

Abstract
While many decades of scientific research studies have gone into harnessing the power of the immune system to fight cancer, only recently have cancer immunotherapeutic approaches begun to show robust clinical responses in patients with a variety of cancers. These treatments are adding to the current arsenal of cancer treatments; surgery, radiation and chemotherapy, and increasing the therapeutic options for cancer patients. Despite these advances, issues associated with these therapies include that not all patients respond to these therapies, and some patients who respond experience varying degrees of toxicities. One of the major issues affecting immunotherapy is the inability to evaluate trafficking of activated T-cells into sites of tumor. The current diagnostic imaging based on conventional anatomic imaging, which is the mainstay to monitor response to cytotoxic chemotherapy or radiation, is not adequate to assess initial response to immunotherapy or disease evolution. Patients’ prognosis by histological analysis has limited use in regards to immunotherapy. Thus, there is a crucial need for noninvasive biomarkers for screening patients that show long term response to therapy. Here, we provide a brief account of emerging molecular magnetic resonance imaging biomarkers that have potential to exploit the metabolism and metabolic products of activated T cells.
How cancer cheats the immune system
Effective anti-cancer treatments are largely affected by the cross-talk between cancer and the patient’s immune system. Studies have demonstrated that tumors evade the host immune response via a number of mechanisms [1–3]. In down regulating the major histocompatibility complex I, cancer cells make the detection of the antigens on their surface by the immune system less effective [4, 5]. Further, they produce immunosuppressive cytokines, such as (TGF-β) and interleukin-10 (IL-10) that down regulate the cytotoxic immune cells and shift the immune response towards a suppressive phenotype [4, 6]. Finally, they upregulate surface proteins such as programmed cell death-ligand 1 (PD-L1), an important protein of normal cells in preventing autoimmune phenomena [6–8]. When the PD-1 receptor on cytotoxic T cells interacts with PD-L1, the T-cells become anergic and do not destroy them.
Immune system for cancer therapy
In the past few years, one of the most exciting advances in the treatment of tumors is boosting the body’s immune response against cancer [9]. There are different approaches to boost or restore immune function against cancer, which are broadly classified into four categories: immune check point blockade [8, 10], adoptive T-cell therapy [11, 12], exogenous cytokines [13, 14] and therapeutic vaccines [15, 16].
Check point blockade
Recent advances have demonstrated that blockade of immune checkpoints is one of the most promising approaches for activating therapeutic antitumor immunity [8]. Immune checkpoints are the receptor-ligand pairs on the cell surface that are involved in regulating T-cell activation.
It is now established that tumors utilize certain immune-checkpoint pathways as a mechanism of immune resistance against T cells that are specific for tumor antigens. Since many of the immune checkpoints involve ligand-receptor interactions, they can be readily blocked by antibodies or modulated by recombinant forms of ligands or receptors. Immunotherapeutics based on antibodies of cytotoxic T-lymphocyte-associated antigen 4 (CTLA4) [17, 18] and programmed cell death protein-1 (PDCD1/PD1) are showing promising results of antitumor immunity [19, 20]. In fact, the immunomodulatory monoclonal antibody of CTLA4, Ipilimumab, is the first Food and Drug Administration (FDA) approved immunotherapeutic agent for treating cancer [10, 21]. More recently, Nivolumab and Pembrolizumab, humanized IgG4 antibodies, which block PD-1 and inhibits its interaction with PD-L1 and PD-L2 have also been approved as immunotherapeutic agents for treatment of cancer by the US FDA [22–24].
Adoptive T-cell therapies
Adoptive T-cell therapies include expanded autologous T cells and T cells with engineered T-cell receptors (TCRs) and chimeric antigen receptors (CARs) [25, 26]. Specifically, tumor-infiltrating lymphocytes (TILs) are isolated from tumor biopsies and expanded before being reinfused into the patient, based on the premise that these TILs are tumor cell specific. The most effective T-cell therapies explored in clinical trials currently focus on leukemia, but are also used to treat patients with solid tumors.
Cytokines
Cytokines play important roles in the body’s normal immune responses and also in the immune system’s ability to respond to cancer. Interferons and interleukins are two main types of cytokines used to treat cancer [13, 14]. These cytokines bind to their receptors on T cells, and stimulate the activation and proliferation of T cells and downstream production of more cytokines [13, 14].
Treatment vaccines
These vaccines stimulate an active immune response against tumor by eliciting adaptive immunity through the patient’s own immune system. After injection of peptide or protein vaccines, the body’s antigen presenting cells (APCs) process vaccines as antigenic fragments to be recognized and stimulate the patient’s naïve T cells, which in turn may stimulate an endogenous immune response against cancer [15, 16].
Problems
While these immunotherapy methods provide tremendous hope for patients, they also present significant challenges. Treatment with immunotherapies is showing new patterns of treatment response and side effects. Specifically, after immunotherapy the response can be manifested different ways: (1) a decrease in size of known tumors without the presence of new tumor after completion of treatment, (2) clinically stable disease after completion of treatment and significantly delayed decrease in tumor size, (3) new or enlarging tumors observed soon after completion of treatment, which may not reflect disease progression, preceding a later decrease in tumor burden (4) autoimmune-mediated toxic effects that could be mistaken for metastatic disease or misdiagnosed as a non-treatment-related process and delay appropriate clinical management [27].
Currently, there are no robust biomarkers to identify the patients who will most likely benefit from these treatments. In the absence of a predictive biomarker, many patients may receive these expensive treatments without any benefit. These unconventional treatment response patterns and the wide range of autoimmune toxic effects make it rather challenging to monitor the effects of immunotherapies using Response Evaluation Criteria in Solid Tumors (RECIST) [28] criteria, which are based on the conventional anatomical imaging by computed tomography and magnetic resonance imaging (MRI) [29]. Hence, there is need for robust technology, which not only characterizes the immune microenvironment of tumors but also screen for patients who can potentially respond to immunotherapies. Imaging methods targeting T cell metabolism have the potential for providing molecular imaging biomarkers to assess immunotherapy response. To develop molecular imaging biomarkers, understanding the T cell metabolism and its changes upon activation are crucial.
Regulation of T cell metabolism
Like all non-proliferating cells, naïve T cells (T cells that have not yet encountered antigen) adopt a basal level of nutrient uptake and primarily use oxidative phosphorylation (OXPHOS) for adenosine triphosphate (ATP) production. When the T cells encounter antigen (on tumors) they become activated and respond by extensive proliferation and differentiation into effector T cells (TEFF), which identify and eradicate pathogenic threats to the host systems. In the activated state, the TEFF cells switch to anabolic growth and biomass accumulation to generate daughter cells that increases the demand for ATP. To support their high energy demand, activated T cells shifts to aerobic glycolysis, which involves conversion of glucose derived pyruvate to lactate even in the presence of oxygen for glucose oxidation-also known as Warburg effect. Although both CD4+ and CD8+ TEFF cells still engage OXPHOS, they predominantly employ aerobic glycolysis [30, 31]. After clearing the pathogens, most TEFF cells die, and a small population of long-lived antigen-specific memory T cells (TM) are left behind. Like naïve cells, the TM cells engage OXPHOS and maintain lower rates of nutrient uptake and biosynthesis when compared to TEFF cells (Fig. 1) [32].
Fig. 1.
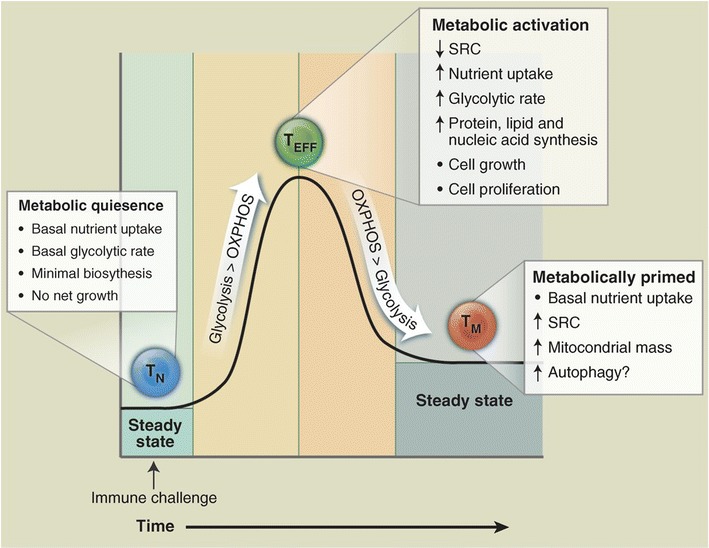
Showing the diagrammatic representation of metabolic switch in different stages of T cells. Naïve T cells (TN) are predominantly depending on the OXPHOS for the energy needs. The proliferative T cells (TEFF) shifts to aerobic glycolysis to support their high energy demand. The memory T cells (TM) use OXPHOS pathway for their metabolic requirement [32]. Reprinted with permission from AAAS
The above discussion implies that in the activated form T cells (TEFF) predominantly engage aerobic glycolysis, just like proliferating cancer cells. It is well known that predominant product of aerobic glycolysis is lactic acid generation. So, cancer cells in a tissue, as a result of aerobic glycolysis, produce lactate and maintain an acidic environment. We hypothesize that upon activation, the TEFF cells will also go through the glycolysis and produce significant amount of lactic acid and increases the overall lactate level in the cancer tissue. This increased lactate level may serve as a biomarker for T cell activation and engaging the cancer cells. In addition to lactate increase, TEFF cells will also show significant increase in Alanine (Fig. 2). Upon successful immunotherapy, both cancer cells as well as the TEFF cell decrease substantially and overall lactate levels will also decrease accordingly and normalize to basal levels over a period of time. So, rapid changes in glycolysis, amino acids and proteins in CAR TEFF cells, lactate production and pH changes potentially serve as molecular biomarkers for therapeutic response and disease progression.
Fig. 2.
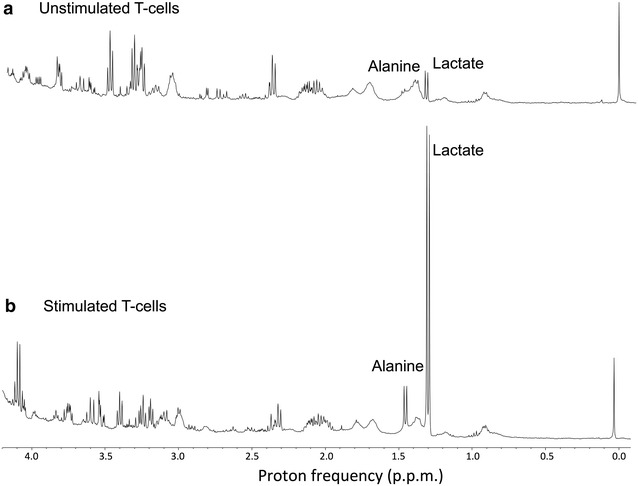
1H NMR spectra from cultured T cells obtained at 9.4 T. a Spectrum from previously non-stimulated resting T-cells shows lactate and other intra cellular metabolites, and very little alanine. b Spectrum from stimulated T cells with CD3/CD28 obtained under identical conditions of the spectra in a, which shows substantially (over 12-fold increase) higher lactate and alanine (3-fold increase) compared to that from non-stimulated resting T-cells (unpublished results from author’s laboratory)
Among the molecular imaging techniques, 18F-fluorodeoxyglucose (18F-FDG) positron emission tomography (PET) imaging of cancer is the most studied modality in oncologic nuclear imaging [33]. It is utilized primarily to assess tumor glycolysis among other things. However, primary challenges with 18F-FDG-PET include its inability to differentiate between cancer and infectious or inflammatory processes. Specifically, this becomes major shortcoming when evaluating response to therapy amid immune-related adverse events after treatment with immunotherapy agents. While 18F-fluorothymidine, a marker of cell proliferation, which was developed to identify viable tumor, it is beset by lower signal to background ratio compared with 18F-FDG-PET and uptake in background structures, accumulation in sites of infection and inflammation can limit detection and quantification of tumor activity [34, 35]. Furthermore, frequent imaging with 18F-FDG-PET is not feasible as it involves radiation.
MRI can provide high resolution anatomical imaging along with an array of functional measures: tumor perfusion, diffusion, cell membrane permeability via contrast enhanced MRI, immune cell tracking using magnetic iron oxide particles, etc. However, for the reasons mentioned above, these measures are not adequate for assessing response to immunotherapy.
Chemical exchange saturation transfer (CEST) MRI of T cell metabolites
One of the challenges in the diagnosis of response to immunotherapy is distinguishing between new tumor and inflammation or edema. CEST methods potentially address this issue. Recent developments in CEST methods show that it is feasible to image metabolites such as glutamate [36, 37], creatine [38], glucose [39], glycogen [40], myoinositol [41], lactate [42] and glycosaminoglycans [43]. In the activated state, the T cells go through the glycolysis to support rapid energy required for biosynthesis of daughter cells. This leads to accumulation of metabolites such as lactate, creatine, choline, glutamate and alanine in TEFF cells. Monitoring changes in these metabolites level pre- and post immune therapy has potential to assess the relative changes in the TEFF cell density.
CEST MRI of lactate: a biomarker for immunotherapy
Currently there are two major methods employed in measuring lactate in vivo. One is traditional magnetic resonance spectroscopy (MRS; both 1H and 13C) [44–46], which has been used to measure both static lactate levels and dynamic changes. However, these are limited by inadequate sensitivity and spatial resolution. The other method involves infusion of dynamic nuclear polarized (DNP)13C-labeled pyruvate, which provides greater than 10,000-fold signal enhancement compared to conventional MRS [47–49]. Despite its high sensitivity, this method only probes fast kinetics (<1 min) of lactate turnover from 13C-labeled pyruvate and it requires special equipment and complex modeling for data analysis.
Recently, MRI method based on lactate CEST (LATEST) [42] to image lactate was described. LATEST method utilizes standard proton MRI and requires neither 13C labeled pyruvate nor DNP polarization. The feasibility of measuring LATEST in vivo was demonstrated in a lymphoma tumor model (Fig. 3), and in human skeletal muscle [42]. Dynamic changes in LATEST are reported in tumors pre- and post-infusion of pyruvate, and in exercising human skeletal muscle [42]. LATEST measurements are compared to lactate measured with multiple quantum filtered proton MRS [42]. LATEST provides over two orders of higher sensitivity compared to the 1H MRS based lactate detection methods.
Fig. 3.
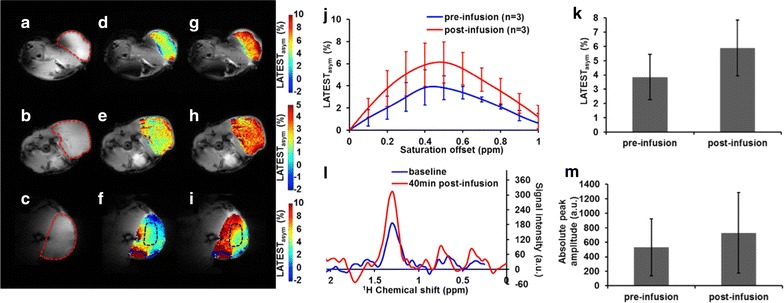
LATEST from lymphoma tumors [42]. a–c Anatomical image from three animals, with flank tumor region indicated by dotted red line, and the LATEST maps (d–f) pre-infusion and (g–i) post-infusion with (j) corresponding asymmetry plots (asymmetry from Animal 3 in the third row is taken from region indicated in dotted black line), (k) LATEST change at 0.4 ppm from three animals pre- and post-infusion, (l) representative SEL-MQC1H-MRS pre- and 40 min post-infusion from flank tumor showing (m) increase in lactate peak amplitude from three animals (~40%) from spectroscopy. Reproduced with permission
It was reported that lactic acid produced by the tumor cells blunts the tumor immunosurveillance by T and natural killer cells. This implies that the higher levels of lactate in tumor cells may adversely affect the immunotherapy and basal levels of lactate itself might give a clue regarding the response to immunotherapy [50]. In the context of immunotherapy, pre-therapy LATEST images provide the basal levels of lactate in tumor regions which are largely glycolytic and produce more lactate. Immediately, post-therapy (12–24 h), if the immune cells have identified receptors on the tumors cell surface and get activated then the TEFF cells switch their metabolism to glycolysis and begin to proliferate rapidly and dump lots of lactate into the tumor microenvironment (at this time tumor cells may be still be producing lactate, although with a different/slower rate). This rapid increase in lactate can be measured using LATEST. In addition to this rapid increase in LATEST, T cells activation may also lead to side effects associated with autoimmunity. As the T cell rapidly proliferates, it may lead to an increase in the size of the T cells mass in the tumor region, which is often mistaken for tumor growth. This elevation in the lactate levels remains until the T cells completely destroy the tumor cells and then levels begin to drop, as the TEFF cells die and convert to TM cells, to basal values.
On the other hand, if the immune cells do not get activated then their metabolism remains OXPHOS and there would not be any change in the lactate levels due to immune cells and as the tumor cells are continuously proliferating, lactate levels and tumor size increase gradually. So the kinetics of the lactate measured shed light on the therapeutic efficacy.
The slopes of the lactate concentration vs. time curves, especially hours after the treatment, will serve as a measure of the response. Response to therapy is expected to produce a steeper slope in the curve than no response.
CEST MRI of glutamate, alanine and creatine
Higher concentration of glutamate, alanine and creatine during the T cell proliferation in response to immunotherapy can also be monitored using CEST. Studies have shown that the changes in these metabolites level in cancer tissue can be monitored non-invasively through CEST. Different CEST based approaches (GluCEST, glutamate; AlaCEST, alanine; CrCEST, creatine) have been developed to image these metabolites in vivo. In addition, another CEST method, amide proton transfer (APT), which primarily depends on the mobile protein content, has been shown to be useful in discriminating between tumor regrowth and radiation necrosis [51]. It has been shown that the glutamate released by the dendritic cells mediates the T cell activation/proliferation [52]. Higher expression of glutamate metabotropic receptor on activated T cells further confirms the role of the glutamate in T cells mediated immunity [52]. The increase in alanine concentration in an in vitro stimulated T cell line (Fig. 2) suggests that the activation of T cells result in more alanine synthesis. Changes in the in vivo glutamate, alanine and creatine level post-immunotherapy as measured by CEST may also serve as potential biomarkers to evaluate the treatment response.
Conclusion
Overall, immediately after the immunotherapy administration, rapid increase in lactate (derived via LATEST) and glutamate, alanine, and creatine from tumor regions is indicative of response to immunotherapy. Successful therapy eventually will be manifested in favorable clinical symptoms as well as low values of LATEST and CrCEST (or GluCEST/AlaCEST) compared to post-treatment. If during post treatment phase inflammation occurs or edema builds up then neither LATEST nor CrCEST (or GluCEST/AlaCEST) would increase. Thus compared to pre-treatment, unchanged or small changes in LATEST and CrCEST (or GluCEST/AlaCEST) values, in tumor region post treatment, may point to unresponsiveness. The CEST MRI methods potentially provide an early biomarker to monitor the immunotherapy response in vivo and to evaluate the patients who will respond to immunotherapy.
Authors’ contributions
MH collected literatures and helped with manuscript writing. PB, HH, BM, LJ discussed the previous findings and helped with manuscript editing. RR conceived of and designed the study and wrote the manuscripts. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Please contact author for data requests.
Funding
This work was supported by the NIBIB under Grant No. P41 EB015893.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- MRI
magnetic resonance imaging
- PD-L1
programmed cell death-ligand 1
- CTLA4
cytotoxic T-lymphocyte-associated antigen 4
- PDCD1/PD1
programmed cell death protein-1
- FDA
Food and Drug Administration
- TCRs
engineered T cell receptors
- CARs
chimeric antigen receptors
- TILs
tumor-infiltrating lymphocytes
- APCs
antigen presenting cells
- RECIST
Response Evaluation Criteria in Solid Tumors
- ATP
adenosine triphosphate
- TEFF
effector T cells
- OXPHOS
oxidative phosphorylation
- TM
memory T cells
- 18F-FDG
18F-fluorodeoxyglucose
- PET
positron emission tomography
- CEST
chemical exchange saturation transfer
- DNP
dynamic nuclear polarization
- MRS
magnetic resonance spectroscopy
- LATEST
lactate CEST
- APT
amide proton transfer
Contributor Information
Mohammad Haris, Email: mharis@sidra.org.
Puneet Bagga, Email: puneetb@mail.med.upenn.edu.
Hari Hariharan, Email: harih@mail.med.upenn.edu.
Bevin McGettigan-Croce, Email: bevinmc@mail.med.upenn.edu.
Laura A. Johnson, Email: ljohnso@upenn.edu
Ravinder Reddy, Phone: (215) 898-9357, Email: krr@upenn.edu.
References
- 1.Muenst S, Laubli H, Soysal SD, Zippelius A, Tzankov A, Hoeller S. The immune system and cancer evasion strategies: therapeutic concepts. J Intern Med. 2016;279:541–562. doi: 10.1111/joim.12470. [DOI] [PubMed] [Google Scholar]
- 2.Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E, Lichtor T, Decker WK, Whelan RL, Kumara HM, et al. Immune evasion in cancer: mechanistic basis and therapeutic strategies. Semin Cancer Biol. 2015;35(Suppl):S185–S198. doi: 10.1016/j.semcancer.2015.03.004. [DOI] [PubMed] [Google Scholar]
- 3.Mapara MY, Sykes M. Tolerance and cancer: mechanisms of tumor evasion and strategies for breaking tolerance. J Clin Oncol. 2004;22:1136–1151. doi: 10.1200/JCO.2004.10.041. [DOI] [PubMed] [Google Scholar]
- 4.Kim R, Emi M, Tanabe K, Arihiro K. Tumor-driven evolution of immunosuppressive networks during malignant progression. Cancer Res. 2006;66:5527–5536. doi: 10.1158/0008-5472.CAN-05-4128. [DOI] [PubMed] [Google Scholar]
- 5.Fruci D, Benevolo M, Cifaldi L, Lorenzi S, Lo Monaco E, Tremante E, Giacomini P. Major histocompatibility complex class i and tumour immuno-evasion: how to fool T cells and natural killer cells at one time. Curr Oncol. 2012;19:39–41. doi: 10.3747/co.19.945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rabinovich GA, Gabrilovich D, Sotomayor EM. Immunosuppressive strategies that are mediated by tumor cells. Annu Rev Immunol. 2007;25:267–296. doi: 10.1146/annurev.immunol.25.022106.141609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Iwai Y, Ishida M, Tanaka Y, Okazaki T, Honjo T, Minato N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc Natl Acad Sci USA. 2002;99:12293–12297. doi: 10.1073/pnas.192461099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12:252–264. doi: 10.1038/nrc3239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Couzin-Frankel J. Breakthrough of the year 2013. Cancer immunotherapy. Science. 2013;342:1432–1433. doi: 10.1126/science.342.6165.1432. [DOI] [PubMed] [Google Scholar]
- 10.Sharma P, Wagner K, Wolchok JD, Allison JP. Novel cancer immunotherapy agents with survival benefit: recent successes and next steps. Nat Rev Cancer. 2011;11:805–812. doi: 10.1038/nrc3153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.June CH. Adoptive T cell therapy for cancer in the clinic. J Clin Investig. 2007;117:1466–1476. doi: 10.1172/JCI32446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Houot R, Schultz LM, Marabelle A, Kohrt H. T-cell-based Immunotherapy: adoptive cell transfer and checkpoint inhibition. Cancer Immunol Res. 2015;3:1115–1122. doi: 10.1158/2326-6066.CIR-15-0190. [DOI] [PubMed] [Google Scholar]
- 13.Lee S, Margolin K. Cytokines in cancer immunotherapy. Cancers (Basel) 2011;3:3856–3893. doi: 10.3390/cancers3043856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ardolino M, Hsu J, Raulet DH. Cytokine treatment in cancer immunotherapy. Oncotarget. 2015;6:19346–19347. doi: 10.18632/oncotarget.5095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Finn OJ. Cancer vaccines: between the idea and the reality. Nat Rev Immunol. 2003;3:630–641. doi: 10.1038/nri1150. [DOI] [PubMed] [Google Scholar]
- 16.Banchereau J, Palucka AK. Dendritic cells as therapeutic vaccines against cancer. Nat Rev Immunol. 2005;5:296–306. doi: 10.1038/nri1592. [DOI] [PubMed] [Google Scholar]
- 17.Callahan MK, Wolchok JD, Allison JP. Anti-CTLA-4 antibody therapy: immune monitoring during clinical development of a novel immunotherapy. Semin Oncol. 2010;37:473–484. doi: 10.1053/j.seminoncol.2010.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Peggs KS, Quezada SA, Chambers CA, Korman AJ, Allison JP. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti-CTLA-4 antibodies. J Exp Med. 2009;206:1717–1725. doi: 10.1084/jem.20082492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ, Topalian SL, Hwu P, Drake CG, Camacho LH, Kauh J, Odunsi K, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012;366:2455–2465. doi: 10.1056/NEJMoa1200694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD, Sosman JA, Atkins MB, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012;366:2443–2454. doi: 10.1056/NEJMoa1200690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010;363:711–723. doi: 10.1056/NEJMoa1003466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kyi C, Postow MA. Checkpoint blocking antibodies in cancer immunotherapy. FEBS Lett. 2014;588:368–376. doi: 10.1016/j.febslet.2013.10.015. [DOI] [PubMed] [Google Scholar]
- 23.Weber JS, D’Angelo SP, Minor D, Hodi FS, Gutzmer R, Neyns B, Hoeller C, Khushalani NI, Miller WH, Jr, Lao CD, et al. Nivolumab versus chemotherapy in patients with advanced melanoma who progressed after anti-CTLA-4 treatment (CheckMate 037): a randomised, controlled, open-label, phase 3 trial. Lancet Oncol. 2015;16:375–384. doi: 10.1016/S1470-2045(15)70076-8. [DOI] [PubMed] [Google Scholar]
- 24.Larkin J, Lao CD, Urba WJ, McDermott DF, Horak C, Jiang J, Wolchok JD. Efficacy and safety of nivolumab in patients with BRAF V600 mutant and BRAF wild-type advanced melanoma: a pooled analysis of 4 clinical trials. JAMA Oncol. 2015;1:433–440. doi: 10.1001/jamaoncol.2015.1184. [DOI] [PubMed] [Google Scholar]
- 25.Dai H, Wang Y, Lu X, Han W. Chimeric antigen receptors modified T-cells for cancer therapy. J Natl Cancer Inst 2016, 108. doi: 10.1093/jnci/djv439. [DOI] [PMC free article] [PubMed]
- 26.Fesnak AD, June CH, Levine BL. Engineered T cells: the promise and challenges of cancer immunotherapy. Nat Rev Cancer. 2016;16:566–581. doi: 10.1038/nrc.2016.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hoos A. Evolution of end points for cancer immunotherapy trials. Ann Oncol. 2012;23(Suppl 8):viii47–viii52. doi: 10.1093/annonc/mds263. [DOI] [PubMed] [Google Scholar]
- 28.Eisenhauer EA, Therasse P, Bogaerts J, Schwartz LH, Sargent D, Ford R, Dancey J, Arbuck S, Gwyther S, Mooney M, et al. New response evaluation criteria in solid tumours: revised RECIST guideline (version 1.1) Eur J Cancer. 2009;45:228–247. doi: 10.1016/j.ejca.2008.10.026. [DOI] [PubMed] [Google Scholar]
- 29.Wang Q, Ornstein M, Kaufman HL. Imaging the immune response to monitor tumor immunotherapy. Expert Rev Vaccines. 2009;8:1427–1437. doi: 10.1586/erv.09.100. [DOI] [PubMed] [Google Scholar]
- 30.van der Windt GJ, Pearce EL. Metabolic switching and fuel choice during T-cell differentiation and memory development. Immunol Rev. 2012;249:27–42. doi: 10.1111/j.1600-065X.2012.01150.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chang CH, Pearce EL. Emerging concepts of T cell metabolism as a target of immunotherapy. Nat Immunol. 2016;17:364–368. doi: 10.1038/ni.3415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pearce EL, Poffenberger MC, Chang CH, Jones RG. Fueling immunity: insights into metabolism and lymphocyte function. Science. 2013;342:1242454. doi: 10.1126/science.1242454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhu A, Lee D, Shim H. Metabolic positron emission tomography imaging in cancer detection and therapy response. Semin Oncol. 2011;38:55–69. doi: 10.1053/j.seminoncol.2010.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Schoder H, Gonen M. Screening for cancer with PET and PET/CT: potential and limitations. J Nucl Med. 2007;48(Suppl 1):4S–18S. [PubMed] [Google Scholar]
- 35.Chen W, Cloughesy T, Kamdar N, Satyamurthy N, Bergsneider M, Liau L, Mischel P, Czernin J, Phelps ME, Silverman DH. Imaging proliferation in brain tumors with 18F-FLT PET: comparison with 18F-FDG. J Nucl Med. 2005;46:945–952. [PubMed] [Google Scholar]
- 36.Cai K, Haris M, Singh A, Kogan F, Greenberg JH, Hariharan H, Detre JA, Reddy R. Magnetic resonance imaging of glutamate. Nat Med. 2012;18:302–306. doi: 10.1038/nm.2615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Haris M, Singh A, Mohammed I, Ittyerah R, Nath K, Nanga RP, Debrosse C, Kogan F, Cai K, Poptani H, et al. In vivo magnetic resonance imaging of tumor protease activity. Sci Rep. 2014;4:6081. doi: 10.1038/srep06081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Haris M, Nanga RP, Singh A, Cai K, Kogan F, Hariharan H, Reddy R. Exchange rates of creatine kinase metabolites: feasibility of imaging creatine by chemical exchange saturation transfer MRI. NMR Biomed. 2012;25:1305–1309. doi: 10.1002/nbm.2792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Walker-Samuel S, Ramasawmy R, Torrealdea F, Rega M, Rajkumar V, Johnson SP, Richardson S, Goncalves M, Parkes HG, Arstad E, et al. In vivo imaging of glucose uptake and metabolism in tumors. Nat Med. 2013;19:1067–1072. doi: 10.1038/nm.3252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.van Zijl PC, Jones CK, Ren J, Malloy CR, Sherry AD. MRI detection of glycogen in vivo by using chemical exchange saturation transfer imaging (glycoCEST) Proc Natl Acad Sci USA. 2007;104:4359–4364. doi: 10.1073/pnas.0700281104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Haris M, Cai K, Singh A, Hariharan H, Reddy R. In vivo mapping of brain myo-inositol. Neuroimage. 2011;54:2079–2085. doi: 10.1016/j.neuroimage.2010.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.DeBrosse C, Nanga RP, Bagga P, Nath K, Haris M, Marincola F, Schnall MD, Hariharan H, Reddy R. Lactate chemical exchange saturation transfer (LATEST) imaging in vivo A biomarker for LDH activity. Sci Rep. 2016;6:19517. doi: 10.1038/srep19517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Singh A, Haris M, Cai K, Kassey VB, Kogan F, Reddy D, Hariharan H, Reddy R. Chemical exchange saturation transfer magnetic resonance imaging of human knee cartilage at 3 T and 7 T. Magn Reson Med. 2012;68:588–594. doi: 10.1002/mrm.23250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Madan A, Ganji SK, An Z, Choe KS, Pinho MC, Bachoo RM, Maher EM, Choi C. Proton T2 measurement and quantification of lactate in brain tumors by MRS at 3 Tesla in vivo. Magn Reson Med. 2015;73:2094–2099. doi: 10.1002/mrm.25352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wijnen JP, Van der Graaf M, Scheenen TW, Klomp DW, de Galan BE, Idema AJ, Heerschap A. In vivo 13C magnetic resonance spectroscopy of a human brain tumor after application of 13C-1-enriched glucose. Magn Reson Imaging. 2010;28:690–697. doi: 10.1016/j.mri.2010.03.006. [DOI] [PubMed] [Google Scholar]
- 46.Terpstra M, Gruetter R, High WB, Mescher M, DelaBarre L, Merkle H, Garwood M. Lactate turnover in rat glioma measured by in vivo nuclear magnetic resonance spectroscopy. Cancer Res. 1998;58:5083–5088. [PubMed] [Google Scholar]
- 47.Gutte H, Hansen AE, Johannesen HH, Clemmensen AE, Ardenkjaer-Larsen JH, Nielsen CH, Kjaer A. The use of dynamic nuclear polarization (13)C-pyruvate MRS in cancer. Am J Nucl Med Mol Imaging. 2015;5:548–560. [PMC free article] [PubMed] [Google Scholar]
- 48.Xu HN, Kadlececk S, Profka H, Glickson JD, Rizi R, Li LZ. Is higher lactate an indicator of tumor metastatic risk? A pilot MRS study using hyperpolarized (13)C-pyruvate. Acad Radiol. 2014;21:223–231. doi: 10.1016/j.acra.2013.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Brindle KM, Bohndiek SE, Gallagher FA, Kettunen MI. Tumor imaging using hyperpolarized 13C magnetic resonance spectroscopy. Magn Reson Med. 2011;66:505–519. doi: 10.1002/mrm.22999. [DOI] [PubMed] [Google Scholar]
- 50.Brand A, Singer K, Koehl GE, Kolitzus M, Schoenhammer G, Thiel A, Matos C, Bruss C, Klobuch S, Peter K, et al. LDHA-associated lactic acid production blunts tumor immunosurveillance by T and NK cells. Cell Metab. 2016;24:657–671. doi: 10.1016/j.cmet.2016.08.011. [DOI] [PubMed] [Google Scholar]
- 51.Zhou J, Tryggestad E, Wen Z, Lal B, Zhou T, Grossman R, Wang S, Yan K, Fu DX, Ford E, et al. Differentiation between glioma and radiation necrosis using molecular magnetic resonance imaging of endogenous proteins and peptides. Nat Med. 2011;17:130–134. doi: 10.1038/nm.2268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pacheco R, Oliva H, Martinez-Navio JM, Climent N, Ciruela F, Gatell JM, Gallart T, Mallol J, Lluis C, Franco R. Glutamate released by dendritic cells as a novel modulator of T cell activation. J Immunol. 2006;177:6695–6704. doi: 10.4049/jimmunol.177.10.6695. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Please contact author for data requests.