

Abstract
Robust evidence exists that certain extreme weather and climate events, especially daily temperature and precipitation extremes, have changed in regard to intensity and frequency over recent decades. These changes have been linked to human-induced climate change, while the degree to which climate change impacts an individual extreme climate event (ECE) is more difficult to quantify. Rapid progress in event attribution has recently been made through improved understanding of observed and simulated climate variability, methods for event attribution and advances in numerical modelling. Attribution for extreme temperature events is stronger compared with other event types, notably those related to the hydrological cycle. Recent advances in the understanding of ECEs, both in observations and their representation in state-of-the-art climate models, open new opportunities for assessing their effect on human and natural systems. Improved spatial resolution in global climate models and advances in statistical and dynamical downscaling now provide climatic information at appropriate spatial and temporal scales. Together with the continued development of Earth System Models that simulate biogeochemical cycles and interactions with the biosphere at increasing complexity, these make it possible to develop a mechanistic understanding of how ECEs affect biological processes, ecosystem functioning and adaptation capabilities. Limitations in the observational network, both for physical climate system parameters and even more so for long-term ecological monitoring, have hampered progress in understanding bio-physical interactions across a range of scales. New opportunities for assessing how ECEs modulate ecosystem structure and functioning arise from better scientific understanding of ECEs coupled with technological advances in observing systems and instrumentation.
This article is part of the themed issue ‘Behavioural, ecological and evolutionary responses to extreme climatic events’.
Keywords: extreme events, climate variability, climate change, detection and attribution, event attribution, ecological impacts
1. Introduction and motivation
A recent publication by the National Academy of Sciences [1] is the latest addition to a series of focused summary reports [2–4] that highlight mounting evidence that extreme weather and climate events have been changing in regard to intensity, frequency and duration in the last few decades. Daily temperature and precipitation extremes in particular have been observed to increase in frequency and intensity, which has been linked to human-induced climate change [5–7].
By contrast, the degree to which climate change impacts an individual extreme weather or climatic event is more difficult to determine and quantify. This applies especially when considering that a variety of natural and anthropogenic factors, such as internal modes of climate variability, various man-made emissions, land-use change, and so on, need to be taken into account when attributing individual weather or climate events to causal factors. Solow [8] cautioned that the concept of attributable risk for single events in a climate change context is inherently difficult given the rarity of extreme climatic events (ECEs) and the limited reliable climatic record. Extreme events by definition are rare occurrences and in most places few examples of past events are seen in the observational record [1].
However, recent rapid progress in event attribution has been made through improved understanding of observed and simulated climate variability, increasing observational capabilities, methods for event attribution and advances in numerical modelling. The American Meteorological Society's annual report of extreme events that occurred in the previous year was first published in 2012 with six extreme event studies for the year 2011 [9]. The number of studies rose sharply to 32 extreme events last year [10], covering all continents and much broader types of events and impacts during 2014 [7]. The National Academy of Sciences report thus concluded that ‘in many cases, it is now often possible to make and defend quantitative statements about the extent to which human-induced climate change (or another causal factor, such as a specific mode of natural variability) has influenced either the magnitude or the probability of occurrence of specific types of events or event classes [1, p. 16]’.
There has also been substantial progress made recently in assessing ECEs according to the latest Intergovernmental Panel on Climate Change (IPCC) assessment report 5 (AR5), compared with the previous AR4 report [3,11]. Recent advances in the understanding of ECEs, both in observations and their representation in state-of-the-art climate model simulations, open new opportunities for assessing the effect of ECEs on human and natural systems at relevant scales. In particular, improved spatial resolution in global climate models combined with advances in statistical and dynamical downscaling (e.g. regional model configurations) now provide climatic information at the appropriate spatial and temporal scales: with these, it is possible to develop a mechanistic understanding of how ECEs in the physical climate system affect biological processes, ecosystem functioning and adaptation capabilities.
It is evident that ECEs, especially large-scale events such as subcontinental-scale drought or heatwaves, can have profound effects on ecosystems [12]. This was shown for example for arid and semi-arid ecosystems in response to hydroclimatic disturbances associated with El Niño-Southern Oscillation (ENSO) events in Australia and the Americas ([13] and references therein): more specifically, ECEs can trigger ecosystem-level disturbances through changing species composition and diversity (i.e. organization) and functional attributes [14]. For marine ecosystems, ECEs are also considered a key driver of biodiversity patterns (e.g. [15]). Addressing how such episodic events affect species distribution is regarded as crucial to advance predictive models of species distribution and ecosystem structure in the future, beyond their current basis on gradual warming trends [16]. However, when assessing 238 widespread species in England, Palmer et al. [17] found extreme biological responses linked individualistically to climate, but long-term trends of widespread species were not (yet) simultaneously dominated by ECEs.
The representation of large-scale to regional-scale ECEs is within the capabilities of current-generation climate models, while ECEs on local and subgrid scale (on the order of metres to dozens of kilometres) still pose challenges. The specifics of the ecosystem response, including initial resistance, evolution of the response and the system's resilience to return to its original condition, depend on the characteristics of the biological system and the level of the disturbance [14]. When considering ECEs as disturbances, these include frequency, intensity, duration, seasonality and preconditioning. Recent improvements have been made in the observational networks to evaluate ECE characteristics with regard to data homogeneity [11], as well as spatial and temporal coverage and resolution through advanced technologies. Remotely sensed datasets of the climate system with short return intervals at identical locations and near-global coverage [18] provide major advances in understanding changes by quantifying processes and spatio-temporal states of the atmosphere, land and oceans [19]. This applies to biological systems for example through global satellite monitoring of climate-induced vegetation disturbances [20] or ocean colour remote sensing for phytoplankton blooms since the 1970s [21]. As such, satellite-based datasets are becoming long enough to be used in detection–attribution studies for ECEs [22].
This review provides an overview of the current understanding of changes in ECEs and how they are quantified. For a detailed assessment of ECEs, the reader is referred to recent papers providing an in-depth review of various aspects of changes in ECEs, their detection and attribution (e.g. [11,22–24]). Here, particular focus is on ECEs with ecological relevance and covering different realms of the climate system and across a range of spatial and temporal scales. It is by no means meant as an exhaustive accounting of changes in ECEs or their ecological impacts, but focuses mostly on ecosystems and community ecology. The remainder of the review is structured as follows: §2 defines ECEs and highlights how their definition might affect the assessment of changes, while §3 reviews detection and attribution of events. Section 4 details observed and simulated changes in ECEs with ecological relevance, along with examples from the terrestrial/atmosphere and ocean realms. Section 5 discusses current challenges and opportunities in understanding how ECEs affect biological systems, followed by conclusions provided in §6.
2. Extreme climatic events
Different definitions for ECEs exist, such as those detailed in table 1 in van de Pol et al. [25] in this issue. In this review, we follow the climatological definition used by the IPCC Special Report on Extreme events [2] for a climate extreme (extreme weather or climate event; figure 1): i.e. the occurrence of a weather or climate variable above (below) a threshold value near the upper (lower) end of the range of observed records of the variable. Definitions of thresholds can vary, but are typically 10%, 5% or 1% relative to a reference period, though absolute thresholds are also sometimes considered (e.g. for critical threshold temperatures for physiological responses; [27]). However, in absolute terms, what is considered an extreme event will vary in different locations: ECE characteristics are intimately tied to a location's mean climatic condition and its variability. For example, a record daily maximum temperature in the Sahara is higher than for Alaska; for the latter, the absolute value for this record temperature will also exhibit much greater seasonal dependence given the temperature range between winter and summer at higher latitudes.
Figure 1.
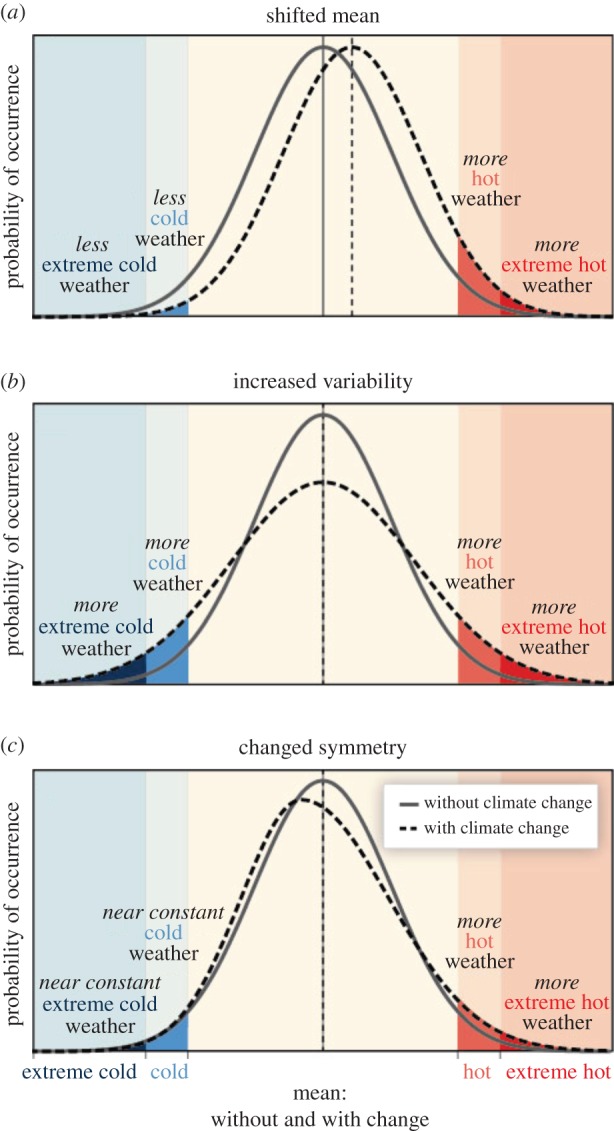
Schematic highlighting the effect of changes in the temperature distribution on ECE occurrence between present and future climate conditions: (a) effects of a simple shift of the entire distribution toward a warmer climate; (b) effects of an increase in temperature variability with no shift in the mean; (c) effects of an altered shape of the distribution, in this example a change in asymmetry toward the hotter part of the distribution. Reproduced from IPCC [26].
Several characteristics of extreme events are of interest. These include the magnitude of ECEs, the probability or return frequency, the duration of the ECE, the spatial extent, timing, onset date or seasonality, and preconditioning [27]. Preconditioning in this context refers to antecedent conditions that facilitate or enable a certain extreme event to occur or modify its characteristics. For example, antecedent soil moisture deficits that accumulate over the course of several months in winter/spring have been shown to exacerbate summer heatwave and drought conditions (e.g. [28]), as dry soils may amplify extreme maximum temperatures through feedback with evapotranspiration [29,30]. Changes in these ECE characteristics have been investigated in climate change studies, though the majority of research has focused on ECE changes with regard to magnitude and probability/return frequency.
To define ECEs, thresholds, percentiles or return values are defined with respect to a reference period, which is often historical, i.e. 1961–1990. The choice of reference period can affect the assessed changes and whether it is considered to be static or transient [27,31]: if ECEs are defined based on a percentile of the probability distribution, shifts in the mean (without any change in the shape of the distribution) will not lead to a relative change in the frequency of the extremes. Sippel et al. [31] cautioned, though, that standardization to a reference period can introduce an inhomogeneity when calculating temperature variability and extremes, with a risk of arbitrarily inflating extremes, and suggested an analytical correction. A series of studies demonstrate that observed changes in the frequency of extremes are consistent with overall shifts in the distribution (e.g. [32–34]). While counts of threshold exceedance, such as frequency and duration, closely follow mean changes, variations in intensity or severity are considerably more sensitive to changes in the shape of the probability distribution (figure 1; e.g. [33,35]). There is also ongoing debate about the role of changes in the variance and higher order moments, such as skewness, in addition to the mean [33]. Furthermore, the statistics of ECEs are particularly sensitive to data availability, quality and consistency.
Some limitations were previously noted with regard to defining ECEs as probability-based or threshold-based [27]: events from the extreme tails of the probability distribution do not necessarily have to be extreme in terms of impact. Impact-related thresholds are variable in space and time, such that definitions for ECEs need to be modified for different locations and time periods (e.g. seasons). To account for this, ECEs can be defined quantitatively in two ways: (i) related to a specific threshold (possibly impact-related); or (ii) related to their probability of occurrence [27]. These definitions are not necessarily diametric; impacts on society or ecosystem responses are often extreme, irrespective of whether a probability or threshold definition has been used.
3. Detection and attribution of extreme climate events
Reliable detection and attribution of changes in specific climatic events and their impacts are key for understanding the scientific basis of climate change and for successful decision-making to enable adaptation and mitigation [22,36]. Similarly, to assess how ECEs impact structure and functioning of populations, individual species or entire ecosystems, detection and attribution approaches are crucial.
Detection refers to the process of demonstrating that an ECE characteristic (or climate variable more broadly) has changed with respect to some previous period in a defined statistical sense; however, no reason for that change is provided [22,36]. In observations, such a change is identified if its likelihood of occurrence by chance due to internal variability alone is considered to be small [5]. By contrast, attribution is the process of assessing the relative contributions from multiple causal factors to an ECE change or event; it also assigns a statistical confidence to this conclusion. Attribution is thus more complex than detection, as it combines statistical analysis with physical understanding [5,37]. One can attribute an observed ECE change to a specific causal factor through demonstrating that the change is consistent with a process-based model that includes this factor and is inconsistent with an alternate model that is otherwise identical, but excludes the factor. This assessment also needs to take into account internal chaotic variability and uncertainties in observations and responses to external causal factors [5].
Approaches for detection and attribution for a particular event can be classified into two categories [3]: (i) studies that use the observational record to assess whether a change in the probability or magnitude of an ECE has occurred; (ii) studies based on climate model simulations (coupled or using a subset of components only) to compare characteristics of the event in simulations with and without anthropogenic climate change; a combination of both approaches is also often used [3]. Stott [7] further divides the latter into two types of methods as follows: (i) exceeding a certain climate index in extended coupled climate model simulations with and without climate change; (ii) using a large ensemble of atmosphere-only simulations that use observed boundary conditions, such as sea surface temperatures (SST), to evaluate if the climate index of interest, with or without the factor included, occurs at a changed frequency [7,24]. This approach is not limited to atmosphere-only configurations, where an atmospheric model is forced by observed boundary conditions (e.g. SST), but could also be applied to ocean models forced with atmospheric boundary conditions (e.g. winds, precipitation, heat fluxes).
Overall, the evidence for detecting a human influence on temperature extremes has strengthened since the IPCC Special Report on Extreme events [2]. Global-scale daily temperature extremes have increased in frequency and intensity since the 1950s, very likely due to anthropogenic influences, and heat-wave probabilities have doubled in some locations [3]. Stott [7] also highlights that specific event attribution for extreme temperature events, such as record daily temperatures or heatwaves, is stronger compared with other types of events, notably those related to changes in the hydrological cycle.
Despite substantial recent improvements in models, reanalyses1 and satellite records, detection and attribution of human influence on the water cycle and in particular regional precipitation remains challenging [6,11,24,39]. To disentangle the complex regional-scale changes in precipitation, a highly noisy variable, several factors are likely to affect the anthropogenic response: (i) how external forcing affects internal modes of climate variability, which can alter the frequency or amplitude of the mode or in turn modify its precipitation teleconnections; (ii) responses to different external drivers (e.g. aerosols, ozone) vary for precipitation; and (iii) the spatial expression of the precipitation response to external forcing contains signals that are due to thermodynamic as well as dynamic changes, which arise due to altered atmospheric energetics, moisture content and large-scale circulation [39].
4. Changes in extreme climate events with ecological relevance
Robust changes in many ECEs have been observed in the second half of the twentieth century. Shifts in the frequency of ECEs can arise due to different changes, as highlighted exemplarily for a temperature distribution in figure 1: ECEs can occur more frequently due to a shift in the mean, be associated with shifts in the variability of the distribution, as well as changes in its symmetry or skewness (figure 1; [26]).
(a). Terrestrial/atmosphere
(i). Temperature extremes
For temperature, changes in the distribution are especially pronounced for increases in maximum and minimum temperatures [40]. There is robust evidence across multiple datasets that minimum temperatures across the globe are rising faster, which could lead to a decline in the daily temperature range [3]. Globally, a decrease in the number of cold days and nights is considered very likely, while the number of warm days and nights increased. Regional variations exist, with Europe and Asia exhibiting especially pronounced decreases in cold nights [40–42]. For North America, it is projected that the total area experiencing freezing days will contract by approximately 6% by 2070, with the number of freezing days declining by 10–90 days depending on the region [43]. It has been shown for the USA [44,45], Australia [46] and Europe [47] that the ratio of daily record high temperatures to daily record low temperatures has been increasing, with the average for the first decade of the twenty-first century for these regions being about two to one (i.e. two daily record highs are set for every one daily record low). The shift in the chances for more record highs than record lows relates to the increase in average temperatures over this time period in these locations. This ratio is projected to increase in the future as the climate continues to warm [30,44,47].
Confidence in trends in temperature extremes is high for North and Central America (e.g. [48]), Europe (e.g. [49]), Asia (e.g. [41]), Southeast Asia and Oceania (e.g. [50]). Trends in the climate models with regard to the frequency of extreme warm and cold days and nights since the 1950s are consistent with observations and are projected to continue to change into the twenty-first century [3]. A decrease of frost days, an increase in growing season length, an increase in the number of warm nights, and an increase in heatwave intensity over the USA have been attributed mostly to increases in human-produced greenhouse gases [51]. Heat-related extremes and some precipitation extremes have also been attributed to human influences on climate [52]. Changes in regional temperature extremes have been associated with changing global and regional atmospheric circulation and SST patterns [33,53]. High temperature extremes have been shown to substantially affect individual species, as shown for example for negative effects on clutch size for a particular bird species [54] and on overwintering success of butterfly species in the UK [55], or ecosystems through overall contractions or shifts in the distribution range of species habitat, as critical temperature thresholds are exceeded [56,57].
(ii). Precipitation extremes
According to theoretical thermodynamic understanding, the water-holding capacity of the atmosphere scales with temperature and an intensification of extreme precipitation is expected in a warming world (e.g. [33,58–63]). According to the well-known Clausius–Clapeyron relationship, the saturation-specific humidity increases by approximately 7% per °C of warming, with higher levels of moisture available and intensifying rainfall. This was also found in observed annual maximum daily rainfall rate increases of 5.9–7.7% for globally averaged surface temperatures [64]. However, in their review of subdaily extreme rainfall changes, Westra et al. [23] described observed and simulated rates of extreme precipitation increases double of that suggested by Clausius–Clapeyron at temperatures below 20°C. Short-duration (less than 1 day) storms were most likely to occur more often, which might increase the frequency and magnitude of flash floods [23]. Since the 1950s, heavy precipitation events have probably increased in frequency over land globally, especially over North America and Europe, while confidence in heavy precipitation over other land areas was only medium [3]. A shift to more extreme precipitation patterns with more heavy rainfall and longer dry intervals [65] has been shown to decrease the rain use efficiency across biomes. This is most pronounced for arid grasslands and Mediterranean forests (16–20%), due to higher water stress conditions and reduced vegetation production [66]. Feng et al. [67] also highlighted changes in the rainfall seasonality in the tropics over the twentieth century: they observed increasing interannual variability of seasonality over large parts of the dry tropics (arid and semi-arid regions), with shifts in seasonal magnitude, timing and duration, all factors that are of importance for local ecological processes.
Changes in regional precipitation can arise due to both a thermodynamic as well as a dynamic contribution (i.e. including changes in circulation, modes of variability and teleconnections; [39]). Westra et al. [23] highlighted the importance of research efforts focusing on improving our understanding of local-scale thermodynamic effects and large-scale atmospheric circulation in modulating subdaily extreme rainfall intensity. A series of studies have recently found that regional SST warming played a role in intensifying extreme precipitation events [68–71]. For example, Australia experienced extreme rainfall conditions during the 2010/2011 La Niña event that led to extensive flooding in the northeast of the country ([71] and references therein). As demonstrated in atmospheric circulation model experiments based on 2010/2011 ocean conditions with and without long-term warming included, both dynamic and thermodynamic factors led to the intensification of the rain-producing atmospheric circulation conditions [71]. The resultant widespread wet conditions in the interior of the Australian continent, which included a rare filling of Lake Eyre [72], led to unusually high growth of the semi-arid vegetation, accounting for record global terrestrial carbon uptake in 2010/2011 [73].
(iii). Heatwaves and droughts
Heatwaves over large areas of Europe, Australia and Asia have probably become more frequent, with droughts more intense and/or longer in many regions since the 1970s [3]. Severe heatwaves, such as the one in Europe in 2003 [28,74–76], Australia in 2009, Russia in 2010 [77–79] and the USA in 2010/2011 [80], are often associated with persistent blocking high pressure systems [81]. These circulation regimes are projected to become more frequent, intense and persistent in the twenty-first century, leading to an intensification of heatwaves in Europe and North America [81]. Summers like 2003 over Europe are likely to occur twice a decade in the late twenty-first century [82]. Assessing seasonal temperature and circulation regime changes over Europe for 1960–2000, Cassou & Cattiaux [83] found an earlier onset of summer: they related this to an earlier disappearance of winter snow in Eastern Europe that hastened the typical summertime formation of a blocking high pressure system over Europe. This was associated with more clear-sky days with increased incoming short-wave radiation and anomalous easterly advection of warm air from the continental interior earlier in the year [83].
During the 2003 heatwave, unprecedented reduction in Europe's gross primary productivity occurred that reversed the effect of 4 years of net ecosystem carbon sequestration, with a 30% reduced primary productivity and lower ecosystem respiration [84]. Several recent reviews [18,85] investigated how ECEs affect terrestrial ecosystems and, in particular, the carbon cycle. ECEs, such as droughts and heatwaves, severely affect forests and grasslands through changes in plant physiology, phenology and carbon allocation. They also lead to increased tree mortality, shifts in vegetation composition, degradation and desertification, along with erosion [85]. Advances have been made recently in understanding how droughts affect tropical forests on molecular, cellular, individual, species, community and landscape level [86]. In a review on ecological impacts of droughts on the Amazon, Asner & Alencar [87] found the hydrological function of floodplains significantly affected by droughts and fires, and burn scars were more frequent during drought years; they also highlighted the importance of integrating multiple lines of evidence from remote sensing of hydrological, disturbance-fire and physiological impacts with field measurements, to reduce uncertainty of basin-level responses to drought [87]. Droughts are most likely to have the largest and most long-lasting impacts globally due to large indirect and lagged impacts and long recovery especially for forest ecosystems [18]. Examples include the rapid subcontinental die-off of woody plants during the 2000–2004 drought in the Southwest United States, when more than 90% of the dominant pine species died as a consequence of 15 months of soil water content deficits [88,89]; the intense 2005 drought affecting the Amazon forest ecosystem to a degree that reversed its role as a long-term carbon sink [90]; and a global overview of drought and heat-induced tree mortality studies from the Americas, Australia, Europe and Asia, indicating that no forest type or climate zone is invulnerable to ECEs even in environments not normally considered water-limited [91].
Focusing on the 2003 European summer, Garcia-Herrera et al. [76] concluded that a northward displacement of the North Atlantic subtropical high and anomalous Mediterranean SST contributed to the heatwave. Investigating the role of soil moisture–atmosphere interactions, Fischer et al. [28] found an early spring soil moisture deficit to be instrumental in accounting for the severity of the summer heatwave in Europe. They linked the decrease in springtime soil moisture to a precipitation deficit along with strong positive radiative anomalies and early vegetation green-up [28]. Similarly, Wolf et al. [92] found increased vegetation growth and carbon uptake during the record-breaking warmth and early arrival of spring 2012. Increased carbon uptake in spring could have enhanced depletion of soil water through higher evapotranspiration and exacerbated summer drought conditions, highlighting the importance of land-atmosphere feedback during ECEs [93]. For the Great Plains in the Midwest US, enhanced local land–atmosphere feedback are likely associated with an amplification of future heatwaves due to stronger subseasonal summertime temperature variability [94].
Droughts can also affect freshwater ecosystems, such as streams, rivers, lakes and wetlands, stressing and depleting both fauna and flora as shown for the so-called Big Dry or Millennium Drought for Southeast Australia ([95] and references therein), floodplains in the Amazon [87] and in a review based on studies from the US, Europe and Australia, while impacts particularly in Asia, Africa, and South America have not been documented in the published literature [96]. Droughts can result in poor water quality, habitat loss and changed biotic interactions, which will impact aquatic biota and ecosystem functioning in both flowing and standing water systems, where the effects of drought on population and community structure are better understood than impacts on ecosystem processes [95].
(iv). Wildfires
The frequency of wildfires is related to temperature, moisture and fuel loads, which in turn are affected by species composition and age structure [14]. Littell et al. [97] found antecedent climatic conditions, such as winter precipitation for shrub and grassland ecosystems and summer droughts in forests, to be an important factor accounting for trends in the areal extent burned in the western US. Extended periods of drought, especially if they are followed by extreme heat and low humidity, provide ideal conditions for wildfires, often incited by lightning associated with thunderstorms at the end of the drought [14]. Assessing changes in fire severity in the western US, Miller et al. [98] found substantial increases in mean and maximum fire size and the annual area burned since the 1980s, indicating that forest fuels were no longer a limiting factor for fire occurrence. Sudden changes in wildfire activity in forests in the northern Rocky Mountains in the mid-1980s, with more frequent large-scale fires, longer-lasting wildfires and a longer wildfire season, were associated with earlier spring snowmelt and increased spring and summer temperatures [99]. In the light of warming regional temperatures and changing precipitation, along with current trends in increasing wildfire severity, it is important to address the implications of ongoing fire suppression. This is especially so as severe wildfires can have extensive ecological impacts, including on forest fragmentation, erosion rates, carbon sequestration, wildlife habitat availability and post-fire seedling recruitment [98]. Several studies emphasized the importance of the interaction of the physical climate system and biological processes across temporal and spatial scales to explain climate–wildfire interactions: Marlon et al. [100] ascertained that improved understanding of the causes and consequences of forest wildfires in the western US is crucially dependent on integrated information of climate change and human activity across a range of temporal scales. Parisien & Moritz [101] advocated further work for improved understanding of direct causal factors that control wildfires across a range of spatial scales. Alencar et al. [102] linked increased fire incidence in dense forests in the Amazon basin to severe, ENSO-related droughts, when the end of the dry season was delayed by a month, resulting in larger burn scars and overall extent of the area burned; by contrast, open and transitional forests with higher deforestation rates burned more frequently, suggesting that climate-mediated forest flammability was exacerbated by landscape fragmentation [102]. Investigating preconditioning of devastating bushfires in Southeast Australia in February of 1983 and 2009, Cai et al. [103] associated these with characteristic Indian Ocean conditions with a positive Indian Ocean Dipole, rather than El Niño events, through impacts on soil moisture, as shown for prolonged Southeast Australian droughts [104].
(b). Ocean
(i). Marine heatwaves and cold spells
In analogy to heatwaves in the atmosphere, the marine environment also experiences sustained extreme temperature events, so-called ‘marine heatwaves’. It is defined as a prolonged discrete event with anomalously warm water and characteristics include its spatial extent, intensity, duration and rate of evolution [105]. Marine heatwaves can have extensive ecological implications, including shifts in species range [106], local extinctions [15] and economic impacts when affecting aquaculture or important fishery species [107]. Some of the recent observed marine heatwaves with extensive ecological implications occurred in the northern Mediterranean in 2003 [108], the 2011 ‘Ningaloo Niño’ in Western Australia [16], and in the northwest Atlantic in 2012 [107]. Investigating the frequency of marine heatwaves in the North Pacific and Atlantic since the 1950s, Scannell et al. [109] found the probability of marine heatwaves to be a trade-off between size, duration and intensity, which are modulated by a region's specific variability, as well as by modes of climate variability and anthropogenic warming. Marine heatwaves have been observed more frequently in the last decades and are projected to become more frequent in a warming climate [105], though decadal variability probably plays a role as well [110]. Wernberg et al. [15,16] linked regime shifts of Western Australian temperate reef ecosystems to continuing ocean warming and extreme marine heatwaves. These resulted in a significant range contraction of kelp forests, which were being replaced by communities typical of subtropical and tropical waters [15,16]. Di Lorenzo & Ohman [111] showed that cumulative responses to atmospheric forcing can help explain large-amplitude state transitions in marine ecosystems, allowing better interpretation of both abrupt responses and gradual changes (e.g. to long-term warming) in biological systems.
An example of a record-setting ocean heatwave was the one that occurred in the North Pacific 2013–2015 [112]. It was one of the largest marine heatwaves ever recorded, with SST anomalies exceeding three standard deviations in consecutive years [112]. It is possible that this record-breaking ocean heatwave could have been the most ecologically significant in recorded history [112]. Impacts included sea lion, whale and sea bird mortality events [113–115], very low ocean primary productivity [116], and the largest algal bloom on record that negatively impacted shellfish along the western coast of North America [117]. This event was characterized by an overall warming trend in the North Pacific Ocean superimposed with anomalously warm interannual SSTs in the Gulf of Alaska and along the west coast of North America that persisted and grew due to alternating mid-latitude-tropical and tropical-mid-latitude interactions, with the possibility that such events could increase in the future in association with an increase in winter-time variance of climate over the North Pacific [112].
Another example of an ocean heatwave impact was associated with the 2015–2016 El Niño that produced extremely high tropical SSTs in regions where coral reefs experienced the third mass bleaching event in recorded history [118]. All these mass bleaching events have occurred in conjunction with El Niño events since 1997 (first mass bleaching was 1998, the second was 2010; [118]), and occurred when steadily rising SSTs from human-caused warming had warm El Niño SST anomalies superimposed, thus crossing the tolerance threshold and causing reefs to bleach. In previous mass bleaching events, a certain percentage of bleached reefs died, thus raising the prospect of large-scale coral reef mortality arising from this ECE related to the 2015–2016 El Niño event. For example, there was a reported bleaching of over 90% of the Great Barrier Reef in Australia by early 2016 [119]. The prospects for even greater mass bleaching events increase during future El Niño events as the climate continues to warm, with dire consequences for the overall health and sustainability of coral reef ecosystems [120].
By contrast, marine cold spells can also severely impact ecosystem structure (e.g. [121,122]). Subtropical Florida experienced a severe cold spell in early 2010, with severe impacts for terrestrial and marine species, including coral reef communities [121], non-native crabs [123] and important gamefish [124]. According to Lirman et al. [121], the Florida Reef Tract experienced the most severe coral mortality on record in response to the cold-water anomaly in January 2010, which disproportionately affected shallow reef habitats that had exhibited resilience to prior disturbance events. However, such abnormal cold winters may be a critical ‘reset’ mechanism for marine invasive species, as the cold snap can limit the range expansion of subtropical species [123]. For a non-native crab that had extended its range into the southern US and mid-Atlantic coast from the Caribbean, Canning-Clode et al. [123] suggested that this explained the crab's sudden disappearance after 2010, as the subtropical species had been unable to tolerate the prolonged extreme cold temperatures in early 2010.
(ii). Other extreme climate events affecting marine biological systems
Reviewing how marine organisms in the coastal environment are affected by climate change, Harley et al. [125] distinguished changes in the physical environment related to sea-level rise, changing circulation, pH, CO2 and UV. Emergent ecological responses could be divided into distributional shifts (e.g. zonation patterns and biogeographic ranges), changes in species composition, diversity and community structure, changing primary and secondary production and population dynamics [125]. ECEs are likely a controlling factor of how changes in the physical environment exert their influence on biological systems in the marine environment. In addition to the temperature-related marine heatwaves and cold snaps, these ECEs include, for example, severe storms [126–128], extreme wave activity [129], extreme sea level [130,131] and salinity changes and floods [132–134].
In a synthesis study, Vose et al. [135] examined changes in ECEs associated with extratropical storms, winds and waves, and found that storm frequency and intensity had increased in the Northern Hemisphere cold season since the 1950s, along with an increase in extreme winds over the oceans since the 1980s. Extreme waves along the Pacific US coast have increased moderately since the 1950s, while the evidence for other US coastlines is inconclusive [135]. Extreme wave heights have been observed to increase in many regions around the world, such as for the US Pacific Northwest [135,136], along the South American Pacific coast since the 1980s [137] and for the North Atlantic over the twentieth century [138]. Using a multi-model ensemble, Hemer et al. [139] found the annual mean significant wave height to decrease by 25% globally by 2070–2100, while only 7% of ocean areas, mostly in the Southern Ocean, exhibited an increase over the same timeframe. By contrast, according to Mori et al. [140], significant wave height is projected to increase globally by 15% by the end of the twenty-first century, exceeding the projected changes in surface pressure and wind speed. Similarly, Wang et al. [141] found significant wave height increases in the eastern Pacific and for the Southern Hemisphere extratropics by the end of the twenty-first century. Extreme wave heights are also likely to double and triple in coastal regions, such as for Chile, the Gulf of Bengal, South and East Asian coasts and the Gulf of Mexico due to increased sea-level pressure gradients and surface winds [141].
Changing storm characteristics can have considerable impact on natural systems, as coastal and near-shore biological communities experience physical disturbances in response to extreme wave action. For example, kelp forest structure can be modified by changes in severe storms and the associated wave activity [129]. The 2013–2014 storm season in the Northeast Atlantic was unusually severe, resulting in extensive flooding and exhibiting extreme wave activity [142–144]. Smale & Vance [129] found the warm-water kelp species to be more affected by the stormy 2013–2014 conditions than the more hardy cold-water kelp species. They cautioned that climate-driven shifts towards more mixed canopies in the Northeast Atlantic due to warming temperatures might erode the kelp communities’ resistance to such storm disturbances [129]. For the California coast, Byrnes et al. [126] showed that, while moderate levels of severe storms (i.e. one storm every 3–4 years) help maintain complexity in kelp forest food webs, more frequent severe storms (i.e. at annual frequency) lead to a decrease in diversity in giant kelp forests. In the Adriatic, Perkol-Finkel & Airoldi [145] attributed a loss of subtidal algal forests to several extreme storm events, compounded by long-term human-induced habitat instability.
Sanchez-Vidal et al. [127] highlighted that severe coastal storms do not only affect the shoreline communities, but also have the potential to affect deep-sea ecosystems. This was observed during an exceptionally strong storm along the Spanish coast in December 2008 that initiated shelf sediment movement and redistribution across the adjacent deep basin that caused abrasion and burial of the benthic communities in the Western Mediterranean [127]. The storm also affected the biodiversity of a coralligenous outcrop in the Northwest Mediterranean, with exposed and impacted sites experiencing major shifts in species composition immediately following the storm and loss of cover of benthic species in the range of 22–58%, with fragile species impacted more [146].
Furthermore, extremes in biogeochemical properties in the marine environment can also affect ecosystem structure and functioning. Investigating episodes of high CO2 concentrations in seawater, McNeil & Sasse [147] found that the amplitude of the annual CO2 cycle is increasing with rising greenhouse gas emissions. By the second half of the twenty-first century, major fisheries in the Southern Ocean, Pacific and Atlantic may be periodically exposed to CO2 concentrations that have detrimental physiological and neurological effects on marine animals [147].
5. Challenges and opportunities in assessing how extreme climate events impact biological systems
Despite advances in the field of event attribution in recent decades, challenges remain. Improvements in statistical methodology, observations, climate and weather modelling will probably allow for better understanding of ECEs and event attribution [1]. Event attribution is most skilful when combining evidence based on theory (sound physical understanding of the processes involved), observations (long-term observational records exist that allow placing the event in a historical context) and numerical model simulations (adequately simulated by models to allow replicating the event and for the right reason). In addition, event types purely meteorological in nature, i.e. not confounded by factors, such as resource management or infrastructure, allow for more reliable event attribution [1].
Irrespective of the approach, the success of detection and attribution relies on a model's ability to represent the relevant processes and their interactions over the region and season of interest [39]. Confidence in attributing changes in ECEs to anthropogenic forcing is most pronounced when considering event types related to regional and global temperature [7], such as extreme heat and cold events, hydrological drought and intense precipitation [1]. Improved process representation through better model dynamics, improved model parametrizations, and higher horizontal and vertical model resolution have led to improved representation of regional-scale climate variability. However, considerable further advances are required to represent fine temporal and spatial scales, at which ECEs in precipitation are experienced at a local level ([39], and references therein). In addition, low-frequency natural variability, such as that associated with Atlantic or Pacific Decadal Variability, can affect the reliability of event attributions [1]. Given the shortness of the observational record relative to the multi-decadal nature of these modes of variability, they remain a challenging aspect also for climate model simulations of ECEs [148]. Furthermore, this is not just a challenge for model simulations of ECEs, but also when considering the length of the instrumental record: as recently shown by Abram et al. [149], industrial-era warming commenced as early as the mid-nineteenth century and therefore instrumental records in many regions are too short to comprehensively assess anthropogenic climate change. This has to be taken into consideration when addressing detection and attribution to assess anthropogenic contributions to specific events (cf. also recent reviews in [22] and [24] on event attribution).
On the biological side, data have traditionally been gathered at single sites (e.g. field stations) or more rarely within a region [14]. Extended cross-regional long-term observations are limited, even though sustained monitoring is important for assessing integrated responses of ecosystems to ECEs to account for long-term effects in subsequent years [93]. For example, maintaining long time series has been the key problem in understanding variability and change in marine biodiversity and ecosystems in response to environmental factors [150]. In their review, Jentsch et al. [151] concluded that long-term observations and experimental studies in different ecosystem types and across a range of spatial and temporal scales are crucial for advancing the understanding of how ECEs affect biological systems. The largely local and site-specific nature of existing long-term biological records, paired with still comparatively coarse climatic information from current-generation reanalysis products and climate models leads to a mismatch in the spatial and temporal scale of available data for addressing how biological systems respond to climate variability and change. This is even more exacerbated in the case of ECEs, which are rare events by definition. To sufficiently sample a distribution to allow inferences about its tails, extended time series are required.
Improved understanding of bio-physical interactions across a range of spatial and temporal scales [152] is not only important for quantifying how ECEs affect biological systems. Parmesan [153] considers the current lack of mechanistic understanding of the effect of ecological, behavioural and evolutionary responses to ECEs a crucial limitation in assessing ecosystem adaptation to climate change more generally. In particular, developing process-based concepts of the biological systems’ response to ECEs is crucial for predicting the impacts of changes in the climate system on ecosystem functioning in the future [14].
Recent advances and enhanced capabilities in observing systems provide new avenues for developing a mechanistic framework to understand interactions between the physical climate system and biological processes. To advance understanding of how ECEs affect ecosystem functioning, remote sensing in particular allows for concurrent observations of physical and biological parameters at comparable spatial and temporal resolution [152]. For example, remote sensing with short return intervals at identical locations and near-global coverage facilitates monitoring of soil properties, concurrent vegetation states (e.g. biomass, leaf area index) and radiative properties like fractions of absorbed radiation [18]. Concurrent impacts on plant physiology, photosynthesis, respiration, mechanical damage for trees (e.g. snow and ice breakage, wind throw) and effects on topsoil erosion can thus be documented, as well as lagged impacts like changes in plant phenology, reduced plant growth, increased mortality and changes in plant species composition [18]. Vrieling et al. [154] for example used a remotely sensed normalized difference vegetation index to predict seasonal forage availability ahead of time to cover livestock losses by pastoralist households in East Africa during drought periods through early insurance payments to allow purchase of forage, water or medicines to protect livestock.
When investigating how ECEs impact the terrestrial carbon cycle, Frank et al. [18] found the (sub)tropics to be largely understudied in regard to ground-based case studies when compared with those obtained via remote sensing. To be able to upscale how ECEs affect biological systems and, more specifically, the carbon-climate feedback on a global scale, more extensive regional studies are required [18]. Zscheischler et al. [155] presented a methodological framework to assess how ECEs affect state and functionality of terrestrial ecosystems on a global scale by identifying spatio-temporally contiguous signals of extremes in different Earth observation products. Using the fraction of absorbed photosynthetically active radiation to detect extremes in vegetation activity over the past 30 years, they demonstrated that the size distribution of extremes follows a distinct power law [85]. Furthermore, based on a hierarchy of models ranging from purely data-driven to semi-empirical and dynamic vegetation, land-surface models and remote sensing products, Zscheischler et al. [156] found that the total effect of negative extremes in global primary production is of a similar magnitude as the mean terrestrial carbon sink. Furthermore, carbon cycle extremes exhibit an uneven spatial distribution with ‘hotspot’ regions in many semi-arid monsoon-affected regions and are strongly associated with water scarcity [18,156,157]. The lack of biological observations is particularly true for marine ecosystems, as changes in the ecosystem structure of many habitats (e.g. kelp forests and seagrass meadows) cannot be remotely sensed. Considering that these habitats play a key role in marine carbon cycling and are affected by ECEs, monitoring of marine ecosystems at appropriate spatial and temporal scales is even more lacking than for terrestrial ecosystems.
Meta-analysis is considered a powerful method of quantitative data synthesis in ecological research [158]. This could be combined with extended observational records, remote sensing capabilities and climate model output at increasingly finer resolution, both from global climate models and regional model configurations, to address interactions of the physical and biological systems across a range of temporal and spatial scales.
6. Summary
How ECEs affect ecosystems largely depends on the magnitude, spatial and temporal extent, as well as timing of the anomalous climatic event [93]. As such, perspectives spanning across spatial and temporal scales on how biological and physical systems interact (figure 2) are crucial [152] for improved process understanding of ecosystem responses to ECEs. Limitations in our current observational network (e.g. [11]), both for physical climate system parameters and even more so for long-term ecological monitoring, have hampered progress in this regard. This is especially pronounced when considering ECEs, which by definition are rare events. The observing systems of the physical climate, and even more so for the biological system, are limited with regard to depth (time dimension) or breadth (spatial scales), or both. The mismatch of the available and required scales in observations has been compounded in (global) climate models, suffering from a discrepancy in the explicitly resolved spatial and temporal scales and those required for ecological impact research.
Figure 2.

Spatial and temporal scales of typical ECEs and scales of biological systems (shaded grey). Individuals, populations and ecosystems within these respond to environmental stressors. Red (blue) labels indicate an increase (decrease) in the frequency or intensity of the event, with bold font reflecting confidence in the change. For each ECE type indicated in the figure, ECEs are likely to affect biological systems at all temporal and spatial scales located to the left and below the specific ECE position in the figure (modified from Leonard et al. [159] and Sheehan [160]).
New opportunities for assessing how ECEs modulate structure and functioning of ecosystems arise from recent technological advances in observing systems and instrumentation (e.g. through advanced remote sensing capabilities). These allow for monitoring at increased spatial and temporal resolution for both physical and biological parameters concurrently at appropriate resolution [152]. Parmesan et al. [14] further saw potential for advances in ecological and evolutionary theory (population dynamics, physiological energetics and community structure) leading to greater descriptive and predictive power as a result of better alignment of ECE analyses of biological and physical system parameters. They saw this as a potential outcome of improved coupling of in-depth climatological analyses and biological processes that would allow us to better characterize the complex interactions between climatic conditions and natural systems spanning the spatial and temporal spectrum across which these interactions can occur (cf. figure 2). Bailey & van de Pol [161] also pointed out that multi-event studies that combine long-term field studies and experiments with modelling are crucial for a better understanding of the mechanisms and for improving the predictions of how ECEs affect natural systems. This is especially the case given the rarity of such events and the challenges with collecting ecological time series of sufficient length. Extensions of the historical observational record through climate quality reanalyses or through longer term archives from palaeo proxies (e.g. tree rings, stalagmites and sediments) are also crucial for ensuring a record of sufficient length to reliably quantify trends and sample the characteristics of the ECEs in the physical climate system across a range of spatial and temporal scales [1,93]. Improvements in statistical methodology and in numerical modelling, including but not limited to model resolution and improved parametrizations, provide the necessary tools to advance our understanding of the physical mechanisms that lead to ECEs [1,22] and how their characteristics are changing in a warming world. Stott [7] stressed the importance of developing new methods to conclusively link the changes in ECEs to their meteorological and climatic drivers. This applies similarly to addressing ECEs and their impacts on species, populations and ecosystems as a whole. Current-to-next generation global climate models, along with higher-resolution regional models, provide new tools and opportunities for developing a mechanistic, process-based understanding of where, when and how ECEs impact biological systems.
Endnote
Reanalyses combine observations with an unchanging data assimilation scheme and model to provide a dynamically consistent estimate of the climate state over the instrumental period [38].
Authors' contributions
The paper was written by C.C.U. with contributions from G.A.M.
Competing interests
We have no competing interests.
Funding
Portions of this study were supported by the Regional and Global Climate Modeling Program (RGCM) of the US Department of Energy's Office of Biological & Environmental Research (BER) Cooperative Agreement #DE-FC02-97ER62402, and the National Science Foundation. The National Center for Atmospheric Research is sponsored by the National Science Foundation. C.C.U. acknowledges support from the Ocean Climate Change Institute at WHOI.
References
- 1.NAS. 2016. Attribution of extreme weather events in the context of climate change. Washington, DC: National Academies Press. [Google Scholar]
- 2.IPCC. 2012. Managing the risks of extreme events and disasters to advance climate change adaptation. In A special report of working groups I and II of the intergovernmental panel on climate change (eds Field CB, et al.), p. 582. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 3.IPCC. 2013. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker TF, et al.). Cambridge, UK: Cambridge University Press. [Google Scholar]
- 4.Grotjahn R, et al. 2014. US CLIVAR workshop on analyses, dynamics, and modeling of large-scale meteorological patterns associated with extreme temperature and precipitation events. US CLIVAR Report 2014-2. Washington, DC: US CLIVAR Project Office.
- 5.Bindoff NL, et al. 2013. Detection and attribution of climate change: from global to regional. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker TF, et al.) Cambridge, UK: Cambridge University Press. [Google Scholar]
- 6.Zwiers FW, et al. 2013. Climate extremes: challenges in estimating and understanding recent changes in the frequency and intensity of extreme climate and weather events. In Climate Science Serving Society: Research, Modelling and Prediction Priorities (eds GR Asrar, JW Hurrell), pp. 339–389. Dordrecht, The Netherlands: Springer. [Google Scholar]
- 7.Stott P. 2016. How climate change affects extreme weather events. Science 352, 1517–1518. ( 10.1126/science.aaf7271) [DOI] [PubMed] [Google Scholar]
- 8.Solow AR. 2015. Extreme weather, made by us? Science 349, 1444–1445. ( 10.1126/science.aad2132) [DOI] [PubMed] [Google Scholar]
- 9.Peterson TC, Stott PA, Herring S. 2012. Explaining extreme events of 2011 from a climate perspective. Bull. Am. Meteorol. Soc. 93, 1041–1067. ( 10.1175/BAMS-D-12-00021.1) [DOI] [Google Scholar]
- 10.Herring SC, Hoerling MP, Kossin JP, Peterson TC, Stott PA. 2015. Explaining extreme events of 2014 from a climate perspective. Bull. Am. Meteorol. Soc. 96, S1–S172. [Google Scholar]
- 11.Alexander LV. 2016. Global observed long-term changes in temperature and precipitation extremes: a review of progress and limitations in IPCC assessments and beyond. Weather Clim. Extremes 11, 4–16. ( 10.1016/j.wace.2015.10.007) [DOI] [Google Scholar]
- 12.Smith MD. 2011. The ecological role of climate extremes: current understanding and future prospects. J. Ecol. 99, 651–655. ( 10.1111/j.1365-2745.2011.01833.x) [DOI] [Google Scholar]
- 13.Holmgren M, et al. 2006. Extreme climatic events shape arid and semi-arid ecosystems. Front. Ecol. Environ. 4, 87–95. ( 10.1890/1540-9295(2006)004%5B0087:ECESAA%5D2.0.CO;2) [DOI] [Google Scholar]
- 14.Parmesan C, Root TL, Willig MR. 2000. Impacts of extreme weather and climate on terrestrial biota. Bull. Am. Meteorol. Soc. 81, 443–450. ( 10.1175/1520-0477(2000)081%3C0443:IOEWAC%3E2.3.CO;2) [DOI] [Google Scholar]
- 15.Wernberg T, et al. 2016. Climate-driven regime shift of a temperate marine ecosystem. Science 353, 169–172. ( 10.1126/science.aad8745) [DOI] [PubMed] [Google Scholar]
- 16.Wernberg T, Smale DA, Tuya F, Thomsen MS, Langlois TJ, de Bettignies T, Bennett S, Rousseaux CS. 2013. An extreme climatic event alters marine ecosystem structure in a global diversity hotspot. Nat. Clim. Change 3, 78–82. ( 10.1038/nclimate1627) [DOI] [Google Scholar]
- 17.Palmer G, et al. 2017. Climate change, climatic variation and extreme biological responses. Phil. Trans. R. Soc. B 372, 20160144 ( 10.1098/rstb.2016.0144) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Frank D, et al. 2015. Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Glob. Change Biol. 21, 2861–2880. ( 10.1111/gcb.12916) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yang J, Gong P, Fu R, Zhang M, Chen J, Liang S, Xu B, Shi J, Dickinson R. 2013. The role of satellite remote sensing in climate change studies. Nat. Clim. Change 3, 875–1001. ( 10.1038/nclimate1908) [DOI] [Google Scholar]
- 20.McDowell NG, et al. 2015. Global satellite monitoring of climate-induced vegetation disturbances. Trends Plant Sci. 20, 114–123. ( 10.1016/j.tplants.2014.10.008) [DOI] [PubMed] [Google Scholar]
- 21.Blondeau-Patissier D, Gower JFR, Dekker AG, Phinn SR, Brando VE. 2014. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans. Progr. Oceanogr. 123, 123–144. ( 10.1016/j.pocean.2013.12.008) [DOI] [Google Scholar]
- 22.Easterling DR, Kunkel KE, Wehner MF, Sun L. 2016. Detection and attribution of climate extremes in the observed record. Weather Clim. Extremes 11, 17–27. ( 10.1016/j.wace.2016.01.001) [DOI] [Google Scholar]
- 23.Westra S, Fowler HJ, Evans JP, Alexander LV, Berg P, Johnson F, Kendon EJ, Lenderink G, Roberts NM. 2014. Future changes to the intensity and frequency of short-duration extreme rainfall. Rev. Geophys. 52, 522–555. ( 10.1002/2014RG000464) [DOI] [Google Scholar]
- 24.Stott PA, et al. 2016. Attribution of extreme weather and climate-related events. Wiley Interdiscip. Rev. Clim. Change 7, 23–41. ( 10.1002/wcc.380) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.van de Pol M, Jenouvrier S, Cornelissen JHC, Visser ME. 2017. Behavioural, ecological and evolutionary response to extreme climatic events: challenges and directions. Phil. Trans. R. Soc. B 372, 20160134 ( 10.1098/rstb.2016.0134) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.IPCC. 2012. Summary for policymakers. In Managing the risks of extreme events and disasters to advance climate change adaptation (eds Field CB, et al.), pp. 3–21. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change Cambridge, UK: Cambridge University Press. [Google Scholar]
- 27.Seneviratne SI, et al. 2012. Changes in climate extremes and their impacts on the natural physical environment. In Managing the risks of extreme events and disasters to advance climate change adaptation (eds Field CB, et al.), pp. 109–230. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC) Cambridge, UK: Cambridge University Press. [Google Scholar]
- 28.Fischer EM, Seneviratne SI, Vidale PL, Luthi D, Schär C. 2007. Soil moisture–atmosphere interactions during the 2003 European summer heat wave. J. Clim. 20, 5081–5099. ( 10.1175/JCLI4288.1) [DOI] [Google Scholar]
- 29.Whan K, Zscheischler J, Orth R, Shongwe M, Rahimi M, Asare EO, Seneviratne SI. 2015. Impact of soil moisture on extreme maximum temperatures in Europe. Weather Clim. Extremes 9, 57–67. ( 10.1016/j.wace.2015.05.001) [DOI] [Google Scholar]
- 30.Meehl GA, Tebaldi C, Adams-Smith D. 2016. US daily temperature records past, present and future. Proc. Natl Acad. Sci. USA 113, 13 977–13 982. ( 10.1073/pnas.1606117113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sippel S, Zscheischler J, Heimann M, Otto FEL, Peters J, Mahecha MD. 2015. Quantifying changes in climate variability and extremes: pitfalls and their overcoming. Geophys. Res. Lett. 42, 9990–9998. ( 10.1002/2015GL066307) [DOI] [Google Scholar]
- 32.Ballester J, Giorgi F, Rodo X. 2010. Changes in European temperature extremes can be predicted from changes in PDF central statistics. Clim. Change 98, 277–284. ( 10.1007/s10584-009-9758-0) [DOI] [Google Scholar]
- 33.Hartmann DL, et al. 2013. Observations: atmosphere and surface. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker TF, et al.). Cambridge, UK: Cambridge University Press. [Google Scholar]
- 34.Rhines A, Huybers P. 2013. Frequent summer temperature extremes reflect changes in the mean, not the variance. Proc. Natl Acad. Sci. USA 110, E546 ( 10.1073/pnas.1218748110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fischer EM, Schär C. 2010. Consistent geographical patterns of changes in high-impact European heatwaves. Nat. Geosci. 3, 398–403. ( 10.1038/ngeo866) [DOI] [Google Scholar]
- 36.Hegerl GC, Hoegh-Guldberg O, Casassa G, Hoerling MP, Kovats RS, Parmesan C, Pierce DW, Stott PA. 2010. Good practice guidance paper on detection and attribution related to anthropogenic climate change. In Meeting Report of the Intergovernmental Panel on Climate Change Expert Meeting on Detection and Attribution of Anthropogenic Climate Change (eds Stocker TF, Field CB, Qin D, Barros V, Plattner G-K, Tignor M, Midgley PM, Ebi KL). Bern, Switzerland: University of Bern. [Google Scholar]
- 37.Solow AR. 2017. On detecting ecological impacts of extreme climate events and why it matters. Phil. Trans. R. Soc. B 372, 20160136 ( 10.1098/rstb.2016.0136) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.NCAR. 2016. Climate Data Guide on Atmospheric Reanalysis: overview & comparison tables. https://climatedataguide.ucar.edu/climate-data/atmospheric-reanalysis-overview-comparison-tables (accessed 9 October 2016).
- 39.Sarojini BB, Stott PA, Black E. 2016. Detection and attribution of human influence on regional precipitation. Nat. Clim. Change 6, 669–675. ( 10.1038/nclimate2976) [DOI] [Google Scholar]
- 40.Donat MG, et al. 2013. Updated analyses of temperature and precipitation extreme indices since the beginning of the twentieth century: the HadEX2 dataset. J. Geophys. Res. Atmos. 118, 2098–2118. ( 10.1002/jgrd.50150) [DOI] [Google Scholar]
- 41.Choi G, et al. 2009. Changes in means and extreme events of temperature and precipitation in the Asia-Pacific Network region, 1955–2007. Int. J. Climatol. 29, 1906–1925. ( 10.1002/joc.1979) [DOI] [Google Scholar]
- 42.Donat MG, Alexander LV, Yang H, Durre I, Vose R, Caesar J. 2013. Global land-based datasets for monitoring climatic extremes. Bull. Am. Meteorol. Soc. 94, 997–1006. ( 10.1175/BAMS-D-12-00109.1) [DOI] [Google Scholar]
- 43.Rawlins M, Bradley R, Diaz H, Kimball J, Robinson D. 2016. Future decreases in freezing days across North America. J. Clim. 29, 6923–6935. ( 10.1175/JCLID-15-0802.1) [DOI] [Google Scholar]
- 44.Meehl GA, Tebaldi C, Walton G, Easterling D, McDaniel L. 2009. The relative increase of record high maximum temperatures compared to record low minimum temperatures in the U.S. Geophys. Res. Lett. 36, L23701 ( 10.1029/2009GL040736) [DOI] [Google Scholar]
- 45.Rowe CM, Derry LE. 2012. Trends in record-breaking temperatures for the conterminous United States. Geophys. Res. Lett. 39, L16703 ( 10.1029/2012GL052775) [DOI] [Google Scholar]
- 46.Trewin B, Vermont H. 2010. Change in the frequency of record temperatures in Australia, 1957–2009. Aust. Meteorol. Oceanogr. J. 60, 113–119. ( 10.22499/2.6002.003) [DOI] [Google Scholar]
- 47.Beniston M. 2015. Ratios of record high to record low temperatures in Europe exhibit sharp increases since 2000 despite a slowdown in the rise of mean temperatures. Clim. Change 129, 225–237. ( 10.1007/s10584-015-1325-2) [DOI] [Google Scholar]
- 48.Peterson TC, Zhang XB, Brunet-India M, Vazquez-Aguirre JL. 2008. Changes in North American extermes derived from daily weather data. J. Geophys. Res. Atmos. 113, D07113 ( 10.1029/2007JD009453) [DOI] [Google Scholar]
- 49.Andrade C, Leite S, Santos J. 2012. Temperature extremes in Europe: overview of their driving atmospheric patterns. Nat. Hazards Earth Syst. Sci. 12, 1671–1691. ( 10.5194/nhess-12-1671-2012) [DOI] [Google Scholar]
- 50.Caesar J, et al. 2011. Changes in temperature and precipitation extremes over the Indo-Pacific region from 1971 to 2005. Int. J. Climatol. 31, 791–801. ( 10.1002/joc.2118) [DOI] [Google Scholar]
- 51.Meehl GA, Arblaster JM, Tebaldi C. 2007. Contributions of natural and anthropogenic forcing to changes in temperature extremes over the U.S. Geophys. Res. Lett. 34, L19709 ( 10.1029/2007GL030948) [DOI] [Google Scholar]
- 52.Coumou D, Rahmstorf S. 2012. A decade of weather extremes. Nat. Clim. Change 2, 491–496. ( 10.1038/NCLIMATE1452) [DOI] [Google Scholar]
- 53.Scaife A, Folland C, Alexander L, Moberg A, Knight J. 2008. European climate extremes and the North Atlantic Oscillation. J. Clim. 21, 72–83. ( 10.1175/2007JCLI1631.1) [DOI] [Google Scholar]
- 54.Marrot P, Garant D, Charmantier A. 2017. Multiple extreme climatic events strengthen selection for earlier breeding in a wild passerine. Phil. Trans. R. Soc. B 372, 20160372 ( 10.1098/rstb.2016.0372) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Long OMC, Warren R, Price J, Brereton TM, Botham MS, Franco MA. 2016. Sensitivity of UK butterflies to local climatic extremes: which life stages are most at risk? J. Anim. Ecol. 86, 108–116. ( 10.1111/1365-2656.12594) [DOI] [PubMed] [Google Scholar]
- 56.Fischlin A, et al. 2007. Ecosystems, their properties, goods, and services. In Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change (eds Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE), pp. 211–272. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 57.Handmer J, et al. 2012. Changes in impacts of climate extremes: human systems and ecosystems. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation (eds Field CB, et al.), pp. 231–290. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC) Cambridge, UK: Cambridge University Press. [Google Scholar]
- 58.Trenberth KE, Dai A, Rasmussen RM, Parsons DB. 2003. The changing character of precipitation. Bull. Am. Meteorol. Soc. 84, 1205–1217. ( 10.1175/BAMS-84-9-1205) [DOI] [Google Scholar]
- 59.Meehl GA, Arblaster JM, Tebaldi C. 2005. Understanding future patterns of precipitation in-tensity in climate model simulations. Geophys. Res. Lett. 32, L18719 ( 10.1029/2005GL023680) [DOI] [Google Scholar]
- 60.Held IM, Soden BJ. 2006. Robust responses of the hydrological cycle to global warming. J. Clim. 19, 5686–5699. ( 10.1175/JCLI3990.1) [DOI] [Google Scholar]
- 61.Wentz FJ, Ricciardulli L, Hilburn K, Mears C. 2007. How much more rain will global warming bring? Science 317, 233–235. ( 10.1126/science.1140746) [DOI] [PubMed] [Google Scholar]
- 62.Pall P, Aina T, Stone DA, Stott PA, Nozawa T, Hilberts AGJ, Lohmann D, Allen MR. 2011. Anthropogenic greenhouse gas contribution to flood risk in England and Wales in autumn 2000. Nature 470, 382–385. ( 10.1038/nature09762) [DOI] [PubMed] [Google Scholar]
- 63.Lehmann J, Coumou D, Frieler K. 2015. Increased record-breaking precipitation events under global warming. Clim. Change 132, 501–515. ( 10.1007/s10584-015-1434-y) [DOI] [Google Scholar]
- 64.Westra S, Alexander LV, Zwiers FW. 2013. Global increasing trends in annual maximum daily precipitation. J. Clim. 26, 3904–3918. ( 10.1175/JCLI-D-12-00502.1) [DOI] [Google Scholar]
- 65.Tebaldi C, Hayhoe K, Arblaster JM, Meehl GA. 2006. Going to the extremes: an intercomparison of model-simulated historical and future changes in extreme events. Clim. Change 79, 185–211. ( 10.1007/s10584-006-9051-4) [DOI] [Google Scholar]
- 66.Zhang Y, et al. 2013. Extreme precipitation patterns and reductions of terrestrial ecosystem production across biomes. J. Geophys. Res. Biogeosci. 118, 148–157. ( 10.1029/2012JG002136) [DOI] [Google Scholar]
- 67.Feng X, Porporato A, Rodriguez-Iturbe I. 2013. Changes in rainfall seasonality in the tropics. Nat. Clim. Change 3, 811–815. ( 10.1038/nclimate1907) [DOI] [Google Scholar]
- 68.Evans JP, Boyer-Souchet I. 2012. Local sea surface temperatures add to extreme precipitation in northeast Australia during La Nina. Geophys. Res. Lett. 39, L10803. [Google Scholar]
- 69.Meredith EP, Semenov VA, Maraun D, Park W, Chernokulsky AV. 2015. Crucial role of Black Sea warming in amplifying the 2012 Krymsk precipitation extreme. Nat. Geosci. 8, 615–619. ( 10.1038/ngeo2483) [DOI] [Google Scholar]
- 70.Trenberth KE, Fasullo JT, Shepherd TG. 2015. Attribution of climate extreme events. Nat. Clim. Change 5, 725–730. ( 10.1038/nclimate2657) [DOI] [Google Scholar]
- 71.Ummenhofer CC, Sen Gupta A, England MH, Taschetto AS, Briggs P, Raupach MR. 2015. How did ocean warming affect Australian rainfall extremes during the 2010/2011 La Nina event? Geophys. Res. Lett. 42, 9942–9951. ( 10.1002/2015GL065948) [DOI] [Google Scholar]
- 72.Pook M, Risbey JS, Ummenhofer CC, Briggs PR, Cohen TJ. 2014. A synoptic climatology of heavy rain events in the Lake Eyre and Lake Frome catchments. Front. Environ. Sci. 2, 54 ( 10.3389/fenvs.2014.00054) [DOI] [Google Scholar]
- 73.Poulter B, et al. 2014. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 509, 600–603. ( 10.1038/nature13376) [DOI] [PubMed] [Google Scholar]
- 74.Beniston M. 2004. The 2003 heat wave in Europe: a shape of the things to come? An analysis based on Swiss climatological data and model simulations. Geophys. Res. Lett. 31, L02202 ( 10.1029/2003GL018857) [DOI] [Google Scholar]
- 75.Stott PA, Stone DA, Allen MR. 2004. Human contribution to the European heatwave of 2003. Nature 432, 610–614. ( 10.1038/nature03089) [DOI] [PubMed] [Google Scholar]
- 76.Garcia-Herrera R, Diaz J, Trigo RM, Luterbacher J, Fischer EM. 2010. A review of the European summer heat wave of 2003. Crit. Rev. Environ. Sci. Technol. 40, 267–306. ( 10.1080/10643380802238137) [DOI] [Google Scholar]
- 77.Matsueda M. 2011. Predictability of Euro-Russian blocking in summer of 2010. Geophys. Res. Lett. 38, L06801 ( 10.1029/2010GL046557) [DOI] [Google Scholar]
- 78.Barriopedro D, Fischer EM, Luterbach J, Trigo R, Garcia-Herrera R. 2011. The hot summer of 2010: redrawing the temperature record map of Europe. Science 332, 220–224. ( 10.1126/science.1201224) [DOI] [PubMed] [Google Scholar]
- 79.Trenberth KE, Fasullo JT. 2012. Climate extremes and climate change: the Russian heat wave and other climate extremes of 2010. J. Geophys. Res. 117, D17103 ( 10.1029/2012JD018020) [DOI] [Google Scholar]
- 80.Hoerling M, et al. 2013. Anatomy of an extreme event. J. Clim. 26, 2811–2832. ( 10.1175/JCLI-D-12-00270.1) [DOI] [Google Scholar]
- 81.Meehl GA, Tebaldi C. 2004. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 305, 994–997. ( 10.1126/science.1098704) [DOI] [PubMed] [Google Scholar]
- 82.Christidis N, Jones GS, Stott PA. 2015. Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Clim. Change 5, 46–50. ( 10.1038/nclimate2468) [DOI] [Google Scholar]
- 83.Cassou C, Cattiaux J. 2016. Disruption of the European climate seasonal clock in a warming world. Nat. Clim. Change 6, 589–594. ( 10.1038/nclimate2969) [DOI] [Google Scholar]
- 84.Ciais P, et al. 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 437, 529–533. ( 10.1038/nature03972) [DOI] [PubMed] [Google Scholar]
- 85.Reichstein M, et al. 2013. Climate extremes and the carbon cycle. Nature 500, 287–295. ( 10.1038/nature12350) [DOI] [PubMed] [Google Scholar]
- 86.Corlett RT. 2016. The impacts of droughts on tropical forests. Trends Plant Sci. 21, 584–593. ( 10.1016/j.tplants.2016.02.003) [DOI] [PubMed] [Google Scholar]
- 87.Asner GP, Alencar A. 2010. Drought impacts on the Amazon forest: the remote sensing perspective. New Phytol. 187, 569–578. ( 10.1111/j.1469-8137.2010.03310.x) [DOI] [PubMed] [Google Scholar]
- 88.Breshears DD, et al. 2005. Regional vegetation die-off in response to global-change-type drought. Proc. Natl Acad. Sci. USA 102, 15 144–15 148. ( 10.1073/pnas.0505734102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Rich PM, Breshears DD, White AB. 2008. Phenology of mixed woody herbaceous ecosystems following extreme events: net and differential responses. Ecology 89, 342–352. ( 10.1890/06-2137.1) [DOI] [PubMed] [Google Scholar]
- 90.Phillips OL, et al. 2009. Drought sensitivity of the Amazon rainforest. Science 323, 1344–1347. ( 10.1126/science.1164033) [DOI] [PubMed] [Google Scholar]
- 91.Allen CD, et al. 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manage. 259, 660–684. ( 10.1016/j.foreco.2009.09.001) [DOI] [Google Scholar]
- 92.Wolf S, et al. 2016. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl Acad. Sci. USA 113, 5880–5885. ( 10.1073/pnas.1519620113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Sippel S, Zscheischler J, Reichstein M. 2016. Ecosystem impacts of climate extremes crucially depend on the timing. Proc. Natl Acad. Sci. USA 113, 5768–5770. ( 10.1073/pnas.1605667113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Teng H, Branstator G, Meehl GA, Washington WM. 2016. Projected intensification of subseasonal temperature variability and heat waves in the Great Plains. Geophys. Res. Lett. 43, 2165–2173. ( 10.1002/2015GL067574) [DOI] [Google Scholar]
- 95.Bond NR, Lake PS, Arthington AH. 2008. The impacts of drought on freshwater ecosystems: an Australian perspective. Hydrobiologia 600, 3–16. ( 10.1007/s10750-008-9326-z) [DOI] [Google Scholar]
- 96.Mosley LM. 2015. Drought impacts on the water quality of freshwater systems; review and integration. Earth Sci. Rev. 140, 203–214. ( 10.1016/j.earscirev.2014.11.010) [DOI] [Google Scholar]
- 97.Little JS, McKenzie D, Peterson DL, Westerling AL. 2009. Climate and wildfire area burned in western U.S. ecoprovinces, 1916–2003. Ecol. Appl. 19, 1003–1021. ( 10.1890/07-1183.1) [DOI] [PubMed] [Google Scholar]
- 98.Miller JD, Safford HD, Crimmins M, Thode AE. 2009. Quantative evidence for increasing forest fire severity in the Sierra Nevada and Southern Cascade Mountains, California and Nevada, USA. Ecosystems 12, 16–32. ( 10.1007/s10021-008-9201-9) [DOI] [Google Scholar]
- 99.Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW. 2006. Warming and earlier spring increase western U.S. forest wildfire activity. Science, 313, 940–943. ( 10.1126/science.1128834) [DOI] [PubMed] [Google Scholar]
- 100.Marlon JR, et al. 2012. Long-term perspectives on wildfires in the western USA. Proc. Natl Acad. Sci. USA 109, E535–E543. ( 10.1073/pnas.1112839109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Parisien M-A, Moritz MA. 2009. Environmental control on the distribution of wildfire at multiple spatial scales. Ecol. Monogr. 79, 127–154. ( 10.1890/07-1289.1) [DOI] [Google Scholar]
- 102.Alencar AA, Brando PM, Asner GP, Putz FE. 2015. Landscape fragmentation, severe drought, and the new Amazon forest fire regime. Ecol. Appl. 25, 1493–1505. ( 10.1890/14-1528.1) [DOI] [PubMed] [Google Scholar]
- 103.Cai W, Cowan T, Raupach MR. 2009. Positive Indian Ocean Dipole events precondition southeast Australia bushfires. Geophys. Res. Lett. 36, L19710 ( 10.1029/2009GL039902) [DOI] [Google Scholar]
- 104.Ummenhofer CC, England MH, McIntosh PC, Meyers GA, Pook MJ, Risbey JS, Sen Gupta A, Taschetto AS. 2009. What causes southeast Australia's worst droughts? Geophys. Res. Lett. 36, L04706 ( 10.1029/2008GL036801) [DOI] [Google Scholar]
- 105.Hobday AJ, et al. 2016. A hierarchical approach to defining marine heatwaves. Prog. Oceanog. 141, 227–238. ( 10.1016/j.pocean.2015.12.014) [DOI] [Google Scholar]
- 106.Smale DA, Wernberg T. 2013. Extreme climatic event drives range contraction of a habitat-forming species. Proc. R. Soc. B 280, 20122829 ( 10.1098/rspb.2012.2829) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Mills KE, et al. 2013. Fisheries management in a changing climate: lessons from the 2012 ocean heat wave in the Northwest Atlantic. Oceanography 26, 191–195. ( 10.5670/oceanog.2013.27) [DOI] [Google Scholar]
- 108.Garrabou J, et al. 2009. Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global Change Biology 15, 1090–1103. ( 10.1111/j.1365-2486.2008.01823.x) [DOI] [Google Scholar]
- 109.Scannel HA, Pershing AJ, Alexander MA, Thomas AC, Mills KE. 2016. Frequency of marine heatwaves in the North Atlantic and North Pacific since 1950. Geophys. Res. Lett. 43, 2069–2076. ( 10.1002/2015GL067308) [DOI] [Google Scholar]
- 110.Feng M, Hendon HH, Xie S-P, Marshall AG, Schiller A, Kosaka Y, Caputi N, Pearce A. 2015. Decadal increase in Ningaloo Niño since the late 1990s. Geophys. Res. Lett. 42, 104–112. ( 10.1002/2014GL062509) [DOI] [Google Scholar]
- 111.Di Lorenzo E, Ohman MD. 2013. A double-integration hypothesis to explain ecosystem response to climate forcing. Proc. Natl Acad. Sci. USA 110, 2496–2499. ( 10.1073/pnas.1218022110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Di Lorenzo E, Mantua N. 2016. Multi-year persistence of the 2014–2015 North Pacific marine heatwave. Nat. Clim. Change 6, 1042–1047. ( 10.1038/nclimate3082) [DOI] [Google Scholar]
- 113.NOAA. 2016. 2013–2016 California sea lion unusual mortality event in California. http://www.nmfs.noaa.gov/pr/health/mmume/californiasealions2013.html.
- 114.NOAA. 2015–2016 Large whale unusual mortality event in the Western Gulf of Alaska. http://www.nmfs.noaa.gov/pr/health/mmume/large_whales_2015.html.
- 115.Opar A.2015. Lost at sea: starving birds in a warming world. Audubon, March-April 2015. https://www.audubon.org/magazine/march-april-2015/lost-seastarving-birds-warming-world .
- 116.Whitney FA. 2015. Anomalous winter winds decrease 2014 transition zone productivity in the NE Pacific. Geophys. Res. Lett. 42, 428–431. ( 10.1002/2014GL062634) [DOI] [Google Scholar]
- 117.NOAA. 2016. NOAA Fisheries mobilizes to gauge unprecedented West Coast toxic algal bloom. http://www.nwfsc.noaa.gov/news/features/west_coast_algal_bloom/index.cfm.
- 118.NOAA. 2015. NOAA declares third ever coral bleaching event. http://www.noaanews.noaa.gov/stories2015/100815-noaa-declares-third-ever-global-coral-bleaching-event.html.
- 119.ARC. 2016. Only 7% of the Great Barrier Reef has avoided coral bleaching. https://www.coralcoe.org.au/only-7-of-the-great-barrier-reef-has-avoided-coral-bleaching/.
- 120.Buddemeier RW, Kleypas JA, Aronson RB. 2004. Coral reefs and global climate change: potential contributions of climate change to stresses on coral reef ecosystems . Pew Center on Global Climate Change; http://www.c2es.org/docUploads/Coral_Reefs.pdf. [Google Scholar]
- 121.Lirman D, et al. 2011. Severe 2010 cold-water event caused unprecedented mortality to corals of the Florida Reef Tract and reversed previous survivorship patterns. PLoS ONE 6, e23047 ( 10.1371/journal.pone.0023047) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Firth LB, Knights AM, Bell SS. 2011. Air temperature and winter mortality: implications for the persistence of the invasive mussel, Perna viridis in the intertidal zone of the south-eastern United States. J. Exp. Mar. Biol. Ecol. 400, 250–256. ( 10.1016/j.jembe.2011.02.007) [DOI] [Google Scholar]
- 123.Canning-Clode J, Fowler AE, Byers JE, Carlton JT, Ruiz GM. 2011. ‘Caribbean Creep’ chills out: climate change and marine invasive species. PLoS ONE 6, e29657 ( 10.1371/journal.pone.0029657) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Adams AJ, Hill JE, Kurth BN, Barbour AB. 2012. Effects of a severe cold event on the subtropical, estuarine-dependent common snook, Centropomus undecimalis. Gulf Caribbean Res. 24, 13–21. ( 10.18785/gcr.2401.03) [DOI] [Google Scholar]
- 125.Harley CDG, Hughes AR, Hultgren KM, Miner BG, Sorte CJB, Thornber CS, Rodriguez LF, Tomanek L, Williams SL. 2006. The impacts of climate change on coastal marine systems. Ecol. Lett. 9, 228–241. ( 10.1111/j.1461-0248.2005.00871.x) [DOI] [PubMed] [Google Scholar]
- 126.Byrnes JE, Reed DC, Cardinale BJ, Cavanaugh KC, Holbrooks SJ, Schmitt RJ. 2011. Climate-driven increases in storm frequency simplify kelp forest food webs. Glob. Change Biol. 17, 2513–2524. ( 10.1111/j.1365-2486.2011.02409.x) [DOI] [Google Scholar]
- 127.Sanchez-Vidal A, et al. 2012. Impacts on the deep-sea ecosystem by a severe coastal storm. PLoS ONE 7, e30395 ( 10.1371/journal.pone.0030395) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.De'ath G, Fabricius KE, Sweatman H, Puotinen M. 2012. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl Acad. Sci. USA 109, 17 995–17 999. ( 10.1073/pnas.1208909109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Smale DA, Vance T. 2016. Climate-driven shifts in species’ distributions may exacerbate the impacts of storm disturbances on North-east Atlantic kelp forests. Mar. Freshw. Res. 67, 65–74. ( 10.1071/MF14155) [DOI] [Google Scholar]
- 130.Woodworth PL, Menendez M, Gehrels WR. 2011. Evidence for century-timescale acceleration in mean sea levels and for recent changes in extreme sea levels. Surveys Geophys. 32, 603–618. ( 10.1007/s10712-011-9112-8) [DOI] [Google Scholar]
- 131.Rhein M, et al. 2013. Observations: ocean. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker TF, et al.), pp. 255–316. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 132.Gillanders BM, Kingsford MJ. 2002. Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms. Oceanogr. Mar. Biol. Annu. Rev. 40, 233–309. ( 10.1201/9780203180594.ch5) [DOI] [Google Scholar]
- 133.Marques SC, Azeiteiro UM, Martinho F, Pardal MA. 2007. Climate variability and planktonic communities: the effect of an extreme event (severe drought) in a southern European estuary. Estuar. Coast. Shelf Sci. 73, 725–734. ( 10.1016/j.ecss.2007.03.010) [DOI] [Google Scholar]
- 134.Lejeusne C, Chevaldonne P, Pergent-Martini C, Boudouresque CF, Perez T. 2009. Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 25, 250–260. ( 10.1016/j.tree.2009.10.009) [DOI] [PubMed] [Google Scholar]
- 135.Vose RS, et al. 2014. Monitoring and understanding changes in extremes: extratropical storms, winds, and waves. Bull. Am. Meteorol. Soc. 95, 377–386. ( 10.1175/BAMS-D-12-00162.1) [DOI] [Google Scholar]
- 136.Ruggiero P, Komar PD, Allan JC. 2010. Increasing wave heights and extreme value projections: the wave climate of the U.S. Pacific Northwest. Coastal Eng. 57, 539–552. ( 10.1016/j.coastaleng.2009.12.005) [DOI] [Google Scholar]
- 137.Izaguirre C, Mendez FJ, Espejo A, Losada IJ, Reguero BG. 2013. Extreme wave climate change in Central-South America. Clim. Change 119, 277–290. ( 10.1007/s10584-013-0712-9) [DOI] [Google Scholar]
- 138.Bertin X, Prouteau E, Letetrel C. 2013. A significant increase in wave height in the North Atlantic Ocean over the 20th century. Glob. Planet. Change 106, 77–83. ( 10.1016/j.gloplacha.2013.03.009) [DOI] [Google Scholar]
- 139.Hemer MA, Fan Y, Mori N, Semedo A, Wang XL. 2013. Projected changes in wave climate from a multi-model ensemble. Nat. Clim. Change 3, 471–476. ( 10.1038/nclimate1791) [DOI] [Google Scholar]
- 140.Mori N, Shimura T, Yasuda T, Mase H. 2013. Multi-model climate projections of ocean surface variables under different climate scenarios—future change of waves, sealevel and wind. Ocean Eng. 71, 122–129. ( 10.1016/j.oceaneng.2013.02.016) [DOI] [Google Scholar]
- 141.Wang XL, Feng Y, Swail VR. 2014. Changes in global ocean wave heights as projected using multimodel CMIP5 simulations. Geophys. Res. Lett. 41, 1026–1034. ( 10.1002/2013GL058650) [DOI] [Google Scholar]
- 142.Huntingford C, et al. 2014. Potential influences on the United Kingdom's floods of winter 2013/14. Nat. Clim. Change 4, 769–777. ( 10.1038/nclimate2314) [DOI] [Google Scholar]
- 143.Matthews T, Murphy C, Wilby RL, Harrigan S. 2014. Stormiest winter on record for Ireland and UK. Nat. Clim. Change 4, 738–740. ( 10.1038/nclimate2336) [DOI] [Google Scholar]
- 144.Masselink G, Castelle B, Scott T, Dodet G, Suanez S, Jackson D, Floc'h F. 2016. Extreme wave activity during 2013/2014 winter and morphological impacts along the Atlantic coast of Europe. Geophys. Res. Lett. 43, 2135–2143. ( 10.1002/2015GL067492) [DOI] [Google Scholar]
- 145.Perkol-Finkel S, Airoldi L. 2010. Loss and recovery potential of marine habitats: an experimental study of factors maintaining resilience in subtidal algal forests at the Adriatic Sea. PLoS ONE 5, e10791 ( 10.1371/journal.pone.0010791) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Teixido N, Casas E, Cebrian E, Linares C, Garrabou J. 2013. Impacts on coralligenous outcrop biodiversity of a dramatic coastal storm. PLoS ONE 8, e53742 ( 10.1371/journal.pone.0053742) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.McNeil BI, Sasse TP. 2016. Future ocean hypercapnia driven by anthropogenic amplification of the natural CO2 cycle. Nature 529, 383–386. ( 10.1038/nature16156) [DOI] [PubMed] [Google Scholar]
- 148.Meehl GA, Zwiers F, Evans J, Knutson T, Mearns LO, Whetton P. 2000. Trends in extreme weather and climate events: issues related to modeling extremes in projections of future climate change. Bull. Am. Meteorol. Soc. 81, 427–436. ( 10.1175/1520-0477(2000)081%3C0427:TIEWAC%3E2.3.CO;2) [DOI] [Google Scholar]
- 149.Abram, et al. 2016. Early onset of industrial-era warming across the oceans and continents. Nature 536, 411–418. ( 10.1038/nature19082) [DOI] [PubMed] [Google Scholar]
- 150.Mieszkowska N, Sugden H, Firth LB, Hawkins SJ. 2014. The role of sustained observations in tracking impacts of environmental change on marine biodiversity and ecosystems. Phil. Trans. R. Soc. A 372, 20130339 ( 10.1098/rsta.2013.0339) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Jentsch A, Kreyling J, Beierkuhnlein C. 2007. A new generation of climate-change experiments: events, not trends. Front. Ecol. Environ. 5, 365–374. ( 10.1890/1540-9295(2007)5%5B365:ANGOCE%5D2.0.CO;2) [DOI] [Google Scholar]
- 152.Prairie JC, Sutherland KR, Nickols KJ, Kaltenberg AM. 2012. Biophysical interactions in the plankton: a cross-scale review. Limnol. Oceanogr. Fluids Environ. 2, 121–145. ( 10.1215/21573689-1964713) [DOI] [Google Scholar]
- 153.Parmesan C. 2006. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669. ( 10.1146/annurev.ecolsys.37.091305.110100) [DOI] [Google Scholar]
- 154.Vrieling A, Meroni M, Mude AG, Chantarat S, Ummenhofer CC, de Bie CAJM. 2016. Early assessment of seasonal forage availability for mitigating the impact of drought on East African pastoralists. Remote Sensing Environ. 174, 44–55. ( 10.1016/j.rse.2015.12.003) [DOI] [Google Scholar]
- 155.Zscheischler J, Mahecha MD, Harmeling S, Reichstein M. 2013. Detection and attribution of large spatiotemporal extreme events in Earth observation data. Ecol. Informatics 15, 66–73. ( 10.1016/j.ecoinf.2013.03.004) [DOI] [Google Scholar]
- 156.Zscheischler J, et al. 2014. A few extreme events dominate interannual variability in gross primary production. Environ. Res. Lett. 9, 035001 ( 10.1088/1748-9326/9/3/035001) [DOI] [Google Scholar]
- 157.Zscheischler J, Reichstein M, Harmeling S, Rammig A, Tomelleri E, Mahecha MD. 2014. Extreme events in gross primary production: a characterization across continents. Biogeosciences 11, 2909–2924. ( 10.5194/bg-11-2909-2014) [DOI] [Google Scholar]
- 158.Hays GC, Richardson AJ, Robinson C. 2005. Climate change and marine plankton. Trends Ecol. Evol. 20, 337–344. ( 10.1016/j.tree.2005.03.004) [DOI] [PubMed] [Google Scholar]
- 159.Leonard M, et al. 2014. A compound event framework for understanding extreme impacts. WIREs Clim. Change 5, 113–128. ( 10.1002/wcc.252) [DOI] [Google Scholar]
- 160.Sheehan P. 1995. Assessments of ecological impacts on a regional scale. In SCOPE 53—Methods to assess the effects of chemicals on ecosystems. http://www.scopenvironment.org/downloadpubs/scope53/chapter14.html.
- 161.Bailey LD, van de Pol M. 2016. Tackling extremes: challenges for ecological and evolutionary research on extreme climatic events. J. Anim. Ecol. 85, 85–96. ( 10.1111/1365-2656.12451) [DOI] [PubMed] [Google Scholar]