

Abstract
Objective
Fibroblast growth factor 21 (FGF21) shows great potential for the treatment of obesity and type 2 diabetes, as its long-acting analogue reduces body weight and improves lipid profiles of participants in clinical studies; however, the intracellular mechanisms mediating these effects are poorly understood. AMP-activated protein kinase (AMPK) is an important energy sensor of the cell and a molecular target for anti-diabetic medications. This work examined the role of AMPK in mediating the glucose and lipid-lowering effects of FGF21.
Methods
Inducible adipocyte AMPK β1β2 knockout mice (iβ1β2AKO) and littermate controls were fed a high fat diet (HFD) and treated with native FGF21 or saline for two weeks. Additionally, HFD-fed mice with knock-in mutations on the AMPK phosphorylation sites of acetyl-CoA carboxylase (ACC)1 and ACC2 (DKI mice) along with wild-type (WT) controls received long-acting FGF21 for two weeks.
Results
Consistent with previous studies, FGF21 treatment significantly reduced body weight, adiposity, and liver lipids in HFD fed mice. To add, FGF21 improved circulating lipids, glycemic control, and insulin sensitivity. These effects were independent of adipocyte AMPK and were not associated with changes in browning of white (WAT) and brown adipose tissue (BAT). Lastly, we assessed whether FGF21 exerted its effects through the AMPK/ACC axis, which is critical in the therapeutic benefits of the anti-diabetic medication metformin. ACC DKI mice had improved glucose and insulin tolerance and a reduction in body weight, body fat and hepatic steatosis similar to WT mice in response to FGF21 administration.
Conclusions
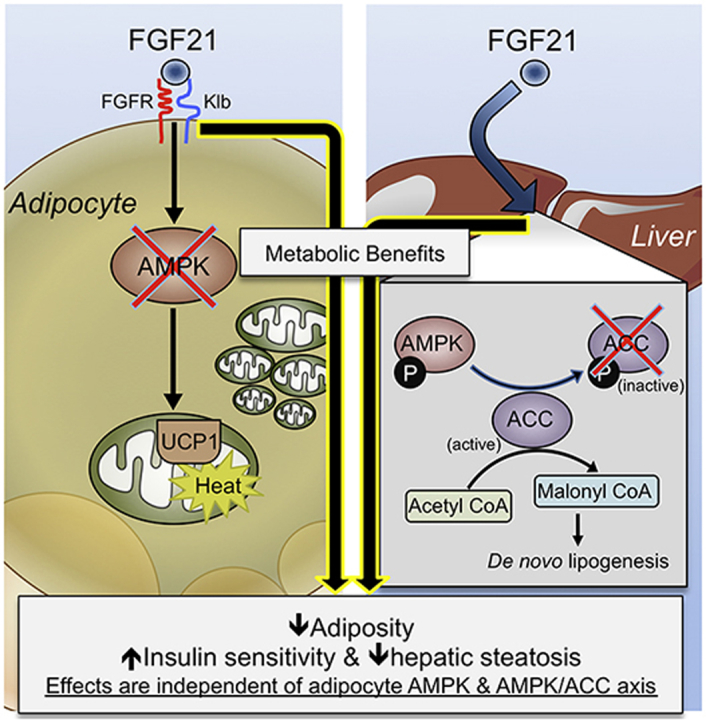
These data illustrate that the metabolic improvements upon FGF21 administration are independent of adipocyte AMPK, and do not require the inhibitory action of AMPK on ACC. This is in contrast to the anti-diabetic medication metformin and suggests that the treatment of obesity and diabetes with the combination of FGF21 and AMPK activators merits consideration.
Keywords: FGF21, AMPK, ACC, Adipocyte, Brown fat, Obesity, Diabetes
Abbreviations: ACC, acetyl-CoA carboxylase; ACC DKI, ACC1-S79A and ACC2-S212A double knock-in; AKT, protein kinase B; AMPK, AMP-activated protein kinase; BAT, brown adipose tissue; COX, cytochrome c oxidase; CreERT2, Cre recombinase – estrogen receptor T2; CNS, central nervous system; DAG, diacylglycerol; FFA, free fatty acid; FGF21, fibroblast growth factor 21; FGFR1c, fibroblast growth factor receptor 1c; GTT, glucose tolerance test; gWAT, gonadal white adipose tissue; H&E, hematoxylin and eosin; HFD, high fat diet; iβ1β2AKO, inducible AMPK β1β2 adipocyte knockout; ITT, insulin tolerance test; iWAT, inguinal white adipose tissue; KLB, beta klotho; RER, respiratory exchange ratio; mTORC1, mammalian target of rapamycin; NAFLD, non-alcoholic fatty liver disease; TAG, triacylglycerol; UCP1, uncoupling protein 1; WAT, white adipose tissue; WT, wildtype
Graphical abstract
Highlights
-
•
FGF21 reduces adiposity and improves insulin resistance in mice fed a high-fat diet.
-
•
FGF21 improves insulin sensitivity and hepatic steatosis independent of adipocyte AMPK.
-
•
FGF21 treatment does not elicit an increase in browning of BAT or WAT.
-
•
In contrast to metformin, FGF21's intracellular mechanism is not through AMPK/ACC.
-
•
Findings suggest that combination of FGF21 and AMPK activators could be of benefit.
1. Introduction
Initially identified as a hormone secreted by the liver that could potently stimulate glucose uptake in adipocytes [1], fibroblast growth factor 21 (FGF21) is an endocrine factor that exerts potent anti-obesity and anti-diabetic effects in rodents and non-human primates [2], [3], [4], [5], [6], [7], [8]. Furthermore, long-acting FGF21 analogues in phase 1 clinical trials decrease body weight and improve the lipid profile of patients with type 2 diabetes [6], [9]. In pre-clinical rodent models, FGF21 administration increases energy expenditure and improves glucose and lipid homeostasis [2]. These effects have been related to increases in glucose uptake and triglyceride clearance in white (WAT) and brown adipose tissue (BAT) [10] and require the expression of the FGF21 receptor, fibroblast growth factor receptor 1c (FGFR1c), and cofactor beta klotho (KLB) [11], [12]. Findings from several FGF21 studies have also shown increases in the browning of WAT; a process whereby WAT acquires characteristics of brown fat such as increases in mitochondrial number and the expression of uncoupling protein 1 (UCP1). While recent studies have suggested that metabolic improvements mediated by FGF21 do not require UCP1 [13], [14] nor intrascapular BAT [2], other groups have shown that UCP1 is required for FGF21-mediated glucose disposal [15] (see Straub and Wolfrum for a minireview [16]). Although it has recently been recognized that FGF21 may exert many of its pharmacological effects through the central nervous system (CNS) [17], the intracellular mechanisms mediating the beneficial metabolic effects of FGF21 on adipose tissue metabolism in vivo are currently not fully elucidated.
Previous studies have shown that FGF21 administration activates the energy sensor AMP-activated protein kinase (AMPK) to promote mitochondrial biogenesis and greater mitochondrial oxidative function in cultured rodent and human adipocytes [18]. AMPK is a central regulator of the energy status of the cell, regulating a myriad of metabolic pathways [19], [20], [21], [22]. AMPK in WAT and BAT is activated by sympathetic drive through mechanisms that are not fully understood but are thought to occur, at least in part, by increased lipolysis [23], [24], [25]. Our previous work demonstrated that adipocyte AMPK is required for acute BAT-mediated thermogenesis, the browning of WAT in response to β3-adrenergic stimuli, and protection against the deleterious effects of a high fat diet (HFD) via the regulation of the mitochondrial quality control pathway (mitophagy) [23]. Furthermore, the inducible deletion of adipocyte AMPK in adult mice did not alter adipocyte lipolysis [23]. However, whether adipocyte AMPK is important for the glucose and lipid-lowering effects of FGF21 remains to be determined.
Chronic FGF21 treatment reduces hepatic triglyceride levels by promoting fat oxidation [26], [27] and suppressing de novo lipogenesis in the liver of rodents, effects associated with an increase in AMPK activation [2]. In addition, discrete effects of FGF21, such as suppression of hepatic glucose output [28], are thought to involve action on the liver [2], [29], [30]. Interestingly, the therapeutic effects of metformin, a first line therapy for the control of blood glucose levels, requires AMPK regulation of acetyl-CoA carboxylase (ACC) to suppress de novo lipogenesis and promote hepatic fat oxidation [31]. Together, these data suggest that FGF21 might also mediate improvements in blood glucose levels through the AMPK/ACC axis in the liver.
Considering that FGF21 is in clinical trials for the treatment of obesity and diabetes, a deeper understanding of the responsible in vivo signaling pathways has obvious medical benefits. In particular, both clinical studies published to date do not demonstrate robust glycemic control, despite weight loss [6], [9]. Since adipose tissue is a key organ that directly mediates the effects of FGF21 [6], [17], [32], [33], a better understanding of adipocyte-specific signaling events is required in the context of improvements in whole-body metabolism. Moreover, since metformin is a first line therapy for glycemic control in diabetic patients, it would be of great benefit to understand if FGF21 also requires the AMPK/ACC axis to improve blood glucose levels. To this end, we utilized an inducible model for the deletion of AMPK in adipocytes of adult mice [23] to examine whether the metabolic improvements of FGF21 therapy require adipocyte AMPK in vivo. To circumvent the deleterious effects of AMPK removal prior to HFD challenge [23], we performed deletion of adipocyte AMPK following HFD treatment. We also tested whether FGF21 requires AMPK-mediated phosphorylation of ACC to improve blood glucose and hepatic lipid levels by utilizing a knock-in model in which AMPK lacks the ability to inactivate ACC. Overall, we demonstrated that the beneficial metabolic effects of pharmacological administration of FGF21 on energy metabolism, fuel selection, body weight, insulin tolerance, and liver lipids are not mediated through adipocyte AMPK or the upregulation of a thermogenic program in BAT or inguinal WAT (iWAT). Additionally, FGF21 administration does not require the AMPK/ACC pathway to reduce hepatic lipid content or improve glucose homeostasis and insulin sensitivity. Taken together, these findings offer new insights into the mechanism of FGF21 action and open up the possibility of understanding the disconnect between rodent and clinical data on glucose endpoints.
2. Materials and methods
2.1. Animals
All experiments were approved by the McMaster University Animals Ethics Committee and conducted under appropriate Canadian guidelines for animal research. Mice lacking adipocyte AMPK β1β2 (iβ1β2AKO) along with littermate controls were generated as previously described [23]. Tamoxifen (Cayman Chemical; MI, USA), dissolved in sunflower oil, was given via oral gavage for five consecutive days in adult males to induce the deletion of adipocyte AMPK β1β2 after 10 weeks of a high fat diet (HFD, 45% kcal fat) (D12451, Research Diets; New Brunswick, New Jersey). At week 14 of HFD, control and iβ1β2AKO mice were anesthetized with Avertin (2,2,2-tribromoethanol dissolved in methyl-2-butanol, Sigma-Aldrich; ON, Canada; 0.5 mg/g body) and implanted dorsally with two week mini-osmotic pumps (1002, Alzet; CA, USA) delivering 0.35 mg/kg/day of native FGF21 or 0.9% sodium chloride (saline) and incisions closed with perma-hand silk suture (Ethicon; ON, Canada). Food and water were provided ad libitum and mice were housed at 23 °C with a 12-hour light/dark cycle.
Male ACC1-S79A and ACC2-S212A knock-in mutation mice (ACC DKI) were as previously described and were put on a 45% HFD at 3–4 months of age [31]. At week 10 of HFD, wildtype and ACC DKI mice received subcutaneous injections of long-acting FGF21 (PF-05231023) at 10 mg/kg twice a week for 2 weeks.
2.2. Metabolic parameters and blood measurements
Metabolic monitoring was performed using a Comprehensive Lab Animal Monitoring System (CLAMS, Columbus Instruments; OH, USA). Body fat was determined using a time-domain NMR whole-body composition analyzer (minispec LF90II, Bruker; MA, USA) and normalized to body weight. A glucose tolerance test (GTT) or insulin tolerance test (ITT) was performed with 6-hour fasted mice on day 13 of treatment, after intraperitoneal injection of 1.0 g/kg of glucose or 1 U/kg insulin, respectively, for HFD-fed mice [34]. Blood glucose levels were determined using a One Touch Ultra Glucometer (LifeScan, Canada). FFA were measured by isolating plasma via centrifugation at 4 °C, diluting in PBS at a 1:5 ratio, and using a non-esterified fatty acid colorimetric assay kit (NEFA-HR 2, Wako Diagnostics; VA, USA). Plasma insulin and adiponectin were quantified by ELISA (Millipore).
2.3. In-vivo glucose clearance
Tissue-specific glucose uptake, stimulated by 1.0 U/kg of insulin via intraperitoneal injection, was performed by tail vein injection of 2 μCi of 3H-2-deoxy-d-glucose (2-DG) 15 min prior to sacrifice, as previously described [35]. Rates of tissue-specific glucose uptake were measured by analyzing radioactivity from tissues in both the deproteinized and proteinized state using 0.3 mM BaOH and 0.3 mM Zn(SO4) (Sigma-Aldrich; ON, Canada).
2.4. Immunoblotting and cytochrome c oxidase (COX) activity
Tissue lysates were prepared in lysis buffer (50 mM HEPES pH 7.4, 150 mM NaCl, 100 mM NaF, 10 Na-pyrophosphate, 5 EDTA mM, 250 mM sucrose, 1 mM DTT, and 1 mM Na-orthovanadate, 1% Triton X, and Complete protease inhibitor cocktail (Roche)). Western samples were prepared using 4x SDS sample buffer (40% glycerol, 240 mM, Tris–HCl pH 6.8, 8% SDS, 0.04% bromophenol blue, 5% β-mercaptoethanol, with a 1:50 dilution of 1 M DTT) and boiled at 95 °C for 5 min. 20 μg of protein was loaded per well for iWAT and 5 μg of protein for BAT, as this amount avoids oversaturation of the UCP1 signal. Immunoblotting was performed as previously described [23]. Using antibodies purchased from Cell Signaling Technology (AKT, #9272; pAKT Ser473, #4058; anti-Rabbit IgG horseradish peroxidase (HRP)-linked, #7074; and anti-Mouse IgG HRP-linked; #7076), Alpha Diagnostic International (UCP1, #UCP11-A), and Invitrogen (β-tubulin, #32-2600), blocked membranes were incubated in primary antibody overnight at 4 °C (5% BSA in TBST).
In order to determine phospho/total protein ratios, membranes were first probed for the phospho protein, stripped for 30 min at 37 °C using Restore Plus (Thermo Scientific; ON, Canada) to ensure removal of phospho signal, and re-probed overnight using antibody against the total protein. UCP1 protein levels were normalized to the loading control β-tubulin. Densitometrical analysis was performed using Image J Software (NIH; MD, USA).
COX activity was measured in tissue lysates as prepared above and as previously described [23].
2.5. Histology
Samples harvested from mice were initially stored and fixed in 10% formalin (24–48 h) and subsequently processed for paraffin embedding and stained with H&E by the Department of Pathology and Molecular Medicine at McMaster University Children's Hospital. Images of the liver (20x) and BAT (10x) were taken using a Nikon 90i Eclipse (Nikon Inc.; NY, USA).
2.6. Lipid analysis
Plasma and lipid analysis was performed as previously described [6]. Briefly, liver tissue samples were homogenized in Methanol:Water (1:1 v/v) and then lipids were extracted with Dichloromethane: Isopropanol: Methanol (25:10:65, v/v/v) and analyzed by mass spec against internal standards. Liver triglyceride levels in ACC-DKI mice were measured using a Triglyceride Assay Kit (Cayman Chemical) as previously described [23].
2.7. Real-time quantitative PCR (RT-qPCR)
RT-qPCR was performed to determine mRNA expression of browning and mitochondrial markers as previously described [36]. Briefly, RNA was isolated using TRIzol reagent (Invitrogen; CA, USA) and applied to columns (RNeasy kit, Qiagen; CA, USA) for purification. All Taqman primers were purchased from Invitrogen [23] (Hadh, Mm00492535_m1; Cpt1b, Mm00487191_g1; Ppargc1a, Mm01208835_m1; Ppara, Mm00440939_m1; Ucp1, Mm01244861_m1; Ppia, Mm02342430_g1; Mt-co2, Mm03294838_g1; Cox8b, Mm00432648_m1; Cidea, Mm00432554_m1), and relative gene expression was calculated using the comparative Ct (2-ΔCT) method. Values were normalized to the housekeeping gene Ppia and subsequently expressed as relative to control saline.
2.8. Statistical analyses
Results were analyzed by two-way ANOVA using GraphPad Prism software. Repeated measures two-way ANOVA was used for GTT and ITT data. All values are presented as mean ± SEM and significance was accepted at p ≤ 0.05.
3. Results
3.1. FGF21 increases energy expenditure, reduces adiposity, and improves blood parameters independent of adipocyte AMPK
To examine the role of adipocyte AMPK in mediating the metabolic effects of FGF21, double floxed AMPK β1 and β2 mice with or without the Adiponectin Cre recombinase – estrogen receptor T2 (CreERT2) transgene were subjected to HFD treatment for 10 weeks followed by tamoxifen-induced deletion of AMPK (Figure 1A). We evaluated the efficiency of deletion by determining phosphorylation of ACC, the most sensitive method for detecting cellular AMPK activity, and found no detectable phosphorylation of ACC in BAT and iWAT of mice deficient for AMPK (Supplemental Figure 1). Following three weeks to allow for the appropriate genetic recombination and recovery, mice treated with native FGF21 for two weeks showed increased oxygen consumption and energy expenditure (Figure 1B–D) and lowered respiratory exchange ratio (R.E.R.) (Figure 1E) in both control and mice deficient for AMPK in adipocytes. The increase in metabolic activity by FGF21 was similar when the data were normalized to lean body mass (Figure 1B vs. C) and was not due to differences in food intake and physical activity, as these parameters were unchanged upon FGF21 administration (Figure 1F and G). Concomitant with greater metabolic activity, FGF21 treatment reduced body weight (Figure 1H) and adiposity (Figure 1I) in both genotypes. Overall, these data indicate that FGF21 does not require adipocyte AMPK to mediate an increase in energy expenditure and a reduction in body fat.
Figure 1.

Native FGF21 increases energy metabolism, fuel selection, and body weight regulation in both control and iβ1β2AKO mice. (A) timeline of experiment with 45% high fat diet (HFD), in weeks, consisting of a week of tamoxifen (TMX) treatment to induce the deletion of both AMPK beta subunits (β1 and β2) only in adipocytes, and two weeks of native FGF21 administration using implanted osmotic mini-pumps. Mice were maintained on a HFD throughout the illustrated timeline and FGF21 treatment was administered for two weeks. (B–G) oxygen consumption (VO2) normalized to whole body mass (B) and lean body mass (C), energy expenditure normalized to lean body mass (D) respiratory exchange ratio (R.E.R.) (E), food intake (F), and activity levels (G) of control and iβ1β2AKO mice treated with saline or native FGF21 for 14 days (n = 4 per group). Change in body weights (H) and fat percentage (I) of control and iβ1β2AKO mice treated with saline or native FGF21 for 14 days (n = 4–7 per group). Data are means ± SEM with ††† p < 0.001, †† p < 0.01, † p < 0.05 denoting a general treatment effect as determined by a two-way ANOVA.
Adiponectin has been suggested to be an important mediator for the insulin sensitizing effects of FGF21 [3], [37] and is an activator of AMPK [38]. Indeed, FGF21 administration increases circulating adiponectin in humans [6], [9]. FGF21 administration caused a modest trend of increased circulating adiponectin levels in control (p = 0.068) (Figure 2A) but not in iβ1β2AKO mice; however, serum insulin levels were dramatically reduced in both genotypes (Figure 2B). Despite the much lower insulin levels, WT and iβ1β2AKO mice treated with FGF21 had lower fed (Figure 2C) and fasted blood glucose levels (Figure 2D). FGF21 treatment also lowered plasma free fatty acid (FFA) and triacylglycerol (TAG) levels in both genotypes to a similar extent (Figure 2E and F). Overall, these data indicate that FGF21 lowers serum insulin, blood glucose, and plasma TAGs and FFAs independently of adipocyte AMPK.
Figure 2.

The effects of native FGF21 on specific blood parameters in control and iβ1β2AKO mice. Circulating adiponectin (A) and insulin (B) levels of control and iβ1β2AKO mice treated with saline or native FGF21 for 14 days (n = 4–7 per group). Blood glucose levels in the fed (C) and 12 h fasted (D) state (n = 4–7 per group). E and F: end-point plasma triacylglycerol (E) and free fatty acids (FFA) (F) of Control and iβ1β2AKO mice treated with saline or native FGF21 (n = 4–7 per group). Data are means ± SEM with ††† p < 0.001 denoting a general treatment effect and * p < 0.05 denoting a general genotype effect as determined by a two-way ANOVA.
3.2. FGF21 improves insulin sensitivity and promotes glucose uptake in BAT independently of adipocyte AMPK
Consistent with lower blood glucose and plasma insulin, FGF21 improved insulin-mediated glucose disposal in both control and iβ1β2AKO mice (Figure 3A and B). To determine which tissues were mediating the improvements in insulin-mediated glucose uptake, we assessed 2-deoxyglucose uptake following an intraperitoneal injection of insulin. FGF21 increased glucose uptake in BAT (Figure 3C), but this effect was not significant in iWAT (Figure 3D). Interestingly, FGF21 had an overall effect of lowering glucose uptake in gonadal WAT (gWAT), liver and quadriceps muscle (Figure 3E–G). To determine if FGF21 treatment was associated with improvements in adipose tissue insulin signaling, and whether there were differences in iβ1β2AKO mice, we examined AKT phosphorylation status in BAT and iWAT. FGF21 treatment increased phosphorylation of AKT in both BAT (Figure 3H and I) and iWAT (Figure 3H and J) following an acute injection of insulin. These data indicate that FGF21 treatment improves BAT insulin action and that these effects do not require AMPK.
Figure 3.

Control and iβ1β2AKO mice treated with native FGF21 for two weeks have improved insulin sensitivity. A and B: insulin tolerance test (ITT: 1 U/kg) (A) and area under the curve (AUC) (B) of control and iβ1β2AKO mice treated with native FGF21 on indicated diet (n = 4–7 per group). C–G: tissue glucose uptake measured using 2-DG of BAT (C), iWAT (D), gWAT (E), liver (F), and quadriceps muscle (G) in control and iβ1β2AKO mice treated with saline or native FGF21 (n = 4–7 per group). H–J: representative total and phospho (S473) AKT immunoblotting (H) with quantification in BAT (I: n = 4–7 per group) and iWAT (J: n = 3–7 per group) of 10-week HFD and 2-week treated (saline or native FGF21) control and iβ1β2AKO mice. Data are means ± SEM with ††† p < 0.001, †† p < 0.01, † p < 0.05 denoting a general treatment effect as determined by a two-way ANOVA (A: two-way repeated measures ANOVA).
3.3. FGF21 reduces hepatic steatosis without increasing WAT browning
FGF21 reduces non-alcoholic fatty liver disease (NAFLD) [2], [5] while inducible deletion of adipocyte AMPK prior to HFD treatment promotes NAFLD [23]. Consistent with previous reports [2], [29], FGF21 treatment resulted in lower hepatic lipid content, as indicated by lower lipid-laden white area present within cells in hematoxylin and eosin (H&E) stains (Figure 4A), which was further confirmed by a reduction in liver diacylglycerol (DAG) (Figure 4B) and TAG (Figure 4C) levels. In contrast to DAG and TAG levels, liver ceramide levels were increased with FGF21 treatment (Figure 4D).
Figure 4.

Treatment with native FGF21 for two weeks reduces liver lipids in mice fed a high fat diet (45%) for 10 weeks. (A) representative H&E liver stains (20x) of control and iβ1β2AKO mice treated with saline or native FGF21. B and C: Liver levels of diacylglycerol (B) and triacylglycerol (C) as measured by mass spec (n = 4–7 per group). (D) Liver ceramides as measured by mass spec (n = 4–7 per group). Data are means ± SEM with †† p < 0.01 denoting a general treatment effect as determined by a two-way ANOVA.
FGF21 has been associated with improvements in BAT function and increased WAT browning; however, the role of UCP1 and WAT browning in mediating the metabolic improvements induced by pharmacological administration of FGF21 are somewhat dichotomous [13], [15], [16]. FGF21 treatment reduced both BAT and iWAT mass (Figure 5A and B, respectively) and reduced the presence of lipid in BAT, as indicated by H&E staining (Figure 5C). FGF21 treatment had a modest overall effect on increasing UCP1 and MT-CO2 mRNA levels in BAT of control and iβ1β2AKO mice (Figure 5D), but not in iWAT (Figure 5F). Consistent with an increase in MT-CO2, FGF21 increased mitochondrial cytochrome c oxidase (COX) activity in the BAT of both control and iβ1β2AKO mice (Figure 5E). FGF21 increased Ppargc1a mRNA levels in iWAT, but the expression of other browning markers was unaffected (Figure 5F). FGF21 treatment did not affect UCP1 protein levels in BAT, while there was an overall effect for lower UCP1 levels in the BAT of iβ1β2AKO mice (Figure 5G and H). The expression of UCP1 protein in iWAT, while highly variable, was unaffected by FGF21 treatment in either Control or iβ1β2AKO mice (Figure 5G and I).
Figure 5.

The beneficial effects of native FGF21 in control and iβ1β2AKO mice are not driven by the upregulation of a thermogenic program in vivo. A and B: tissue weights of BAT (A) and iWAT (B) of control and iβ1β2AKO mice treated with saline or native FGF21 for 14 days (n = 4–7 per group). (C) representative H&E stains (10x) of BAT from control and iβ1β2AKO mice fed a high fat diet (45%) for 10 weeks and treated with saline or native FGF21 for 14 days. D and E: mRNA expression of mitochondrial and browning markers (Hadh, MT-CO2, Cox8b, Cpt1b, Ppargc1α, Pparα, Cidea, Ucp1) in BAT (D) and iWAT (F) (n = 4–7 per group). (E) mitochondrial COX activity in whole tissue BAT lysates from control and iβ1β2AKO mice treated with FGF21 (n = 4–7 per group). G–H: representative UCP1 and β-tubulin immunoblotting (G) with quantification in BAT (H) and iWAT (I) of high fat diet-fed and two week treated (saline or native FGF21) control and iβ1β2AKO mice (n = 4–7 per group). Data are means ± SEM with ††† p < 0.001, †† p < 0.01, † p < 0.05 denoting a general treatment effect and * p < 0.05 denoting a general genotype effect as determined by a two-way ANOVA.
3.4. FGF21-mediated improvements in glucose homeostasis, insulin sensitivity and hepatic lipid content are independent of phosphorylation of ACC by AMPK
The accumulation of hepatic lipids is thought to be critical in liver insulin resistance and causative in the dysregulation of whole-body glucose homeostasis [39]. One of the molecular mechanisms for hepatic lipid accumulation is the process of de novo lipogenesis. FGF21 has been shown to suppress hepatic glucose output [28] and de novo lipogenesis, effects that were associated with the activation of AMPK [2]. To examine whether the metabolic improvements resulting from FGF21 administration are due to AMPK's inhibitory action on ACC – a key rate-limiting enzyme in the lipogenesis pathway and regulator of hepatic fat oxidation – we utilized a previously described knock-in mutation mouse model for ACC where AMPK lacks the ability to phosphorylate and inactivate ACC [31]. In addition, we utilized a long-acting form of FGF21 (PF-05231023) that demonstrated efficacy in humans [6]. Consistent with previous studies [2], [5] and the current data in iβ1β2AKO mice, FGF21 significantly improved glucose tolerance (Figure 6A and B) and insulin sensitivity (Figure 6C and D). However, these effects were independent of the ability of AMPK to phosphorylate and inactivate ACC. Additionally, FGF21 administration led to reduced body weight (Figure 6E), adiposity (Figure 6F), and lowered fed (Figure 6G) and fasted-state (Figure 6H) blood glucose levels, irrespective of genotype. Lastly, the administration of long-acting FGF21 substantially lowered hepatic lipid content, marked by the reduced lipid containing area within H&E stained liver sections (Figure 6I) and reduced liver triacylglycerol levels (Figure 6J).
Figure 6.

Long-acting (PF-05231023) FGF21-mediated improvements in insulin sensitivity and hepatic lipid content are not due to AMPK's suppression of ACC. A–D: glucose tolerance test (GTT; 1 g/kg) (A) with area under the curve (AUC) (B) and insulin tolerance test (ITT; 1 U/kg) (C) with area under the curve (D) of wildtype and ACC DKI fed a 45% high fat diet for 10 weeks and treated with saline or long-acting FGF21 for two weeks (n = 4 per group). Change in body weights (E) and in fat percent (F) in wildtype and ACC DKI mice treated with saline or PF-05231023 (n = 8 per group). Blood glucose levels in the fed (G) and 12-hr fasted (H) state of wildtype and ACC DKI mice treated with saline or long-acting FGF21 for two weeks (n = 8 per group). I and J: representative H&E liver stains (20x) (I) and levels of triacylglycerol (J) in the liver of wildtype and ACC DKI mice treated with saline or long-acting FGF21 for 2 weeks (n = 8 per group). Data are means ± SEM with †††† p < 0.0001, ††† p < 0.001, †† p < 0.01, † p < 0.05 denoting a general treatment effect and ** p < 0.01 denoting a general genotype effect as determined by a two-way ANOVA (A and C: two-way repeated measures ANOVA).
4. Discussion
In the current study, we examined whether pharmacological administration of FGF21 requires adipocyte AMPK to mediate its beneficial effects on metabolic homeostasis. Our previous work demonstrated that adipocyte AMPK was necessary to maintain BAT function, mediate increases in WAT browning and protect against the development of hepatic steatosis in rodents on a HFD [23]. Our current results demonstrate that FGF21 administration does not require adipocyte AMPK to mediate improvements in glucose homeostasis, lipid homeostasis, insulin sensitivity, and reductions in body weight in vivo. Furthermore, we tested whether AMPK's inhibitory phosphorylation of ACC mediated the metabolic benefits of FGF21 administration and showed no genotype differences in the treated groups.
In mice, FGF21-mediated improvements in metabolic homeostasis have also been associated with the browning of WAT [2], [14]. We found that FGF21 treatment increased energy expenditure, reduced body fat mass, and improved insulin sensitivity without increases in BAT or iWAT UCP1 protein levels. Indeed, as previously reported [23], we found that BAT UCP1 levels were lower in mice deficient for adipocyte AMPK, yet FGF21 improved metabolic homeostasis in these mice. FGF21 increased mitochondrial COX activity in both genotypes, which may explain how FGF21 might be mediating its effects independently of adipocyte AMPK, possibly via UCP1-indepdendent mechanisms (see below). We did not observe changes in overall WAT browning, as measured by gene expression analyses and UCP1 protein levels, although FGF21 treatment did increase Ppargc1α mRNA levels, which could still be involved in mediating the effects of FGF21 [18], [40] in this model.
We found that FGF21 increased glucose uptake in BAT and lowered uptake in the liver and quadriceps muscle, effects which were independent of adipocyte AMPK. Since we did not perform the hyperinsulinemic–euglycemic clamp technique, we cannot rule out the potential effects of FGF21 on suppressing hepatic glucose output [2], [28]. FGF21 also increased tissue insulin action in BAT and iWAT independently of adipocyte AMPK; however, FGF21 did not significantly increase tissue glucose uptake in iWAT. These data suggest that the target tissue for the glucose lowering effects of FGF21 is likely BAT, although this does not require increased BAT UCP1 protein levels or AMPK. These data could explain the lack of glucose lowering in humans since the subjects recruited for the clinical studies were not measured for the presence of functional BAT. An alternate pathway is that FGF21 might promote UCP1-independent thermogenic mechanisms to mediate some of its beneficial metabolic effects. Recently, a phospho-proteomic approach to identify the signaling pathways activated downstream of FGF21 in adipocytes uncovered the metabolic sensor mammalian target of rapamycin (mTORC1) as an important mediator of the effects of FGF21 in vitro [41]. mTORC1 was required for FGF21-mediated increase in UCP1 mRNA, adiponectin secretion, and glucose uptake [41]. Future work should determine whether FGF21 requires adipocyte mTORC1 to mediate improvements in whole-body metabolism.
FGF21 likely improves hepatic steatosis and liver insulin action by reducing lipogenesis and enhancing lipid oxidation [2], [27], [29], [30]. As metabolic improvements upon FGF21 administration are associated with reduced expression of ACC1 and ACC2 in liver [27], we used previously described knock-in mutation mice (ACC DKI) to test whether AMPK's inhibitory phosphorylation of ACC (ACC1-S79 and ACC2-S212) mediated the metabolic benefits of FGF21 administration. When ACC is phosphorylated by AMPK, as occurs with metformin treatment, malonyl-CoA levels are reduced, leading to the suppression of de novo lipogenesis and the reduction of hepatic lipid content and insulin resistance [31], [42]. However, we observed no differences between wildtype and ACC DKI mice in response to FGF21 administration over two weeks. These data indicate that FGF21 does not share a similar mechanism to metformin for the amelioration of lipid content and insulin sensitivity [31].
In conclusion, we report that FGF21 improves metabolic homeostasis in HFD-fed mice, effects that are independent of adipocyte AMPK or its downstream substrate ACC, and which were not associated with increases in BAT or WAT browning. These data suggest that combinatorial treatment of obesity and diabetes with FGF21 and AMPK activators [31], [43] may have enhanced efficacy since they function through different signaling pathways. Future work is required to determine whether FGF21 mediates its effects via UCP1-independent futile cycling pathways, such as greater lipid turnover [44] or a creatine substrate cycle [45].
Author contributions
E.P.M., E.M.D., A.M.F., S.T., and G.R.S. designed the experiments. E.P.M., E.M.D., A.M.F., V.Z.Z., J.D.C., J.M.Y., D.M.E., and A.L. performed the experiments and testing. E.P.M., E.M.D., A.M.F., V.Z.Z., J.D.C., B.K., J.G.G., D.M.E., A.L., and S.T. provided technical expertise and analyzed data. E.P.M., E.M.D., and G.R.S. wrote the manuscript. All authors edited the manuscript and provided comments.
Acknowledgments
We thank Dr. Thomas J. Hawke for allowing access to the microscope. We thank Dr. Bruce E. Kemp for providing the β1/β2 floxed mice and the ACC1/2 DKI mice. We thank Pfizer Inc. for providing native FGF21 and PF-05231023. D.M.E., A.L. and S.T. were employees of Pfizer Inc. at the time of the study. This study was supported by a grant from the Canadian Diabetes Association (G.R.S.) (OG-3-13-4103-GS). E.P.M. was a Canadian Diabetes Association Post-doctoral research fellow. E.M.D. was a recipient of an Ontario Graduate Scholarship and a Queen Elizabeth II Graduate Scholarship in Science and Technology. A.M.F. was funded by the Danish Diabetes Academy, supported by the Novo Nordisk Foundation. G.R.S. is a Canada Research Chair in Metabolism and Obesity and the J. Bruce Duncan Chair in Metabolic Diseases.
Footnotes
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.molmet.2017.04.001.
Conflict of interest
The authors declare no competing financial interests.
Appendix A. Supplementary data
The following is the supplementary data related to this article:
Lack of ACC phosphorylation in BAT and iWAT of mice deficient for adipocyte AMPK. The efficiency of AMPK deletion was evaluated by immunoblotting BAT and iWAT protein samples from control (CreERT2 −) and mice deficient for adipocyte AMPK (CreERT2 +) for the AMPK consensus site phospho-ACC S79. Blots were stripped and also probed for total ACC and β-tubulin.
References
- 1.Kharitonenkov A., Shiyanova T.L., Koester A., Ford A.M., Micanovic R., Galbreath E.J. FGF-21 as a novel metabolic regulator. The Journal of Clinical Investigation. 2005;115(6):1627–1635. doi: 10.1172/JCI23606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bernardo B., Lu M., Bandyopadhyay G., Li P., Zhou Y., Huang J. FGF21 does not require interscapular brown adipose tissue and improves liver metabolic profile in animal models of obesity and insulin-resistance. Scientific Reports. 2015;5:11382. doi: 10.1038/srep11382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Holland W.L., Adams A.C., Brozinick J.T., Bui H.H., Miyauchi Y., Kusminski C.M. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell Metabolism. 2013;17(5):790–797. doi: 10.1016/j.cmet.2013.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kharitonenkov A., DiMarchi R. Fibroblast growth factor 21 night watch: advances and uncertainties in the field. Journal of Internal Medicine. 2016 doi: 10.1111/joim.12580. [DOI] [PubMed] [Google Scholar]
- 5.Wu A.-L., Kolumam G., Stawicki S., Chen Y., Li J., Zavala-Solorio J. Amelioration of type 2 diabetes by antibody-mediated activation of fibroblast growth factor receptor 1. Science Translational Medicine. 2011;3(113) doi: 10.1126/scitranslmed.3002669. 113ra126. [DOI] [PubMed] [Google Scholar]
- 6.Talukdar S., Zhou Y., Li D., Rossulek M., Dong J., Somayaji V. A long-acting FGF21 molecule, PF-05231023, decreases body weight and improves lipid profile in non-human primates and type 2 diabetic subjects. Cell Metabolism. 2016;23(3):427–440. doi: 10.1016/j.cmet.2016.02.001. [DOI] [PubMed] [Google Scholar]
- 7.Veniant M.M., Komorowski R., Chen P., Stanislaus S., Winters K., Hager T. Long-acting FGF21 has enhanced efficacy in diet-induced obese mice and in obese rhesus monkeys. Endocrinology. 2012;153(9):4192–4203. doi: 10.1210/en.2012-1211. [DOI] [PubMed] [Google Scholar]
- 8.Kharitonenkov A., Wroblewski V.J., Koester A., Chen Y.-F., Clutinger C.K., Tigno X.T. The metabolic state of diabetic monkeys is regulated by fibroblast growth factor-21. Endocrinology. 2007;148(2):774–781. doi: 10.1210/en.2006-1168. [DOI] [PubMed] [Google Scholar]
- 9.Gaich G., Chien J.Y., Fu H., Glass L.C., Deeg M.A., Holland W.L. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metabolism. 2013;18(3):333–340. doi: 10.1016/j.cmet.2013.08.005. [DOI] [PubMed] [Google Scholar]
- 10.Schlein C., Talukdar S., Heine M., Fischer A.W., Krott L.M., Nilsson S.K. FGF21 lowers plasma triglycerides by accelerating lipoprotein catabolism in white and brown adipose tissues. Cell Metabolism. 2016;23(3):441–453. doi: 10.1016/j.cmet.2016.01.006. [DOI] [PubMed] [Google Scholar]
- 11.Adams A.C., Cheng C.C., Coskun T., Kharitonenkov A. FGF21 requires βklotho to act in vivo. PLoS One. 2012;7(11):e49977. doi: 10.1371/journal.pone.0049977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Adams A.C., Yang C., Coskun T., Cheng C.C., Gimeno R.E., Luo Y. The breadth of FGF21's metabolic actions are governed by FGFR1 in adipose tissue. Molecular Metabolism. 2013;2(1):31–37. doi: 10.1016/j.molmet.2012.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Samms R.J., Smith D.P., Cheng C.C., Antonellis P.P., Perfield J.W., Kharitonenkov A. Discrete aspects of FGF21 in vivo pharmacology do not require UCP1. Cell Reports. 2015;11(7):991–999. doi: 10.1016/j.celrep.2015.04.046. [DOI] [PubMed] [Google Scholar]
- 14.Véniant M.M., Sivits G., Helmering J., Komorowski R., Lee J., Fan W. Pharmacologic effects of FGF21 are independent of the “Browning” of white adipose tissue. Cell Metabolism. 2015;21(5):731–738. doi: 10.1016/j.cmet.2015.04.019. [DOI] [PubMed] [Google Scholar]
- 15.Kwon M.M., O'Dwyer S.M., Baker R.K., Covey S.D., Kieffer T.J. FGF21-Mediated improvements in glucose clearance require uncoupling protein 1. Cell Reports. 2015;13(8):1521–1527. doi: 10.1016/j.celrep.2015.10.021. [DOI] [PubMed] [Google Scholar]
- 16.Straub L., Wolfrum C. FGF21, energy expenditure and weight loss – how much brown fat do you need? Molecular Metabolism. 2015;4(9):605–609. doi: 10.1016/j.molmet.2015.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bookout A.L., de Groot M.H.M., Owen B.M., Lee S., Gautron L., Lawrence H.L. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nature Medicine. 2013;19(9):1147–1152. doi: 10.1038/nm.3249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chau M.D.L., Gao J., Yang Q., Wu Z., Gromada J. Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK–SIRT1–PGC-1α pathway. Proceedings of the National Academy of Sciences. 2010;107(28):12553–12558. doi: 10.1073/pnas.1006962107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Steinberg G.R., Kemp B.E. AMPK in health and disease. Physiological Reviews. 2009;89(3):1025–1078. doi: 10.1152/physrev.00011.2008. [DOI] [PubMed] [Google Scholar]
- 20.Schaffer B.E., Levin R.S., Hertz N.T., Maures T.J., Schoof M.L., Hollstein P.E. Identification of AMPK phosphorylation sites reveals a network of proteins involved in cell invasion and facilitates large-scale substrate prediction. Cell Metabolism. 2015;22(5):907–921. doi: 10.1016/j.cmet.2015.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ducommun S., Deak M., Sumpton D., Ford R.J., Núñez Galindo A., Kussmann M. Motif affinity and mass spectrometry proteomic approach for the discovery of cellular AMPK targets: identification of mitochondrial fission factor as a new AMPK substrate. Cellular Signalling. 2015;27(5):978–988. doi: 10.1016/j.cellsig.2015.02.008. [DOI] [PubMed] [Google Scholar]
- 22.Hoffman N.J., Parker B.L., Chaudhuri R., Fisher-Wellman K.H., Kleinert M., Humphrey S.J. Global phosphoproteomic analysis of human skeletal muscle reveals a network of exercise-regulated kinases and AMPK substrates. Cell Metabolism. 2015;22(5):922–935. doi: 10.1016/j.cmet.2015.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mottillo E.P., Desjardins E.M., Crane J.D., Smith B.K., Green A.E., Ducommun S. Lack of adipocyte AMPK exacerbates insulin resistance and hepatic steatosis through brown and beige adipose tissue function. Cell Metabolism. 2016;24(1):118–129. doi: 10.1016/j.cmet.2016.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gauthier M.-S., Miyoshi H., Souza S.C., Cacicedo J.M., Saha A.K., Greenberg A.S. AMP-activated protein kinase is activated as a consequence of lipolysis in the adipocyte potential mechanism and physiological relevance. Journal of Biological Chemistry. 2008;283(24):16514–16524. doi: 10.1074/jbc.M708177200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mulligan J.D., Gonzalez A.A., Stewart A.M., Carey H.V., Saupe K.W. Upregulation of AMPK during cold exposure occurs via distinct mechanisms in brown and white adipose tissue of the mouse: AMPK in brown and white adipose tissue. The Journal of Physiology. 2007;580(2):677–684. doi: 10.1113/jphysiol.2007.128652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Camporez J.P.G., Jornayvaz F.R., Petersen M.C., Pesta D., Guigni B.A., Serr J. Cellular mechanisms by which FGF21 improves insulin sensitivity in male mice. Endocrinology. 2013;154(9):3099–3109. doi: 10.1210/en.2013-1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Coskun T., Bina H.A., Schneider M.A., Dunbar J.D., Hu C.C., Chen Y. Fibroblast growth factor 21 corrects obesity in mice. Endocrinology. 2008;149(12):6018–6027. doi: 10.1210/en.2008-0816. [DOI] [PubMed] [Google Scholar]
- 28.Berglund E.D., Li C.Y., Bina H.A., Lynes S.E., Michael M.D., Shanafelt A.B. Fibroblast growth factor 21 controls glycemia via regulation of hepatic glucose flux and insulin sensitivity. Endocrinology. 2009;150(9):4084–4093. doi: 10.1210/en.2009-0221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Emanuelli B., Vienberg S.G., Smyth G., Cheng C., Stanford K.I., Arumugam M. Interplay between FGF21 and insulin action in the liver regulates metabolism. Journal of Clinical Investigation. 2014;124(2):515–527. doi: 10.1172/JCI67353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fisher F.M., Chui P.C., Antonellis P.J., Bina H.A., Kharitonenkov A., Flier J.S. Obesity is a fibroblast growth factor 21 (FGF21)-resistant state. Diabetes. 2010;59(11):2781–2789. doi: 10.2337/db10-0193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fullerton M.D., Galic S., Marcinko K., Sikkema S., Pulinilkunnil T., Chen Z.-P. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nature Medicine. 2013;19(12):1649–1654. doi: 10.1038/nm.3372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Owen B.M., Ding X., Morgan D.A., Coate K.C., Bookout A.L., Rahmouni K. FGF21 acts centrally to induce sympathetic nerve activity, energy expenditure, and weight loss. Cell Metabolism. 2014;20(4):670–677. doi: 10.1016/j.cmet.2014.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Veniant M.M., Hale C., Helmering J., Chen M.M., Stanislaus S., Busby J. FGF21 promotes metabolic homeostasis via white adipose and leptin in mice. PloS One. 2012;7(7):e40164. doi: 10.1371/journal.pone.0040164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Steinberg G.R., O'Neill H.M., Dzamko N.L., Galic S., Naim T., Koopman R. Whole body deletion of AMP-activated protein kinase 2 reduces muscle AMPK activity and exercise capacity. Journal of Biological Chemistry. 2010;285(48):37198–37209. doi: 10.1074/jbc.M110.102434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Henriksbo B.D., Lau T.C., Cavallari J.F., Denou E., Chi W., Lally J.S. Fluvastatin causes NLRP3 inflammasome-mediated adipose insulin resistance. Diabetes. 2014;63(11):3742–3747. doi: 10.2337/db13-1398. [DOI] [PubMed] [Google Scholar]
- 36.O'Neill H.M., Maarbjerg S.J., Crane J.D., Jeppesen J., Jorgensen S.B., Schertzer J.D. AMP-activated protein kinase (AMPK) 1 2 muscle null mice reveal an essential role for AMPK in maintaining mitochondrial content and glucose uptake during exercise. Proceedings of the National Academy of Sciences. 2011;108(38):16092–16097. doi: 10.1073/pnas.1105062108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lin Z., Tian H., Lam K.S.L., Lin S., Hoo R.C.L., Konishi M. Adiponectin mediates the metabolic effects of FGF21 on glucose homeostasis and insulin sensitivity in mice. Cell Metabolism. 2013;17(5):779–789. doi: 10.1016/j.cmet.2013.04.005. [DOI] [PubMed] [Google Scholar]
- 38.Yamauchi T., Kamon J., Minokoshi Y., Ito Y., Waki H., Uchida S. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nature Medicine. 2002;8(11):1288–1295. doi: 10.1038/nm788. [DOI] [PubMed] [Google Scholar]
- 39.Perry R.J., Samuel V.T., Petersen K.F., Shulman G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature. 2014;510(7503):84–91. doi: 10.1038/nature13478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fisher F.M., Kleiner S., Douris N., Fox E.C., Mepani R.J., Verdeguer F. FGF21 regulates PGC-1 and browning of white adipose tissues in adaptive thermogenesis. Genes & Development. 2012;26(3):271–281. doi: 10.1101/gad.177857.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Minard A.Y., Tan S.-X., Yang P., Fazakerley D.J., Domanova W., Parker B.L. mTORC1 is a major regulatory node in the FGF21 signaling network in adipocytes. Cell Reports. 2016;17(1):29–36. doi: 10.1016/j.celrep.2016.08.086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Smith B.K., Marcinko K., Desjardins E.M., Lally J.S., Ford R.J., Steinberg G.R. Treatment of nonalcoholic fatty liver disease: role of AMPK. American Journal of Physiology, Endocrinology and Metabolism. 2016;311(4):E730–E740. doi: 10.1152/ajpendo.00225.2016. [DOI] [PubMed] [Google Scholar]
- 43.Hawley S.A., Fullerton M.D., Ross F.A., Schertzer J.D., Chevtzoff C., Walker K.J. The ancient drug salicylate directly activates AMP-activated protein kinase. Science. 2012;336(6083):918–922. doi: 10.1126/science.1215327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mottillo E.P., Balasubramanian P., Lee Y.-H., Weng C., Kershaw E.E., Granneman J.G. Coupling of lipolysis and de novo lipogenesis in brown, beige, and white adipose tissues during chronic 3-adrenergic receptor activation. The Journal of Lipid Research. 2014;55(11):2276–2286. doi: 10.1194/jlr.M050005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kazak L., Chouchani E.T., Jedrychowski M.P., Erickson B.K., Shinoda K., Cohen P. A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell. 2015;163(3):643–655. doi: 10.1016/j.cell.2015.09.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Lack of ACC phosphorylation in BAT and iWAT of mice deficient for adipocyte AMPK. The efficiency of AMPK deletion was evaluated by immunoblotting BAT and iWAT protein samples from control (CreERT2 −) and mice deficient for adipocyte AMPK (CreERT2 +) for the AMPK consensus site phospho-ACC S79. Blots were stripped and also probed for total ACC and β-tubulin.