

Abstract
Purpose
Previously, a genome-wide association study (GWAS) identified rs13382811 (near ZFHX1B) and rs6469937 (near SNTB1) to be associated with high myopia. The present study evaluates the association of these two single nucleotide polymorphisms (SNPs) with moderate to high myopia in two Chinese cohorts and two cohorts of European populations.
Methods
Two Chinese university student cohorts, including one with 300 unrelated subjects with high myopia and 308 emmetropic controls from Guangzhou and a second with 96 unrelated individuals with moderate to high myopia and 96 emmetropic controls of Chaoshanese origin in Guangzhou, were enrolled in this study. Two SNPs, rs6469937 and rs13382811, were selected for genotyping based on their reported associations with severe myopia. The SNPs were genotyped via DNA sequencing. In addition, association analysis of both SNPs was performed using genotype data from the database of Genotypes and Phenotypes (dbGaP) involving a total of 2,423 samples in two independent cohorts of European-derived populations, as follows: Kooperative Gesundheitsforschung in der Region Augsburg (KORA) and TwinsUK. The allelic and genotypic distribution among cases and controls were analyzed using the Chi-square test. Logistic regression was used to evaluate the SNP–SNP interaction. Fisher’s exact test was used for two-SNP comparisons.
Results
In the Guangzhou cohort, SNP rs13382811 near ZFHX1B showed significant association with high myopia (pallelic = 0.0001, pgenotypic = 4.07 × 10−5), with the minor T allele showing an increased risk of high myopia (odds ratio [OR] = 1.68, 95% confidence interval [CI] = 1.28–2.20). SNP rs6469937 near SNTB1 showed nominal evidence of association (pallelic = 0.0085, pgenotypic = 0.0166), which did not withstand correction for multiple testing. No significant association was detected in the smaller Chaoshan cohort alone. The association of SNPs rs13382811 and rs6469937 remained significant when both Han Chinese cohorts were combined (pallelic = 0.0033 and 0.0016, respectively), and it was also significant under the genotypic test (pgenotypic = 0.0036 and 0.0053, respectively). When both SNPs were considered together under a recessive model, their significance increased (p = 8.37 × 10−4), as did their effect (OR = 4.09, 95%CI = 1.7–9.8). The association between either of these two SNPs alone and myopia did not replicate significantly in the combined cohorts of European descent, providing only suggestive results (pallelic = 0.0088 for rs13382811 and pallelic = 0.0319 for rs6469937). However, the effects of the combined SNPs showed significant association (p = 8.2 × 10−4; OR = 1.56, 95%CI = 1.2–2.0). While the risk for myopia increased with risk alleles from both SNPs, the increase was additive rather representing a multiplicative interaction in both populations.
Conclusions
Our study confirms that the two susceptibility loci ZFHX1B and SNTB1 are associated with moderate to high myopia in a Han Chinese population, as well as in a European population, when both SNPs are combined. These results confirm previous reports of their associations, extend these observations to a European population, and suggest that additional interactive and possibly population-specific genetic or environmental factors may affect their contribution to myopia.
Introduction
Myopia is the greatest cause of visual impairment worldwide, and both genetic and environmental factors contribute to its development [1]. Efforts to decipher the hereditary determinants of myopia began in the 1960s and established the importance of genetic factors for myopia, especially for Mendelian forms of high myopia. Family-based linkage studies focusing on Mendelian form of myopia have identified 18 loci contributing to myopia [2-17], while population-based association studies—typically genome-wide association studies (GWAS)—have identified many susceptibility loci, although some are controversial and need further confirmation. Currently, although next-generation sequencing has elucidated several candidate genes associated with high myopia [18-26], causative genes for most identified loci have yet to be determined.
Recently, Khor et al. performed a genome-wide meta-analysis of four GWAS and a replication analysis of three cohorts [27], involving a total of 2,835 subjects with high myopia and 6,986 emmetropic controls of East Asian descent. Two susceptibility loci, rs13382811 near ZFHX1B and rs6469937 near SNTB1, showed evidence of associations with high myopia, with p = 5.79 × 10−10 and 2.01 × 10−9, respectively. However, the association studies of both loci were performed in a mixed population of Asian origin [27], and their associations with high myopia have not yet been replicated in additional studies. Therefore, the present research aims to evaluate the associations of these two SNPs (rs6469937 and rs13382811) with moderate to high myopia in two Chinese university student cohorts from Guangzhou and Chaoshan. Further replication is conducted in two additional cohorts of European ethnicity.
Methods
This study was approved by the Institutional Review Board of Zhongshan Ophthalmic Center (Sun Yat-sen University, Guangzhou, China) and the CNS Review Board (National Institutes of Health, Bethesda, MD). The two Chinese university student cohorts were recruited from the clinic of the Zhongshan Ophthalmic Center, and the laboratory part of the study was performed at the National Eye Institution (NEI). Written informed consent was obtained from participants in according with the tenets of the Declaration of Helsinki.
Subjects
The two Chinese university student cohorts (Guangzhou cohort and Chaoshan cohort) enrolled in this study were studied previously and have been described in detail [28]. Briefly (Table 1), the Guangzhou cohort was composed of 300 unrelated Han Chinese individuals with high myopia and 308 emmetropic controls recruited in Guangzhou of other than Chaoshanese origin. The Chaoshan cohort consisted of 96 unrelated subjects with moderate to high myopia and 96 emmetropic controls in Guangzhou but from Chaoshan, a linguistically and to some degree ethnically distinct population from the eastern part of Guangdong province. The inclusion criteria were described previously [28]; the associated parameters were as follows: 1) moderate to high myopia was defined as spherical equivalent (SE) ≤ −4.0 D; 2) high myopia was defined as SE ≤ −6.0 D; 3) all individuals in the case group had a best unaided visual acuity of 0.8 or better without other known ocular or systemic diseases; and 4) emmetropic controls were defined as bilateral SEM between −0.50 D and +1.0 D with a best unaided visual acuity of 1.0 or better and no other known ocular or systemic diseases. All individuals underwent a complete ophthalmological examination as described previously [28]. Power calculations using the Power for Genetic Association (PGA) analysis program [29] and a high myopia and disease allele prevalence of 20% [30], as well as the marker allele frequencies for controls in each population shown in Table 2, Table 3, and Table 4, showed 75% and 62% power for the Guangzhou sample set alone, 34% and 24% for the Chaoshan sample set alone, and 87% and 73% power for the combined Chinese sample set in association with markers respectively rs13382811 and rs6469937 with a p-value < 0.05 under a codominant model and an odds ratio (OR) = 2.
Table 1. Baseline characteristics of each cohort in this study.
| Parameters |
Chaoshan |
Guangzhou |
Combined Han Chinese4 |
KORA |
TwinsUK |
||||
|---|---|---|---|---|---|---|---|---|---|
| Cases1 | Controls | Cases2 | Controls | Cases | Controls | Cases3 | Controls | Cases3 | |
| N |
96 |
96 |
300 |
308 |
396 |
404 |
526 |
413 |
562 |
| Sex (%Male) |
65.63 |
65.63 |
49.17 |
63.64 |
53.15 |
64.11 |
50.38 |
48.42 |
16.20 |
| Mean age (SD) |
21.80 (1.27) |
21.68 (1.30) |
22.19 (1.67) |
21.66 (1.54) |
22.10 (4.77) |
21.67 (1.48) |
49.25(10.21) |
57.23 (10.72) |
46.46 (14.99) |
|
Refraction (Diotpers) |
|
|
|
|
|
|
|
|
|
| OD |
−6.54±1.31 |
0.27±0.51 |
−7.82±0.87 |
0.19±0.48 |
−7.51±1.14 |
0.21±0.49 |
−2.50±1.87 |
0.23±0.42 |
−3.26±2.73 |
| OS | −6.28±1.84 | 0.33±0.44 | −7.79±1.12 | 0.31±0.44 | −7.42±1.48 | 0.31±0.44 | −2.49±1.87 | 0.23±0.42 | −3.16±2.67 |
1. Cases with moderate to high myopia were defined as SEM ≤ −4.0 D. 2. Cases with high myopia were defined as SEM ≤ −6.0 D 3. Cases with myopia were defined as SE< −0.5 D. 4. Han Chinese group=Chaoshan + Guangzhou. 5. European group=KORA+TwinsUK 2.
Table 2. Association results of rs13382811 and rs6469937 in the Guangzhou cohort.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=300) | controls (n=308) | myopia (n=299) | controls (n=308) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C |
437 (72.8%) |
504 (81.8%) |
A |
136 (22.7%) |
181 (29.4%) |
| |
T* |
163 (27.2%) |
112 (18.2%) |
G* |
462 (77.3%) |
435 (70.6%) |
| HWE P |
|
0.05 |
0.05 |
|
0.42 |
0.35 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.0001 |
1.68 (1.28–2.20) |
|
0.0085 |
0.71 (0.55–0.92) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
166 (55.3%) |
201 (65.3%) |
AA |
13 (4.3%) |
30 (9.7%) |
| |
CT |
105 (35.0%) |
102 (33.1%) |
AG |
110 (36.8%) |
121 (39.3%) |
| |
TT |
29 (9.7%) |
5 (1.6%) |
GG |
176 (58.9%) |
157 (51.0%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
4.07×10−5 |
|
|
0.0166 |
|
| Additive (Trend) |
|
0.0002 |
|
|
0.0090 |
|
| Dominant |
|
0.0124 |
1.52 (1.09–2.10) |
|
0.0509 |
0.73 (0.53–1.00) |
| Recessive | 1.59×10−5 | 6.48 (2.48–16.99) | 0.0096 | 0.42 (0.22–0.82) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Table 3. Association results of rs13382811 and rs6469937 in the Chaoshan cohort.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=96) | controls (n=96) | myopia (n=95) | controls (n=95) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C* |
158 (82.3%) |
152 (79.2%) |
A |
41 (21.6%) |
56 (29.5%) |
| |
T |
34 (17.7%) |
40 (20.8%) |
G* |
149 (78.4%) |
134(70.5%) |
| HWE P |
|
0.16 |
0.26 |
|
0.80 |
0.11 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.4376 |
0.82 (0.49–1.36) |
|
0.0780 |
0.66 (0.41–1.05) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
63 (65.6%) |
62 (64.6%) |
AA |
4 (4.2%) |
5 (5.3%) |
| |
CT |
32 (33.3%) |
28 (29.2%) |
AG |
33 (34.7%) |
46 (48.4%) |
| |
TT |
1 (1.0%) |
6 (6.3%) |
GG |
58 (61.1%) |
44 (46.3%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
0.1462 |
|
|
0.1242 |
|
| Additive (Trend) |
|
0.4378 |
|
|
0.0645 |
|
| Dominant |
|
0.8797 |
0.96 (0.53–1.73) |
|
0.0417 |
0.55 (0.31–0.98) |
| Recessive | 0.0542 | 0.16 (0.02–1.34) | 0.7327 | 0.79 (0.21–3.04) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Table 4. Association results of rs13382811 and rs6469937 in combined cohorts of Guangzhou and Chaoshan.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=396) | controls (n=404) | myopia (n=394) | controls (n=403) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C |
595 (75.1%) |
656 (81.2%) |
A |
177 (22.5%) |
237 (29.4%) |
| |
T* |
197 (24.9%) |
152 (18.8%) |
G* |
611 (77.5%) |
569 (70.6%) |
| HWE P |
|
0.14 |
0.28 |
|
0.40 |
0.97 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.0033 |
1.42 (1.13–1.81) |
|
0.0016 |
0.70 (0.55–0.87) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
229 (57.8%) |
263 (65.1%) |
AA |
17 (4.3%) |
35 (8.7%) |
| |
CT |
137 (34.6%) |
130 (32.2%) |
AG |
143 (36.3%) |
167 (41.4%) |
| |
TT |
30 (7.6%) |
11 (2.7%) |
GG |
234 (59.4%) |
201 (49.9%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
0.0036 |
|
|
0.0053 |
|
| Additive (Trend) |
|
0.0037 |
|
|
0.0015 |
|
| Dominant |
|
0.0346 |
1.36 (1.02–1.81) |
|
0.0070 |
0.68 (0.51–0.90) |
| Recessive | 0.0019 | 2.93 (1.45–5.93) | 0.0125 | 0.47 (0.26–0.86) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Additional replication in European individuals included 526 unrelated subjects with myopia and 413 emmetropic controls from the Kooperative Gesundheitsforschung in der Region Augsburg (KORA; Cooperative Health Research in the Region of Augsburg) cohort and 562 unrelated subjects with myopia and 922 emmetropic controls from the TwinsUK cohort. Phenotype data of both cohorts were publicly available through the database of Genotypes and Phenotypes (dbGaP) after approval of controlled access to the data. Individuals with bilateral SE < −0.5 D were included in the case group. Criteria for emmetropic controls were the same as those described above. Power calculations using the PGA program [29] and a myopia and disease allele prevalence of 30% [31], as well as the marker allele frequencies for controls in each population shown in Table 5, Table 6, and Table 7, exhibited 99% and 99% power for the KORA sample set alone, 97% and 99% for the TwinsUK sample set alone, and 99% and 99% power for the combined European sample set to detect the association with markers rs13382811 and rs6469937, respectively, with a p < 0.05 under a codominant model assuming an OR = 2.
Table 5. Association results of rs13382811 and rs6469937 in the KORA cohort.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=526) | controls (n=413) | myopia (n=526) | controls (n=413) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C* |
928 (88.2%) |
718 (86.9%) |
A* |
670 (63.7%) |
510 (61.7%) |
| |
T |
124 (11.8%) |
108 (13.1%) |
G |
382 (36.3%) |
316 (38.3%) |
| HWE P |
|
0.77 |
0.68 |
|
0.21 |
0.91 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.3998 |
0.89 (0.67–1.17) |
|
0.3867 |
0.92 (0.76–1.11) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
410 (78.0%) |
313 (75.8%) |
AA |
220 (41.8%) |
158 (38.3%) |
| |
CT |
108 (20.5%) |
92 (22.3%) |
AG |
230 (43.7%) |
194 (47.0%) |
| |
TT |
8 (1.5%) |
8 (1.9%) |
GG |
76 (14.4%) |
61 (14.8%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
0.7027 |
|
|
0.5251 |
|
| Additive (Trend) |
|
0.4039 |
|
|
0.3947 |
|
| Dominant |
|
0.4350 |
0.89 (0.65–1.20) |
|
0.2684 |
0.86 (0.66–1.12) |
| Recessive | 0.6248 | 0.78 (0.30–2.10) | 0.8900 | 0.97(0.68–1.40) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Table 6. Association results of rs13382811 and rs6469937 in the TwinsUK cohort.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=562) | controls (n=922) | myopia (n=562) | controls (n=922) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C* |
963 (85.68%) |
1531 (83.03%) |
A |
720 (64.06%) |
1239 (67.19%) |
| |
T |
161 (14.32%) |
313 (16.97%) |
G* |
404 (35.94%) |
605 (32.81%) |
| HWE P |
|
0.06 |
0.42 |
|
0.24 |
0.68 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.0559 |
0.82 (0.66–1.00) |
|
0.0804 |
1.15 (0.98–1.34) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
418 (74.38%) |
639 (69.31%) |
AA |
237 (42.10%) |
419 (45.44%) |
| |
CT |
127 (22.56%) |
253 (27.44%) |
AG |
246 (43.77%) |
401 (43.49%) |
| |
TT |
17 (3.02%) |
30 (3.25%) |
GG |
79 (14.03%) |
102 (11.06%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
0.1046 |
|
|
0.1809 |
|
| Additive (Trend) |
|
0.0617 |
|
|
0.0848 |
|
| Dominant |
|
0.0364 |
0.78 (0.61–0.98) |
|
0.218 |
1.14 (0.92–1.41) |
| Recessive | 0.807 | 0.93 (0.51–1.70) | 0.0873 | 1.31 (0.96–1.80) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Table 7. Association results of rs13382811 and rs6469937 in combined KORA and TwinsUK cohorts.
|
SNPs |
rs13382811 (ZFHX1B) |
rs6469937 (SNTB1) |
||||
|---|---|---|---|---|---|---|
| Phenotype | myopia (n=1088) | controls (n=1335) | myopia (n=1088) | controls (n=1335) | ||
| Allelic Analysis |
|
|
|
|
|
|
| Alleles (%) |
C* |
1891 (86.90%) |
2249 (84.23%) |
A* |
1445 (66.41%) |
1694 (63.45%) |
| |
T |
285 (13.10%) |
421 (15.77%) |
G |
731 (33.59%) |
976 (36.55%) |
| HWE P |
|
0.09 |
0.32 |
|
0.76 |
0.08 |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Allelic P Values |
|
0.0088 |
0.80 (0.68–0.95) |
|
0.0319 |
0.88 (0.78–0.99) |
| Genotypic and Model Based Analysis |
|
|
|
|
||
| Genotypes (%) |
CC |
828 (76.10%) |
952 (71.31%) |
AA |
482 (44.30%) |
552 (41.35%) |
| |
CT |
235 (21.60%) |
345 (25.84%) |
AG |
481 (44.21%) |
590 (44.19%) |
| |
TT |
25 (2.30%) |
38 (2.85%) |
GG |
125 (11.49%) |
193 (14.46%) |
| |
|
P# |
OR§ (95%CI) |
|
P# |
OR§ (95%CI) |
| Genotypic test |
|
0.0290 |
|
|
0.0725 |
|
| Additive (Trend) |
|
0.0101 |
|
|
0.0346 |
|
| Dominant |
|
0.0076 |
0.78 (0.65–0.94) |
|
0.1438 |
0.89 (0.75–1.04) |
| Recessive | 0.3986 | 0.80 (0.48–1.34) | 0.0314 | 0.77 (0.60–0.98) | ||
*risk allele. # Chi-square p value. The correected significance level was 0.005 (α<0.05/10). §Odds ratios are calculated for the minor allele and genotypes.
Genotyping
Genotyping of the Guangzhou and Chaoshan cohorts was performed as described previously [28]. Primers for SNPs rs13382811 and rs6469937 were designed using Primer 3 v.0.4.0. For rs13382811, the forward primer was 5′-TCC TCT ACA CCG TCG ATT GC-3′, and the reverse was 5′-CCC CCT AAG GGA GAT ACA CTG-′3; for rs6469937, the forward primer was 5′-GCT CAC TGG GAA AAC TAT GTG C-′3, and the reverse was 5′-TGC AGA GAT GAG GAT GAA GAA G-3′. Genotype data from the KORA and TwinsUK cohorts were retrieved from dbGaP, except that the rs6469937 alleles of the KORA cohort were imputed from the 1000 Genomes Project phase 1 reference panel using BEAGLE version 3.3.2 with default values using 900 adjacent SNPs [32].
Statistical analysis
Genotype association tests were implemented in Golden Helix SVS software version 8.4.1 (Golden Helix, Bozeman, MT). Because age and gender were relatively closely matched between cases, and controls and initial analysis using logistic regression showed only small effects of control for these differences, basic allelic, genotypic, and model-based (additive, dominant, and recessive) associations of each SNP with myopia were analyzed using the Chi-square test. The OR and 95% confidence interval (CI) were also estimated. The Chi-square test was used to evaluate the deviation of Hardy–Weinberg equilibrium for each SNP in the case and control groups. Multiple testing corrections were done using a Bonferroni correction. Principal components analysis (PCA) was used to assess possible population stratification in the KORA and TwinsUK cohorts, with a subset of 311,792 autosomal SNPs. Heterogeneity of the ORs was tested using the Breslow–Day test. SNP–SNP interactions were evaluated using logistic regression based on a recessive model, and p-values for association of both SNPs combined were estimated using Fisher’s exact test.
Results
A total of 800 unrelated subjects of Han Chinese ancestry were enrolled in this study, including 300 unrelated individuals with high myopia and 308 emmetropic controls in Guangzhou of other than Chaoshanese origin, and 96 unrelated subjects with moderate to high myopia and 96 emmetropic controls in Guangzhou but from Chaoshan. Two additional cohorts of European ancestry available through dbGAP included 526 unrelated subjects with myopia and 413 emmetropic controls from the KORA cohort and 562 unrelated subjects with myopia and 922 emmetropic controls from the TwinsUK cohort.
The baseline characteristics of the subjects are shown in Table 1. The details of the clinical data from individuals of Han Chinese ancestry (Guangzhou and Chaoshan) were described previously [28]. Briefly, the sex and the age between cases and controls were similar, at 50–65% male and with an average age of 21.6–22.2 years. The refraction between the left and right eyes showed no statistical difference. The refraction in diopters in cases between the Chaoshan and Guangzhou cohorts were similar. The average refraction in Chaoshan was −6.54 D for the right eyes and −6.28 D for the left eyes, while that of Guangzhou was −7.82 D for the right eyes and −7.79 D for the left eyes. The clinical data of subjects in the KORA and TwinsUK cohorts are also summarized in Table 1. No significant difference in average age was observed between cases and controls for each cohort, although the percentage of males was higher in controls than cases in the Guangzhou group (63.6% versus 49.2%). The average refractions in the right and left eyes in cases from the KORA study were −2.50 D and −2.49 D, while those from the TwinsUK study were −3.26 D and −3.16 D, respectively, so that the myopia was of a lower grade in the European compared with the Chinese group. The refractions in controls for all cohorts were close to plano.
SNP rs13382811, near ZFHX1B, was genotyped in all individuals in the Guangzhou and Chaoshan cohorts, while genotyping of SNP rs6469937, near SNTB1, failed in three samples, including one case in the Guangzhou cohort, one case in the Chaoshan cohort, and one control in the Chaoshan cohort. Genotype data from the KORA and TwinsUK cohorts were available to be downloaded or imputed for all included individuals. Both SNPs were in the Hardy–Weinberg equilibrium (p > 0.05) in both cases and controls for each cohort (Table 2, Table 3, Table 4, Table 5, Table 6, and Table 7).
In the Han Chinese group, both SNPs (rs13382811 and rs6469937) showed significant association with high myopia in the Guangzhou cohort (Table 2) but not with moderate to high myopia in the Chaoshan cohort (Table 3). In the Guangzhou cohort, the minor allele (T) of SNP rs13382811 was significantly associated with high myopia (p = 0.0001, OR = 1.68, 95%CI=1.28–2.20), and the genotypic test gave p = 4.07 × 10−5. Since the allelic and genotypic models showed significant associations, specific models were tested, and these also showed significant associations. The recessive model showed the highest level of association (p = 1.59 × 10−5; OR = 6.48, 95%CI=2.48–16.99), the additive model gave p = 2×10−4, and the dominant model showed only a suggestive association, with p = 0.0124. The second SNP, rs6469937, showed nominal evidence of association with high myopia: The minor allele (A) of rs6469937 showed a protective effect on high myopia (p = 0.0085; OR = 0.71, 95%CI=0.55–0.92), the genotypic test gave p = 0.0166, the additive model gave p = 0.0090, the dominant model gave p = 0.0509, and the recessive model gave p = 0.0096. However, this suggestive association did not withstand a Bonferroni correction for multiple testing (α < 0.05/10). While analysis of the Chaoshanese group did not show significant association, there was a suggestive protective effect of the A allele of rs6469937 in a dominant model that did not withstand multiple testing (p = 0.0417; OR = 0.55, 95% CI=0.31–0.98). Because of the small size of the Chaoshan group and their common Han heritage, these two groups were combined and analyzed together, as in our previous work [28].
When the Guangzhou and Chaoshan cohorts were combined into a single analysis, the Breslow–Day test showed no significant evidence of heterogeneity in the ORs for each SNP (p > 0.05), and both SNPs showed significant associations with moderate to high myopia (Table 4). For rs13382811, the basic allelic test gave p = 0.0033, OR = 1.42 (CI=1.13–1.81), the genotypic test gave p = 0.0036, and tests of specific models for additive, dominant, and recessive gave p = 0.0037, 0.0346, and 0.0019, respectively. The association results of rs6469937 showed p = 0.0016 and OR = 0.70 (0.55–0.87) for allelic association; 0.0053 for the genotypic test; and 0.0015, 0.0070, and 0.0125, for the additive, dominant, and recessive models, respectively. Both SNPs remained significant after adjusting for multiple testing (α < 0.05/10), except for rs6469937 under the dominant and recessive models.
Examination of the ORs for genotypes of rs6469937 and rs13382811 considered together in the combined Han Chinese Guangdong and Chaoshan groups showed that the risk of myopia is much higher in individuals with homozygous risk alleles (GG for rs6469937 and TT for rs13382811) for both genes than for homozygous protective alleles for both, p = 8.37 × 10−4 and OR = 6.57 (2.3–20.5), with the ORs increasing with additional copies of risk alleles from either locus (Figure 1). It also confirmed that the risk for the rs6469937 G allele is additive, while that for rs13382811 lies between the additive and recessive models. However, ignoring individuals with double homozygote minor alleles, of whom there were too few to provide statistical significance in this group, this suggests that the rs13382811 T allele only increases the risk of myopia in individuals with at least one rs6469937 G allele, and it does not increase this risk in individuals that are homozygous for the rs6469937 A allele. In addition, the rs6469937–rs13382811 interaction effect was evaluated using logistic regression, with a recessive model giving the best results (p = 0.001 for the full model). When both SNPs were considered together under the recessive model, individuals with rs13382811 risk genotype TT and rs6469937 risk genotype GG together had an increased risk of moderate to high myopia compared to individuals with at least one C allele for rs13382811 and at least one A allele for rs6469937 (p = 8.37 × 10−4; OR = 4.09, 95%CI = 1.7–9.8; Figure 2). Again, there was no evidence of a multiplicative effect between these two SNPs (p = 0.9).
Figure 1.
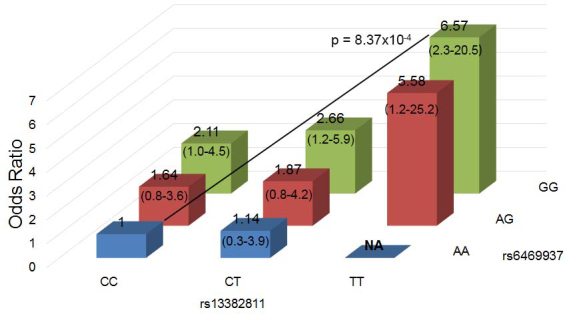
Interaction between the rs13382811 and rs6469937 alleles in terms of myopia risk in the combined Han Chinese groups. The odds ratios (ORs) of combinations of genotypes at rs13382811 and rs6469937 are given relative to the homozygous protective genotypes, namely CC for rs13382811 and AA for rs6469937. NA: the odds ratio was not available because of the small number of samples with TT for rs13382811 and AA for rs6469937 (none in the case group and only one sample in the control group).
Figure 2.

Interaction between rs13382811 and rs6469937 in a Han Chinese population under a recessive model. Individuals are grouped by the presence of the homozygous risk genotypes and compared with risks with at least one protective allele for rs13382811 (risk allele = T, protective allele = C) or rs6469937 (risk allele = G, protective allele = A). The odds ratios (ORs) are given relative to those samples with at least one protective allele for both markers.
Additional analysis of both SNPs in European-derived populations did not show significant associations with myopia in KORA (Table 5), and it was only suggestive of an association with the dominant model for rs13382811 (p = 0.0364; OR = 0.78, 95%CI = 0.61–0.98) that could not withstand correction for multiple testing in the TwinsUK cohort (Table 6). The Breslow–Day test showed no evidence of heterogeneity of the OR (p > 0.05), and a PCA showed no evidence of population stratification between the KORA and TwinsUK cohorts (Figure 3). The combined European cohorts showed a suggestive association of myopia, with pallelic = 0.0088 (OR = 0.8, 95%CI = 0.68–0.95) for rs13382811 and pallelic = 0.0319 (OR=0.88, 95%CI 0.78–0.99) for rs6469937 (Table 7). Examining specific inheritance models, rs13382811 showed the best p(0.0076) with a dominant model and a protective effect of homozygous TT combined with CT genotypes on myopia (OR=0.78, 95%CI= 0.65–0.94). With rs6469937, a best p of 0.0314 and a protective effect of the GG genotype on myopia (OR=0.77, 95%CI=0.60–0.98) was seen under a recessive model (Table 7). The results did not remain significant for either SNP after Bonferroni correction. However, when the effects of both SNPs were considered together under a recessive model, individuals’ homozygous AA genotypes for rs6469937 showed a suggestive increase in risk (p = 0.01; OR = 1.53, 95%CI = 1.1–2.1) in the presence of at least one T allele of rs13382811, while CC homozygotes for rs13382811 showed a significant increase in risk in the presence of at least one G allele for rs6469937 (p = 7.2 × 10−4; OR = 1.52, 95% CI = 1.1–2.1). Double homozygotes for the risk alleles, CC for rs13382811 and AA for rs6469937, also showed a significant increase in risk for myopia (p = 8.2 × 10−4, OR = 1.56, 95%CI = 1.2–2.0), so that the presence of a single homozygous risk allele for either or both markers appeared to provide an approximately similar increase in risk as homozygosity for both (Figure 4). SNP–SNP interaction was also accessed using logistic regression, showing a similar pattern to that seen in the Han Chinese cohorts, with no multiplicative interactive effect between these two SNPs (p > 0.05).
Figure 3.

Plot of the principal component analysis for the Kooperative Gesundheitsforschung in der Region Augsburg (KORA) and TwinsUK cohorts. The KORA cohort is shown in blue, while the TwinsUK cohort is shown in green. Principal component 1 (EV = 4.33712) was plotted against principal component 2 (EV = 1.15219) in these two cohorts.
Figure 4.

Interaction between rs13382811 and rs6469937 in the combined European population groups under a recessive model. Individuals are grouped by the presence of the homozygous risk genotypes and compared to risks with at least one protective allele for rs13382811 (risk allele = C, protective allele = T) or rs6469937 (risk allele = A, protective allele = G). The odds ratios (ORs) are given relative to those samples with at least one protective allele for both markers. It should be noted that these results are reversed from those in the Han Chinese population.
Discussion
In this study, we confirmed the significant associations of rs13382811 and rs6469937 with moderate to high myopia in a Han Chinese population, although additional replication analyses were only confirmed in European patients with lower levels of myopia when the genotypes at both loci were considered. Although the effects of rs13382811 appeared to be limited to individuals with at least one rs6469937 G allele, the effect of each SNP on the risk of myopia was independent and did not exhibit a multiplicative interaction effect.
Our results showed significant associations of SNPs rs6469937 and rs13382811 with high myopia in the Guangzhou cohort and moderate to high myopia in a combined cohort (Guangzhou and Chaoshan), but not in the Chaoshan cohort, a distinct population that was used for association analysis in our previous study [28]. The absence of significant associations detected in this cohort may simply be due to the relatively small sample size, resulting in a lack of power to detect a significant association.
SNP rs13382811, located in the intron of ZFHX1B, was reported to show significant evidence of association with high myopia in a total of 2,835 unrelated subjects with high myopia and 6,986 controls (p = 5.79 × 10−10; OR = 1.26) from an Asian population [27]. Four SNPs located in SNTB1 were previously reported to be associated with high myopia in two GWAS [27,33]. The first GWAS meta-analysis showed three SNPs (rs7839488, rs4395927, and rs4455882) in the same linkage disequilibrium block associated with high myopia from Han Chinese ancestry [33]. In the second GWAS meta-analysis, which reported on associations of rs13382811 in ZFHX1B [27], these three SNPs only showed a suggestive p < 10−7 at the GWAS stage, but another SNP (rs6469937) in the same linkage disequilibrium group showed a significant association with high myopia (p = 2.01 × 10−9; OR = 0.79) in Asian populations. The current study of SNPs rs13382811 and rs6469937 showed significant associations with moderate to high myopia, and these results were consistent with those of previous studies, confirming that ZFHX1B and SNTB1 are susceptibility loci for high myopia in Han Chinese populations.
In contrast, the individual evaluation of each locus in a European-derived population showed only nominal evidence of association with myopia, which could not withstand correction for multiple testing. However, when their effects were considered together, they did reach statistical significance. Part of the explanation for the lack of effect of the individual SNPs could relate to the apparently similar effects of homozygosity for the risk allele from either locus to homozygosity for both loci, which could dilute the individual effects of each locus when examined separately (Figure 4). While the trends for both markers were weak, they were in the opposite direction from those in Han Chinese individuals. The C and A alleles were protective in the Asian groups, while both nominally increased the risk in Europeans, and the 95% CIs for the ORs did not overlap for any test, suggesting that the difference may be real. It should be noted that since the ORs are given for the minor alleles in Table 4 and Table 7 (for rs6469937 A in Han Chinese and G in Europeans), they need to be inverted for comparison between these two population. It is well established that genetic associations are different among populations, often due to differences in the background haplotype of novel mutations or variations in allele frequencies caused by genetic drift or founder effects in separate populations [34]. Thus, in Asian populations, the T allele of rs13382811 was associated with an increase in risk for myopia [27] and Table 4, but appeared nominally protective in European populations (Table 7). Similarly, the G allele rs6469937 increased myopia risk as the major allele in Asian populations, while it showed a slight protective effect as a minor allele in European populations (Table 4 and Table 7). A second and critical part of the explanation for the lack of effect of the individual SNPs is that, the phenotype used in the KORA and TwinsUK cohorts is myopia with bilateral SE < −0.5 D, which is considerably broader than that used in the Guangdong and Chaoshan groups, SE < −6.0 and −4.5 D, respectively. Finally, myopia is a complex trait caused by an interplay of genetic and environmental factors. The inconsistency in associations may have been due to differences in environmental exposure between the populations interacting with the diverse genetic factors in each group.
The zinc finger E-box-binding homeobx2 protein (ZEB2), encoded by ZFHX1B, is a transcriptional repressor that interacts with activated SMAD proteins to influence the transforming growth factor β (TGF-β) signaling pathway, which has been implicated in several myopiagenic mechanisms, including ocular growth and scleral metabolism [35,36]. Consistent with this, ZFHX1B is strongly expressed in the retina, RPE, sclera, and lens [27,33,37]. In experimental myopia, differential ZFHX1B expression was observed between induced myopic eyes, fellow eyes, and untreated control eyes. Moreover, somatic deletion of Zeb2 in mouse retinal progenitors resulted in abnormal numbers of retinal cells in the inner nuclear layer, suggesting that it has a critical role in retinal development [38]. Mutations in ZFHX1B are associated with Mowat–Wilson syndrome/syndromic Hirschsprung disease, which is characterized by multiple congenital abnormalities, commonly including typical facial features, intellectual disability, delayed development, Hirschsprung disease, and other birth defects. Although ocular disorders are rarely seen in this syndrome, a patient with Down syndrome, Hirschsprung disease with high myopia, and ocular coloboma was reported to carry a de novo missense mutation (p.R953G) in ZFHX1B in addition to trisomy 21 [39]. Combined with its genetic association with high myopia, these findings suggest that ZFHX1B may have a potential role in the pathogenicity of high myopia.
SNTB1 was also reported to be associated with high myopia in two previous studies [27,33]. Differential expressions of SNTB1 in the mouse retina, RPE, and sclera were also observed in induced myopic eyes compared with fellow eyes and control eyes [27]. Although no multiplicative interactive effect was found between ZFHX1B and SNTB1, in this work, their apparently true dominant interaction in Europeans and the apparent requirement for at least one rs6469937 G allele for association of the rs13382811 T allele with high myopia in the Guangdong cohort suggests that these two loci may affect common pathways in the development of high myopia. However, no common pathways were identified using the STRING program [40], and their functional relationship with high myopia is currently unclear and needs to be studied.
In conclusion, we confirmed significant associations of two SNPs, rs13382811 in ZFHX1B and rs6469937 in SNTB1, with moderate to high myopia in the Han Chinese population. However, we were unable to confirm this association for each individual SNP in patients from two studies in the European population, although significant associations were observed when genotypes from both SNPs were considered together. The risk for myopia increases markedly in the presence of risk alleles from both loci, to the extent that the effect of rs13382811 is only significant in the presence of at least one rs7479937 risk allele. However, their interaction appears to be primarily additive, as no multiplicative interaction could be identified with logistic regression. Taken together with related functional evidence, confirmation of these two susceptibility loci in Asian populations strongly supports their having a potential role in the pathogenesis of high myopia.
Acknowledgments
The authors thank all of the patients and controls for their participation in this study. Supported by NEI project EYE000272, the National Natural Science Foundation of China (U1201221), the Natural Science Foundation of Guangdong (S2013030012978), and the Fundamental Research Funds of the State Key Laboratory of Ophthalmology.
References
- 1.Wojciechowski R. Nature and nurture: the complex genetics of myopia and refractive error. Clin Genet. 2011;79:301–20. doi: 10.1111/j.1399-0004.2010.01592.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Schwartz M, Haim M, Skarsholm D. X-linked myopia: Bornholm eye disease. Linkage to DNA markers on the distal part of Xq. Clin Genet. 1990;38:281–6. [PubMed] [Google Scholar]
- 3.Young TL, Ronan SM, Alvear AB, Wildenberg SC, Oetting WS, Atwood LD, Wilkin DJ, King RA. A second locus for familial high myopia maps to chromosome 12q. Am J Hum Genet. 1998;63:1419–24. doi: 10.1086/302111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Young TL, Ronan SM, Drahozal LA, Wildenberg SC, Alvear AB, Oetting WS, Atwood LD, Wilkin DJ, King RA. Evidence that a locus for familial high myopia maps to chromosome 18p. Am J Hum Genet. 1998;63:109–19. doi: 10.1086/301907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Paluru P, Ronan SM, Heon E, Devoto M, Wildenberg SC, Scavello G, Holleschau A, Makitie O, Cole WG, King RA, Young TL. New locus for autosomal dominant high myopia maps to the long arm of chromosome 17. Invest Ophthalmol Vis Sci. 2003;44:1830–6. doi: 10.1167/iovs.02-0697. [DOI] [PubMed] [Google Scholar]
- 6.Stambolian D, Ibay G, Reider L, Dana D, Moy C, Schlifka M, Holmes T, Ciner E, Bailey-Wilson JE. Genomewide Linkage Scan for Myopia Susceptibility Loci among Ashkenazi Jewish Families Shows Evidence of Linkage on Chromosome 22q12. Am J Hum Genet. 2004;75:448–59. doi: 10.1086/423789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hammond CJ, Andrew T, Mak YT, Spector TD. A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11: a genomewide scan of dizygotic twins. Am J Hum Genet. 2004;75:294–304. doi: 10.1086/423148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang Q, Guo X, Xiao X, Jia X, Li S, Hejtmancik JF. A new locus for autosomal dominant high myopia maps to 4q22-q27 between D4S1578 and D4S1612. Mol Vis. 2005;11:554–60. [PubMed] [Google Scholar]
- 9.Paluru PC, Nallasamy S, Devoto M, Rappaport EF, Young TL. Identification of a novel locus on 2q for autosomal dominant high-grade myopia. Invest Ophthalmol Vis Sci. 2005;46:2300–7. doi: 10.1167/iovs.04-1423. [DOI] [PubMed] [Google Scholar]
- 10.Zhang Q, Guo X, Xiao X, Jia X, Li S, Hejtmancik JF. Novel locus for X linked recessive high myopia maps to Xq23-q25 but outside MYP1. J Med Genet. 2006;43:e20. doi: 10.1136/jmg.2005.037853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Guo X, Xiao X, Li S, Wang P, Jia X, Zhang Q. Nonsyndromic high myopia in a Chinese family mapped to MYP1: linkage confirmation and phenotypic characterization. Arch Ophthalmol. 2010;128:1473–9. doi: 10.1001/archophthalmol.2010.270. [DOI] [PubMed] [Google Scholar]
- 12.Wojciechowski R, Moy C, Ciner E, Ibay G, Reider L, Bailey-Wilson JE, Stambolian D. Genomewide scan in Ashkenazi Jewish families demonstrates evidence of linkage of ocular refraction to a QTL on chromosome 1p36. Hum Genet. 2006;119:389–99. doi: 10.1007/s00439-006-0153-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nallasamy S, Paluru PC, Devoto M, Wasserman NF, Zhou J, Young TL. Genetic linkage study of high-grade myopia in a Hutterite population from South Dakota. Mol Vis. 2007;13:229–36. [PMC free article] [PubMed] [Google Scholar]
- 14.Lam CY, Tam PO, Fan DS, Fan BJ, Wang DY, Lee CW, Pang CP, Lam DS. A genome-wide scan maps a novel high myopia locus to 5p15. Invest Ophthalmol Vis Sci. 2008;49:3768–78. doi: 10.1167/iovs.07-1126. [DOI] [PubMed] [Google Scholar]
- 15.Paget S, Julia S, Vitezica ZG, Soler V, Malecaze F, Calvas P. Linkage analysis of high myopia susceptibility locus in 26 families. Mol Vis. 2008;14:2566–74. [PMC free article] [PubMed] [Google Scholar]
- 16.Yang Z, Xiao X, Li S, Zhang Q. Clinical and linkage study on a consanguineous Chinese family with autosomal recessive high myopia. Mol Vis. 2009;15:312–8. [PMC free article] [PubMed] [Google Scholar]
- 17.Ma JH, Shen SH, Zhang GW, Zhao DS, Xu C, Pan CM, Jiang H, Wang ZQ, Song HD. Identification of a locus for autosomal dominant high myopia on chromosome 5p13.3-p15.1 in a Chinese family. Mol Vis. 2010;16:2043–54. [PMC free article] [PubMed] [Google Scholar]
- 18.Aldahmesh MA, Khan AO, Alkuraya H, Adly N, Anazi S, Al-Saleh AA, Mohamed JY, Hijazi H, Prabakaran S, Tacke M, Al-Khrashi A, Hashem M, Reinheckel T, Assiri A, Alkuraya FS. Mutations in LRPAP1 Are Associated with Severe Myopia in Humans. Am J Hum Genet. 2013 doi: 10.1016/j.ajhg.2013.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guo H, Jin X, Zhu T, Wang T, Tong P, Tian L, Peng Y, Sun L, Wan A, Chen J, Liu Y, Li Y, Tian Q, Xia L, Zhang L, Pan Y, Lu L, Liu Q, Shen L, Li Y, Xiong W, Li J, Tang B, Feng Y, Zhang X, Zhang Z, Pan Q, Hu Z, Xia K. SLC39A5 mutations interfering with the BMP/TGF-beta pathway in non-syndromic high myopia. J Med Genet. 2014;51:518–25. doi: 10.1136/jmedgenet-2014-102351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jiang D, Li J, Xiao X, Li S, Jia X, Sun W, Guo X, Zhang Q. Detection of mutations in LRPAP1, CTSH, LEPREL1, ZNF644, SLC39A5, and SCO2 in 298 families with early-onset high myopia by exome sequencing. Invest Ophthalmol Vis Sci. 2014;56:339–45. doi: 10.1167/iovs.14-14850. [DOI] [PubMed] [Google Scholar]
- 21.Tran-Viet KN, Powell C, Barathi VA, Klemm T, Maurer-Stroh S, Limviphuvadh V, Soler V, Ho C, Yanovitch T, Schneider G, Li YJ, Nading E, Metlapally R, Saw SM, Goh L, Rozen S, Young TL. Mutations in SCO2 are associated with autosomal-dominant high-grade myopia. Am J Hum Genet. 2013;92:820–6. doi: 10.1016/j.ajhg.2013.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mordechai S, Gradstein L, Pasanen A, Ofir R, El Amour K, Levy J, Belfair N, Lifshitz T, Joshua S, Narkis G, Elbedour K, Myllyharju J, Birk OS. High myopia caused by a mutation in LEPREL1, encoding prolyl 3-hydroxylase 2. Am J Hum Genet. 2011;89:438–45. doi: 10.1016/j.ajhg.2011.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li J, Gao B, Xiao X, Li S, Jia X, Sun W, Guo X, Zhang Q. Exome sequencing identified null mutations in LOXL3 associated with early-onset high myopia. Mol Vis. 2016;22:161–7. [PMC free article] [PubMed] [Google Scholar]
- 24.Li J, Gao B, Guan L, Xiao X, Zhang J, Li S, Jiang H, Jia X, Yang J, Guo X, Yin Y, Wang J, Zhang Q. Unique Variants in OPN1LW Cause Both Syndromic and Nonsyndromic X–Linked High Myopia Mapped to MYP1. Invest Ophthalmol Vis Sci. 2015;56:4150–5. doi: 10.1167/iovs.14-16356. [DOI] [PubMed] [Google Scholar]
- 25.Xiao X, Li S, Jia X, Guo X, Zhang Q. X-linked heterozygous mutations in ARR3 cause female-limited early onset high myopia. Mol Vis. 2016;22:1257–66. [PMC free article] [PubMed] [Google Scholar]
- 26.Shi Y, Li Y, Zhang D, Zhang H, Lu F, Liu X, He F, Gong B, Cai L, Li R, Liao S, Ma S, Lin H, Cheng J, Zheng H, Shan Y, Chen B, Hu J, Jin X, Zhao P, Chen Y, Zhang Y, Lin Y, Li X, Fan Y, Yang H, Wang J, Yang Z. Exome sequencing identifies ZNF644 mutations in high myopia. PLoS Genet. 2011;7:e1002084. doi: 10.1371/journal.pgen.1002084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Khor CC, Miyake M, Chen LJ, Shi Y, Barathi VA, Qiao F, Nakata I, Yamashiro K, Zhou X, Tam PO, Cheng CY, Tai ES, Vithana EN, Aung T, Teo YY, Wong TY, Moriyama M, Ohno-Matsui K, Mochizuki M, Matsuda F. Nagahama Study G, Yong RY, Yap EP, Yang Z, Pang CP, Saw SM, Yoshimura N. Genome-wide association study identifies ZFHX1B as a susceptibility locus for severe myopia. Hum Mol Genet. 2013;22:5288–94. doi: 10.1093/hmg/ddt385. [DOI] [PubMed] [Google Scholar]
- 28.Jiao X, Wang P, Li S, Li A, Guo X, Zhang Q, Hejtmancik JF. Association of markers at chromosome 15q14 in Chinese patients with moderate to high myopia. Mol Vis. 2012;18:2633–46. [PMC free article] [PubMed] [Google Scholar]
- 29.Menashe I, Rosenberg PS, Chen BE. PGA: power calculator for case-control genetic association analyses. BMC Genet. 2008;9:36. doi: 10.1186/1471-2156-9-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rose KA, French AN, Morgan IG. Environmental Factors and Myopia: Paradoxes and Prospects for Prevention. Asia Pac J Ophthalmol (Phila) 2016;5:403–10. doi: 10.1097/APO.0000000000000233. [DOI] [PubMed] [Google Scholar]
- 31.Williams KM, Verhoeven VJ, Cumberland P, Bertelsen G, Wolfram C, Buitendijk GH, Hofman A, van Duijn CM, Vingerling JR, Kuijpers RW, Hohn R, Mirshahi A, Khawaja AP, Luben RN, Erke MG, von Hanno T, Mahroo O, Hogg R, Gieger C, Cougnard-Gregoire A, Anastasopoulos E, Bron A, Dartigues JF, Korobelnik JF, Creuzot-Garcher C, Topouzis F, Delcourt C, Rahi J, Meitinger T, Fletcher A, Foster PJ, Pfeiffer N, Klaver CC, Hammond CJ. Prevalence of refractive error in Europe: the European Eye Epidemiology (E(3)) Consortium. Eur J Epidemiol. 2015;30:305–15. doi: 10.1007/s10654-015-0010-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Browning SR, Browning BL. Rapid and accurate haplotype phasing and missing-data inference for whole-genome association studies by use of localized haplotype clustering. Am J Hum Genet. 2007;81:1084–97. doi: 10.1086/521987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shi Y, Gong B, Chen L, Zuo X, Liu X, Tam PO, Zhou X, Zhao P, Lu F, Qu J, Sun L, Zhao F, Chen H, Zhang Y, Zhang D, Lin Y, Lin H, Ma S, Cheng J, Yang J, Huang L, Zhang M, Zhang X, Pang CP, Yang Z. A genome-wide meta-analysis identifies two novel loci associated with high myopia in the Han Chinese population. Hum Mol Genet. 2013;22:2325–33. doi: 10.1093/hmg/ddt066. [DOI] [PubMed] [Google Scholar]
- 34.Lin PI, Vance JM, Pericak-Vance MA, Martin ER. No gene is an island: the flip-flop phenomenon. Am J Hum Genet. 2007;80:531–8. doi: 10.1086/512133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rohrer B, Stell WK. Basic fibroblast growth factor (bFGF) and transforming growth factor beta (TGF-beta) act as stop and go signals to modulate postnatal ocular growth in the chick. Exp Eye Res. 1994;58:553–61. doi: 10.1006/exer.1994.1049. [DOI] [PubMed] [Google Scholar]
- 36.McBrien NA. Regulation of scleral metabolism in myopia and the role of transforming growth factor-beta. Exp Eye Res. 2013;114:128–40. doi: 10.1016/j.exer.2013.01.014. [DOI] [PubMed] [Google Scholar]
- 37.Manthey AL, Lachke SA, FitzGerald PG, Mason RW, Scheiblin DA, McDonald JH, Duncan MK. Loss of Sip1 leads to migration defects and retention of ectodermal markers during lens development. Mech Dev. 2014;131:86–110. doi: 10.1016/j.mod.2013.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Menuchin-Lasowski Y, Oren-Giladi P, Xie Q, Ezra-Elia R, Ofri R, Peled-Hajaj S, Farhy C, Higashi Y, Van de Putte T, Kondoh H, Huylebroeck D, Cvekl A, Ashery-Padan R. Sip1 regulates the generation of the inner nuclear layer retinal cell lineages in mammals. Development. 2016;143:2829–41. doi: 10.1242/dev.136101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gregory-Evans CY, Vieira H, Dalton R, Adams GG, Salt A, Gregory-Evans K. Ocular coloboma and high myopia with Hirschsprung disease associated with a novel ZFHX1B missense mutation and trisomy 21. Am J Med Genet A. 2004;131:86–90. doi: 10.1002/ajmg.a.30312. [DOI] [PubMed] [Google Scholar]
- 40.Szklarczyk D, Morris JH, Cook H, Kuhn M, Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, Jensen LJ, von Mering C. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017;45(D1):D362–8. doi: 10.1093/nar/gkw937. [DOI] [PMC free article] [PubMed] [Google Scholar]