

Abstract
Alcohol consumption and stress increase brain levels of known innate immune signaling molecules. Microglia, the innate immune cells of the brain, and neurons respond to alcohol, signaling through Toll-like receptors (TLRs), high-mobility group box 1 (HMGB1), miRNAs, pro-inflammatory cytokines and their associated receptors involved in signaling between microglia, other glia and neurons. Repeated cycles of alcohol and stress cause a progressive, persistent induction of HMGB1, miRNA and TLR receptors in brain that appear to underlie the progressive and persistent loss of behavioral control, increased impulsivity and anxiety, as well as craving, coupled with increasing ventral striatal responses that promote reward seeking behavior and increase risk of developing alcohol use disorders. Studies employing anti-oxidant, anti-inflammatory, anti-depressant, and innate immune antagonists further link innate immune gene expression to addiction-like behaviors. Innate immune molecules are novel targets for addiction and affective disorders therapies.
Keywords: HMGB1, TLR, cytokines, miRNA-Let-7, alcohol, addiction
1. Introduction
Neuroimmune signaling is emerging as a contributor to the development of alcohol use disorders. Alcohol consumption and stress increase brain levels of known innate immune signaling molecules. Microglia, the innate immune cells of the brain, play an important role in brain development and function. As mesodermal tissue specific monocytes, microglia express signaling molecules first characterized as “acute phase proteins” that increase in blood rapidly in response to infections. More recently, Toll-like receptors (TLRs), members of a superfamily of receptors, i.e. - interleukin-1 receptor / toll-like receptor superfamily, were found to initiate pro-inflammatory responses to infectious agents by sensing large molecules with lipid, sugar, protein and nucleic acid components. Pro-inflammatory cytokines and their associated receptors share complex signaling pathways with TLRs that converge upon NF-κB, a key innate immune transcription factor in monocyte-like cells, and other transcription factors regulated by TLR-kinase cascades. Recent studies in brain, which is generally sterile, indicate endogenous TLR agonists, i.e. danger-associated molecular patterns (DAMPs), represent important signaling systems between microglia, other glia and neurons. Further, emerging studies find cytokines and DAMPs may signal directly between neurons. High-mobility group box 1 (HMGB1) is a cytokine-like molecule highly expressed in neurons that can directly activate TLRs as well as enhancing responses at other pro-inflammatory receptors. Alcohol- and stress-induced increases in Toll-like receptors, and other neuroimmune signaling molecules in brain sensitize the brain to further innate immune gene induction, resulting in progressive and persistent increases in HMGB1 and TLR signaling as evidenced by prefrontal cortical expression levels correlating with lifetime alcohol consumption across alcoholic and moderate drinking controls (Crews et al., 2013a). Mechanistic studies of epilepsy find neuronal firing results in the rapid release of HMGB1 and IL-1β preceding the seizure, inducing changes in excitatory receptors that sensitize neurons to excitation, lowering seizure threshold and contributing to the “kindling” progressive increase in seizure frequency and severity with each event (Maroso et al., 2011). Although different brain regions and circuits distinguish addiction from epilepsy, HMGB1 and TLR signaling and cycles of stress and drug abuse appear to share the common mechanisms of sensitization of circuitry through increased HMGB1 and TLR signaling.
Alcohol administration to mice and rats increases expression of HMGB1 and multiple TLR receptors in brain that persist for long periods of abstinence. Furthermore, human post-mortem alcoholic brains show increased expression of microglial markers, cytokines, TLR receptors and HMGB1, with the latter two being correlated with lifetime alcohol consumption. Repeated alcohol, drug use and stress in rodents sensitize microglia toward hyperactivity, likely through induction of immune signaling (see Sections 2 and 5). Cytokine infusions into specific rodent brain regions initiate craving and alcohol consumption, and human alcoholics have increased plasma pro-inflammatory cytokines that correlate with alcohol craving and severity of dependence (see Section 5). Further, pro-inflammatory cytokines TNFα, IL-6, and IL-1β play critical roles in the pathologies of mood disorders (Bhattacharya et al., 2016; Bhattacharya and Drevets, 2017). These observations, detailed further below, support the hypothesis that stress and alcohol induction of neuroimmune signaling contribute to the progressive, persistent increase in craving and mood dysfunction in addiction. Recent discoveries indicate broad induction of TLR receptors in brain as well as endogenous agonists for TLR receptors, including miRNA that activate nucleic acid sensing TLRs and HMGB1 that activates TLR4, a key signaling TLR in the brain. Although poorly understood, studies suggest that activation and escalated signaling of this system leads to progressive and persistent loss of behavioral control, increased impulsivity and anxiety, as well as negative affect and craving coupled with increasing ventral striatal responses that promote reward seeking behavior and increase risk of developing alcohol use disorders. Studies employing anti-oxidant, anti-inflammatory, anti-depressant, and innate immune antagonists further link innate immune gene expression to addiction-like behaviors. In this review we discuss both preclinical rodent and human studies that examine the role of neuroimmune signaling in alcoholism. Unless otherwise noted the presented studies were performed in rodents. From these studies, we surmise that innate immune molecules are novel targets for addiction and affective disorders therapies.
2. Alcohol and Stress, Microglia and Neuroimmune Signaling
Repeated cycles of alcohol, drugs of abuse and stress interact, contributing to a progressive sequence of stages leading to addiction (Volkow et al., 2016). Alcohol, drugs of abuse and stress also trigger neuroinflammatory responses. Inflammation may have beneficial or maladaptive consequences on brain function (Fig 1). Physical stressors, such as injury and infection, increase pro-inflammatory cytokines and other leukocyte derived proteins and prostaglandins as part of the acute inflammatory phase response. Stressor-induced inflammation also triggers adaptive behavioral changes known as “sickness behavior.” Sickness behavior includes social withdrawal, decreased activity, somnolence/sleepiness, anhedonia, etc. that help to conserve energy and facilitate recovery (Dantzer et al., 2008). Interestingly, even psychological stressors (e.g. – social stress) can cause inflammation (Steptoe et al., 2007). The acute pro-inflammatory response, often referred to as Th1, includes signals that activate a delayed wound healing and cell growth through “anti-inflammatory” Th2 innate immune signaling molecules. While the benefit of the Th1 to Th2 progression following trauma or infection may be apparent, what happens in brain is poorly understood. Therefore, both physical and psychological stressors may enhance inflammation, initiating molecular and behavioral changes that facilitate recovery. While stress and inflammation can lead to the adaptive response of sickness behavior, intense or chronic stress and inflammation can become maladaptive and lead to neuropsychiatric disease such as depression. Indeed, the similarities between sickness behavior and depression have been noted (Maes et al., 2012). Models of depression include endotoxin treatments of rodents followed beyond the 24 hour “sickness behavior” time course (Dantzer et al., 2008). The inflammation caused by excessive alcohol consumption may have similar maladaptive consequences.
Figure 1. Immune Activation and the Development of Substance Use Disorders.
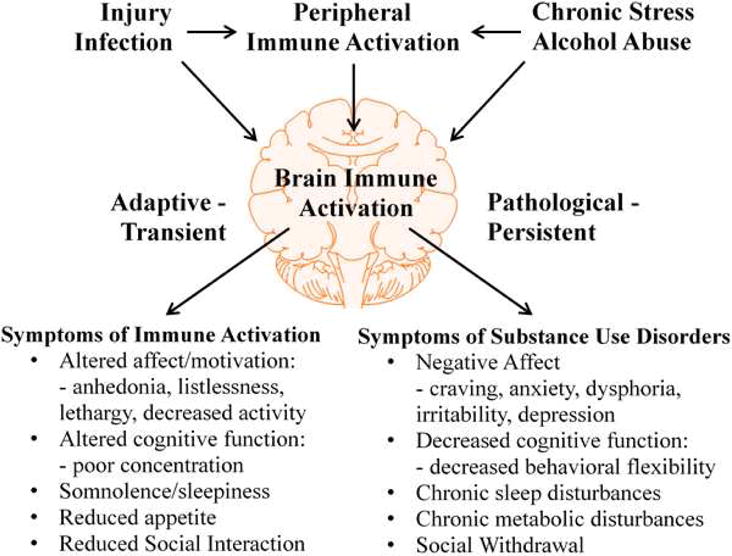
A variety of common, naturally occurring stressors such as injury or infection cause immune activation. Immune activation occurs in both the periphery and central nervous system and leads to inflammatory cytokine production. Activation of the immune system causes adaptive behavioral changes known as sickness behavior, which includes anhedonia, listlessness, lethargy and decreased activity, poor concentration, somnolence/sleepiness, loss of appetite and reduced social interaction. These changes are adaptive, transient and facilitate the recovery of the organism by reducing activity for healing. However, under conditions of chronic repeated stress and/or alcohol abuse, increased brain innate immune activation can lead to pathological and persistent changes in mood, cognition and other physiological factors such as sleep. Many of the behavioral characteristics of substance abuse disorders have been linked to peripheral and central immune activation, including substance self-administration, negative affect, decreased cognitive function and social withdrawal.
Studies have found similar mechanisms through which stress and alcohol contribute to inflammation (Fig 2). Stress enhances intestinal permeability and bacterial translocation from the gut lumen (Garate et al., 2013). The immune system mounts a response to the leaked bacteria, thereby causing peripheral inflammation. Alcohol also contributes to gut leakiness and translocation of bacteria from the intestinal lumen to the periphery (Adachi et al., 1995). Leaked bacterial products cause inflammation in the liver and release of pro-inflammatory (Giraldo et al., 2010)cytokines into the systemic circulation (Mayfield et al., 2013). Therefore, stress and alcohol can cause gut leakiness, leading to peripheral inflammation. Both stress and alcohol (Ellis, 1966) also increase glucocorticoids. Intriguingly, glucocorticoids have pro-inflammatory effects under the right circumstances (Sorrells et al., 2009). While glucocorticoids are acutely anti-inflammatory at high doses, glucocorticoids also have a priming effect on the inflammatory response. That is to say, prior exposure to glucocorticoids enhances the inflammatory response to subsequent stimuli (Frank et al., 2010). Therefore, both stress and alcohol may exert inflammatory/priming effects through glucocorticoid signaling. Stressors also enhance the systemic release of cytokines (e.g. IL-1 and IL-6) (Merlot et al., 2002) as well as danger-associated molecular patterns (DAMPs) such as high mobility group box 1 (HMGB1), uric acid and heat-shock protein 72 (Hsp72) (Maslanik et al., 2013). Physical stressors such as trauma can release DAMPs directly via cellular damage. Immune activation due to psychological stressors might involve multiple mechanisms including sympathetic nervous system activation, as noradrenaline causes Hsp72 release from neutrophils (Giraldo et al., 2010) or endotoxin release due to increased intestinal permeability (Ait-Belgnaoui et al., 2012; Ferrier et al., 2003) that may involve CRF release (Overman et al., 2012; Saunders et al., 2002; Teitelbaum et al., 2008). Released DAMPs and cytokines can activate innate immune cells, thereby increasing inflammatory signaling. Peripheral inflammation can impact the brain and behavior in many ways (Fig 2) (Miller and Raison, 2016). One route is the neural route: the vagus nerve can sense peripheral inflammatory cytokines such as IL-1β through IL-1 receptors (Ek et al., 1998). Activation of vagal afferents transmits signals to the central nervous system where neural centers that promote sickness behavior are activated. Indeed, vagotomy reduces the sickness behavior response to a peripheral injection of LPS (Bluthe et al., 1994). Another means by which peripheral inflammation can impact the brain and behavior is the humoral route. This includes diffusion of cytokines from the periphery through leaky regions of the blood brain barrier, such as the circumventricular organs and by transport of cytokines across the blood brain barrier. Indeed, TNFα transporters on the blood brain barrier are essential for systemic inflammation to cause brain inflammation (Qin et al., 2007). A third route is the cellular route, by which activated immune cells such as monocytes traffic into the brain. The brain recruits monocytes in response to peripheral inflammation (D’Mello et al., 2009). In each case, the peripheral inflammatory signal is transmitted to the brain, where it can impact central inflammation and behavior. Thus, alcohol abuse and stress share common mechanisms of inducing neuroimmune signaling that could contribute to the progression and persistence of the neuropathology of addiction by cycles of stress and substance abuse
Figure 2. Mechanisms of Stress- and Ethanol-induced Immune Activation.
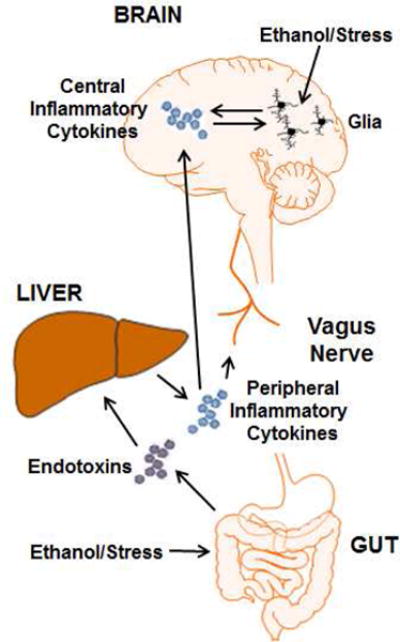
Stress and ethanol activate the peripheral and central immune systems in multiple ways. Both stress and ethanol can enhance gut leakiness. This causes increased translocation of bacterial products such as endotoxins from the intestinal lumen to the periphery. Leaked bacterial products make their way to the liver via the portal system where they induce an inflammatory response from resident macrophages. The production of peripheral inflammatory cytokines such as TNFα, IL-1β and IL-6 impact the brain and behavior through multiple mechanisms. One way is the neural route. The vagus nerve expresses cytokines receptors and is activated by peripheral inflammation. The signal of peripheral inflammation is transmitted to central brain regions involved in the regulation of sickness behavior. Another route is the humoral route. Peripheral cytokines can cross the blood brain barrier either by transport proteins or by diffusion in regions where the barrier is leaky. This can lead to a central immune response. Stress and ethanol also activate glia through more direct mechanisms. Glucocorticoids are important for the priming effect of stress on microglia. Also, ethanol exposure can directly activate glia.
3. Sensitization of Glia in the Stressed and Addicted Brain
Microglia are the resident macrophages of the brain and the primary cells of the neuroimmune system. They are involved in many physiological processes in the healthy brain, including synaptic pruning, debris clearance, immune surveillance/defense, neurogenesis and more (Kettenmann et al., 2011; Kettenmann et al., 2013). Microglia normally exist in a “resting” state, but can become activated in response to insults. Activation of microglia has been categorized according to either morphological or functional standards. Under the morphological classification, “resting” microglia are called “ramified” microglia. Morphological activation of microglia progresses through multiple states of increasing activation, including the hyper-ramified state, the bushy state and finally the amoeboid state (Fig 3) (Beynon and Walker, 2012). Fully activated amoeboid microglia are phagocytic cells with a rounded morphology. To date, no known studies have observed alcohol causing an amoeboid microglial activation. Rather, alcohol seems to sensitize microglia, inducing a hyper-ramified activation state characterized by modest morphological changes and increased release of cytokines and other signaling molecules (Fig 3). Functional microglial activation is traditionally understood as occurring along a spectrum between two extremes, although recent studies suggest functional activation is a more complex phenomenon (Xue et al., 2014). Functional activation states range from the pro-inflammatory, destructive M1 state (comparable to systemic Th1) to the anti-inflammatory, reparative M2 state (comparable to systemic Th2). M1 microglia secrete pro-inflammatory cytokines such as TNFα, IL-1β and IL-6 and produce reactive oxygen species through increased iNOS and NADPH oxidase expression. M2 microglia secrete anti-inflammatory cytokines such as IL-10 and synthesize extracellular matrix products to assist with healing (Cherry et al., 2014). The exact relationship between the morphological and functional activation states is currently unclear. Microglia also undergo another phenomenon known as priming, or sensitization. Priming occurs when an initial stimulus causes a greater microglial response to a subsequent stimulus (Perry and Holmes, 2014). Primed microglia produce a greater inflammatory response to immune stimuli than non-primed microglia.
Figure 3. Microglial Activation Following Stress and Ethanol.
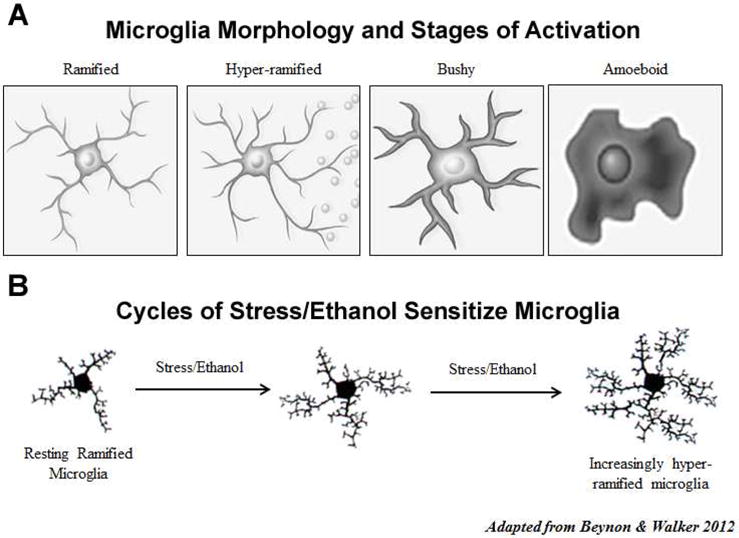
A. In the healthy brain, microglia normally exist in a “resting” or ramified state. However, in response to insults microglia undergo a process known as activation which involves changes in microglial morphology, gene expression and function. Microglial activation can be morphologically classified according to stages of increasing activation, including hyper-ramified, bushy and amoeboid (illustrations adapted from Beynon & Walker, 2012). Hyper-ramified microglial activation occurs in response to relatively mild insults and is thought to be associated with cytokine release. Bushy and amoeboid microglia are observed in more overt forms of brain damage, such as seizure, stroke or trauma. B. Hyper-ramified microglial activation has been observed in chronically stressed brains. It is also the predominant form of microglial activation following alcohol abuse. Repeated cycles of stress and ethanol abuse result in increasingly sensitized/activated hyper-ramified microglia, contributing to the neurobiology of substance use disorders.
Interestingly, astrocytes are also an important component of the brain immune system (Farina et al., 2007). Astrocytes are involved in numerous physiological processes, such as metabolic support of neurons and modulation of synaptic transmission (Khakh and Sofroniew, 2015). Astrocytes express multiple immune receptors and cytokines that allow them to participate in immune processes (Jensen et al., 2013). In response to insults, astrocytes also undergo a process of activation or reactive gliosis that serves to limit tissue damage (Pekny and Pekna, 2014). Some studies suggest that, like microglia, astrocytes are capable of adopting pro-inflammatory and anti-inflammatory states (Jang et al., 2013). Together, microglia and astrocytes are the primary components of the brain immune system. Both microglia and astrocytes interact with neuronal synapses, positioning these cells to regulate synaptic transmission. Therefore, changes in neuronal functioning may occur through altered glial functioning.
Both stress and alcohol can affect glia. Various types of acute and chronic stress, including restraint (Tynan et al., 2010), foot shock (Frank et al., 2007), social defeat (Wohleb et al., 2011) and chronic variable stress (Kreisel et al., 2014), increase microglial markers such as CD11b and Iba1 across multiple brain regions, consistent with microglial activation. Acute stress also primes microglia. Exposure to acute foot shock enhanced the ex vivo microglial inflammatory response to the inflammogen lipopolysaccharide (LPS) (Frank et al., 2007). In addition to microglia, stress can also affect astrocytes. Sub-chronic stress increased the astrocyte marker GFAP in the hippocampus, suggestive of astrocyte activation (Lambert et al., 2000). Therefore, stress activates/primes glia. Various alcohol treatment protocols also alter glia. Ethanol increases microglial markers such as CD11b (Fernandez-Lizarbe et al., 2009a) and Iba1 (Qin and Crews, 2012b) in vivo, consistent with microglial activation. Alcohol exposure can also prime the microglial response to peripheral inflammation (Qin and Crews, 2012b). Astrocytes are also affected by alcohol, as evidenced by increased GFAP (Alfonso-Loeches et al., 2010b). Other studies show that alcohol consumption can initially increase astrocytic GFAP in the hippocampus, but decrease GFAP after extended exposure (Franke, 1995). Therefore, alcohol can also impact glia. Repeated bouts of exposure to these stimuli may lead to increasing glial activation that persists over time. Indeed, two alcohol binge treatments activate microglia to a greater extent than a single binge treatment (Marshall et al., 2016b). Binge ethanol also increases microglial markers up to four weeks following the binge, indicative of persistent activation (Marshall et al., 2013). Thus, repeated episodes of stress and/or alcohol abuse may cause escalating and persistent glial activation (Fig 3), and this pathological activation/sensitization may contribute to the neurobiology of alcohol use disorders.
4. Stress, Inflammation and Affective Disorders
Stress and the resulting inflammation impact the neuroimmune system of the CNS. Stress sensitizes microglia to inflammation in an HMGB1-dependent manner (Weber et al., 2015). Chronic stress activates microglia in multiple brain regions (Tynan et al., 2010) and causes depression-like behavior. Administration of microglial-modulating agents can block the development of depression-like behavior, suggesting that microglia may have a causal role in the development of depression (Kreisel et al., 2014). Human studies also support a role for chronic stress and inflammation in affective disorders. Administration of inflammatory agents (such as interferon-α for the treatment of hepatitis C) can cause depression in previously non-depressed patients (Bonaccorso et al., 2002). Human studies find individuals with increased plasma levels of CRP, an acute phase marker of inflammation, have an increased risk for depression (Young et al., 2014). Interestingly, increased CRP in adolescents has also been found to predict addiction later in life (Costello et al., 2013). Other studies find peripheral inflammation can increase brain expression of innate immune signals and contribute to depression and negative affect (Harrison et al., 2014). Imaging studies in humans show that a marker of microglial activation, TSPO, is elevated in patients with depression (Setiawan et al., 2015). Depressed patients who committed suicide showed increased expression of microglial markers Iba1 and CD45 and increased levels of the inflammatory cytokine Ccl2 in the cingulate cortex (Torres-Platas et al., 2014). Studies also find changes in astrocyte markers in the brains of depressed patients (Rajkowska and Stockmeier, 2013). These findings suggest that stress contributes to increased brain innate immune signaling induction and psychopathology.
Stress-induced CNS inflammation can contribute to affective disorders through many mechanisms. Acute inflammation alters the activity of neural circuitry implicated in anxiety and depression. For example, acute inflammation changes activity in the cingulate cortex, medial prefrontal cortex and amygdala (Harrison et al., 2009). Acute inflammation can also impair spatial memory in humans (Harrison et al., 2014), potentially contributing to the cognitive changes seen in affective disorders. Furthermore, studies find that a variety of cytokine receptors, such as those for TNFα, IL-1β, IL-6 and the interferons, are expressed on neurons (Khairova et al., 2009), suggesting that cytokines act directly on neurons to influence their activity. Inflammation can also act on the serotonergic system to cause depression-like behavior. LPS increases activity of the serotonin transporter (SERT) and increases depression-like behavior. The ability of inflammation to cause depression-like behavior is lost in SERT knock-out animals (Zhu et al., 2010). Inflammation also increases the activity of an enzyme known as indoleamine 2,3-dioxygenase (IDO) in microglia. IDO generates the compound kynurenine, which has been shown to induce depression-like behavior (O’Connor et al., 2009). Pharmacological inhibition of IDO blocked the development of depression-like behavior (O’Connor et al., 2009) and blocks microglial activation. The trafficking of peripheral inflammatory monocytes into the CNS has also been implicated in stress-induced behaviors. Stress causes trafficking of peripheral monocytes into the brain that, in turn, promotes anxiety-like behavior (Wohleb et al., 2013). Thus, stress-induced microglial activation contributes to increased depression-like negative affect and anxiety-like morbidity that overlaps with and contributes to progressive stages of addiction.
Inflammatory cytokines such as TNFα and IL-1β can also impact neuronal plasticity. Interestingly, low physiological levels of TNFα and IL-1β seem to be important for long-term potentiation (LTP); however, high pathological levels of these cytokines disrupt the process. Blocking TNFα in hippocampal slices decreased synaptic strength (Beattie et al., 2002). However, excessive TNFα can inhibit LTP in hippocampal slices (Tancredi et al., 1992). The effects of cytokines on plasticity are behaviorally relevant, as TNFα overexpressing mice show decreased performance on spatial learning and memory tasks (Aloe et al., 1999). Also, physiological doses of IL-1β are needed for memory. Blockade of IL-1β signaling by its antagonist, IL-1ra, impairs memory. However, addition of excessive IL-1 β also impairs memory (Goshen et al., 2007). Therefore, immune signaling may alter neurons and neural circuitry and change behavioral phenotypes by impacting plasticity through changes in cytokine levels.
Inflammation can also impact neurogenesis, which is implicated in affective disorders. Neurogenesis occurs throughout adulthood in discrete brain regions, including the forebrain subventricular zone and the subgranular zone of the hippocampal dentate gyrus. Hippocampal neurogenesis contributes not only to learning and memory (Shors et al., 2001), but also mood and affective state (Malberg et al., 2000). Decreased neurogenesis contributes both to anxiety and depression (Hill et al., 2015). The ability of anti-depressants to alleviate depression-like behavior is dependent on hippocampal neurogenesis (Santarelli et al., 2003). Both inflammation (Ryan and Nolan, 2016) and chronic stress (Kreisel et al., 2014) reduce neurogenesis and cause depression-like behavior. Indeed, stress induces IL-1β in the hippocampus which decreases neurogenesis and contributes to depression. Inhibition of IL-1β blocks stress-induced decreases in neurogenesis and depression-like behavior (Koo and Duman, 2008). Furthermore, enhanced neurogenesis is sufficient to reverse the anxiety-like and depression-like behavior caused by chronic corticosterone treatment (Hill et al., 2015). Therefore, stress can increase CNS immune signaling, which contributes to affective disorders.
5. Innate Immune Molecules Mimic Addiction-like Behavior
Like stress, excessive alcohol use can impact the neuroimmune system, contributing to the development of alcohol use disorders and related psychopathology in many ways. There are multiple mechanisms through which this can occur. Alcohol is capable of acting directly on immune cells to increase inflammation. Alcohol can directly activate microglia in vitro, increasing expression of pro-inflammatory genes such as TNFα, IL-1β and iNOS (Fernandez-Lizarbe et al., 2009a). Alcohol also increases neuroimmune signaling molecules in vivo. These effects in vitro and in vivo are dependent on TLR4, which seems to play a pivotal role in alcohol-induced neuroimmune signaling (Alfonso-Loeches et al., 2010b). Repeated binge alcohol treatment persistently increases neuroimmune molecules such as TLR3, TLR4, HMGB1, RAGE, etc. (Vetreno and Crews, 2012; Vetreno et al., 2013a). Studies on the post-mortem brains of human alcoholics also find increased microglial markers (He and Crews, 2008; Rubio-Araiz et al., 2016) and increased astrocyte markers (Rubio-Araiz et al., 2016). Post-mortem human alcoholic brains show increased neuroimmune molecules, such as MCP-1, TL2, TLR3, TLR4 and HMGB1 (Crews et al., 2013b; He and Crews, 2008). Furthermore, alcohol appears to cause a pro-inflammatory M1 activation of microglial cells, as alcohol treatment induces expression of pro-inflammatory cytokines, such as TNFα and IL-1β, and enzymes, such as NADPH oxidase (Qin and Crews, 2012b; Qin et al., 2008b). Further, alcoholic hepatic encephalopathy patients have increased microglial proliferation (Dennis et al., 2014). Thus, alcohol increases microglia and glial signaling contributing to lasting changes.
Alcohol also sensitizes the neuroimmune system to subsequent inflammatory stimuli. Prior alcohol treatment enhanced brain levels of pro-inflammatory cytokines to immune challenges such as lipopolysaccharide (LPS) or polyinosine-polycytidylic acid (poly I:C) (Qin and Crews, 2012b; Qin et al., 2008b). It is important to note that LPS itself does not enter the brain, but causes systemic release of immune mediators that do enter the brain that are required for central LPS responses such as TNFα (Qin et al., 2007). It is unclear whether poly I:C crosses into the brain, but it does cause TNFα release similar to LPS (Qin and Crews, 2012a). The sensitization or priming of the neuroimmune response has important consequences for the pathogenesis of alcohol use disorders. Alcohol neuroimmune sensitization increases neurodegeneration and loss of neurogenesis following an immune challenge (Qin and Crews, 2012b, d; Qin et al., 2008b). Neurodegeneration and decreased neurogenesis have been linked to a variety of neuropsychiatric diseases. Therefore, repeated exposure to alcohol enhances and sensitizes neuroimmune signaling, thereby contributing to molecular and cellular causes of alcohol use disorders and co-morbidities of alcoholism, such as affective disorders.
Current theories of addiction posit a three-staged cycle that contributes to substance use disorders: binge/intoxication, withdrawal/negative affect and preoccupation/craving (Koob and Volkow, 2010). Interestingly, the neuroimmune system can impact the neurobiology of addiction at multiple points in this process. Regarding the binge/intoxication, multiple immune interventions have been found to regulate alcohol consumption. Injection of the IL-1 receptor antagonist or the anti-inflammatory cytokine IL-10 into the basolateral amygdala reduced alcohol self-administration (Marshall et al., 2016a; Marshall et al., 2016c). Furthermore, microglial-inhibiting compounds blocked conditioned place preference to cocaine, suggesting a role for the neuroimmune system in the subjective reward of drugs (Northcutt et al., 2015). Manipulations of the immune system have also been found to change alcohol consumption in experimental animals. Injection with the inflammogen LPS increases ethanol consumption in mice (Blednov et al., 2011). Furthermore, deletion of certain neuroimmune genes, including cytokine genes encoding IL-6 and IL-1ra, decreased ethanol consumption (Blednov et al., 2012). Local knockdown of TLR4 in the central amygdala also decreased ethanol self-administration (Liu et al., 2011). Administration of the cytokine MCP-1 increased self-administration of alcohol in rats (Valenta and Gonzales, 2016). These data suggest that neuroimmune signaling impacts alcohol consumption. Regarding the craving/preoccupation stage, which often precedes binge intoxication, blocking IL-1β signaling in the VTA was found to block cocaine-induced dopamine release in the nucleus accumbens (Northcutt et al., 2015). Furthermore, human alcoholics have increased plasma levels of multiple pro-inflammatory cytokines such as TNFα (Heberlein et al., 2014), which correlates with severity of alcoholism, IL-1β, IL-6 and IL-8 which correlate with alcohol craving (Heberlein et al., 2014; Leclercq et al., 2014). TNFα, IL-6, and IL-1β each cross the blood brain barrier and might contribute to central effects (Banks et al., 1995; Banks et al., 1994). Though the precise signaling mechanisms need to be further expounded, central pro-inflammatory cytokines modulate alcohol craving and consumption and peripheral cytokines are indices of craving and severity of alcoholism.
The immune system also impacts the withdrawal/negative affect stage. Indeed, a significant body of literature suggests that substance abuse contributes to the development of negative affective states such as anxiety and dysphoria (Koob and Le Moal, 2005). Additional substance abuse aimed at alleviating these negative states contributes to a cycle of abuse and dependence. Inflammation and cytokine induction are known to contribute to negative affect. Withdrawal from alcohol increases inflammatory cytokines in the brain (Freeman et al., 2012). Also, intracerebroventricular injection of cytokines sensitized anxiety-like behavior during withdrawal from ethanol (Breese et al., 2008). These data suggest that cytokines are involved in the ethanol withdrawal process and contribute to the negative affect of alcohol use disorders. Alcohol can also decrease neurogenesis, which is associated with negative affect. Chronic voluntary alcohol consumption decreases neurogenesis during withdrawal and is accompanied by the development of depression-like behavior (Stevenson et al., 2009). Both the decreased neurogenesis and depression-like behavior were reversed by treatment with anti-depressants. Therefore, there are multiple mechanisms by which alcohol increases CNS neuroimmune signaling, contributing to the molecular and cellular causes of alcohol use disorders.
6. HMGB1, Toll-Like Receptors and Innate Immune Signaling in the Brain
Understanding the mechanism of signaling for many hormones and neurotransmitters has benefited from manipulations in chemical structure allowing agonist and antagonist pharmacological distinction of responses. However, receptors that respond to larger molecules are more difficult to fully characterize. In the innate immune system, this has been the case. Initial studies characterized many receptors as responding to pathogens such as bacteria and virus components. These pattern recognition receptors (PRRs) and somewhat promiscuous and recently, studies involving “sterile” inflammation i.e. innate immune activation in the absence of a foreign pathogen have identified additional roles for these receptors. This is especially true for the CNS, which is normally a sterile environment, yet PRRs play critical roles in addiction pathology. PRRs have revolutionized the understanding of the innate immune system signaling. PRRs recognize specific molecular patterns associated with either foreign pathogens (Pathogen Associated Molecular Patterns-PAMPs) or endogenous molecules associated with cell death, damage or stress (Danger Associated Molecular Patterns-DAMPs). Five classes of PRRs have been described: Toll-like receptors (TLRs), C-type lectin receptors, nucleotide binding domain receptors (leucine-rich repeat containing or NOD-like receptors), RIG-I-like receptors, and AIM 2-like receptors (Brubaker et al., 2015). TLRs are the most well characterized PRRs, and have been implicated in addiction. TLRs are characterized by an N-terminal extracellular leucine-rich repeat sequence and an intracellular Toll/Interleukin-1 receptor/resistance motif (TIR) (Takeuchi and Akira, 2010). These receptors recognize PAMPs and DAMPs that include a variety of molecules from bacterial endotoxin to mammalian HMGB1 and heat shock proteins (Vabulas et al., 2002). To date ten TLRs have been identified in humans and 12 in mice (Brubaker et al., 2015). Activation of TLRs by endogenous DAMPs has been implicated in several non-infectious neurological disorders, including alcoholism (Table 1). Transcriptome analyses in Drosophila have implicated Toll genes in the alcohol response, as discussed by Park et al. in their review in this special edition (Park et al., 2017). The expression of several TLRs are upregulated in the brains of human alcoholics. TLRs 2–4 as well as the PRR RAGE (Receptor for advanced glycation End-products) were each increased in the frontal cortex of postmortem human alcoholics (Crews et al., 2013a). Ethanol treatment in vivo has also been shown to increase the expression of TLRs 2–4 in cortex and cerebellum with subsequent NF-κb activation and cytokine induction (Crews et al., 2013a; Lippai et al., 2013b). Furthermore, ethanol sensitizes neuroimmune responses to TLR3 and TLR4 agonists (Qin and Crews, 2012a; Qin et al., 2008a). Chronic ethanol enhanced responses to systemic TLR3 and TLR4 agonists. This increased systemic and brain pro-inflammatory response is in part due to binge alcohol increasing intestinal permeability, resulting in endotoxin/LPS as well as bacterial RNA and other gut molecules crossing into the blood inducing innate immune responses (Bala et al., 2014; Keshavarzian et al., 2009; Szabo et al., 2010). Thus, there are multiple mechanisms by which ethanol leads to TLR activation and induction in the brain.
Table 1.
Selected Toll-like Receptors (TLRs) implicated in Neurological Diseases
| TLR | Foreign PAMP | Endogenous DAMP | Neurological Condition |
|---|---|---|---|
| 2 | Bacterial di- and tri-aceylated polypeptides (Buwitt-Beckmann et al., 2006) Gram (+) lipoglyans (Blanc et al., 2013) | α-synuclein | Parkinson’s Disease (Kim et al., 2013) |
| 3 | dsRNA | stathmin | Alzheimer’s Disease (Jackson et al., 2006) Multiple Sclerosis (Bsibsi et al., 2010) |
| 4 | Bacterial endotoxin Peptidoglycans |
HMGB1 HSPs 60, 70/72 (Vabulas et al., 2002) | Stroke, Traumatic Brain Injury Chronic Pain Alcoholism (Crews et al., 2013a) |
| 7 | ssRNA (Lehmann et al., 2012b) | Let-7, miR-21 | Alcoholism (Coleman et al., 2017) Alzheimer’s Disease (Lehmann et al., 2012a) Chronic Pain (Park et al., 2014) |
High mobility group box 1 protein (HMGB1) has been identified as one such critical immune mediator in alcohol abuse. HMGB1 is increased in the cortex of human alcoholics (Crews et al., 2013a). Similar to TLRs 2-4, ethanol treatment in vivo increases HMGB1 in cortex and cerebellum (Crews et al., 2013a; Lippai et al., 2013b). HMGB1 is a nuclear chromatin binding protein that can also be released as an innate immune mediator (Muller et al., 2004). HMGB1 consists of two basic L-shaped ‘HMG box’ domains and an acidic 30 aa tail domain joined by a connecting linker (Stott et al., 2010). In the nucleus, HMGB1 stabilizes chromatin structure and acts as a chaperone for certain transcription factors (Thomas and Stott, 2012). Extracellular HMGB1 activates innate immune responses through direct binding to TLR4 or RAGE depending on its redox state (Janko et al., 2014) (Figure 4). The Cys106 thiol/Cys23-Cys45 disulfide bond form of HMGB1 acts at TLR4 receptors (Figure 4a), while fully reduced HMGB1 is active at the RAGE receptor (Figure 4b) (Venereau et al., 2012; Yang et al., 2012). Fully oxidized HMGB1 is inactive at immune receptors (Figure 4c) (Liu et al., 2012). Both ethanol and tobacco smoke extract cause HMGB1 translocation from the nucleus with subsequent release in vitro (Chen et al., 2016; Zou and Crews, 2014). Thus, ethanol and possibly other drugs of abuse lead to ‘sterile inflammation’ through the release of HMGB1. The release of other endogenous DAMPs likely also occurs, with recent reports indicating ethanol-induced release of TLR7-activating miRNA. The complex structure of HMGB1 allows it bind to a large number of molecules that also sensitizes other agonist responses through their associated receptors in a complex manner that is poorly understood. The double box and cysteine-disulfide structure of HMGB1 creates unique heterodimers with lipoglycans, endotoxin, and cytokines (e.g. IL-1β, Figure 4d) and nucleic acids (e.g. miRNA, Figure 4e) (Bianchi, 2009; Hreggvidsdottir et al., 2009). HMGB1 enhances the potency of cytokines at their respective receptors leading to greater immune responses (Figure 4c). Furthermore, HMGB1 is required for immune responses to endosomal nucleic acid binding TLRs (3, 7 and 9) (Yanai et al., 2009), though the mechanism is unclear. Recent evidence has found that ethanol causes increasing binding of HMGB1 to the miRNA let-7 while reducing its association with the RISC complex protein Ago2. HMGB1 might aid in the presentation of nucleic acids to their endosomal TLR receptors. Thus, the modulation of HMGB1 by ethanol might activate signaling through multiple TLRs in addition to TLR4.
Figure 4. Mechanisms of HMGB1 signaling.
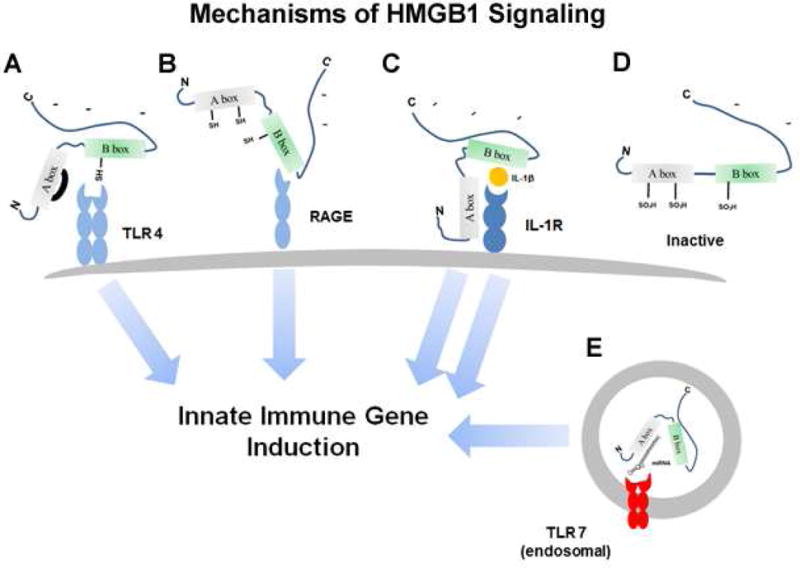
The HMGB1 protein is comprised of two similar Box structures, A and B, and a long C terminal negatively charged tail. There are several cysteines, some of which can form disulfide bonds. HMGB1 is secreted from activated or stressed cells or from dying necrotic cells and acts as an immune modulator. A. TLR4 Agonist HMGB1. The reduced Cys106 thiol/Cys23-Cys45 disulfide bond form of HMGB1 acts directly as an agonist at TLR4 receptors to cause innate immune activation. B. RAGE agonist-HMGB1. The fully reduced non-disulfide form of HMGB1 acts at RAGE receptors to cause innate immune induction and neurite outgrowth. C. Heterodimer HMGB1/IL-1 agonist: HMGB1 forms pro-inflammatory complexes with cytokines such as IL-1β to cause enhanced IL-1β signaling through the IL-1 receptor and greater innate immune induction. D. The fully oxidized form of HMGB1 is inactive at immune receptors. E. HMGB1-miRNA-chaperone: HMGB1 binds miRNA and activates the RNA sensing TLR7 receptor increasing immune activation.
7. TLR4 - A key brain receptor involved in ethanol induced neuropathology
Ethanol likely affects the signaling of multiple TLRs. However, TLR4 in particular has been found to play a critical role in the neuroimmune activation by ethanol. TLR4 knockout mice are protected from ethanol induced glial cell activation, NF-κB activation, and caspase-3 activation (Alfonso-Loeches et al., 2010a, b; Blanco et al., 2005; Fernandez-Lizarbe et al., 2009a, b; Pascual et al., 2011a; Valles et al., 2004). Cultured astrocytes with siRNA against TLR4 or MD-2 and CD14 (critical adaptor molecules for TLR4 signaling) were also protected from ethanol induced NF-κB induction. Mice lacking TLR4 were also protected against ethanol withdrawal associated anxiety-like behavior and memory impairment (Pascual et al., 2011b). Ethanol not only increases the expression of TLR4 (Crews et al., 2013a; Fernandez-Lizarbe et al., 2013), it increases the formation of TLR4/TLR2 heterodimers in lipid rafts of microglia, leading to iNOS induction and MAPK activation (Fernandez-Lizarbe et al., 2013). Importantly, both TLR4 and TLR2 KO microglia were protected in this study. In another genetic model, the alcohol preferring p-rats have increased expression of TLR4 in the VTA that was related to binge ethanol responding (June et al., 2015). TLR4 expression in the VTA was regulated by GABA(A)α2 receptor (Liu et al., 2011) and corticotropin-releasing factor (CRF) expression (June et al., 2015). Pharmacological studies also illustrate the role of TLR4 in the immune pathology of ethanol. TLR4 stimulation with LPS leads to a prolonged increase in alcohol self-administration (Blednov et al., 2011). Mice lacking CD14, a critical adapter protein for TLR4, were protected from this effect. Naltrexone and Naloxone, opioid antagonists used to treat alcoholism, also have anti-TLR4 actions. These antagonists reduce superoxide generation, cytokine production, and dopaminergic neurotoxicity by the TLR4 agonist LPS (Liu et al., 2000a; Liu et al., 2000b). In Silico analyses indicate that naltrexone and naloxone fill the LPS binding site of the MD2 co-receptor for TLR4 (Hutchinson et al., 2010). Furthermore, the opioid inactive stereoisomers (+)-naltrexone and (+)-naloxone have been shown recently to possess anti-TLR4 properties (Wang et al., 2016) indicating an immune mechanism that is independent of the opioid effects. These studies have identified a link between neuroimmune activation by ethanol via TLR4 signaling. This, coupled with the aforementioned studies regarding the endogenous TLR4 agonist HMGB1, emphasizes the importance of studying the efficacy of targeted TLR4 antagonists for the treatment of alcohol use disorders. Thus, significant opportunities exist for identifying unique drug targets for neuroimmune pathology in alcoholism.
8. Induction of Innate Immune Genes, and miRNA
Innate immune signaling is complex and involves multiple intracellular signaling cascades that lead to activation of key transcription factors. TLR signaling is mediated through key adapter proteins. Upon recognition of the ligand by the TLR, these adapter proteins initiate the signaling cascade. The TIRAP/MyD88 adapter protein complex interacts with all the TLRs, except for TLR3. TIRAP/MyD88 complex formation leads to activation of IL-1 receptor-associated kinases (IRAKs) and TNF receptor-associated factor 6 (TRAF6) which cause IκB and MAPK activation. IκB and MAPK ultimately lead to activation of the transcription factors Nuclear Factor kappa-light-chain-enhancer of Activated B cells (NF-κB) and Activated Protein-1 (AP-1) respectively. These transcription factors regulate the expression of pro-inflammatory cytokines that propagate and magnify the immune response. Furthermore, these transcription factors are also linked to both stress and addictive behaviors. For instance, NF-κB activation occurs in the nucleus accumbens in response to cocaine, and was required for increased conditioned place preference (Ang et al., 2001; Russo et al., 2009). Restraint stress increases the expression of NF-κB, cytokines, prostaglandin E2, and cyclooxygenase-2 (COX-2) in the CNS (Madrigal et al., 2003; Madrigal et al., 2002). Also, ethanol has been found to induce AP-1 transcription in primary cortical neurons (Qiang and Ticku, 2005). Subcellular location and receptor trafficking also contribute to TLR responses, with nucleic acid sensing TLRs being endosomal and cell surface trafficking to endosomal TLRs (including TLR4 after its endocytosis) also bind to the TRIF adapter protein leading to transcription factor IRF3 activation and type I interferon gene induction. It is important to note that responses vary across cell type and most signaling has been characterized in monocyte and other immune cells that likely differ from that of brain cells.
The precise signaling pathways for the TLRs in each brain cell type have yet to be delineated. Both microglia and astrocytes clearly show canonical TLR4 responses to ethanol leading to NF-κB activation and further innate immune induction. However, TLR responses in neurons are poorly understood. Some have suggested that neurons are not capable of activating NF-κB gene transcription (Mao et al., 2009). Others, however, find NF-κB activation in glutamatergic neurons that may regulate plasticity, learning and memory (Kaltschmidt and Kaltschmidt, 2015). However, RAGE was originally identified as a neuronal receptor that bound HMGB1, originally called amphoterin, involved in neurite outgrowth (Hori et al., 1995). Ethanol is known to activate NF-κB in neurons in brain slice culture (Zou and Crews, 2006) and in vivo (Ward et al., 1996). The activated p-65 NF-κB subunit has been shown to colocalize with dorsal horn spinal neurons (Bai et al., 2014) and both P19 and SH-SY5Y neuronal cell lines show NF-κB dependent regulation of μ-opioid receptor expression (Borner et al., 2012; Wagley et al., 2013). These responses are complex in part due to the release of HMGB1 by stimuli that can subsequently activate multiple TLR and RAGE signals difficult to distinguish from the primary stimuli. For example, ethanol responses in brain involve signaling across microglia, astroglia and neurons. HMGB1 is one of the signals that contribute to neuroimmune activation and other responses. A comparison of ethanol activation of neuroimmune signaling in neuron-like SH-SY5Y and microglia-like BV cells finds ethanol induces TLR7 mRNA in both cell types, but neurons respond at concentrations below binge drinking blood levels whereas microglia-like BV cells do not respond until high concentrations of alcohol well above the binge drinking BAC, but had a more robust response (Figure 5). Though the unique cellular roles need to be further identified, it is clear that ethanol activates NF-κB signaling in brain. We have found that ethanol increases NF-κB–DNA binding both in in vivo in mice (Crews et al., 2006) and in vitro in rat hippocampal-entorhinal cortex slice culture (Zou and Crews, 2006). Furthermore, we and others have found that ethanol causes induction of NF-κB target genes in brain, such as the chemokine monocyte chemoattractant protein-1 (MCP-1, CCL2) (He and Crews, 2008), proinflammatory cytokines (TNFα, IL-1β, and IL-6) (Qin et al., 2007), proinflammatory oxidases (inducible nitric oxide synthase (Alfonso-Loeches et al., 2010b; Zou and Crews, 2010), COX (Alfonso-Loeches et al., 2010b; Knapp and Crews, 1999), and NOX (Qin et al., 2008b)), and proteases (TNFα-converting enzyme [TACE] and tissue plasminogen activator [tPA]) (Zou and Crews, 2010).
Figure 5. Neuron-like SH-SY5Y cells are more sensitive to ethanol induction of TLR7 than microglia-like BV cells.
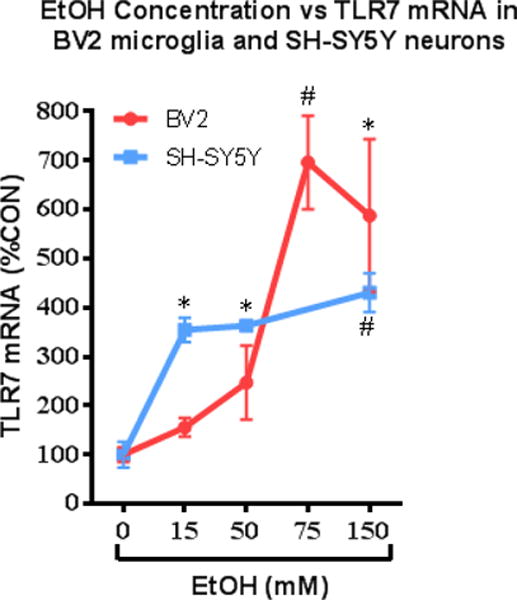
Shown are TLR7 mRNA levels following 24 hr. of treatment with various concentrations of ethanol in cultures of microglia-like BV2 and neuron-like RA-differentiated SH-SY5Y cells. TLR7 mRNA was determined using RT-PCR. Data is represented as percent control (%CON) for each respective cell type, with CON set at 100%. Note neuron-like SHSY-5Y show a maximal response at about 15 mM ethanol, whereas microglia show little response at this concentration, but show far larger responses at higher concentrations of ethanol. (*p<0.05, #p<0.01 vs. CON.) Adapted from: (Lawrimore and Crews, 2017)
In addition to activation of TLRs via release of DAMPs, ethanol also modulates immune function through release of microRNAs (miRNA). MicroRNAs are small non-coding RNAs (~22 nucleotides). Neuroimmune regulation of miRNAs is extensive with numerous miRNAs involved (Thounaojam et al., 2013). These molecules regulate immune function in two known fashions. First, they regulate the stability of target mRNAs in the cytosol via formation of an RNA-induced silencing complex (RISC) and binding to the 3′-untranslated region of mRNA (Ambros, 2004; Bartel, 2004; Czech and Hannon, 2011). The role of microRNAs as transcriptional regulators in alcoholism is discussed by Warden and Mayfield in their article in this special edition (Warden and Mayfield, 2017). Another newly identified mechanism whereby ethanol modulates immune activation is through the extracellular release of miRNA (Turchinovich et al., 2013). Extracellular miRNA are contained either in extracellular vesicles (EVs) or bound to either protein or lipoprotein chaperones. MicroRNA-containing vesicles are capable of being endocytosed by other cells where they can modulate their function. Recently, microRNA in EVs from ethanol-treated monocytes were found to modulate the activation state of naïve monocytes (Saha et al., 2016). Further, the miRNAs let-7 and miR-21 were found to activate TLR7, leading to neuroimmune activation and neurodegeneration (Lehmann et al., 2012a; Yelamanchili et al., 2015). Thus, these miRNAs can have diverse actions to modulate neuroimmunity, both through regulation of mRNA stability and TLR activation. Ethanol exposure causes significant alterations to the miRNA milieu in the brain. In both human alcoholics and mice (20 day treatment), ethanol altered the expression of dozens of miRNAs in the frontal cortex (Lewohl et al., 2011; Nunez and Mayfield, 2012; Nunez et al., 2013). Interestingly, members of the miRNA let-7 family were induced across species. Recently, ethanol has been found to potentiate TLR7-mediated neurotoxicity via release of let-7 in MVs (Coleman et al., 2017). The miR-155 is also found in vesicles and promotes TLR4 associated neuroimmune responses after chronic ethanol (Lippai et al., 2013a). The miR-155 KO was protected from ethanol-induction of cytokines. Thus, ethanol modulation of miRNA release is an important aspect of innate immune induction. These pathways represent new pathological mechanisms that are poorly understood, but are targets for potential intervention in the pathology of alcoholism.
9. Autocrine and Paracrine Processes amplify NF-κB Innate Immune Gene Induction
As has been discussed above, regulation of innate immune responses in the brain is complex. This involves microglial priming and changes in activation state, microglia-neuronal interactions, and positive and negative feedback of intracellular signaling cascades. The transcription factor NF-κB is a critical modulator of immune function. Both stress and alcohol result in NF-κB activation through the immune mechanisms discussed above. Various models of stress in humans and rodents result in central and peripheral NF-κB activation in immune cells. Human psychosocial stress causes NF-κB activation in peripheral blood mononuclear cells (Bierhaus et al., 2003). Restraint stress in rodents causes NF-κB translocation into the nucleus with subsequent production of TNFα and pro-inflammatory prostaglandins (Bierhaus et al., 2003; Madrigal et al., 2002). Ethanol also activates NF-κB in rat and mouse brain (Qin and Crews, 2012c; Ward et al., 1996), as well as human astrocytes (Davis and Syapin, 2004). The system itself is well-regulated. However, in the context of repeated or prolonged activation, dysregulation occurs leading to many disease states (de Jesus et al., 2015). Recurrent and persistent NF-κB activation by repeated alcohol consumption appears to mimic a chronic inflammatory state. This is evidenced by increased innate immune markers in the post-mortem brains of human alcoholics (Coleman et al., 2017; Crews et al., 2013a) Further, NF-κB target genes are upregulated in the prefrontal cortex of alcoholics (Okvist et al., 2007). This persistent and chronic immune response has also been seen after one dose of systemic LPS. One administration of high dose LPS (5mg/kg) resulted in persistent microglial activation with ROS generation that led to dopaminergic neuron loss over a period of months (Qin et al., 2013). Though on a lower scale, binge-ethanol doses cause similar pathophysiology (Bala et al., 2014; Qin et al., 2008a; Zou and Crews, 2010).
The prolonged amplification of neuroimmune responses occurs on multiple levels. TLR activation in response to ethanol causes NF-κB activation, leading to increased TLR expression, cytokine production and DAMP release. This amplifies immune responses in an autocrine and paracrine fashion, as secreted DAMPs and cytokines activate both their cell of origin and surrounding cells (Figure 6). Paracrine signaling induced by ethanol also proceeds through the release of pro-inflammatory miRNA-containing microvesicles (Bala et al., 2011; Coleman et al., 2017; Szabo and Lippai, 2014). This method of intercellular paracrine signaling has been implicated in many conditions such as cancer, cardiovascular disease, and neurodegeneration (Hulsmans and Holvoet, 2013; Lehmann et al., 2012a; Ohshima et al., 2010). Also, ethanol induces ROS generation that activates NF-κB and leads to further cytokine release and paracrine signaling (Pietri et al., 2005; Qin and Crews, 2012c; Thakur et al., 2007). A ‘smoldering’ level of inflammation due to repeated binge-ethanol, as is typical with alcoholism, might underlie the transition from alcohol abuse to addiction. Thus, pharmacological strategies should target the amplification and persistence of the immune response, as this likely underlies disease pathology.
Figure 6. Autocrine and Paracrine Innate Immune Activation by Ethanol.
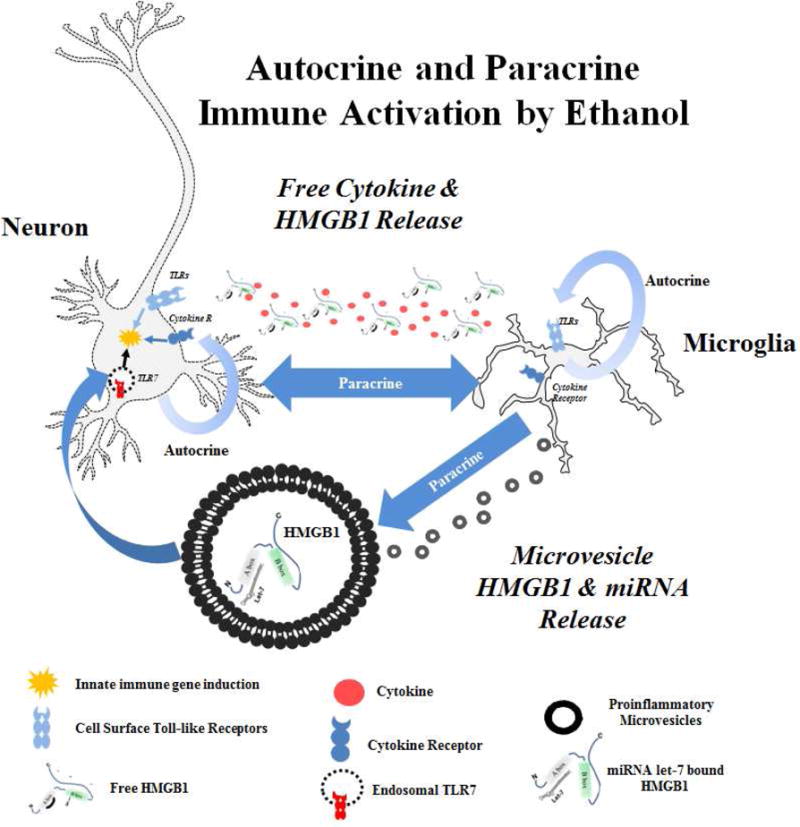
Ethanol causes Toll-like Receptor (TLR) activation in both autocrine and paracrine fashions. This occurs through release of endogenous TLR agonists. HMGB1 acts at TLRs directly and also acts as a chaperone for miRNA such as let-7 in microvesicles, promoting TLR7 signaling. Cytokines such as IL-1β bind cytokine receptors. Immune mediators act in both autocrine and paracrine fashions on neurons and microglia to amplify neuroimmune responses.
10. New treatments strategies based on immune pharmacology
The discovery of immune mechanisms of alcohol presents a new approach for the treatment of alcohol use disorders. It is currently unclear whether immune therapies would be of highest benefit for prevention or recovery from alcohol addiction. Neuroimmune activation can contribute to neurodegeneration in many disease settings (Crews and Vetreno, 2014; Qin et al., 2013; Rocha et al., 2015; von Bernhardi et al., 2015; Wang et al., 2015). Microglia and cytokines might also alter synaptic signaling (Lacagnina et al., 2016). For example, IL-1β reduces eIPSCs in the central amygdala to modulate ethanol effects on GABA receptors (Bajo et al., 2015). Other immune mediators might also impact synaptic signaling. Though immune therapies would likely not be of benefit in brain regions where neurodegeneration has already occurred, there might yet still be benefit if they allow for recovery of normal synaptic function. Therefore, significant exploration of immune therapies in various models of alcohol abuse is warranted. Many commercially available drugs have been found to have anti-inflammatory actions in the CNS. Certain antibiotics in particular have immune actions in addition to their antimicrobial functions. Table 2 details some candidate drugs that have been found to be beneficial for alcohol use disorders or other neuroimmune conditions. Minocycline, for example, is a tetracycline antibiotic and microglial inhibitor (Plane et al., 2010) that prevents ethanol induced-microglial activation and reduces alcohol self-administration (Agrawal et al., 2011; Qin and Crews, 2012a). Phosphodiesterase 4 (PDE4) inhibitors have traditionally been used to treat psoriasis and psoriatic arthritis. These drugs are thought to reduce inflammation by increasing cAMP concentration resulting in reduced NF-κB activation (Jimenez et al., 2001). Recently, these agents have been found to reduce ethanol intake in rodents (Bell et al., 2015; Blednov et al., 2014; Hu et al., 2011). A phase I clinical trial involving the PDE4 inhibitor ibudilast was recently completed but has not yet been reported. A recent placebo-controlled trial found that ibudilast reduced some of the subjective reward effects of methamphetamine (Worley et al., 2016). PPARγ agonists, historically used for the treatment of type II diabetes, might also be helpful in alcohol use disorders through their anti-inflammatory activities. The PPARγ agonist pioglitazone is a microglial inhibitor (Storer et al., 2005) that reduces toxicity in models of fetal alcohol spectrum disorders (Drew et al., 2015; Kane et al., 2011) and may potentially play a beneficial role in alcoholism. Each of these drugs, and others with anti-inflammatory actions should be investigated for their potential efficacy in alcohol use disorders.
Table 2.
Antibiotics and Other Drugs with Anti-Neuroinflammatory Actions
| Drug | Mechanism Primary Immune |
CNS activity |
|---|---|---|
| Minocycline | Tetracycline antibiotic Microglial inhibitor |
Reduces alcohol self-administration (Agrawal et al., 2011) Reduces ethanol microglia activation (Qin and Crews, 2012a) Prevents reinstatement of morphine and amphetamine seeking (Arezoomandan and Haghparast, 2016; Attarzadeh-Yazdi et al., 2014) |
| Rapamycin | Macrolide antibiotic mTORC1 inhibitor | Reduces binge ethanol intake in males (Cozzoli et al., 2016) Neuroprotection via Autophagy Promotion (Chen et al., 2012; Pla et al., 2016) |
| Azithromycin | Macrolide antibiotic Microglial inhibitor |
Promotes anti-inflammatory M2 microglial activation state (Zhang et al., 2015) |
| Rifampin | Bacterial RNA polymerase inhibitor TLR4 inhibition | Inhibits microglia activation to TLR4 (Bi et al., 2011; Wang et al., 2013) |
| Indomethacin | COX-2 inhibitor | Reduces alcohol self-administration (George, 1989) Reduces ethanol neurotoxicity (Pascual et al., 2007b) |
| Simvastatin | HMG-CoA Reductase Inhibitor NF-κB inhibition | Reduces inflammation and neurotoxicity to ischemia and injury (Lim et al., 2016; Sironi et al., 2006) |
| Glycyrrhizin | HMGB1 inhibition | Blocks ethanol-induced cytokine release (Zou and Crews, 2014) Reduces neuroinflammation after traumatic brain injury (Okuma et al., 2014) |
| Pioglitazone, DHA | PPARγ Agonists | Reduce toxicity and pro-inflammatory cytokines in fetal alcohol spectrum disorder model (Drew et al., 2015; Kane et al., 2011) |
| Ibudilast, Mesopram, Rolipram, CDP 840 | Phosphodiesterase 4 inhibition | Reduce ethanol intake in C57BL/6J mice (Blednov et al., 2014) Reduces ethanol self-administration in rats (Bell et al., 2015) |
| Naltrexone/Naloxone | μ-opioid antagonist TLR4 inhibition | Reduces alcohol self-administration Binds TLR4 adaptor protein MD2 (Hutchinson et al., 2010; Wang et al., 2016) |
11. Neuroimmune basis of Addiction
A hypothesis of stress- and drug-induced neuroimmune signaling that inactivates PFC and sensitizes limbic circuitry that suggest the progressive and persistent increases in HMGB1 and TLR with cycles of stress and drug abuse are the mechanisms underlying the progressive and persistent nature of addiction. Innate immune signaling is known to have a major impact on cognitive and emotive functioning, leading to dysfunction (Dantzer et al., 2008; Hanke and Kielian, 2011; Okun et al., 2010; Yirmiya and Goshen, 2011). Ethanol and other drugs promote innate immune gene induction (Crews and Vetreno, 2016; He and Crews, 2008; Qin et al., 2008b) that are linked to changes in executive function, reinforcement-reward and negative affect-craving-anxiety that promote alcohol abuse and addiction (Vetreno and Crews, 2014). Indeed, innate immune activation and TLR signaling via ethanol appear to be essential for ethanol induction of addiction-like behaviors. For instance, Guerri and colleagues (Pascual et al., 2011a) demonstrated that ethanol-induced innate immune activation in mice impaired short- and long-term memory for object recognition. This behavioral impairment was accompanied by a reduction of H3 and H4 histone acetylation as well as histone acetyltransferase activity in the frontal cortex, striatum, and hippocampus. Interestingly, ethanol neither impaired behavioral performance nor altered histone activity in TLR4 knockout mice. Another inflammatory pathway, mediated by the RAGE receptor, might be involved in the memory impairments associated with chronic alcohol exposure since neuroinflammation associated with increased expression of this receptor is implicated in the memory impairments that accompany Alzheimer’s disease (Arancio et al., 2004; Fang et al., 2010; Maczurek et al., 2008; Wilson et al., 2009). Adolescent intermittent ethanol treatment of rats has been found to increase expression of RAGE that persists into adulthood and mimics the increase found in post-mortem human alcoholic brain (Vetreno et al., 2013b). Thus, HMGB1, TLRs, RAGE, and other innate immune signaling are likely to drive changes in neurobiology related to addiction.
The innate immune system can modulate alcohol consumptive behavior. Analysis of genetically paired rats and mice that differ primarily in the amount of ethanol consumption reveal that NF-κB and other pro-inflammatory genes have high expression in high ethanol drinking animals (Mulligan et al., 2006). Beta-2 microglobulin, a NF-κB target gene involved in MHC immune signaling (Pahl, 1999), was elevated in high ethanol preferring brain transcriptomes (Mulligan et al., 2006). Interestingly, adolescent intermittent ethanol treatment results in increased expression of beta-2 microglobulin in the adult hippocampus (Figure 7). Beta-2 microglobulin progressively increases following adolescent alcohol exposure (Figure 7) in a manner similar to that found with HMGB1 (Vetreno and Crews, 2012) and RAGE (Vetreno et al., 2013b). In addition, work from Harris, Blednov and colleagues provided interesting and novel data supporting the hypothesis that innate immune genes regulate ethanol drinking behavior (Blednov et al., 2005; Blednov et al., 2012). Multiple strains of mice with immune gene deletion universally drink significantly less ethanol than matched controls across multiple ethanol drinking paradigms. Recently, Blednov and colleagues (Blednov et al., 2011) also demonstrated that innate immune activation through LPS can cause long-lasting increases in ethanol drinking. Indeed, strains of mice show varied innate immune responses to LPS that correspond to increases in the consumption of ethanol. Furthermore, a single injection of LPS is capable of producing a delayed, but long-lasting increase in ethanol consumption even in strains of high drinking mice. Recent data has revealed that the TLR family, especially TLR4, modulates ethanol intake. Administration of a GABAAα2 siRNA vector into the central nucleus of the amygdala of alcohol-preferring rats diminished binge drinking, which was associated with reduced expression of TLR4 (Liu et al., 2011). Interestingly, the neuronal location of GABAAα2 receptors suggests that the influence of TLR4 on binge drinking is at least partially mediated by neurons. Taken together, these findings are consistent with a modulatory role of innate immune genes in alterations in GABA signaling as well as alcohol preference and consumption.
Figure 7. β2-microglobulin is induced in adult hippocampus following adolescent intermittent ethanol treatment.
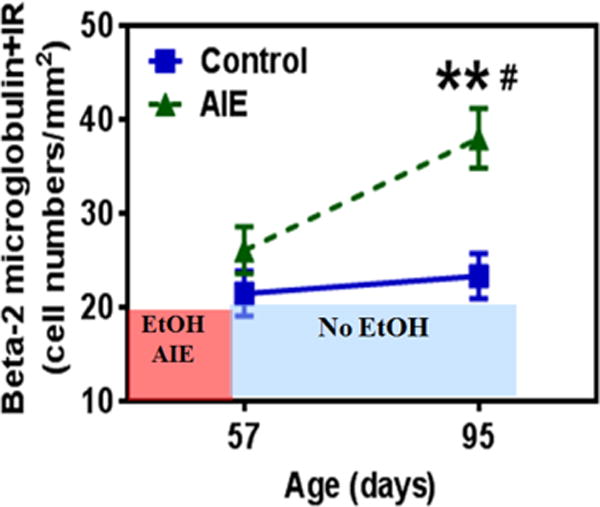
Shown are levels of β2 microglobin immunoreativity (β2M+IR) determined by immunohistochemistry in hippocampal dentate gyrus. Rats were treated with ethanol on an intermittent schedule through adolescence, i.e. adolescent intermittent ethanol treatment as described previously (Liu and Crews, 2015). β2M+IR was assessed on post-natal day 57, 24 hours after the last AIE ethanol treatment, and in adulthood on post-natal day 95. Note the ethanol-triggered induction of β2M during abstinence, but not at P57, just after ethanol treatment ended. (**p<0.01 compared with control at P95; #p<0.05 compared with AIE at P57).
Frontal cortical executive functions include: motivation, planning and goal setting, and behavioral flexibility. Among social drinkers the heaviest binge drinkers report more negative mood and perform worse on executive functioning tasks (Townshend and Duka, 2003; Weissenborn and Duka, 2003). The frontal cortex regulates mood and cognition through reciprocal glutamatergic connections with multiple brain regions. In astrocytes, ethanol exposure induces NF-κB transcription leading to increased expression of pro-inflammatory innate immune genes (Pascual et al., 2007a; Zou and Crews, 2006, 2010) and reduced astrocyte glutamate transport (Zou and Crews, 2005). The increased extracellular glutamate levels lead to increased neuronal excitation, microglial activation, and excitotoxicity (Ward et al., 2009; Zou and Crews, 2006). Indeed, increased glutamate excitotoxicity contributes to ethanol-induced increases of caspase-3 and COX-2 in the frontal cortex that require TLR4 signaling (Alfonso-Loeches et al., 2010b; Knapp and Crews, 1999). Glutamatergic hyper-excitability has also been demonstrated to occur in the cocaine- and stimulant-addicted brain (Reissner and Kalivas, 2010). The involvement of innate immune genes in the reduction of glutamate transporters and subsequent hyper-excitability reduces frontal cortical executive function that contributes to the neurobiology of addiction (Crews et al., 2006; Crews et al., 2011). Frontal cortical dysfunction is common in the alcoholic brain (Crews and Boettiger, 2009) and is manifest in impulsivity and behavioral inflexibility. Reversal learning is an index of behavioral flexibility that is especially relevant to addiction. Reversal learning involves adjusting learned responses to changes in reinforcement-reward (Stalnaker et al., 2009). Indeed, proper frontal cortex function is necessary to weigh the value of decisions and is important when new learning and/or responses are reinforced. The involvement of the frontal cortex in reversal learning is supported by lesion-induced reversal learning deficits similar in nature to chronic drug abuse-induced dysfunction (Schoenbaum et al., 2006). The entire reversal learning circuitry involves loops between the prefrontal cortex, striatum and amygdala (Izquierdo et al., 2016). This paradigm models the changes that occur when the initial reinforcement, intoxication, and binge drinking stages of alcohol abuse progress to obsessive-compulsive stages that are not able to avoid negative consequences of drinking due to persistent perseveration and inability to learn new approaches to reinforcement when situations change. Indeed, both human alcoholics (Fortier et al., 2008; Jokisch et al., 2014) and cocaine addicts (Stalnaker et al., 2009) show reversal learning deficits. Using a rat model of binge drinking, we found persistent deficits in reversal learning in rats (Obernier et al., 2002) and in adult mice following adolescent binge drinking (Coleman et al., 2011). Others have also seen reversal learning deficits in adult mice after chronic ethanol (Badanich et al., 2011). Interestingly, these reversal learning deficits are also found in rats exposed to self-administered cocaine or passive cocaine injections (Calu et al., 2007; Schoenbaum et al., 2004). Frontal cortical dysfunction is established as resulting in perseveration and repetition of previously learned behaviors due to failure to associate new information (e.g. negative consequences) into decision-making. Alcohol-induced damage to other regions associated with reversal learning (i.e. striatum and amygdala) might also contribute to reversal learning deficits and should be investigated. Further, the efficacy of immune modulating therapies to treat cognitive dysfunction associate with alcohol abuse should be investigated. Considerable emerging evidence supports a role for HMGB1/TLR signaling, innate immune gene induction and alterations in epigenetics and neurotransmission as culminating in the neurobiology of addiction (Crews et al., 2011; Cui et al., 2015; Vetreno and Crews, 2014).
Highlights.
Neuroimmune activation is a critical part of the pathology of alcohol use disorders
Repeated alcohol abuse and stress contributes to cycles of neuroimmune activation
DAMPs and secreted miRNAs are involved in alcohol-induced neuroinflammation signals
Neuroimmune therapies may represent the next generation of therapies for alcoholism
Acknowledgments
We would like to thank NIAAA for their financial support. (AA019767, AA11605, AA007573, and AA021040)
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Chemical compounds studied in this article: Minocycline (PubChem CID: 54675783); Rapamycin (PubChem CID: 5284616); Azithromycin (PubChem CID: 447043); Rifampin (PubChem CID: 5381226); Indomethacin (PubChem CID: 3715); Simvastatin (PubChem CID: 54454); Glycyrrhizin (PubChem CID: 14982); Pioglitazone (PubChem CID: 4829); Ibudilast (PubChem CID: 3671); Naltrexone (PubChem CID: 5360515)
References
- Adachi Y, Moore LE, Bradford BU, Gao W, Thurman RG. Antibiotics prevent liver injury in rats following long-term exposure to ethanol. Gastroenterology. 1995;108:218–224. doi: 10.1016/0016-5085(95)90027-6. [DOI] [PubMed] [Google Scholar]
- Agrawal RG, Hewetson A, George CM, Syapin PJ, Bergeson SE. Minocycline reduces ethanol drinking. Brain Behav Immun. 2011;25(Suppl 1):S165–169. doi: 10.1016/j.bbi.2011.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ait-Belgnaoui A, Durand H, Cartier C, Chaumaz G, Eutamene H, Ferrier L, Houdeau E, Fioramonti J, Bueno L, Theodorou V. Prevention of gut leakiness by a probiotic treatment leads to attenuated HPA response to an acute psychological stress in rats. Psychoneuroendocrinology. 2012;37:1885–1895. doi: 10.1016/j.psyneuen.2012.03.024. [DOI] [PubMed] [Google Scholar]
- Alfonso-Loeches S, Pascual-Lucas M, Blanco AM, Sanchez-Vera I, Guerri C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. J Neurosci. 2010a;30:8285–8295. doi: 10.1523/JNEUROSCI.0976-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alfonso-Loeches S, Pascual-Lucas M, Blanco AM, Sanchez-Vera I, Guerri C. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2010b;30:8285–8295. doi: 10.1523/JNEUROSCI.0976-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aloe L, Properzi F, Probert L, Akassoglou K, Kassiotis G, Micera A, Fiore M. Learning abilities, NGF and BDNF brain levels in two lines of TNF-alpha transgenic mice, one characterized by neurological disorders, the other phenotypically normal. Brain Research. 1999;840:125–137. doi: 10.1016/s0006-8993(99)01748-5. [DOI] [PubMed] [Google Scholar]
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- Ang E, Chen J, Zagouras P, Magna H, Holland J, Schaeffer E, Nestler EJ. Induction of nuclear factor-kappaB in nucleus accumbens by chronic cocaine administration. J Neurochem. 2001;79:221–224. doi: 10.1046/j.1471-4159.2001.00563.x. [DOI] [PubMed] [Google Scholar]
- Arancio O, Zhang HP, Chen X, Lin C, Trinchese F, Puzzo D, Liu S, Hegde A, Yan SF, Stern A, Luddy JS, Lue LF, Walker DG, Roher A, Buttini M, Mucke L, Li W, Schmidt AM, Kindy M, Hyslop PA, Stern DM, Du Yan SS. RAGE potentiates Abeta-induced perturbation of neuronal function in transgenic mice. The EMBO journal. 2004;23:4096–4105. doi: 10.1038/sj.emboj.7600415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arezoomandan R, Haghparast A. Administration of the glial cell modulator, minocycline, in the nucleus accumbens attenuated the maintenance and reinstatement of morphine-seeking behavior. Can J Physiol Pharmacol. 2016;94:257–264. doi: 10.1139/cjpp-2015-0209. [DOI] [PubMed] [Google Scholar]
- Attarzadeh-Yazdi G, Arezoomandan R, Haghparast A. Minocycline, an antibiotic with inhibitory effect on microglial activation, attenuates the maintenance and reinstatement of methamphetamine-seeking behavior in rat. Prog Neuropsychopharmacol Biol Psychiatry. 2014;53:142–148. doi: 10.1016/j.pnpbp.2014.04.008. [DOI] [PubMed] [Google Scholar]
- Badanich KA, Becker HC, Woodward JJ. Effects of chronic intermittent ethanol exposure on orbitofrontal and medial prefrontal cortex-dependent behaviors in mice. Behav Neurosci. 2011;125:879–891. doi: 10.1037/a0025922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai L, Zhai C, Han K, Li Z, Qian J, Jing Y, Zhang W, Xu JT. Toll-like receptor 4-mediated nuclear factor-kappaB activation in spinal cord contributes to chronic morphine-induced analgesic tolerance and hyperalgesia in rats. Neurosci Bull. 2014;30:936–948. doi: 10.1007/s12264-014-1483-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajo M, Varodayan FP, Madamba SG, Robert AJ, Casal LM, Oleata CS, Siggins GR, Roberto M. IL-1 interacts with ethanol effects on GABAergic transmission in the mouse central amygdala. Front Pharmacol. 2015;6:49. doi: 10.3389/fphar.2015.00049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bala S, Marcos M, Gattu A, Catalano D, Szabo G. Acute binge drinking increases serum endotoxin and bacterial DNA levels in healthy individuals. PLoS One. 2014;9:e96864. doi: 10.1371/journal.pone.0096864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bala S, Marcos M, Kodys K, Csak T, Catalano D, Mandrekar P, Szabo G. Up-regulation of microRNA-155 in macrophages contributes to increased tumor necrosis factor {alpha} (TNF{alpha}) production via increased mRNA half-life in alcoholic liver disease. J Biol Chem. 2011;286:1436–1444. doi: 10.1074/jbc.M110.145870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banks WA, Kastin AJ, Broadwell RD. Passage of cytokines across the blood-brain barrier. Neuroimmunomodulation. 1995;2:241–248. doi: 10.1159/000097202. [DOI] [PubMed] [Google Scholar]
- Banks WA, Kastin AJ, Gutierrez EG. Penetration of interleukin-6 across the murine blood-brain barrier. Neurosci Lett. 1994;179:53–56. doi: 10.1016/0304-3940(94)90933-4. [DOI] [PubMed] [Google Scholar]
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- Beattie EC, Stellwagen D, Morishita W, Bresnahan JC, Ha BK, Von Zastrow M, Beattie MS, Malenka RC. Control of synaptic strength by glial TNFalpha. Science. 2002;295:2282–2285. doi: 10.1126/science.1067859. [DOI] [PubMed] [Google Scholar]
- Bell RL, Lopez MF, Cui C, Egli M, Johnson KW, Franklin KM, Becker HC. Ibudilast reduces alcohol drinking in multiple animal models of alcohol dependence. Addict Biol. 2015;20:38–42. doi: 10.1111/adb.12106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beynon SB, Walker FR. Microglial activation in the injured and healthy brain: what are we really talking about? Practical and theoretical issues associated with the measurement of changes in microglial morphology. Neuroscience. 2012;225:162–171. doi: 10.1016/j.neuroscience.2012.07.029. [DOI] [PubMed] [Google Scholar]
- Bhattacharya A, Derecki NC, Lovenberg TW, Drevets WC. Role of neuro-immunological factors in the pathophysiology of mood disorders. Psychopharmacology (Berl) 2016;233:1623–1636. doi: 10.1007/s00213-016-4214-0. [DOI] [PubMed] [Google Scholar]
- Bhattacharya A, Drevets WC. Role of Neuro-Immunological Factors in the Pathophysiology of Mood Disorders: Implications for Novel Therapeutics for Treatment Resistant Depression. Curr Top Behav Neurosci. 2017;31:339–356. doi: 10.1007/7854_2016_43. [DOI] [PubMed] [Google Scholar]
- Bi W, Zhu L, Wang C, Liang Y, Liu J, Shi Q, Tao E. Rifampicin inhibits microglial inflammation and improves neuron survival against inflammation. Brain Res. 2011;1395:12–20. doi: 10.1016/j.brainres.2011.04.019. [DOI] [PubMed] [Google Scholar]
- Bianchi ME. HMGB1 loves company. J Leukoc Biol. 2009;86:573–576. doi: 10.1189/jlb.1008585. [DOI] [PubMed] [Google Scholar]
- Bierhaus A, Wolf J, Andrassy M, Rohleder N, Humpert PM, Petrov D, Ferstl R, von Eynatten M, Wendt T, Rudofsky G, Joswig M, Morcos M, Schwaninger M, McEwen B, Kirschbaum C, Nawroth PP. A mechanism converting psychosocial stress into mononuclear cell activation. Proc Natl Acad Sci U S A. 2003;100:1920–1925. doi: 10.1073/pnas.0438019100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanc L, Castanier R, Mishra AK, Ray A, Besra GS, Sutcliffe I, Vercellone A, Nigou J. Gram-positive bacterial lipoglycans based on a glycosylated diacylglycerol lipid anchor are microbe-associated molecular patterns recognized by TLR2. PLoS One. 2013;8:e81593. doi: 10.1371/journal.pone.0081593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanco AM, Valles SL, Pascual M, Guerri C. Involvement of TLR4/type I IL-1 receptor signaling in the induction of inflammatory mediators and cell death induced by ethanol in cultured astrocytes. Journal of immunology. 2005;175:6893–6899. doi: 10.4049/jimmunol.175.10.6893. [DOI] [PubMed] [Google Scholar]
- Blednov YA, Benavidez JM, Black M, Harris RA. Inhibition of phosphodiesterase 4 reduces ethanol intake and preference in C57BL/6J mice. Front Neurosci. 2014;8:129. doi: 10.3389/fnins.2014.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blednov YA, Benavidez JM, Geil C, Perra S, Morikawa H, Harris RA. Activation of inflammatory signaling by lipopolysaccharide produces a prolonged increase of voluntary alcohol intake in mice. Brain, behavior, and immunity. 2011;25(Suppl 1):S92–S105. doi: 10.1016/j.bbi.2011.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blednov YA, Bergeson SE, Walker D, Ferreira VM, Kuziel WA, Harris RA. Perturbation of chemokine networks by gene deletion alters the reinforcing actions of ethanol. Behavioural brain research. 2005;165:110–125. doi: 10.1016/j.bbr.2005.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blednov YA, Ponomarev I, Geil C, Bergeson S, Koob GF, Harris RA. Neuroimmune regulation of alcohol consumption: behavioral validation of genes obtained from genomic studies. Addiction biology. 2012;17:108–120. doi: 10.1111/j.1369-1600.2010.00284.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bluthe RM, Walter V, Parnet P, Laye S, Lestage J, Verrier D, Poole S, Stenning BE, Kelley KW, Dantzer R. Lipopolysaccharide induces sickness behaviour in rats by a vagal mediated mechanism. C R Acad Sci III. 1994;317:499–503. [PubMed] [Google Scholar]
- Bonaccorso S, Marino V, Biondi M, Grimaldi F, Ippoliti F, Maes M. Depression induced by treatment with interferon-alpha in patients affected by hepatitis C virus. Journal of affective disorders. 2002;72:237–241. doi: 10.1016/s0165-0327(02)00264-1. [DOI] [PubMed] [Google Scholar]
- Borner C, Hollt V, Kraus J. Mechanisms of the inhibition of nuclear factor-kappaB by morphine in neuronal cells. Mol Pharmacol. 2012;81:587–597. doi: 10.1124/mol.111.076620. [DOI] [PubMed] [Google Scholar]
- Breese GR, Knapp DJ, Overstreet DH, Navarro M, Wills TA, Angel RA. Repeated lipopolysaccharide (LPS) or cytokine treatments sensitize ethanol withdrawal-induced anxiety-like behavior. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2008;33:867–876. doi: 10.1038/sj.npp.1301468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brubaker SW, Bonham KS, Zanoni I, Kagan JC. Innate immune pattern recognition: a cell biological perspective. Annu Rev Immunol. 2015;33:257–290. doi: 10.1146/annurev-immunol-032414-112240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bsibsi M, Bajramovic JJ, Vogt MH, van Duijvenvoorden E, Baghat A, Persoon-Deen C, Tielen F, Verbeek R, Huitinga I, Ryffel B, Kros A, Gerritsen WH, Amor S, van Noort JM. The microtubule regulator stathmin is an endogenous protein agonist for TLR3. J Immunol. 2010;184:6929–6937. doi: 10.4049/jimmunol.0902419. [DOI] [PubMed] [Google Scholar]
- Buwitt-Beckmann U, Heine H, Wiesmuller KH, Jung G, Brock R, Akira S, Ulmer AJ. TLR1-and TLR6-independent recognition of bacterial lipopeptides. J Biol Chem. 2006;281:9049–9057. doi: 10.1074/jbc.M512525200. [DOI] [PubMed] [Google Scholar]
- Calu DJ, Roesch MR, Stalnaker TA, Schoenbaum G. Associative encoding in posterior piriform cortex during odor discrimination and reversal learning. Cerebral cortex. 2007;17:1342–1349. doi: 10.1093/cercor/bhl045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen G, Ke Z, Xu M, Liao M, Wang X, Qi Y, Zhang T, Frank JA, Bower KA, Shi X, Luo J. Autophagy is a protective response to ethanol neurotoxicity. Autophagy. 2012;8:1577–1589. doi: 10.4161/auto.21376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Li G, Liu Y, Werth VP, Williams KJ, Liu ML. Translocation of Endogenous Danger Signal HMGB1 From Nucleus to Membrane Microvesicles in Macrophages. J Cell Physiol. 2016;231:2319–2326. doi: 10.1002/jcp.25352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cherry JD, Olschowka JA, O’Banion MK. Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. Journal of neuroinflammation. 2014;11:98. doi: 10.1186/1742-2094-11-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coleman LG, Jr, He J, Lee J, Styner M, Crews FT. Adolescent binge drinking alters adult brain neurotransmitter gene expression, behavior, brain regional volumes, and neurochemistry in mice. Alcoholism, clinical and experimental research. 2011;35:671–688. doi: 10.1111/j.1530-0277.2010.01385.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coleman LG, Jr, Zou J, Crews F. Microglial-derived miRNA Let-7 and HMGB1 contribute to ethanol-induced neurotoxicity via TLR7. J Neuroinflammation In Press. 2017 doi: 10.1186/s12974-017-0799-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costello EJ, Copeland WE, Shanahan L, Worthman CM, Angold A. C-reactive protein and substance use disorders in adolescence and early adulthood: a prospective analysis. Drug Alcohol Depend. 2013;133:712–717. doi: 10.1016/j.drugalcdep.2013.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cozzoli DK, Kaufman MN, Nipper MA, Hashimoto JG, Wiren KM, Finn DA. Functional regulation of PI3K-associated signaling in the accumbens by binge alcohol drinking in male but not female mice. Neuropharmacology. 2016;105:164–174. doi: 10.1016/j.neuropharm.2016.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews F, Nixon K, Kim D, Joseph J, Shukitt-Hale B, Qin L, Zou J. BHT blocks NF-kappaB activation and ethanol-induced brain damage. Alcoholism, clinical and experimental research. 2006;30:1938–1949. doi: 10.1111/j.1530-0277.2006.00239.x. [DOI] [PubMed] [Google Scholar]
- Crews FT, Boettiger CA. Impulsivity, frontal lobes and risk for addiction. Pharmacology, biochemistry, and behavior. 2009;93:237–247. doi: 10.1016/j.pbb.2009.04.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Qin L, Sheedy D, Vetreno RP, Zou J. High mobility group box 1/Toll-like receptor danger signaling increases brain neuroimmune activation in alcohol dependence. Biol Psychiatry. 2013a;73:602–612. doi: 10.1016/j.biopsych.2012.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Qin L, Sheedy D, Vetreno RP, Zou J. High mobility group box 1/Toll-like receptor danger signaling increases brain neuroimmune activation in alcohol dependence. Biological psychiatry. 2013b;73:602–612. doi: 10.1016/j.biopsych.2012.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Vetreno RP. Neuroimmune basis of alcoholic brain damage. Int Rev Neurobiol. 2014;118:315–357. doi: 10.1016/B978-0-12-801284-0.00010-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Vetreno RP. Mechanisms of neuroimmune gene induction in alcoholism. Psychopharmacology (Berl) 2016;233:1543–1557. doi: 10.1007/s00213-015-3906-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Zou J, Qin L. Induction of innate immune genes in brain create the neurobiology of addiction. Brain Behav Immun. 2011;25(Suppl 1):S4–S12. doi: 10.1016/j.bbi.2011.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui C, Noronha A, Warren KR, Koob GF, Sinha R, Thakkar M, Matochik J, Crews FT, Chandler LJ, Pfefferbaum A, Becker HC, Lovinger D, Everitt BJ, Egli M, Mandyam CD, Fein G, Potenza MN, Harris RA, Grant KA, Roberto M, Meyerhoff DJ, Sullivan EV. Brain pathways to recovery from alcohol dependence. Alcohol. 2015;49:435–452. doi: 10.1016/j.alcohol.2015.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czech B, Hannon GJ. Small RNA sorting: matchmaking for Argonautes. Nat Rev Genet. 2011;12:19–31. doi: 10.1038/nrg2916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Mello C, Le T, Swain MG. Cerebral microglia recruit monocytes into the brain in response to tumor necrosis factoralpha signaling during peripheral organ inflammation. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2009;29:2089–2102. doi: 10.1523/JNEUROSCI.3567-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nature reviews. Neuroscience. 2008;9:46–56. doi: 10.1038/nrn2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis RL, Syapin PJ. Ethanol increases nuclear factor-kappa B activity in human astroglial cells. Neuroscience letters. 2004;371:128–132. doi: 10.1016/j.neulet.2004.08.051. [DOI] [PubMed] [Google Scholar]
- de Jesus AA, Canna SW, Liu Y, Goldbach-Mansky R. Molecular mechanisms in genetically defined autoinflammatory diseases: disorders of amplified danger signaling. Annu Rev Immunol. 2015;33:823–874. doi: 10.1146/annurev-immunol-032414-112227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennis CV, Sheahan PJ, Graeber MB, Sheedy DL, Kril JJ, Sutherland GT. Microglial proliferation in the brain of chronic alcoholics with hepatic encephalopathy. Metab Brain Dis. 2014;29:1027–1039. doi: 10.1007/s11011-013-9469-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drew PD, Johnson JW, Douglas JC, Phelan KD, Kane CJ. Pioglitazone blocks ethanol induction of microglial activation and immune responses in the hippocampus, cerebellum, and cerebral cortex in a mouse model of fetal alcohol spectrum disorders. Alcohol Clin Exp Res. 2015;39:445–454. doi: 10.1111/acer.12639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ek M, Kurosawa M, Lundeberg T, Ericsson A. Activation of vagal afferents after intravenous injection of interleukin-1beta: role of endogenous prostaglandins. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1998;18:9471–9479. doi: 10.1523/JNEUROSCI.18-22-09471.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis FW. Effect of ethanol on plasma corticosterone levels. The Journal of pharmacology and experimental therapeutics. 1966;153:121–127. [PubMed] [Google Scholar]
- Fang F, Lue LF, Yan S, Xu H, Luddy JS, Chen D, Walker DG, Stern DM, Schmidt AM, Chen JX, Yan SS. RAGE-dependent signaling in microglia contributes to neuroinflammation, Abeta accumulation, and impaired learning/memory in a mouse model of Alzheimer’s disease. The FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2010;24:1043–1055. doi: 10.1096/fj.09-139634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farina C, Aloisi F, Meinl E. Astrocytes are active players in cerebral innate immunity. Trends in immunology. 2007;28:138–145. doi: 10.1016/j.it.2007.01.005. [DOI] [PubMed] [Google Scholar]
- Fernandez-Lizarbe S, Montesinos J, Guerri C. Ethanol induces TLR4/TLR2 association, triggering an inflammatory response in microglial cells. J Neurochem. 2013;126:261–273. doi: 10.1111/jnc.12276. [DOI] [PubMed] [Google Scholar]
- Fernandez-Lizarbe S, Pascual M, Guerri C. Critical role of TLR4 response in the activation of microglia induced by ethanol. Journal of immunology. 2009a;183:4733–4744. doi: 10.4049/jimmunol.0803590. [DOI] [PubMed] [Google Scholar]
- Fernandez-Lizarbe S, Pascual M, Guerri C. Critical role of TLR4 response in the activation of microglia induced by ethanol. J Immunol. 2009b;183:4733–4744. doi: 10.4049/jimmunol.0803590. [DOI] [PubMed] [Google Scholar]
- Ferrier L, Mazelin L, Cenac N, Desreumaux P, Janin A, Emilie D, Colombel JF, Garcia-Villar R, Fioramonti J, Bueno L. Stress-induced disruption of colonic epithelial barrier: role of interferon-gamma and myosin light chain kinase in mice. Gastroenterology. 2003;125:795–804. doi: 10.1016/s0016-5085(03)01057-6. [DOI] [PubMed] [Google Scholar]
- Fortier CB, Steffen EM, Lafleche G, Venne JR, Disterhoft JF, McGlinchey RE. Delay discrimination and reversal eyeblink classical conditioning in abstinent chronic alcoholics. Neuropsychology. 2008;22:196–208. doi: 10.1037/0894-4105.22.2.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank MG, Baratta MV, Sprunger DB, Watkins LR, Maier SF. Microglia serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine responses. Brain, behavior, and immunity. 2007;21:47–59. doi: 10.1016/j.bbi.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Frank MG, Miguel ZD, Watkins LR, Maier SF. Prior exposure to glucocorticoids sensitizes the neuroinflammatory and peripheral inflammatory responses to E. coli lipopolysaccharide. Brain, behavior, and immunity. 2010;24:19–30. doi: 10.1016/j.bbi.2009.07.008. [DOI] [PubMed] [Google Scholar]
- Franke H. Influence of chronic alcohol treatment on the GFAP-immunoreactivity in astrocytes of the hippocampus in rats. Acta Histochem. 1995;97:263–271. doi: 10.1016/S0065-1281(11)80187-X. [DOI] [PubMed] [Google Scholar]
- Freeman K, Brureau A, Vadigepalli R, Staehle MM, Brureau MM, Gonye GE, Hoek JB, Hooper DC, Schwaber JS. Temporal changes in innate immune signals in a rat model of alcohol withdrawal in emotional and cardiorespiratory homeostatic nuclei. Journal of neuroinflammation. 2012;9:97. doi: 10.1186/1742-2094-9-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garate I, Garcia-Bueno B, Madrigal JL, Caso JR, Alou L, Gomez-Lus ML, Mico JA, Leza JC. Stress-induced neuroinflammation: role of the Toll-like receptor-4 pathway. Biological psychiatry. 2013;73:32–43. doi: 10.1016/j.biopsych.2012.07.005. [DOI] [PubMed] [Google Scholar]
- George FR. The role of arachidonic acid metabolites in mediating ethanol self-administration and intoxication. Ann N Y Acad Sci. 1989;559:382–391. doi: 10.1111/j.1749-6632.1989.tb22624.x. [DOI] [PubMed] [Google Scholar]
- Giraldo E, Multhoff G, Ortega E. Noradrenaline increases the expression and release of Hsp72 by human neutrophils. Brain, behavior, and immunity. 2010;24:672–677. doi: 10.1016/j.bbi.2010.02.003. [DOI] [PubMed] [Google Scholar]
- Goshen I, Kreisel T, Ounallah-Saad H, Renbaum P, Zalzstein Y, Ben-Hur T, Levy-Lahad E, Yirmiya R. A dual role for interleukin-1 in hippocampal-dependent memory processes. Psychoneuroendocrinology. 2007;32:1106–1115. doi: 10.1016/j.psyneuen.2007.09.004. [DOI] [PubMed] [Google Scholar]
- Hanke ML, Kielian T. Toll-like receptors in health and disease in the brain: mechanisms and therapeutic potential. Clinical science. 2011;121:367–387. doi: 10.1042/CS20110164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison NA, Brydon L, Walker C, Gray MA, Steptoe A, Critchley HD. Inflammation causes mood changes through alterations in subgenual cingulate activity and mesolimbic connectivity. Biological psychiatry. 2009;66:407–414. doi: 10.1016/j.biopsych.2009.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison NA, Doeller CF, Voon V, Burgess N, Critchley HD. Peripheral inflammation acutely impairs human spatial memory via actions on medial temporal lobe glucose metabolism. Biological psychiatry. 2014;76:585–593. doi: 10.1016/j.biopsych.2014.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He J, Crews FT. Increased MCP-1 and microglia in various regions of the human alcoholic brain. Experimental neurology. 2008;210:349–358. doi: 10.1016/j.expneurol.2007.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heberlein A, Kaser M, Lichtinghagen R, Rhein M, Lenz B, Kornhuber J, Bleich S, Hillemacher T. TNF-alpha and IL-6 serum levels: neurobiological markers of alcohol consumption in alcohol-dependent patients? Alcohol. 2014;48:671–676. doi: 10.1016/j.alcohol.2014.08.003. [DOI] [PubMed] [Google Scholar]
- Hill AS, Sahay A, Hen R. Increasing Adult Hippocampal Neurogenesis is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2015;40:2368–2378. doi: 10.1038/npp.2015.85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hori O, Brett J, Slattery T, Cao R, Zhang J, Chen JX, Nagashima M, Lundh ER, Vijay S, Nitecki D, et al. The receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin. Mediation of neurite outgrowth and co-expression of rage and amphoterin in the developing nervous system. J Biol Chem. 1995;270:25752–25761. doi: 10.1074/jbc.270.43.25752. [DOI] [PubMed] [Google Scholar]
- Hreggvidsdottir HS, Ostberg T, Wahamaa H, Schierbeck H, Aveberger AC, Klevenvall L, Palmblad K, Ottosson L, Andersson U, Harris HE. The alarmin HMGB1 acts in synergy with endogenous and exogenous danger signals to promote inflammation. J Leukoc Biol. 2009;86:655–662. doi: 10.1189/jlb.0908548. [DOI] [PubMed] [Google Scholar]
- Hu W, Lu T, Chen A, Huang Y, Hansen R, Chandler LJ, Zhang HT. Inhibition of phosphodiesterase-4 decreases ethanol intake in mice. Psychopharmacology (Berl) 2011;218:331–339. doi: 10.1007/s00213-011-2290-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hulsmans M, Holvoet P. MicroRNA-containing microvesicles regulating inflammation in association with atherosclerotic disease. Cardiovasc Res. 2013;100:7–18. doi: 10.1093/cvr/cvt161. [DOI] [PubMed] [Google Scholar]
- Hutchinson MR, Zhang Y, Shridhar M, Evans JH, Buchanan MM, Zhao TX, Slivka PF, Coats BD, Rezvani N, Wieseler J, Hughes TS, Landgraf KE, Chan S, Fong S, Phipps S, Falke JJ, Leinwand LA, Maier SF, Yin H, Rice KC, Watkins LR. Evidence that opioids may have toll-like receptor 4 and MD-2 effects. Brain, behavior, and immunity. 2010;24:83–95. doi: 10.1016/j.bbi.2009.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izquierdo A, Brigman JL, Radke AK, Rudebeck PH, Holmes A. The neural basis of reversal learning: An updated perspective. Neuroscience. 2016 doi: 10.1016/j.neuroscience.2016.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson AC, Rossiter JP, Lafon M. Expression of Toll-like receptor 3 in the human cerebellar cortex in rabies, herpes simplex encephalitis, and other neurological diseases. J Neurovirol. 2006;12:229–234. doi: 10.1080/13550280600848399. [DOI] [PubMed] [Google Scholar]
- Jang E, Kim JH, Lee S, Seo JW, Jin M, Lee MG, Jang IS, Lee WH, Suk K. Phenotypic polarization of activated astrocytes: the critical role of lipocalin-2 in the classical inflammatory activation of astrocytes. Journal of immunology. 2013;191:5204–5219. doi: 10.4049/jimmunol.1301637. [DOI] [PubMed] [Google Scholar]
- Janko C, Filipovic M, Munoz LE, Schorn C, Schett G, Ivanovic-Burmazovic I, Herrmann M. Redox modulation of HMGB1-related signaling. Antioxid Redox Signal. 2014;20:1075–1085. doi: 10.1089/ars.2013.5179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen CJ, Massie A, De Keyser J. Immune players in the CNS: the astrocyte. Journal of neuroimmune pharmacology: the official journal of the Society on NeuroImmune Pharmacology. 2013;8:824–839. doi: 10.1007/s11481-013-9480-6. [DOI] [PubMed] [Google Scholar]
- Jimenez JL, Punzon C, Navarro J, Munoz-Fernandez MA, Fresno M. Phosphodiesterase 4 inhibitors prevent cytokine secretion by T lymphocytes by inhibiting nuclear factor-kappaB and nuclear factor of activated T cells activation. J Pharmacol Exp Ther. 2001;299:753–759. [PubMed] [Google Scholar]
- Jokisch D, Roser P, Juckel G, Daum I, Bellebaum C. Impairments in learning by monetary rewards and alcohol-associated rewards in detoxified alcoholic patients. Alcohol Clin Exp Res. 2014;38:1947–1954. doi: 10.1111/acer.12460. [DOI] [PubMed] [Google Scholar]
- June HL, Liu J, Warnock KT, Bell KA, Balan I, Bollino D, Puche A, Aurelian L. CRF-amplified neuronal TLR4/MCP-1 signaling regulates alcohol self-administration. Neuropsychopharmacology. 2015;40:1549–1559. doi: 10.1038/npp.2015.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaltschmidt B, Kaltschmidt C. NF-KappaB in Long-Term Memory and Structural Plasticity in the Adult Mammalian Brain. Front Mol Neurosci. 2015;8:69. doi: 10.3389/fnmol.2015.00069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kane CJ, Phelan KD, Han L, Smith RR, Xie J, Douglas JC, Drew PD. Protection of neurons and microglia against ethanol in a mouse model of fetal alcohol spectrum disorders by peroxisome proliferator-activated receptor-gamma agonists. Brain Behav Immun. 2011;25(Suppl 1):S137–145. doi: 10.1016/j.bbi.2011.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keshavarzian A, Farhadi A, Forsyth CB, Rangan J, Jakate S, Shaikh M, Banan A, Fields JZ. Evidence that chronic alcohol exposure promotes intestinal oxidative stress, intestinal hyperpermeability and endotoxemia prior to development of alcoholic steatohepatitis in rats. J Hepatol. 2009;50:538–547. doi: 10.1016/j.jhep.2008.10.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kettenmann H, Hanisch UK, Noda M, Verkhratsky A. Physiology of microglia. Physiological Reviews. 2011;91:461–553. doi: 10.1152/physrev.00011.2010. [DOI] [PubMed] [Google Scholar]
- Kettenmann H, Kirchhoff F, Verkhratsky A. Microglia: new roles for the synaptic stripper. Neuron. 2013;77:10–18. doi: 10.1016/j.neuron.2012.12.023. [DOI] [PubMed] [Google Scholar]
- Khairova RA, Machado-Vieira R, Du J, Manji HK. A potential role for pro-inflammatory cytokines in regulating synaptic plasticity in major depressive disorder. The international journal of neuropsychopharmacology / official scientific journal of the Collegium Internationale Neuropsychopharmacologicum. 2009;12:561–578. doi: 10.1017/S1461145709009924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khakh BS, Sofroniew MV. Diversity of astrocyte functions and phenotypes in neural circuits. Nature neuroscience. 2015;18:942–952. doi: 10.1038/nn.4043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim C, Ho DH, Suk JE, You S, Michael S, Kang J, Joong Lee S, Masliah E, Hwang D, Lee HJ, Lee SJ. Neuron-released oligomeric alpha-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia. Nat Commun. 2013;4:1562. doi: 10.1038/ncomms2534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knapp DJ, Crews FT. Induction of cyclooxygenase-2 in brain during acute and chronic ethanol treatment and ethanol withdrawal. Alcoholism, clinical and experimental research. 1999;23:633–643. [PubMed] [Google Scholar]
- Koo JW, Duman RS. IL-1beta is an essential mediator of the antineurogenic and anhedonic effects of stress. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:751–756. doi: 10.1073/pnas.0708092105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koob GF, Le Moal M. Plasticity of reward neurocircuitry and the ‘dark side’ of drug addiction. Nature neuroscience. 2005;8:1442–1444. doi: 10.1038/nn1105-1442. [DOI] [PubMed] [Google Scholar]
- Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2010;35:217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreisel T, Frank MG, Licht T, Reshef R, Ben-Menachem-Zidon O, Baratta MV, Maier SF, Yirmiya R. Dynamic microglial alterations underlie stress-induced depressive-like behavior and suppressed neurogenesis. Molecular Psychiatry. 2014;19:699–709. doi: 10.1038/mp.2013.155. [DOI] [PubMed] [Google Scholar]
- Lacagnina MJ, Rivera PD, Bilbo SD. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug use and Abuse. Neuropsychopharmacology. 2016 doi: 10.1038/npp.2016.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert KG, Gerecke KM, Quadros PS, Doudera E, Jasnow AM, Kinsley CH. Activity-stress increases density of GFAP-immunoreactive astrocytes in the rat hippocampus. Stress. 2000;3:275–284. doi: 10.3109/10253890009001133. [DOI] [PubMed] [Google Scholar]
- Lawrimore C, Crews F. Ethanol, TLR3, and TLR4 agonists have unique innate immune responses in neuron-like SH-SY5Y and microglia-like BV2. Alcoholism Clinical Experimental Research In Press. 2017 doi: 10.1111/acer.13368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leclercq S, De Saeger C, Delzenne N, de Timary P, Starkel P. Role of inflammatory pathways, blood mononuclear cells, and gut-derived bacterial products in alcohol dependence. Biol Psychiatry. 2014;76:725–733. doi: 10.1016/j.biopsych.2014.02.003. [DOI] [PubMed] [Google Scholar]
- Lehmann SM, Kruger C, Park B, Derkow K, Rosenberger K, Baumgart J, Trimbuch T, Eom G, Hinz M, Kaul D, Habbel P, Kalin R, Franzoni E, Rybak A, Nguyen D, Veh R, Ninnemann O, Peters O, Nitsch R, Heppner FL, Golenbock D, Schott E, Ploegh HL, Wulczyn FG, Lehnardt S. An unconventional role for miRNA: let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat Neurosci. 2012a;15:827–835. doi: 10.1038/nn.3113. [DOI] [PubMed] [Google Scholar]
- Lehmann SM, Rosenberger K, Kruger C, Habbel P, Derkow K, Kaul D, Rybak A, Brandt C, Schott E, Wulczyn FG, Lehnardt S. Extracellularly delivered single-stranded viral RNA causes neurodegeneration dependent on TLR7. J Immunol. 2012b;189:1448–1458. doi: 10.4049/jimmunol.1201078. [DOI] [PubMed] [Google Scholar]
- Lewohl JM, Nunez YO, Dodd PR, Tiwari GR, Harris RA, Mayfield RD. Up-regulation of microRNAs in brain of human alcoholics. Alcohol Clin Exp Res. 2011;35:1928–1937. doi: 10.1111/j.1530-0277.2011.01544.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim SW, Shiue YL, Liao JC, Wee HY, Wang CC, Chio CC, Chang CH, Hu CY, Kuo JR. Simvastatin Therapy in the Acute Stage of Traumatic Brain Injury Attenuates Brain Trauma-Induced Depression-Like Behavior in Rats by Reducing Neuroinflammation in the Hippocampus. Neurocrit Care. 2016 doi: 10.1007/s12028-016-0290-6. [DOI] [PubMed] [Google Scholar]
- Lippai D, Bala S, Csak T, Kurt-Jones EA, Szabo G. Chronic alcohol-induced microRNA-155 contributes to neuroinflammation in a TLR4-dependent manner in mice. PLoS One. 2013a;8:e70945. doi: 10.1371/journal.pone.0070945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lippai D, Bala S, Petrasek J, Csak T, Levin I, Kurt-Jones EA, Szabo G. Alcohol-induced IL-1beta in the brain is mediated by NLRP3/ASC inflammasome activation that amplifies neuroinflammation. J Leukoc Biol. 2013b;94:171–182. doi: 10.1189/jlb.1212659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu A, Fang H, Dirsch O, Jin H, Dahmen U. Oxidation of HMGB1 causes attenuation of its pro-inflammatory activity and occurs during liver ischemia and reperfusion. PLoS One. 2012;7:e35379. doi: 10.1371/journal.pone.0035379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Du L, Hong JS. Naloxone protects rat dopaminergic neurons against inflammatory damage through inhibition of microglia activation and superoxide generation. J Pharmacol Exp Ther. 2000a;293:607–617. [PubMed] [Google Scholar]
- Liu B, Du L, Kong LY, Hudson PM, Wilson BC, Chang RC, Abel HH, Hong JS. Reduction by naloxone of lipopolysaccharide-induced neurotoxicity in mouse cortical neuron-glia co-cultures. Neuroscience. 2000b;97:749–756. doi: 10.1016/s0306-4522(00)00057-9. [DOI] [PubMed] [Google Scholar]
- Liu J, Yang AR, Kelly T, Puche A, Esoga C, June HL, Jr, Elnabawi A, Merchenthaler I, Sieghart W, June HL, Sr, Aurelian L. Binge alcohol drinking is associated with GABAA alpha2-regulated Toll-like receptor 4 (TLR4) expression in the central amygdala. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:4465–4470. doi: 10.1073/pnas.1019020108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W, Crews FT. Adolescent intermittent ethanol exposure enhances ethanol activation of the nucleus accumbens while blunting the prefrontal cortex responses in adult rat. Neuroscience. 2015;293:92–108. doi: 10.1016/j.neuroscience.2015.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maczurek A, Shanmugam K, Munch G. Inflammation and the redox-sensitive AGE-RAGE pathway as a therapeutic target in Alzheimer’s disease. Annals of the New York Academy of Sciences. 2008;1126:147–151. doi: 10.1196/annals.1433.026. [DOI] [PubMed] [Google Scholar]
- Madrigal JL, Garcia-Bueno B, Moro MA, Lizasoain I, Lorenzo P, Leza JC. Relationship between cyclooxygenase-2 and nitric oxide synthase-2 in rat cortex after stress. The European journal of neuroscience. 2003;18:1701–1705. doi: 10.1046/j.1460-9568.2003.02888.x. [DOI] [PubMed] [Google Scholar]
- Madrigal JL, Moro MA, Lizasoain I, Lorenzo P, Leza JC. Stress-induced increase in extracellular sucrose space in rats is mediated by nitric oxide. Brain research. 2002;938:87–91. doi: 10.1016/s0006-8993(02)02467-8. [DOI] [PubMed] [Google Scholar]
- Maes M, Berk M, Goehler L, Song C, Anderson G, Galecki P, Leonard B. Depression and sickness behavior are Janus-faced responses to shared inflammatory pathways. BMC medicine. 2012;10:66. doi: 10.1186/1741-7015-10-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2000;20:9104–9110. doi: 10.1523/JNEUROSCI.20-24-09104.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mao XR, Moerman-Herzog AM, Chen Y, Barger SW. Unique aspects of transcriptional regulation in neurons–nuances in NFkappaB and Sp1-related factors. Journal of neuroinflammation. 2009;6:16. doi: 10.1186/1742-2094-6-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maroso M, Balosso S, Ravizza T, Liu J, Bianchi ME, Vezzani A. Interleukin-1 type 1 receptor/Toll-like receptor signalling in epilepsy: the importance of IL-1beta and high-mobility group box 1. J Intern Med. 2011;270:319–326. doi: 10.1111/j.1365-2796.2011.02431.x. [DOI] [PubMed] [Google Scholar]
- Marshall SA, Casachahua JD, Rinker JA, Blose AK, Lysle DT, Thiele TE. IL-1 receptor signaling in the basolateral amygdala modulates binge-like ethanol consumption in male C57BL/6J mice. Brain Behav Immun. 2016a;51:258–267. doi: 10.1016/j.bbi.2015.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall SA, Geil CR, Nixon K. Prior Binge Ethanol Exposure Potentiates the Microglial Response in a Model of Alcohol-Induced Neurodegeneration. Brain Sci. 2016b;6 doi: 10.3390/brainsci6020016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall SA, McClain JA, Kelso ML, Hopkins DM, Pauly JR, Nixon K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiology of disease. 2013;54:239–251. doi: 10.1016/j.nbd.2012.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall SA, McKnight KH, Blose AK, Lysle DT, Thiele TE. Modulation of Binge-like Ethanol Consumption by IL-10 Signaling in the Basolateral Amygdala. J Neuroimmune Pharmacol. 2016c doi: 10.1007/s11481-016-9709-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maslanik T, Mahaffey L, Tannura K, Beninson L, Greenwood BN, Fleshner M. The inflammasome and danger associated molecular patterns (DAMPs) are implicated in cytokine and chemokine responses following stressor exposure. Brain, behavior, and immunity. 2013;28:54–62. doi: 10.1016/j.bbi.2012.10.014. [DOI] [PubMed] [Google Scholar]
- Mayfield J, Ferguson L, Harris RA. Neuroimmune signaling: a key component of alcohol abuse. Current opinion in neurobiology. 2013;23:513–520. doi: 10.1016/j.conb.2013.01.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merlot E, Moze E, Dantzer R, Neveu PJ. Suppression of restraint-induced plasma cytokines in mice pretreated with LPS. Stress. 2002;5:131–135. doi: 10.1080/10253890290027921. [DOI] [PubMed] [Google Scholar]
- Miller AH, Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nature reviews. Immunology. 2016;16:22–34. doi: 10.1038/nri.2015.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller S, Ronfani L, Bianchi ME. Regulated expression and subcellular localization of HMGB1, a chromatin protein with a cytokine function. J Intern Med. 2004;255:332–343. doi: 10.1111/j.1365-2796.2003.01296.x. [DOI] [PubMed] [Google Scholar]
- Mulligan MK, Ponomarev I, Hitzemann RJ, Belknap JK, Tabakoff B, Harris RA, Crabbe JC, Blednov YA, Grahame NJ, Phillips TJ, Finn DA, Hoffman PL, Iyer VR, Koob GF, Bergeson SE. Toward understanding the genetics of alcohol drinking through transcriptome meta-analysis. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:6368–6373. doi: 10.1073/pnas.0510188103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northcutt AL, Hutchinson MR, Wang X, Baratta MV, Hiranita T, Cochran TA, Pomrenze MB, Galer EL, Kopajtic TA, Li CM, Amat J, Larson G, Cooper DC, Huang Y, O’Neill CE, Yin H, Zahniser NR, Katz JL, Rice KC, Maier SF, Bachtell RK, Watkins LR. DAT isn’t all that: cocaine reward and reinforcement require Toll-like receptor 4 signaling. Mol Psychiatry. 2015;20:1525–1537. doi: 10.1038/mp.2014.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunez YO, Mayfield RD. Understanding Alcoholism Through microRNA Signatures in Brains of Human Alcoholics. Front Genet. 2012;3:43. doi: 10.3389/fgene.2012.00043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunez YO, Truitt JM, Gorini G, Ponomareva ON, Blednov YA, Harris RA, Mayfield RD. Positively correlated miRNA-mRNA regulatory networks in mouse frontal cortex during early stages of alcohol dependence. BMC Genomics. 2013;14:725. doi: 10.1186/1471-2164-14-725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connor JC, Lawson MA, Andre C, Moreau M, Lestage J, Castanon N, Kelley KW, Dantzer R. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Molecular Psychiatry. 2009;14:511–522. doi: 10.1038/sj.mp.4002148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obernier JA, White AM, Swartzwelder HS, Crews FT. Cognitive deficits and CNS damage after a 4-day binge ethanol exposure in rats. Pharmacology, biochemistry, and behavior. 2002;72:521–532. doi: 10.1016/s0091-3057(02)00715-3. [DOI] [PubMed] [Google Scholar]
- Ohshima K, Inoue K, Fujiwara A, Hatakeyama K, Kanto K, Watanabe Y, Muramatsu K, Fukuda Y, Ogura S, Yamaguchi K, Mochizuki T. Let-7 microRNA family is selectively secreted into the extracellular environment via exosomes in a metastatic gastric cancer cell line. PLoS One. 2010;5:e13247. doi: 10.1371/journal.pone.0013247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okuma Y, Liu K, Wake H, Liu R, Nishimura Y, Hui Z, Teshigawara K, Haruma J, Yamamoto Y, Yamamoto H, Date I, Takahashi HK, Mori S, Nishibori M. Glycyrrhizin inhibits traumatic brain injury by reducing HMGB1-RAGE interaction. Neuropharmacology. 2014;85:18–26. doi: 10.1016/j.neuropharm.2014.05.007. [DOI] [PubMed] [Google Scholar]
- Okun E, Griffioen K, Barak B, Roberts NJ, Castro K, Pita MA, Cheng A, Mughal MR, Wan R, Ashery U, Mattson MP. Toll-like receptor 3 inhibits memory retention and constrains adult hippocampal neurogenesis. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:15625–15630. doi: 10.1073/pnas.1005807107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okvist A, Johansson S, Kuzmin A, Bazov I, Merino-Martinez R, Ponomarev I, Mayfield RD, Harris RA, Sheedy D, Garrick T, Harper C, Hurd YL, Terenius L, Ekstrom TJ, Bakalkin G, Yakovleva T. Neuroadaptations in human chronic alcoholics: dysregulation of the NF-kappaB system. PLoS One. 2007;2:e930. doi: 10.1371/journal.pone.0000930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overman EL, Rivier JE, Moeser AJ. CRF induces intestinal epithelial barrier injury via the release of mast cell proteases and TNF-alpha. PLoS One. 2012;7:e39935. doi: 10.1371/journal.pone.0039935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999;18:6853–6866. doi: 10.1038/sj.onc.1203239. [DOI] [PubMed] [Google Scholar]
- Park A, Ghezzi A, Thilini P, Wijesekera, Atkinson N. Genetics and Genomics of Alcohol Responses in Drosophila. Neuropharmacology. 2017 doi: 10.1016/j.neuropharm.2017.01.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park CK, Xu ZZ, Berta T, Han Q, Chen G, Liu XJ, Ji RR. Extracellular microRNAs activate nociceptor neurons to elicit pain via TLR7 and TRPA1. Neuron. 2014;82:47–54. doi: 10.1016/j.neuron.2014.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pascual M, Balino P, Alfonso-Loeches S, Aragon CM, Guerri C. Impact of TLR4 on behavioral and cognitive dysfunctions associated with alcohol-induced neuroinflammatory damage. Brain, behavior, and immunity. 2011a;25(Suppl 1):S80–91. doi: 10.1016/j.bbi.2011.02.012. [DOI] [PubMed] [Google Scholar]
- Pascual M, Balino P, Alfonso-Loeches S, Aragon CM, Guerri C. Impact of TLR4 on behavioral and cognitive dysfunctions associated with alcohol-induced neuroinflammatory damage. Brain Behav Immun. 2011b;25(Suppl 1):S80–91. doi: 10.1016/j.bbi.2011.02.012. [DOI] [PubMed] [Google Scholar]
- Pascual M, Blanco AM, Cauli O, Minarro J, Guerri C. Intermittent ethanol exposure induces inflammatory brain damage and causes long-term behavioural alterations in adolescent rats. The European journal of neuroscience. 2007a;25:541–550. doi: 10.1111/j.1460-9568.2006.05298.x. [DOI] [PubMed] [Google Scholar]
- Pascual M, Blanco AM, Cauli O, Minarro J, Guerri C. Intermittent ethanol exposure induces inflammatory brain damage and causes long-term behavioural alterations in adolescent rats. Eur J Neurosci. 2007b;25:541–550. doi: 10.1111/j.1460-9568.2006.05298.x. [DOI] [PubMed] [Google Scholar]
- Pekny M, Pekna M. Astrocyte reactivity and reactive astrogliosis: costs and benefits. Physiological Reviews. 2014;94:1077–1098. doi: 10.1152/physrev.00041.2013. [DOI] [PubMed] [Google Scholar]
- Perry VH, Holmes C. Microglial priming in neurodegenerative disease. Nature reviews. Neurology. 2014;10:217–224. doi: 10.1038/nrneurol.2014.38. [DOI] [PubMed] [Google Scholar]
- Pietri M, Schneider B, Mouillet-Richard S, Ermonval M, Mutel V, Launay JM, Kellermann O. Reactive oxygen species-dependent TNF-alpha converting enzyme activation through stimulation of 5-HT2B and alpha1D autoreceptors in neuronal cells. FASEB J. 2005;19:1078–1087. doi: 10.1096/fj.04-3631com. [DOI] [PubMed] [Google Scholar]
- Pla A, Pascual M, Guerri C. Autophagy Constitutes a Protective Mechanism against Ethanol Toxicity in Mouse Astrocytes and Neurons. PLoS One. 2016;11:e0153097. doi: 10.1371/journal.pone.0153097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plane JM, Shen Y, Pleasure DE, Deng W. Prospects for minocycline neuroprotection. Archives of neurology. 2010;67:1442–1448. doi: 10.1001/archneurol.2010.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiang M, Ticku MK. Role of AP-1 in ethanol-induced N-methyl-D-aspartate receptor 2B subunit gene up-regulation in mouse cortical neurons. J Neurochem. 2005;95:1332–1341. doi: 10.1111/j.1471-4159.2005.03464.x. [DOI] [PubMed] [Google Scholar]
- Qin L, Crews FT. Chronic ethanol increases systemic TLR3 agonist-induced neuroinflammation and neurodegeneration. J Neuroinflammation. 2012a;9:130. doi: 10.1186/1742-2094-9-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Crews FT. Chronic ethanol increases systemic TLR3 agonist-induced neuroinflammation and neurodegeneration. Journal of neuroinflammation. 2012b;9:130. doi: 10.1186/1742-2094-9-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Crews FT. NADPH oxidase and reactive oxygen species contribute to alcohol-induced microglial activation and neurodegeneration. J Neuroinflammation. 2012c;9:5. doi: 10.1186/1742-2094-9-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Crews FT. NADPH oxidase and reactive oxygen species contribute to alcohol-induced microglial activation and neurodegeneration. Journal of neuroinflammation. 2012d;9:5. doi: 10.1186/1742-2094-9-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, He J, Hanes RN, Pluzarev O, Hong JS, Crews FT. Increased systemic and brain cytokine production and neuroinflammation by endotoxin following ethanol treatment. J Neuroinflammation. 2008a;5:10. doi: 10.1186/1742-2094-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, He J, Hanes RN, Pluzarev O, Hong JS, Crews FT. Increased systemic and brain cytokine production and neuroinflammation by endotoxin following ethanol treatment. Journal of neuroinflammation. 2008b;5:10. doi: 10.1186/1742-2094-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Liu Y, Hong JS, Crews FT. NADPH oxidase and aging drive microglial activation, oxidative stress, and dopaminergic neurodegeneration following systemic LPS administration. Glia. 2013;61:855–868. doi: 10.1002/glia.22479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Wu X, Block ML, Liu Y, Breese GR, Hong JS, Knapp DJ, Crews FT. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia. 2007;55:453–462. doi: 10.1002/glia.20467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajkowska G, Stockmeier CA. Astrocyte pathology in major depressive disorder: insights from human postmortem brain tissue. Current drug targets. 2013;14:1225–1236. doi: 10.2174/13894501113149990156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reissner KJ, Kalivas PW. Using glutamate homeostasis as a target for treating addictive disorders. Behavioural pharmacology. 2010;21:514–522. doi: 10.1097/FBP.0b013e32833d41b2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocha NP, de Miranda AS, Teixeira AL. Insights into Neuroinflammation in Parkinson’s Disease: From Biomarkers to Anti-Inflammatory Based Therapies. Biomed Res Int. 2015;2015:628192. doi: 10.1155/2015/628192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubio-Araiz A, Porcu F, Perez-Hernandez M, Garcia-Gutierrez MS, Aracil-Fernandez MA, Gutierrez-Lopez MD, Guerri C, Manzanares J, O’Shea E, Colado MI. Disruption of blood-brain barrier integrity in postmortem alcoholic brain: preclinical evidence of TLR4 involvement from a binge-like drinking model. Addiction biology. 2016 doi: 10.1111/adb.12376. [DOI] [PubMed] [Google Scholar]
- Russo SJ, Wilkinson MB, Mazei-Robison MS, Dietz DM, Maze I, Krishnan V, Renthal W, Graham A, Birnbaum SG, Green TA, Robison B, Lesselyong A, Perrotti LI, Bolanos CA, Kumar A, Clark MS, Neumaier JF, Neve RL, Bhakar AL, Barker PA, Nestler EJ. Nuclear factor kappa B signaling regulates neuronal morphology and cocaine reward. J Neurosci. 2009;29:3529–3537. doi: 10.1523/JNEUROSCI.6173-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryan SM, Nolan YM. Neuroinflammation negatively affects adult hippocampal neurogenesis and cognition: can exercise compensate? Neuroscience and biobehavioral reviews. 2016;61:121–131. doi: 10.1016/j.neubiorev.2015.12.004. [DOI] [PubMed] [Google Scholar]
- Saha B, Momen-Heravi F, Kodys K, Szabo G. MicroRNA Cargo of Extracellular Vesicles from Alcohol-exposed Monocytes Signals Naive Monocytes to Differentiate into M2 Macrophages. J Biol Chem. 2016;291:149–159. doi: 10.1074/jbc.M115.694133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science. 2003;301:805–809. doi: 10.1126/science.1083328. [DOI] [PubMed] [Google Scholar]
- Saunders PR, Santos J, Hanssen NP, Yates D, Groot JA, Perdue MH. Physical and psychological stress in rats enhances colonic epithelial permeability via peripheral CRH. Dig Dis Sci. 2002;47:208–215. doi: 10.1023/a:1013204612762. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Roesch MR, Stalnaker TA. Orbitofrontal cortex, decision-making and drug addiction. Trends in neurosciences. 2006;29:116–124. doi: 10.1016/j.tins.2005.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoenbaum G, Saddoris MP, Ramus SJ, Shaham Y, Setlow B. Cocaine-experienced rats exhibit learning deficits in a task sensitive to orbitofrontal cortex lesions. The European journal of neuroscience. 2004;19:1997–2002. doi: 10.1111/j.1460-9568.2004.03274.x. [DOI] [PubMed] [Google Scholar]
- Setiawan E, Wilson AA, Mizrahi R, Rusjan PM, Miler L, Rajkowska G, Suridjan I, Kennedy JL, Rekkas PV, Houle S, Meyer JH. Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes. JAMA psychiatry. 2015;72:268–275. doi: 10.1001/jamapsychiatry.2014.2427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, Gould E. Neurogenesis in the adult is involved in the formation of trace memories. Nature. 2001;410:372–376. doi: 10.1038/35066584. [DOI] [PubMed] [Google Scholar]
- Sironi L, Banfi C, Brioschi M, Gelosa P, Guerrini U, Nobili E, Gianella A, Paoletti R, Tremoli E, Cimino M. Activation of NF-kB and ERK1/2 after permanent focal ischemia is abolished by simvastatin treatment. Neurobiol Dis. 2006;22:445–451. doi: 10.1016/j.nbd.2005.12.004. [DOI] [PubMed] [Google Scholar]
- Sorrells SF, Caso JR, Munhoz CD, Sapolsky RM. The stressed CNS: when glucocorticoids aggravate inflammation. Neuron. 2009;64:33–39. doi: 10.1016/j.neuron.2009.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stalnaker TA, Takahashi Y, Roesch MR, Schoenbaum G. Neural substrates of cognitive inflexibility after chronic cocaine exposure. Neuropharmacology. 2009;56(Suppl 1):63–72. doi: 10.1016/j.neuropharm.2008.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steptoe A, Hamer M, Chida Y. The effects of acute psychological stress on circulating inflammatory factors in humans: a review and meta-analysis. Brain, behavior, and immunity. 2007;21:901–912. doi: 10.1016/j.bbi.2007.03.011. [DOI] [PubMed] [Google Scholar]
- Stevenson JR, Schroeder JP, Nixon K, Besheer J, Crews FT, Hodge CW. Abstinence following alcohol drinking produces depression-like behavior and reduced hippocampal neurogenesis in mice. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2009;34:1209–1222. doi: 10.1038/npp.2008.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storer PD, Xu J, Chavis J, Drew PD. Peroxisome proliferator-activated receptor-gamma agonists inhibit the activation of microglia and astrocytes: implications for multiple sclerosis. J Neuroimmunol. 2005;161:113–122. doi: 10.1016/j.jneuroim.2004.12.015. [DOI] [PubMed] [Google Scholar]
- Stott K, Watson M, Howe FS, Grossmann JG, Thomas JO. Tail-mediated collapse of HMGB1 is dynamic and occurs via differential binding of the acidic tail to the A and B domains. J Mol Biol. 2010;403:706–722. doi: 10.1016/j.jmb.2010.07.045. [DOI] [PubMed] [Google Scholar]
- Szabo G, Bala S, Petrasek J, Gattu A. Gut-liver axis and sensing microbes. Dig Dis. 2010;28:737–744. doi: 10.1159/000324281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabo G, Lippai D. Converging actions of alcohol on liver and brain immune signaling. Int Rev Neurobiol. 2014;118:359–380. doi: 10.1016/B978-0-12-801284-0.00011-7. [DOI] [PubMed] [Google Scholar]
- Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140:805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- Tancredi V, D’Arcangelo G, Grassi F, Tarroni P, Palmieri G, Santoni A, Eusebi F. Tumor necrosis factor alters synaptic transmission in rat hippocampal slices. Neuroscience letters. 1992;146:176–178. doi: 10.1016/0304-3940(92)90071-e. [DOI] [PubMed] [Google Scholar]
- Teitelbaum AA, Gareau MG, Jury J, Yang PC, Perdue MH. Chronic peripheral administration of corticotropin-releasing factor causes colonic barrier dysfunction similar to psychological stress. Am J Physiol Gastrointest Liver Physiol. 2008;295:G452–459. doi: 10.1152/ajpgi.90210.2008. [DOI] [PubMed] [Google Scholar]
- Thakur V, McMullen MR, Pritchard MT, Nagy LE. Regulation of macrophage activation in alcoholic liver disease. Journal of gastroenterology and hepatology. 2007;22(Suppl 1):S53–56. doi: 10.1111/j.1440-1746.2006.04650.x. [DOI] [PubMed] [Google Scholar]
- Thomas JO, Stott K. H1 and HMGB1: modulators of chromatin structure. Biochem Soc Trans. 2012;40:341–346. doi: 10.1042/BST20120014. [DOI] [PubMed] [Google Scholar]
- Thounaojam MC, Kaushik DK, Basu A. MicroRNAs in the brain: it’s regulatory role in neuroinflammation. Mol Neurobiol. 2013;47:1034–1044. doi: 10.1007/s12035-013-8400-3. [DOI] [PubMed] [Google Scholar]
- Torres-Platas SG, Cruceanu C, Chen GG, Turecki G, Mechawar N. Evidence for increased microglial priming and macrophage recruitment in the dorsal anterior cingulate white matter of depressed suicides. Brain, behavior, and immunity. 2014 doi: 10.1016/j.bbi.2014.05.007. [DOI] [PubMed] [Google Scholar]
- Townshend JM, Duka T. Mixed emotions: alcoholics’ impairments in the recognition of specific emotional facial expressions. Neuropsychologia. 2003;41:773–782. doi: 10.1016/s0028-3932(02)00284-1. [DOI] [PubMed] [Google Scholar]
- Turchinovich A, Samatov TR, Tonevitsky AG, Burwinkel B. Circulating miRNAs: cell-cell communication function? Front Genet. 2013;4:119. doi: 10.3389/fgene.2013.00119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tynan RJ, Naicker S, Hinwood M, Nalivaiko E, Buller KM, Pow DV, Day TA, Walker FR. Chronic stress alters the density and morphology of microglia in a subset of stress-responsive brain regions. Brain, behavior, and immunity. 2010;24:1058–1068. doi: 10.1016/j.bbi.2010.02.001. [DOI] [PubMed] [Google Scholar]
- Vabulas RM, Braedel S, Hilf N, Singh-Jasuja H, Herter S, Ahmad-Nejad P, Kirschning CJ, Da Costa C, Rammensee HG, Wagner H, Schild H. The endoplasmic reticulum-resident heat shock protein Gp96 activates dendritic cells via the Toll-like receptor 2/4 pathway. The Journal of biological chemistry. 2002;277:20847–20853. doi: 10.1074/jbc.M200425200. [DOI] [PubMed] [Google Scholar]
- Valenta JP, Gonzales RA. Chronic Intracerebroventricular Infusion of Monocyte Chemoattractant Protein-1 Leads to a Persistent Increase in Sweetened Ethanol Consumption During Operant Self-Administration But Does Not Influence Sucrose Consumption in Long-Evans Rats. Alcoholism, clinical and experimental research. 2016;40:187–195. doi: 10.1111/acer.12928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valles SL, Blanco AM, Pascual M, Guerri C. Chronic ethanol treatment enhances inflammatory mediators and cell death in the brain and in astrocytes. Brain pathology. 2004;14:365–371. doi: 10.1111/j.1750-3639.2004.tb00079.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venereau E, Casalgrandi M, Schiraldi M, Antoine DJ, Cattaneo A, De Marchis F, Liu J, Antonelli A, Preti A, Raeli L, Shams SS, Yang H, Varani L, Andersson U, Tracey KJ, Bachi A, Uguccioni M, Bianchi ME. Mutually exclusive redox forms of HMGB1 promote cell recruitment or proinflammatory cytokine release. J Exp Med. 2012;209:1519–1528. doi: 10.1084/jem.20120189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetreno RP, Crews FT. Adolescent binge drinking increases expression of the danger signal receptor agonist HMGB1 and Toll-like receptors in the adult prefrontal cortex. Neuroscience. 2012;226:475–488. doi: 10.1016/j.neuroscience.2012.08.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetreno RP, Crews FT. Current hypotheses on the mechanisms of alcoholism. Handb Clin Neurol. 2014;125:477–497. doi: 10.1016/B978-0-444-62619-6.00027-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetreno RP, Qin L, Crews FT. Increased receptor for advanced glycation end product expression in the human alcoholic prefrontal cortex is linked to adolescent drinking. Neurobiology of disease. 2013a;59:52–62. doi: 10.1016/j.nbd.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetreno RP, Qin L, Crews FT. Increased receptor for advanced glycation end product expression in the human alcoholic prefrontal cortex is linked to adolescent drinking. Neurobiol Dis. 2013b;59:52–62. doi: 10.1016/j.nbd.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow ND, Koob GF, McLellan AT. Neurobiologic Advances from the Brain Disease Model of Addiction. N Engl J Med. 2016;374:363–371. doi: 10.1056/NEJMra1511480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Bernhardi R, Eugenin-von Bernhardi L, Eugenin J. Microglial cell dysregulation in brain aging and neurodegeneration. Front Aging Neurosci. 2015;7:124. doi: 10.3389/fnagi.2015.00124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagley Y, Hwang CK, Lin HY, Kam AF, Law PY, Loh HH, Wei LN. Inhibition of c-Jun NH2-terminal kinase stimulates mu opioid receptor expression via p38 MAPK-mediated nuclear NF-kappaB activation in neuronal and non-neuronal cells. Biochim Biophys Acta. 2013;1833:1476–1488. doi: 10.1016/j.bbamcr.2013.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang WY, Tan MS, Yu JT, Tan L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann Transl Med. 2015;3:136. doi: 10.3978/j.issn.2305-5839.2015.03.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Grace PM, Pham MN, Cheng K, Strand KA, Smith C, Li J, Watkins LR, Yin H. Rifampin inhibits Toll-like receptor 4 signaling by targeting myeloid differentiation protein 2 and attenuates neuropathic pain. FASEB J. 2013;27:2713–2722. doi: 10.1096/fj.12-222992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Zhang Y, Peng Y, Hutchinson MR, Rice KC, Yin H, Watkins LR. Pharmacological characterization of the opioid inactive isomers (+)-naltrexone and (+)-naloxone as antagonists of toll-like receptor 4. Br J Pharmacol. 2016;173:856–869. doi: 10.1111/bph.13394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward RJ, Colivicchi MA, Allen R, Schol F, Lallemand F, de Witte P, Ballini C, Corte LD, Dexter D. Neuro-inflammation induced in the hippocampus of ‘binge drinking’ rats may be mediated by elevated extracellular glutamate content. Journal of neurochemistry. 2009;111:1119–1128. doi: 10.1111/j.1471-4159.2009.06389.x. [DOI] [PubMed] [Google Scholar]
- Ward RJ, Zhang Y, Crichton RR, Piret B, Piette J, de Witte P. Identification of the nuclear transcription factor NFkappaB in rat after in vivo ethanol administration. FEBS letters. 1996;389:119–122. doi: 10.1016/0014-5793(96)00545-5. [DOI] [PubMed] [Google Scholar]
- Warden A, Mayfield RD. Gene expression profiling in the human alcoholic brain. Neuropharmacology. 2017 doi: 10.1016/j.neuropharm.2017.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber MD, Frank MG, Tracey KJ, Watkins LR, Maier SF. Stress induces the danger-associated molecular pattern HMGB-1 in the hippocampus of male Sprague Dawley rats: a priming stimulus of microglia and the NLRP3 inflammasome. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2015;35:316–324. doi: 10.1523/JNEUROSCI.3561-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weissenborn R, Duka T. Acute alcohol effects on cognitive function in social drinkers: their relationship to drinking habits. Psychopharmacology. 2003;165:306–312. doi: 10.1007/s00213-002-1281-1. [DOI] [PubMed] [Google Scholar]
- Wilson JS, Mruthinti S, Buccafusco JJ, Schade RF, Mitchell MB, Harrell DU, Gulati NK, Miller LS. Anti-RAGE and Abeta immunoglobulin levels are related to dementia level and cognitive performance. The journals of gerontology Series A, Biological sciences and medical sciences. 2009;64:264–271. doi: 10.1093/gerona/gln002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wohleb ES, Hanke ML, Corona AW, Powell ND, Stiner LM, Bailey MT, Nelson RJ, Godbout JP, Sheridan JF. beta-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity induced by repeated social defeat. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2011;31:6277–6288. doi: 10.1523/JNEUROSCI.0450-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wohleb ES, Powell ND, Godbout JP, Sheridan JF. Stress-induced recruitment of bone marrow-derived monocytes to the brain promotes anxiety-like behavior. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2013;33:13820–13833. doi: 10.1523/JNEUROSCI.1671-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worley MJ, Heinzerling KG, Roche DJ, Shoptaw S. Ibudilast attenuates subjective effects of methamphetamine in a placebo-controlled inpatient study. Drug Alcohol Depend. 2016;162:245–250. doi: 10.1016/j.drugalcdep.2016.02.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue J, Schmidt SV, Sander J, Draffehn A, Krebs W, Quester I, De Nardo D, Gohel TD, Emde M, Schmidleithner L, Ganesan H, Nino-Castro A, Mallmann MR, Labzin L, Theis H, Kraut M, Beyer M, Latz E, Freeman TC, Ulas T, Schultze JL. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity. 2014;40:274–288. doi: 10.1016/j.immuni.2014.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanai H, Ban T, Wang Z, Choi MK, Kawamura T, Negishi H, Nakasato M, Lu Y, Hangai S, Koshiba R, Savitsky D, Ronfani L, Akira S, Bianchi ME, Honda K, Tamura T, Kodama T, Taniguchi T. HMGB proteins function as universal sentinels for nucleic-acid-mediated innate immune responses. Nature. 2009;462:99–103. doi: 10.1038/nature08512. [DOI] [PubMed] [Google Scholar]
- Yang H, Lundback P, Ottosson L, Erlandsson-Harris H, Venereau E, Bianchi ME, Al-Abed Y, Andersson U, Tracey KJ, Antoine DJ. Redox modification of cysteine residues regulates the cytokine activity of high mobility group box-1 (HMGB1) Mol Med. 2012;18:250–259. doi: 10.2119/molmed.2011.00389. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Yelamanchili SV, Lamberty BG, Rennard DA, Morsey BM, Hochfelder CG, Meays BM, Levy E, Fox HS. MiR-21 in Extracellular Vesicles Leads to Neurotoxicity via TLR7 Signaling in SIV Neurological Disease. PLoS Pathog. 2015;11:e1005032. doi: 10.1371/journal.ppat.1005032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yirmiya R, Goshen I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain, behavior, and immunity. 2011;25:181–213. doi: 10.1016/j.bbi.2010.10.015. [DOI] [PubMed] [Google Scholar]
- Young JJ, Bruno D, Pomara N. A review of the relationship between proinflammatory cytokines and major depressive disorder. J Affect Disord. 2014;169:15–200. doi: 10.1016/j.jad.2014.07.032. [DOI] [PubMed] [Google Scholar]
- Zhang B, Bailey WM, Kopper TJ, Orr MB, Feola DJ, Gensel JC. Azithromycin drives alternative macrophage activation and improves recovery and tissue sparing in contusion spinal cord injury. J Neuroinflammation. 2015;12:218. doi: 10.1186/s12974-015-0440-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu CB, Lindler KM, Owens AW, Daws LC, Blakely RD, Hewlett WA. Interleukin-1 receptor activation by systemic lipopolysaccharide induces behavioral despair linked to MAPK regulation of CNS serotonin transporters. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2010;35:2510–2520. doi: 10.1038/npp.2010.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou J, Crews F. CREB and NF-kappaB transcription factors regulate sensitivity to excitotoxic and oxidative stress induced neuronal cell death. Cellular and molecular neurobiology. 2006;26:385–405. doi: 10.1007/s10571-006-9045-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou J, Crews F. Induction of innate immune gene expression cascades in brain slice cultures by ethanol: key role of NF-kappaB and proinflammatory cytokines. Alcoholism, clinical and experimental research. 2010;34:777–789. doi: 10.1111/j.1530-0277.2010.01150.x. [DOI] [PubMed] [Google Scholar]
- Zou JY, Crews FT. TNF alpha potentiates glutamate neurotoxicity by inhibiting glutamate uptake in organotypic brain slice cultures: neuroprotection by NF kappa B inhibition. Brain research. 2005;1034:11–24. doi: 10.1016/j.brainres.2004.11.014. [DOI] [PubMed] [Google Scholar]
- Zou JY, Crews FT. Release of neuronal HMGB1 by ethanol through decreased HDAC activity activates brain neuroimmune signaling. PLoS One. 2014;9:e87915. doi: 10.1371/journal.pone.0087915. [DOI] [PMC free article] [PubMed] [Google Scholar]