

Abstract
Medaka Oryzias latipes is a well-recognized biomedical fish model because of advantageous features such as small body size, transparency of embryos, and established techniques for gene knockout and modification. The goal of this study was to evaluate two critical factors, cryoprotectant and cooling rate, for sperm cryopreservation in 0.25-ml French straws. The objectives were to: 1) evaluate the acute toxicity of methanol, 2-methoxyethanol (ME), dimethyl sulfoxide (Me2SO), N, N- dimethylacetamide (DMA), N, N,-dimethyl formamide (DMF), and glycerol with concentrations of 5, 10, and 15% for 60 min of incubation at 4 °C; 2) evaluate cooling rates from 5 to 25 °C/min for freezing and their interaction with cryoprotectants, and 3) test fertility of thawed sperm cryopreserved with selected cryoprotectants and associated cooling rates. Evaluation of cryoprotectant toxicity showed that methanol and ME (5 and 10%) did not change the sperm motility after 30 min; Me2SO, DMA, and DMF (10 and 15%) and glycerol (5, 10 and 15%) significantly decreased the motility of sperm within 1 min after mixing. Based on these results, methanol and ME were selected as cryoprotectants (10%) to evaluate with different cooling rates (from 5 °C/min to 25 °C/min) and were compared to Me2SO and DMF (10%) (based on their use as cryoprotectants in previous publications). Post-thaw motility was affected by cryoprotectant, cooling rate, and their interaction (P ≤ 0.000). The highest post-thaw motility (50 ± 10%) was observed at a cooling rate of 10 °C/min with methanol as cryoprotectant. Comparable post-thaw motility (37 ± 12%) was obtained at a cooling rate of 15 °C/min with ME as cryoprotectant. With DMF, post-thaw motility at all cooling rates was ≤ 10% which was significantly lower than that of methanol and ME. With Me2SO, post-thaw motilities were less than 1% at all cooling rates, and significantly lower compared to the other three cryoprotectants (P ≤ 0.000). When sperm from individual males were cryopreserved with 10% methanol at a cooling rate of 10 °C/min and 10% ME with a rate of 15 °C/min, no difference was found in post-thaw motility. Fertility testing of thawed sperm cryopreserved with 10% methanol at a rate of 10 °C/min showed average hatching of 70 ± 30% which was comparable to that of fresh sperm (86 ± 15%). Overall, this study established a baseline for high-throughput sperm cryopreservation of medaka provides an outline for protocol standardization and use of automated processing equipment in the future.
Keywords: Medaka Oryzias latipes, Sperm cryopreservation, Cryoprotectant, Cooling rate
Introduction
The application of small-bodied fish species including medaka Oryzias latipes as animal models in biomedical and environmental research has increased dramatically over the past two decades. Medaka has an extensive list of attributes including ease of laboratory culture, short generation time (4-6 weeks), prolific reproduction, transparent embryos, wide tolerance of temperature and salinities, low basal rates of tumor formation, and responsiveness to a broad range of toxicant exposures that have contributed to the widespread use of this species [8,11,23,24]. Whereas various aspects of medaka biology, including physiology, genetics, and embryology have been studied for more than a century [25], recent advances in genomics and genetics research have intensified interest in the use of medaka as a comparative animal model. The recent completion of the draft medaka genome sequence and subsequent releases of additional genome coverage [13] have contributed to the placement of medaka among the leading model organisms for genomics and developmental genetics research.
Advances in genetic modification procedures, from the first transgenic medaka [20] to the most recent disruption (or knock-out) of specific gene function [21] have further increased the attractiveness of this species. In particular, procedures such as tilling (Targeted Induced Lesions In Genomes), in which random mutations are chemically introduced in the male genome followed by breeding with wild-type females to establish a library of genomic DNA and sperm from the F1 generation male offspring hold significant promise in the use of medaka as an animal model [21]. These gene-targeted mutants are retrieved using in vitro fertilization with cryopreserved sperm, and the lineages are established by subsequent out crossing. The generation of gene knockout models by target-selected mutagenesis represents a significant advance to develop new lineages of mutant medaka to explore mechanisms of gene function and related disease processes. The continued expansion of genetic resources including specialized lineages such as targeted gene knockouts and transgenics will contribute thousands of new lineages over the next decades. Coupled with the needs to preserve diminishing native populations of medaka, the demands for increased use of this species highlight the need to improve standardization, efficiency, and high-throughput capabilities for cryopreservation of medaka stocks.
To date, sperm cryopreservation of medaka has been reported in two publications [1,15], and a recently published book chapter [14] based on the combination of these two publications. The first publication reported fetal bovine serum as an extender and 10% dimethyl sulfoxide (Me2SO) and dimethylformamide (DMF) as cryoprotectants [1]; higher fertilization was reported with DMF (96-100%) than that with Me2SO (60%). The second publication reported 0.6 M sucrose as extender, 10% Me2SO as cryoprotectant, and fertilization of 85% [15]. Whereas these protocols are currently used [14], there remains a need to evaluate critical factors such as cryoprotectant, cooling rate, and male characteristics to improve procedures with an emphasis on increased efficiency, standardization, and potential for high-throughput applications. Also, the packaging used in these reports was plastic cryovials or glass capillary tubes. These containers lack capability for automated processing, which is desirable for establishment of large sperm repositories, and present inefficiencies in labeling and storage. In addition, the cooling of samples was not quantified due to the nature of the methods used (i. e. suspension over dry ice, liquid nitrogen or liquid nitrogen vapor), and thus are difficult to repeat. Therefore, a standardized protocol that is readily reproducible among different laboratories is required for this species with assessment of the critical factors in cryopreservation using methods suitable for adoption in high-throughput approaches.
Generally, cryopreservation involves a series of steps including sample collection, sperm extension (dilution), cryoprotectant selection, cooling, cryogenic storage, thawing, and viability assessment [16]. Development of protocols for cryopreservation requires suitable choices at each step and consideration of the interactions among the factors. Inefficiencies at each step can lead to considerable (cumulative) loss of viability in thawed sperm [16]. This study was intended to develop a protocol for medaka sperm cryopreservation based on evaluation of cryoprotectant and cooling rate. The objectives were to: 1) evaluate the acute toxicity of methanol, 2-methoxyethanol (ME), Me2SO, N, N- dimethylacetamide (DMA), DMF, and glycerol with concentrations of 5, 10, and 15% for 60 min of incubation at 4 °C; 2) evaluate cooling rates from 5 to 25 °C/min for freezing and their interaction with cryoprotectants, and 3) test fertility of thawed sperm cryopreserved with selected cryoprotectants and associated cooling rates. In this study, commercially available 0.25-ml French straws were used for packaging of samples, enabling permanent labeling and reliable sealing; Hanks’ balanced salt solution (HBSS, without protein addition) was used as extender, and a programmable freezer was used for precise control of cooling rates.
Materials and Methods
Fish care
Medaka (obtained from Carolina Biological, derived originally from a strain native to Southern Japan), 6-8 months old, were obtained from the Aquatic Biotechnology and Environmental Laboratory of the University of Georgia (Athens, GA) by overnight shipping to Baton Rouge, LA. At the University of Georgia, the fish were cultured in a freshwater (30 mOsmol/kg) system with a photoperiod of 16 h light: 8 h dark at 26 °C with routine disease screening, and were fed 2-3 times daily with live Artemia larvae hatched from cysts (Brine Shrimp Direct, Ogden, UT) and commercial flakes (Zeigler Aquatox Food or Otohime B1). Upon arrival, the fish were maintained in 5-L tanks with 2-3 fish/L in a freshwater aquarium system (Aquatic Habitats™, Aquatic Eco-systems, Inc. Apopka, FL), and were fed twice daily with commercial flakes (Aquatic Eco-system, Apopka, FL) and live hatched Artemia larvae (INVE group; Grantsville, UT). The photoperiod was maintained at 14 h light: 10 h dark at 26 °C. The water quality for the systems at both locations was: dissolved oxygen > 6.5 mg/l; total ammonia-N < 3.0 mg/l, nitrite-N <0.1 mg/l, and hardness (as Ca2CO3) > 100 mg/l. Guidelines from the Institutional Animal Care and Use Committees of Louisiana State University Agricultural Center and University of Georgia were followed for animal care in this study.
Sperm collection and processing
Male fishes were anesthetized with tricaine methanesulfonate (MS-222, 0.05%, Western Chemical Inc., Ferndale, WA) or ice slurry for 1 min (to avoid MS-222 effects on sperm quality), and blotted with a paper towel to dry the body. Before dissection, standard length (from snout tip to the base of the caudal fin) and body weight were measured, and fish were killed by severing of the spinal cord. The testes were removed from surrounding tissues while viewing with a dissecting microscope (10-× magnification) and were transferred to tared 1.5-ml centrifuge tubes for weighing. Hanks’ balanced salt solution (HBSS) with an osmolality of 350 mOsmol/kg (HBSS350) was used as extender in this study. The HBSS350 was prepared by adjusting the water volume using the recipe for HBSS300 (0.137 M NaCl, 5.4 mM KCl, 1.3 mM CaCl2, 1.0 mM MgSO4, 0.25 mM Na2HPO4, 0.44 mM KH2PO4, 4.2 mM NaHCO3, and 5.5 mM glucose, pH = 7.2), and the final osmolality was measured by a vapor pressure osmometer (model 5520, Wescor Inc., Logan, Utah). Sperm suspensions were collected by crushing testes in HBSS350 with a volume (μl) of approximately 40 times the testis weight (mg) yielding approximately 4-8 × 107 cells/ml (estimated from our preliminary data from ten males for sperm concentration determination) except the samples in Experiment III in which the sperm concentrations were determined by use of computer-assisted sperm analysis (CASA) (Hamilton Thorne, Inc., Beverly, MA; CEROS model) while estimating the sperm motility (described below).
Motility estimation
To estimate motility in Experiments I and II, 2 μl of sperm suspension were placed on a glass slide, and 18 μl of HBSS with an osmolality of 100 mOsmol/kg were added to yield a final osmolality of 125 mOsmol/kg which activated medaka sperm motility [26]. The motility was observed at 200-x magnification using a dark-field microscope (Optiphot 2, Nikon Inc., Garden City, NY, USA). The motility was expressed as the percentage of sperm that moved actively in a forward direction; sperm that vibrated in place were not counted as motile. For each sample, sperm motility was estimated at least two times with three fields observed each time. For the sperm samples used for in vitro fertilization in this study (Experiment III), the motility was also evaluated by use of CASA with 20-μm 2-cell slides (Hamilton Thorne, Inc.). The parameter settings of CASA were: minimum contrast, 30; minimum cell size, 4; number of frames for recording, 60; average path (VAP) cut off, 15 μ/s; straight line (VSL) cut off, 10 μ/s; intensity gate, 0.10-1.68; cell gate, 0.72-2.25; elongation gate, 50-100, and slow cell, static.
Freezing and thawing procedures
Prior to cooling, the sperm samples were mixed with an equal volume of double-strength (of target concentration) cryoprotectant in HBSS350, and held on ice for 15 min of equilibration (defined as the time from addition of cryoprotectant to sperm suspension to the time when samples were at 0 °C during the cooling process). Aliquots (100 μl) of sperm suspension were loaded into each 250-μl French straw (IMV International, Minneapolis, MN, USA) by pipetting. The samples were cooled from 5 to -80 °C at the specified cooling rates in a programmable freezer (Kryo 10 series II; Planer Products, Sunbury-on-Thames, UK), and transferred to liquid nitrogen for storage. Samples were evaluated after a minimum of 7 d storage in liquid nitrogen. Motility was estimated at Aquaculture Research Station of the Louisiana State University Agricultural Center. Fertility testing by in vitro fertilization was performed at the University of Georgia following transport of samples (coded for blind analysis) in a 10-L shipping dewar (LD-10, Taylor-Wharton, Theodore, AL). Each cryopreserved sperm sample was thawed individually in a water bath at 40 °C for 5 s, wiped dry with a paper towel, and was released into a sterile 1.5-ml centrifuge tube by cutting of the end (with the cotton plug).
In vitro fertilization
In vitro fertilization was performed at the University of Georgia. Females were separated from males on the preceding day. Within the first 4 h following onset of light, females were killed with an overdose of 0.1% MS-222, dissected, and the ovary removed and placed in a dish with balanced salt solution for medaka (BSS: NaCl, 111.23 mM; KCl, 5.37mM; CaCl2, 1.80 mM; MgSO4·7H2O, 0.81mM; NaHCO3, 0.10 mM, pH = 7.4) with an osmolality of 220 mOsmol/kg [12] measured by osmometer. While viewing with a dissecting microscope, mature eggs were identified and removed from the ovarian tissues by use of fine forceps, and single eggs were separated by cutting their attachment filaments near the surface of the chorion of each egg by use of small scissors. The eggs were held in BSS prior to fertilization for as long as 75 min. To perform in vitro fertilization, individual cryopreserved sperm samples were thawed and added to the eggs within 10 s. BSS (900 μl) was added within 1 min to the egg-sperm mixture (990 µl total solution volume). The fertilized eggs were incubated for 5 min at 26 °C to allow the chorion to become rigid enough for handling. The fertilized eggs were transferred to egg-rearing solution (NaCl, 102.67 mM; KCl, 0.40 mM; CaCl2, 0.27 mM; MgSO4, 0.65 mM) in a Petri dish for incubation at 26 °C for approximately 10-14 d until hatching.
Pooled eggs from 2-4 females were divided into two groups to provide matched samples for direct comparison of egg quality for each cryopreserved sample. One group was fertilized with fresh sperm as a control for egg quality, and the other group was fertilized with thawed sperm for fertility evaluation of cryopreserved samples. Fresh sperm were prepared by crushing of dissected testis in HBSS350 in an 80 times volume (μl) of the testis weight (mg) yielding an concentration of 2-4 × 107 cells/ml. Fertility was recorded as the percentage of fertilized eggs observed at 6 hr (early blastula stage) out of the total number of eggs, development of embryos was monitored daily, and hatching was recorded as the number of hatched fingerlings at 14 d after fertilization divided by the total number of eggs.
Experiment I: Toxicity of six cryoprotectants on sperm motility without freezing
Evaluation of the effect of six cryoprotectants (methanol, ME, Me2SO, DMA, DMF, and glycerol) on sperm motility was performed at final concentrations of 5, 10, and 15%. Sperm suspensions were prepared by pooling sperm from the testes of three males. Three replicates were prepared and measured for each concentration and incubation time. Sperm suspensions were mixed with double-strength cryoprotectant in HBSS350 at a final concentration of 2-4 × 107 cells/ml, and motility was monitored at 0, 10, 20, 30, 40, 50, and 60 min on ice.
Experiment II: Effect of cryoprotectants and cooling rates on sperm motility after thawing
Based on the evaluation of cryoprotectants on sperm motility in Experiment I, methanol, and ME (which showed the least reduction of sperm motility), and Me2SO and DMF (used as cryoprotectants in the two previous publications) [1,15] at a final concentration of 10% were chosen for evaluation of the effect of cooling rate on post-thaw sperm motility. The sperm concentration at freezing was 2-4 × 107 cells/ml. In the first trial, sperm were frozen with three cooling rates (5, 19, and 23 °C/min) with each cryoprotectant. In the second trial, tightened interval spacing was chosen for five cooling rates (5, 10, 15, 20, and 25 °C/min) for each cryoprotectant except Me2SO (which was dropped from consideration based on results in the first trial). Again, three replicates were produced with testes from 5 individuals pooled for each replicate.
Experiment III: Fertility testing of cryopreserved sperm with two freezing protocols
Based on the results of Experiment II, two combinations -- methanol (10%) as cryoprotectant with a cooling rate of 10 °C/min, and ME (10%) as cryoprotectant with a cooling rate of 15 °C/min -- were selected to cryopreserve sperm for fertility evaluation. In the first trial, sperm samples from 10 males were individually cryopreserved by each of the two combinations, and the final sperm concentration at freezing was 2-4 × 107 cells/ml. In the second trial, sperm samples from 20 males were individually cryopreserved with methanol (10%) and a cooling rate of 10 °C/min only. After storage for 3 months, frozen samples were thawed for motility and fertility analysis as described above. Egg quality in the fertility test was evaluated by use of fresh sperm (control) collected from males of the same strain as described above.
Data analysis
Data were analyzed using SYSTAT 12 (Systat Software Inc., MI). Treatment effects were tested by using a two-sample T-test, analysis of variance (ANOVA), or repeated-measures ANOVA. The relationships among fish characteristics, sperm motility and fertility after thawing were analyzed by SYSTAT correlations. Gonadosomatic index (GSI) was calculated as the percentage of testis weight divided by the body weight, and condition factor was calculated as the percentage of the body weight (g) divided by the cube of standard body length (cm). Percentage data were arcsine-square-root transformed for normalization before analysis. The significance level was set at P < 0.050.
Results
Biological characteristics
In total, the 74 males used in this study possessed a standard body length of 2.6 ± 0.1 cm (mean ± SD), body weight of 0.315 ± 0.050 g, and testis weight of 2.0 ± 0.6 (mg).
Experiment I. Toxicity of six cryoprotectants to sperm motility without freezing
The motility of fresh sperm before incubation with cryoprotectants was 84 ± 4% (mean ± SD). The capacity for sperm motility activation decreased with increased cryoprotectant concentration (5, 10, and 15%) and incubation time (5 to 60 min) for all six cryoprotectants (methanol, ME, Me2SO, DMA, DMF, and glycerol) (Fig. 1). Overall, the motility of sperm varied significantly depending on the cryoprotectants tested (P = 0.000), and the incubation time (P = 0.000) at all three test concentrations.
Figure 1.
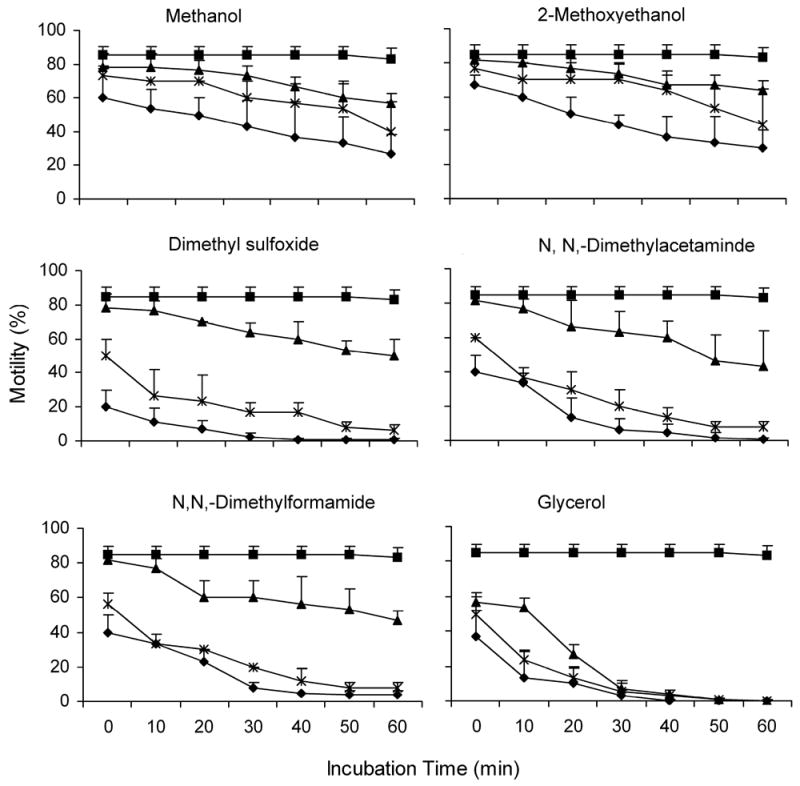
The motility of sperm (mean ± SD, n = 3) of medaka Oryzias latipes after exposure to six cryoprotectants during 60 min at final concentrations of 0% (squares), 5% (triangles), 10% (stars), and 15% (diamonds) in Hanks’ balanced salt solution at an osmolality of 350 mOsmol/kg on ice.
Glycerol reduced motility the most of the six cryoprotectants tested. The ability to activate motility of sperm decreased significantly immediately after mixing in all three final concentrations (P ≤ 0.003). After incubation for 30 min, sperm motility activation decreased to < 10% in all concentrations, and continued to decrease to 0% at 60 min with 5% and 10%, and at 40 min with 15%.
Dimethylsulfoxide, DMA and DMF were of intermediate toxicity. The ability to activate motility of sperm decreased significantly immediately after mixing at final concentrations of 10% and 15% (P ≤ 0.009), but not at 5% (P = 1.000). After 50 min of incubation, motility at the 10% concentrations decreased to < 10%, and after 30 min, motility at the 15% concentrations decreased to < 10%. When a concentration of 5% was evaluated, motility decreased significantly compared to the motility in control sperm (without cryoprotectants) after 30 min with Me2SO (P ≤ 0.043), after 40 min with DMA (P ≤ 0.029), and after 10 min with DMF (P ≤ 0.029).
Methanol and ME were the least toxic of the cryoprotectants tested. The ability to activate motility at all three concentrations did not change after immediate mixing (P ≥ 0.187). When methanol was used, the motility of sperm did not decrease compared with that of the control treatment until after 40 min with the 5% concentration (P ≤ 0.010), 30 min with the 10% concentration (P ≤ 0.012), and 10 min with 15% concentration (P ≤ 0.030). When ME was used, the motility of sperm after mixing was the same as that of the control after 40 min with concentrations of 5% and 10% (P ≥ 0.062), and 10 min with a concentration of 15% (P ≥ 0.073).
Experiment II. Post-thaw motility of sperm cryopreserved with different combinations of cryoprotectants and cooling rates
In the first trial, the cryoprotectants interacted with cooling rate. After cryopreservation with Me2SO, methanol, ME, and DMF (10%) at three cooling rates (5, 19, and 23 °C/min), the motility of sperm after thawing was significantly decreased compared to that of fresh sperm (85%) before freezing (P ≤ 0.000), and the motility decrease was affected by cryoprotectant, cooling rate, and their interaction (ANOVA; P ≤ 0.000) (Fig. 2; upper panel). Analysis of the four cryoprotectants tested in the first trial showed that Me2SO reduced the post-thaw motility significantly compared to the other three cryoprotectants (P ≤ 0.000); the motility at all three cooling rates were all less than 1%, and were not different from one another (P ≥ 0.358). When 10% methanol was used, motility at a rate of 5 °C/min (23 ± 6%) was significantly higher (P ≤ 0.000) than that for 19 °C/min (1 ± 1%) and 23 °C/min (0 ± 1%). When 10% ME was used, motility at 19 °C/min (23 ± 6%) was significantly higher (P ≤ 0.000) than that for 5 °C/min (5 ± 0%) and 23 °C/min (0 ± 0%). The same pattern was observed when DMF was used. A rate of 19 °C/min yielded higher motility (10 ± 0%) than did 5 °C/min (1 ± 1%) (P = 0.001) and 23 °C/min (4 ± 2%) (P = 0.001).
Figure 2.
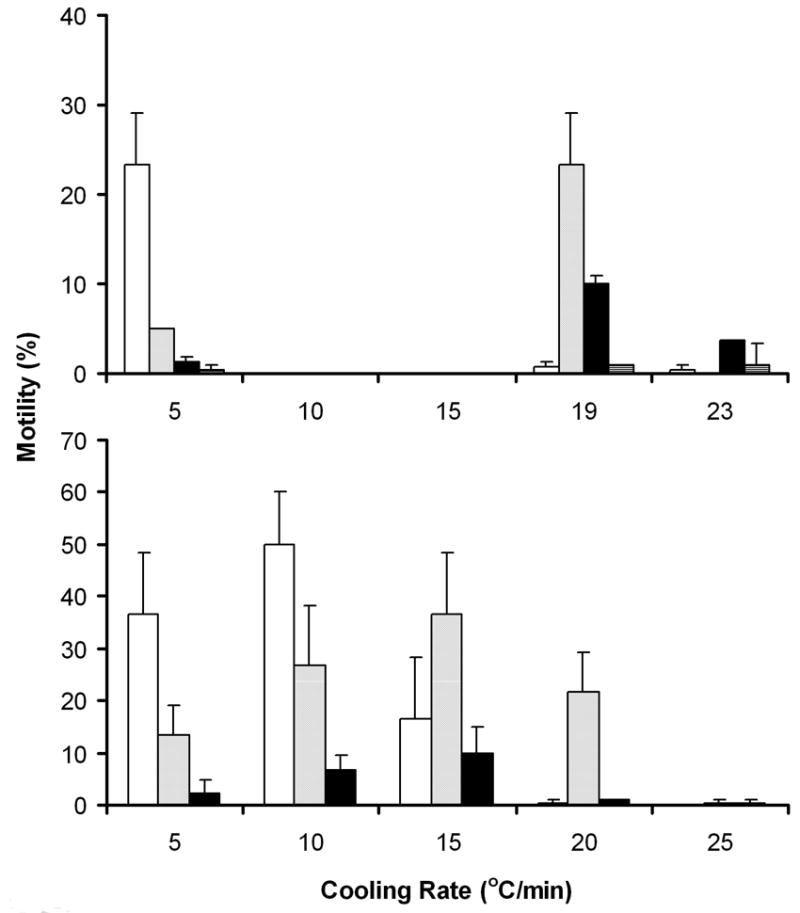
Motility (mean ± SD) of thawed sperm with four cryoprotectants (at 10% concentration) at different cooling rates from 5 to -80 °C in Trial 1(top panel), and Trial 2 (bottom panel): methanol, white; 2-methoxyethanol (ME), light gray; dimethyl sulfoxide (Me2SO), dark gray (Trial 1 only), and N, N-dimethyl formamide (DMF), black. The motility of fresh sperm (before addition of cryoprotectants) was 85 ± 0 % in Trial 1, and 83 ± 3% in Trial 2.
In the second trial, a tighter spacing of cooling rates between 5 and 25 °C/min was used to cryopreserve sperm with methanol, ME, and DMF as cryoprotectants (Me2SO was not included because of the low post-thaw motility in the first trial), and the interaction of cryoprotectants and cooling rate was clearly evident. The post-thaw motility was significantly affected by cryoprotectant (P = 0.000), cooling rate (P = 0.000), and their interaction (P = 0.000) (Fig. 2; lower panel) compared to that of fresh sperm (83 ± 3%) before freezing. When methanol (10%) was used as cryoprotectant, motility at a cooling rate of 10 °C/min (50 ± 10%) was highest (P ≤ 0.003), and motility (37 ± 12%) did not change significantly (P = 0.490) at 5 °C/min. However, with the cooling rate increased to 15 °C/min, motility declined significantly to 17 ± 12% (P = 0.003), and to 0% when rate increased to 20 and 25 °C/min (P ≤ 0.000). When ME was used, motility was not significantly different at rates of 10 °C/min (27 ± 12%), 15 °C/min (37 ± 12%), and 20 °C/min (22 ± 8%) (P ≤ 0.287). But, at rates of 5 °C/min and 25 °C/min, motility was significantly lower (P ≤ 0.027) compared to that at 15 °C/min. When DMF was used, motility at all rates tested was ≤ 10%. The highest motility was observed at rates of 10 °C/min (7 ± 3%) and 15 °C/min (10 ± 5%).
Experiment III. Evaluation of methanol (10%) with a cooling rate of 10°C/min, and ME (10%) with a cooling rate of 15°C/min
In the first trial, no difference was detected (P = 0.546) in the motility of post-thaw sperm cryopreserved with methanol (10%) with a cooling rate of 10 °C/min (50 ± 9%) and ME (10%) with a rate of 15 °C/min (58 ± 11%) (Table 1). Also, no difference was observed in motility before freezing (P = 0.117). Of the fish used for those two treatments, there were no differences in the body characteristics (including standard body length, body weight, and testis weight) (Table 1). However, during the process of in vitro fertilization in this trial, BSS buffer was added ~2 hr after the mixture of post-thaw sperm and eggs (rather than after the usual 1 min). For fresh sperm, this is not a necessary step; but for cryopreserved sperm, it meant that the gametes were exposed to the cryoptotectant (10% methanol or ME) for 2 hr. The development at day 7 and hatch at day 14 from cryopreserved groups were all less than 10%. Based on these and our other studies, we concluded that these data were not representative of the actual fertility of cryopreserved sperm. In addition, we performed a separate study (data not shown) to evaluate the effect of 10% methanol and ME on in vitro fertilization by using fresh sperm. Abnormal embryos were again observed in the eggs fertilized with sperm after ME incubation for 10 min or longer before adding BSS. We suspect that prolonged exposure to this cryoprotectant exerted a teratogenic or mutagenic effect. This observation merits further investigation given reports of mutagenesis for this chemical [5]. This cryoprotectant was eliminated from further study.
Table 1.
Sperm motility (mean ± SD) of medaka Oryzias latipes (n = 10) before freezing and after thawing with two freezing treatments: methanol (10%) with a cooling rate of 10 °C/min (Method 1), and 2-methoxyethanol (10%) with a cooling rate of 15 °C/min (Method 2), and the associated male body characteristics. The P values are for comparisons made within columns (there were no significant differences).
| Motility (%)
|
Body characteristics
|
||||
|---|---|---|---|---|---|
| Fresh | After thawing | Length (cm) | Weight (g) | Testis (mg) | |
| Method 1 | 83 ± 9 | 50 ± 9 | 2.6 ± 0.1 | 0.309 ± 0.035 | 2.1 ± 0.4 |
| Method 2 | 85 ± 7 | 58 ± 11 | 2.7 ± 0.1 | 0.349 ± 0.080 | 2.1 ± 0.4 |
| P values | 0.117 | 0.546 | 0.080 | 0.315 | 0.842 |
In the second fertilization trial, the sperm concentration from each individual varied from 7 ×106 to 1 × 108 cells/ml (Table 2). The initial motility of fresh sperm (before freezing) from 20 males was 45 ± 22% ranging from 5 to 83%, and after thawing, the motility was 21 ± 10% ranging from 1 to 37% (Table 2) (note: the sample from fish number 20 did not include sperm, but was processed for fertilization as part of the single-blind test). There was a significant correlation between initial motility of fresh sperm and the post-thaw motility (R2 = 0.521) (P = 0.017) (Table 3). Thawed sperm showed an average fertilization of 82 ± 26% (from 10 to 100%), and an average hatching of 70 ± 30 (from 0 to 100%) (n = 19; excluding fish number 20). Correlation analysis showed that fertilization and hatch produced by these thawed sperm were significantly related to each other (R2 = 0.664) (P = 0.001) (Table 3). Egg quality evaluation with fresh sperm showed an average fertilization of 97 ± 7% (from 70 to 100%), and an average hatching of 86 ± 15% (from 50 to 100%) (Table 2). For the 19 males used in this experiment, the standard body length (2.6 ± 0.1 cm) showed a significant relationship with body weight (0.3131 ± 0.0547 g) (P = 0.008), and with testis weight (2.1 ± 0.6 mg) (P = 0.026). Also, the testis weight had a significant correlation with post-thaw motility (R2 = 0.494) (P = 0.029) (Table 3).
Table 2.
Fertility evaluation (in Trial 2) of cryopreserved sperm of medaka Oryzias latipes by using methanol (10%) as cryoprotectant combined with a cooling rate of 10 °C/min from 5°C to -80 °C. The characteristics reported are standard length (cm), body weight (g), and testis weight (mg), motility before freezing and after thawing, and fertility of cryopreserved sperm including fertilization (percentage of eggs developed to blastula stage) and hatching (percentage of hatched fry in relation to total number of eggs) of the 20 individual males used. Quality of the eggs was evaluated by using matched batches of eggs for fresh and cryopreserved sperm.
| Fish No. |
Body length (cm) |
Body weight (g) |
Testis weight (mg) |
Sperm density (×107/ml) |
Sperm motility (%)
|
Cryopreserved sperm
|
Egg quality with fresh sperm
|
|||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before freezing |
After thawing |
Eggs used |
Fertilization (%) |
Hatch (%) |
Eggs used |
Fertilization (%) |
Hatch (%) |
|||||
| 1 | 2.6 | 0.3567 | 2.7 | 2.95 | 58 | 37 | 12 | 92 | 83 | 12 | 100 | 92 |
| 2 | 2.5 | 0.2857 | 1.5 | 6.92 | 36 | 12 | 11 | 100 | 100 | 10 | 100 | 100 |
| 3 | 2.7 | 0.3249 | 3.4 | 3.78 | 58 | 25 | 12 | 100 | 100 | 11 | 100 | 100 |
| 4 | 2.5 | 0.2768 | 1.0 | 0.70 | 26 | 5 | 11 | 18 | 18 | 10 | 100 | 100 |
| 5 | 2.4 | 0.2716 | 1.9 | 2.24 | 32 | 12 | 13 | 85 | 92 | 12 | 92 | 67 |
| 6 | 2.7 | 0.4041 | 2.4 | 10.25 | 83 | 41 | 10 | 100 | 100 | 10 | 100 | 50 |
| 7 | 2.6 | 0.3646 | 1.6 | 10.16 | 83 | 20 | 10 | 100 | 70 | 10 | 100 | 80 |
| 8 | 2.4 | 0.2890 | 2.1 | 2.13 | 74 | 26 | 10 | 90 | 40 | 10 | 100 | 100 |
| 9 | 2.4 | 0.3097 | 2.0 | 1.81 | 16 | 11 | 10 | 90 | 90 | 10 | 100 | 90 |
| 10 | 2.5 | 0.3061 | 2.0 | 3.28 | 29 | 18 | 10 | 100 | 100 | 10 | 100 | 80 |
| 11 | 2.6 | 0.2540 | 2.2 | 0.96 | 66 | 23 | 10 | 100 | 80 | 10 | 90 | 90 |
| 12 | 2.6 | 0.3177 | 2.2 | 1.41 | 45 | 21 | 10 | 80 | 50 | 10 | 100 | 70 |
| 13 | 2.8 | 0.4026 | 2.5 | 2.95 | 30 | 28 | 10 | 90 | 90 | 10 | 100 | 100 |
| 14 | 2.6 | 0.2900 | 2.0 | 1.81 | 35 | 19 | 10 | 80 | 60 | 10 | 100 | 100 |
| 15 | 2.5 | 0.2384 | 2.2 | 6.40 | 40 | 22 | 10 | 100 | 80 | 10 | 100 | 90 |
| 16 | 2.9 | 0.4318 | 3.1 | 2.28 | 48 | 22 | 10 | 80 | 80 | 10 | 70 | 70 |
| 17 | 2.3 | 0.2388 | 1.2 | 0.72 | 5 | 1 | 10 | 10 | 0 | 10 | 90 | 80 |
| 18 | 2.6 | 0.2714 | 2.1 | 2.78 | 29 | 22 | 10 | 80 | 30 | 10 | 100 | 70 |
| 19 | 2.7 | 0.2970 | 2.6 | 3.60 | 56 | 26 | 10 | 70 | 70 | 10 | 100 | 100 |
| 20 | 2.7 | 0.3302 | 1.2 | * | * | * | 10 | 0 | 0 | 10 | 100 | 100 |
| Mean | 2.6 | 0.3122 | 2.1 | 3.53 | 45 | 21 | 10 | 82 | 70 | 10 | 97 | 86 |
| SD | 0.1 | 0.0561 | 0.6 | 2.87 | 22 | 10 | 1 | 26 | 30 | 1 | 7 | 15 |
Fish number 20 not included in calculation of these means and SD because there were no sperm included in this sample.
Table 3.
The Pearson correlation (R2 value) and significant Bonferroni P values (within parentheses) among the standard body length (cm), body weight (g), condition factor, testis weight (mg), gonadosomatic index (GSI), pre-freeze motility (before adding methanol) (%), post-thaw motility (%), fertilization (%) and hatch (%) from thawed sperm of 19 males of medaka Oryzias latipes. Sperm from individual males were cryopreserved with 10% methanol at a cooling rate of 10 °C/min.
| Length | Body weight | Condition factor | Testis weight | GSI | Pre-freeze motility | Post-thaw motility | Fertility | Hatch | |
|---|---|---|---|---|---|---|---|---|---|
| Length | 1 | ||||||||
| Body weight | 0.561 (0.008) | 1 | |||||||
| Condition factor | 0.135 | 0.114 | 1 | ||||||
| Testis weight | 0.498 (0.026) | 0.276 | 0.056 | 1 | |||||
| GSI | 0.077 | 0.017 | 0.294 | 0.588 (0.005) | 1 | ||||
| Pre-freeze motility | 0.162 | 0.166 | 0.000 | 0.158 | 0.049 | 1 | |||
| Post-thaw motility | 0.373 | 0.325 | 0.003 | 0.494 (0.029) | 0.197 | 0.521 (0.017) | 1 | ||
| Fertility | 0.105 | 0.102 | 0.000 | 0.282 | 0.212 | 0.286 | 0.384 | 1 | |
| Hatch | 0.141 | 0.184 | 0.008 | 0.281 | 0.116 | 0.097 | 0.213 | 0.664 (0.001) | 1 |
Discussion
In nature medaka are typically found in rice fields, streams, and various freshwater habitats in East Asia. However, because this species can thrive in brackish or even full-strength sea water, it is considered to be a euryhaline fish [10]. As a consequence of tolerance to a wide range of salinity, medaka sperm possess a mode of motility activation that markedly differs from that of typical freshwater fishes such as zebrafish [27]. Sperm motility can be initiated by distilled water without electrolytes (25 mOsmol/kg) and by HBSS with an osmolality spanning from 92 to 686 mOsmol/kg, a range including hypotonic, isotonic, and hypertonic osmolalities [26]. Upon activation, the sperm remain continuously motile for as long as one week. This combination of sperm characteristics is unique in the aquatic species literature, yet may be representative of euryhaline fishes [26] which have not been well studied in this area. To develop protocols for sperm cryopreservation and subsequent artificial insemination, those sperm characteristics need to be considered and identified. In the present study, HBSS350 was used as the extender for sample collection because previous research has shown the sperm retains motility for as long as 7 d during refrigerated storage [26]. For in vitro fertilization, BSS (osmolality of 220 mOsmol/kg) was added at a volume of 9 times the thawed sperm volume to lower the osmolality of thawed sperm from 350 mOsmol/kg (10% methanol does not contribute to osmolality) to around 230 mOsmol/kg to allow fertilization [26], while the addition of BSS reduced the concentration of methanol (to ~1%) and related toxicity.
Toxicity of cryoprotectants to medaka sperm
Cryoprotectants are chemicals that can interact with water (polar molecules) to inhibit ice crystal formation and solution effect during freezing and thawing, and cryoprotectant addition has been widely used in most protocols for sperm cryopreservation [6,18]. Generally, cryoprotectants are classified as: permeating (such as Me2SO, glycerol, and methanol), and non-permeating (such as sucrose, glucose, and proteins) [17]. High water solubility and low toxicity of cryoprotectants are the most important factors to consider for use in cryopreservation. Cryoprotectants can suppress cell injuries when used at appropriate concentrations. However, some cryoprotectants such as Me2SO and glycerol are known to be toxic to various cell types. The effects of protection and toxicity of these chemicals need to be balanced for cryoprotectant selection. Basically, the toxicity of cryoprotectants used in sperm cryopreservation has been found to vary in different species [6]. For example, among other biomedical model fish species, glycerol was used successfully in sperm cryopreservation in Xiphophorus fishes [9,28,29], but was considered to be too toxic for use in zebrafish [27].
In this study, the six cryoprotectants tested showed different effects. Methanol and ME were the least toxic, and the other four, including Me2SO and DMF, showed toxicity immediately after mixing with sperm at concentrations of 10 and 15%. However, Me2SO and DMF were previously used as cryoprotectants for sperm cryopreservation in medaka with a reported fertility of 85-99% [1,15]. To compare our study with these previous publications, Me2SO and DMF were tested and found not to be suitable for medaka sperm in terms of post-thaw motility under the conditions we used. Meanwhile, in agreement with the results of the toxicity estimation, methanol and ME showed better protection during freezing. Despite our efforts to test previously reported cryoprotectants, it is difficult to directly compare the results from different publications because many factors not controlled or reported are involved in the process of cryopreservation that can affect the results (Table 4). It is necessary to identify and quantify these factors when developing standardized cryopreservation protocols, and it is also necessary to report detailed information about these factors. Medaka in this case serves as an illustrative example. It is virtually impossible to make justifiable direct comparisons of the results in this report with the other two previous reports due to a lack of standardization (e.g. containers, freezing methods) and the absence of specific relevant information regarding methods (Table 4). This lack of identification, quantification and reporting are general problems characterizing the cryopreservation literature for all aquatic species [22].
Table 4.
Major factors (under-appreciated for aquatic species in general) that need to be defined, controlled, and reported to enable direct comparisons of sperm cryopreservation results for protocol standardization and application.
| Step or process | Factor to be defined | Relevance to cryopreservation |
|---|---|---|
| Source, housing, and conditioning of fish | Strain and source | Variation among populations or mutant lines can influence results |
| Size and age | Can affect gamete quality and quantity: report body weight, length, age | |
| Maturity | Reproductive condition: report sperm volume, or testis mass, and GSI* | |
| Fish culture | Water quality parameters, temperature, salinity, and light:dark cycles | |
| Male selection | Using best males may not represent normal variation: report selection criteria | |
|
| ||
| Sample preparation and freezing process | Initial quality | Major influence on post-thaw quality: requires assessment and definition |
| Sperm density | Can affect cryopreservation and fertilization: a major uncontrolled variable unless set | |
| Cryoprotectant | Type and final concentration is a critical factor: should be defined and reported | |
| Motility | Sperm motility prior to addition of cryoprotectant: should be defined and reported | |
| Equilibration time | Requires tight control of methods and temperature: should be defined and reported | |
| Packaging | Affects heat transfer: type, size, and materials should be defined and reported | |
| Cooling rate | Critical factor: method should be defined with start and finish temperatures | |
| Storage time | Duration can differentiate freezing and super-cooling: should be defined and reported | |
|
| ||
| Egg collection and use of thawed sperm samples | Pooling of eggs | Should be reported if used to provide sufficient numbers for experiments |
| Thawing process | Warming temperature and duration (rate): should be defined and reported | |
| Post-thaw motility | Necessary to estimate effects of cryopreservation: should be defined and reported | |
| Fertilization | Can influence gamete activation and fertilization especially for thawed sperm | |
| Sperm-to-egg ratio | Concentration and volume of sperm for fertilization: should be defined and reported | |
| Egg quality | Should report if fresh sperm was used to evaluate fertilization of eggs | |
| Fertilization rate | Reporting of exact definition of fertility criteria should be compulsory | |
| Hatching rate | Reporting of absolute or relative values should be identified (report both) | |
GSI, gonadosomatic index, the percentage of testis weight in relation to the body weight.
Cooling rate is a sensitive factor in medaka sperm cryopreservation
Cooling rate is a critical factor in process of cryopreservation. When the cooling rate is too slow, cells lose water to remain in osmotic equilibrium, and face dehydration and prolonged exposure to concentrated solutions; whereas when the cooling rate is too fast, cells do not have sufficient time for water to leave, and intracellular ice crystals can be initiated [18,19]. The consequences from each situation are damaging or lethal to the cells. A balanced situation that allows cell survival is to identify a cooling rate that is rapid enough to minimize the time of exposure to concentrated solutions, yet slow enough to minimize intracellular ice crystal formation [17,19]. This suitable cooling rate can be determined empirically or can be estimated by techniques such as differential scanning calorimetry [3]. In this study, the cooling rate and cell survival displayed an inverted “U” shaped relationship, and suitable rates were related to the choice of cryoprotectants, which is true in most species tested [19].
Another interesting finding in this experiment was the sensitivity of cooling rate especially when methanol was used as cryoprotectant: a 5°C change in rate (from 10 to 15 °C/min) resulted in a significant reduction of post-thaw motility (from 50 ± 10% to 17 ± 12%). In zebrafish, a 10 °C difference from cooling rate (from 10 to 20 °C/min) also yielded a significant drop of post-thaw sperm motility (from 35 ± 23% to 1 ± 1%) with methanol as cryoprotectant [27]. However, in Xiphophorus helleri, the cooling rate yielding highest post-thaw motility fell across a wide range from 20-35 °C/min with glycerol as cryoprotectant [9]. Overall, it is critical to control cooling rate to produce dependable results in sperm cryopreservation, and as indicated in this study, cooling rate needs to be strictly controlled for medaka. In the previous two publications, cooling was produced by placing the samples on dry ice with no quantified data reported. The mechanism that explains the sensitivity to cooling rate observed in the present study remains unknown, but medaka sperm would seem to provide a useful model to explore this. It should be noted that responses to nominal cryoprotectant concentrations and cooling rates can be significantly affected by the sperm cell concentration, and thus attention should be given to adjusting and reporting the cell concentration [4].
Factors that influence the fertility estimation of cryopreserved sperm
Reliable high fertilization and hatching success are the primary aims for cryopreserved sperm. As such, fertility testing is a necessary step for validating the effectiveness of protocols for sperm cryopreservation. Therefore, it is important to present relevant information to evaluate fertility values especially for small fishes such as medaka. Unlike large-bodied fishes that can produce hundreds or thousands of eggs at a time, medaka females produced 10 ± 4 (mean ± SD) (from 1 to 19) mature eggs (data from 40 females used in this study). Thus the egg numbers used for fertility testing in this study were 10-15, and each batch of eggs required pooling from 2-4 females. This low number of eggs used to evaluate fertility success could produce variation in fertility calculations because each individual egg represents 7-10% of the potential fertilization value. Also, in this study, sperm concentration from each male was not strictly standardized, and the 19 males used for fertility testing had varied initial motility (from 5 to 83%), and accordingly the post-thaw motility of these 19 samples varied from 1 to 37%. Inclusion of this variability was done by design to represent the actual wild-type male viability present in stock centers. This is relevant because inbred, transgenic or knockout mutant lines may have reproductive impairments, high variability or low mean sample quality. As such, no pre-selection was made for the males and post-thaw samples used for fertility testing in this study. Overall, the hatch percentage of thawed sperm in this study was comparable to that produced from matched egg batches with fresh sperm (used for egg quality testing). In previous publications [15], the egg numbers used for fertility testing were not reported, and the hatching was calculated as the percentage of hatched fry relative to fertilized embryos (raising the values), not the total number of embryos (The absolute values were not reported), and it was not described if males were selected (by quality) for inclusion in the study. Therefore it is problematic to directly compare the fertility of thawed sperm from this study with that reported in these previous studies.
In this study, the testis weight was observed to significantly correlate with the post-thaw motility, and the testis weight was correlated with body length. This indicated that large fish which have larger testes could provide a preliminary screening factor for sperm cryopreservation. It is true for mammals that a large testis can produce copious sperm with high fertility [7] and this may hold true for medaka as well.
Post-thaw motility can often be a useful index to evaluate the general effects of cryopreservation and to predict the fertility of sperm. However, post-thaw motility was not significantly correlated with fertility (R2 = 0.384; P = 0.166), or hatching (R2 = 0.213; P = 1.000) in this study. This was possibly due to variation in sperm concentration and sperm numbers in each test. For in vitro fertilization, the ratio of eggs and sperm is an important factor that influences fertilization and embryo survival [2]. Therefore, further investigation is needed to study the effect of sperm concentration on cryopreservation and fertilization in medaka. Also, optimized standardization of sperm concentration would assist efficient handling and use of cryopreserved samples especially for valuable strains or lines. The use of sperm collected by non-lethal stripping rather than crushing of testes would yield lower sperm volumes and will place even greater emphasis on the efficient use of sperm.
In summary, this study systematically evaluated in medaka the effect of six cryoprotectants on sperm motility before freezing, the effect of cooling rate and its interaction with cryoprotectants for sperm cryopreservation, and provided a protocol for use of standardized packaging (0.25-ml straw). With the protocol developed in this study, cryopreserved sperm produced results comparable to fresh sperm in terms of fertilization and hatching. The optimized protocol is as follows: 1) dissecting of males to collect testis; 2) crushing of the testes in a volume of 40 times of the testis weight in HBSS with an osmolality of 350 mOsmol/kg (estimated as ~ 4-8 × 107 cells/ml); 3) mixing with an equal volume of 20% methanol in HBSS350; 4) loading of 100 μl of sperm suspension into 250-μl straws; 5) initiation of the freezing program at a cooling rate of 10 °C/min from 5 °C to -80 °C at between 15 and 20 min after mixing with methanol; 6) transfer of samples into liquid nitrogen for storage; 7) thawing at 40 °C for 5 s; 8) addition of thawed sperm onto batches of 10-15 eggs; 9) addition of 900 μl (a volume of 10 times the sperm volume) of BSS buffer (egg culture medium) within 30 sec (total volume of 990 μl); 10) transfer of embryos into fresh BSS buffer after the chorion became rigid (after 5 min), and 11) culture of the embryo at 26 °C and evaluation of fertilization at 6 hr, and hatching at day 14 after fertilization.
This study evaluated the two major factors, cryoprotectant and cooling rate, for sperm cryopreservation of medaka. Because of the limited size of testis from medaka (2.0 ± 0.6 mg, N = 74), it is desirable to maximize the use of cryopreserved sperm samples by identifying suitable sperm concentrations and minimal loading volumes in each straw. Also, standardization of these factors during cryopreservation would assist the efficiency and reliability of processing and use, especially in different laboratories. Based on the foundation developed in this study, future studies should focus on refining the protocol in the following aspects by establishing the: 1) effect of sperm concentration on cryopreservation and fertilization; 2) effect of loading volume in each straw on cryopreservation, fertilization, and repository activities; 3) effect of extender osmolality on cryopreservation, and 4) assessment and control of sperm quality by use of computer-assisted sperm analysis parameters and flow cytometry analysis. Future efforts should also focus on development of database capabilities, rules and agreements for labeling and coding, establishment of standard practices for quality evaluation, mechanisms for sample transport among locations, and biosecurity provisions to address the potential for transfer of pathogens or other contaminants in frozen samples.
Acknowledgments
We thank E Hu, E. Tan, and N. Novelo for technical assistance during the experiments. This work was supported in part by funding from the National Institutes of Health, National Center for Research Resources (R24RR023998), the U.S. Department of Agriculture, and the National and Louisiana Sea Grant College Programs. This manuscript has been approved for publication by the Director of the Louisiana Agricultural Experiment Station as number 2009-244-3897.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Aoki K, Okamoto M, Tatsumi K, Ishikawa Y. Cryopreservation of medaka spermatozoa. Zoo Sci. 1997;14:641–644. [Google Scholar]
- 2.Butts IAE, Trippel EA, Litvak MK. The effect of sperm to egg ratio and gamete contact time on fertilization success in Atlantic cod Gadus morhua L. Aquaculture. 2009;286:89–94. [Google Scholar]
- 3.Devireddy RV, Raha D, Bischof JC. Measurement of water transport during freezing in cell suspensions using a differential scanning calorimeter. Cryobiology. 1998;36:124–155. doi: 10.1006/cryo.1997.2071. [DOI] [PubMed] [Google Scholar]
- 4.Dong Q, Huang C, Tiersch TR. Control of sperm concentration is necessary for standardization of sperm cryopreservation in aquatic species: Evidence from sperm agglutination in oysters. Cryobiology. 2007;54:87–98. doi: 10.1016/j.cryobiol.2006.11.007. [DOI] [PubMed] [Google Scholar]
- 5.Eisses KT. Concurrent teratogenic and mutagenic action of 2-methoxyethanol in Drosophila melanogaster larvae resulted in similar phenotypes: Close resemblance to directed mutations. Teratog Carcinog Mutag. 1999;19:183–204. [PubMed] [Google Scholar]
- 6.Fuller BJ. Cryoprotectants: the essential antifreezes to protect life in the frozen state. Cryoletters. 2004;25:375–388. [PubMed] [Google Scholar]
- 7.Harcourt AH, Harvey PH, Larson SG, Short RV. Testis weight, body-weight and breeding system in primates. Nature. 1981;293:55–57. doi: 10.1038/293055a0. [DOI] [PubMed] [Google Scholar]
- 8.Hawkins WE, Overstreet RM, Walker WW. Small fish models for identifying carcinogens in the aqueous environment. Water Resour Bull. 1988;24:941–949. [Google Scholar]
- 9.Huang C, Dong Q, Walter RB, Tiersch TR. Sperm cryopreservation of green swordtail Xiphophorus helleri, a fish with internal fertilization. Cryobiology. 2004;48:295–308. doi: 10.1016/j.cryobiol.2004.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Inoue K, Takei Y. Diverse adaptability in Oryzias species to high environmental salinity. Zoo Sci. 2002;19:727–734. doi: 10.2108/zsj.19.727. [DOI] [PubMed] [Google Scholar]
- 11.Ishikawa Y. Medaka fish as a model system for vertebrate developmental genetics. Bioessays. 2000;22:487–495. doi: 10.1002/(SICI)1521-1878(200005)22:5<487::AID-BIES11>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 12.Iwamatsu T. A new technique for dechorionation and observations on the development of the naked eggs in Oryzias latipes. J Exp Zool. 1983;228:83–89. [Google Scholar]
- 13.Kasahara M, Naruse K, Sasaki S, Nakatani Y, Qu W, Ahsan B, Yamada T, Nagayasu Y, Doi K, Kasai Y, Jindo T, Kobayashi D, Shimada A, Toyoda A, Kuroki Y, Fujiyama A, Sasaki T, Shimizu A, Asakawa S, Shimizu N, Hashimoto SI, Yang J, Lee Y, Matsushima K, Sugano S, Sakaizumi M, Narita T, Ohishi K, Haga S, Ohta F, Nomoto H, Nogata K, Morishita T, Endo T, Shin-I T, Takeda H, Morishita S, Kohara Y. The medaka draft genome and insights into vertebrate genome evolution. Nature. 2007;447:714–719. doi: 10.1038/nature05846. [DOI] [PubMed] [Google Scholar]
- 14.Kinoshita M, Murate K, Naruse K, Tanaka M. Medaka: Biology, Management, and Experimental Protocols. Wiley-Blackwell; Ames, Iowa: 2009. [Google Scholar]
- 15.Krone A, Wittbrodt J. A simple and reliable protocol for cryopreservation of medaka Oryzias latipes spermatozoa. Fish Biol J Medaka. 1997;9:47–48. [Google Scholar]
- 16.Leibo SP. Sources of variation in cryopreservation. In: Tiersch TR, M MP, editors. Cryopreservation in Aquatic Species. World Aquaculture Society; Baton Rouge: 2000. pp. 75–83. [Google Scholar]
- 17.Leung LKP, Jamieson BGM. Principles of biological cryopreservation. In: Jamieson BGM, editor. Fish Evolution and Systmatics: Evidence from Spermatozoa. Cambridge University Press; Cambridge, UK: 1991. pp. 231–244. [Google Scholar]
- 18.Mazur P. Freezing of living cells - mechanisms and implications. Am J Physiol. 1984;247:C125–C142. doi: 10.1152/ajpcell.1984.247.3.C125. [DOI] [PubMed] [Google Scholar]
- 19.Mazur P. Principal of cryobiology. In: Fuller BJ, Lane N, Benson EE, editors. Life in the Frozen State. CRC Press; New York: 2004. pp. 1–65. [Google Scholar]
- 20.Ozato K, Kondoh H, Inohara H, Iwamatsu T, Wakamatsu Y, Okada TS. Production of transgenic fish - introduction and expression of chicken delta-crystallin gene in medaka embryos. Cell Differentiation. 1986;19:237–244. doi: 10.1016/0045-6039(86)90100-4. [DOI] [PubMed] [Google Scholar]
- 21.Taniguchi Y, Takeda S, Furutani-Seiki M, Kamei Y, Todo T, Sasado T, Deguchi T, Kondoh H, Mudde J, Yamazoe M, Hidaka M, Mitani H, Toyoda A, Sakaki Y, Plasterk RHA, Cuppen E. Generation of medaka gene knockout models by target-selected mutagenesis. Genome Biol. 2006;7:R116. doi: 10.1186/gb-2006-7-12-r116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tiersch TR, Yang H, Jenkins JA, Dong Q. Sperm cryopreservation in fish and shellfish. In: Roldan ERS, Gomendio M, editors. Spermatology (Society of Reproduction and Fertility supplement 65) Nottingham University Press; Nottingham, U.K: 2007. pp. 493–508. [PubMed] [Google Scholar]
- 23.Winn RN. Transgenic fish as models in environmental toxicology. ILAR J. 2001;42:322–328. doi: 10.1093/ilar.42.4.322. [DOI] [PubMed] [Google Scholar]
- 24.Wittbrodt J, Shima A, Schartl M. Medaka - A model organism from the Far East. Nature Reviews Genetics. 2002;3:53–64. doi: 10.1038/nrg704. [DOI] [PubMed] [Google Scholar]
- 25.Yamamoto T. Medaka (killifish): Biology and Strains. Keigaku Publishing Co; Tokyo, Japan: 1975. [Google Scholar]
- 26.Yang H, Tiersch TR. Sperm motility initiation and duration in a euryhaline fish, medaka Oryzias latipes. Theriogenology. 2009;72:386–392. doi: 10.1016/j.theriogenology.2009.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yang H, Carmichael C, Varga ZM, Tiersch TR. Development of a simplified and standardized protocol with potential for high-throughput for sperm cryopreservation in zebrafish Danio rerio. Theriogenology. 2007;68:128–136. doi: 10.1016/j.theriogenology.2007.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yang H, Hazlewood L, Walter RB, Tiersch TR. Sperm cryopreservation of a live-bearing fish, Xiphophorus couchianus: Male-to-male variation in post-thaw motility and production of F-1 hybrid offspring. Comp Biochem Physiol C. 2009;149:233–239. doi: 10.1016/j.cbpc.2008.10.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yang H, Hazlewood L, Heater SJ, Guerrero PA, Walter RB, Tiersch TR. Production of F-1 interspecies hybrid offspring with cryopreserved sperm from a live-bearing fish, the swordtail Xiphophorus helleri. Biol Reprod. 2007;76:401–406. doi: 10.1095/biolreprod.106.056549. [DOI] [PMC free article] [PubMed] [Google Scholar]