

Supplemental Digital Content is available in the text.
Keywords: adenoma, aldosterone, extracellular matrix, hypertension, Wnt signaling pathway
Abstract
Primary aldosteronism is a common cause of hypertension, which becomes refractory if undiagnosed, but potentially curable when caused by an aldosterone-producing adenoma (APA). The discovery of somatic mutations and differences in clinical presentations led to recognition of small but common zona glomerulosa (ZG)–like adenomas, distinct from classical large zona fasciculata–like adenomas. The inverse correlation between APA size and aldosterone synthase expression prompted us to undertake a systematic study of genotype–phenotype relationships. After a microarray comparing tumor subtypes, in which NPNT (nephronectin) was the most highly (>12-fold) upregulated gene in ZG-like APAs, we aimed to determine its role in physiological and pathological aldosterone production. NPNT was identified by immunohistochemistry as a secreted matrix protein expressed exclusively around aldosterone-producing glomeruli in normal adrenal ZG and in aldosterone-dense ZG-like APAs; the highest expression was in ZG-like APAs with gain-of-function CTNNB1 mutations, whose removal cured hypertension in our patients. NPNT was absent from normal zona fasciculata, zona fasciculata–like APAs, and ZG adjacent to an APA. NPNT production was regulated by canonical Wnt pathway, and NPNT overexpression or silencing increased or reduced aldosterone, respectively. NPNT was proadhesive in primary adrenal and APA cells but antiadhesive and antiapoptotic in immortalized adrenocortical cells. The discovery of NPNT in the adrenal helped recognition of a common subtype of APAs and a pathway by which Wnt regulates aldosterone production. We propose that this arises through NPNT’s binding to cell-surface integrins, stimulating cell–cell contact within glomeruli, which define ZG. Therefore, NPNT or its cognate integrin could present a novel therapeutic target.
Five percent to 13% of all hypertension and 20% of resistant hypertension can be attributed to primary aldosteronism, of which unilateral aldosterone-producing adenoma (APA) is the most common curable cause.1,2 Early detection of APA is important because of significant increases in cardiovascular morbidity and mortality: congestive cardiac failure and ischemic heart disease are 2 to 5 times more prevalent,3 with an increase in 14-year mortality in these patients compared with matched patients with essential hypertension.4 Whether this additional risk is due directly to aldosterone excess, independent of high blood pressure, or reflects greater average duration of hypertension before diagnosis, clinical outcome is considered to benefit from prompt recognition and removal of APAs.5
Over the past decade, new molecular stratifications have enabled the recognition of a group of smaller zona glomerulosa (ZG)–like APAs.6–8 Compared with the classical large lipid-laden zona fasciculata (ZF)–like APA with mutations in inward rectifier potassium channel 4 (KCNJ5),6 not only is this ZG-like subtype of APA histologically and biochemically different, it also harbors hallmark somatic mutations in genes encoding a subunit of the voltage-gated calcium channel (CACNA1D),8 Na+/K+-ATPase (ATP1A1),8,9 Ca2+-ATPase (ATP2B3),9 or the Wnt pathway mediator β-catenin (CTNNB1).10 Biochemically, these smaller ZG-like APAs have a higher capacity for aldosterone production; semiquantitative analysis of immunohistochemical staining has revealed that CYP11B2 score is inversely correlated with tumor size and volume.11,12 In addition, ZG-like APAs are more responsive to angiotensin II, with higher levels of type 1 angiotensin II receptor mRNA.13,14 However, because of their small size, they are readily overlooked on cross-sectional imaging because computed tomography is unable to reliably detect adrenal tumors <5 to 6 mm.15 We hypothesized that specific gene products are responsible for the increased hormone production in these aldosterone-dense APAs and can potentially be used as a diagnostic biomarker when imaging proves inconclusive.
In seeking transcriptomic evidence to identify possible pathways of aldosterone production specific to the more compact, aldosterone synthase–dense APAs, we compared CACNA1D/ATP1A1-mutant with KCNJ5-mutant APAs.8 Extracellular matrix (ECM) gene NPNT (nephronectin) was found to be the most upregulated, by 12-fold, in the former and confirmed a categorical difference between APA genotypes by immunohistochemistry. Discovered in the kidney in 2001,16 NPNT was recently found to be a downstream Wnt target in the epidermis.17 This may be relevant in the adrenal, where normal adrenocortical development and steroidogenic activity of the ZG are dependent on the canonical Wnt pathway.18
Further examination of NPNT’s distribution presented in this article led us to hypothesize a key role in bringing ZG cells together to form functional units for aldosterone production through intercellular communication. This is supported by the observation, in rat ZG cells, of numerous tight junctions likely to be important in the establishment of electrical coupling,19 and the more recent discovery, only in intact adrenal slices, of oscillating membrane potentials regulating aldosterone secretion.20
In this study, we have found that NPNT is directly involved in the physiological secretion of aldosterone in the adrenal. By using both cell lines and primary human adrenal cells, we have also uncovered a previously unknown role of NPNT in adhesion and cell survival.
Methods
Human Subjects
Human adrenal tissues from patients who underwent adrenalectomy after being diagnosed with unilateral APA or pheochromocytoma were obtained from Cambridge University Hospitals’ Human Research Tissue Bank postsurgery at Addenbrooke’s Hospital, Cambridge, United Kingdom. All tissues were obtained with approval from the Cambridgeshire Research Ethics Committee with written informed consent before surgery. Further details are provided in the online-only Data Supplement and clinical features in Table S1 in the online-only Data Supplement.
Cell Culture
Human adrenocortical carcinoma H295R cells were obtained from the American Type Culture Collection (ATCC CRL-2128), and grown in DMEM/Nutrient F-12 Ham supplemented with 10% FBS, 100 U penicillin, 0.1 mg/mL streptomycin, 0.4 mmol/L l-glutamine, and insulin-transferrin-selenium at 37°C in 5% CO2. Human embryonic kidney cells were obtained from the American Type Culture Collection (ATCC CRL-1573) and grown in DMEM supplemented with 10% FBS.
Gene Overexpression and Silencing
Gene overexpression was performed using lipid-mediated cell transfection lipofectamine 3000 (Thermo Fisher), whereas gene silencing was achieved using DharmaFECT 1 lipid transfection reagent (Dharmacon), both according to manufacturer’s instructions. Cells were harvested for analysis of mRNA and protein expression after 48 hours. Further details are provided in the online-only Data Supplement.
RNA Extraction, Reverse Transcription, and Quantitative Real-Time Polymerase Chain Reaction
Fifty to 100 mg of tissue or 1×105 cells were used for each RNA extraction. Further details are provided in the online-only Data Supplement. Quantitative real-time polymerase chain reaction was performed using TaqMan ABI probes (Applied Biosystems) for NPNT (Hs01568126), ITGB1 (Hs00559595), and BCL2 (Hs00608023). CYP11B2 expression was quantified using custom-made TaqMan probes (Invitrogen) previously validated for specificity.21
Immunohistochemistry
Immunohistochemistry was performed using the peroxidase–antiperoxidase method on fresh frozen human tissue. In cases where fresh frozen tissue was unavailable, immunohistochemistry was performed on formalin-fixed, paraffin-embedded adrenal sections (4 μm) using an automated immunostainer with cover tile technology (Bond-III system; Leica Biosystems). NPNT antibody (HPA003711, Sigma; 1:50 dilution) and CYP11B2 antibody (custom mouse antihuman antibody from Dr Celso E. Gomez-Sanchez)22 were used as the primary antibodies. Further details are provided in the online-only Data Supplement.
Aldosterone Measurement
Supernatant from cultured cells was used for aldosterone quantification using the homogenous time resolved fluorescence assay (Cisbio assays) based on the fluorescence resonance energy transfer technology, according to manufacturer’s instructions. The final fluorescence readout was conducted using a Pherastar FS microplate reader (BMG Labtech). Aldosterone concentrations were then normalized to total cell protein, quantified by the bicinchoninic acid protein assay (Pierce Biotechnology).
Firefly/Renilla Luciferase Assay
To measure the activity of Wnt transcriptional complex T-cell factor/lymphoid enhancer factor (TCF/LEF), firefly luciferase and renilla luciferase activities were measured 48 hours after cotransfection with the Dual-Glo Luciferase Assay System (Promega) and normalized to the empty pCMV6 vector as described in the manufacturer’s protocol. Canonical Wnt signaling was quantified using the Cignal TCF/LEF reporter (luc) kit (SABiosciences).
Cell Confluency and Cytotoxicity Assay
To measure changes in H295R cell confluency post-NPNT silencing, time-lapsed images were obtained using an Incucyte system (Essen BioScience). To differentiate changes in cell proliferation from cytotoxicity, cell-impermeant cyanine dimer nucleic acid stain YOYO-1 (Y3601; Life Technologies) was used. Further details are provided in the online-only Data Supplement.
Annexin V–Propidium Iodide Dual Stain
To assess apoptosis over time, cells were double labeled with annexin V-APC (550474; BD Pharmingen) and propidium iodide (Sigma). After silencing, adherent cells were trypsinized, added to any detached cells in the supernatant as previously described,23 stained with annexin V–propidium iodide, and analyzed with the FACSCanto II flow cytometer (Becton-Dickinson). Further details are provided in the online-only Data Supplement.
Xcelligence Cell Impedance Measurement and Hoechst Stain Assay
To evaluate changes in adhesion in response to NPNT, wells of an E-Plate 16 (ACEA Biosciences) were precoated with PBS, or 10 μg/mL of BSA, NPNT, or laminin for 1 hour at 37°C as previously described.24 Cell adhesion was also measured by Hoechst dye quantification of cells remaining on precoated wells post-wash. Further details are provided in the online-only Data Supplement.
Proteins and Chemicals
Proteins used in this study were BSA (A9576; Sigma), NPNT (4298-NP-050; R&D systems), fibronectin (FC010; Millipore), and laminin (AG56P; Millipore). The selective porcupine inhibitor, LGK-974 (1 μmol/L; Selleck Chemicals), as used previously,25 was used to analyze the effect of blocking all Wnt secretion.
Statistical Analysis
Results are expressed as mean values with SEM and compared using the 2-sided Student t test or by 1-way ANOVA followed by Tukey post hoc test. Significance level of P<0.05 was considered to indicate statistical significance. Statistical analysis was performed using Graphpad Prism (Graphpad Software).
Results
NPNT Is Selectively Expressed in the Subtype of Smaller ZG-Like APAs and Able to Distinguish Between the 2 APA Classes
Microarray analysis revealed NPNT to be 12.2-fold upregulated in the smaller ZG-like APAs with higher aldosterone synthetic capacity (Figure 1A). Validation with quantitative real-time polymerase chain reaction revealed a 9-fold difference; both ZG-like APAs and adrenocortical carcinomas (ACCs) had significantly elevated levels of NPNT compared with the ZF-like APAs (Figure 1B). The 2 APAs with the highest levels of NPNT harbored gain-of-function mutations in the Wnt gene CTNNB1.
Figure 1.
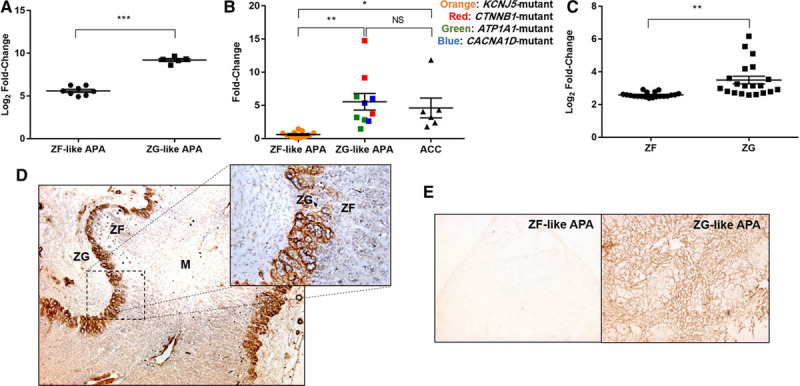
NPNT (nephronectin) is selectively expressed in normal adrenal zona glomerulosa (ZG), ZG-like aldosterone-producing adenomas (APAs), and adrenocortical carcinomas (ACCs). A, Microarray analysis of NPNT, comparing 8 zona fasciculata (ZF)-like adenomas with 5 ZG-like adenomas. B, Quantitative real-time polymerase chain reaction of NPNT, on mRNA extracted from 11 ZF-like APAs and 10 ZG-like APAs differentiated based on their genotypic mutations, as well as 6 ACCs. C, Microarray analysis of NPNT, comparing 20 paired ZF and ZG (each pair from the same patient), isolated via laser capture microdissection. D, Representative immunohistochemistry of NPNT showing selective extracellular localization in the ZG of adrenal adjacent to a pheochromocytoma (4× magnification; inset: 20× magnification). E, Representative immunohistochemistry of NPNT comparing staining between ZF-like APA and ZG-like APA mounted on the same slide (4× magnification). In (A)–(C), bars represent mean expression per group±SEM. Statistical analyses were conducted by Student t test (A and C) or 1-way ANOVA followed by Tukey post hoc test (B). *P<0.05; **P<0.005; ***P<0.0005. M indicates medulla; and NS, not significant.
In a further microarray comparing the ZG and ZF of 20 human adrenals isolated via laser capture microdissection,26 NPNT was on average 2-fold more highly expressed in the outer, aldosterone-producing zone of the ZG (Figure 1C). This selective expression was evident in protein immunohistochemistry, in which NPNT staining is localized exclusively to the ZG (Figure 1D; Figure S1). When mounted on the same slide, NPNT also makes it easy to differentiate between the 2 APA subtypes as there is negligible staining in ZF-like APAs (Figure 1E; Figure S2). In all cases, NPNT expression appeared extracellular and strikingly periglomerular, surrounding clusters of cells.
NPNT Expression Corresponds to Aldosterone Synthase CYP11B2 Expression
Although the ZG is the zone of physiological aldosterone production, CYP11B2 staining is patchy.22 The overall analysis of 20 normal adrenal and APA samples revealed a significant positive correlation between NPNT and CYP11B2 encoding aldosterone synthase (r=0.82; P<0.0001; Figure 2A). At the protein level, areas of CYP11B2 expression also corresponded consistently with NPNT staining (Figure 2B; Figure S3). We also compared the ZG expression of genes in adrenal adjacent to a pheochromocytoma versus that next to an APA (ie, in a state of aldosterone excess). NPNT was 3.6-fold upregulated in ZG, compared with ZF, when adjacent to a pheochromocytoma, but diminished or absent when adjacent to an APA (Figure 2C). The same observations were made at the protein level (Figure 2D). Similarly, CYP11B2 was 2-fold upregulated in ZG next to a pheochromocytoma versus next to an APA and 7.8-fold higher on quantitative real-time polymerase chain reaction.
Figure 2.
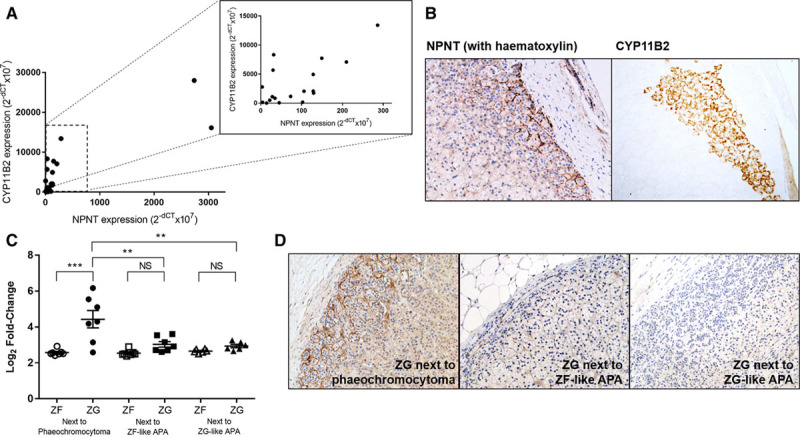
NPNT (nephronectin) expression corresponds to CYP11B2 expression, with presence of negative feedback. A, Strong positive linear correlation between NPNT and CYP11B2 expression in 10 pairs of adenomas and their adjacent adrenal, r(18)=0.8273; P<0.0001. Inset: correlation plot excluding the 2 samples with highest NPNT expression, r(16)=0.6806; P=0.0019. Statistical analysis was conducted by Pearson product-moment correlation. B, Representative immunohistochemistry of NPNT and CYP11B2 in corresponding zona glomerulosa (ZG) areas in serial sections of the same adrenal tissue (20× magnification). C, Microarray expression of NPNT in 7 paired ZG and zona fasciculata (ZF) samples adjacent to a pheochromocytoma and 13 pairs next to an aldosterone-producing adenoma (APA). Bars represent mean expression per group±SEM. Statistical analysis was conducted by 1-way ANOVA followed by Tukey post hoc test. *P<0.05; **P<0.005; ***P<0.0005; NS, not significant. D, Negative feedback shown by representative immunohistochemistry of NPNT, in ZG adjacent to pheochromocytoma compared with that adjacent to ZF-like or ZG-like APAs (20× magnification).
NPNT Drives Aldosterone Production
NPNT overexpression in H295R cells increased aldosterone synthesis compared with control (Figure 3A). Similarly, silencing NPNT by >75% reduced hormone production (Figure 3B). NPNT has been previously found to bind strongly and specifically to integrin receptor α8β1,16 with ≈100-fold higher affinity compared with other RGD motif-containing proteins such as fibronectin or vitronectin.27 In our study, silencing of ITGB1, encoding integrin subunit β1, by ≈80%, caused a similar reduction in aldosterone production comparable to silencing of NPNT (Figure 3C). This receptor silencing was accompanied by a 3.8-fold increase in NPNT mRNA expression (P=0.01).
Figure 3.
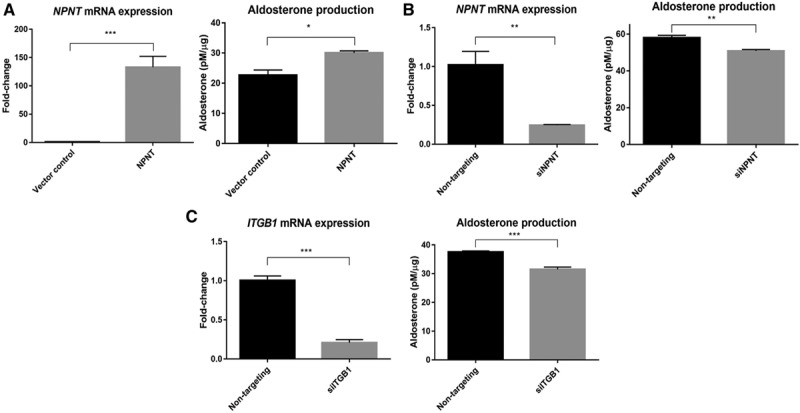
NPNT (nephronectin) drives aldosterone production, likely through receptor ITGB1. A, NPNT overexpression increases protein-normalized aldosterone production, compared with vector control (n=4). B, NPNT silencing decreases protein-normalized aldosterone production, compared with nontargeting control (n=4). C, ITGB1 silencing decreases protein-normalized aldosterone production, compared with nontargeting control (n=4). Bars represent mean expression per group±SEM. Statistical analyses were conducted by Student t test. *P<0.05; **P<0.005; ***P<0.0005.
NPNT Is a Wnt Target Gene and Produces Aldosterone via This Pathway
NPNT was found to be a Wnt/β-catenin target gene in skin, being induced by Wnt activation in the bulge and hair germ cells.17 We investigated the influence of Wnt on NPNT mRNA expression in H295R cells, using plasmids modulating Wnt signaling. To activate the Wnt canonical pathway, ΔN47 β-catenin, a strong constitutive inducer encoding an N-terminally truncated form of β-catenin resistant to proteolysis,28 was expressed. This led to a near doubling of NPNT expression levels. Conversely, to inhibit β-catenin–dependent gene transcription, we expressed ΔN-TCF4, a Wnt constitutive repressor, because of an N-terminally truncated, dominant-negative TCF4 protein lacking the β-catenin interaction domain.29 Wnt repression caused NPNT mRNA levels to halve (Figure 4A). To investigate the potential for negative feedback of NPNT on its own release, TCF/LEF activity was measured after changes in NPNT expression. Overexpressing NPNT caused a reduction in Wnt transcriptional activity, whereas silencing NPNT had the opposite effect (Figure 4B).
Figure 4.
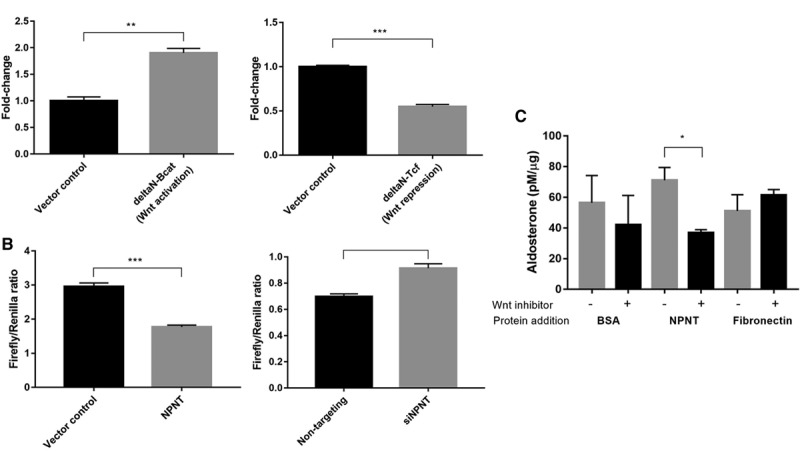
NPNT (nephronectin) is a Wnt target gene and produces aldosterone via Wnt. A, Constitutive Wnt activation (ΔN-Bcat) induces NPNT mRNA expression, whereas constitutive Wnt repression (ΔN-TCF4) decreased NPNT expression compared with vector control (n=3). B, Wnt transcriptional complex T-cell factor/lymphoid enhancer factor (TCF/LEF) activity decreased in NPNT-overexpressed and increased in NPNT-silenced samples, as measured by firefly/renilla luciferase assay (n=6; n=4). C, Wnt inhibitor LGK-974 attenuated the increase in protein-normalized aldosterone production with addition of NPNT protein, compared with negative controls BSA and fibronectin (n=3). Bars represent mean expression per group±SEM. Statistical analyses were conducted by Student t test. *P<0.05; **P<0.005; ***P<0.0005.
In cases where NPNT protein was added to the cell medium, on addition of Wnt inhibitor LGK-974, which blocks Wnt pathways upstream by binding the Wnt chaperone, porcupine, aldosterone production was nearly diminished by half compared with the controls (Figure 4C).
NPNT Is Proadhesive in Normal Adrenal and APA Cells
NPNT promotes cell adhesion in kidney mesangial cells30 and cardiomyocytes.31 Cell impedance was recorded as a measure of cell adhesion in wells coated with PBS, negative control BSA, NPNT, or positive control laminin. NPNT was proadhesive in human embryonic kidney cells, normal primary adrenal cells, and cells cultured from ZF-like and ZG-like APAs (Figure 5Ai through Aiv). These findings are consistent with the hypothesized physiological role of NPNT in adrenal cell clustering for aldosterone production.
Figure 5.
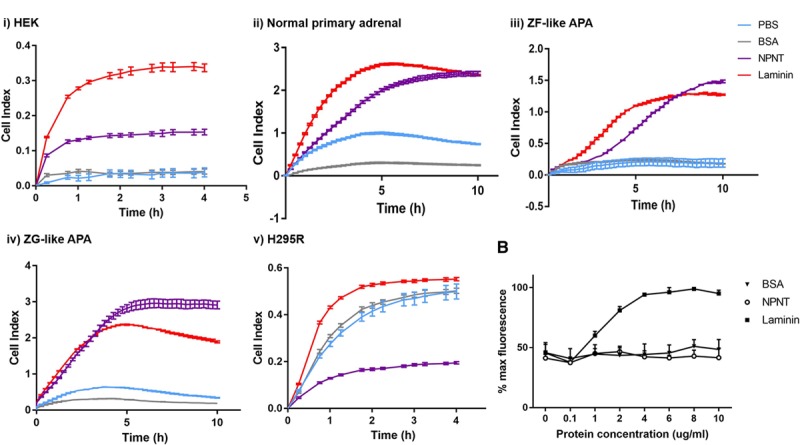
NPNT (nephronectin) is proadhesive in normal adrenal and aldosterone-producing adenoma (APA) cells, but antiadhesive in H295R. A, NPNT is proadhesive in (i) HEK, (ii) normal primary adrenal, (iii) zona fasciculata (ZF)–like APA, and (iv) zona glomerulosa (ZG)–like APA, but antiadhesive in (v) H295R, as demonstrated by wells precoated with PBS, BSA, NPNT, or laminin, and measured by Xcelligence cell impedance as cell index over time (4 h for cell lines, 10 h for primary adrenal cells; n=2 for ZF-like APA and ZG-like APA, n=4 for the rest). B, NPNT is antiadhesive in H295R cells, as confirmed by Hoechst fluorescent stain assay measuring % maximum fluorescence as a representation of number of cells adhered to well with increasing concentrations of BSA, NPNT, and laminin precoating (n=3 for each protein at each concentration). Bars represent mean values per group±SEM.
Intriguingly, NPNT had the opposite effect on H295R cells, with cell index reaching <50% of that in BSA-coated wells even after 4 hours (Figure 5Av). This suggested that NPNT was antiadhesive toward H295R cells, with cells tending to repel from the well surface. To reaffirm this finding, Hoechst stain assays were performed independently, with increasing concentrations of matrix coating on wells. Post-wash, there was a concentration-dependent increase in number of cells remaining (as measured by fluorescence) in positive control laminin–coated wells. However, in NPNT-coated wells, the number of remaining cells showed no significant difference from that in BSA-coated wells at every protein concentration, indicating only background levels of adhesion (Figure 5B).
NPNT Protects H295R Cells From Apoptosis
Cell confluency was 41% in the nontargeting controls compared with 26% in the NPNT-silenced samples at 48 hours, as observed using kinetic live-cell imaging (Figure 6A). Kinetic measurement of cytotoxicity via the YOYO-1-iodide reagent revealed >3-fold increase in fluorescent (dead) cells when NPNT was silenced (Figure 6B). Annexin V–propidium iodide dual assay was used to monitor H295R cell staining over time. NPNT-silenced cells exhibited greater apoptosis over time, whereas samples with nontargeting siRNA consistently showed basal levels of apoptosis (Figure 6C).
Figure 6.
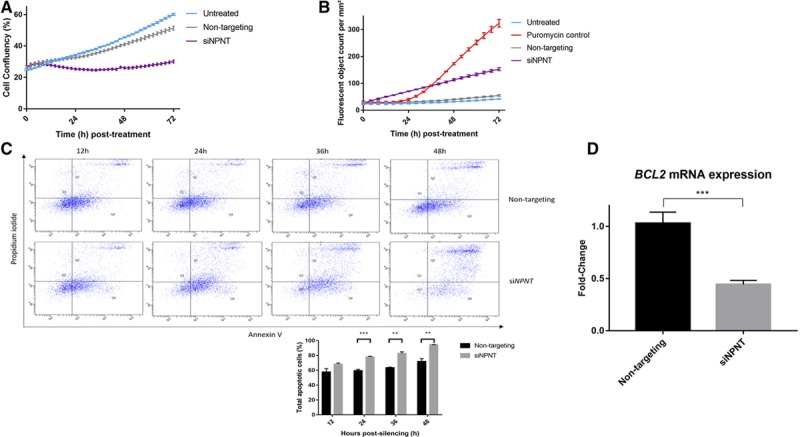
NPNT (nephronectin) protects H295R cells from apoptosis. A, NPNT silencing causes cell confluency to remain low over 72 h, as measured by kinetic live-cell imaging in groups that are untreated or treated with nontargeting siRNA (n=3; P<0.0001 between siNPNT and nontargeting). B, NPNT silencing drives active cell death, as shown by kinetic measurement of cytotoxicity using YOYO-1 iodide reagent, comparing number of fluorescent (dead) cells in groups that are untreated, treated with positive control puromycin, nontargeting siRNA, or specific siRNA against NPNT (n=4; P<0.0001 between siNPNT and nontargeting). C, NPNT silencing causes cell death via apoptosis, as shown by flow cytometric analysis using annexin V-APC and propidium iodide double staining in groups that are treated with nontargeting siRNA or specific siRNA against NPNT. Quadrant analysis of the gated cells in FL-1 versus FL-2 channels was from 10 000 events. Annexin V+/PI− (lower right quadrant) areas stand for early apoptotic cells, and Annexin V+/PI+ (upper right quadrant) areas stand for late apoptotic or necrotic cells. Graph below shows percentage of total apoptotic cells at 12, 24, 36, and 48 h post-silencing. D, NPNT silencing causes apoptosis through reduction of BCL2, a prosurvival factor in the intrinsic apoptotic pathway, as shown by mRNA expression in groups that are treated with nontargeting siRNA or specific siRNA against NPNT (n=6). Bars represent mean expression per group±SEM. Statistical analyses were conducted by Student t test. *P<0.05; **P<0.005; ***P<0.0005.
A prosurvival factor in the intrinsic apoptotic pathway,32 transcription of BCL2 (B-cell lymphoma 2) was greatly suppressed (>56%) in NPNT-silenced (siNPNT) cells (Figure 6D).
Discussion
The ZG of human adrenal is unusual among endocrine organs in that few cells produce its signature hormone, aldosterone, and yet there is a high incidence of APA occurrence, which is a common curable cause of hypertension. The ZG has likely evolved primarily to protect mammals from the scarcity of salt that has been the prevailing natural state, including for most of human history. It clearly also has the ability to adapt to chronic salt excess, to which the sparsity of aldosterone synthase is usually attributed,25 but perhaps imperfectly, and hence the frequent somatic mutations permitted by high rates of ZG cell migration and renewal. Our discovery of NPNT, and its putative roles, in the adrenal may help to explain the link between physiology and pathology.
We first discovered NPNT in the adrenal as the most upregulated gene in the smaller ZG-like APAs with higher aldosterone synthetic capacity, harboring mutations of CACNA1D or ATP1A1, when compared with those with a ZF-like phenotype and mutations of KCNJ5. We also noted the exquisite ZG selectivity of its distribution in normal adrenal cortex.8 These original findings have been reproduced by Åkerström et al.33 Interest in finding a mechanistic link between NPNT expression and blood pressure control has also been raised by a large-scale genome-wide association study in which a common single-nucleotide polymorphism in NPNT was associated with blood pressure regulation.34
The investigations that we now report show that NPNT is not just a marker of ZG cells but plays an essential role in normal adrenal physiology as a periglomerular ECM protein. Coupled with its steroidogenic and proadhesive properties, this is consistent with a physiological role in adrenal cell clustering to form functional aldosterone-producing units in the ZG. Our work provides evidence that a matrix protein can play a role in driving hormone synthesis and, together with the regulation of ZG cell behavior, helps us understand why the ZG may be structured as it is and why tumors that resemble ZG are able to have a higher density of aldosterone production.
The ZG-selective staining suggests that the ECM of which NPNT is a major constituent supports zone-specific cellular behavior. In 2012, Hu et al20 made the crucial observation that isolated mouse ZG cells are too hyperpolarized to permit calcium entry; however, ZG cells in an intact adrenal slice generate spontaneous membrane oscillations sufficient for recurrent Ca2+ signals, whose periodicity could sustain aldosterone production. Therefore, these findings suggest that the aldosterone-producing ability of adrenal cells requires them to coexist in whole glomeruli, and this process could be regulated by NPNT.
Although the ZG is the zone of physiological aldosterone production, this is not performed by all cells in the area, as evidenced by previous reports of patchy CYP11B2 staining.22 But, the mechanism underlying why some ZG cells express CYP11B2, whereas others do not, is yet to be determined. Together with the consistent correlation between NPNT and CYP11B2 staining, and evidence showing NPNT increases aldosterone production, we propose that the role of this matrix protein is to cluster ZG cells together to form a functional unit as indicated by its periglomerular staining. This is supported by the recent proposal that Ca2+ and Ca2+-activated K+ channels in ZG cells, when grouped in rosette structures, act as a pacemaker generating the oscillations that regulate aldosterone production.35 On NPNT silencing, although the cell-corrected fall in aldosterone appears relatively small (Figure 3B), the absolute change consequent on reduction in both secretion and cell number is more substantial and likely to have considerable impact in intact ZG.
In addition, our own finding of NPNT as an exquisitely selective ZG protein, 3.6-fold upregulated in ZG compared with ZF when adjacent to a pheochromocytoma, but diminished or absent when adjacent to an APA, suggests the disappearance of NPNT when physiological aldosterone secretion is not required. Analogous negative feedback has been reported in ZG adjacent to APA, where both subtypes of 3β-hydroxysteroid dehydrogenase-isomerase (3β-HSD), responsible for synthesizing progesterone from pregnenolone, were found to be suppressed.36
Our results also showed that NPNT promotes adhesion in normal adrenal cells and in both subtypes of APAs. This is consistent with previous reports of NPNT expression in hepatitis, inducing the development of granuloma-like cell clusters of hepatocytes.37 Cardiomyocytes grown on NPNT not only exhibited increased adhesion but also expressed high amounts of connexin-43 along their intercellular junctions, indicating well-established intercellular communication, and couple electrically with each other resulting in synchronous beating.31
The link between increased cell adhesion and aldosterone production lies within the Wnt signaling system. Intracellular Wnt signaling diversifies into several major pathways, including (1) the β-catenin/TCF–LEF pathway (canonical Wnt), which activates nuclear target genes; (2) the planar cell polarity pathway; and (3) the Wnt/Ca2+ pathway, with the last 2 being classified as the noncanonical Wnt pathways.38 Although NPNT expression is itself under control of the canonical pathway, the noncanonical Wnt/planar cell polarity pathway is likely to be involved in mediating NPNT’s control of cell–cell adhesion and the localized assembly of ECM (Figure 4C).39 It has been shown that an aberrant planar cell polarity pathway leads to disruption of integrin β1–mediated interactions and, in turn, disorganization of the ECM.40 In addition, in line with the proposed role of NPNT, integrin β1 expression was reported to be crucial for adhesion of endothelial cells, with its absence causing focal adhesions to become short and disorganized.41
Although our experiments concentrated on the physiological roles of NPNT in normal adrenal and benign APAs, its antiadhesive and antiapoptotic effect on H295R cells have drawn our attention to a potential role in malignancy. ACCs, although rare, are much more devastating than APAs. NPNT has been shown to confer apoptosis resistance in H295R cells by modulating the expression of prosurvival protein BCL2, whose role is to block caspase activation.32 This detachment of cells from the ECM often results in apoptotic cell death known as anoikis.42 Excess secretion of ECM components suppresses the physiological induction of anoikis in maintaining normal tissue architecture43 and could explain the high levels of NPNT expression in adrenocortical carcinoma and immortalized H295R cells (Figure 1B).
However, benign APAs are much commoner than ACCs, and the main translational potential of NPNT may lie in its use as a diagnostic marker in patients with subcentimeter APAs, whose computed tomographic scan and adrenal vein sampling results are inconclusive. Recent publications have reported only 50% concordance between adrenal vein sampling and computed tomography, with >10% of patients aged >50 years lateralizing to opposite sides.44,45 Therefore, in vivo measurement of adrenal NPNT could be a more accurate predictor of APA presence and even genotype, as the secreted protein may be measurable in adrenal vein samples routinely collected for unilateral APA diagnoses.
A limitation of the work to date is that the H295R cell line is not a perfect model for native ZG cells or ZG-like APAs, even though H295R cells have proven invaluable in studying mechanisms involved in the physiological regulation of aldosterone production.46 Therefore, in the adhesion studies, primary adrenal cell types were also used. In addition, the H295R cell line is already known to harbor a CTNNB1 mutation47 with high levels of NPNT; so, whenever applicable, silencing NPNT was prioritized. Furthermore, our work on malignant cells in this study has been performed only on H295R, because of the lack of other human adrenal carcinoma cell lines amenable to transfection. A further limitation is that centripetal migration of adrenocortical cells cannot be studied in human adrenal. Because of the dispersed-cell nature of the experiments which we can currently undertake, the effects of NPNT’s various roles in the adrenal may be underestimated in this study. The full impact of NPNT will become apparent during experiments on intact adrenal with preserved cell–cell contacts. Future work may involve a ZG-selective conditional-knockout of NPNT, by crossing a floxed-NPNT mouse48 with an aldosterone synthase-Cre recombinase mouse.49
In conclusion, we have discovered NPNT to be an exquisitely ZG-selective ECM protein in the adrenal. The distribution of NPNT defines the glomeruli anatomically, and its actions explain the critical role of glomerular structure in regulation of aldosterone production and hence blood pressure control. The high levels of NPNT in the smaller aldosterone-dense ZG-like APA subtype, as well as in ACCs, suggest future potential as a diagnostic marker or target for novel therapies.
Perspectives
Primary aldosteronism is the most common secondary cause of hypertension, of which APAs make up 30% to 50% of cases, with hypertension potentially curable by adrenalectomy. Recently, the discovery of somatic mutations in CACNA1D/ATP1A1/ATP2B3/CTNNB1 characterize a subtype of small ZG-like APAs that are also histologically and biochemically distinct from the classical large KCNJ5-mutant ZF-like APAs. Investigations into transcriptomic differences between the 2 subtypes revealed NPNT, encoding the matrix protein nephronectin, to be the most upregulated gene in ZG-like versus ZF-like APAs. Subsequent investigations have shown NPNT not only to be just a biomarker of ZG-type APAs but also to play an important role in normal ZG. Found to be under canonical Wnt control, the distribution and effects of NPNT suggest that it defines the anatomy and function of normal adrenal glomeruli, driving steroidogenesis and adhesion physiologically. In contrast, in immortalized adrenocortical carcinoma cells, NPNT exerts antiadhesive and antiapoptotic effects. Clinical measurement of NPNT in adrenal vein blood may have application in diagnosis of unilateral APAs. Complete cure of hypertension, on removal of CTNNB1-mutant APAs, may be predicted through unilateral detection of secreted proteins such as NPNT during adrenal vein sampling. Apart from its potential as a diagnostic marker, the high levels of NPNT in the smaller aldosterone-dense ZG-like APA subtype, and in ACCs, make it an attractive molecular target for novel therapies.
Acknowledgments
We thank the Cambridge NIHR BRC Cell Phenotyping Hub, in particular, Anna Petrunkina Harrison and Simon McCallum and for their advice and support in flow cytometry.
Sources of Funding
This research was funded by grants from the National Institute for Health Research (NIHR) Senior Investigator award (NF-SI-0512-10052) to M.J. Brown. A.E.D. Teo is supported by the Agency for Science, Technology and Research (A*STAR) Singapore. This study is also supported by Wellcome Trust Translational Medicine and Therapeutics award to M.J. Brown (085686/Z/08/A). S. Garg is supported by the British Heart Foundation (FS/14/75/31134). J. Zhou is supported by the Cambridge Overseas Trust. Additional support was provided by the NIHR Cambridge Biomedical Research Centre (Cardiovascular and Metabolic, and Human Tissue Bank).
Disclosures
None.
Supplementary Material
Footnotes
The online-only Data Supplement is available with this article at http://hyper.ahajournals.org/lookup/suppl/doi:10.1161/HYPERTENSIONAHA.117.09156/-/DC1.
Novelty and Significance
What Is New?
NPNT (nephronectin), selectively expressed in the zona glomerulosa of human adrenal cortex and of aldosterone-producing adenomas arising from this, is shown to have roles in steroidogenesis, cell adhesion, and protection of adrenocortical cells from apoptosis.
What Is Relevant?
Aldosterone-producing adenomas are greatly underdiagnosed, one reason being the small size of those appearing to arise in the zona glomerulosa of adrenal cortex. Recognition of their separate identity and of the role of NPNT in steroidogenesis will encourage clinicians to give greater consideration to small adenomas.
Summary
NPNT is an exquisitely selective, Wnt-driven zona glomerulosa protein, which seems to play an important role in steroidogenesis, by promoting adhesion of ZG cells and preventing apoptosis.
References
- 1.Mulatero P, Stowasser M, Loh KC, Fardella CE, Gordon RD, Mosso L, Gomez-Sanchez CE, Veglio F, Young WF., Jr. Increased diagnosis of primary aldosteronism, including surgically correctable forms, in centers from five continents. J Clin Endocrinol Metab. 2004;89:1045–1050. doi: 10.1210/jc.2003-031337. doi: 10.1210/jc.2003-031337. [DOI] [PubMed] [Google Scholar]
- 2.Rossi GP, Bernini G, Caliumi C, et al. PAPY Study Investigators. A prospective study of the prevalence of primary aldosteronism in 1,125 hypertensive patients. J Am Coll Cardiol. 2006;48:2293–2300. doi: 10.1016/j.jacc.2006.07.059. doi: 10.1016/j.jacc.2006.07.059. [DOI] [PubMed] [Google Scholar]
- 3.Savard S, Amar L, Plouin PF, Steichen O. Cardiovascular complications associated with primary aldosteronism: a controlled cross-sectional study. Hypertension. 2013;62:331–336. doi: 10.1161/HYPERTENSIONAHA.113.01060. doi: 10.1161/HYPERTENSIONAHA.113.01060. [DOI] [PubMed] [Google Scholar]
- 4.Reincke M, Fischer E, Gerum S, et al. German Conn’s Registry-Else Kröner-Fresenius-Hyperaldosteronism Registry. Observational study mortality in treated primary aldosteronism: the German Conn’s registry. Hypertension. 2012;60:618–624. doi: 10.1161/HYPERTENSIONAHA.112.197111. doi: 10.1161/HYPERTENSIONAHA.112.197111. [DOI] [PubMed] [Google Scholar]
- 5.Funder JW, Carey RM, Mantero F, Murad MH, Reincke M, Shibata H, Stowasser M, Young WF., Jr. The management of primary aldosteronism: case detection, diagnosis, and treatment: an endocrine society clinical practice guideline. J Clin Endocrinol Metab. 2016;101:1889–1916. doi: 10.1210/jc.2015-4061. doi: 10.1210/jc.2015-4061. [DOI] [PubMed] [Google Scholar]
- 6.Choi M, Scholl UI, Yue P, et al. K+ channel mutations in adrenal aldosterone-producing adenomas and hereditary hypertension. Science. 2011;331:768–772. doi: 10.1126/science.1198785. doi: 10.1126/science.1198785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Azizan EA, Lam BY, Newhouse SJ, Zhou J, Kuc RE, Clarke J, Happerfield L, Marker A, Hoffman GJ, Brown MJ. Microarray, qPCR, and KCNJ5 sequencing of aldosterone-producing adenomas reveal differences in genotype and phenotype between zona glomerulosa- and zona fasciculata-like tumors. J Clin Endocrinol Metab. 2012;97:E819–E829. doi: 10.1210/jc.2011-2965. doi: 10.1210/jc.2011-2965. [DOI] [PubMed] [Google Scholar]
- 8.Azizan EA, Poulsen H, Tuluc P, et al. Somatic mutations in ATP1A1 and CACNA1D underlie a common subtype of adrenal hypertension. Nat Genet. 2013;45:1055–1060. doi: 10.1038/ng.2716. doi: 10.1038/ng.2716. [DOI] [PubMed] [Google Scholar]
- 9.Beuschlein F, Boulkroun S, Osswald A, et al. Somatic mutations in ATP1A1 and ATP2B3 lead to aldosterone-producing adenomas and secondary hypertension. Nat Genet. 2013;45:440–4, 444e1. doi: 10.1038/ng.2550. doi: 10.1038/ng.2550. [DOI] [PubMed] [Google Scholar]
- 10.Teo AE, Garg S, Shaikh LH, Zhou J, Karet Frankl FE, Gurnell M, Happerfield L, Marker A, Bienz M, Azizan EA, Brown MJ. Pregnancy, primary aldosteronism, and adrenal CTNNB1 mutations. N Engl J Med. 2015;373:1429–1436. doi: 10.1056/NEJMoa1504869. doi: 10.1056/NEJMoa1504869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nanba K, Tsuiki M, Sawai K, Mukai K, Nishimoto K, Usui T, Tagami T, Okuno H, Yamamoto T, Shimatsu A, Katabami T, Okumura A, Kawa G, Tanabe A, Naruse M. Histopathological diagnosis of primary aldosteronism using CYP11B2 immunohistochemistry. J Clin Endocrinol Metab. 2013;98:1567–1574. doi: 10.1210/jc.2012-3726. doi: 10.1210/jc.2012-3726. [DOI] [PubMed] [Google Scholar]
- 12.Ono Y, Nakamura Y, Maekawa T, Felizola SJ, Morimoto R, Iwakura Y, Kudo M, Seiji K, Takase K, Arai Y, Gomez-Sanchez CE, Ito S, Sasano H, Satoh F. Different expression of 11β-hydroxylase and aldosterone synthase between aldosterone-producing microadenomas and macroadenomas. Hypertension. 2014;64:438–444. doi: 10.1161/HYPERTENSIONAHA.113.02944. doi: 10.1161/HYPERTENSIONAHA.113.02944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen YM, Wu KD, Hu-Tsai MI, Chu JS, Lai MK, Hsieh BS. Differential expression of type 1 angiotensin II receptor mRNA and aldosterone responsiveness to angiotensin in aldosterone-producing adenoma. Mol Cell Endocrinol. 1999;152:47–55. doi: 10.1016/s0303-7207(99)00059-3. [DOI] [PubMed] [Google Scholar]
- 14.Azizan EA, Murthy M, Stowasser M, Gordon R, Kowalski B, Xu S, Brown MJ, O’Shaughnessy KM. Somatic mutations affecting the selectivity filter of KCNJ5 are frequent in 2 large unselected collections of adrenal aldosteronomas. Hypertension. 2012;59:587–591. doi: 10.1161/HYPERTENSIONAHA.111.186239. doi: 10.1161/HYPERTENSIONAHA.111.186239. [DOI] [PubMed] [Google Scholar]
- 15.Omura M, Sasano H, Saito J, Yamaguchi K, Kakuta Y, Nishikawa T. Clinical characteristics of aldosterone-producing microadenoma, macroadenoma, and idiopathic hyperaldosteronism in 93 patients with primary aldosteronism. Hypertens Res. 2006;29:883–889. doi: 10.1291/hypres.29.883. doi: 10.1291/hypres.29.883. [DOI] [PubMed] [Google Scholar]
- 16.Brandenberger R, Schmidt A, Linton J, Wang D, Backus C, Denda S, Müller U, Reichardt LF. Identification and characterization of a novel extracellular matrix protein nephronectin that is associated with integrin alpha8beta1 in the embryonic kidney. J Cell Biol. 2001;154:447–458. doi: 10.1083/jcb.200103069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fujiwara H, Ferreira M, Donati G, Marciano DK, Linton JM, Sato Y, Hartner A, Sekiguchi K, Reichardt LF, Watt FM. The basement membrane of hair follicle stem cells is a muscle cell niche. Cell. 2011;144:577–589. doi: 10.1016/j.cell.2011.01.014. doi: 10.1016/j.cell.2011.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kim AC, Reuter AL, Zubair M, Else T, Serecky K, Bingham NC, Lavery GG, Parker KL, Hammer GD. Targeted disruption of beta-catenin in Sf1-expressing cells impairs development and maintenance of the adrenal cortex. Development. 2008;135:2593–2602. doi: 10.1242/dev.021493. doi: 10.1242/dev.021493. [DOI] [PubMed] [Google Scholar]
- 19.Palacios G. Cell junctions in the adrenal cortex of the postnatal rat. J Anat. 1979;129(pt 4):695–701. [PMC free article] [PubMed] [Google Scholar]
- 20.Hu C, Rusin CG, Tan Z, Guagliardo NA, Barrett PQ. Zona glomerulosa cells of the mouse adrenal cortex are intrinsic electrical oscillators. J Clin Invest. 2012;122:2046–2053. doi: 10.1172/JCI61996. doi: 10.1172/JCI61996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fallo F, Pezzi V, Barzon L, Mulatero P, Veglio F, Sonino N, Mathis JM. Quantitative assessment of CYP11B1 and CYP11B2 expression in aldosterone-producing adenomas. Eur J Endocrinol. 2002;147:795–802. doi: 10.1530/eje.0.1470795. [DOI] [PubMed] [Google Scholar]
- 22.Gomez-Sanchez CE, Qi X, Velarde-Miranda C, Plonczynski MW, Parker CR, Rainey W, Satoh F, Maekawa T, Nakamura Y, Sasano H, Gomez-Sanchez EP. Development of monoclonal antibodies against human CYP11B1 and CYP11B2. Mol Cell Endocrinol. 2014;383:111–117. doi: 10.1016/j.mce.2013.11.022. doi: 10.1016/j.mce.2013.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.van Engeland M, Ramaekers FC, Schutte B, Reutelingsperger CP. A novel assay to measure loss of plasma membrane asymmetry during apoptosis of adherent cells in culture. Cytometry. 1996;24:131–139. doi: 10.1002/(SICI)1097-0320(19960601)24:2<131::AID-CYTO5>3.0.CO;2-M. doi: 10.1002/(SICI)1097-0320(19960601)24:2<131::AID-CYTO5>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 24.Atienza JM, Zhu J, Wang X, Xu X, Abassi Y. Dynamic monitoring of cell adhesion and spreading on microelectronic sensor arrays. J Biomol Screen. 2005;10:795–805. doi: 10.1177/1087057105279635. doi: 10.1177/1087057105279635. [DOI] [PubMed] [Google Scholar]
- 25.Shaikh LH, Zhou J, Teo AE, Garg S, Neogi SG, Figg N, Yeo GS, Yu H, Maguire JJ, Zhao W, Bennett MR, Azizan EA, Davenport AP, McKenzie G, Brown MJ. LGR5 activates noncanonical Wnt signaling and inhibits aldosterone production in the human adrenal. J Clin Endocrinol Metab. 2015;100:E836–E844. doi: 10.1210/jc.2015-1734. doi: 10.1210/jc.2015-1734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhou J, Shaikh LH, Neogi SG, McFarlane I, Zhao W, Figg N, Brighton CA, Maniero C, Teo AE, Azizan EA, Brown MJ. DACH1, a zona glomerulosa selective gene in the human adrenal, activates transforming growth factor-β signaling and suppresses aldosterone secretion. Hypertension. 2015;65:1103–1110. doi: 10.1161/HYP.0000000000000025. doi: 10.1161/HYP.0000000000000025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sato Y, Uemura T, Morimitsu K, Sato-Nishiuchi R, Manabe R, Takagi J, Yamada M, Sekiguchi K. Molecular basis of the recognition of nephronectin by integrin alpha8beta1. J Biol Chem. 2009;284:14524–14536. doi: 10.1074/jbc.M900200200. doi: 10.1074/jbc.M900200200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kolligs FT, Hu G, Dang CV, Fearon ER. Neoplastic transformation of RK3E by mutant beta-catenin requires deregulation of Tcf/Lef transcription but not activation of c-myc expression. Mol Cell Biol. 1999;19:5696–5706. doi: 10.1128/mcb.19.8.5696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Korinek V, Barker N, Morin PJ, van Wichen D, de Weger R, Kinzler KW, Vogelstein B, Clevers H. Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC-/- colon carcinoma. Science. 1997;275:1784–1787. doi: 10.1126/science.275.5307.1784. [DOI] [PubMed] [Google Scholar]
- 30.Bieritz B, Spessotto P, Colombatti A, Jahn A, Prols F, Hartner A. Role of alpha8 integrin in mesangial cell adhesion, migration, and proliferation. Kidney Int. 2003;64:119–127. doi: 10.1046/j.1523-1755.2003.00057.x. doi: 10.1046/j.1523-1755.2003.00057.x. [DOI] [PubMed] [Google Scholar]
- 31.Patra C, Ricciardi F, Engel FB. The functional properties of nephronectin: an adhesion molecule for cardiac tissue engineering. Biomaterials. 2012;33:4327–4335. doi: 10.1016/j.biomaterials.2012.03.021. doi: 10.1016/j.biomaterials.2012.03.021. [DOI] [PubMed] [Google Scholar]
- 32.Kelly PN, Strasser A. The role of Bcl-2 and its pro-survival relatives in tumourigenesis and cancer therapy. Cell Death Differ. 2011;18:1414–1424. doi: 10.1038/cdd.2011.17. doi: 10.1038/cdd.2011.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Åkerström T, Willenberg HS, Cupisti K, et al. Novel somatic mutations and distinct molecular signature in aldosterone-producing adenomas. Endocr Relat Cancer. 2015;22:735–744. doi: 10.1530/ERC-15-0321. doi: 10.1530/ERC-15-0321. [DOI] [PubMed] [Google Scholar]
- 34.Warren HR, Evangelou E, Cabrera CP, et al. International Consortium of Blood Pressure (ICBP) 1000G Analyses; BIOS Consortium; Lifelines Cohort Study; Understanding Society Scientific group; CHD Exome+ Consortium; ExomeBP Consortium; T2D-GENES Consortium; GoT2DGenes Consortium; Cohorts for Heart and Ageing Research in Genome Epidemiology (CHARGE) BP Exome Consortium; International Genomics of Blood Pressure (iGEN-BP) Consortium; UK Biobank CardioMetabolic Consortium BP working group. Genome-wide association analysis identifies novel blood pressure loci and offers biological insights into cardiovascular risk. Nat Genet. 2017;49:403–415. doi: 10.1038/ng.3768. [Google Scholar]
- 35.Barrett PQ, Guagliardo NA, Klein PM, Hu C, Breault DT, Beenhakker MP. Role of voltage-gated calcium channels in the regulation of aldosterone production from zona glomerulosa cells of the adrenal cortex. J Physiol. 2016;594:5851–5860. doi: 10.1113/JP271896. doi: 10.1113/JP271896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Okamura H, Doi M, Goto K, Kojima R. Clock genes and salt-sensitive hypertension: a new type of aldosterone-synthesizing enzyme controlled by the circadian clock and angiotensin II. Hypertens Res. 2016;39:681–687. doi: 10.1038/hr.2016.91. doi: 10.1038/hr.2016.91. [DOI] [PubMed] [Google Scholar]
- 37.Inagaki FF, Tanaka M, Inagaki NF, Yagai T, Sato Y, Sekiguchi K, Oyaizu N, Kokudo N, Miyajima A. Nephronectin is upregulated in acute and chronic hepatitis and aggravates liver injury by recruiting CD4 positive cells. Biochem Biophys Res Commun. 2013;430:751–756. doi: 10.1016/j.bbrc.2012.11.076. doi: 10.1016/j.bbrc.2012.11.076. [DOI] [PubMed] [Google Scholar]
- 38.Rao TP, Kühl M. An updated overview on Wnt signaling pathways: a prelude for more. Circ Res. 2010;106:1798–1806. doi: 10.1161/CIRCRESAHA.110.219840. doi: 10.1161/CIRCRESAHA.110.219840. [DOI] [PubMed] [Google Scholar]
- 39.Dzamba BJ, Jakab KR, Marsden M, Schwartz MA, DeSimone DW. Cadherin adhesion, tissue tension, and noncanonical Wnt signaling regulate fibronectin matrix organization. Dev Cell. 2009;16:421–432. doi: 10.1016/j.devcel.2009.01.008. doi: 10.1016/j.devcel.2009.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Astudillo P, Larraín J. Wnt signaling and cell-matrix adhesion. Curr Mol Med. 2014;14:209–220. doi: 10.2174/1566524014666140128105352. [DOI] [PubMed] [Google Scholar]
- 41.Abraham S, Kogata N, Fässler R, Adams RH. Integrin beta1 subunit controls mural cell adhesion, spreading, and blood vessel wall stability. Circ Res. 2008;102:562–570. doi: 10.1161/CIRCRESAHA.107.167908. doi: 10.1161/CIRCRESAHA.107.167908. [DOI] [PubMed] [Google Scholar]
- 42.Frisch SM, Francis H. Disruption of epithelial cell-matrix interactions induces apoptosis. J Cell Biol. 1994;124:619–626. doi: 10.1083/jcb.124.4.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gabbiani G. The myofibroblast in wound healing and fibrocontractive diseases. J Pathol. 2003;200:500–503. doi: 10.1002/path.1427. doi: 10.1002/path.1427. [DOI] [PubMed] [Google Scholar]
- 44.Dekkers T, Prejbisz A, Kool LJ, et al. SPARTACUS Investigators. Adrenal vein sampling versus CT scan to determine treatment in primary aldosteronism: an outcome-based randomised diagnostic trial. Lancet Diabetes Endocrinol. 2016;4:739–746. doi: 10.1016/S2213-8587(16)30100-0. doi: 10.1016/S2213-8587(16)30100-0. [DOI] [PubMed] [Google Scholar]
- 45.Zhu L, Zhang Y, Zhang H, Zhou W, Shen Z, Zheng F, Tang X, Tao B, Zhang J, Lu X, Xu J, Chu S, Zhu D, Gao P, Wang JG. Comparison between adrenal venous sampling and computed tomography in the diagnosis of primary aldosteronism and in the guidance of adrenalectomy. Medicine (Baltimore) 2016;95:e4986. doi: 10.1097/MD.0000000000004986. doi: 10.1097/MD.0000000000004986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bird IM, Hanley NA, Word RA, Mathis JM, McCarthy JL, Mason JI, Rainey WE. Human NCI-H295 adrenocortical carcinoma cells: a model for angiotensin-II-responsive aldosterone secretion. Endocrinology. 1993;133:1555–1561. doi: 10.1210/endo.133.4.8404594. doi: 10.1210/endo.133.4.8404594. [DOI] [PubMed] [Google Scholar]
- 47.Tissier F, Cavard C, Groussin L, Perlemoine K, Fumey G, Hagneré AM, René-Corail F, Jullian E, Gicquel C, Bertagna X, Vacher-Lavenu MC, Perret C, Bertherat J. Mutations of beta-catenin in adrenocortical tumors: activation of the Wnt signaling pathway is a frequent event in both benign and malignant adrenocortical tumors. Cancer Res. 2005;65:7622–7627. doi: 10.1158/0008-5472.CAN-05-0593. doi: 10.1158/0008-5472.CAN-05-0593. [DOI] [PubMed] [Google Scholar]
- 48.Linton JM, Martin GR, Reichardt LF. The ECM protein nephronectin promotes kidney development via integrin alpha8beta1-mediated stimulation of Gdnf expression. Development. 2007;134:2501–2509. doi: 10.1242/dev.005033. doi: 10.1242/dev.005033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Freedman BD, Kempna PB, Carlone DL, Shah MS, Guagliardo NA, Barrett PQ, Gomez-Sanchez CE, Majzoub JA, Breault DT. Adrenocortical zonation results from lineage conversion of differentiated zona glomerulosa cells. Dev Cell. 2013;26:666–673. doi: 10.1016/j.devcel.2013.07.016. doi: 10.1016/j.devcel.2013.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]