

Abstract
The photoreceptor photoactive yellow protein (PYP) was used as a model system to study receptor activation and protein folding. Refolding was studied by stopped-flow absorbance spectroscopy for PYP with either a trans or a cis chromophore. Chromophore trans to cis isomerization, the mechanism of light detection by PYP, greatly affects the protein folding process. When the cis chromophore is present, refolding from the unfolded state proceeds through the putative signaling state of PYP as an on-pathway intermediate. In addition, moderate denaturant concentrations result in the specific unfolding of the signaling state of PYP. Thus, the signaling state is common to the pathways of folding and signaling. This result provides an avenue for the study of protein folding. We demonstrate how this approach can be used to establish whether a folding intermediate is on-pathway or off-pathway. The results also reveal transient partial unfolding as a molecular mechanism for signaling.
To become functionally active, a protein needs to fold into its native three-dimensional structure. Protein folding has been investigated extensively, but many of its aspects remain unresolved. The properties of folding intermediates and their role in the folding mechanism as either productive on-pathway intermediates or dispensable off-pathway intermediates have proven to be difficult to unambiguously determine for many proteins (see refs. 1–4). Here evidence is reported for the hypothesis that protein folding and signaling are directly related. We demonstrate that this provides an approach to experimental studies on protein folding, using a small water soluble receptor protein that is activated by an external stimulus: unfolding and refolding reactions can be triggered not only by using traditional rapid mixing methods, but also by activation of the receptor. Thus, two independent pathways are available to populate the same folding intermediate. The existence, properties, and position in the folding pathway of the intermediate as deduced from rapid mixing studies therefore can be directly and independently tested. The use of a photoreceptor in this approach provides the additional benefit that signaling is triggered by exposure of the receptor to light, allowing for time-resolved measurements on the receptor activation process.
To study the relationship between protein folding and signaling, we used photoactive yellow protein (PYP) as a model system. PYP is a small water-soluble photoreceptor from purple bacteria (5–7) that displays rhodopsin-like photochemistry (5, 8) based on its unique p-coumaric acid chromophore (9, 10). PYP is a member of the PAS domain family that is involved in regulation, sensing, and the circadian rhythm (11, 12). The involvement of PAS domains in a wide range of biological responses has triggered studies on PYP aimed at understanding the signaling mechanism of this ubiquitous signaling module. Absorbance of a photon initiates a photocycle in PYP by the trans to cis photoisomerization of its chromophore (13–16). This triggers a number of transitions in the photoreceptor that result in its activation. Subsequently, PYP recovers spontaneously to its initial state within a second (5, 8). The longest lived photocycle intermediate, characterized by a blue-shifted absorbance spectrum, is considered to be the signaling state of PYP (5, 6, 14). This proposal is supported by the resemblance of the blue-shifted photocycle intermediate of PYP to the S373 photocycle intermediate of the archaeal sensory rhodopsin I, which has been experimentally determined to be a signaling state (see ref. 17). Here we address the question to what extent the pathways for protein folding and light-triggered signaling in PYP are common.
Materials and Methods
Nomenclature.
We use the following nomenclature for the different states of PYP: the protein can be in the initial groundstate (pG), or in photocycle intermediates with a red- or blue-shifted absorbance spectrum (pR and pB, respectively), or may be fully unfolded (pU). The isomerization state of the chromophore is indicated as a superscript. Thus, fully unfolded PYP containing the chromophore in the cis configuration is indicated as pUcis.
Materials.
PYP was overexpressed in Escherichia coli and purified as described (14). All kinetic and equilibrium experiments were carried out by using 8 μM PYP in 20 mM potassium phosphate buffer (pH 7.3).
Equilibrium Denaturant Titrations.
Equilibrium denaturant titrations of PYP with guanidinium-hydrochloride (GdmHCl) and urea were performed at 25°C by using a Cary 300 UV-Vis spectrophotomer (Varian). The observed transitions were analyzed by using Eq. 1, as derived in ref. 18:
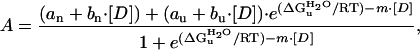 |
1 |
where A is the measured absorbance at 446 nm, 375, or 340 nm; [D] is the denaturant concentration; bn and bu are the slopes of the baselines before and after the transition; an and au are the signal intensities of these sloping baseline at zero denaturant; ΔG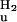
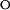 is the free energy of unfolding in water, m is the denaturant dependence of free energy per mol of denaturant; R is the gas constant; and T is the temperature in Kelvin. The two sloping baselines describe the dependence of the absorbance of the native and unfolded states of PYP on denaturant concentration. The transition between the native and unfolded states of PYP is described by using the approximation that the free energy for unfolding ΔGu depends linearly (with slope m) on denaturant concentration.
is the free energy of unfolding in water, m is the denaturant dependence of free energy per mol of denaturant; R is the gas constant; and T is the temperature in Kelvin. The two sloping baselines describe the dependence of the absorbance of the native and unfolded states of PYP on denaturant concentration. The transition between the native and unfolded states of PYP is described by using the approximation that the free energy for unfolding ΔGu depends linearly (with slope m) on denaturant concentration.
For the analysis of pBcis denaturation, steady-state light-induced difference spectra were measured and normalized to equal amounts of pGtrans bleaching at 446 nm. In these normalized difference spectra pBcis denaturation was monitored at 375 nm, where the difference in absorbance between pBcis and pUcis is maximal. We investigated the effect of a small correction of the resulting absorbance values at 375 nm as a function of denaturant concentration by taking into account the low level of pGtrans denaturation at the highest denaturant concentrations. The values for ΔG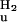
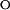 and m were essentially unaffected by this correction of the data.
and m were essentially unaffected by this correction of the data.
Measurement of Photocycle Kinetics.
Recovery kinetics of the PYP photocycle at different denaturant concentrations at 25°C after 20 sec of continuous illumination with actinic light were recorded by using either a Hewlett–Packard 8453 diode array spectrophotometer or a Cary 300 spectrophotomer at 446 and 340 nm.
CD Spectroscopy.
CD spectroscopy was performed with a 1-mm pathlength cuvette and a PYP concentration of 16 μM by using a Jasco J-715 spectropolarimeter. The recovery of pG after 20 sec actinic light illumination on closure of an optical shutter was monitored at various wavelengths. A second optical shutter was used to protect the photomultiplier tube from scattered actinic light.
Stopped-Flow Absorbance Spectroscopy.
Stopped-flow absorbance spectroscopy of PYP unfolding and refolding was performed by using a SX-18MV stopped-flow spectrophotometer (Applied Photophysics, Surrey, U.K.). Jumps to a range of GdmHCl concentrations were obtained by using a mixing ratio of 1:1 or 1:10. To investigate the effect of the chromophore isomerization state on the refolding process, pUtrans was exposed to UV-A light from a Cuda I-150 light source for 4 min to photoisomerize the chromophore, yielding pUcis. Stopped-flow absorbance traces were recorded at 445 and 340 nm, and analyzed by using the SX-18MV software.
The refolding kinetics of pUtrans were found to usually contain a slow component of small amplitude (≈10%). This slow phase may be because of proline isomerization (PYP contains four Pro residues). In this paper the fast phase of the kinetics is investigated. Stopped-flow experiments performed at 5°C revealed indications of an intermediate on the folding pathway for pUtrans.
Analysis of Chevron Plots.
A widely used approach to investigate the pathway of protein folding is to perform rapid mixing experiments in which the denaturant concentration is increased to a high value, revealing the kinetics of unfolding, or decreased to a low value, showing the kinetics of refolding. The dependence of the kinetics of unfolding and refolding is measured as a function of the final denaturant concentration, yielding a chevron plot. The dependence of kinetics on denaturant concentration reveals the nature of a transition: unfolding reactions involving the exposure of groups to solvent are accelerated at increased denaturant concentrations, whereas refolding transitions are decelerated. In the absence of folding intermediates, the logarithm of the (un)folding rate constants has been found for a wide range of proteins to depend linearly on denaturant concentration. The chevron plots for the pG to pUtrans transitions in PYP reported here were fitted to a two-state model (U ↔ F) by using Eq. 2, where kobs is the observed rate constant at different GdmHCl concentration and [D] is the GdmHCl concentration. In this two-state model, k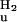
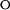 and k
and k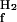
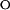 are the unfolding and refolding rate constants in the absence of denaturant, and mF and mU are the denaturant dependence of these rate constants:
are the unfolding and refolding rate constants in the absence of denaturant, and mF and mU are the denaturant dependence of these rate constants:
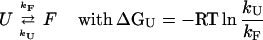 |
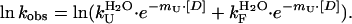 |
2 |
The chevron plot for the pUcis to pG transition deviated from linearity (see below) and was analyzed by using two different models involving a folding intermediate I. Two fundamentally different models for such three-state folding exist: one in which the folding intermediate is on the productive folding pathway from U to F, the second in which the intermediate I is a nonproductive off-pathway intermediate. The three-state on-pathway (U ↔ I → F) and off-pathway (I ↔ U → F) intermediate models were described by Eqs. 3 and 4, respectively, as derived in refs. 19–21:
 |
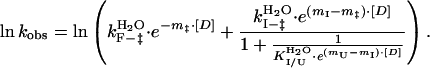 |
3 |
 |
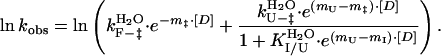 |
4 |
In the case of a three-state on-pathway intermediate model, the k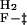
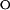 and k
and k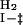
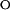 are the unfolding and refolding rate constants in the absence of denaturant in which transitions between the intermediate (I) and the native state (F) occur. The mU, mI, and m‡ describe the denaturant dependence of the free energy levels of U, I, and the transition state between F and I. K
are the unfolding and refolding rate constants in the absence of denaturant in which transitions between the intermediate (I) and the native state (F) occur. The mU, mI, and m‡ describe the denaturant dependence of the free energy levels of U, I, and the transition state between F and I. K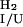
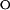 is the preequilibrium constant of the rapid U to I transition ([I]/[U]). For the three-state off-pathway model (see the scheme above) an analogous nomenclature is used.
is the preequilibrium constant of the rapid U to I transition ([I]/[U]). For the three-state off-pathway model (see the scheme above) an analogous nomenclature is used.
Results and Discussions
Probing Protein Folding in PYP.
On the conversion of the native state of PYP to the fully unfolded state by the addition of denaturants, the chromophore of PYP becomes solvent exposed and is protonated, causing a strong blue-shift in the absorbance spectrum of PYP (5, 8, 10). We used this large spectral shift to monitor protein folding transitions in PYP in both equilibrium and kinetic rapid mixing experiments. The effect of the isomerization state of the chromophore on the refolding of PYP was investigated by exploiting its intrinsic photochemical activity. Fully unfolded PYP was first exposed to light to photoisomerize the chromophore and subsequently was used in rapid mixing refolding experiments. The observed transitions were compared with those triggered by the photoexcitation of PYP. Both the rapid-mixing folding and light-triggered photocycle experiments were performed as a function of denaturant concentration (chevron analysis; ref. 22) by using urea and GmdHCl, the two most common denaturants.
The kinetics of unfolding and refolding of PYP containing a trans chromophore were determined by stopped-flow absorbance spectroscopy. Folding transitions between the fully unfolded and native states of PYP initiated by rapid mixing display monoexponential kinetics (Fig. 1A). The dependence of the kinetics of these transitions on denaturant concentration were found to exhibit a typical V-shaped pattern (22), in which the logarithm of the kinetics depend linearly on denaturant concentration (Fig. 1B). At 2.72 M GdmHCl the rates for protein folding and unfolding are identical, corresponding to the midpoint of the equilibrium titration (Fig. 1C). Both the monoexponential kinetics and the good correspondence between equilibrium and kinetic experiments indicate that folding of PYP containing a trans chromophore is a two-state transition without the involvement of folding intermediates.
Figure 1.

Protein folding in PYP. (A) The kinetics of refolding in PYP observed by stopped-flow absorbance spectroscopy. Refolding was initiated by a jump in denaturant concentration, obtained by rapid mixing, from 3.5 to 1.0 M GdmHCl at pH 7.0. The resulting kinetics were monitored at 340 (dashed lines) and 445 (solid lines) nm for the refolding of pUtrans (monoexponential decay with k = 31.4 s−1) and a mixture of pUtrans and pUcis generated by the illumination of pUtrans with near-UV light [biexponential decay with k1 = 31.4 s−1 (68%) and k2 = 0.9 s−1 (32%)], The kinetics of the pBcis to pGtrans photocycle transition (○; monoexponential decay with k = 0.9 s−1) were determined under identical conditions, and normalized to match the amplitude of the rapid mixing signals. (B) Chevron analysis of protein folding in PYP. The kinetics of pGtrans unfolding (●) and of pUtrans (●) and pUcis (▴) refolding were determined as a function of GdmHCl concentration. The kinetics of the pBcis to pGtrans photocycle kinetics (○) were determined under identical conditions. The rapid mixing data were fit (solid lines, see Tables 1 and 2) to a two-state model for PYP containing a trans chromophore, and to a model containing an on-pathway intermediate for PYP containing a cis chromophore. (C) Equilibrium denaturation titration of pGtrans to pUtrans with GdmHCl, followed by visible absorbance spectroscopy at 340 nm (○) and 446 nm (▵).
The Chromophore Isomerization State Greatly Affects Folding Kinetics in PYP.
Refolding of PYP containing a cis chromophore was studied by UV-A illumination of fully unfolded PYP to generate a stable mixture of the trans and cis conformations of the chromophore. After illumination, refolding exhibited biexponential kinetics. The fast phase is identical to that observed for PYP containing a trans chromophore, whereas the new slow phase reports the kinetics of refolding for PYP with a cis chromophore (Fig. 1A). The dependence of the kinetics of the slow phase on denaturant concentration shows that isomerization of the chromophore to the cis conformation dramatically affects the refolding process (Fig. 1B). First, refolding is slowed down by 1–3 orders of magnitude. Second, the dependence of the kinetics on the concentration of GdmHCl no longer is linear. This demonstrates that chromophore isomerization not only initiates the PYP photocycle, but also strongly affects the folding pathway of PYP.
Our interpretation of the nonlinear dependence of the kinetics of refolding of the unfolded state containing a cis chromophore is the occurrence of an on-pathway folding intermediate that is in rapid preequilibrium with the unfolded state (19, 20). The data are accurately described by this proposal (Fig. 1B) with realistic parameters (Table 1). Analysis of the data with the on-pathway model indicates that the free energy difference between the unfolded state and the folding intermediate is zero at 1.75 M GdmHCl, resulting in the observed kink in the chevron plot.
Table 1.
Dependence of protein-folding kinetics in PYP on the concentration of GdmHCl
| Process | m‡ (M−1) |
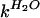 (s−1) (s−1) |
|---|---|---|
| Two-state transition | ||
| pGtrans→pUtrans | mU = 2.3 ± 0.05 |
k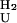 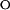 = (8.0 ± 1.5)* 10−5 = (8.0 ± 1.5)* 10−5
|
| pUtrans→pGtrans | mF = 3.4 ± 0.05 |
k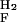 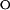 = 814 ± 66 = 814 ± 66 |
| On-pathway intermediate | ||
| I*→pGtrans | mI − m‡ = 0.9 ± 0.2 |
k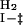 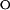 = 2.5 ± 0.3 = 2.5 ± 0.3 |
| pBcis→pGtrans† | mpB − m‡ = 0.5 ± 0.2 |
k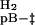 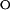 = 1.7 ± 0.2 = 1.7 ± 0.2 |
| Off-pathway intermediate | ||
| Ucis→pGtrans | mU − m‡ = 5.1 ± 0.6 |
k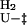  = 4211 ± 5258 = 4211 ± 5258 |
The folding intermediate I observed by rapid mixing is shown here to be identical to the blue-shifted photocycle intermediate.
Obtained from analysis of photocycle kinetics.
An alternative model is the occurrence of an off-pathway folding intermediate. For a number of proteins it has proven to be difficult to conclusively determine whether a folding intermediate is on-pathway or off-pathway, and the same difficulty is encountered in interpreting the rapid mixing data reported here for PYP. The off-pathway model results in an estimation of a protein refolding rate in water of 4,211 s−1 for PYP containing a cis chromophore. The folding rate for PYP containing a trans chromophore in water is five times slower (814 s−1). Because the native state of PYP contains a trans chromophore, it seems unlikely that isomerization of the chromophore to cis would result in a 5-fold acceleration of the folding rate. This consideration provides an argument supporting, but not demonstrating (see ref. 21), the on-pathway nature of the folding intermediate observed for PYP containing a cis chromophore. Below we show how a direct link between folding and light-triggered signaling in PYP can be used to demonstrate the on-pathway nature of the folding intermediate.
Folding and Signaling Share the Same Pathway in PYP.
We examined the possibility that the intermediate in PYP refolding from the fully unfolded state with a cis chromophore would be identical to the signaling state of PYP. To test this proposal, we determined the kinetics of the final photocycle transition, from the blue-shifted intermediate to native PYP, as a function of GdmHCl concentration. The observed photocycle kinetics are essentially indistinguishable from those observed for refolding (Fig. 1). This provides strong support for the proposal that folding and signaling share the same pathway in PYP.
A possibility that needs to be considered is that both the signaling state and the native state of PYP are fully folded, and that the last PYP photocycle transition involves structural rearrangements but not refolding. In this case, denaturants would not slow down this transition (see ref. 23). This is in contrast with the observed deceleration in the recovery of native PYP at increased denaturant concentrations (Fig. 1B). Therefore, under physiological conditions refolding of PYP containing a cis chromophore proceeds via a partially folded intermediate. This demonstrates that the final PYP photocycle transition is a protein refolding process, in line with a number of results obtained on the signaling state (24–26).
The link between protein folding and receptor activation in PYP was exploited to examine whether the observed folding intermediate is on-pathway or off-pathway. Continuous illumination of PYP results in the steady-state accumulation of the signaling state. This allowed us to directly study the preequilibrium, deduced from the chevron analysis, between the signaling state and the fully unfolded state containing a cis chromophore. To probe this preequilibrium, the UV/vis absorbance spectroscopic properties of the blue-shifted intermediate were determined at a range of denaturant concentrations. At each denaturant concentration PYP was photoexcited to yield the blue-shifted photocycle intermediate, and the resulting absorbance difference spectrum was measured. Under native conditions, the signaling state displays an absorbance maximum at 355 nm, whereas fully unfolded PYP absorbs maximally at 338 nm (8, 10). Therefore, a shift in the absorbance spectrum of the blue-shifted photocycle intermediate from 355 nm to 338 nm was expected at denaturant concentrations, which fully unfold the signaling state. Indeed, at moderate denaturant concentrations such a shift was observed (data not shown), revealing the cooperative unfolding of the signaling state (Fig. 2A). The midpoint for the denaturation of the signaling state, i.e., the GdmHCl concentration at which unfolding free energy for the signaling state is zero, was found to be 1.75 M. This is identical to the value obtained from the analysis of the denaturant dependence of the refolding kinetics obtained by using the on-pathway intermediate model.
Figure 2.

Specific denaturation of the pBcis photocycle intermediate. (A) Equilibrium denaturant titrations of pGtrans (squares) and pBcis (circles) with GdmHCl (closed symbols) and urea (open symbols). The pGtrans denaturation curve was determined from absorbance data at 340 and 446 nm; pBcis denaturation was monitored at 375 nm in the pBcis-pGtrans absorbance difference spectrum. (B) CD spectroscopy of the pUcis state (●) was performed by probing the CD signal during pGtrans recovery after PYP photoexcitation at various wavelengths. The depicted values were obtained by extrapolation to t = 0 (i.e., immediately after closure of the optical shutter for sample illumination) for pUcis (●) and t = ∞ (i.e., after complete pGtrans recovery) for pGtrans (○). The pUcis values were scaled to 100% pGtrans photobleaching, as determined from the measured absorbance values at 446 nm. For comparison, the steady-state CD spectrum of the pGtrans state in the presence of 0 (solid line) and 6.0 (dotted line) M urea are shown. (C) The monoexponential recovery of pGtrans in the presence of 6.0 M urea, detected by CD spectroscopy at 222 nm. (D) The amplitude of the light-induced loss of CD signal at 222 nm extrapolated to t = 0 (●) was determined as a function of urea concentration and scaled to 100% pGtrans photobleaching. ○ depict the dependence of the CD signal for pGtrans at 222 nm.
Based on the chevron analysis of the refolding kinetics of the fully unfolded state containing a cis chromophore, it is difficult to determine whether the native state is formed directly from the unfolded state or from the signaling state as an on-pathway folding intermediate. However, extensive investigations on PYP have shown that under native conditions the signaling state is an on-pathway intermediate for the photocycle, without the involvement of the fully unfolded state. Therefore, the signaling state of PYP also is on on-pathway folding intermediate for the refolding from fully unfolded PYP with a cis chromophore (see Fig. 3).
Figure 3.

Integrating folding and signaling in PYP. The folding and photocycle transitions in PYP are depicted schematically. Note that pUcis is an off-pathway intermediate for the photocycle, whereas pBcis is an on-pathway intermediate for pUcis refolding.
The denaturation curves of native PYP and its signaling state were analyzed to determine the unfolding free energy of these two states in the absence of denaturants (Table 2). The GdmHCl concentration at which unfolding free energy is zero is reduced from 2.72 M for native PYP to 1.75 M for the signaling state of PYP. For native PYP the unfolding free energy in water is 37 ± 1 kJ/mol. This value is reduced by ≈20% for the signaling state, yielding an energy content for the signaling state of 7 ± 2 kJ/mol in water at pH 7.0 relative to native PYP. Note that this calculation involves the simplifying assumption that the ΔG between the unfolded states containing trans and cis chromophores is close to zero.
Table 2.
Denaturant dependence of equilibria in PYP
| Process | ΔG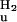  (kJ/mol) (kJ/mol) |
m (M−1) |
|---|---|---|
| GdmHCl | ||
| pGtrans↔pUtrans | 36.4 ± 1.1*/39.9 ± 7.0† | 5.4 ± 0.2*/5.7 ± 0.1† |
| pBcis↔pUcis | 28.2 ± 2.0* | 6.4 ± 0.5* |
| I↔pUcis† | 18.3 ± 18.6‡ | 4.2 ± 0.7‡ |
| Urea | ||
| pGtrans↔pUtrans | 37.6 ± 1.0* | 1.98 ± 0.06* |
| pBcis↔pUcis | 32.3 ± 1.5* | 2.48 ± 0.12* |
The reported values were obtained from equilibrium titrations
and from chevron analysis of refolding of PYP using the relationship ΔGu = −RT lnK = −RT ln(kU/kF) for pUtrans
and Eq. 3 for pUcis
.
The crystal structure of the signaling state as it accumulates in PYP crystals has been reported (27). This structure does not reveal any unfolding on the formation of the signaling state, in contrast to the results reported here. We have recently shown that the protein crystal lattice greatly reduces the structural changes that occur on PYP photoexcitation (28). Apparently, the unfolding event during the PYP photocycle occurs when PYP is in solution, but not in PYP crystals.
Specific Unfolding of a Functional Intermediate.
A central assumption in the analysis described above is that the denaturant-induced transition observed by steady-state illumination of PYP corresponds to the preequilibrium between the fully unfolded state and the signaling state. Time-resolved CD spectroscopy (Fig. 2 B and C) was performed during the last photocycle step to confirm that the blue-shift in the absorbance spectrum of the signaling state from 355 to 338 nm is caused by the complete unfolding of this state. The presence of 6.0 M urea does not affect the CD spectrum of native PYP (see Fig. 2B). However, on photoexcitation of PYP under these conditions the peptide CD spectrum greatly decreases in amplitude and becomes featureless, confirming that the signaling state is converted to a fully denatured state (Fig. 2B). The denaturant dependence of the light-induced loss of CD signal matches the UV/vis absorbance data (Fig. 2D). This demonstrates the specific and cooperative denaturation of a functional intermediate.
PYP as a Model System to Study Protein Folding.
The results reported here show that the signaling state of PYP can be transiently accumulated via two independent routes: (i) by rapid mixing refolding of the fully unfolded state of PYP containing a cis chromophore (Fig. 1B), and (ii) by photoexitation of PYP at moderate denaturant concentrations (Fig. 2A). The unfolding of the blue-shifted photocycle intermediate, formed by photoexcitation of PYP, and the refolding of PYP from the fully unfolded state containing a cis chromophore, observed by rapid mixing, follow the same denaturant dependence. This provides strong evidence for the role of the signaling state as an on-pathway folding intermediate (Fig. 3). Both the occurrence of folding intermediates and the position of such intermediates as on-pathway or off-pathway states have been found to be difficult to determine conclusively in many proteins (1–4). By studying protein folding in PYP not only by rapid mixing techniques but also by photoexcitation, we provide an unusually strong case for the occurrence of an on-pathway folding intermediate in PYP folding that causes a rapid preequilibrium in kinetic studies.
We demonstrate a close correspondence between the kinetics for PYP refolding in a chevron plot, the PYP photocycle kinetics, and the data on the steady-state denaturation of the blue-shifted photocycle intermediate on PYP photoexcitation in the presence of moderate denaturant concentrations. This provides strong evidence that the signaling state is shared by the energy surfaces for folding and function in PYP.
Until recently, the kinetics of protein folding were mainly studied by using commercially available rapid mixing devices, resulting in a time resolution limited to a few milliseconds (but see ref. 29). The possibility of reversible, light-triggered, and complete protein unfolding in PYP will allow future flash-photolysis experiments to study these folding reactions with picosecond time resolution (30). In combination with the possibility of studying the same protein folding transitions by two independent pathways, this establishes PYP as a valuable model system to study protein folding. The same approach can be applied to other (photo)receptor systems.
Transient Partial Protein Unfolding as a Signaling Mechanism.
This study demonstrates a direct link between receptor activation and protein folding in PYP. We show how this result provides an avenue for the study of protein folding that can yield solid information on the pathway of folding. In addition, this result implies that the light-triggered transient partial protein unfolding event that occurs during signaling state formation under native conditions is required for the generation of a biological signal during the PYP photocycle. The transient unfolding process can provide the receptor with the conformational plasticity needed for signaling state formation. Such transient unfolding has not been observed in well-studied transmembrane receptors such as rhodopsin and could be limited to water-soluble receptor proteins. Because PYP serves as a prototype for the PAS domain family (11, 12), it is expected that transient partial protein unfolding also is involved in other proteins containing this ubiquitous signaling module. The same mechanism also may function in unrelated signaling systems (31), for example in the Wiskott–Aldrich syndrome protein (32). This emphasizes the importance of future studies on the role of transient partial protein unfolding in signal transduction in the living cell.
Acknowledgments
We thank Applied Photophysics for excellent technical support, Kevin Meisner for help in the initial stage of the project, Tobin Sosnick for invaluable help with the time-resolved CD measurements and illuminating discussions, and Philippe Cluzel for valuable comments on the manuscript. This work is supported by grants to W.D.H. from the American Cancer Society and the Cancer Research Foundation.
Abbreviations
- GmdHCl
guanidinium-hydrochloride
- pBcis
blue-shifted photocycle intermediate with cis chromophore
- pGtrans
initial state of PYP with trans chromophore
- pUcis
fully unfolded state of PYP with cis chromophore
- pUtrans
fully unfolded state of PYP with trans chromophore
- PYP
photoactive yellow protein
References
- 1.Takei J, Chu R-A, Bai Y. Proc Natl Acad Sci USA. 2000;97:10796–10801. doi: 10.1073/pnas.190265797. . (First Published September 12, 2000; 10.1073/pnas.190265797) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dalby P A, Oliveberg M, Fersht A R. J Mol Biol. 1998;276:625–646. doi: 10.1006/jmbi.1997.1546. [DOI] [PubMed] [Google Scholar]
- 3.Krantz B A, Sosnick T R. Biochemistry. 2000;39:11696–11701. doi: 10.1021/bi000792+. [DOI] [PubMed] [Google Scholar]
- 4.Fersht A R. Proc Natl Acad Sci USA. 2001;97:14121–14126. doi: 10.1073/pnas.260502597. . (First Published December 12, 2000; 10.1073/pnas.260502597) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Meyer T E, Yakali E, Cusanovich M A, Tollin G. Biochemistry. 1987;26:418–423. doi: 10.1021/bi00376a012. [DOI] [PubMed] [Google Scholar]
- 6.Sprenger W W, Hoff W D, Armitage J P, Hellingwerf K J. J Bacteriol. 1993;175:3096–3104. doi: 10.1128/jb.175.10.3096-3104.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jiang Z, Swem L R, Rushing B G, Devanathan S, Tollin G, Bauer C E. Science. 1999;285:406–409. doi: 10.1126/science.285.5426.406. [DOI] [PubMed] [Google Scholar]
- 8.Hoff W D, Van Stokkum I H M, Van Ramesdonk H J, Van Brederode M E, Brouwer A M, Fitch J C, Meyer T E, Van Grondelle R, Hellingwerf K J. Biophys J. 1994;67:1691–1705. doi: 10.1016/S0006-3495(94)80643-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hoff W D, Düx P, Hård K, Devreese B, Nugteren-Roodzant I M, Crielaard W, Boelens R, Van Beeumen J, Hellingwerf K J. Biochemistry. 1994;33:13959–13962. doi: 10.1021/bi00251a001. [DOI] [PubMed] [Google Scholar]
- 10.Baca M, Borgstahl G E O, Boissinot M, Burke P M, Williams D R, Slater K A, Getzoff E D. Biochemistry. 1994;33:14369–14377. doi: 10.1021/bi00252a001. [DOI] [PubMed] [Google Scholar]
- 11.Pellequer J L, Wager-Smith K A, Kay S A, Getzoff E D. Proc Natl Acad Sci USA. 1998;95:5884–5890. doi: 10.1073/pnas.95.11.5884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Taylor B L, Zhulin I B. Microbiol Mol Biol Rev. 1999;63:479–506. doi: 10.1128/mmbr.63.2.479-506.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kort R, Vonk H, Xu X, Hoff W D, Crielaard W, Hellingwerf K J. FEBS Lett. 1996;382:73–78. doi: 10.1016/0014-5793(96)00149-4. [DOI] [PubMed] [Google Scholar]
- 14.Xie A, Hoff W D, Kroon A R, Hellingwerf K J. Biochemistry. 1996;35:14671–14678. doi: 10.1021/bi9623035. [DOI] [PubMed] [Google Scholar]
- 15.Genick U K, Soltis S M, Kuhn P, Canestrelli I L, Getzoff E D. Nature (London) 1998;392:206–209. doi: 10.1038/32462. [DOI] [PubMed] [Google Scholar]
- 16.Perman B, Šrajer V, Ren Z, Teng T, Pradervand C, Ursby T, Schotte F, Wulff M, Kort R, Hellingwerf K J, Moffat K. Science. 1998;279:1946–1950. doi: 10.1126/science.279.5358.1946. [DOI] [PubMed] [Google Scholar]
- 17.Hoff W D, Jung K, Spudich J L. Annu Rev Biophys Biomol Struct. 1997;26:223–258. doi: 10.1146/annurev.biophys.26.1.223. [DOI] [PubMed] [Google Scholar]
- 18.Santoro M M, Bolen D W. Biochemistry. 1992;31:4901–4907. doi: 10.1021/bi00135a022. [DOI] [PubMed] [Google Scholar]
- 19.Matouschek A, Kellis J T, Serrano L, Bycroft M, Fersht A R. Nature (London) 1990;346:440–450. doi: 10.1038/346440a0. [DOI] [PubMed] [Google Scholar]
- 20.Parker M J, Spencer J, Clarke A R. J Mol Biol. 1995;253:771–786. doi: 10.1006/jmbi.1995.0590. [DOI] [PubMed] [Google Scholar]
- 21.Capaldi A P, Ferguson S J, Radford S E. J Mol Biol. 1999;286:1621–1632. doi: 10.1006/jmbi.1998.2588. [DOI] [PubMed] [Google Scholar]
- 22.Matthews C R. Methods Enzymol. 1987;154:498–511. doi: 10.1016/0076-6879(87)54092-7. [DOI] [PubMed] [Google Scholar]
- 23.Bai Y. Proc Natl Acad Sci USA. 1999;96:477–480. doi: 10.1073/pnas.96.2.477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Van Brederode M E, Hoff W D, Van Stokkum I H M, Groot M L, Hellingwerf K J. Biophys J. 1996;71:365–380. doi: 10.1016/S0006-3495(96)79234-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rubinstenn G, Vuister G W, Mulder F A A, Düx P E, Boelens R, Hellingwerf K J, Kaptein R. Nat Struct Biol. 1998;5:568–570. doi: 10.1038/823. [DOI] [PubMed] [Google Scholar]
- 26.Hoff W D, Xie A, Van Stokkum I H M, Tang X-J, Gural J, Kroon A R, Hellingwerf K J. Biochemistry. 1999;38:1009–1017. doi: 10.1021/bi980504y. [DOI] [PubMed] [Google Scholar]
- 27.Genick U K, Borgstahl G E, Ng K, Ren Z, Pradervand C, Burke P M, Šrajer V, Teng T Y, Schildkamp W, McRee D E, et al. Science. 1997;275:1471–1475. doi: 10.1126/science.275.5305.1471. [DOI] [PubMed] [Google Scholar]
- 28.Xie A, Kelemen L, Hendriks J, White B J, Hellingwerf K J, Hoff W D. Biochemistry. 2001;40:1510–1517. doi: 10.1021/bi002449a. [DOI] [PubMed] [Google Scholar]
- 29.Eaton W A, Muñoz V, Hagen S J, Jas G S, Lapidus L J, Henry E R, Hofrichter J. Annu Rev Biophys Biomol Struct. 2000;29:327–359. doi: 10.1146/annurev.biophys.29.1.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jones C M, Henry E R, Hu Y, Chan C K, Luck S D, Bhuyan A, Roder H, Hofrichter J, Eaton W A. Proc Natl Acad Sci USA. 1993;90:11860–1864. doi: 10.1073/pnas.90.24.11860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wright P E, Dyson H J. J Mol Biol. 1999;293:321–331. doi: 10.1006/jmbi.1999.3110. [DOI] [PubMed] [Google Scholar]
- 32.Kim A S, Kakalis L T, Abdul Manan N, Liu G A, Rosen M K. Nature (London) 2000;404:151–158. doi: 10.1038/35004513. [DOI] [PubMed] [Google Scholar]