli L, Bacci A, Cestari V, Tirone F. The timing of differentiation of adult hippocampal neurons is crucial for spatial memory. PLoS Biol. 2008;6:e246. doi: 10.1371/journal.pbio.0060246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farnsworth DR, Bayraktar OA, Doe CQ. Aging Neural Progenitors Lose Competence to Respond to Mitogenic Notch Signaling. Curr Biol. 2015 doi: 10.1016/j.cub.2015.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Favaro R, Valotta M, Ferri AL, Latorre E, Mariani J, Giachino C, Lancini C, Tosetti V, Ottolenghi S, Taylor V, et al. Hippocampal development and neural stem cell maintenance require Sox2-dependent regulation of Shh. Nat Neurosci. 2009;12:1248–1256. doi: 10.1038/nn.2397. [DOI] [PubMed] [Google Scholar]
- Ferri AL, Cavallaro M, Braida D, Di Cristofano A, Canta A, Vezzani A, Ottolenghi S, Pandolfi PP, Sala M, DeBiasi S, et al. Sox2 deficiency causes neurodegeneration and impaired neurogenesis in the adult mouse brain. Development. 2004;131:3805–3819. doi: 10.1242/dev.01204. [DOI] [PubMed] [Google Scholar]
- Filippov V, Kronenberg G, Pivneva T, Reuter K, Steiner B, Wang LP, Yamaguchi M, Kettenmann H, Kempermann G. Subpopulation of nestin-expressing progenitor cells in the adult murine hippocampus shows electrophysiological and morphological characteristics of astrocytes. Mol Cell Neurosci. 2003;23:373–382. doi: 10.1016/s1044-7431(03)00060-5. [DOI] [PubMed] [Google Scholar]
- Gama-Norton L, Ferrando E, Ruiz-Herguido C, Liu Z, Guiu J, Islam AB, Lee SU, Yan M, Guidos CJ, Lopez-Bigas N, et al. Notch signal strength controls cell fate in the haemogenic endothelium. Nat Commun. 2015;6:8510. doi: 10.1038/ncomms9510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ganat YM, Silbereis J, Cave C, Ngu H, Anderson GM, Ohkubo Y, Ment LR, Vaccarino FM. Early postnatal astroglial cells produce multilineage precursors and neural stem cells in vivo. J Neurosci. 2006;26:8609–8621. doi: 10.1523/JNEUROSCI.2532-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia AD, Doan NB, Imura T, Bush TG, Sofroniew MV. GFAP-expressing progenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat Neurosci. 2004;7:1233–1241. doi: 10.1038/nn1340. [DOI] [PubMed] [Google Scholar]
- Gheusi G, Cremer H, McLean H, Chazal G, Vincent JD, Lledo PM. Importance of newly generated neurons in the adult olfactory bulb for odor discrimination. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:1823–1828. doi: 10.1073/pnas.97.4.1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giachino C, Basak O, Lugert S, Knuckles P, Obernier K, Fiorelli R, Frank S, Raineteau O, Alvarez-Buylla A, Taylor V. Molecular diversity subdivides the adult forebrain neural stem cell population. Stem Cells. 2014;32:70–84. doi: 10.1002/stem.1520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giachino C, Taylor V. Notching up neural stem cell homogeneity in homeostasis and disease. Front Neurosci. 2014;8:32. doi: 10.3389/fnins.2014.00032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glowatzki E, Cheng N, Hiel H, Yi E, Tanaka K, Ellis-Davies GC, Rothstein JD, Bergles DE. The glutamate-aspartate transporter GLAST mediates glutamate uptake at inner hair cell afferent synapses in the mammalian cochlea. J Neurosci. 2006;26:7659–7664. doi: 10.1523/JNEUROSCI.1545-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Schambra UB, Nowak NJ, Joyner A, Leblanc G, Hatten ME, et al. A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature. 2003;425:917–925. doi: 10.1038/nature02033. [DOI] [PubMed] [Google Scholar]
- Hartfuss E, Galli R, Heins N, Gotz M. Characterization of CNS precursor subtypes and radial glia. Dev Biol. 2001;229:15–30. doi: 10.1006/dbio.2000.9962. [DOI] [PubMed] [Google Scholar]
- Hatakeyama J, Bessho Y, Katoh K, Ookawara S, Fujioka M, Guillemot F, Kageyama R. Hes genes regulate size shape and histogenesis of the nervous system by control of the timing of neural stem cell differentiation. Development. 2004;131:5539–5550. doi: 10.1242/dev.01436. [DOI] [PubMed] [Google Scholar]
- Hayashi A, Koob JW, Liu DZ, Tong AY, Hunter DA, Parsadanian A, Mackinnon SE, Myckatyn TM. A double-transgenic mouse used to track migrating Schwann cells and regenerating axons following engraftment of injured nerves. Exp Neurol. 2007;207:128–138. doi: 10.1016/j.expneurol.2007.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hegedus B, Dasgupta B, Shin JE, Emnett RJ, Hart-Mahon EK, Elghazi L, Bernal-Mizrachi E, Gutmann DH. Neurofibromatosis-1 regulates neuronal and glial cell differentiation from neuroglial progenitors in vivo by both cAMP- and Ras-dependent mechanisms. Cell Stem Cell. 2007;1:443–457. doi: 10.1016/j.stem.2007.07.008. [DOI] [PubMed] [Google Scholar]
- Heine VM, Zareno J, Maslam S, Joels M, Lucassen PJ. Chronic stress in the adult dentate gyrus reduces cell proliferation near the vasculature and VEGF and Flk–1 protein expression. Eur J Neurosci. 2005;21:1304–1314. doi: 10.1111/j.1460-9568.2005.03951.x. [DOI] [PubMed] [Google Scholar]
- Hirrlinger PG, Scheller A, Braun C, Quintela-Schneider M, Fuss B, Hirrlinger J, Kirchhoff F. Expression of reef coral fluorescent proteins in the central nervous system of transgenic mice. Mol Cell Neurosci. 2005;30:291–303. doi: 10.1016/j.mcn.2005.08.011. [DOI] [PubMed] [Google Scholar]
- Hockfield S, McKay RD. Identification of major cell classes in the developing mammalian nervous system. J Neurosci. 1985;5:3310–3328. doi: 10.1523/JNEUROSCI.05-12-03310.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodge RD, Kowalczyk TD, Wolf SA, Encinas JM, Rippey C, Enikolopov G, Kempermann G, Hevner RF. Intermediate progenitors in adult hippocampal neurogenesis: Tbr2 expression and coordinate regulation of neuronal output. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2008;28:3707–3717. doi: 10.1523/JNEUROSCI.4280-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt RF, Dinday MT, Hindle-Katel W, Baraban SC. LIS1 deficiency promotes dysfunctional synaptic integration of granule cells generated in the developing and adult dentate gyrus. J Neurosci. 2012;32:12862–12875. doi: 10.1523/JNEUROSCI.1286-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imayoshi I, Ohtsuka T, Metzger D, Chambon P, Kageyama R. Temporal regulation of Cre recombinase activity in neural stem cells. Genesis. 2006;44:233–238. doi: 10.1002/dvg.20212. [DOI] [PubMed] [Google Scholar]
- Imayoshi I, Sakamoto M, Ohtsuka T, Takao K, Miyakawa T, Yamaguchi M, Mori K, Ikeda T, Itohara S, Kageyama R. Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci. 2008;11:1153–1161. doi: 10.1038/nn.2185. [DOI] [PubMed] [Google Scholar]
- Jacobs BL, van Praag H, Gage FH. Adult brain neurogenesis and psychiatry: a novel theory of depression. Molecular psychiatry. 2000;5:262–269. doi: 10.1038/sj.mp.4000712. [DOI] [PubMed] [Google Scholar]
- Jessberger S, Toni N, Clemenson GD, Jr, Ray J, Gage FH. Directed differentiation of hippocampal stem/progenitor cells in the adult brain. Nat Neurosci. 2008;11:888–893. doi: 10.1038/nn.2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joels M, Karst H, Krugers HJ, Lucassen PJ. Chronic stress: implications for neuronal morphology, function and neurogenesis. Front Neuroendocrinol. 2007;28:72–96. doi: 10.1016/j.yfrne.2007.04.001. [DOI] [PubMed] [Google Scholar]
- Johansson CB, Lothian C, Molin M, Okano H, Lendahl U. Nestin enhancer requirements for expression in normal and injured adult CNS. J Neurosci Res. 2002;69:784–794. doi: 10.1002/jnr.10376. [DOI] [PubMed] [Google Scholar]
- Josephson R, Muller T, Pickel J, Okabe S, Reynolds K, Turner PA, Zimmer A, McKay RD. POU transcription factors control expression of CNS stem cell-specific genes. Development. 1998;125:3087–3100. doi: 10.1242/dev.125.16.3087. [DOI] [PubMed] [Google Scholar]
- Jourdon A, Gresset A, Spassky N, Charnay P, Topilko P, Santos R. Prss56, a novel marker of adult neurogenesis in the mouse brain. Brain Struct Funct. 2015 doi: 10.1007/s00429-015-1171-z. [DOI] [PubMed] [Google Scholar]
- Jump DB, Oppenheimer JH. High basal expression and 3,5,3′-triiodothyronine regulation of messenger ribonucleic acid S14 in lipogenic tissues. Endocrinology. 1985;117:2259–2266. doi: 10.1210/endo-117-6-2259. [DOI] [PubMed] [Google Scholar]
- Kageyama R, Ohtsuka T. The Notch-Hes pathway in mammalian neural development. Cell Res. 1999;9:179–188. doi: 10.1038/sj.cr.7290016. [DOI] [PubMed] [Google Scholar]
- Kamachi Y, Kondoh H. Sox proteins: regulators of cell fate specification and differentiation. Development. 2013;140:4129–4144. doi: 10.1242/dev.091793. [DOI] [PubMed] [Google Scholar]
- Kang W, Hebert JM. A Sox2 BAC transgenic approach for targeting adult neural stem cells. PLoS One. 2012;7:e49038. doi: 10.1371/journal.pone.0049038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karow M, Sanchez R, Schichor C, Masserdotti G, Ortega F, Heinrich C, Gascon S, Khan MA, Lie DC, Dellavalle A, et al. Reprogramming of pericyte-derived cells of the adult human brain into induced neuronal cells. Cell Stem Cell. 2012;11:471–476. doi: 10.1016/j.stem.2012.07.007. [DOI] [PubMed] [Google Scholar]
- Kawaguchi A, Miyata T, Sawamoto K, Takashita N, Murayama A, Akamatsu W, Ogawa M, Okabe M, Tano Y, Goldman SA, et al. Nestin-EGFP transgenic mice: visualization of the self-renewal and multipotency of CNS stem cells. Mol Cell Neurosci. 2001;17:259–273. doi: 10.1006/mcne.2000.0925. [DOI] [PubMed] [Google Scholar]
- Kim EJ, Ables JL, Dickel LK, Eisch AJ, Johnson JE. Ascl1 (Mash1) defines cells with long-term neurogenic potential in subgranular and subventricular zones in adult mouse brain. PLoS One. 2011;6:e18472. doi: 10.1371/journal.pone.0018472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim EJ, Leung CT, Reed RR, Johnson JE. In vivo analysis of Ascl1 defined progenitors reveals distinct developmental dynamics during adult neurogenesis and gliogenesis. J Neurosci. 2007;27:12764–12774. doi: 10.1523/JNEUROSCI.3178-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitamura T, Saitoh Y, Takashima N, Murayama A, Niibori Y, Ageta H, Sekiguchi M, Sugiyama H, Inokuchi K. Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell. 2009;139:814–827. doi: 10.1016/j.cell.2009.10.020. [DOI] [PubMed] [Google Scholar]
- Knobloch M, Braun SM, Zurkirchen L, von Schoultz C, Zamboni N, Arauzo-Bravo MJ, Kovacs WJ, Karalay O, Suter U, Machado RA, et al. Metabolic control of adult neural stem cell activity by Fasn-dependent lipogenesis. Nature. 2013;493:226–230. doi: 10.1038/nature11689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knobloch M, von Schoultz C, Zurkirchen L, Braun SM, Vidmar M, Jessberger S. SPOT14-positive neural stem/progenitor cells in the hippocampus respond dynamically to neurogenic regulators. Stem Cell Reports. 2014;3:735–742. doi: 10.1016/j.stemcr.2014.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuo CT, Mirzadeh Z, Soriano-Navarro M, Rasin M, Wang D, Shen J, Sestan N, Garcia-Verdugo J, Alvarez-Buylla A, Jan LY, et al. Postnatal deletion of Numb/Numblike reveals repair and remodeling capacity in the subventricular neurogenic niche. Cell. 2006;127:1253–1264. doi: 10.1016/j.cell.2006.10.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon GS, Hadjantonakis AK. Eomes∷GFP-a tool for live imaging cells of the trophoblast, primitive streak, and telencephalon in the mouse embryo. Genesis. 2007;45:208–217. doi: 10.1002/dvg.20293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagace DC, Whitman MC, Noonan MA, Ables JL, DeCarolis NA, Arguello AA, Donovan MH, Fischer SJ, Farnbauch LA, Beech RD, et al. Dynamic contribution of nestin-expressing stem cells to adult neurogenesis. J Neurosci. 2007;27:12623–12629. doi: 10.1523/JNEUROSCI.3812-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai K, Kaspar BK, Gage FH, Schaffer DV. Sonic hedgehog regulates adult neural progenitor proliferation in vitro and in vivo. Nat Neurosci. 2003;6:21–27. doi: 10.1038/nn983. [DOI] [PubMed] [Google Scholar]
- Lendahl U, Zimmerman LB, McKay RD. CNS stem cells express a new class of intermediate filament protein. Cell. 1990;60:585–595. doi: 10.1016/0092-8674(90)90662-x. [DOI] [PubMed] [Google Scholar]
- Leung CT, Coulombe PA, Reed RR. Contribution of olfactory neural stem cells to tissue maintenance and regeneration. Nat Neurosci. 2007;10:720–726. doi: 10.1038/nn1882. [DOI] [PubMed] [Google Scholar]
- Li D, Takeda N, Jain R, Manderfield LJ, Liu F, Li L, Anderson SA, Epstein JA. Hopx distinguishes hippocampal from lateral ventricle neural stem cells. Stem Cell Res. 2015;15:522–529. doi: 10.1016/j.scr.2015.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Mignone J, Yang M, Matic M, Penman S, Enikolopov G, Hoffman RM. Nestin expression in hair follicle sheath progenitor cells. Proc Natl Acad Sci U S A. 2003;100:9958–9961. doi: 10.1073/pnas.1733025100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Zhao X, Fang Y, Jiang X, Duong T, Fan C, Huang CC, Kain SR. Generation of destabilized green fluorescent protein as a transcription reporter. J Biol Chem. 1998;273:34970–34975. doi: 10.1074/jbc.273.52.34970. [DOI] [PubMed] [Google Scholar]
- Lobo MV, Arenas MI, Alonso FJ, Gomez G, Bazan E, Paino CL, Fernandez E, Fraile B, Paniagua R, Moyano A, et al. Nestin, a neuroectodermal stem cell marker molecule, is expressed in Leydig cells of the human testis and in some specific cell types from human testicular tumours. Cell Tissue Res. 2004;316:369–376. doi: 10.1007/s00441-003-0848-4. [DOI] [PubMed] [Google Scholar]
- Lois C, Alvarez-Buylla A. Long-distance neuronal migration in the adult mammalian brain. Science. 1994;264:1145–1148. doi: 10.1126/science.8178174. [DOI] [PubMed] [Google Scholar]
- Lothian C, Prakash N, Lendahl U, Wahlstrom GM. Identification of both general and region-specific embryonic CNS enhancer elements in the nestin promoter. Exp Cell Res. 1999;248:509–519. doi: 10.1006/excr.1999.4417. [DOI] [PubMed] [Google Scholar]
- Lucassen PJ, Oomen CA, Naninck EF, Fitzsimons CP, van Dam AM, Czeh B, Korosi A. Regulation of Adult Neurogenesis and Plasticity by (Early) Stress, Glucocorticoids, and Inflammation. Cold Spring Harb Perspect Biol. 2015;7:a021303. doi: 10.1101/cshperspect.a021303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lugert S, Basak O, Knuckles P, Haussler U, Fabel K, Gotz M, Haas CA, Kempermann G, Taylor V, Giachino C. Quiescent and active hippocampal neural stem cells with distinct morphologies respond selectively to physiological and pathological stimuli and aging. Cell Stem Cell. 2010;6:445–456. doi: 10.1016/j.stem.2010.03.017. [DOI] [PubMed] [Google Scholar]
- Lugert S, Vogt M, Tchorz JS, Muller M, Giachino C, Taylor V. Homeostatic neurogenesis in the adult hippocampus does not involve amplification of Ascl1(high) intermediate progenitors. Nat Commun. 2012;3:670. doi: 10.1038/ncomms1670. [DOI] [PubMed] [Google Scholar]
- Luskin MB. Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron. 1993;11:173–189. doi: 10.1016/0896-6273(93)90281-u. [DOI] [PubMed] [Google Scholar]
- Machold R, Hayashi S, Rutlin M, Muzumdar MD, Nery S, Corbin JG, Gritli-Linde A, Dellovade T, Porter JA, Rubin LL, et al. Sonic hedgehog is required for progenitor cell maintenance in telencephalic stem cell niches. Neuron. 2003;39:937–950. doi: 10.1016/s0896-6273(03)00561-0. [DOI] [PubMed] [Google Scholar]
- Mainen ZF, Maletic-Savatic M, Shi SH, Hayashi Y, Malinow R, Svoboda K. Two-photon imaging in living brain slices. Methods. 1999;18:231–239. doi: 10.1006/meth.1999.0776. [DOI] [PubMed] [Google Scholar]
- Mak GK, Enwere EK, Gregg C, Pakarainen T, Poutanen M, Huhtaniemi I, Weiss S. Male pheromone-stimulated neurogenesis in the adult female brain: possible role in mating behavior. Nature neuroscience. 2007;10:1003–1011. doi: 10.1038/nn1928. [DOI] [PubMed] [Google Scholar]
- Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2000;20:9104–9110. doi: 10.1523/JNEUROSCI.20-24-09104.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maletic-Savatic M, Vingara LK, Manganas LN, Li Y, Zhang S, Sierra A, Hazel R, Smith D, Wagshul ME, Henn F, et al. Metabolomics of neural progenitor cells: a novel approach to biomarker discovery. Cold Spring Harb Symp Quant Biol. 2008;73:389–401. doi: 10.1101/sqb.2008.73.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manganas LN, Maletic-Savatic M. Stem cell therapy for central nervous system demyelinating disease. Curr Neurol Neurosci Rep. 2005;5:225–231. doi: 10.1007/s11910-005-0050-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manganas LN, Zhang X, Li Y, Hazel RD, Smith SD, Wagshul ME, Henn F, Benveniste H, Djuric PM, Enikolopov G, et al. Magnetic resonance spectroscopy identifies neural progenitor cells in the live human brain. Science. 2007;318:980–985. doi: 10.1126/science.1147851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marino S, Vooijs M, van Der Gulden H, Jonkers J, Berns A. Induction of medulloblastomas in p53-null mutant mice by somatic inactivation of Rb in the external granular layer cells of the cerebellum. Genes Dev. 2000;14:994–1004. [PMC free article] [PubMed] [Google Scholar]
- Mayer EJ, Hughes EH, Carter DA, Dick AD. Nestin positive cells in adult human retina and in epiretinal membranes. Br J Ophthalmol. 2003;87:1154–1158. doi: 10.1136/bjo.87.9.1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell J, Petrie L, Stennard F, Ryan K, Nichols J. Eomesodermin is expressed in mouse oocytes and pre-implantation embryos. Mol Reprod Dev. 2005;71:399–404. doi: 10.1002/mrd.20318. [DOI] [PubMed] [Google Scholar]
- McHugh TJ, Jones MW, Quinn JJ, Balthasar N, Coppari R, Elmquist JK, Lowell BB, Fanselow MS, Wilson MA, Tonegawa S. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science. 2007;317:94–99. doi: 10.1126/science.1140263. [DOI] [PubMed] [Google Scholar]
- Mignone JL, Kukekov V, Chiang AS, Steindler D, Enikolopov G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J Comp Neurol. 2004;469:311–324. doi: 10.1002/cne.10964. [DOI] [PubMed] [Google Scholar]
- Ming GL, Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011;70:687–702. doi: 10.1016/j.neuron.2011.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuhashi T, Aoki Y, Eksioglu YZ, Takahashi T, Bhide PG, Reeves SA, Caviness VS., Jr Overexpression of p27Kip1 lengthens the G1 phase in a mouse model that targets inducible gene expression to central nervous system progenitor cells. Proc Natl Acad Sci U S A. 2001;98:6435–6440. doi: 10.1073/pnas.111051398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyagi S, Nishimoto M, Saito T, Ninomiya M, Sawamoto K, Okano H, Muramatsu M, Oguro H, Iwama A, Okuda A. The Sox2 regulatory region 2 functions as a neural stem cell-specific enhancer in the telencephalon. J Biol Chem. 2006;281:13374–13381. doi: 10.1074/jbc.M512669200. [DOI] [PubMed] [Google Scholar]
- Mori T, Tanaka K, Buffo A, Wurst W, Kuhn R, Gotz M. Inducible gene deletion in astroglia and radial glia–a valuable tool for functional and lineage analysis. Glia. 2006;54:21–34. doi: 10.1002/glia.20350. [DOI] [PubMed] [Google Scholar]
- Morshead CM, Reynolds BA, Craig CG, McBurney MW, Staines WA, Morassutti D, Weiss S, van der Kooy D. Neural stem cells in the adult mammalian forebrain: a relatively quiescent subpopulation of subependymal cells. Neuron. 1994;13:1071–1082. doi: 10.1016/0896-6273(94)90046-9. [DOI] [PubMed] [Google Scholar]
- Mouret A, Lepousez G, Gras J, Gabellec MM, Lledo PM. Turnover of newborn olfactory bulb neurons optimizes olfaction. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:12302–12314. doi: 10.1523/JNEUROSCI.3383-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakashiba T, Cushman JD, Pelkey KA, Renaudineau S, Buhl DL, McHugh TJ, Rodriguez Barrera V, Chittajallu R, Iwamoto KS, McBain CJ, et al. Young dentate granule cells mediate pattern separation, whereas old granule cells facilitate pattern completion. Cell. 2012;149:188–201. doi: 10.1016/j.cell.2012.01.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namba T, Mochizuki H, Onodera M, Mizuno Y, Namiki H, Seki T. The fate of neural progenitor cells expressing astrocytic and radial glial markers in the postnatal rat dentate gyrus. Eur J Neurosci. 2005;22:1928–1941. doi: 10.1111/j.1460-9568.2005.04396.x. [DOI] [PubMed] [Google Scholar]
- Ninov N, Borius M, Stainier DY. Different levels of Notch signaling regulate quiescence, renewal and differentiation in pancreatic endocrine progenitors. Development. 2012;139:1557–1567. doi: 10.1242/dev.076000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nolte C, Matyash M, Pivneva T, Schipke CG, Ohlemeyer C, Hanisch UK, Kirchhoff F, Kettenmann H. GFAP promoter-controlled EGFP-expressing transgenic mice: a tool to visualize astrocytes and astrogliosis in living brain tissue. Glia. 2001;33:72–86. [PubMed] [Google Scholar]
- Ohtsuka T, Sakamoto M, Guillemot F, Kageyama R. Roles of the basic helix-loop-helix genes Hes1 and Hes5 in expansion of neural stem cells of the developing brain. J Biol Chem. 2001;276:30467–30474. doi: 10.1074/jbc.M102420200. [DOI] [PubMed] [Google Scholar]
- Palmer TD, Takahashi J, Gage FH. The adult rat hippocampus contains primordial neural stem cells. Mol Cell Neurosci. 1997;8:389–404. doi: 10.1006/mcne.1996.0595. [DOI] [PubMed] [Google Scholar]
- Pan YW, Chan GC, Kuo CT, Storm DR, Xia Z. Inhibition of adult neurogenesis by inducible and targeted deletion of ERK5 mitogen-activated protein kinase specifically in adult neurogenic regions impairs contextual fear extinction and remote fear memory. J Neurosci. 2012;32:6444–6455. doi: 10.1523/JNEUROSCI.6076-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastrana E, Cheng LC, Doetsch F. Simultaneous prospective purification of adult subventricular zone neural stem cells and their progeny. Proc Natl Acad Sci U S A. 2009;106:6387–6392. doi: 10.1073/pnas.0810407106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira AC, Huddleston DE, Brickman AM, Sosunov AA, Hen R, McKhann GM, Sloan R, Gage FH, Brown TR, Small SA. An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:5638–5643. doi: 10.1073/pnas.0611721104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perl AK, Wert SE, Nagy A, Lobe CG, Whitsett JA. Early restriction of peripheral and proximal cell lineages during formation of the lung. Proc Natl Acad Sci U S A. 2002;99:10482–10487. doi: 10.1073/pnas.152238499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pevny LH, Sockanathan S, Placzek M, Lovell-Badge R. A role for SOX1 in neural determination. Development. 1998;125:1967–1978. doi: 10.1242/dev.125.10.1967. [DOI] [PubMed] [Google Scholar]
- Pimeisl IM, Tanriver Y, Daza RA, Vauti F, Hevner RF, Arnold HH, Arnold SJ. Generation and characterization of a tamoxifen-inducible Eomes(CreER) mouse line. Genesis. 2013;51:725–733. doi: 10.1002/dvg.22417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platel JC, Gordon V, Heintz T, Bordey A. GFAP-GFP neural progenitors are antigenically homogeneous and anchored in their enclosed mosaic niche. Glia. 2009;57:66–78. doi: 10.1002/glia.20735. [DOI] [PubMed] [Google Scholar]
- Pollak J, Wilken MS, Ueki Y, Cox KE, Sullivan JM, Taylor RJ, Levine EM, Reh TA. ASCL1 reprograms mouse Muller glia into neurogenic retinal progenitors. Development. 2013;140:2619–2631. doi: 10.1242/dev.091355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinones-Hinojosa A, Sanai N, Soriano-Navarro M, Gonzalez-Perez O, Mirzadeh Z, Gil-Perotin S, Romero-Rodriguez R, Berger MS, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular composition and cytoarchitecture of the adult human subventricular zone: a niche of neural stem cells. The Journal of comparative neurology. 2006;494:415–434. doi: 10.1002/cne.20798. [DOI] [PubMed] [Google Scholar]
- Raposo AA, Vasconcelos FF, Drechsel D, Marie C, Johnston C, Dolle D, Bithell A, Gillotin S, van den Berg DL, Ettwiller L, et al. Ascl1 Coordinately Regulates Gene Expression and the Chromatin Landscape during Neurogenesis. Cell Rep. 2015 doi: 10.1016/j.celrep.2015.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regan MR, Huang YH, Kim YS, Dykes-Hoberg MI, Jin L, Watkins AM, Bergles DE, Rothstein JD. Variations in promoter activity reveal a differential expression and physiology of glutamate transporters by glia in the developing and mature CNS. J Neurosci. 2007;27:6607–6619. doi: 10.1523/JNEUROSCI.0790-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahay A, Hen R. Hippocampal neurogenesis and depression. Novartis Found Symp. 2008;289:152–160. doi: 10.1002/9780470751251.ch12. discussion 160–154, 193–155. [DOI] [PubMed] [Google Scholar]
- Sahay A, Scobie KN, Hill AS, O’Carroll CM, Kheirbek MA, Burghardt NS, Fenton AA, Dranovsky A, Hen R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature. 2011;472:466–470. doi: 10.1038/nature09817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakamoto M, Ieki N, Miyoshi G, Mochimaru D, Miyachi H, Imura T, Yamaguchi M, Fishell G, Mori K, Kageyama R, et al. Continuous postnatal neurogenesis contributes to formation of the olfactory bulb neural circuits and flexible olfactory associative learning. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2014;34:5788–5799. doi: 10.1523/JNEUROSCI.0674-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science. 2003;301:805–809. doi: 10.1126/science.1083328. [DOI] [PubMed] [Google Scholar]
- Saxe MD, Battaglia F, Wang JW, Malleret G, David DJ, Monckton JE, Garcia AD, Sofroniew MV, Kandel ER, Santarelli L, et al. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:17501–17506. doi: 10.1073/pnas.0607207103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid RS, Yokota Y, Anton ES. Generation and characterization of brain lipid-binding protein promoter-based transgenic mouse models for the study of radial glia. Glia. 2006;53:345–351. doi: 10.1002/glia.20274. [DOI] [PubMed] [Google Scholar]
- Seri B, Garcia-Verdugo JM, Collado-Morente L, McEwen BS, Alvarez-Buylla A. Cell types, lineage, and architecture of the germinal zone in the adult dentate gyrus. J Comp Neurol. 2004;478:359–378. doi: 10.1002/cne.20288. [DOI] [PubMed] [Google Scholar]
- Seri B, Garcia-Verdugo JM, McEwen BS, Alvarez-Buylla A. Astrocytes give rise to new neurons in the adult mammalian hippocampus. J Neurosci. 2001;21:7153–7160. doi: 10.1523/JNEUROSCI.21-18-07153.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen Q, Wang Y, Kokovay E, Lin G, Chuang SM, Goderie SK, Roysam B, Temple S. Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions. Cell Stem Cell. 2008;3:289–300. doi: 10.1016/j.stem.2008.07.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata T, Watanabe M, Tanaka K, Wada K, Inoue Y. Dynamic changes in expression of glutamate transporter mRNAs in developing brain. Neuroreport. 1996;7:705–709. doi: 10.1097/00001756-199602290-00006. [DOI] [PubMed] [Google Scholar]
- Shibata T, Yamada K, Watanabe M, Ikenaka K, Wada K, Tanaka K, Inoue Y. Glutamate transporter GLAST is expressed in the radial glia-astrocyte lineage of developing mouse spinal cord. J Neurosci. 1997;17:9212–9219. doi: 10.1523/JNEUROSCI.17-23-09212.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimojo H, Ohtsuka T, Kageyama R. Oscillations in notch signaling regulate maintenance of neural progenitors. Neuron. 2008;58:52–64. doi: 10.1016/j.neuron.2008.02.014. [DOI] [PubMed] [Google Scholar]
- Shin J, Berg DA, Zhu Y, Shin JY, Song J, Bonaguidi MA, Enikolopov G, Nauen DW, Christian KM, Ming GL, et al. Single-Cell RNA-Seq with Waterfall Reveals Molecular Cascades underlying Adult Neurogenesis. Cell Stem Cell. 2015;17:360–372. doi: 10.1016/j.stem.2015.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E. Neurogenesis may relate to some but not all types of hippocampal-dependent learning. Hippocampus. 2002;12:578–584. doi: 10.1002/hipo.10103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sierra A, Encinas JM, Maletic-Savatic M. Adult human neurogenesis: from microscopy to magnetic resonance imaging. Front Neurosci. 2011;5:47. doi: 10.3389/fnins.2011.00047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slezak M, Goritz C, Niemiec A, Frisen J, Chambon P, Metzger D, Pfrieger FW. Transgenic mice for conditional gene manipulation in astroglial cells. Glia. 2007;55:1565–1576. doi: 10.1002/glia.20570. [DOI] [PubMed] [Google Scholar]
- Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature. 2011;476:458–461. doi: 10.1038/nature10287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soriano P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat Genet. 1999;21:70–71. doi: 10.1038/5007. [DOI] [PubMed] [Google Scholar]
- Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M, Huttner HB, Bostrom E, Westerlund I, Vial C, Buchholz BA, et al. Dynamics of hippocampal neurogenesis in adult humans. Cell. 2013;153:1219–1227. doi: 10.1016/j.cell.2013.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh H, Consiglio A, Ray J, Sawai T, D’Amour KA, Gage FH. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2+ neural stem cells in the adult hippocampus. Cell Stem Cell. 2007;1:515–528. doi: 10.1016/j.stem.2007.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sultan S, Gebara E, Toni N. Doxycycline increases neurogenesis and reduces microglia in the adult hippocampus. Front Neurosci. 2013;7:131. doi: 10.3389/fnins.2013.00131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun MY, Yetman MJ, Lee TC, Chen Y, Jankowsky JL. Specificity and efficiency of reporter expression in adult neural progenitors vary substantially among nestin-CreER(T2) lines. J Comp Neurol. 2014;522:1191–1208. doi: 10.1002/cne.23497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surget A, Tanti A, Leonardo ED, Laugeray A, Rainer Q, Touma C, Palme R, Griebel G, Ibarguen-Vargas Y, Hen R, et al. Antidepressants recruit new neurons to improve stress response regulation. Mol Psychiatry. 2011;16:1177–1188. doi: 10.1038/mp.2011.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherland ML, Delaney TA, Noebels JL. Glutamate transporter mRNA expression in proliferative zones of the developing and adult murine CNS. J Neurosci. 1996;16:2191–2207. doi: 10.1523/JNEUROSCI.16-07-02191.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- Takeda N, Jain R, Leboeuf MR, Padmanabhan A, Wang Q, Li L, Lu MM, Millar SE, Epstein JA. Hopx expression defines a subset of multipotent hair follicle stem cells and a progenitor population primed to give rise to K6+ niche cells. Development. 2013;140:1655–1664. doi: 10.1242/dev.093005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeda N, Jain R, LeBoeuf MR, Wang Q, Lu MM, Epstein JA. Interconversion between intestinal stem cell populations in distinct niches. Science. 2011;334:1420–1424. doi: 10.1126/science.1213214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tavazoie M, Van der Veken L, Silva-Vargas V, Louissaint M, Colonna L, Zaidi B, Garcia-Verdugo JM, Doetsch F. A specialized vascular niche for adult neural stem cells. Cell Stem Cell. 2008;3:279–288. doi: 10.1016/j.stem.2008.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tseng YY, Gruzdeva N, Li A, Chuang JZ, Sung CH. Identification of the Tctex–1 regulatory element that directs expression to neural stem/progenitor cells in developing and adult brain. J Comp Neurol. 2010;518:3327–3342. doi: 10.1002/cne.22402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uwanogho D, Rex M, Cartwright EJ, Pearl G, Healy C, Scotting PJ, Sharpe PT. Embryonic expression of the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role in neuronal development. Mech Dev. 1995;49:23–36. doi: 10.1016/0925-4773(94)00299-3. [DOI] [PubMed] [Google Scholar]
- Venere M, Han YG, Bell R, Song JS, Alvarez-Buylla A, Blelloch R. Sox1 marks an activated neural stem/progenitor cell in the hippocampus. Development. 2012;139:3938–3949. doi: 10.1242/dev.081133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Sudhof TC, Wernig M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010;463:1035–1041. doi: 10.1038/nature08797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker Tara L, Overall Rupert W, Vogler S, Sykes Alex M, Ruhwald S, Lasse D, Ichwan M, Fabel K, Kempermann G. Lysophosphatidic Acid Receptor Is a Functional Marker of Adult Hippocampal Precursor Cells. Stem Cell Reports. 2016 doi: 10.1016/j.stemcr.2016.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker TL, Yasuda T, Adams DJ, Bartlett PF. The doublecortin-expressing population in the developing and adult brain contains multipotential precursors in addition to neuronal-lineage cells. J Neurosci. 2007;27:3734–3742. doi: 10.1523/JNEUROSCI.5060-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Qiu R, Tsark W, Lu Q. Rapid promoter analysis in developing mouse brain and genetic labeling of young neurons by doublecortin-DsRed-express. J Neurosci Res. 2007;85:3567–3573. doi: 10.1002/jnr.21440. [DOI] [PubMed] [Google Scholar]
- Wiese C, Rolletschek A, Kania G, Blyszczuk P, Tarasov KV, Tarasova Y, Wersto RP, Boheler KR, Wobus AM. Nestin expression–a property of multi-lineage progenitor cells? Cell Mol Life Sci. 2004;61:2510–2522. doi: 10.1007/s00018-004-4144-6. [DOI] [PubMed] [Google Scholar]
- Williams SM, Sullivan RK, Scott HL, Finkelstein DI, Colditz PB, Lingwood BE, Dodd PR, Pow DV. Glial glutamate transporter expression patterns in brains from multiple mammalian species. Glia. 2005;49:520–541. doi: 10.1002/glia.20139. [DOI] [PubMed] [Google Scholar]
- Wojtowicz JM, Askew ML, Winocur G. The effects of running and of inhibiting adult neurogenesis on learning and memory in rats. Eur J Neurosci. 2008;27:1494–1502. doi: 10.1111/j.1460-9568.2008.06128.x. [DOI] [PubMed] [Google Scholar]
- Yamaguchi M, Saito H, Suzuki M, Mori K. Visualization of neurogenesis in the central nervous system using nestin promoter-GFP transgenic mice. Neuroreport. 2000;11:1991–1996. doi: 10.1097/00001756-200006260-00037. [DOI] [PubMed] [Google Scholar]
- Yang SM, Alvarez DD, Schinder AF. Reliable Genetic Labeling of Adult-Born Dentate Granule Cells Using Ascl1 CreERT2 and Glast CreERT2 Murine Lines. J Neurosci. 2015;35:15379–15390. doi: 10.1523/JNEUROSCI.2345-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yaworsky PJ, Kappen C. Heterogeneity of neural progenitor cells revealed by enhancers in the nestin gene. Dev Biol. 1999;205:309–321. doi: 10.1006/dbio.1998.9035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu TS, Dandekar M, Monteggia LM, Parada LF, Kernie SG. Temporally regulated expression of Cre recombinase in neural stem cells. Genesis. 2005;41:147–153. doi: 10.1002/gene.20110. [DOI] [PubMed] [Google Scholar]
- Zappone MV, Galli R, Catena R, Meani N, De Biasi S, Mattei E, Tiveron C, Vescovi AL, Lovell-Badge R, Ottolenghi S, et al. Sox2 regulatory sequences direct expression of a (beta)-geo transgene to telencephalic neural stem cells and precursors of the mouse embryo, revealing regionalization of gene expression in CNS stem cells. Development. 2000;127:2367–2382. doi: 10.1242/dev.127.11.2367. [DOI] [PubMed] [Google Scholar]
- Zecevic N. Specific characteristic of radial glia in the human fetal telencephalon. Glia. 2004;48:27–35. doi: 10.1002/glia.20044. [DOI] [PubMed] [Google Scholar]
- Zhang CL, Zou Y, He W, Gage FH, Evans RM. A role for adult TLX-positive neural stem cells in learning and behaviour. Nature. 2008;451:1004–1007. doi: 10.1038/nature06562. [DOI] [PubMed] [Google Scholar]
- Zhao C, Deng W, Gage FH. Mechanisms and functional implications of adult neurogenesis. Cell. 2008;132:645–660. doi: 10.1016/j.cell.2008.01.033. [DOI] [PubMed] [Google Scholar]
- Zhuo L, Sun B, Zhang CL, Fine A, Chiu SY, Messing A. Live astrocytes visualized by green fluorescent protein in transgenic mice. Dev Biol. 1997;187:36–42. doi: 10.1006/dbio.1997.8601. [DOI] [PubMed] [Google Scholar]
- Zimmerman L, Parr B, Lendahl U, Cunningham M, McKay R, Gavin B, Mann J, Vassileva G, McMahon A. Independent regulatory elements in the nestin gene direct transgene expression to neural stem cells or muscle precursors. Neuron. 1994;12:11–24. doi: 10.1016/0896-6273(94)90148-1. [DOI] [PubMed] [Google Scholar]