

Abstract
Pien Tze Huang Gan Bao (PZH-GB), a traditional Chinese medicine, has been used for thousands of years as a protective remedy effective against liver injury induced by excessive alcohol and smoking. The present study aimed to evaluate the protective effects and potential mechanisms of PZH-GB against carbon tetrachloride (CCl4)-induced hepatic injury. Rats were pre-treated with silymarin (50 mg/kg) or different doses of PZH-GB (150, 300 or 600 mg/kg) orally administered for 7 days. At the end of treatment, the rats were intraperitoneally injected with CCl4, or control rats received a corn oil injection. The lactate dehydrogenase (LDH) levels in serum were evaluated. Apoptosis was assessed via terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. p53, B-cell lymphoma 2 (Bcl-2), B cell-lymphoma 2-associated X protein (Bax), cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS) and cytochrome P450 family 2 subfamily E member 1 (CYP2E1) were measured by reverse transcription-quantitative polymerase chain reaction and western blotting. The activity of caspase-9 and caspase-3 were measured by a colorimetric assay. The results indicated that silymarin and PZH-GB prevented CCl4-induced serum LDH elevations, and CCl4 induced high levels of LDH. Compared with the CCl4 group, silymarin and PZH-GB treatment significantly decreased LDH levels. Histopathological results revealed that silymarin and PZH-GB ameliorated the CCl4-induced liver histological alterations. The TUNEL results showed that compared with the control group, CCl4 induced liver cell apoptosis, while silymarin and PZH-GB treatment inhibited apoptosis and the TUNEL-positive cells. The elevated expression of Bax, p53, iNOS, COX-2 and CYP2E1 were reduced by silymarin or PZH-GB pretreatment, whereas reduced Bcl-2 expression levels were increased. CCl4 increased the activity of caspase-9 and −3 by 6.86- and 7.42-fold, respectively; however, silymarin and PZH-GB ameliorated this effect. In conclusion, silymarin and PZH-GB treatment prevented the deleterious effects on liver functions by attenuation of oxidative stress, inflammation and mitochondrial apoptosis via the p53 signaling pathway.
Keywords: hepatic injury, mitochondria
Introduction
The liver is a vital organ involved in regulating many important functions, including metabolism, secretion and storage. It serves a key role in regulating many physiological processes (1). Furthermore, it is involved in detoxification and biotransformation of drugs and xenobiotics; therefore, has demonstrated increased susceptibility to toxicity from these agents (2). Liver diseases have become a worldwide problem as a result of extremely poor prognoses and high mortality rates attributed to the lack of effective prevention methods or therapeutic drugs (3). As such, attempts are perpetually being made to discover novel treatments for liver diseases. The drug discovery process has paid great attention to the investigation of the efficacy of plant-based drugs used in traditional medicine, as they are cheaper and have fewer side effects. Natural products extracted from medicinal plants are considered to be an effective and safe alternative to conventional treatments for hepatotoxicity. Therefore, present study aimed to investigate the hepatoprotective activity of Pien Tze Huang Gan Bao (PZH-GB) against carbon tetrachloride (CCl4)-induced hepatotoxicity in rats. PZH-GB, a classical traditional Chinese medicine, has been used as a protective remedy for thousands of years across China and Southeast Asian countries. PZH-GB has demonstrated therapeutic effects against liver injury induced by excessive alcohol and smoking. To date, however, there are no studies available regarding the protective effect of PZH-GB in response to chemical-induced liver injury.
The present study investigated the protective effect of PZH-GB against CCl4-induced hepatotoxicity, and aimed to determine the mechanisms by which it may confer its protection on the rat liver against CCl4-induced chemical damage. The effect of PZH-GB on liver injury was compared to that of silymarin treatment, a substance used clinically in Europe and Asia for the treatment of liver diseases. For evaluation of the hepatoprotective mechanisms of PZH-GB, liver oxidative damage and anti-oxidant defense potential, proinflammatory mediators including inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), and apoptosis-associated factors were determined.
Materials and methods
Reagents
PZH-GB was obtained from and authenticated by the sole manufacturer Zhangzhou Pien Tze Huang Pharmaceutical Co., Ltd. (Chinese FDA approval no. HPK-08411; Zhangzhou, China). A lactate dehydrogenase assay kit (cat no. 2P56-21) was purchased from Abbott Pharmaceutical Co., Ltd. (Lake Bluff, IL, USA). TRIzol reagents were purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA). A PrimeScript™ RT reagent kit with gDNA Eraser was purchased from Takara Bio, Inc. (Otsu, Japan). Caspase-3 (cat no. KHZ0022) and −9 (cat no. KHZ0102) colorimetric protease assays were purchased from Invitrogen; Thermo Fisher Scientific, Inc. Rabbit monoclonal antibodies for COX-2 (cat no. 12282S; 1:1,000 dilution) and GAPDH (cat no. 5174S; 1:1,000 dilution) were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA), and rabbit monoclonal antibodies for iNOS (cat no. ab3523), cytochrome P450 family 2 subfamily E member 1 (CYP2E1; cat no. ab151544), p53 (cat no. ab109396), B-cell lymphoma 2 (Bcl-2; cat no. ab59348) and Bcl-2 X-associated protein (Bax; cat no. ab32503) were obtained from Abcam (1:1,000 dilution; Cambridge, MA, USA). Horseradish peroxidase-conjugated secondary antibodies (cat no. ab205718) were purchased from Abcam (1:2,000 dilution). CCl4 was purchased from Shanghai Ling Feng Chemical Reagent Co., Ltd. (Shanghai, China).
Animals
Male Sprague-Dawley rats (age, 8 weeks; weight, 180–200 g; n=60; Slike Co., Ltd., Shanghai, China), were housed five per cage in an environmentally-controlled room at a temperature of 22±1°C in a relative humidity 40–60%. Animals were housed in an air ventilation setting of 12–18 times/h and a 12-h light/dark cycle of 150–300 lux conditions, with feed and water provided ad libitum for one week before the start of the experiments. The animal studies were approved by the Fujian Institute of Traditional Chinese Medicine Animal Ethics Committee (Fuzhou, China). The experimental procedures were carried out in accordance with the Guidelines for Animal Experimentation of Fujian University of Traditional Chinese Medicine (Fuzhou, China).
CCl4-induced hepatotoxicity and experimental design
The rats were divided into six groups according to the following conditions: Group 1: control group (NO); group 2: CCl4 model group (vehicle); group 3 (SM+CCl4): silymarin treated CCl4 group (pre-treated with silymarin, 50 mg/kg), a positive control; group 4 (LoGB+CCl4): low dose PZH-GB treatment CCl4 group (pre-treated with PZH-GB, 150 mg/kg); group 5 (MeGB+CCl4): medium dose PZH-GB treatment CCl4 group (pre-treated with PZH-GB, 300 mg/kg); and group 6 (HiGB+CCl4): high dose PZH-GB treatment CCl4 group (pre-treated with PZH-GB, 600 mg/kg). Each group contained 10 rats. Rats from groups 3 to 6 were administered an oral dose of either silymarin or PZH-GB while the rats from control and CCl4 model groups were administered an oral dose of PBS for 7 days before initial CCl4 exposure. On the last day of pre-treatment, rats in groups 2 to 6 were intraperitoneally (i.p.) injected with CCl4 at a dose of 2.0 ml/kg, provided as a 50% corn oil solution, while group 1 received 2.0 ml/kg of corn oil. A total of 24 h after CCl4 administration, the animals were anesthetized (40 mg/kg pentobarbital i.p.; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), sacrificed by cervical dislocation, and subjected to laparotomy, which involved opening the abdominal cavity and exposing the abdominal aorta. Blood (15 ml) was collected by aorta abdominalis into non-heparinized tubes and centrifuged at 980 × g at 4°C for 10 min to obtain serum for biochemical tests. The livers were quickly excised and washed, and part of each liver was cut and fixed in formaldehyde saline (4%) solution for histological analysis; the remaining liver was snap frozen in liquid nitrogen and stored at −70°C until required for molecular analysis.
Measurement of LDH
The quantification of LDH5 was achieved by using the pyruvate to lactate kinetic method, a two-point colometric method for determination of LDH.
Histopathology
Liver samples were fixed in 10% buffered formalin for at least 48 h, processed and embedded in paraffin. Next, the paraffin sections were prepared using an automatic tissue processor and cut into 5-µm thick sections by a rotary microtome. The sections were stained with hematoxylin and eosin (H&E) and mounted on slides for examination of histopathological changes. The results were recorded as photomicrographs using a Leica DMRB/E light microscope (Leica Microsystems GmbH, Wetzlar, Germany). A semi-quantitative scoring system was used to assess the severity of the hepatic steatosis and inflammatory cell infiltration in 10 microscopic fields. In brief, the following criteria were used for scoring hepatic steatosis: grade 0 (−), no inflammation or ballooning degeneration; grade 1 (+), inflammation or ballooning degeneration occupying 33% of the hepatic parenchyma; grade 2 (++), inflammation or ballooning degeneration occupying 33–66% of the hepatic parenchyma; grade 3 (+++), inflammation or ballooning degeneration occupying >66% of the hepatic parenchyma.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from the remaining liver samples were isolated using TRIzol reagent. The isolation was performed according to the manufacturer's protocol, and RNA was subsequently quantified by measuring the absorbance of the preparation at a wavelength of 260 nm. cDNA synthesis was performed using the PrimeScript™ RT reagent kit according to the manufacturer's protocol. Total RNA (1.0 µg from each sample) was added to a mixture of 4.0 µl 5X PrimeScript buffer, 1.0 µl PrimeScript RT enzyme mix I, 1.0 µl Oligo dT primer (50 µM), 1.0 µl random 6 mers (100 µM), and RNase-free water was added to a final volume of 20 µl. The final reaction mixture was kept at 25°C for 10 min, heated to 37°C for 15 min, heated to 85°C for 5 sec, and then cooled to 4°C.
cDNA from the above preparation was subjected to PCR amplification using SYBR Premix Ex Taq I in an ABI 7500 Fast instrument. mRNA expression values were determined using the 2−∆∆Cq method (4). GAPDH served as an internal reference control. All qPCR reactions were conducted in triplicate. Primer sequences were as follows: Forward, GCATCCTGTCCCCATCACCA and reverse, CCCAGCAACTACCAACCCATTC for p52; forward, GAACTGTATCCCGCCCTGCTGGT and reverse, CTTGCGTTGATGGTGGCTGTCTT for COX-2; forward, CCTCCTTGTTCAACTCACCTTCG and reverse, ACCTCTGCCTGTGCGTCTCTTC for iNOS; forward, CCTTTTGACCCCACATTTCTGAT and reverse, ATGGCTTCCAGGTAGGTATCGTAG for CYP2E1; forward, GTGTATTTCACGGGACCTGGCT and reverse, GATGCTCTTGAAGGTCTCGTAGGT for nuclear factor (NF)-κB; forward, GCTGATGGCAACTTCAACTGGG and reverse, TTCTTCCAGATGGTGAGCGAGG for Bax; forward, TACCGTCGTGACTTCGCAGAGAT and reverse, AGGAGAAATCAAACAGAGGTCGC for Bcl-2; forward, CGGTCAGGTCATCACTATCGGC and reverse, GTGTTGGCATAGAGGTCTTTACGG for GAPDH.
Western blot analysis
Livers (n=4/group) were homogenized and total protein lysate was prepared in Radioimmunoprecipitation Assay Lysis and Extraction Buffer (Thermo Fisher Scientific, Inc.) containing protease and phosphatase inhibitor cocktails, which was followed by centrifugation at 12,000 × g for 15 min at 4°C. Protein concentrations were determined with Bicinchoninic Acid protein assay kit (Pierce; Thermo Fisher Scientific, Inc.). Protein samples (50 µg) were separated by 10% SDS-PAGE and transferred onto polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA, USA). Membranes were blocked with TBS overnight at 4°C followed by incubation with primary rabbit monoclonal antibodies against COX-2, iNOS, CYP2E1, p53, Bax, Bcl-2 and GAPDH for 2 h at room temperature. After three washes in TBS with 0.1% Tween-20 (TBST), membranes were each probed with an anti-rabbit horseradish peroxidase-conjugated secondary antibody (1:2,000) for 1 h at room temperature, and the membranes were washed again in TBST followed by enhanced chemiluminescence detection using SuperSignal™ West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Inc.).
Analysis of caspase-3 and −9 activation
The activities of caspase-3 and −9 were determined by caspase-3 and −9 colorimetric activation kits following the manufacturer's protocol. Briefly, six liver tissues from each group were homogenized and lysed with lysis buffer for 30 min on ice. The lysed tissues were centrifuged at 16,000 × g for 10 min at 4°C. The protein concentration of the clarified supernatant was determined and 100 µg protein was incubated with 50 µl colorimetric tetrapeptides, Asp-Glue-Val-Asp (DEAD)-p-nitroaniline (pNA, specific substrate of caspase-3) or Leu-Glue-His-Asp (LEHD)-pNA (specific substrate of caspase-9) at 37°C in the dark for 2 h. Samples were read at 405 nm on an ELISA plate reader (model ELX800; BioTek, Winooski, VT, USA). The data were normalized to caspase activity in control group liver tissue and are presented as a fold of control.
High performance liquid chromatography (HPLC)
Samples were analyzed on an Agilent 1200 HPLC system (Agilent Technologies, Santa Clara, CA, USA) using a Welch Ultimate XB-C18 (250×4.60 mm; 5 µm). The absorbance was measured at 274 nm. The mobile phase consisted of methanol and 0.1% phosphoric acid (45:55) at a flow rate of 0.9 ml/min with an injection volume of 5 µl. The column temperature was 20°C. Baicalin (Sigma-Aldrich; Merck KGaA) served as a positive control.
Statistical analysis
All data are expressed as the means ± standard deviation of three independent experiments. Data were analyzed using SPSS version 11.5 (SPSS, Inc., Chicago, IL, USA). Statistical analysis was performed by one-way analysis of variance followed by a post hoc Fisher's least significant difference test, and qualitative data was analyzed by Chi-squared test. P<0.05 was considered to indicate a statistically significant difference.
Results
Effect of PZH-GB on serum LDH5
The effects of PZH-GB on CCl4-induced hepatotoxicity in rats was evaluated by changes in serum LDH5. The activities of LDH5 in control and all treated groups are presented in Fig. 1. The data demonstrated increased levels of serum LDH5 in the CCl4-treated animals compared with the control group. This effect was reversed in the animal groups that were given PZH-GB or silymarin pretreatment. Treatment with 150, 300 and 600 mg/kg PZH-GB, and silymarin at a dose of 50 mg/kg, exhibited a reduction of 19.29, 20.46, 27.97 and 56.80% on LDH5 levels, respectively.
Figure 1.
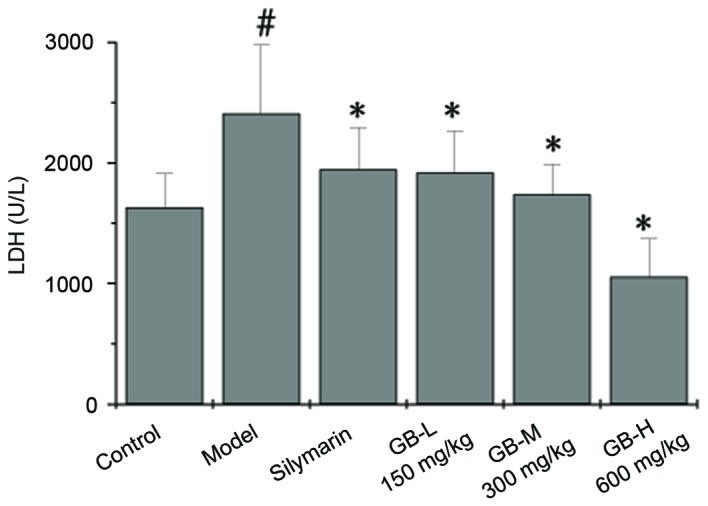
Effects of pre-treatment of PZH-GB and silymarin on serum LDH in CCl4-intoxicated rats. Animals were pretreated with PZH-GB (150, 300 and 600 mg/kg), silymarin (50 mg/kg) or vehicle. Data are expressed as the mean ± standard deviation (n=10/group). #P<0.05 vs. control group; *P<0.05 vs. CCl4-exposed group. PZH-GB, Pien Tze Huang Gan Bao; LDH, lactate dehydrogenase; CCl4, carbon tetrachloride; GB-L, GB-M, GB-H, PZH-GB low, medium and high-dose treatment groups, respectively.
Effect of GB on histopathological changes
Histological analysis with H&E revealed advanced centrilobular necrosis, broad infiltration of inflammatory cells, ballooning degeneration and the loss of cellular boundaries in liver sections of CCl4 model rats. Both PZH-GB and silymarin groups exhibited an apparent reversal of these histopathological effects, especially PZH-GB at the 600 mg/kg dose, as presented in Fig. 2 and Table I.
Figure 2.

Effects of PZH-GB on histopathological changes induced by CCl4 in rats. The rats were pretreated with PZH-GB (150, 300 and 600 mg/kg), silymarin (50 mg/kg) or vehicle for 7 days before administration of CCl4 (2 ml/kg body weight and 1:1 diluted in corn oil) to induce acute liver injury. Representative pictures of liver sections stained with hematoxylin and eosin (magnification, ×200) are presented. (−), no inflammation or ballooning degeneration; (+), inflammation or ballooning degeneration occupying 33% of the hepatic parenchyma; (+++), inflammation or ballooning degeneration occupying >66% of the hepatic parenchyma. PZH-GB, Pien Tze Huang Gan Bao; CCl4, carbon tetrachloride; GB, PZH-GB.
Table I.
Quantification analysis for histopathologic changes.
| Liver injury | |||||
|---|---|---|---|---|---|
| Group | Rats (n) | − | + | ++ | +++ |
| Control | 10 | 10 | 0 | 0 | 0 |
| Model | 10 | 0 | 0 | 2 | 8a |
| Silymarin | 10 | 1 | 3 | 4 | 2b |
| GB-150 mg/kg | 10 | 0 | 4 | 3 | 3b |
| GB-300 mg/kg | 10 | 0 | 4 | 4 | 2b |
| GB-600 mg/kg | 10 | 2 | 4 | 2 | 2b |
To evaluate the degree of necrosis after acute liver injury, an injury grading score based on severity of necrotic lesions in the liver parenchyma was created. Data were analyzed by Chi-squared test.
P<0.05 vs. control group
P<0.05 vs. CCl4-treated group. (−), grade 0, no inflammation or ballooning degeneration; (+), grade 1, inflammation or ballooning degeneration occupying 33% of the hepatic parenchyma; (++), grade 2, inflammation or ballooning degeneration occupying 33-66% of the hepatic parenchyma; (+++), grade 3, inflammation or ballooning degeneration occupying >66% of the hepatic parenchyma.
Effect of PZH-GB on apoptosis
Cell apoptosis in liver tissues was determined via immunohistochemical staining for terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL). As presented in Fig. 3, the percentage of TUNEL-positive cells in the CCl4 model group was 86.33±15.62%, which was increased significantly compared with the control group (11.83±2.32%). Alternatively, the percentage of TUNEL-positive cells in the silymarin and low-, medium- and high-PZH-GB-pretreated groups was 66.67±18.19, 37.17±11.50, 41.00±3.03 and 36.33±14.46%, respectively. This reflected a significant decrease compared with the CCl4 untreated model group.
Figure 3.

Effect of Pien Tze Huang Gan Bao on apoptosis in carbon tetrachloride-exposed rats. (A) Representative immunohistochemical images and (B) quantification of TUNEL staining in liver tissues. Magnification, ×400. Data are presented as the mean ± standard deviation (n=10/group). #P<0.05 vs. control group; *P<0.05 vs. model group. GB, Pien Tze Huang Gan Bao; L, low-dose; M, medium-dose; H, high-dose.
To identify the downstream effectors involved in the CCl4 apoptotic signaling pathway, the activation of caspase-9 and caspase-3 was examined by a colorimetric assay using specific chromophores. Activation of caspase-9 (Fig. 4A) and caspase-3 (Fig. 4B) in the liver tissue was significantly induced by CCl4. Both PZH-GB and silymarin treatment inhibited the activation of caspase-9 and caspase-3 (P<0.05 vs. model group).
Figure 4.

Effect of Pien Tze Huang Gan Bao on the activity of caspases in carbon tetrachloride-induced rats. (A) Caspase-9 and (B) caspase-3 activities were determined by a colorimetric assay. Data are presented as the mean ± standard deviation of three independent experiments. #P<0.05 vs. control group; *P<0.05 vs. model group. GB, Pien Tze Huang Gan Bao.
To further investigate the mechanism by which PZH-GB confers its anti-apoptotic activity, RT-qPCR and western blot analyses were performed to determine the mRNA and protein expression levels of p53, Bcl-2 and Bax in liver tissue. As presented in Fig. 5A, the mRNA expression levels of p53 and Bax were upregulated in CCl4-induced liver, whereas both PZH-GB and silymarin treatment reversed the expression of p53, Bcl-2 and Bax, albeit in slightly different patterns. The western blotting results were consistent with this (Fig. 5B). Collectively, these results suggested that CCl4 induced apoptosis of liver cells, whereas PZH-GB and silymarin inhibited apoptosis by decreasing the expression of the Bax/Bcl-2 ratio and p53.
Figure 5.
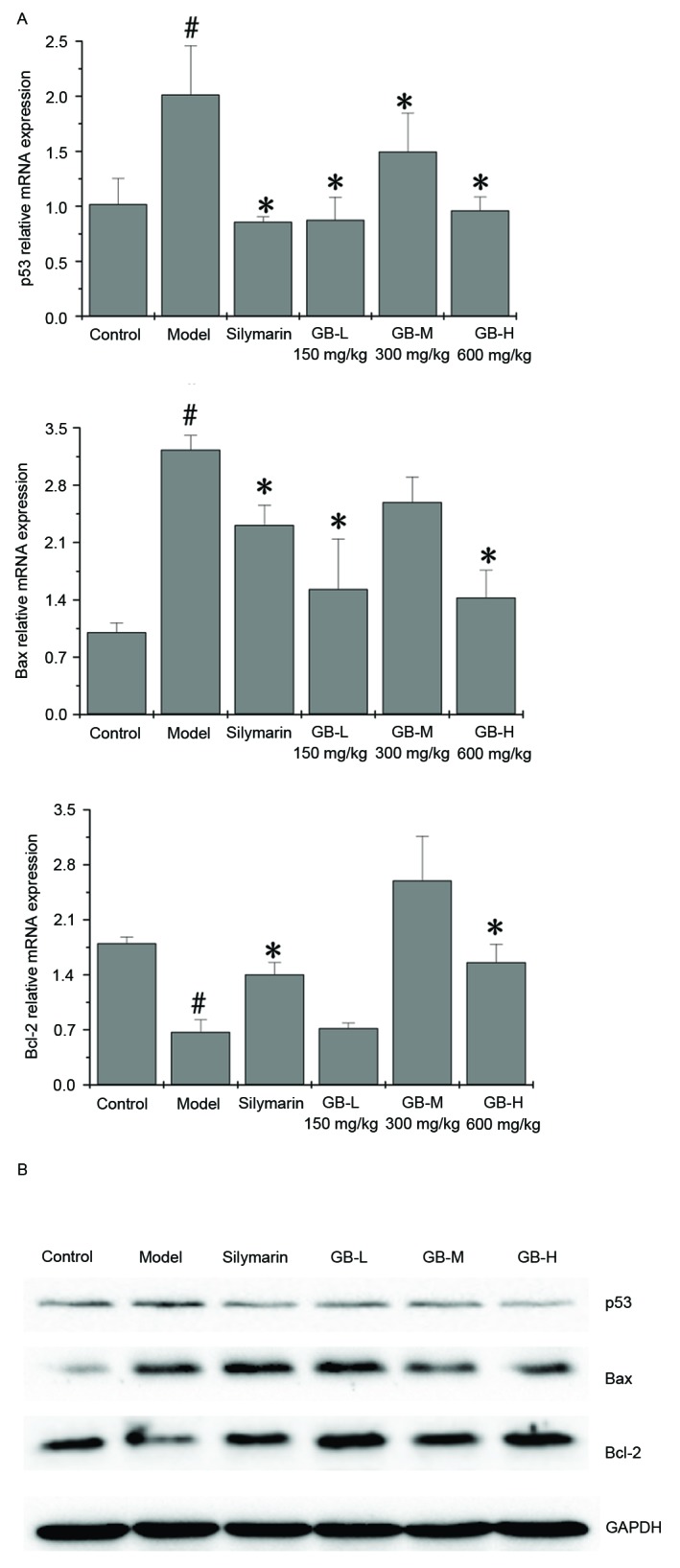
Effect of Pien Tze Huang Gan Bao on the expression of p53, Bax and Bcl-2 in carbon tetrachloride-exposed rats. (A) mRNA and (B) protein expression levels of p53, Bcl-2 and Bax, as determined by reverse transcription-quantitative polymerase chain reaction and western blotting, respectively. GAPDH served as the internal control. Data are presented as the mean ± standard deviation (n=3/group). #P<0.05 vs. control group; *P<0.05 vs. model group. Bax, B cell lymphoma-2 X-associated protein; Bcl-2, B-cell lymphoma 2; GB, Pien Tze Huang Gan Bao; L, low-dose; M, medium-dose; H, high-dose.
Effect of PZH-GB on oxidative stress and inflammation
The effect of PZH-GB on iNOS, COX-2 and CYP2E1 expression was examined by RT-qPCR and western blotting. CCl4 upregulated the mRNA (Fig. 6A) and protein (Fig. 6B) expression levels of iNOS, COX-2 and CYP2E1; however, pre-treatment with PZH-GB and silymarin ameliorated this effect for CYP2E1 and iNOS. Silymarin had no effect on COX-2 expression levels; however, 600 mg/kg PZH-GB significantly reduced them.
Figure 6.
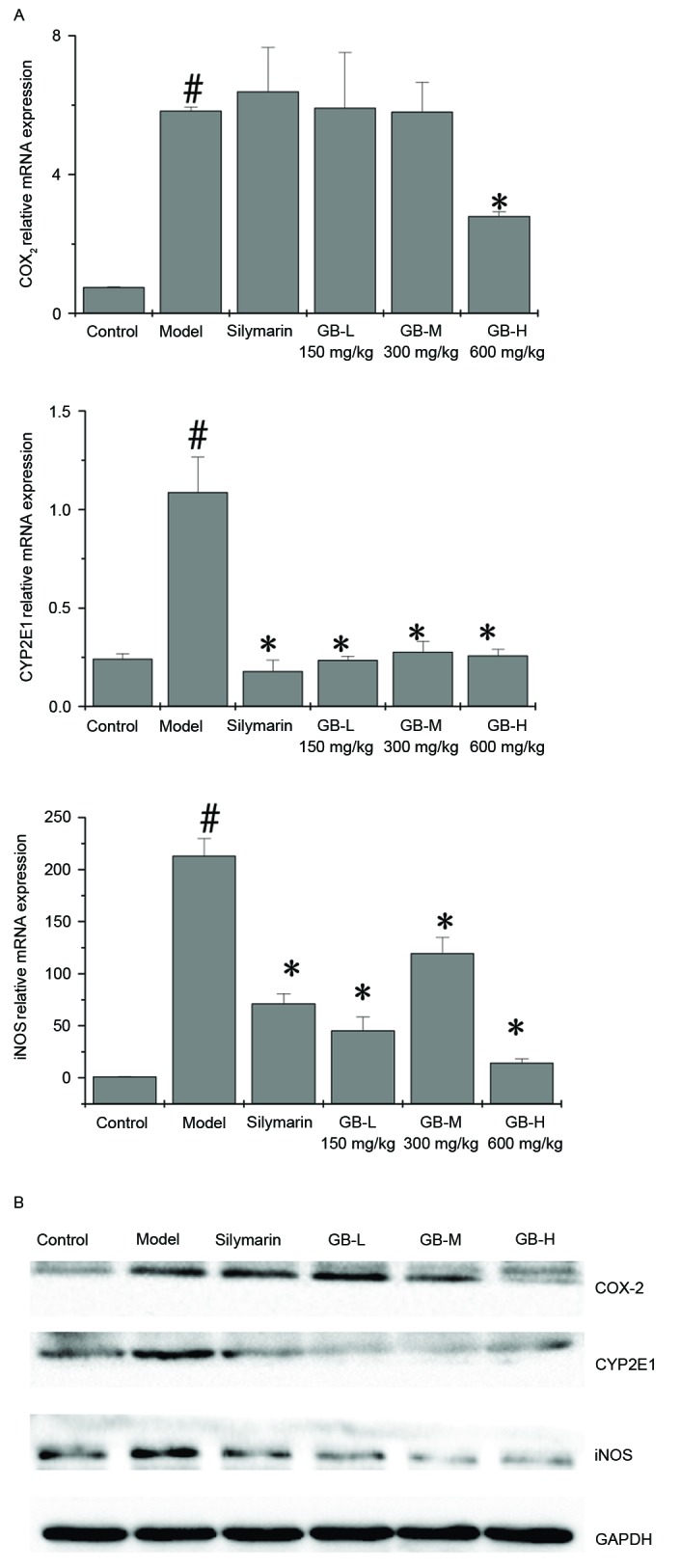
Effect of Pien Tze Huang Gan Bao on the expression of COX-2, CYP2E1 and iNOS in carbon tetrachloride-exposed rats. (A) mRNA and (B) protein expression levels of COX-2, CYP2E1 and iNOS, as determined by reverse transcription-quantitative polymerase chain reaction and western blotting, respectively. GAPDH served as the internal control. Data are presented as the mean ± standard deviation (n=3/group). #P<0.05 vs. control group; *P<0.05 vs. model group. COX-2, cyclooxygenase 2; iNOS, inducible nitric oxide; CYP2E1, cytochrome P450 family 2 subfamily E member 1; GB, Pien Tze Huang Gan Bao; L, low-dose; M, medium-dose; H, high-dose.
HPLC
Based on the fingerprint as presented in Fig. 7, an optimum and easily controlled procedure for preparing PZH-GB was established, as mentioned above. The control sample was composed of baicalin.
Figure 7.

High performance liquid chromatography profiles of Pien Tze Huang Gan Bao and a control sample. The mobile phase consisted of methanol: 0.1% phosphoric acid=45:55. The control sample was composed of baicalin.
Discussion
CCl4, a well-known compound for producing chemical hepatic injury, is metabolized by the hepatic microsomal cytochrome P450 2E1 system (5,6). During metabolism, an unstable trichloromethyl (CCl3) free radical is formed and rapidly converted to trichloromethyl peroxide (Cl3COO−) (3,7). This results in a large amount of reactive oxygen species (ROS) generation, which in turn may increase oxidative stress (8). Previous studies have revealed that the ROS accumulation can overwhelm the intrinsic antioxidant defense system and trigger oxidative stress representative of significant cytotoxic effects in liver and other tissues (9).
Mitochondria are the main organelles involved in intracellular oxidation reactions and are also the main location for ROS production (10). During the CCl4 metabolism process, nicotinamide adenine dinucleotide (NAD) serves as a co-enzyme in the presence of large free radical consumption, resulting in a reduction in the ratio of intracellular NAD/NADH. In turn, this event may promote mitochondrial respiratory chain circulation and generate excess ROS (11). When mitochondrial ROS generation occurs beyond the antioxidant capacity of scavengers, ROS can damage the mitochondria and overflow into the cytoplasm, leading to cellular damage (12). Mitochondrial damage can then trigger the release of caspase-3 into the cytoplasm, which in turn signals the onset of apoptosis. A growing number of studies suggest that apoptosis serves an important role in liver disease etiology (13,14). The Bcl-2 family of proteins (including the anti-apoptotic protein Bcl-2 and the pro-apoptotic protein Bax) serve a key regulatory role in the apoptotic process. The ratio of Bax/Bcl-2 represents the degree of apoptosis to a certain extent (15).
In the current study, evidence that pre-treatment with PZH-GB had a protective effect against CCl4-induced hepatic apoptosis in rats was provided. It was demonstrated that PZH-GB attenuates chemical-induced liver injury, and that histopathological changes caused by CCl4, such as cellular ballooning, are clearly ameliorated by PZH-GB. This finding was consistent with observed changes of liver enzyme levels, such as LDH5 (a LDH isoenzyme which is synthesized by the liver). LDH is a key enzyme marker associated with liver injury. Following liver injury, LDH levels were elevated. In the present study, increased LDH was induced by CCl4. PZH-GB treatment could decrease the LDH level, but the LDH level of GB-H group was lower than that of the control group. This result may indicate that GB-H treatment can excessively reduce the stress response induced by CCl4. The anti-apoptotic effect of PZH-GB was further evidenced by the demonstration that PZH-GB decreased the number of TUNEL-positive hepatic cells, and reduced caspase-3 activation. Steatosis and ballooning of hepatocytes are the earliest, most frequent and most striking pathological changes observed in CCl4-induced liver injury (16,17). The present study confirmed this pathological change by H&E staining, and determined that PZH-GB reversed this change.
Apoptosis of hepatocytes is a critical mechanism of liver injury (18). To date, the TUNEL assay had been used as a marker of apoptosis. In the present study, TUNEL-positive cells in the CCl4 group were significantly increased compared with the control group. More importantly, PZH-GB reduced the number of TUNEL-labeled cells. However, the TUNEL assay is not a specific marker of apoptosis-multiple factors activate apoptotic cascade proteins. Previous evidence supports the role of caspase-3 as an executioner of apoptosis, and it is understood that caspase-3 serves a dominant role after hepatic injury (19). In addition, the Bcl-2 family serves an important role in hepatocyte intrinsic apoptosis, particularly Bcl-2 and Bax. The Bcl-2/Bax ratio is considered to determine cell fate after an apoptotic stimulus (20,21). The present study demonstrated that the caspase-9 and −3 activity in the group treated with PZH-GB was remarkably decreased compared with the untreated CCl4-induced group. Furthermore, the PZH-GB treated group exemplified a restored Bcl-2/Bax ratio. These data collectively suggested that PZH-GB alleviated CCl4-induced hepatic injury by suppressing apoptosis and the associated members of the mitochondrial signaling pathway which confer this apoptotic stimulus.
It has been demonstrated that hepatocyte apoptosis can induce liver injury, and p53 is accumulated in hepatocytes in several liver diseases (22–25). It has been reported that CCl4 upregulates the p53 expression (26). Therefore, transcriptional mechanisms are indeed involved in p53 activation by CCl4. RT-qPCR and western blot analysis further demonstrated that p53 was markedly upregulated in the CCl4 group compared with the control group. This indicated that p53 was activated after CCl4 administration, and that PZH-GB was able to normalize that activation. However, the mRNA expression levels of p53, Bax and Bcl-2 were increased in the GB-M group compared with the GB-L and GB-H groups. These data indicated that the effect of PZH-GB was unstable at the transcription level. These findings suggested that intervention of PZH-GB along the mitochondrial apoptotic pathway was p53-mediated in a rat model of CCl4-induced liver disease.
ROS formed during the biotransformation of CCl4 are more reactive and toxic than the parental compound. Biotransformation of CCl4 occurs in the endoplasmic reticulum and the isoenzyme implicated in this process is CYP2E1 (27,28). Our preliminary studies have demonstrated that the rats exposed to CCl4 generated more oxidative products, including malondialdehyde (MDA) and TBARs thiobarbituric acid reactive substances, and PZH-GB treatment could decrease the level of MDA and TBARs. In this study, the results revealed that the active free radical/intermediate CCl4 caused a reduction in CYP2E1, which was markedly restored by PZH-GB pre-treatment. Taken together, this suggested that ROS was generated upon exposure to CCl4.
The liver is a major inflammatory organ, and inflammatory processes contribute to a number of pathological events following exposure to various hepatotoxins (29). NO is a highly reactive oxidant that is produced via the action of iNOS, and serves important roles as a vasodilator and neurotransmitter, and in the immunological system as a defense against tumor cells, parasites and bacteria (30). However, there is evidence that excessive NO production by iNOS may also lead to hepatic damage (31,32). COX-2 is the enzyme responsible for the catalysis of prostaglandin E2 from arachidonic acid (33), and the induction of COX-2 is closely associated with NO production (34). The current study confirmed that significant increases in hepatic iNOS and COX-2 expression are exhibited after CCl4 administration. These alterations were attenuated by PZH-GB pre-treatment, which suggests that PZH-GB suppressed iNOS and COX-2 protein secretion and/or enhanced the degradation of these proteins. Accordingly, at least one other potential mechanism of PZH-GB protection against CCl4-induced hepatotoxicity appears to be, at least in part, due to the suppression of inflammatory responses.
In conclusion, the results of the present study demonstrated that PZH-GB exerted anti-apoptotic effects via a p53-dependent mitochondrial pathway. It also protected against CCl4-induced hepatocyte apoptosis by regulating the Bcl-2 family of proteins and caspase activity. These anti-apoptotic effects were associated with decreases in the expression of pro-apoptotic proteins in the cytoplasm and the inhibition of proteins associated with apoptosis in the mitochondria. Additionally, PZH-GB exhibited a protective effect against CCl4-induced acute histological liver injury, likely via the suppression of inflammatory responses. Coupled with an anti-apoptotic capacity and its ability as a free radical scavenger, PZH-GB has exhibited an effective therapeutic quality in a rat model of chemical-induced liver disease that may be clinically useful as a safer alternative to present treatment options.
Acknowledgements
The present study was supported by the National Natural Science Foundation of China (grant no. 81303125), and the Developmental Fund of Chen Keji Integrative Medicine (CKJ2013015). The authors would like to thank Clarity Manuscript Consultant LLC (Indianapolis, IN, USA) for language editing.
References
- 1.Kumar CH, Ramesh A, Kumar JN Suresh, Ishaq BM. A review on hepatoprotective activity of medicinal plants. Int J Pharmaceut Sci Res. 2011;23:501–515. [Google Scholar]
- 2.Ahsan R, Islam KM, Bulbul IJ, Musaddik A, Haque E. Hepatoprotective activity of methanol extract of some medicinal plants against carbon tetrachloride-induced hepatotoxicity in rats. Eur J Sci Res. 2009;37:302–310. [Google Scholar]
- 3.Chattopadhyay RR. Possible mechanism of hepatoprotective activity of Azadirachta indica leaf extract: part II. J Ethnopharmacol. 2003;89:217–219. doi: 10.1016/j.jep.2003.08.006. [DOI] [PubMed] [Google Scholar]
- 4.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 5.Recknagel RO, Glende EA, Jr, Dolak JA, Waller RL. Mechanisms of carbon tetrachloride toxicity. Pharmacol Ther. 1989;43:139–154. doi: 10.1016/0163-7258(89)90050-8. [DOI] [PubMed] [Google Scholar]
- 6.Williams AT, Burk RF. Carbon tetrachloride hepatotoxicity: An example of free radical-mediated injury. Semin Liver Dis. 1990;10:279–284. doi: 10.1055/s-2008-1040483. [DOI] [PubMed] [Google Scholar]
- 7.Slater TF. Free radicals as reactive intermediates in tissue injury. Adv Exp Med Biol. 1981;136:575–589. doi: 10.1007/978-1-4757-0674-1_42. [DOI] [PubMed] [Google Scholar]
- 8.Brattin WJ, Glende EA, Jr, Recknagel RO. Pathological mechanisms in carbon tetrachloride hepatotoxicity. J Free Radic Biol Med. 1985;1:27–38. doi: 10.1016/0748-5514(85)90026-1. [DOI] [PubMed] [Google Scholar]
- 9.Cederbaum AI. Role of CYP2E1 in ethanol-induced oxidant stress, fatty liver and hepatotoxicity. Dig Dis. 2010;28:802–811. doi: 10.1159/000324289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bartosz G. Reactive oxygen species: Destroyers or messengers? Biochem Pharmacol. 2009;77:1303–1315. doi: 10.1016/j.bcp.2008.11.009. [DOI] [PubMed] [Google Scholar]
- 11.Cadenas E, Davies KJ. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic Biol Med. 2000;29:222–230. doi: 10.1016/S0891-5849(00)00317-8. [DOI] [PubMed] [Google Scholar]
- 12.Circu ML, Aw TY. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic Biol Med. 2010;48:749–762. doi: 10.1016/j.freeradbiomed.2009.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shi J, Aisaki K, Ikawa Y, Wake K. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am J Pathol. 1998;153:515–525. doi: 10.1016/S0002-9440(10)65594-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hoek JB, Cahill A, Pastorino JG. Alcohol and mitochondria: A dysfunctional relationship. Gastroenterology. 2002;122:2049–2063. doi: 10.1053/gast.2002.33613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Volkmann N, Marassi FM, Newmeyer DD, Hanein D. The rheostat in the membrane: BCL-2 family proteins and apoptosis. Cell Death Differ. 2014;21:206–215. doi: 10.1038/cdd.2013.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Takebe H, Sato I, Tajima S, Ikeda Y, Ito K, Nose T. Effects of cianidanol (KB-53) on liver cirrhosis induced by CCl4 in rats: A pathological investigation. Nihon Yakurigaku Zasshi. 1983;81:585–591. doi: 10.1254/fpj.81.585. (In Japanese) [DOI] [PubMed] [Google Scholar]
- 17.Merino N, González R, González A, Remirez D. Histopathological evaluation on the effect of red propolis on liver damage induced by CCl4 in rats. Arch Med Res. 1996;27:285–289. [PubMed] [Google Scholar]
- 18.Yadav SS, Sindram D, Perry DK, Clavien PA. Ischemic preconditioning protects the mouse liver by inhibition of apoptosis through a caspase-dependent pathway. Hepatology. 1999;30:1223–1231. doi: 10.1002/hep.510300513. [DOI] [PubMed] [Google Scholar]
- 19.Perry DK, Smyth MJ, Stennicke HR, Salvesen GS, Duriez P, Poirier GG, Hannun YA. Zinc is a potent inhibitor of the apoptotic protease, caspase-3. A novel target for zinc in the inhibition of apoptosis. J Biol Chem. 1997;272:18530–18533. doi: 10.1074/jbc.272.30.18530. [DOI] [PubMed] [Google Scholar]
- 20.Yao Y, Huang C, Li ZF, Wang AY, Liu LY, Zhao XG, Luo Y, Ni L, Zhang WG, Song TS. Exogenous phosphatidylethanolamine induces apoptosis of human hepatoma HepG2 cells via the bcl-2/Bax pathway. World J Gastroenterol. 2009;15:1751–1758. doi: 10.3748/wjg.15.1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jin S, Dai CL. Attenuation of reperfusion-induced hepatocyte apoptosis is associated with reversed bcl-2/bax ratio in hemi-hepatic artery-preserved portal occlusion. J Surg Res. 2012;174:298–304. doi: 10.1016/j.jss.2010.12.030. [DOI] [PubMed] [Google Scholar]
- 22.Liu B, Shi Y, Peng W, Zhang Q, Liu J, Chen N, Zhu R. Diosmetin induces apoptosis by upregulating p53 via the TGF-β signal pathway in HepG2 hepatoma cells. Mol Med Rep. 2016;14:159–164. doi: 10.3892/mmr.2016.5258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ortega JF, de Conti A, Tryndyak V, Furtado KS, Heidor R, Horst MA, Fernandes LH, Tavares PE, Pogribna M, Shpyleva S, et al. Suppressing activity of tributyrin on hepatocarcinogenesis is associated with inhibiting the p53-CRM1 interaction and changing the cellular compartmentalization of p53 protein. Oncotarget. 2016;7:24339–24347. doi: 10.18632/oncotarget.8248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Allam A, Gabr S, Ajarem J, Abdel-Maksoud M. Bcl-2 and p53 expression in hepatic tissues of Egyptian patients with Chronic Hepatitis C. J Pak Med Assoc. 2015;65:1186–1192. [PubMed] [Google Scholar]
- 25.Li X, Yu J, Brock MV, Tao Q, Herman JG, Liang P, Guo M. Epigenetic silencing of BCL6B inactivates p53 signaling and causes human hepatocellular carcinoma cell resist to 5-FU. Oncotarget. 2015;6:11547–11560. doi: 10.18632/oncotarget.3413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Guo XL, Liang B, Wang XW, Fan FG, Jin J, Lan R, Yang JH, Wang XC, Jin L, Cao Q. Glycyrrhizic acid attenuates CCl4-induced hepatocyte apoptosis in rats via a p53-mediated pathway. World J Gastroenterol. 2013;19:3781–3791. doi: 10.3748/wjg.v19.i24.3781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rechnagel RO, Glende EA., Jr Carbon tetrachloride hepatotoxicity: An example of lethal cleavage. CRC Crit Rev Toxicol. 1973;2:263–297. doi: 10.3109/10408447309082019. [DOI] [PubMed] [Google Scholar]
- 28.Al-Shabanah OA, Alam K, Nagi MN, Al-Rikabi AC, Al-Bekairi AM. Protective effect of aminoguanidine, a nitric oxide synthase inhibitor, against carbon tetrachloride induced hepatotoxicity in mice. Life Sci. 2000;66:265–270. doi: 10.1016/S0024-3205(99)00589-5. [DOI] [PubMed] [Google Scholar]
- 29.Badger DA, Sauer JM, Hoglen NC, Jolley CS, Sipes IG. The role of inflammatory cells and cytochrome P450 in the potentiation of CCl4-induced liver injury by a single dose of retinol. Toxicol Appl Pharmacol. 1996;141:507–519. doi: 10.1006/taap.1996.0316. [DOI] [PubMed] [Google Scholar]
- 30.Lowenstein CJ, Snyder SH. Nitric oxide, a novel biologic messenger. Cell. 1992;70:705–707. doi: 10.1016/0092-8674(92)90301-R. [DOI] [PubMed] [Google Scholar]
- 31.Inoue T, Kwon AH, Oda M, Kaibori M, Kamiyama Y, Nishizawa M, Ito S, Okumura T. Hypoxia and heat inhibit inducible nitric oxide synthase gene expression by different mechanisms in rat hepatocytes. Hepatology. 2000;32:1037–1044. doi: 10.1053/jhep.2000.18715. [DOI] [PubMed] [Google Scholar]
- 32.Nadler EP, Dickinson EC, Beer-Stolz D, Alber SM, Watkins SC, Pratt DW, Ford HR. Scavenging nitric oxide reduces hepatocellular injury after endotoxin challenge. Am J Physiol Gastrointest Liver Physiol. 2001;281:G173–G181. doi: 10.1152/ajpgi.2001.281.1.G173. [DOI] [PubMed] [Google Scholar]
- 33.Chun KS, Surh YJ. Signal transduction pathways regulating cyclooxygenase-2 expression: Potential molecular targets for chemoprevention. Biochem Pharmacol. 2004;68:1089–1100. doi: 10.1016/j.bcp.2004.05.031. [DOI] [PubMed] [Google Scholar]
- 34.Chang YC, Li PC, Chen BC, Chang MS, Wang JL, Chiu WT, Lin CH. Lipoteichoic acid-induced nitric oxide synthase expression in RAW 264.7 macrophages is mediated by cyclooxygenase-2, prostaglandin E2, protein kinase A, p38 MAPK, and nuclear factor-κB pathways. Cell Signal. 2006;18:1235–1243. doi: 10.1016/j.cellsig.2005.10.005. [DOI] [PubMed] [Google Scholar]