

Abstract
Protecting coastal communities has become increasingly important as their populations grow, resulting in increased demand for engineered shore protection and hardening of over 50% of many urban shorelines. Shoreline hardening is recognized to reduce ecosystem services that coastal populations rely on, but the amount of hardened coastline continues to grow in many ecologically important coastal regions. Therefore, to inform future management decisions, we conducted a meta-analysis of studies comparing the ecosystem services of biodiversity (richness or diversity) and habitat provisioning (organism abundance) along shorelines with versus without engineered-shore structures. Seawalls supported 23% lower biodiversity and 45% fewer organisms than natural shorelines. In contrast, biodiversity and abundance supported by riprap or breakwater shorelines were not different from natural shorelines; however, effect sizes were highly heterogeneous across organism groups and studies. As coastal development increases, the type and location of shoreline hardening could greatly affect the habitat value and functioning of nearshore ecosystems.
Keywords: biodiversity, bulkhead, ecosystem function, seawall, shoreline hardening
Over the last two centuries, humans have rapidly and dramatically altered the global landscape, causing many to refer to this period as the Anthropocene epoch (Steffen et al. 2007). Some of the strongest examples of anthropogenic change can be found along coastlines. With roughly one-third of human populations living within 100 kilometers of a coastline and continued migration toward coastal areas expected to increase this proportion to one-half by 2030 (Small and Nicholls 2003, MEA 2005), coastal ecosystems are among the most modified and threatened globally (Adger et al. 2005). In efforts to protect people, property, and critical infrastructure from coastal hazards (e.g., erosive waves, storms, and flooding), as well as achieve other human aspirations (e.g., maritime docking, and navigation), coastal societies have historically armored or hardened shorelines with a variety of engineering structures (Dugan et al. 2011). Shoreline hardening, defined as the installation of engineered-shore structures to (a) stabilize sediment and prevent erosion and/or (b) provide flood protection, is a common practice worldwide, with over 22,000 kilometers (roughly 14%) of shoreline hardened in the United States alone (Gittman et al. 2015). Major coastal cities such as New York, Sydney, and Hong Kong have 50% or more of their shorelines hardened (Chapman and Bulleri 2003, Lam et al. 2009, Gittman et al. 2015). Given the current levels of shoreline hardening and the projected growth of coastal populations, understanding the ecological effects of these structures is crucial for developing sustainable coastal management and climate-adaptation strategies (Titus et al. 1998, Gittman et al. 2015). Specifically, understanding how shoreline hardening affects biodiversity and ecosystem functioning is necessary for evaluating the consequences of these activities on associated ecosystem services, such as fisheries production, property protection, and water quality benefits, to coastal communities (Arkema et al. 2015, Scyphers et al. 2015, Gittman et al. 2016).
Although conservation and restoration practitioners have been advocating for the implementation of “living shorelines” or “nature-based” strategies in lieu of traditional “hard” approaches, such as seawalls or bulkheads, over the last three decades (see Broome et al. 1988, Currin et al. 2007), the science on the ecological consequences of various shore-protection structures has lagged behind (NRC 2007). Recent narrative reviews have identified many of the impacts of engineered-shore structures on coastal ecosystems and have recommended ways to minimize these impacts (Chapman and Underwood 2011, Dugan et al. 2011, Perkins et al. 2015); however, a comparative and quantitative synthesis of the effects of engineered-shore structures on coastal ecosystem services has yet to be conducted. The purpose of this systematic review and meta-analysis was to synthesize, quantify, and compare the effects of commonly used engineered-shore structures on the coastal ecosystem services of biodiversity and habitat provision. Moreover, such a synthesis can help inform the development of effective coastal conservation policies and management actions.
Methods
To evaluate the biodiversity and habitat provision effects of different engineered-shore structures, we conducted a systematic review of all studies comparing the biodiversity or abundance of organisms on shorelines with engineered structures versus unmodified shorelines. Three categories of engineered-shore structures were considered: (1) seawalls and bulkheads (figure 1a); (2) riprap revetments (figure 1b); and (3) breakwaters and sills (figure 1c). For the purposes of this review, all vertical walls constructed parallel to shore in or above the high intertidal zone are termed seawalls (figure 1a). Shore-parallel, sloped structures constructed of unconsolidated rock or rubble in or above the high intertidal zone are referred to as riprap revetments (figure 1b). Structures constructed within the low intertidal or subtidal zones are referred to as breakwaters (figure 1c). We have elected to use the term breakwater in lieu of sill in accordance with the terminology used by the United States Army Corps of Engineer (USACE) in their guidance document Low Cost Shore Protection (2001). The materials used to construct the structures evaluated in the selected studies vary and include concrete, granite or sandstone rock, marl, wood, and vinyl sheeting. We defined natural shorelines as rocky, soft-sediment, or biogenic (e.g., marshes, mangroves, oyster reefs, or coral reefs present) shorelines without any engineered-shore structures or modifications (figure 1d–f).
Figure 1.

Example of engineered-shore structures: (a) a seawall; (b) riprap revetment; (c) breakwater; and natural shorelines compared in this study: (d) rocky shoreline (granite platforms); (e) soft-sediment shoreline (sand beach); and (f) biogenic shoreline (salt marsh). Rocky shorelines consist of consolidated rocky platforms and/or cobbles and boulders. Soft-sediment shorelines consist of unconsolidated sediments (sands, muds, silts, clays) without intertidal vegetation. Biogenic shorelines can include intertidal and shallow subtidal marsh, mangrove, bivalve or coral reef, or seagrass.
Peer-reviewed literature search
Using the Web of Science database and the Google Scholar search engine, we searched the literature with the following search terms: structure type (seawall OR bulkhead OR riprap, OR breakwater OR sill) AND response metric (richness OR diversity OR abundance OR density OR cover OR growth OR fitness OR “ecosystem service∗” OR habitat) AND shoreline hardening indicators (“shore∗ hard∗” OR “shore∗ armor∗” OR “shore∗ stabilization” OR “shore protection”) to account for all literature available by 5 November 2015. A total of 121 studies were selected after reviewing the title, keywords, and abstract to determine whether each study evaluated the effects of engineered-shore structures on one or more ecological response variables (e.g., species richness, and abundance). Of those studies, we only included those that compared the ecological effects of one or more engineered-shore structures with those of natural shorelines (e.g., unmodified rocky, soft-sediment, or biogenic shores; figure 1d–f). Studies that evaluated the ecological effects of biogenic methods of shore stabilization (e.g., oyster or marsh restoration) alone were not included because they could also be considered biogenic habitat restoration. However, if the study compared the effects of biogenic habitat restoration, such as marsh planting, combined with construction of an engineered-shore structure (e.g., a rock breakwater) with those of a natural shoreline, then the study was included in the analysis. The evaluation of biogenic habitat restoration effectiveness in restoring, enhancing, or sustaining ecosystems functions has been covered elsewhere (e.g., Peterson and Lipcius 2003, Benayas et al. 2009, Shepard et al. 2011, Baggett et al. 2015) and is beyond the scope of this review. Finally, only studies evaluating the effects of engineered-shore structures on coastal shorelines (including open coast, estuarine, bay, lagoon, and tidally influenced riverine shorelines) were included. Studies of nontidal riverine or lake shorelines, such as shorelines along the Great Lakes, United States, were not included. Applying these criteria yielded 54 studies for further review and analysis (supplemental appendix S1). In 52 of the 54 studies considered, a control-impact (CI) approach was used to compare hardened shorelines to natural shorelines, whereas only two studies sampled hardened and natural shorelines before and after hard shoreline structures were installed (BACI design; e.g., Gittman et al. 2016). Studies that converted or experimentally manipulated the configuration or substrate of hardened shorelines (e.g., Bulleri 2005) were beyond the scope of this review and therefore not included.
Data extraction
We extracted the means, standard deviations, and sample sizes of community (e.g., taxonomic richness and diversity) and individual taxa metrics (e.g., abundance, density, percent cover, and biomass) for hardened and natural shorelines from 32 of the 54 studies (table 1). The remaining 22 studies either did not report the means, standard deviations, or sample sizes for community and individual taxa metrics in an extractable format or had no replication (n = 1) at the level of shoreline type (e.g., seawall or natural; supplemental table S1). Data were extracted from the text, tables, and figures, with data extracted from figures using the software program Data Thief (Tummers 2006). Data for each metric were extracted for each structure or natural-shore comparison (figure 1), with means averaged and standard deviations calculated across replicate sites, time, and species within a phylum or subphylum, but separately by shore zone sampled (e.g., high intertidal, low intertidal, and subtidal) and habitat-use group (flora, benthic infauna, birds, epibiota, and nekton) when reported. Responses for flora included marsh plants, mangroves, and upland shore plants; benthic infauna included organisms living within soft sediments (e.g., bivalves, amphipods, and polychaetes); birds included shorebirds, gulls, and other waterfowl; epibiota include both sessile and mobile organisms living on the surface of the shoreline substrate (e.g., algae, bivalves, barnacles, and gastropods); and nekton included fishes and free-swimming crustaceans. Organisms were grouped into these categories on the basis of their habitat use (e.g., benthic infauna versus nekton) and groupings commonly used in the studies (e.g., epibiota).
Table 1.
Studies included in the meta-analyses.
| Authors | Year | Seawall | Riprap | Breakwater | Rocky | Soft Sediment | Biogenic | Flora | Benthic Infauna | Birds | Epibiota | Nekton |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bilkovic and Mitchell | 2013 | X | X | X | A | A | AB | |||||
| Bilkovic and Roggero | 2008 | X | X | X | B | |||||||
| Bozek and Burdick | 2005 | X | X | A | ||||||||
| Bulleri and Chapman | 2004 | X | X | X | A | |||||||
| Bulleri et al. | 2004 | X | X | A | ||||||||
| Bulleri et al. | 2005 | X | X | A | ||||||||
| Burt et al. | 2009 | X | X | AB | AB | |||||||
| Chapman | 2003 | X | X | B | ||||||||
| Chapman | 2005 | X | X | AB | ||||||||
| Currin et al. | 2007 | X | X | A | AB | |||||||
| Diaz-Agras et al. | 2010 | X | X | A | ||||||||
| Drexler at al. | 2013 | X | X | A | ||||||||
| Dugan and Hubbard | 2006 | X | X | AB | ||||||||
| Dugan et al. | 2008 | X | X | AB | AB | |||||||
| Gittman et al. | 2016 | X | X | X | A | A | AB | |||||
| Glasby et al. | 2007 | X | X | B | ||||||||
| Harris and Strayer | 2014 | X | X | X | B | |||||||
| Heatherington and Bishop | 2012 | X | X | A | ||||||||
| Hendon et al. | 2000 | X | X | X | A | |||||||
| Jackson et al. | 2015 | X | X | A | A | |||||||
| Lam et al. | 2009 | X | X | A | ||||||||
| Lawless and Seitz | 2014 | X | X | X | AB | A | ||||||
| Lee and Li | 2013 | X | X | X | A | |||||||
| Long et al. | 2011 | X | X | X | AB | A | ||||||
| Moreira et al. | 2006 | X | X | A | ||||||||
| Morley et al. | 2012 | X | X | A | AB | A | ||||||
| O'Conner et al. | 2010 | X | X | A | AB | |||||||
| Peters et al. | 2015 | X | X | A | AB | |||||||
| Peterson et al. | 2000 | X | X | X | A | |||||||
| Seitz et al. | 2006 | X | X | X | AB | AB | ||||||
| Sobocinski et al. | 2010 | X | X | X | AB | B | ||||||
| Strayer et al. | 2012 | X | X | X | X | B | A | AB | A |
Note: “A” indicates abundance data and “B” indicates biodiversity data.
Statistical analyses
We calculated effect sizes and corresponding sampling variances for community and individual taxa metrics as log response ratios; the proportional difference between the means for three types of hardened shorelines (seawalls, riprap revetments, and breakwaters) and natural shorelines (Hedges et al. 1999). On the log scale, an effect size of zero means no difference, whereas a negative value means that the hardened shorelines had lower community and individual metrics than natural shorelines. We fit meta-analytic random-effects models to pooled community (i.e., overall and organism-group biodiversity) and pooled individual (i.e., overall and organism-group abundance) effect sizes separately for seawall, riprap revetment, and breakwater comparisons to natural shorelines. Because not all response variables were restricted to specific shore zones (e.g., subtidal or intertidal) and shore zonation classifications were not consistent across studies, we did not compare effect sizes across shore zones. The total amount of residual heterogeneity (τ2) was calculated for each model using restricted maximum-likelihood estimation to account for covariance among responses (Viechtbauer 2010). Residual heterogeneity is variability among the true effects that is not accounted for by the model (Viechtbauer 2010). Differences in the functional responses of organism groups measured, the study ecosystems, and study methods could all contribute to heterogeneity in effect sizes. To allow for consideration of the potential sources of heterogeneity, we explored the effect sizes across organism groups, as well as across studies, through forest plots. Forest plots are recommended for visually assessing the number and precision of the studies included in the meta-analysis and the heterogeneity across effect sizes (Vetter et al. 2013). We included several effect sizes from the same publication, which are not independent; therefore, we included a publication-level random effect to account for the interdependency among multiple within-study observations. To determine the percent difference in biodiversity and abundance between hardened and natural shorelines, we back-transformed the log response ratios and then converted the back-transformed value to a percentage.
The potential for biases in favor of “significant effects” in the published literature (the file drawer problem) is a concern when conducting meta-analyses (Gillman and Wright 2010). To test for “file drawer” bias, we constructed funnel plots for each random effects model and evaluated funnel plot asymmetry using a regression test (Egger et al. 1997). A funnel plot assumes that studies with smaller sample sizes and higher sampling variances are more likely to be skewed and are less likely to be published (Duval and Tweedie 2000). Therefore, asymmetry in the published data can be detected by testing whether the observed effects are related to their sampling sizes. In addition, when a significant effect size was observed, we calculated Rosenthal's fail-safe number, which is the number of unpublished studies, with a mean effect of zero, required to eliminate a significant overall effect size (Rosenthal 1979, Møller and Jennions 2001). A fail-safe number of 5K + 10 or higher, with K being the number of number of studies included in the analysis, is considered to be evidence of a robust average effect size (Rosenthal 1991). All analyses were carried out using R 3.0.1 (R Development Core Team 2016) with the R package metaphor (Viechtbauer 2010).
Results
Of the 32 studies included in the analyses, 78% evaluated seawalls, 28% evaluated riprap revetments, and 25% evaluated breakwaters (table 1). More studies compared the ecosystem function of hardened shorelines with that of biogenic shorelines (n = 16) than with that of rocky (n = 12) or soft-sediment shorelines (n = 8). Most studies were conducted along the Atlantic and Gulf of Mexico coasts of the United States. All of the studies were published since the year 2000, with nearly half of the studies published since 2010.
Seawalls
The overall mean log response ratio (LRR) between seawalls and natural shorelines for biodiversity was −0.26 (95% CI: −0.40, −0.12); therefore, we found that biodiversity was 23% (CI: −33, −11) lower along shorelines with seawalls when compared with that of natural shorelines (z = −3.67, df = 10, p < .001, figure 2a and supplemental figure S1a). The mean LRRs with 95% CIs between seawalls and natural shorelines for biodiversity were negative across all natural shoreline types (biogenic: −0.22 [−0.41, −0.03], rocky: −0.12, [−0.24, −0.01], and soft sediment: −0.52, [−0.78, −0.26]) and all organism groups except epibiota (figure 3a). Biodiversity was significantly lower for flora (66%; [−80, −41]), benthic infauna (20%; [−33, −4]), birds (52%; [−66, −34]), and nekton (24%; [−37, −10]; supplemental figure S2a).
Figure 2.
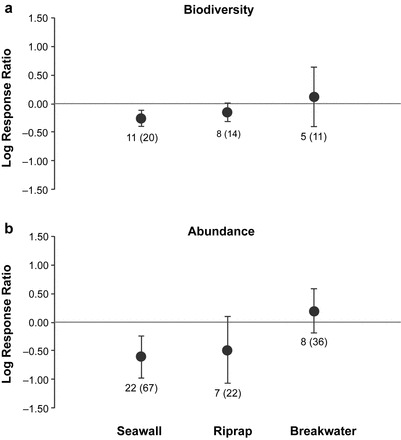
Overall log response ratios between engineered-shore structures (seawall, riprap, breakwater) and natural shorelines for (a) biodiversity and (b) abundance. The error bars represent 95% confidence intervals and data labels show the number of studies and the total number of responses from the studies.
Figure 3.
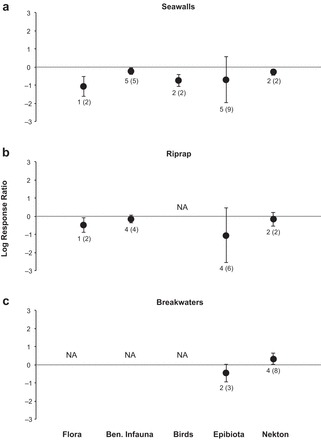
Overall log response ratios between engineered- shore structures (a) seawall, (b) riprap, (c) breakwater and natural shorelines for biodiversity of flora, benthic infauna, birds, epibiota, and nekton. The error bars represent 95% confidence intervals and data labels show the number of studies and the total number of responses from the studies.
The LRR for organism abundance between seawalls and natural shorelines was −0.61 (−0.98, −0.23), corresponding to 45% (−62, −21) lower abundances of organisms along shorelines with seawalls when compared with those along natural shorelines (z = −3.20, df = 21, p = .001, figures 2b and S1b). The mean LRRs with 95% CIs for abundance were negative for biogenic (−0.74, [−1.25, −0.22]) and soft sediment (−1.11, [−1.72, −0.51]), but not rocky (−0.64, [−1.43, 0.16]) shoreline comparisons. All organism groups except flora and epibiota had negative mean LRRs with 95% CIs for abundance between seawalls and natural shorelines (figure 4a). The abundance of benthic infauna, birds, and nekton were 66% (−88, −8), 71% (−86, −41), and 56% (−79, −9) lower, respectively, along shorelines with seawalls when compared with along natural shorelines (supplemental figure S3a).
Figure 4.
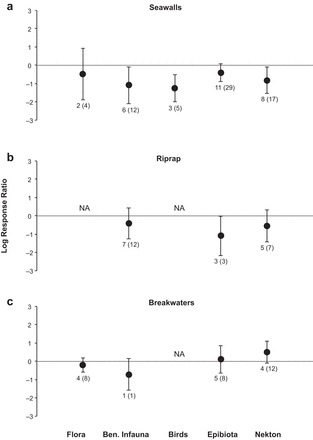
Log response ratios between engineered-shore structures (a) seawall, (b) riprap, (c) breakwater and natural shorelines for abundance of flora, benthic infauna, birds, epibiota, and nekton. The error bars represent 95% confidence intervals and data labels show the number of studies and the total number of responses from the studies.
Our meta-analyses for seawalls included 20 biodiversity and 67 abundance responses from 25 studies. The total heterogeneity (τ2) in the true effect sizes for biodiversity and abundance were estimated to be 0.02 and 0.54, respectively. Fifty-four percent of the total variability in biodiversity effect sizes and 83% of the total variability in abundance effect sizes were attributed to heterogeneity in the true effects (I2). Although true effect size estimates were heterogeneous for both biodiversity (q = 26.25, df = 10, p = .003) and abundance (q = 123.53, df = 21, p < .001), nearly half of the studies found significant, negative effects of seawalls on the biodiversity and abundance of organisms (supplemental figures S4 and S5). There was no evidence of “file drawer bias” or asymmetry in the published data for comparisons of biodiversity or abundance of organisms between seawalls and natural shorelines (z = −1.51, p = .13 and z = 1.04, p = .30, respectively). The Rosenthal fail-safe number for the observed biodiversity effect, or the number of unpublished studies with a mean effect size of zero, needed to eliminate the overall effect size at α = .05 is 122, which is greater than the 65 studies required for a robust effect size estimate. To eliminate the overall observed abundance effect for seawall–natural shoreline comparisons, 651 unpublished studies with a mean effect size of zero would be needed, which is greater than the number of studies required (n = 120) for the effect size to be considered robust.
Riprap revetments
There was no difference in the biodiversity or abundance of organisms found along shorelines with riprap revetments and natural shorelines, with the mean LRRs not being significantly different from zero (z = −1.82, df = 7, p = .07 and z = −1.64, df = 6, p = .10, respectively, figures 2 and S1). Mean biodiversity and abundance did not differ between riprap and natural shorelines across organism groups (figures 3b, 4b, S2, and S3), with the exception of a 39% (CI: −59, −9) reduction in flora biodiversity along riprap shorelines (LRR = −0.49, 95% CI: −0.89, −0.09, figures 3b and S2). The total heterogeneity (τ2) in the true effect sizes for biodiversity and abundance were estimated to be 0.03 and 0.23 respectively. 78% of the total variability in biodiversity effect sizes and 41% the total variability in abundance effect sizes were attributed to heterogeneity in the true effects (I2). The true effect size estimates were heterogeneous for both biodiversity (q = 29.37, df = 7, p < .001) and abundance (q = 0.07, df = 6, p < .001) and varied considerably across studies (supplemental figures S6 and S7). Finally, we did not find evidence of “file drawer bias” or asymmetry (z = −1.84, p = .07 and z = −0.78, p = .43, respectively).
Breakwaters
Similar to the results for riprap revetments, there was no difference in the biodiversity or abundance of organisms found along shorelines with breakwaters when compared with those along natural shorelines (figure 2). The mean LRRs were not significantly different from zero (z = 0.46, df = 4, p = .65 and z = 0.97, df = 7, p = .33, respectively, figures 2 and S1). The 95% CIs for the mean biodiversity and abundance LRRs encompassed zero for all organism groups except for 39% (3, 88) greater biodiversity of nekton (LRR = 0.33, [0.03, 0.63], figures 3c, 4c, S2, and S3) on shorelines with breakwaters compared with that on natural shorelines. The total heterogeneity (τ2) in the true effect sizes for biodiversity and abundance were estimated to be 0.32 and 0.22, respectively. 96% of the total variability in biodiversity effect sizes and 82% of the total variability in abundance effect sizes were attributed to heterogeneity in the true effects (I2). The true effect size estimates were heterogeneous for both biodiversity (q = 50.74, df = 4, p < .001) and abundance (q = 42.86, df = 7, p < .001) and varied considerably across studies (supplemental figures S8 and S9). There was no evidence of “file drawer bias” for published studies comparing breakwaters and natural shorelines (z = 0.81, p = .42 and z = −1.14, p = .25, respectively).
Conclusions
The design of engineered-shore structures and their functional similarity to natural shorelines varies widely across and within structure types (figure 1a–f; Nordstrom 2014, Perkins et al. 2015). Moreover, our analyses revealed some clear distinctions in the quality of habitat provided by the most common engineering alternatives to natural shorelines. Most importantly, seawalls typically supported lower biodiversity and abundance of organisms than did natural shorelines, indicating that these engineered-shore structures are adversely affecting coastal ecosystems (figure 2). Biodiversity and abundance did not differ significantly on riprap and breakwaters from natural shorelines; however, this lack of difference may reflect heterogeneity in the effects of riprap or breakwaters across organism groups, as well as a small number of studies (figures 2–4, S6–S9). Studies included in this meta-analysis proposed that structure complexity and composition of substrate (Chapman and Bulleri 2003, Seitz et al. 2006, Gittman et al. 2016), structure placement within the intertidal or subtidal zones (Bozek and Burdick 2005, Dugan et al. 2008, Bilkovic and Mitchell 2013), and associated wave and sediment dynamics (Bulleri et al. 2004, Bulleri and Chapman 2004, Strayer et al. 2012), may determine whether the biodiversity and abundance of organisms differ between engineered and natural shorelines. Therefore, we explored how reported differences in seawall, riprap, and breakwater structure complexity, composition, and placement related to the biodiversity and abundance of different organism groups below.
Structure complexity and composition
Intertidal and shallow subtidal habitats, particularly structurally complex biogenic habitats (e.g., wetlands, mangroves, and oyster reefs), provide refuge for numerous small and juvenile nekton species (e.g., Fundulus spp., Able et al. 2012; Penaeid shrimp, Boesch and Turner 1984); from abiotic stress (e.g., wave energy, Möller et al. 2014); and from predation (Peterson and Turner 1994). Seawalls can alter the habitat available to nekton by reducing the complexity of intertidal and subtidal habitats (e.g., Chapman and Bulleri 2003, Bilkovic and Roggero 2008). The vertical profile and typically uniform surface of seawalls (figure 1a) does not offer the same refuge for nekton as boulders and camouflaging sediment (Strayer et al. 2012), or dense marsh vegetation (Hendon et al. 2000, Peterson et al. 2000, Seitz et al. 2006, Bilkovic and Roggero 2008, Gittman et al. 2016) characteristic of natural shorelines. The lack of complexity along seawalls likely explains why biodiversity and abundance is lower for many organisms than along natural shorelines.
The biodiversity and abundance of nekton were similar between riprap and natural shorelines (e.g., Seitz et al. 2006, Bilkovic and Roggero 2008, Strayer et al. 2012) and potentially greater at shorelines with breakwaters when compared with those at biogenic shorelines (only type of natural shoreline evaluated, Burt et al. 2009, Peters et al. 2015, Gittman et al. 2016). Because both riprap and breakwaters typically consist of piles of unconsolidated rock and rubble of varying sizes and shapes (figures 1b and 1c; Nordstrom 2014), these structures may provide nekton with equivalent or greater refuge from predation or access to food resources (e.g., epibiota, Clynick et al. 2007; benthic infauna, discussed below) when compared with less structurally complex natural shorelines.
In contrast to nekton, there were no differences in the biodiversity or abundance of epibiota between seawalls or breakwaters and natural shorelines (figures 3 and 4). Epibiota include both sessile organisms such as algae, oysters, mussels, and barnacles, and mobile organisms such as limpets, chitons, snails, and whelks that live on the surface of hard substrates (e.g., shells, rocks, and plants). Despite many studies reporting no differences in epibiota biodiversity (e.g., species richness and Shannon diversity, Glasby et al. 2007) or overall abundance (e.g., Bulleri and Chapman 2004, Bulleri et al. 2005, Lam et al. 2009), several of these same studies did report differences in community composition or dominance (via multivariate ordinations), particularly for mobile grazers, such as limpets, snails, whelks, and chitons (e.g., Bulleri and Chapman 2004, Lam et al. 2009). Further research is needed to understand whether shoreline structures that induce shifts in grazer communities also affect the structure and function of nearshore marine communities.
Of the few studies that reported differences in epibiota, a majority compared engineered-shore structures with soft-sediment or biogenic shorelines. Epibiota biodiversity and abundance were lower along shorelines with seawalls and riprap revetments when compared with those along soft-sediment shores (Sobocinski et al. 2010, Strayer et al. 2012, Harris et al. 2014). In contrast, studies reported higher epibiota diversity and abundance on riprap and breakwaters than on marsh and mangrove shorelines (O'Connor et al. 2010, Drexler et al. 2013, Peters et al. 2015, Gittman et al. 2016). Some shore-protection structures may serve as surrogate habitats for native epibiota where natural hard substrates, such as oyster reefs and mussel beds, have been lost to overharvest, erosion, and poor water quality (Beck et al. 2011). However, the introduction of some types of hard substrates into soft-sediment and biogenic shorelines may also facilitate invasive species. Therefore, the location relative to invasion pathways and substrate type should be carefully considered (Ruiz et al. 1997).
Structure placement and associated wave and sediment dynamics
Intertidal and shallow soft-sediment and biogenic habitats provide refuge for benthic infauna—such as clams (Seitz et al. 2006) and burrowing crustaceans (Dugan et al. 2008)—from predation (Lipcius et al. 2005) and are often occupied by marine flora, such as marsh plants, mangroves, and seagrasses. Larger nektonic predators (e.g., blue crabs) and shorebirds (e.g., sand pipers, willets, and wading birds) forage in soft-sediment and biogenic intertidal and shallow subtidal habitats (Kneib 1982). Lower biodiversity and abundance of benthic infauna and birds were associated with narrower soft-sediment shores along seawalls (Dugan and Hubbard 2006, Dugan et al. 2008), and lower abundances of benthic infauna were also associated with coarser sediments (Sobocinski et al. 2010), leading us to conclude that seawalls reduced both the quantity and quality of habitat available to these organisms. Because they are typically placed in the high intertidal zone (Titus et al. 1998), installation of a seawall and to a lesser extent, a riprap revetment, can severe the connection between upland and intertidal habitat, reflect wave energy and alter sediment transport, and potentially increasing the depth of the intertidal and nearshore subtidal zones reported in several studies (Ruggiero and McDougal 2001, Peregrine 2003).
The loss or disruption of habitat suitable to upland flora species by seawalls and riprap is likely the cause of the reduced biodiversity observed by Strayer and colleagues (2012) and the complete absence of high marsh at seawall sites studied by Bozek and Burdick (2005). The loss of vegetated habitat can alter nutrient cycling in the intertidal (e.g., lower denitrification rates, O'Meara et al. 2015) and reduce pollutant filtration (Reboreda and Cacador 2007), which could have cascading effects via shifts in nutrient availability and the bioaccumulation of toxins in benthic infauna, epibiota, nekton, and birds (Franca et al. 2005). Unlike the studies of seawalls and riprap, studies included in this meta-analysis suggested that breakwaters can decrease the depth of the shoreline via sediment deposition landward of the breakwater, promoting the persistence of intertidal flora such as marsh plants (Currin et al. 2007, Gittman et al. 2014, 2016). Flora abundance effects were only estimated from one short-duration study on marsh dominated by Spartina alterniflora and one short-duration study on the mangrove, Avicennia marina. Both S. alterniflora and A. marina occupy habitat seaward of typical seawall placement, leaving these species vulnerable to loss from reflected, wave-induced erosion or sea-level rise, often termed “coastal squeeze” (Pontee 2013), over longer (e.g., decadal) time scales (Titus et al. 1998), perhaps explaining why the above two short-term studies did not find a difference between shorelines with seawalls versus natural shorelines.
Study limitations
There was significant heterogeneity across organism groups (figures 3, 4, S2, and S3) and studies (figures S4–S9) for all structure types. However, seawalls had a significant negative effect when compared with natural shorelines for at least one metric (biodiversity or abundance) for more than half of all studies and for all organism groups except epibiota. Riprap and breakwater effects were more heterogeneous than seawall effects in both magnitude and direction across organism groups. There were fewer studies on the ecological effects of riprap revetments (n = 9) and breakwaters (n = 8) than seawalls (n = 25), which may have increased heterogeneity in effect sizes and therefore limited our ability (statistical power) to detect the effects of these shore-protection structures relative to seawalls. However, our results do suggest that some organism groups may be adversely affected by riprap (e.g., flora and epibiota) or positively affected by breakwaters (e.g., nekton). Flora, such as marsh plants, seagrasses, and mangroves, were represented by only a single riprap study; however, a study by Patrick and colleagues, which did not meet our criteria to include in the analysis, showed a significant negative correlation between seagrass percent cover and the percentage of riprap shoreline in the estuary (2014). Therefore, research targeting the effects of shore-protection structures on these organisms is needed before more definitive conclusions can be drawn. In general, additional studies examining the ecological effects of riprap revetments and breakwaters are needed to inform future decisions on the consequences of selecting these types of structures.
A majority of studies occurred over a period of 1 year or less and did not replicate their measurements or sampling through time. Observable changes to coastal habitats as a result of shoreline hardening may only be detectable with long-term measurements of multiple characteristics (e.g., Moody et al. 2013) or event-specific monitoring (e.g., storms, Gittman et al. 2014). Studies that track the effects of different shore-protection structures on habitat-forming organisms, such as marsh plants, seagrasses, mangroves, and shellfish reefs, over multiple years to decades would provide valuable insights on the stability and resilience of these shoreline habitats and supported ecosystem services. Using spatially and temporally replicated BACI or beyond-BACI designs (Underwood 1994) may be particularly important for studies of habitat-forming species if changes are a result of direct replacement of habitat with a hard structure (e.g., Bozek and Burdick 2005) or if there is high spatiotemporal variability in the physical environment (e.g., Bilkovic and Mitchell 2013). Finally, studies on the effects of shore-protection structures on the broader suite of ecosystem functions and services (e.g., nutrient cycling, pollutant filtration, carbon sequestration, and sediment stabilization) would allow coastal managers to better compare the overall functionality of shore-protection approaches.
Implications for coastal conservation and management
Shoreline protection will almost certainly continue to be a priority as coastal hazards, such as storms and sea level rise, continute to threaten growing coastal populations and infrastructure. We found that not all shore-protection structures perform equally regarding their ecological impacts on coastal ecosystems. Seawalls have clear negative consequences for coastal biodiversity and habitat quality, and these ecological impacts should be considered by coastal managers and decisionmakers when developing coastal shoreline policies and permitting shoreline protection structures. In addition, a growing body of literature suggests that natural alternatives, such as living or nature-based shore protection or biogenic habitat restoration, can reduce erosion while also enhancing other ecosystem services (e.g., Meyer at al. 1997, Benayas et al. 2009, Scyphers et al. 2011, Gittman et al. 2014). Policymakers and coastal managers should consider the ecological effects of engineered-shore structures when deciding how to best fulfill the need to protect people, property, and infrastructure while also conserving and sustaining coastal ecosystem biodiversity and function.
Supplementary Material
Acknowledgments
We thank the Handling Editor and three anonymous reviewers for the thoughtful comments that greatly improved this manuscript. This research was funded by a contract to R. Gittman from the Pew Charitable Trusts (Award Number12627) and supported by Northeastern University. S. Scyphers was supported by a National Science Foundation SEES Fellowship (OCE-1215825). C. Smith and I. Neylan were supported by a North Carolina Coastal Recreational Fishing License grant to C. Smith and C. Peterson and the University of North Carolina at Chapel Hill.
Supplemental material
The supplemental material is available online at Supplementary Data.
References cited
- Able KW, Vivian DN, Petruzzelli G, Hagan SM. Connectivity among salt marsh subhabitats: Residency and movements of the mummichog (Fundulus heteroclitus) Estuaries and Coasts. 2012;35:743–753. [Google Scholar]
- Adger WN, Hughes TP, Folke C, Carpenter SR, Rockström J. Social–ecological resilience to coastal disasters. Science. 2005;309:1036–1039. doi: 10.1126/science.1112122. [DOI] [PubMed] [Google Scholar]
- Arkema KK, et al. Embedding ecosystem services in coastal planning leads to better outcomes for people and nature. Proceedings of the National Academy of Sciences. 2015;112:7390–7395. doi: 10.1073/pnas.1406483112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baggett LP, et al. Guidelines for evaluating performance of oyster habitat restoration. Restoration Ecology. 2015;23:737–745. [Google Scholar]
- Beck MW, et al. Oyster reefs at risk and recommendations for conservation, restoration, and management. BioScience. 2011;61:107–116. [Google Scholar]
- Benayas JMR, Newton AC, Diaz A, Bullock JM. Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis. Science. 2009;325:1121–1124. doi: 10.1126/science.1172460. [DOI] [PubMed] [Google Scholar]
- Bilkovic DM, Mitchell MM. Ecological tradeoffs of stabilized salt marshes as a shoreline protection strategy: Effects of artificial structures on macrobenthic assemblages. Ecological Engineering. 2013;61:469–481. [Google Scholar]
- Bilkovic DM, Roggero MM. Effects of coastal development on nearshore estuarine nekton communities. Marine Ecology Progress Series. 2008;358:27–39. [Google Scholar]
- Boesch DF, Turner RE. Dependence of fishery species on salt marshes: The role of food and refuge. Estuaries. 1984;7:460–468. [Google Scholar]
- Bozek CM, Burdick DM. Impacts of seawalls on saltmarsh plant communities in the Great Bay Estuary, New Hampshire USA. Wetlands Ecology and Management. 2005;13:553–568. [Google Scholar]
- Broome SW, Seneca ED, Woodhouse WW., Jr Tidal salt marsh restoration. Aquatic Botany. 1988;32:1–22. [Google Scholar]
- Bulleri F. Experimental evaluation of early patterns of colonisation of space on rocky shores and seawalls. Marine Environmental Research. 2005;60:355–374. doi: 10.1016/j.marenvres.2004.12.002. [DOI] [PubMed] [Google Scholar]
- Bulleri F, Chapman MG. Intertidal assemblages on artificial and natural habitats in marinas on the north-west coast of Italy. Marine Biology. 2004;145:381–391. [Google Scholar]
- Bulleri F, Chapman MG, Underwood AJ. Patterns of movement of the limpet Cellana tramoserica on rocky shores and retaining seawalls. Marine Ecology Progress Series. 2004;281:121–129. [Google Scholar]
- Bulleri F, Chapman MG, Underwood AJ. Intertidal assemblages on seawalls and vertical rocky shores in Sydney Harbour, Australia. Austral Ecology. 2005;30:655–667. [Google Scholar]
- Burt J, Bartholomew A, Usseglio P, Bauman A, Sale PF. Are artificial reefs surrogates of natural habitats for corals and fish in Dubai, United Arab Emirates? Coral Reefs. 2009;28:663–675. [Google Scholar]
- Chapman MG, Bulleri F. Intertidal seawalls: New features of landscape in intertidal environments. Landscape and Urban Planning. 2003;62:159–172. [Google Scholar]
- Chapman MG, Underwood AJ. Evaluation of ecological engineering of “armoured” shorelines to improve their value as habitat. Journal of Experimental Marine Biology and Ecology. 2011;400:302–313. [Google Scholar]
- Clynick BG, Chapman MG, Underwood AJ. Effects of epibiota on assemblages of fish associated with urban structures. Marine Ecology Progress Series. 2007;332:201–210. [Google Scholar]
- Currin CA, Delano PC, Valdes-Weaver LM. Utilization of a citizen monitoring protocol to assess the structure and function of natural and stabilized fringing salt marshes in North Carolina. Wetlands Ecology and Management. 2007;16:97–118. [Google Scholar]
- Drexler M, Parker ML, Geiger SP, Arnold WS. Biological assessment of eastern oysters (Crassostrea virginica) inhabiting reef, mangrove, seawall, and restoration substrates. Estuaries and Coasts. 2014;37:962–972. [Google Scholar]
- Dugan JE, Hubbard DM. Ecological responses to coastal armoring on exposed sandy beaches. Shore and Beach. 2006;74:10–16. [Google Scholar]
- Dugan JE, Hubbard DM, Rodil IF, Revell DL, Schroeter S. Ecological effects of coastal armoring on sandy beaches. Marine Ecology. 2008;29:160–170. [Google Scholar]
- Dugan JE, Airoldi L, Chapman MG, Walker SJ, Schlacher T. In: Estuarine and Coastal Structures: Environmental Effects, a Focus on Shore and Nearshore Structures. Treatise on Estuarine and Coastal Science. Wolanski E, McLusky D, editors. Academic Press; 2011. pp. 17–41. [Google Scholar]
- Egger M, Smith GD, Phillips AN. Meta-analysis: Principles and procedures. British Medical Journal. 1997;315:1533–1537. doi: 10.1136/bmj.315.7121.1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- França S, Vinagre C, Caçador I, Cabral HN. Heavy metal concentrations in sediment, benthic invertebrates and fish in three salt marsh areas subjected to different pollution loads in the Tagus Estuary (Portugal) Marine Pollution Bulletin. 2005;50:998–1003. doi: 10.1016/j.marpolbul.2005.06.040. [DOI] [PubMed] [Google Scholar]
- Gillman LN, Wright SD. Mega mistakes in meta-analyses: Devil in the detail. Ecology. 2010;91:2550–2552. doi: 10.1890/09-0339.1. [DOI] [PubMed] [Google Scholar]
- Gittman RK, Fodrie FJ, Popowich AM, Keller DA, Bruno JF, Currin CA, Peterson CH, Piehler MF. Engineering away our natural defenses: An analysis of shoreline hardening in the US. Frontiers in Ecology and the Environment. 2015;13:301–307. [Google Scholar]
- Gittman RK, Peterson CH, Currin CA, Fodrie FJ, Piehler MF, Bruno JF. Living shorelines can enhance the nursery role of threatened estuarine habitats. Ecological Applications. 2016;26:249–263. doi: 10.1890/14-0716. [DOI] [PubMed] [Google Scholar]
- Gittman RK, Popowich AM, Bruno JF, Peterson CH. Marshes with and without sills protect estuarine shorelines from erosion better than bulkheads during a category 1 hurricane. Ocean and Coastal Management. 2014;102:94–102. [Google Scholar]
- Glasby TM, Connell SD, Holloway MG, Hewitt CL. Nonindigenous biota on artificial structures: Could habitat creation facilitate biological invasions? Marine Biology. 2007;151:887–895. [Google Scholar]
- Harris C, Strayer DL, Findlay S. The ecology of freshwater wrack along natural and engineered Hudson River shorelines. Hydrobiologia. 2014;722:233–245. [Google Scholar]
- Hedges LV, Gurevitch J, Curtis PS. The meta-analysis of response ratios in experimental ecology. Ecology. 1999;80:1150–1156. [Google Scholar]
- Hendon JR, Peterson MS, Comyns BH. Spatio–temporal distribution of larval Gobiosoma bosc in waters adjacent to natural and altered marsh-edge habitats of Mississippi coastal waters. Bulletin of Marine Science. 2000;66:143–156. [Google Scholar]
- Kneib RT. Habitat preference, predation, and the intertidal distribution of gammaridean amphipods in a North Carolina salt marsh. Journal of Experimental Marine Biology and Ecology. 1982;59:219–230. [Google Scholar]
- Lam NWY, Huang R, Chan BKK. Variations in Intertidal assemblages and zonation patterns between vertical artificial seawalls and natural rocky shores: A case study from Victoria Harbour, Hong Kong. Zoological Studies. 2009;48:184–195. [Google Scholar]
- Lipcius RN, Seitz RD, Seebo MS, Colón-Carrión D. Density, abundance, and survival of the blue crab in seagrass and unstructured salt marsh nurseries of Chesapeake Bay. Journal of Experimental Marine Biology and Ecology. 2005;319:69–80. [Google Scholar]
- [MEA] Millennium Ecosystem Assessment . Ecosystems and Human Well-Being. Island Press; 2005. [Google Scholar]
- Meyer DL, Townsend EC, Thayer GW. Stabilization and erosion control value of oyster cultch for intertidal marsh. Restoration Ecology. 1997;5:93–99. [Google Scholar]
- Møller AP, Jennions MD. Testing and adjusting for publication bias. Trends in Ecology and Evolution. 2001;16:580–586. [Google Scholar]
- Möller I, et al. Wave attenuation over coastal salt marshes under storm surge conditions. Nature Geoscience. 2014;7:727–731. [Google Scholar]
- Moody RM, Cebrian J, Heck KL., Jr Interannual recruitment dynamics for resident and transient marsh species: Evidence for a lack of impact by the Macondo oil spill. PLOS ONE. 2013;8 doi: 10.1371/journal.pone.0058376. (art. e58376) [DOI] [PMC free article] [PubMed] [Google Scholar]
- [NRC] National Research Council . Mitigating Shore Erosion along Sheltered Coasts. National Academies Press; 2007. [Google Scholar]
- O'Connor MI, Violin CR, Anton A, Ladwig LM, Piehler MF. Salt marsh stabilization affects algal primary producers at the marsh edge. Wetlands Ecology and Management. 2011;19:131–140. [Google Scholar]
- O'Meara T, Thompson SP, Piehler MF. Effects of shoreline hardening on nitrogen processing in estuarine marshes of the US mid-Atlantic coast. Wetlands Ecology and Management. 2015;23:385–394. [Google Scholar]
- Patrick CJ, Weller DE, Li X, Ryder M. Effects of shoreline alteration and other stressors on submerged aquatic vegetation in subestuaries of Chesapeake Bay and the mid-Atlantic coastal bays. Estuaries and Coasts. 2014;37:1516–1531. [Google Scholar]
- Peregrine DH. Water-wave impact on walls. Annual Review of Fluid Mechanics. 2003;35:23–43. [Google Scholar]
- Perkins MJ, Ng TPT, Dudgeon D, Bonebrake TC, Leung KMY. Conserving intertidal habitats: What is the potential of ecological engineering to mitigate impacts of coastal structures? Estuarine, Coastal, and Shelf Science. 2015;167B:1–12. [Google Scholar]
- Peters JR, Yeager LA, Layman CA. Comparison of fish assemblages in restored and natural mangrove habitats along an urban shoreline. Bulletin of Marine Science. 2015;91:125–139. [Google Scholar]
- Peterson CH, Lipcius RN. Conceptual progress towards predicting quantitative ecosystem benefits of ecological restorations. Marine Ecology Progress Series. 2003;264:297–307. [Google Scholar]
- Peterson GW, Turner RE. The value of salt marsh edge vs interior as a habitat for fish and decapod crustaceans in a Louisiana tidal marsh. Estuaries. 1994;17:235–262. [Google Scholar]
- Pontee N. Defining coastal squeeze: A discussion. Ocean and Coastal Management. 2013;84:204–207. [Google Scholar]
- Reboreda R, Cacador I. Halophyte vegetation influences in salt marsh retention capacity for heavy metals. Environmental Pollution. 2007;146:147–154. doi: 10.1016/j.envpol.2006.05.035. [DOI] [PubMed] [Google Scholar]
- Rosenthal R. The file drawer problem and tolerance for null results. Psychological Bulletin. 1979;86:638–641. [Google Scholar]
- Rosenthal R. Meta-Analytic Procedures for Social Research. Sage; 1991. [Google Scholar]
- Ruggiero P, Komar PD, McDougal WG, Marra JJ. Wave runup, extreme water levels and the erosion of properties backing beaches. Journal of Coastal Research. 2001;17:407–419. [Google Scholar]
- Ruiz GM, Carlton JT, Grosholz ED, Hines AH. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. Integrative and Comparative Biology. 1997;37:621–632. [Google Scholar]
- Scyphers SB, Powers SP, Heck KL, Jr., Byron D. Oyster reefs as natural breakwaters mitigate shoreline loss and facilitate fisheries. PLOS ONE. 2011;6 doi: 10.1371/journal.pone.0022396. (art. e22396) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scyphers SB, Gouhier TC, Grabowski JH, Beck MW, Mareska J, Powers SP. Natural shorelines promote the stability of fish communities in an urbanized coastal system. PLOS ONE. 2015;10 doi: 10.1371/journal.pone.0118580. (art. e0118580) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz R, Lipcius R, Olmstead N, Seebo M, Lambert D. Influence of shallow-water habitats and shoreline development on abundance, biomass, and diversity of benthic prey and predators in Chesapeake Bay. Marine Ecology Progress Series. 2006;326:11–27. [Google Scholar]
- Shepard CC, Crain CM, Beck MW. The protective role of coastal marshes: A systematic review and meta-analysis. PLOS ONE. 2011;6 doi: 10.1371/journal.pone.0027374. (art. e27374–11) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Small C, Nicholls R. A global analysis of human settlement in coastal zones. Journal of Coastal Research. 2003;19:584–599. [Google Scholar]
- Sobocinski KL, Cordell JR, Simenstad CA. Effects of shoreline modifications on supratidal macroinvertebrate fauna on Puget Sound, Washington, beaches. Estuaries and Coasts. 2010;33:699–711. [Google Scholar]
- Steffen W, Crutzen J, McNeill JR. The Anthropocene: Are humans now overwhelming the great forces of Nature? Ambio. 2007;36:614–621. doi: 10.1579/0044-7447(2007)36[614:taahno]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Strayer DL, Findlay SEG, Miller D, Malcom HM, Fischer DT, Coote T. Biodiversity in Hudson River shore zones: Influence of shoreline type and physical structure. Aquatic Sciences: Research Across Boundaries. 2012;74:597–610. [Google Scholar]
- Titus J. Rising seas, coastal erosion, and the takings clause: How to save wetlands and beaches without hurting property owners. Maryland Law Review. 1998;57:1279–1318. [Google Scholar]
- Tummers B. DataThief III. 2006. (6 July 2016; http://datathief.org) [Google Scholar]
- Underwood AJ. On beyond BACI: Sampling designs that might reliably detect environmental disturbances. Ecological Applications. 1994;4:3–15. [Google Scholar]
- Unites States Army Corps of Engineers (USACE) Low Cost Shore Protection. USACE; 2001. (6 July 2016; http://chl.erdc.usace.army.mil/Media/2/4/1/sect54eng.pdf) [Google Scholar]
- Vetter D, Rücker G, Storch I. Meta-analysis: A need for well-defined usage in ecology and conservation biology. Ecosphere. 2013;4:1–24. [Google Scholar]
- Viechtbauer W. Conducting meta-analyses in R with the metafor package. Journal of Statistical Software. 2010;36:1–48. (6 July 2016; www.jstatsoft.org/v36/i03) [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.