

Abstract
Despite substantial recent progress, our understanding of the principles and mechanisms underlying complex brain function and cognition remains incomplete. Network neuroscience proposes to tackle these enduring challenges. Approaching brain structure and function from an explicitly integrative perspective, network neuroscience pursues new ways to map, record, analyze and model the elements and interactions of neurobiological systems. Two parallel trends drive the approach: the availability of new empirical tools to create comprehensive maps and record dynamic patterns among molecules, neurons, brain areas and social systems; and the theoretical framework and computational tools of modern network science. The convergence of empirical and computational advances opens new frontiers of scientific inquiry, including network dynamics, manipulation and control of brain networks, and integration of network processes across spatiotemporal domains. We review emerging trends in network neuroscience and attempt to chart a path toward a better understanding of the brain as a multiscale networked system.
Neuroscience is entering a period marked by a rapid expansion in the size, scope and complexity of neural data acquired from large portions of nervous systems and spanning multiple levels of organization1,2. Much of these ‘big data’ represent networks comprising the relations or interconnections that link the many elements of large-scale neurobiological systems3–6. Examples include protein interaction and genetic regulatory networks, synaptic connections and anatomical projections among brain areas, dynamic patterns of neural signaling and communication associated with spontaneous and task-evoked brain activity, and interactions among brain systems and the environment in the course of behavior. Notably, these data often cross multiple levels of organization (neurons, circuits, systems, whole brain) or involve different domains of biology and data types (for example, anatomical and functional connectivity, genetic patterns and disease states, activity in distributed brain regions in relation to behavioral phenotypes). The size and complexity of these data require sophisticated strategies for statistical inference and dimensionality reduction7 and present important challenges for scalable computing8, data sharing, and reproducibility9.
These recent developments not only pose challenges, but they also provide unique opportunities, as sophisticated empirical methods for mapping and recording neurobiological data intersect with theoretical and computational advances in data analysis and modeling of brain networks. We propose to call the research agenda emerging at this intersection ‘network neuroscience’. In this review, we outline the fundamentals of network neuroscience and highlight a selection of current and future topics. Our emphasis is less on past achievements and more on identifying emerging trends that will shape the field going forward. We survey new empirical and computational tools and discuss how these tools can become useful for attacking long-standing complex problems and research frontiers. An important focus is on how such tools may help to uncover network structures and processes that support integrative brain function and span multiple spatial and temporal scales (Fig. 1). We argue that network neuroscience theories and tools will bring substantial changes to the types of questions that we can ask and the hypotheses that we can test, ushering in a new era of network-based inquiry into brain structure and function.
Figure 1.
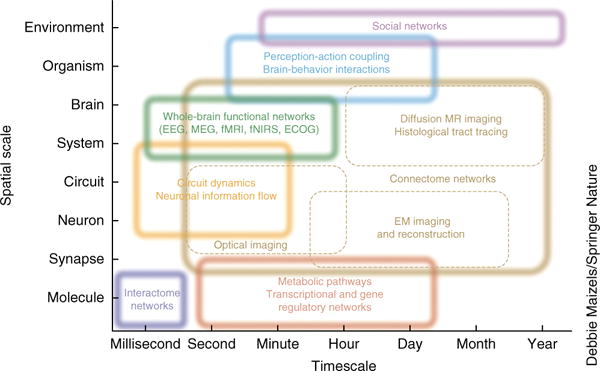
Networks on multiple spatial and temporal scales. Network neuroscience encompasses the study of very different networks encountered across many spatial and temporal scales. Starting from the smallest elements, network neuroscience seeks to bridge information encoded in the relationships between genes and biomolecules to the information shared between neurons. It seeks to build a mechanistic understanding of how neuron-level processes give rise to the structure and function of large-scale circuits, brain systems and whole-brain structure and function. However, network neuroscience does not stop at the brain, but instead asks how these patterns of interconnectivity in the CNS drive and interact with patterns of behavior: how perception and action are mutually linked and how brain-environment interactions influence cognition. Finally, network neuroscience asks how all of these levels of inquiry help us to understand the interactions between social beings that give rise to ecologies, economies and cultures. Rather than reducing systems to a list of parts defined at a particular scale, network neuroscience embraces the complexity of the interactions between the parts and acknowledges the dependence of phenomena across scales. Box dimensions give outer bounds of the spatial and temporal scales at which relational data are measured and interactions unfold, and over which networks exhibit characteristic variations and dynamic changes. Inspired by an iconic image of neuroscience recording methods, last updated in ref. 1. ECOG, intracranial electrocorticography; EEG, electroencephalography; fMRI, functional magnetic resonance imaging; fNIRS, functional near-infrared spectroscopy; MEG, magnetoencephalography.
Network mapping and observation
One of the major developments supporting the growth of network neuroscience involves the creation of powerful methodologies that enable the comprehensive recording of connections and interactions in neurobiological systems. What these methodologies have in common is that they capture large numbers of elements and then record all of their interactions in parallel, resulting in data sets that take the mathematical form of graphs or networks (Fig. 2). Major advances have occurred in the measurements of molecular interactions, the mapping of structural connections among neurons and brain areas (connectomics), the physiological recordings of activity in ensembles of neurons or across the whole brain, the automated classification of behaviors, and the tracking of social dynamics.
Figure 2.
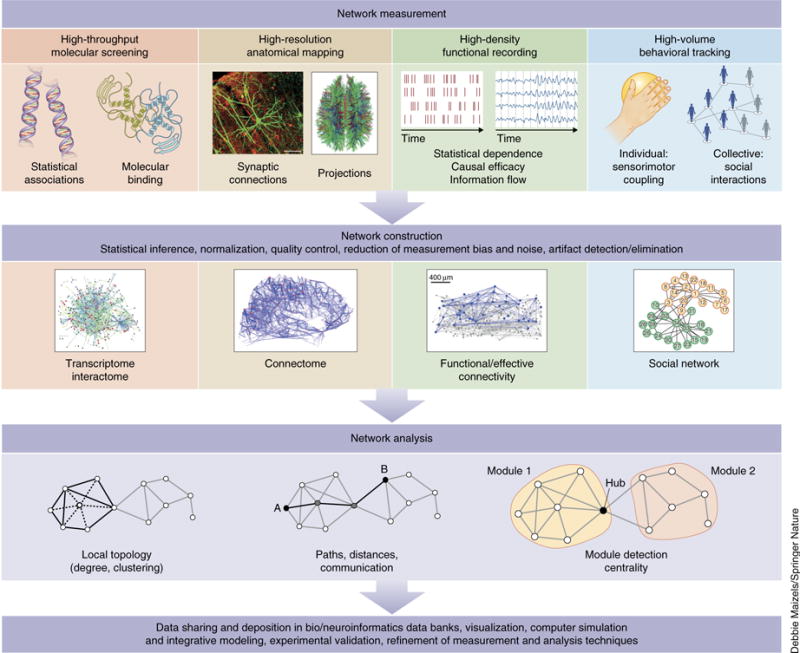
Network measurement, construction and analysis. Top, network neuroscience begins with the collection of relational data among elements of a neurobiological system. These data may refer to statistical associations among genes, physical binding events among macromolecules, anatomical networks of synaptic connections or inter-regional projections, multi-dimensional time series and their statistical dependencies or causal relations, or links in behavior, such as dynamic couplings among sensors and effectors in individuals or collective social interactions. Middle, once collected, relational data are generally subject to normalization, artifact and noise reduction before being assembled into the mathematical form of a graph or network, consisting of nodes (elements) and edges (their relations). Common examples are transcriptome and interactome networks, connectomes, networks of functional and effective connectivity, and social networks. Bottom, the common mathematical framework of graph theory offers a set of measures and tools for network analysis. As we argue in this review, descriptive measures such as the ones shown here are but a first step toward more powerful analysis and modeling approaches, such as generative modeling, prediction and control. Finally, network data are generally shared in large repositories, and numerous follow-up tool kits allow sophisticated visualization and simulation. Continual refinement of measurement, construction and analysis techniques ensures that the shape of this diagram will change as the field of network neuroscience matures. Image of functional/effective connectivity reproduced from ref. 70, Society for Neuroscience.
Building on the as yet unparalleled achievement of mapping the neuronal wiring of the nematode C. elegans10, the reconstruction of synaptic connectivity among neurons at the ultrastructural level has evolved rapidly, as has measurement of extra-synaptic connectivity11. Among recent studies in C. elegans12, as well as in the Drosophila visual system13, are refined mappings of connections among sensory and effector neurons, whose network patterns have offered important insights into the specific behavioral contributions of network elements. However, the enterprise of ultrastructural connectomics has been challenged by the sheer size and volume of vertebrate brains14. One of the largest efforts thus far has resulted in a connection map of a portion of the mouse retina involving the tracing of the neurites of 950 distinct neurons15. Although such ‘dense reconstructions’ capture cellular architecture in fine detail, they remain vulnerable to reconstruction errors and require expert curation for reliable detection of synaptic couplings. Alternative approaches use RNA barcoding to label projections of individual neurons, which may allow for high-volume tracing of neuronal circuits at single-cell resolution16.
Complementary mesoscale efforts to map more extended portions of the nervous systems of several model organisms have employed large-scale optical imaging as well as quantitative histological tract tracing. High-throughput tracing and imaging approaches (which require careful monitoring of sensitivity and specificity) have resulted in network data sets recording the connections among ‘local processing units’ in Drosophila17 as well as interregional projections in the mouse18. A different approach builds on expertly curated neuroinformatics databases of anatomical observations to construct aggregated data sets of network connectivity, such as in rat19 and macaque monkey20. Quantitative assessment of tract tracing experiments reveals relatively dense networks that include projections ranging in strength over six orders of magnitude21. Finally, there is continuous improvement of methodology in diffusion imaging and tractography, which allows the inference of the trajectory and strength of white matter projections in the human brain22. Although subject to many experimental and statistical limitations23 and interpretational pitfalls24, comparative studies have suggested that representations of long-distance connectivity derived from invasive histological methods and noninvasive imaging of neuroanatomical structure are significantly related25.
Advances in optical cellular imaging have enabled recordings of neuronal activity in extended functional circuits26, in some cases spanning the whole organism27. Imaging of Ca2+ dynamics, as well as genetically encoded fluorescent reporters of membrane voltages, generates dynamic functional data from hundreds to thousands of neurons. Following processing steps such as image registration and cell sorting, the data can be represented as time traces that can be subjected to statistical time series analyses. Parallel recordings from hundreds of neurons enable computational strategies that identify and characterize functional interactions and statistical dependencies between neurons. The resulting functional networks can be examined for modular organization, as well as for evidence of coherent network states and patterned temporal dynamics. Functional network analysis has progressed most strongly in applications to noninvasive electromagnetic or functional magnetic resonance imaging recordings from the human brain. Major themes include the definition of coherent subnetworks spanning the whole brain28 that exhibit changing topology in conditions of rest (spontaneous activity) and task demand29, or in relation to visual input30. Methodological advances involve improved sensitivity in measures of statistical dependence, inference of causal links, and greater temporal resolution. An important frontier is the analysis of sequences of functional networks that change across time.
Linking elements and interactions in the brain to different domains of behavior has advanced from classic univariate (one region, one behavior) to bivariate (connectivity, behavior) and finally to multivariate frameworks31. Large-scale studies of brain-behavior relations and behavior-behavior dependencies, although still in their infancy, promise to provide a rich database for mapping the relations among brain processes and their contributions to perception, action and cognition. In one such study carried out in Drosophila, the roles of neurons in triggering a diverse set of behaviors was systematically investigated by optogenetically stimulating individual neuronal cell lines and recording the associated behavioral responses32. Machine-learning techniques were then applied to extract statistically robust relations between neuron lines and behavioral phenotypes, resulting in a neuron-behavior atlas. This work illustrates the utility of using relational data to establish mappings from clusters of neural elements to clusters of behavioral phenotypes. On a very different organizational scale, brain-behavior relations have been approached through meta-analyses of large repositories of human neuroimaging experiments reporting patterns of brain activation in relation to different domains of behavior and cognition. Aggregating brain imaging data from thousands of such studies allowed the construction of ‘co-activation networks’, whose major components and overall network topology strongly resembled functional networks derived from resting-state (task-free) recordings33. Finally, as demonstrated in a study of chemotaxis in C. elegans34, our understanding of the relationships between neural dynamics and observable behaviors can benefit from integrating data capturing the topology of anatomical networks with behavioral data acquired as the organism is interacting with its environment.
Brain networks may be viewed as ‘intermediate phenotypes’35 that are situated between the domains of genetics and molecular systems, and the expression of individual and collective behavior in the environment. As such, brain networks mediate the causal effect of genetics on behavior and vice versa. For example, genetic mutations may cause changes in network topology that in turn drive alterations in behavior. Indeed, network science has made substantial advances into characterizing both molecular and social systems, and the effect of these advances are beginning to be felt at the intermediate scales of mesoscale neural circuits and large-scale brain systems.
At molecular scales, the arrival of comprehensive molecular and genomic data for many organisms, including humans, has ushered in the new discipline of systems biology36, which combines high-throughput measurements of molecular data with modeling and computational analysis. Systems biology recognizes that most biological functions and phenotypes cannot be reduced to the action of individual proteins or biomolecules, but rather emerge from their interactions in molecular complexes and in cells, giving rise to spatiotemporal patterns of gene expression, tissue growth and differentiation, and other integrative biological processes. In systems biology, networks are core ingredients for analysis and modeling37, for example, in proteomics38, mapping of gene-disorder relationships39 and genetics-based comorbidity studies40. The architecture of these networks at the molecular scale affects higher order functions measured at the larger scale of functional brain areas. Here, network interactions offer important insights into the biological mechanisms associated with several common brain disorders, including schizophrenia41 and autism42. Convergent evidence suggests that the biological bases of psychiatric illness cannot be fully accounted for by small numbers of mutations or risk factors. Instead, these disorders involve disturbances in biological networks on multiple spatial scales.
Social data, aggregated into networks, capture important aspects of individual and collective behavior. The ubiquity of digital traces of human behavior has led to a rapid expansion of computational social science43, a new discipline that leverages methods and tools from statistical physics and computer science to observe and predict the collective behavior of organizations and societies. Going beyond small-scale analyses of data laboriously gathered from individuals in laboratory settings, computational social science relies on the parallel and pervasive collection of rich spatiotemporal data capturing the organization and dynamics of large-scale social systems. The approach has promise in providing new means to monitor and enhance behavioral and mental health, such as in individuals with major depressive disorder44.
Network analysis and modeling
Networks are phenomena of the natural, social and technological world that are studied across many disciplines with a common toolset provided by network science45. Perhaps most foundational is graph theory, a branch of mathematics that examines the properties of graphs or networks, defined as sets of nodes and edges that represent system elements and their interrelations46. In neuroscience, descriptive measures that report on local and global features of network topology have been widely applied across structural and functional data sets from multiple species. These analyses have consistently revealed nonrandom topological attributes, such as high clustering and short path length3,47, and network communities (modules) linked by highly connected hub nodes48 that are in turn densely linked, forming an integrative core49 or rich club50. Recent investigations have examined more complex organizational features such as hierarchical organization51,52, the role of geometry and spatial embedding in large-scale anatomical connectivity53, and the importance of considering relationships among multiple structural constraints when accounting for the emergence of significant network attributes54. Indeed, the brain is inherently a spatially embedded network, and physical constraints resulting from that embedding underlie functionally important network characteristics, such as efficient network communication and information processing. The application of graph measures has also been critically examined with respect to sensitivity to node/edge definition, spatial and temporal resolution, and reliability and reproducibility across observations55. Important issues currently under development involve improved and domain-appropriate approaches for module (community) detection56, model-based inference of networks from observational data57, and statistically principled methods for network comparison across different individuals and in different conditions58.
More recently, methods from network science are expanding in new directions, going beyond descriptive accounts of network topology and toward addressing network dynamics, generative principles and higher order dependencies among nodes. One prominent example is the development of methods for assessing multi-scale organization in networks. This includes characterizing fluctuations in community structure of networks across time59, and implementing dynamic processes on networks as a diagnostic tool for explicitly linking micro-scale features of network organization to macro-scale characteristics of neurophysiological dynamics60. Yet another approach uses network science to ask questions about the processes that can potentially generate the topology of an empirical network. Such generative models can clarify the contributions of spatial embedding and other (non-spatial) wiring rules in shaping the network topology of the connectome61, and can also reveal potential factors driving the selection of functionally important network attributes62. Finally, the application of concepts from algebraic topology (Fig. 3) attempts to discern non-random structure in networks by going beyond dyadic relations (two nodes linked by an edge) and considering non-dyadic, higher order relations among network nodes63–65, a goal that is complementary to that motivating the use of graphs in which edges can link any number of nodes, or so-called hypergraphs66. This approach can identify non-random structure in structural connectivity of cortical microcircuits67, such as unexpectedly high numbers of directed ‘all-to-all’ connected cliques of neurons, or cavities in which edges are conspicuously absent68.
Figure 3.
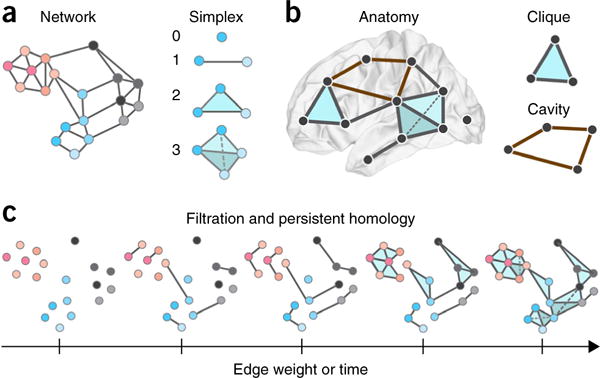
Algebraic topology and simplicial complexes. (a) Left, the traditional way in which to study networked systems including the brain is to examine patterns in pairwise relationships between nodes (dyads). Indeed, the dyad has traditionally been the fundamental unit of interest in graph theory and network science. Here we show an example brain network, composed exclusively of dyads. Right, in many cases, however, neural systems appear to employ higher order interactions139, which increase the complexity of neural codes that produce the wide range of behaviors observed in these systems140. To study these higher order interactions, one must expand one’s worldview to include units of interest that exceed the simple dyad. Using recent advances in the field of applied algebraic topology, we can study so-called simplicial complexes, which are generalizations of graphs that encode non-dyadic relationships63. Here we show representations of a zero-simplex (a node), a one-simplex (an edge between two nodes), a two-simplex (a filled triangle), etc. (b) Left, we can study the location of these simplices in brain networks, from the small scale of neurons to the large scale of brain regions. For example, we show a toy simplicial complex embedded in the human brain, and containing a zero-simplex, several one-simplices, a two-simplex and a three-simplex. Right, when doing so, it is interesting to characterize the locations of cliques (all-to-all connected subgraphs of any size) and cavities (a collection of n-simplices that are arranged so that they have an empty geometric boundary), which are structurally predisposed to be critical for in integrated (cliques) versus segregated (cavities) codes and computations. Here we show a two-clique (top) and a cavity bounded by four one-simplices (bottom). (c) It is also often interesting to study how networks evolve in time or how their internal structure depends on the weight of the edges between nodes. In these and similar scenarios, we can apply a powerful tool from algebraic topology called a filtration, which can represent a weighted simplicial complex as a series of unweighted simplicial complexes. We can then trace the evolution of specific cavities from one complex (one time point or one edge weight value) to another, as well as locate the moment of their creation or collapse. Collectively, this is called the persistent homology of the weighted simplicial complex, and it can be statistically quantified using a collection of functions called Betti curves, which record the number of cavities in each dimension. Here we show a filtration on edge weight, which begins with all nodes being disconnected because no edges exceed a threshold value τ. At subsequent points along the filtration, those nodes are connected by simplices that are composed of edges of weight greater than or equal to τ. This same sort of structure can be observed in a filtration of a growing network over time: the filtration begins with all nodes being disconnected because no edges exist. At subsequent points along the filtration, those nodes are connected by simplices that are composed of edges that have grown at later time points. Thus, filtrations allow the investigator to assess the organization of weighted simplicial complex representations of brain structure and dynamics as a function of edge weight, or as a function of time.
Time series analysis is a common basis for constructing edges in functional networks. Generally, different methods aim to extract covariance or correlation (hence delivering non-causal similarity-based metrics of statistical dependence) and to estimate the direction and strength of causal influence. Although correlation-based metrics are generally simple to compute, the propensity of correlations to ‘fill in’ links among indirectly coupled nodes (transitive closure)69 tends to result in networks that are overly dense, clustered and modular, containing many dependencies that arise from indirect interactions in the underlying anatomy. Although computing partial correlations can address this issue and yield sparser networks, the resulting topologies are difficult to interpret, may regress away true interactions, and require many samples to achieve stability. Measures of directed influence based on temporal precedence cues such as Granger causality and transfer entropy result in directed edges that provide information about the extent to which the past of one time series can predict the future of another. In this narrow sense, these networks portray potential causal relations among network nodes, and they have been shown to have some utility in systems of spiking neurons70 and electrophysiological recordings. In contrast with time series analysis tools for functional networks, effective onnectivity offers a model-based account of directed causal links among network elements that is derived by rigorous model selection criteria. Model-based effective connectivity is central to the framework of dynamic causal modeling71, which allows the estimation of dynamic couplings among brain nodes and their modulation by time, task or sensory input.
As is the case in systems biology, there is a growing role for numerical simulations and computational models of brain circuits and systems in explaining and predicting empirical functional network data. These approaches can provide effective generative models of brain responses that simulate the activity of neural elements and their interactions. Simulation environments, such as The Virtual Brain at the level of whole-brain models72, Neuron at the level of neuronal circuits73, and OpenWorm specifically for the organism C. elegans74, generate neuronal time series resulting from the biophysical properties of neuronal elements that are linked by a network of structural (synaptic) couplings. Such forward modeling can be combined with model inversion that, constrained by model selection, allows inferences on model parameters associated with candidate neurobiological mechanisms75. Similar modeling and simulation approaches are applied to protein interaction and genetic regulatory networks76, can capture the translation of genotype to phenotype in whole cells77, and have become important tools in studies of social dynamics and collective behavior78. The ever accelerating power and decreasing cost of computing has brought network-based simulations and mathematical models of neurobiological systems at all scales within the reach of most investigators.
Current frontiers
Network dynamics
One of the emerging frontiers of network neuroscience is the investigation of network dynamics (Fig. 4). The broad question of how networks change encompasses at least two important and distinct dimensions79: how activity patterns can change on top of a fixed structural network (referred to as dynamics on networks) and how network edges themselves can reconfigure (referred to as dynamics of networks). Although most current investigations are focusing on one or the other of these two dimensions, future work will undoubtedly expand to develop methods and insights that straddle both domains.
Figure 4.

Dynamic and multilayer networks. In the field of network science, two types of dynamic processes are studied in some detail: dynamics on networks and dynamics of networks. (a) Dynamics on networks indicates that the activity (or some other property of interest) of nodes changes as a function of time. Here we illustrate decreasing activity (pink), increasing activity (gray) and changes in the pattern of activity (blue) over time in distinct network modules or communities. (b) Dynamics of networks indicates that the edges of the network themselves change either in their existence/absence or in their strength. Here we illustrate the coalescence of modules (blue and yellow), as well as the transfer of allegiance of a single region from one module (pink) to another (yellow) over time. This latter process can be quantified using the statistic of network flexibility, and has been shown to be an important indicator of human learning141 and a correlate of individual differences in executive function142. (c) Although not yet common, studies addressing the combined problem of dynamics on and of networks will be of increasing importance in the coming years. There is a critical need to better understand the relationships between changes in connectivity and changes in activity, gray matter, neurotransmitter levels, genetic expression or other nodal properties. Methods to bridge these scales will be critical in advancing network neuroscience toward more mechanistic models and insights. (d) One particularly useful construct in the context of dynamic and multimodal networks is that of multilayer networks88. Multilayer networks are networks whose nodes may be connected by different types of edges, with each type being encoded in a different layer90. These layers could, for example, represent different time points, subjects, tasks, brain states, ages or imaging modalities. In multilayer networks, nodes in one layer are connected to corresponding nodes in other layers by identity links (a distinct sort of edge), which hardcode the non-independence of data obtained from these nodes. Here we show the simplest case in which all nodes and all edges exist in all layers, but multilayer network tools can also be used in cases in which nodes and edges change across layers. We also illustrate the simplest inter-layer connection pattern, with identity links connecting consecutive layers; however, alternative connection patterns are possible59.
The concept of dynamics on networks builds on the notion that the complex physiological activity of neural systems is fundamentally constrained by the patterns of connections between their elements. These processes include neuronal activation, the release of neurotransmitters, the editing of genetic expression, the utilization of energy in terms of cellular metabolism and the assembly of information. How these processes depend on underlying connections between neurons, areas or genes is the perennial question of how form constrains function. The first forays into this question have emerged at the level of large-scale neuroimaging in humans, where activation patterns at rest can be partially predicted by the underlying network of white-matter tracts80,81. Modeling the evolution of these patterns can be tackled using first-order difference equations, oscillators, neural mass models and other approaches82. Perhaps equally intriguingly, one can also exercise phenomenological models to gain an intuition regarding processes whose physiology is less well understood, such as the trans-synaptic spread of prions thought to cause dementia83 or the propagation of information packets along parallel structural routes84.
Notably, changes in function can elicit changes in structure, leading to dynamics of networks. The first conception of dynamics of networks grew from the recognition that many physical and biological systems display patterns of connections that change over time, in different contexts or in response to varying external demands. Indeed, this recognition now pervades many areas of neuroscientific investigation, from the changing patterns of functional connectivity85 to the patterns of genome folding over developmental timescales86 or genetic coexpression over evolutionary timescales87. Quantitatively characterizing and mathematically modeling these evolving patterns of connectivity was largely intractable before recent advances in the field of applied mathematics. So-called multilayer networks88 offer mathematically principled models of evolving networks as well as a battery of statistical variables to characterize their evolution. Initial applications of these tools to neuroimaging data have addressed questions of how connectivity between brain regions changes over the lifespan89, during behavioral adaptation or learning90, and across frequency domains91. However, these applications have only begun to scratch the surface of empirical avenues of inquiry in which patterns of connections between neurophysiological elements evolve or change in a manner that is critical for system function. A current fundamental challenge lies in constructing appropriate generative models that can be used to infer the dynamic mechanisms potentially driving these network reconfigurations and how they might be altered in disease states92.
Although dynamics on networks and dynamics of networks are both important areas of research, emerging tools from machine learning are beginning to bring the two areas together to better understand the time-varying expression of sets of networks, where all networks in the set exist at every time point, but each network is expressed to differing amounts93. In addition, developing methods for understanding the dynamics of networks-of-networks that are interconnected with one another across contexts and scales remains challenging94. Could changes in functional connectivity driven by task performance be predicted by an individual’s place in their social network95? Could changes in the interactions among individuals lead to changes in the inter-subject correlation in brain activity96? What are the meaningful (and tractable) classes of networks-of-networks that we can begin to understand? These and related questions will likely compose an increasing focus of network neuroscience in the immediate future.
Prediction
As it is still a relatively young field, network neuroscience has focused on describing neuroscientific data in the language of graph theory and network science, and it has begun to recast avenues of inquiry in terms of explicitly network-based hypotheses. However, this general paradigm shift underscores the necessity of complementing simple descriptions with mechanistic predictions97. Indeed, although network models are traditionally viewed as tools to quantify structure, they have a complementary role as tools to predict system function. Moreover, this predictive power can be used broadly across genetics, systems biology, neuroscience and social science.
According to a standard formal definition, the accuracy of a prediction improves as the difference between the expected behavior and the observed behavior diminishes. Predictions of network dynamics, system function or other network-based phenotypes can be obtained in one of two ways: using black-box approaches such as those afforded by machine learning or using first-principles mechanistic models such as those developed in the field of theoretical physics. Although the former has rarely been explicitly combined with network neuroscience98, the latter has demonstrated initial utility in contexts as far ranging as predicting therapeutic targets in cellular regulatory networks99 and predicting seizure activity in medically refractory epilepsy100. These successes depend on a fundamental understanding of the network interactions and the energy landscape defining how the brain moves between different states101, which predict not only common brain states, but also the probability of transitioning between them. In addition to methodological approaches to understanding these landscapes, open areas of inquiry include whether one can predict cognitive function from structural or functional network architecture102, predict disease onset or progression from early or pre-clinical data103, predict behavioral responses to health messaging104, or predict optimal strategies for early intervention.
Perturbation, manipulation and control
Once a predictive model of a system is built, one has the potential to carefully perturb, manipulate and indeed control the system with an explicit knowledge of the outcome (Fig. 5). Network neuroscience is poised to offer a new conceptual and computational framework in which to guide these interventions in a theoretically principled manner for maximal cognitive enhancement or therapeutic benefit.
Figure 5.
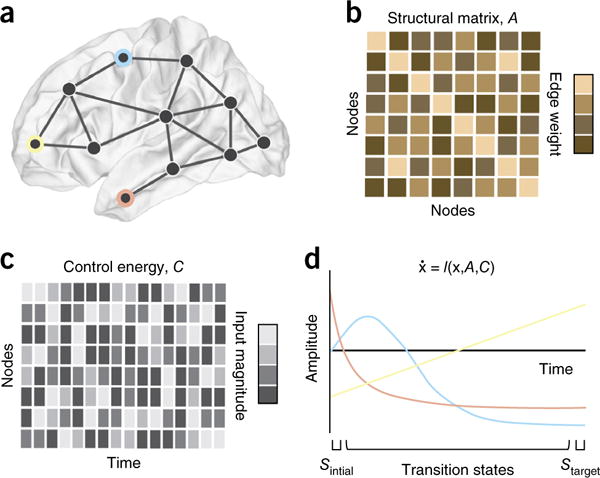
Controlling brain networks. Following a careful description of the network properties of the brain, we might wish to intervene: to push a diseased brain toward health or to enhance the function of a brain that might not be reaching its full potential. In the context of brain networks, this question becomes has provided a question of so-called ‘network control’, for which the field of engineering extensive and carefully validated solutions in recent years112. (a,b) Here we illustrate a problem in which we wish to modulate the activity of three brain regions (blue, peach, gold; a) that are connected both to one another and to other regions in the brain by aset of anatomical links of varying strength or weight (b). (c,d) We wish to determine the amount of control energy (which can take the form of brain stimulation or task demands) that must be injected into brain regions at each time point (c) to affect a continuous change in the amplitude of the three brain regions of interest from a (d), moving them from an initial state characterized by one pattern of activation to a target state characterized by a different pattern of activation. One way to address this problem is to model brain dynamics as a (linear or nonlinear) function of an initial state x, a structural adjacency matrix A and a control energy matrix C. Using techniques from network control theory, we can solve for the optimal control energy, which is usually defined as the smallest amount of energy required to affect the transition (from initial to final state) in a given time period T. Interesting problems include determining which regions can affect which types of control113, which regions form optimal targets or optimal drivers143, how many control points are required, and which transitions are preferredby the system. In many cases, the best model of dynamics is unknown, and in this case an engineering technique known as systems identification can be very useful; this technique reveals not only the model of dynamics, but also the structural matrix A that best explains regional time series. Note that the a matrix uncovered by systems identification might not be identical to an anatomical matrix of synaptic connections. In the coming years, we anticipate that these techniques, and their careful adaptions to neural systems, will prove to be particularly useful in the control of brain networks both for clinical purposes, for example, in optimizing transcranial magnetic stimulation to large-scale brain regions in the human brain144.
Here we take an eclectic view of what constitutes an intervention by including optogenetics, brain stimulation, lesions, neurofeedback, mood induction, task priming, training and task performance. What each of these manipulations has in common is the fact that they can be used to perturb (or change) brain state and its associated dynamics. Although this has been acknowledged for many decades, it has begun to be exploited to address explicitly network-based hypotheses105. For example, what network do we wish to (in)activate? And at what spatial scale does this network operate, or at what spatial scale does this network go awry, for example, in the case of epilepsy (Fig. 6a–c)? What effect will focal perturbations have on network function106 or network reconfiguration? Is there a predictable pattern of network recovery following disturbances resulting from stroke or other injury107?
Figure 6.
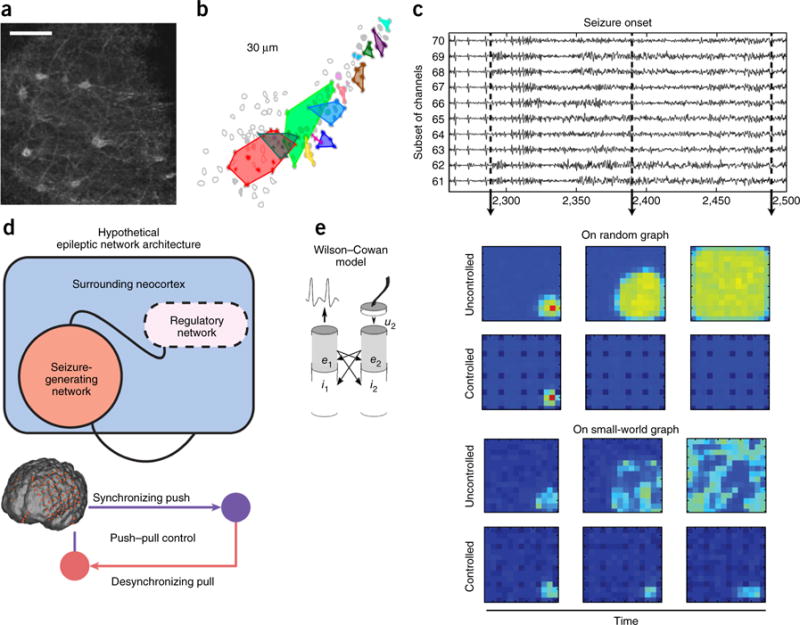
Epilepsy as a multiscale network disorder amenable to control. (a–c) Epileptic activity can be observed across spatial scales, from the level of single neurons up through the level of large-scale areas. (a) Spatial configuration of neurons during an interictal spike in the stratum oriens of a mouse; scale bar represents 10 μm. Reproduced from ref. 145, Sarah Feldt Muldoon, Vincent Villette, Thomas Tressard, Arnaud Malvache, Susanne Reichinnek, Fabrice Bartolomei, Rosa Cossart, GABAergic inhibition shapes interictal dynamics in awake epileptic mice, Brain, 2015, 138, 10, 2875–2890, by permission of Oxford University Press. (b) Cellular activity over a scale of 30-μm clusters together dynamically, forming network modules each shown in a different color, and these clusters are altered in the epileptic brain indicating a microscale network disorder. Over long periods of time, these changes in activity alter the underlying structural connectivity between ensembles, permanently changing the anatomical constraints on neurophysiological processes. Reproduced from ref. 146, National Academy of Sciences. (c) This clustering at the level of neuronal ensembles is accompanied by a large-scale oscillatory signature in intracranial electrocorticographic recordings. Reproduced from ref. 147, National Academy of Sciences. (d,e) Neurological disorders such as epilepsy are amenable to interventions that are informed by notions of network control. (d) The large-scale dynamics of epilepsy are characterized by a spatially intricate and temporally evolving pattern of synchronization and desynchronization that suggests the presence of a homeostatic push-pull control mechanism (bottom), whereby a regulatory network outside of the seizure generating zone controls the spread of seizure activity (top). These data provide two distinct targets for surgical and stimulation-based intervention: the seizure-generating zone, and the seizure-regulating zone. Reproduced from ref. 116, Elsevier. (e) Exogenously controlling the dynamics in either of these zones using stimulation requires careful computational models of control mechanisms, optimal stimulation intensities and optimal targets of that stimulation as a function of time. For example, here we illustrate recent work that models these large-scale dynamics using Wilson-Cowan oscillators. A distributed control mechanism, in which control is enacted by all stimulation sites, may be useful for quieting seizure activity, particularly when the underlying structural network is relatively random (top). However, when the underlying structural connectivity displays a small-world organization, a blanket distributed control mechanism is less effective, and may instead need to be better tuned to control drivers in the regulatory or seizure-generating networks (bottom). Reprinted with permission from ref. 148 as follows: ShiNung Ching, Emery N. Brown & Mark A. Kramer, Phys. Rev. E, 86, 021920 (2012), Copyright 2012 by the American Physical Society.
The ability to conceptualize network-based hypotheses, coupled with the growing potential to modulate neural function across many spatial scales108, brings with it important engineering challenges109. How do we titrate the location, strength or spatiotemporal extent of a perturbation? How do we model the relative contributions of network structure and dynamics on the functional effects of perturbations? Drawing on the wealth of knowledge gleaned across the ancient field of physics, one realizes that one can only hope to answer these translational questions when one is able to construct a meaningful network-based theory of brain function. A particularly promising example of such a theory lies in network control, a nascent field of engineering that has its roots in the 1970s110 and has been re-popularized more recently111. Network control combines estimates of network connectivity with models of system dynamics to predict where in the system one should inject energy to push the system toward a desired target state or target dynamics112. Such a theory offers the fundamental backdrop against which to better understand cognitive control113, optimize stimulation for neurological disorders114, maintain and control levels of anesthesia115, and inform surgical or stimulation-based interventions such as in the case of epilepsy116 (Fig. 6d,e). Future efforts in using network neuroscience for translational purposes will hinge on the development of this or related theories to predict the effects of perturbations on network structure and function.
Crossing levels
Comprehensive multi-scale accounts of brain function are central objectives for neuroscience2, but how can such an understanding be achieved? Networks are not only encountered at virtually all scales of neurobiological systems, they also offer a promising theoretical and analytical framework for bridging these scales and for creating new insights about species commonalities and differences. A case in point is the potential for network models to facilitate integration across micro-, meso- and macro-connectomics. As discussed earlier, improvements in accuracy and scalability of macroconnectome data acquisition and analysis may soon yield large-scale dense reconstructions of neural circuitry at the ultrastructural level (for example, see ref. 15). In parallel, statistics on cell types, laminar profiles, connection probabilities, etc. can inform detailed circuit models with substantial gains in scale and coverage117–119. A combination of dense reconstruction and computational inference could then be used to construct cellular-resolution models of brain areas and eventually entire brain systems. Another case in point is the application of network approaches to the comparison of networks across species120. Thus far, comparative studies have revealed some important commonalities (for example, shared attributes such as modules and hubs), but have also highlighted differences, such as in the organization of inter-areal connectivity in the cerebral cortex of several primate species121,122. In future studies bridging scales and species, increasingly sophisticated network science approaches for graph modeling, dimensionality reduction and community detection, and extensions into multigraphs and spatial networks, will be important adjuncts.
Another bridge is that between structure and function, as illustrated above in the emerging frontier of network dynamics. Networks from connectomics provide a structural skeleton of physical links among neurons and brain areas: networks created by dynamic brain activity, although constrained by the anatomical couplings, form a rich dynamic repertoire of topologies that continually change across time. The fundamental role of structural constraints in defining a ‘sub-space’ or manifold for brain dynamics is encountered in mesoscale neuronal circuits as well as in large-scale brain networks. In circuits, the computational roles and capacities of neurons are defined by network connectivity. In larger systems, coherently active networks and their fluctuations across time are shaped by anatomical projections. It must be noted that topology alone is insufficient to account for all of the observed variance, as the biophysical attributes of nodes and edges have a major role in shaping neural activation and coactivation. Network science offers a set of tools specifically designed to capture different domains of network organization. For example, multiplex or multilayer networks can represent structural and functional connectivity in a single network model123. Computational models of network architecture allow simulations of collective network states and network dynamics across scales from neuronal populations to the whole brain72,80,82,118,119. Models of this kind can become useful tools for predicting the effects of manipulations and perturbations on network function, from simulating genetic knockout experiments to rewiring of circuits, stimulation of specific cell types, effects of brain lesions, and varying conditions of neuromodulation and sensory input.
Networks can also bridge across data of very different types and from different domains of biology. One example is the joint investigation of gene coexpression patterns and patterns of brain connectivity (Fig. 7). Studies in the rodent nervous system have demonstrated that brain areas with similar gene transcription also maintain similar connectivity profiles124, and that transcriptional patterns are more similar among pairs of areas that are directly anatomically connected than among unconnected pairs125. In human cerebral cortex, transcription patterns are more similar in coherently active functional networks126 and exhibit correlations with activity levels observed during rest127. Transcriptional profiles also exhibit relationships with the topological roles of brain areas in the macroscale connectome. In the mouse brain, highly connected and highly central areas exhibit a common and distinct topological profile, specifically involving genes regulating oxidative metabolism, suggestive of links between network topology and metabolic demand128. A separate study found evidence of elevated expression of genes associated with cognition, learning and memory in hub nodes of the mouse brain129. These studies raise important questions about the nature of the mechanisms that tie the topology of structural and functional brain networks to fundamental aspects of basic brain physiology.
Figure 7.
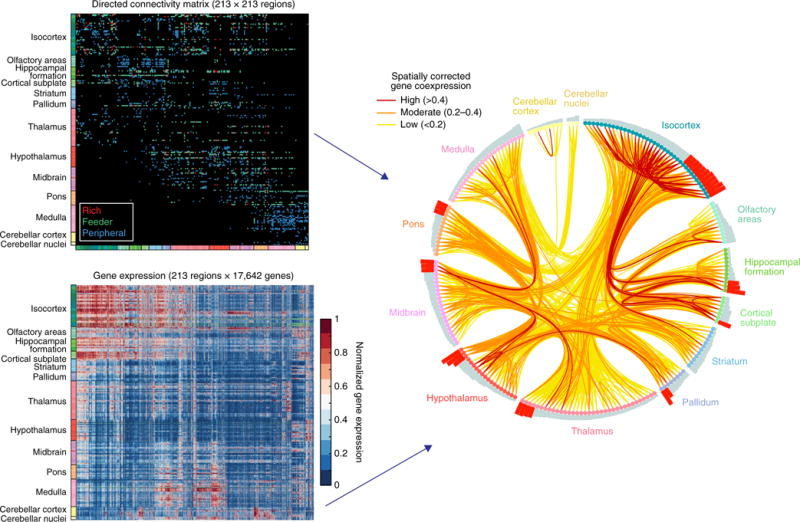
Relations among anatomical connectivity and gene co-expression networks. Top left, matrix of anatomical connections among 213 mouse brain regions. Regions (nodes) with more than 44 distinct connections were considered hubs, and connections were classified as hub→hub (rich), hub→nonhub (feeder) or nonhub→nonhub (peripheral). Bottom left, normalized expression levels of 17,642 genes across 213 brain regions. Genes with highly correlated expression profiles are placed near each other. Right, brain regions have been arranged around a circle, ordered by number of connections (bars) in each anatomical subdivision. Hubs are marked by red bars. The connection diagram traces anatomical connections between pairs of brain regions, color-coded by the corresponding gene coexpression value, after applying a correction for spatial distance. Statistical analysis revealed strongest gene coexpression among pairs of regions linked by reciprocal connections (as compared with unidirectional or unconnected pairs), as well as for rich connections linking hubs (as compared with feeder and peripheral connections). Genes driving correlations in expression in connections involving hub regions are functionally enriched in oxidative energy metabolism. Connectivity data derived from ref. 18. Reproduced from ref. 128, National Academy of Sciences.
Finally, crossing levels with the aid of network science is particularly salient when considering the multiple markers, mechanisms and manifestations of brain disorders. For example, autism spectrum disorders are associated with numerous genetic risk factors, disturbed connectivity among brain regions and heterogeneous behavioral phenotypes130. More complete understanding of autism etiology requires integration across genetic, neuroimaging and phenotypic data131. Here, concepts from multilayer multiscale network science will likely be of use. Tracking cross-level interactions among networks may also offer new approaches for therapeutic intervention and manipulation. For example, a recent meta-analysis suggested a possible association between aspects of social relationships and cognitive decline132. Mechanisms of brain networks and social networks become intertwined (Fig. 8) in phenomena, such as social cognition, that appear to be disturbed in a range of psychiatric conditions133, and in the history of environmental exposures and experiences that constitute human development134. Notably, interactions between levels occur not only from micro- to macroscales, but can also result in top-down causal effects of macro- on micro-scales. For example, genomic profiles and patterns of gene expression can be sensitively influenced by environmental perturbations135 or patterns of behavior136. Such links across levels of networks become critical when considering complex real-world problems such as those posed by the design of effective interventions and treatments in the context of public health137. They are also attractive targets for future investigation employing quantitative tools and methods from network neuroscience.
Figure 8.
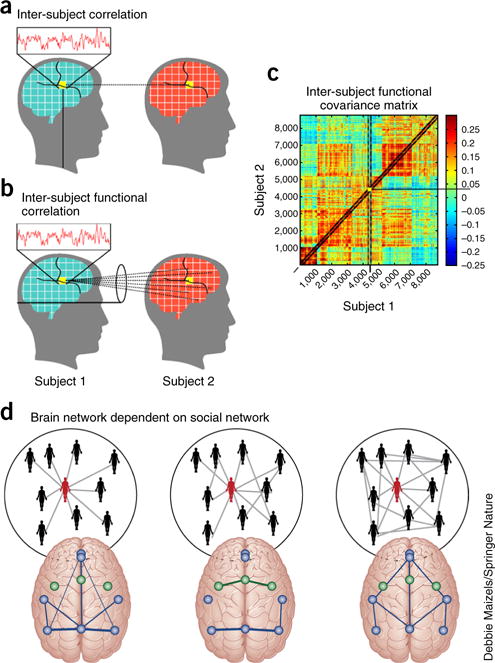
Crossing scales from brain networks to social networks. (a–c) As we interact with one another, our patterns of brain activity can track together, whether in a single voxel (a; inter-subject correlations), from a single voxel to other voxels (b; inter-subject functional correlation) or from any voxel to any other voxel (c; inter-subject functional covariance). These patterns can be studied from a network perspective using the tools of graph theory to better understand how relationships between individuals affect the similarities and differences in our patterns of brain activity. Taking the idea one step further, we can study how the patterns of brain activity in a person who is central in their social network differ from the patterns of brain activity in a person who is less central to their social network. Indeed, how our brains respond to or can be predicted from our social networks is a critical open question with direct import for health interventions at the large-scale of neighborhoods, cities, countries, and cultures (see also ref. 137). a–c adapted from ref. 149, Springer Nature, and d adapted from ref. 150, R. Schmaelzle, M.B. O’Donnell, J.O. Garcia, C.N.C. Cascio, J. Bayer, D. Bassett, J. Vettel and E.B. Falk.
Network science tools are perfectly suited to accomplish the important goal of crossing levels or scales of organization, integrating diverse data sets, and bridging existing disparate analyses. For example, the multilayer formalism recently developed in the field of applied mathematics88 provides a principled way in which two different imaging modalities that each provide estimates of connectivity between the same nodes, such as structural connections and functional connections, can be linked. The same multilayer network formalism can also be used to link data across different levels of spatial scale, where a node in one layer at a large spatial scale becomes a collection of nodes in the next layer at a smaller spatial scale. The multilayer framework allows the collection of statistics of network characteristics that are conserved or variable across modalities or across scales. Another important set of tools, annotated graphs— sometimes also called decorated graphs in mathematics—provide a means of assigning attributes to nodes138. In an annotated graph, a brain region can be characterized by inter-nodal relationships (for example, edges) as well as intra-nodal characteristics (for example, gene expression levels, cytoarchitectural properties, activity magnitudes or morphological features). Statistics can then be used to capture the relationships between connectivity and nodal characteristics in a model-based manner. Notably, this annotated graph approach can be used to bridge common analytical approaches that focus on nodal properties, including both univariate and multivariate approaches, and the emerging analytical approaches from graph theory that focus on inter-nodal relationships. Finally, whether crossing scales, integrating data or bridging analyses, network science also offers tools to capture higher order structure in data, including multi-omics data, using emerging concepts and algorithms from algebraic topology63–68 and topological data analysis.
Conclusion
In this review, we have attempted to sketch the outlines of a new interdisciplinary field, which we call network neuroscience. The field gathers momentum as networks have become ubiquitous phenomena encountered in empirical investigation as well as computational analysis and modeling of neurobiological systems at all scales. The ever-growing volume of big data in neuroscience demands not only advanced analytics and sound statistical inference, but it also calls for theoretical ideas that can unify our understanding of brain structure and function. Theory is indispensable, as it allows us to transform big data into ‘small data’ and, ultimately, knowledge—delivering compact descriptions of regularities, principles and laws that apply to the architecture and functioning of neural systems. We believe that network neuroscience can make an important contribution toward unifying an otherwise fractured discipline by providing a common conceptual framework and a common toolset to meet the challenges of modern neuroscience. Network neuroscience naturally connects with other important theoretical approaches such as dynamical systems, neural coding and statistical physics.
Effectively bridging network theory and empirical neuroscience will require new educational initiatives and training programs that provide strong grounding in both disciplines. If these goals can be accomplished, the network neuroscientist of the future will be in an excellent position to add to our understanding of the network mechanisms that underpin brain structure and function.
Acknowledgments
The authors gratefully acknowledge helpful comments by A. Avena-Koenigsberger, R. Betzel, L. Chai and G. Rosenthal. D.S.B. acknowledges support from the John D. and Catherine T. MacArthur Foundation, the Alfred P. Sloan Foundation, the National Science Foundation (BCS-1430087, NCS BCS-1631550, CAREER PHY-1554488) and the US National Institutes of Health (R01-HD086888, R21-M MH-106799, R01NS099348). O.S. acknowledges support from the Indiana Clinical Translational Sciences Institute (NIH UL1TR0011808), the J.S. McDonnell Foundation (220020387), the National Science Foundation (1636892) and the US National Institutes of Health (R01-AT009036, R01-B022574 and P30-AG010133).
Footnotes
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
References
- 1.Sejnowski TJ, Churchland PS, Movshon JA. Putting big data to good use in neuroscience. Nat Neurosci. 2014;17:1440–1441. doi: 10.1038/nn.3839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jorgenson LA, et al. The BRAIN Initiative: developing technology to catalyse neuroscience discovery. Phil Trans R Soc B. 2015;370:20140164. doi: 10.1098/rstb.2014.0164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bullmore E, Sporns O. Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci. 2009;10:186–198. doi: 10.1038/nrn2575. [DOI] [PubMed] [Google Scholar]
- 4.Sporns O. Contributions and challenges for network models in cognitive neuroscience. Nat Neurosci. 2014;17:652–660. doi: 10.1038/nn.3690. [DOI] [PubMed] [Google Scholar]
- 5.Medaglia JD, Lynall ME, Bassett DS. Cognitive network neuroscience. J Cogn Neurosci. 2015;27:1471–1491. doi: 10.1162/jocn_a_00810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sporns O. Networks of the Brain. MIT Press; 2010. [Google Scholar]
- 7.Cunningham JP, Yu BM. Dimensionality reduction for large-scale neural recordings. Nat Neurosci. 2014;17:1500–1509. doi: 10.1038/nn.3776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Freeman J, et al. Mapping brain activity at scale with cluster computing. Nat Methods. 2014;11:941–950. doi: 10.1038/nmeth.3041. [DOI] [PubMed] [Google Scholar]
- 9.Poldrack RA, Gorgolewski KJ. Making big data open: data sharing in neuroimaging. Nat Neurosci. 2014;17:1510–1517. doi: 10.1038/nn.3818. [DOI] [PubMed] [Google Scholar]
- 10.White JG, Southgate E, Thomson JN, Brenner S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci. 1986;314:1–340. doi: 10.1098/rstb.1986.0056. [DOI] [PubMed] [Google Scholar]
- 11.Bentley B, et al. The multilayer connectome of Caenorhabditis elegans. PLoS Comput Biol. 2016;12:e1005283. doi: 10.1371/journal.pcbi.1005283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jarrell TA, et al. The connectome of a decision-making neural network. Science. 2012;337:437–444. doi: 10.1126/science.1221762. [DOI] [PubMed] [Google Scholar]
- 13.Takemura SY, et al. A visual motion detection circuit suggested by Drosophila connectomics. Nature. 2013;500:175–181. doi: 10.1038/nature12450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lichtman JW, Denk W. The big and the small: challenges of imaging the brain’s circuits. Science. 2011;334:618–623. doi: 10.1126/science.1209168. [DOI] [PubMed] [Google Scholar]
- 15.Helmstaedter M, et al. Connectomic reconstruction of the inner plexiform layer in the mouse retina. Nature. 2013;500:168–174. doi: 10.1038/nature12346. [DOI] [PubMed] [Google Scholar]
- 16.Kebschull JM, et al. High-throughput mapping of single-neuron projections by sequencing of barcoded RNA. Neuron. 2016;91:975–987. doi: 10.1016/j.neuron.2016.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shih CT, et al. Connectomics-based analysis of information flow in the Drosophila brain. Curr Biol. 2015;25:1249–1258. doi: 10.1016/j.cub.2015.03.021. [DOI] [PubMed] [Google Scholar]
- 18.Oh SW, et al. A mesoscale connectome of the mouse brain. Nature. 2014;508:207–214. doi: 10.1038/nature13186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bota M, Sporns O, Swanson LW. Architecture of the cerebral cortical association connectome underlying cognition. Proc Natl Acad Sci USA. 2015;112:E2093–E2101. doi: 10.1073/pnas.1504394112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stephan KE, et al. Advanced database methodology for the Collation of Connectivity data on the Macaque brain (CoCoMac) Phil Trans R Soc B. 2001;356:1159–1186. doi: 10.1098/rstb.2001.0908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Markov NT, et al. A weighted and directed interareal connectivity matrix for macaque cerebral cortex. Cereb Cortex. 2014;24:17–36. doi: 10.1093/cercor/bhs270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jbabdi S, Sotiropoulos SN, Haber SN, Van Essen DC, Behrens TE. Measuring macroscopic brain connections in vivo. Nat Neurosci. 2015;18:1546–1555. doi: 10.1038/nn.4134. [DOI] [PubMed] [Google Scholar]
- 23.Thomas C, et al. Anatomical accuracy of brain connections derived from diffusion MRI tractography is inherently limited. Proc Natl Acad Sci USA. 2014;111:16574–16579. doi: 10.1073/pnas.1405672111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jones DK, Knösche TR, Turner R. White matter integrity, fiber count, and other fallacies: the do’s and don’ts of diffusion MRI. Neuroimage. 2013;73:239–254. doi: 10.1016/j.neuroimage.2012.06.081. [DOI] [PubMed] [Google Scholar]
- 25.Donahue CJ, et al. Using diffusion tractography to predict cortical connection strength and distance: a quantitative comparison with tracers in the monkey. J Neurosci. 2016;36:6758–6770. doi: 10.1523/JNEUROSCI.0493-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hamel EJ, Grewe BF, Parker JG, Schnitzer MJ. Cellular level brain imaging in behaving mammals: an engineering approach. Neuron. 2015;86:140–159. doi: 10.1016/j.neuron.2015.03.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Keller PJ, Ahrens MB. Visualizing whole-brain activity and development at the single-cell level using light-sheet microscopy. Neuron. 2015;85:462–483. doi: 10.1016/j.neuron.2014.12.039. [DOI] [PubMed] [Google Scholar]
- 28.Power JD, et al. Functional network organization of the human brain. Neuron. 2011;72:665–678. doi: 10.1016/j.neuron.2011.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cole MW, Bassett DS, Power JD, Braver TS, Petersen SE. Intrinsic and task-evoked network architectures of the human brain. Neuron. 2014;83:238–251. doi: 10.1016/j.neuron.2014.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rosenthal G, Sporns O, Avidan G. Stimulus dependent dynamic reorganization of the human face processing network. Cereb Cortex. 2016 doi: 10.1093/cercor/bhw279. http://dx.doi.org/10.1093/cercor/bhw279. [DOI] [PMC free article] [PubMed]
- 31.Mišić B, Sporns O. From regions to connections and networks: new bridges between brain and behavior. Curr Opin Neurobiol. 2016;40:1–7. doi: 10.1016/j.conb.2016.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Vogelstein JT, et al. Discovery of brainwide neural-behavioral maps via multiscale unsupervised structure learning. Science. 2014;344:386–392. doi: 10.1126/science.1250298. [DOI] [PubMed] [Google Scholar]
- 33.Crossley NA, et al. Cognitive relevance of the community structure of the human brain functional coactivation network. Proc Natl Acad Sci USA. 2013;110:11583–11588. doi: 10.1073/pnas.1220826110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Izquierdo EJ, Beer RD. Connecting a connectome to behavior: an ensemble of neuroanatomical models of C. elegans klinotaxis. PLoS Comput Biol. 2013;9:e1002890. doi: 10.1371/journal.pcbi.1002890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fornito A, Bullmore ET. Connectomic intermediate phenotypes for psychiatric disorders. Front Psychiatry. 2012;3:32. doi: 10.3389/fpsyt.2012.00032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ideker T, Galitski T, Hood L. A new approach to decoding life: systems biology. Annu Rev Genomics Hum Genet. 2001;2:343–372. doi: 10.1146/annurev.genom.2.1.343. [DOI] [PubMed] [Google Scholar]
- 37.Barabási AL, Oltvai ZN. Network biology: understanding the cell’s functional organization. Nat Rev Genet. 2004;5:101–113. doi: 10.1038/nrg1272. [DOI] [PubMed] [Google Scholar]
- 38.Vidal M, Cusick ME, Barabási AL. Interactome networks and human disease. Cell. 2011;144:986–998. doi: 10.1016/j.cell.2011.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Geschwind DH, Flint J. Genetics and genomics of psychiatric disease. Science. 2015;349:1489–1494. doi: 10.1126/science.aaa8954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Barabási AL, Gulbahce N, Loscalzo J. Network medicine: a network-based approach to human disease. Nat Rev Genet. 2011;12:56–68. doi: 10.1038/nrg2918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fornito A, Zalesky A, Pantelis C, Bullmore ET. Schizophrenia, neuroimaging and connectomics. Neuroimage. 2012;62:2296–2314. doi: 10.1016/j.neuroimage.2011.12.090. [DOI] [PubMed] [Google Scholar]
- 42.de la Torre-Ubieta L, Won H, Stein JL, Geschwind DH. Advancing the understanding of autism disease mechanisms through genetics. Nat Med. 2016;22:345–361. doi: 10.1038/nm.4071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lazer D, et al. Life in the network: the coming age of computational social science. Science. 2009;323:721–723. doi: 10.1126/science.1167742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Onnela JP, Rauch SL. Harnessing smartphone-based digital phenotyping to enhance behavioral and mental health. Neuropsychopharmacology. 2016;41:1691–1696. doi: 10.1038/npp.2016.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Newman M. Networks: An Introduction. Oxford University Press; 2010. [Google Scholar]
- 46.Fornito A, Zalesky A, Bullmore E. Fundamentals of Brain Network Analysis. Academic Press; 2016. [Google Scholar]
- 47.Bassett DS, Bullmore ET. Small-world brain networks revisited. The Neuroscientist. 2016 doi: 10.1177/1073858416667720. http://dx.doi.org/10.1177%2F1073858416667720. [DOI] [PMC free article] [PubMed]
- 48.van den Heuvel MP, Sporns O. Network hubs in the human brain. Trends Cogn Sci. 2013;17:683–696. doi: 10.1016/j.tics.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 49.Zamora-López G, Zhou C, Kurths J. Cortical hubs form a module for multisensory integration on top of the hierarchy of cortical networks. Front Neuroinform. 2010;4:1. doi: 10.3389/neuro.11.001.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.van den Heuvel MP, Sporns O. Rich-club organization of the human connectome. J Neurosci. 2011;31:15775–15786. doi: 10.1523/JNEUROSCI.3539-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Markov NT, et al. Cortical high-density counterstream architectures. Science. 2013;342:1238406. doi: 10.1126/science.1238406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chaudhuri R, Knoblauch K, Gariel MA, Kennedy H, Wang XJ. A large-scale circuit mechanism for hierarchical dynamical processing in the primate cortex. Neuron. 2015;88:419–431. doi: 10.1016/j.neuron.2015.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Horvát S, et al. Spatial embedding and wiring cost constrain the functional layout of the cortical network of rodents and primates. PLoS Biol. 2016;14:e1002512. doi: 10.1371/journal.pbio.1002512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Betzel RF, et al. The modular organization of human anatomical brain networks: accounting for the cost of wiring. Network Neurosci. 2017 doi: 10.1162/NETN_a_00002. http://doi.org/10.1162/NETN_a_00002. [DOI] [PMC free article] [PubMed]
- 55.Fornito A, Zalesky A, Breakspear M. Graph analysis of the human connectome: promise, progress, and pitfalls. Neuroimage. 2013;80:426–444. doi: 10.1016/j.neuroimage.2013.04.087. [DOI] [PubMed] [Google Scholar]
- 56.Sporns O, Betzel RF. Modular brain networks. Annu Rev Psychol. 2016;67:613–640. doi: 10.1146/annurev-psych-122414-033634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hinne M, Heskes T, Beckmann CF, van Gerven MA. Bayesian inference of structural brain networks. Neuroimage. 2013;66:543–552. doi: 10.1016/j.neuroimage.2012.09.068. [DOI] [PubMed] [Google Scholar]
- 58.Zalesky A, Fornito A, Bullmore ET. Network-based statistic: identifying differences in brain networks. Neuroimage. 2010;53:1197–1207. doi: 10.1016/j.neuroimage.2010.06.041. [DOI] [PubMed] [Google Scholar]
- 59.Mucha PJ, Richardson T, Macon K, Porter MA, Onnela JP. Community structure in time-dependent, multiscale, and multiplex networks. Science. 2010;328:876–878. doi: 10.1126/science.1184819. [DOI] [PubMed] [Google Scholar]
- 60.Rosvall M, Bergstrom CT. An information-theoretic framework for resolving community structure in complex networks. Proc Natl Acad Sci USA. 2007;104:7327–7331. doi: 10.1073/pnas.0611034104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Betzel RF, et al. Generative models of the human connectome. Neuroimage. 2016;124(Pt A):1054–1064. doi: 10.1016/j.neuroimage.2015.09.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Avena-Koenigsberger A, Goñi J, Solé R, Sporns O. Network morphospace. J R Soc Interface. 2015;12:20140881. doi: 10.1098/rsif.2014.0881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Giusti C, Ghrist R, Bassett DS. Two’s company, three (or more) is a simplex: Algebraic-topological tools for understanding higher-order structure in neural data. 2016 doi: 10.1007/s10827-016-0608-6. Preprint at https://arxiv.org/abs/1601.01704. [DOI] [PMC free article] [PubMed]
- 64.Courtney OT, Bianconi G. Generalized network structures: The configuration model and the canonical ensemble of simplicial complexes. 2016 doi: 10.1103/PhysRevE.93.062311. Preprint at https://arxiv.org/abs/1602.04110. [DOI] [PubMed]
- 65.Sizemore A, Giusti C, Bassett D. Classification of weighted networks through mesoscale homological features. J Complex Netw. 2015 http://dx.doi.org/10.1093/comnet/cnw013.
- 66.Bassett DS, Wymbs NF, Porter MA, Mucha PJ, Grafton ST. Cross-linked structure of network evolution. Chaos. 2014;24:013112. doi: 10.1063/1.4858457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Dotko P, et al. Topological analysis of the connectome of digital reconstructions of neural microcircuits. 2016 Preprint at https://arxiv.org/abs/1601.01580.
- 68.Sizemore A, Giusti C, Betzel RF, Bassett DS. Closures and Cavities in the Human Connectome. 2016 doi: 10.1007/s10827-017-0672-6. Preprint at https://arxiv.org/abs/1608.03520. [DOI] [PMC free article] [PubMed]
- 69.Zalesky A, Fornito A, Bullmore E. On the use of correlation as a measure of network connectivity. Neuroimage. 2012;60:2096–2106. doi: 10.1016/j.neuroimage.2012.02.001. [DOI] [PubMed] [Google Scholar]
- 70.Nigam S, et al. Rich-club organization in effective connectivity among cortical neurons. J Neurosci. 2016;36:670–684. doi: 10.1523/JNEUROSCI.2177-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Friston KJ, Li B, Daunizeau J, Stephan KE. Network discovery with DCM. Neuroimage. 2011;56:1202–1221. doi: 10.1016/j.neuroimage.2010.12.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Jirsa VK, Sporns O, Breakspear M, Deco G, McIntosh AR. Towards the virtual brain: network modeling of the intact and the damaged brain. Arch Ital Biol. 2010;148:189–205. [PubMed] [Google Scholar]
- 73.Hines ML, Carnevale NT. The NEURON simulation environment. Neural Comput. 1997;9:1179–1209. doi: 10.1162/neco.1997.9.6.1179. [DOI] [PubMed] [Google Scholar]
- 74.Szigeti B, et al. OpenWorm: an open-science approach to modeling Caenorhabditis elegans. Front Comput Neurosci. 2014;8:137. doi: 10.3389/fncom.2014.00137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Stephan KE, Iglesias S, Heinzle J, Diaconescu AO. Translational perspectives for computational neuroimaging. Neuron. 2015;87:716–732. doi: 10.1016/j.neuron.2015.07.008. [DOI] [PubMed] [Google Scholar]
- 76.Karlebach G, Shamir R. Modelling and analysis of gene regulatory networks. Nat Rev Mol Cell Biol. 2008;9:770–780. doi: 10.1038/nrm2503. [DOI] [PubMed] [Google Scholar]
- 77.Karr JR, et al. A whole-cell computational model predicts phenotype from genotype. Cell. 2012;150:389–401. doi: 10.1016/j.cell.2012.05.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Castellano C, Fortunato S, Loreto V. Statistical physics of social dynamics. Rev Mod Phys. 2009;81:591. [Google Scholar]
- 79.Holme P. Temporal networks. Phys Rep. 2012;519:97–125. [Google Scholar]
- 80.Honey CJ, Kötter R, Breakspear M, Sporns O. Network structure of cerebral cortex shapes functional connectivity on multiple time scales. Proc Natl Acad Sci USA. 2007;104:10240–10245. doi: 10.1073/pnas.0701519104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Goñi J, et al. Resting-brain functional connectivity predicted by analytic measures of network communication. Proc Natl Acad Sci USA. 2014;111:833–838. doi: 10.1073/pnas.1315529111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Deco G, Jirsa VK, Robinson PA, Breakspear M, Friston K. The dynamic brain: from spiking neurons to neural masses and cortical fields. PLoS Comput Biol. 2008;4:e1000092. doi: 10.1371/journal.pcbi.1000092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Raj A, Kuceyeski A, Weiner M. A network diffusion model of disease progression in dementia. Neuron. 2012;73:1204–1215. doi: 10.1016/j.neuron.2011.12.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mišić B, Sporns O, McIntosh AR. Communication efficiency and congestion of signal traffic in large-scale brain networks. PLoS Comput Biol. 2014;10:e1003427. doi: 10.1371/journal.pcbi.1003427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Hutchison RM, et al. Dynamic functional connectivity: promise, issues, and interpretations. Neuroimage. 2013;80:360–378. doi: 10.1016/j.neuroimage.2013.05.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Conaco C, et al. Functionalization of a protosynaptic gene expression network. Proc Natl Acad Sci USA. 2012;109(Suppl. 1):10612–10618. doi: 10.1073/pnas.1201890109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Beagan JA, et al. Local genome topology can exhibit an incompletely rewired 3D-folding state during somatic cell reprogramming. Cell Stem Cell. 2016;18:611–624. doi: 10.1016/j.stem.2016.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Kivelä M, et al. Multilayer networks. J Compl Netw. 2014;2:203–271. [Google Scholar]
- 89.Betzel RF, et al. Functional brain modules reconfigure at multiple scales across the human lifespan. 2015 Preprint at https://arxiv.org/abs/1510.08045.
- 90.Bassett DS, Yang M, Wymbs NF, Grafton ST. Learning-induced autonomy of sensorimotor systems. Nat Neurosci. 2015;18:744–751. doi: 10.1038/nn.3993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.De Domenico M, Sasai S, Arenas A. Mapping multiplex hubs in human functional brain networks. Front Neurosci. 2016;10:326. doi: 10.3389/fnins.2016.00326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Calhoun VD, Miller R, Pearlson G, Adali T. The chronnectome: time-varying connectivity networks as the next frontier in fMRI data discovery. Neuron. 2014;84:262–274. doi: 10.1016/j.neuron.2014.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Chai L, et al. Evolution of brain network dynamics in neurodevelopment. Netw Neurosci. 2017 doi: 10.1162/NETN_a_00001. http://doi.org/10.1162/NETN_a_00001. [DOI] [PMC free article] [PubMed]
- 94.Kopell NJ, Gritton HJ, Whittington MA, Kramer MA. Beyond the connectome: the dynome. Neuron. 2014;83:1319–1328. doi: 10.1016/j.neuron.2014.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.O’Donnell MB, Falk EB. Big data under the microscope and brains in social context integrating methods from computational social science and neuroscience. Ann Am Acad Pol Soc Sci. 2015;659:274–289. [Google Scholar]
- 96.Hasson U, Frith CD. Mirroring and beyond: coupled dynamics as a generalized framework for modelling social interactions. Phil Trans R Soc B. 2016;371:20150366. doi: 10.1098/rstb.2015.0366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Proulx SR, Promislow DE, Phillips PC. Network thinking in ecology and evolution. Trends Ecol Evol. 2005;20:345–353. doi: 10.1016/j.tree.2005.04.004. [DOI] [PubMed] [Google Scholar]
- 98.Sacchet MD, Prasad G, Foland-Ross LC, Thompson PM, Gotlib IH. Support vector machine classification of major depressive disorder using diffusion-weighted neuroimaging and graph theory. Front Psychiatry. 2015;6:21. doi: 10.3389/fpsyt.2015.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Albert R, Thakar J. Boolean modeling: a logic-based dynamic approach for understanding signaling and regulatory networks and for making useful predictions. Wiley Interdiscip Rev Syst Biol Med. 2014;6:353–369. doi: 10.1002/wsbm.1273. [DOI] [PubMed] [Google Scholar]
- 100.Wendling F, Benquet P, Bartolomei F, Jirsa V. Computational models of epileptiform activity. J Neurosci Methods. 2016;260:233–251. doi: 10.1016/j.jneumeth.2015.03.027. [DOI] [PubMed] [Google Scholar]
- 101.Watanabe T, Masuda N, Megumi F, Kanai R, Rees G. Energy landscape and dynamics of brain activity during human bistable perception. Nat Commun. 2014;5:4765. doi: 10.1038/ncomms5765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hermundstad AM, et al. Structurally constrained relationships between cognitive states in the human brain. PLoS Comput Biol. 2014;10:e1003591. doi: 10.1371/journal.pcbi.1003591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Harrington DL, et al. Network topology and functional connectivity disturbances precede the onset of Huntington’s disease. Brain. 2015;138:2332–2346. doi: 10.1093/brain/awv145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Falk EB, et al. Self-affirmation alters the brain’s response to health messages and subsequent behavior change. Proc Natl Acad Sci USA. 2015;112:1977–1982. doi: 10.1073/pnas.1500247112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Gratton C, Lee TG, Nomura EM, D’Esposito M. The effect of theta-burst TMS on cognitive control networks measured with resting state fMRI. Front Syst Neurosci. 2013;7:124. doi: 10.3389/fnsys.2013.00124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Crofts JJ, et al. Network analysis detects changes in the contralesional hemisphere following stroke. Neuroimage. 2011;54:161–169. doi: 10.1016/j.neuroimage.2010.08.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Ramsey LE, et al. Normalization of network connectivity in hemispatial neglect recovery. Ann Neurol. 2016;80:127–141. doi: 10.1002/ana.24690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lewis PM, Thomson RH, Rosenfeld JV, Fitzgerald PB. Brain neuromodulation techniques: a review. Neuroscientist. 2016;22:406–421. doi: 10.1177/1073858416646707. [DOI] [PubMed] [Google Scholar]
- 109.Johnson MD, et al. Neuromodulation for brain disorders: challenges and opportunities. IEEE Trans Biomed Eng. 2013;60:610–624. doi: 10.1109/TBME.2013.2244890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kailath T. Linear Systems. Prentice Hall; 1979. [Google Scholar]
- 111.Liu YY, Slotine JJ, Barabási AL. Controllability of complex networks. Nature. 2011;473:167–173. doi: 10.1038/nature10011. [DOI] [PubMed] [Google Scholar]
- 112.Pasqualetti F, Zampieri S, Bullo F. Controllability metrics, limitations and algorithms for complex networks. IEEE Trans Contr Netw Syst. 2014;1:40–52. [Google Scholar]
- 113.Gu S, et al. Controllability of structural brain networks. Nat Commun. 2015;6:8414. doi: 10.1038/ncomms9414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Santaniello S, et al. Therapeutic mechanisms of high-frequency stimulation in Parkinson’s disease and neural restoration via loop-based reinforcement. Proc Natl Acad Sci USA. 2015;112:E586–E595. doi: 10.1073/pnas.1406549111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Ching S, et al. Real-time closed-loop control in a rodent model of medically induced coma using burst suppression. Anesthesiology. 2013;119:848–860. doi: 10.1097/ALN.0b013e31829d4ab4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Khambhati AN, Davis KA, Lucas TH, Litt B, Bassett DS. Virtual cortical resection reveals push-pull network control preceding seizure evolution. Neuron. 2016;91:1170–1182. doi: 10.1016/j.neuron.2016.07.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Wheeler DW, et al. Hippocampome.org: a knowledge base of neuron types in the rodent hippocampus. eLife. 2015;4:e09960. doi: 10.7554/eLife.09960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Markram H, et al. Reconstruction and simulation of neocortical microcircuitry. Cell. 2015;163:456–492. doi: 10.1016/j.cell.2015.09.029. [DOI] [PubMed] [Google Scholar]
- 119.Schneider CJ, Bezaire M, Soltesz I. Toward a full-scale computational model of the rat dentate gyrus. Front Neural Circuits. 2012;6:83. doi: 10.3389/fncir.2012.00083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.van den Heuvel MP, Bullmore ET, Sporns O. Comparative Connectomics. Trends Cogn Sci. 2016;20:345–361. doi: 10.1016/j.tics.2016.03.001. [DOI] [PubMed] [Google Scholar]
- 121.Goulas A, et al. Comparative analysis of the macroscale structural connectivity in the macaque and human brain. PLoS Comput Biol. 2014;10:e1003529. doi: 10.1371/journal.pcbi.1003529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Li L, et al. Mapping putative hubs in human, chimpanzee and rhesus macaque connectomes via diffusion tractography. Neuroimage. 2013;80:462–474. doi: 10.1016/j.neuroimage.2013.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Battiston F, Nicosia V, Chavez M, Latora V. Multilayer motif analysis of brain networks. 2016 doi: 10.1063/1.4979282. Preprint at https://arxiv.org/abs/1606.09115. [DOI] [PubMed]
- 124.Wolf L, Goldberg C, Manor N, Sharan R, Ruppin E. Gene expression in the rodent brain is associated with its regional connectivity. PLoS Comput Biol. 2011;7:e1002040. doi: 10.1371/journal.pcbi.1002040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.French L, Pavlidis P. Relationships between gene expression and brain wiring in the adult rodent brain. PLoS Comput Biol. 2011;7:e1001049. doi: 10.1371/journal.pcbi.1001049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Richiardi J, et al. BRAIN NETWORKS. Correlated gene expression supports synchronous activity in brain networks. Science. 2015;348:1241–1244. doi: 10.1126/science.1255905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Wang GZ, et al. Correspondence between resting-state activity and brain gene expression. Neuron. 2015;88:659–666. doi: 10.1016/j.neuron.2015.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Fulcher BD, Fornito A. A transcriptional signature of hub connectivity in the mouse connectome. Proc Natl Acad Sci USA. 2016;113:1435–1440. doi: 10.1073/pnas.1513302113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Rubinov M, Ypma RJ, Watson C, Bullmore ET. Wiring cost and topological participation of the mouse brain connectome. Proc Natl Acad Sci USA. 2015;112:10032–10037. doi: 10.1073/pnas.1420315112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Uddin LQ. Idiosyncratic connectivity in autism: developmental and anatomical considerations. Trends Neurosci. 2015;38:261–263. doi: 10.1016/j.tins.2015.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hernandez LM, Rudie JD, Green SA, Bookheimer S, Dapretto M. Neural signatures of autism spectrum disorders: insights into brain network dynamics. Neuropsychopharmacology. 2015;40:171–189. doi: 10.1038/npp.2014.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Kuiper JS, et al. Social relationships and cognitive decline: a systematic review and meta-analysis of longitudinal cohort studies. Int J Epidemiol. 2016;45:1169–1206. doi: 10.1093/ije/dyw089. [DOI] [PubMed] [Google Scholar]
- 133.Mier D, Kirsch P. Social-cognitive deficits in schizophrenia. Curr Top Behav Neurosci. 2016 doi: 10.1007/7854_2015_427. https://dx.doi.org/10.1007/7854_2015_427. [DOI] [PubMed]
- 134.Tost H, Champagne FA, Meyer-Lindenberg A. Environmental influence in the brain, human welfare and mental health. Nat Neurosci. 2015;18:1421–1431. doi: 10.1038/nn.4108. [DOI] [PubMed] [Google Scholar]
- 135.Byrge L, Sporns O, Smith LB. Developmental process emerges from extended brain-body-behavior networks. Trends Cogn Sci. 2014;18:395–403. doi: 10.1016/j.tics.2014.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Gibson G. The environmental contribution to gene expression profiles. Nat Rev Genet. 2008;9:575–581. doi: 10.1038/nrg2383. [DOI] [PubMed] [Google Scholar]
- 137.Pescosolido B, et al. The social symbiome framework: linking genes-to-global cultures in public health using network science. In: Neal ZP, editor. Handbook of Applied Systems Science. Routledge; 2015. pp. 25–48. [Google Scholar]
- 138.Murphy AC, et al. Explicitly linking regional activation and function connectivity: community structure of weighted networks with continuous annotation. 2016 Preprint at https://arxiv.org/abs/1611.07962.
- 139.Ganmor E, Segev R, Schneidman E. Sparse low-order interaction network underlies a highly correlated and learnable neural population code. Proc Natl Acad Sci USA. 2011;108:9679–9684. doi: 10.1073/pnas.1019641108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Giusti C, Pastalkova E, Curto C, Itskov V. Clique topology reveals intrinsic geometric structure in neural correlations. Proc Natl Acad Sci USA. 2015;112:13455–13460. doi: 10.1073/pnas.1506407112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bassett DS, et al. Dynamic reconfiguration of human brain networks during learning. Proc Natl Acad Sci USA. 2011;108:7641–7646. doi: 10.1073/pnas.1018985108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Braun U, et al. Dynamic reconfiguration of frontal brain networks during executive cognition in humans. Proc Natl Acad Sci USA. 2015;112:11678–11683. doi: 10.1073/pnas.1422487112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Betzel RF, Gu S, Medaglia JD, Pasqualetti F, Bassett DS. Optimally controlling the human connectome: the role of network topology. Sci Rep. 2016;6:30770. doi: 10.1038/srep30770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Muldoon SF, et al. Stimulation-based control of dynamic brain networks. PLoS Comput Biol. 2016;12:e1005076. doi: 10.1371/journal.pcbi.1005076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Muldoon SF, et al. GABAergic inhibition shapes interictal dynamics in awake epileptic mice. Brain. 2015;138:2875–2890. doi: 10.1093/brain/awv227. [DOI] [PubMed] [Google Scholar]
- 146.Feldt Muldoon S, Soltesz I, Cossart R. Spatially clustered neuronal assemblies comprise the microstructure of synchrony in chronically epileptic networks. Proc Natl Acad Sci USA. 2013;110:3567–3572. doi: 10.1073/pnas.1216958110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Burns SP, et al. Network dynamics of the brain and influence of the epileptic seizure onset zone. Proc Natl Acad Sci USA. 2014;111:E5321–E5330. doi: 10.1073/pnas.1401752111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ching S, Brown EN, Kramer MA. Distributed control in a mean-field cortical network model: implications for seizure suppression. Phys Rev E. 2012;86:021920. doi: 10.1103/PhysRevE.86.021920. [DOI] [PubMed] [Google Scholar]
- 149.Simony E, et al. Dynamic reconfiguration of the default mode network during narrative comprehension. Nat Commun. 2016;7:12141. doi: 10.1038/ncomms12141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Schmaelzle R, et al. Brain connectivity dynamics during social interaction reflect social network structure. 2017 doi: 10.1073/pnas.1616130114. Preprint at https://doi.org/10.1101/096420. [DOI] [PMC free article] [PubMed]