

Abstract
This review based on translational research predicts that the transcription factor p53 is the key effector of all anti-acne therapies. All-trans retinoic acid (ATRA) and isotretinoin (13-cis retinoic acid) enhance p53 expression. Tetracyclines and macrolides via inhibiting p450 enzymes attenuate ATRA degradation, thereby increase p53. Benzoyl peroxide and hydrogen peroxide elicit oxidative stress, which upregulates p53. Azelaic acid leads to mitochondrial damage associated with increased release of reactive oxygen species inducing p53. p53 inhibits the expression of androgen receptor and IGF-1 receptor, and induces the expression of IGF binding protein 3. p53 induces FoxO1, FoxO3, p21 and sestrin 1, sestrin 2, and tumour necrosis factor-related apoptosis-inducing ligand (TRAIL), the key inducer of isotretinoin-mediated sebocyte apoptosis explaining isotretinoin’s sebum-suppressive effect. Anti-androgens attenuate the expression of miRNA-125b, a key negative regulator of p53. It can thus be concluded that all anti-acne therapies have a common mode of action, i.e., upregulation of the guardian of the genome p53. Immortalized p53-inactivated sebocyte cultures are unfortunate models for studying acne pathogenesis and treatment.
Keywords: Acne therapy, Apoptosis, Immortalized sebocytes, p53, SV40, TRAIL
Background
Acne vulgaris is the most common inflammatory skin disease affecting more that 80% of adolescents of developed countries [1]. Four major factors are involved in acne pathogenesis: (1) increased sebum production, (2) hyperkolonization and biofilm formation of Propionibacterium acnes (P. acnes), (3) increased acroinfundibular keratinocyte proliferation with comedo formation, (4) and follicular as well as perifollicular inflammation [2]. Sebum is the secretory product of holocrine secretion of sebocytes derived from sebaceous glands (SGs) [3]. Excessive production of sebum containing higher amounts of monounsaturated pro-inflammatory lipids results from exaggerated sebocyte activity, which is induced by increased insulin-like growth factor-1 (IGF-1) and androgen signalling [2]. Recent evidence underlines that dietary factors, especially hyperglycaemic carbohydrates and milk consumption, increase insulin/IGF-1 signalling promoting acne [4–7]. Enhanced activity of the phosphoinositide-3-kinase (PI3K)/AKT pathway down-regulates the nuclear activity of the metabolic transcription factor FoxO1 [8–11], the transcription factor of starvation [12]. Acne is associated with increased activity of mechanistic target of rapamycin complex 1 (mTORC1) [13, 14], which promotes the expression of two lipogenic transcription factors, sterol regulatory element binding protein-1c (SREBP1c) and peroxisome proliferator-activated receptor-γ (PPARγ) [15]. It has been predicted that mTORC1 is activated in the skin of acne patients [16], which has been confirmed experimentally [10, 17]. SREBP1, which is upregulated via increased AKT/mTORC1 signalling plays a key role in sebaceous lipogenesis [18, 19], and in addition induces sebum fatty acid monounsaturation [20, 21], that plays a crucial role in comedogenesis and inflammation of acne [22, 23]. Activated IGF-1/mTORC1 signalling promotes the expression of the anti-apoptotic protein survivin [24, 25], which has recently been found to be upregulated in the skin of acne patients [26]. Intriguingly, serum IGF-1 levels of acne patients significantly correlate with survivin expression [26]. Morphologically, acne vulgaris is characterized by SG hyperplasia with increased production of sebum with higher amounts of pro-inflammatory and comedogenic monounsaturated fatty acids [22]. Increased IGF-1 signalling of puberty superimposed with insulin signalling of Western diet (hyperglycaemic carbohydrates and milk) provide the input signals for disturbed acne metabolomics including mTORC1-S6K1-mediated insulin resistance [22, 27, 28].
It is the intention of this paper to demonstrate that all these acne-related deviations of molecular signalling will be corrected by increasing the expression of the key transcription factor p53, known as the guardian of the human genome [29, 30]. Translational evidence will be presented showing that all common anti-acne therapies used in today’s clinical practice converge in upregulating the expression of p53.
Retinoic acid
All-trans retinoic acid (ATRA), the prototype of topical retinoids, is comedolytic, resolves precursor microcomedones and is anti-inflammatory [31, 32]. Topical ATRA has been shown to transform the horn-filled utriculi of the rhino mouse into normal follicles [33]. ATRA-mediated upregulation of p53 has been reported in several cells including cervical carcinoma cells, acute myeloblastic leukaemia cells, human embryo carcinoma cells, and glioma cells [34–37]. In human embryonic stem cells, ATRA-mediated upregulation of CBP/p300 acetylated p53 at lysine 373, which leads to p53 dissociation from E3-ubiquitin ligases HDM2 and TRIM24, thereby stabilizing p53 expression [38]. It has been demonstrated in primary human epidermal keratinocytes that ATRA regulates many genes associated with cell cycle arrest and programmed cell death [39]. In human keratinocytes, ATRA increases the expression of p53, pro-apoptotic caspases, and sensitizes keratinocytes to apoptosis [40]. Chronic activation of p53 in mice resulted in the loss of SGs associated with a depletion of B-lymphocyte-induced nuclear maturation protein 1 (BLIMP1) positive SG cells explained by suppressed mTORC1 activity [41]. In fact, two p53 target genes, SESN1 and SESN2, activate the AMP-responsive protein kinase (AMPK) and target it to phosphorylate TSC2 and stimulate its GAP activity, thereby inhibiting mTORC1 [42, 43].
p53 intersects at multiple points with the regulation of inflammation [44]. The pro-inflammatory transcription factor nuclear factor κB (NFκB) and p53 can act as functional antagonists. The E3 ubiquitin ligase mouse double minute homolog 2 (MDM2), whose gene is transcriptionally activated by p53, can act as a direct negative regulator of NFκB by binding and inhibiting p65RelA [45]. Furthermore, ATRA-p53-induced neutrophil apoptosis may reduce inflammation in acne [46–48]. ATRA has also been shown to induce apoptosis and suppression of NFκB signalling in adult T cell leukaemia cells [49]. Both, ATRA-p53-induced inhibition of NFκB and neutrophil as well as T cell apoptosis may contribute to the anti-inflammatory effect of ATRA in the treatment of acne.
Peroxides and photodynamic therapy
Benzoyl peroxide (BPO) is an anti-bacterial topical agent that kills P. acnes through the release of free oxygen radicals and is also mildly keratolytic and comedolytic [50–52]. BPO treatment decreased the size of gold hamster ear SGs and the number of sebocytes entering the S-phase of the cell cycle [53]. The mitotic index of BPO-treated sebocytes exhibited a reduction of 33.8% [53]. Similar results have been obtained in autoradiographic studies of human SGs [54, 55]. Although less efficient than ATRA, BPO decreased the size and numbers of corneocytes [56], and reduced comedo formation in the rabbit ear microcomedo prevention assay [57]. HaCaT keratinocytes incubated for 24 h with BPO exhibited a dose-dependent cytotoxicity at concentrations above 250 μm. It is important to mention that BPO is a potent inducer of oxidative stress increasing the intracellular ratio of oxidized to reduced glutathione (GSSG/GSH) in treated keratinocytes [58]. Notably, BPO interacts with mitochondria, inhibits mitochondrial respiration and induces mitochondrial swelling [59]. In a comparable manner, topical hydrogen peroxide (H2O2) treatment reduced the number of inflammatory and non-inflammatory acne lesions [60–62]. It has been demonstrated in C2C12 muscle cells that H2O2 induced mitochondrial permeability transition pore opening and p53 activation. Intriguingly, testosterone treatment prior to H2O2 administration reduced p53 activation and prevented mitochondrial permeability transition pore opening [63]. After mitochondrial damage, p53 maintains the mitochondrial genome through its translocation into mitochondria and interactions with mitochondrial DNA repair proteins. This mechanism provides a further explanation for the upregulation of p53 after mitochondrial insults such as challenges with BPO or H2O2 [64]. Acting as a signal, H2O2 circumvents antioxidant defence by over-oxidizing peroxiredoxins, the enzymes that metabolize peroxides. Sestrins, a family of proteins whose expression is induced by p53, are required for regeneration of peroxiredoxins containing Cys-SO2H, thus re-establishing the antioxidant firewall [65]. Sestrins accumulate in cells exposed to oxidative stress, potentiate AMPK, which finally inhibits mTORC1 [66]. It is well appreciated that oxidative stress and mitochondrial damage-mediated generation of reactive oxygen species (ROS) promote an immediate p53 response [67]. Oxidative stress activates p53 and in turn inhibits cell proliferation and growth through induction of Sestrin 1 and Sestrin 2, which inhibit mTORC1 [67]. Remarkably, metformin, which as well exhibits beneficial effect in the treatment of acne [68], via activation of AMPK and inhibition of mTORC1 [69], operates on the same pathway as AMPK-activating peroxides. In fact, metformin has been shown to increase p53 expression in patients with polycystic ovary syndrome [70].
The major effect of photodynamic therapy (PDT) in acne is the generation of ROS [71, 72]. Thus, PDT mimics the effects of BPO-mediated upregulation of p53. It should thus be expected that retinoid- and BPO-mediated upregulation of p53 may exert synergistic effects in the treatment of acne. In fact, adapalene and BPO significantly decreased the expression of the proliferation marker Ki67, α2 and α6 integrins, TLR-2, β-defensin-4 and IL-8 in inflammatory acne skin, whereas single treatments with adapalene or BPO alone were less effective [73].
Azelaic acid
Azelaic acid (AZA), a saturated C9-dicarboxylic acid, is mildly effective as a comedolytic, anti-bacterial, and anti-inflammatory topical agent for the treatment of acne vulgaris [74, 75]. In cultured keratinocytes, AZA exerted time- and dose-dependent anti-proliferative effects associated with an early marked swelling and damage of mitochondria [76–78]. AZA and other C8-C13 dicarboxylic acids inhibit mitochondrial respiration and promote mitochondrial damage [79]. It has been shown that phosphatidylcholine esterified with AZA induced mitochondrial apoptosis at low micromolar concentrations in various cell lines [80]. Isolated exposed mitochondria rapidly swelled and released cytochrome c and apoptosis-inducing factor [80]. Mitochondrial damage results in mitochondrial ROS production, which upregulates the expression of p53, which promotes mitochondria-mediated apoptosis [81]. In fact, it has recently been confirmed that AZA induces apoptosis in acute myeloid leukaemia cells in a dose-dependent manner [82]. Thus, AZA shares mechanistic similarities with peroxide-induced mitochondrial disturbances upregulating the p53 response.
Tetracyclines and macrolides
Tetracyclines are considered the first-line therapy in moderate to severe acne [83]. Apart from their anti-bacterial activities against P. acnes and bacterial lipases, non-antibiotic properties of tetracyclines gained recent attention [84]. The observation that sub-antimicrobial dosing of doxycycline showed equal efficacy as conventional anti-bacterial doxycycline treatment of inflammatory lesions in moderate and severe acne underlined the importance of tetracyclines’ non-antibiotic effects in acne [85]. Tetracylines, hypervitaminosis A, and systemic isotretinoin treatment share an increased risk for pseudotumor cerebri, which already points to a common underlying pathogenic mode of action [86]. ATRA homeostasis in the adult CNS is tightly controlled through local ATRA synthesis and cytochrome P450 (CYP450)-mediated inactivation of ATRA [87]. In neuronal cells, minocycline increased ATRA levels via inhibiting p450-mediated ATRA degradation [87]. This observation prompted Hellmann-Regen et al. [88] to speculate that tetracyclines and erythromycin may exert their pharmacological mode of action in acne via suppression of p450-mediated ATRA degradation in the skin. In fact, these investigators provided experimental evidence that minocycline potently blocked ATRA degradation in rat skin microsomes, and strikingly enhanced ATRA levels in ATRA-synthesizing cell cultures in a dose-dependent manner [89]. Several studies underline that tetracyclines and macrolides such as erythromycin and azithromycin suppress ATRA-catabolizing p450 enzymes modifying cellular ATRA homeostasis [90–92]. Intracellular upregulation of ATRA is thus the common denominator of p450-inhibiting agents that finally upregulate p53. A link between p450-regulation and mTORC1 signalling has recently been suggested [93]. In fact, it has been demonstrated that minocycline upregulated p53 and inactivated the AKT/mTORC1 pathway [94].
In contrast, it should be expected that p450-inducing agents such as isoniazid, phenobarbital, rifampicin, phenytoin, glucocorticosteroids and others [95–97] may lower intracellular ATRA levels and thus increase the risk for acne. Indeed, the majority of drugs reported to promote acne and acneiform drug eruption are p450-inducing agents [98–100].
Oral isotretinoin
Oral isotretinoin (13-cis retinoic acid), an isomer of ATRA, has been used for the treatment of severe recalcitrant acne for nearly four decades [101]. Its use has proven successful for most patients with severe acne, resulting in decreased sebum production and marked reduction of inflammatory lesions [102–105]. It is important to realize that the sebum-suppressive action of isotretinoin is not related to decreased lipid synthesis of individual sebocytes but is caused by sebocyte death, which histologically corresponds to the involution of SGs during isotretinoin treatment [106–108]. In pioneering histological and planimetrical studies, a marked decrease in the size of SGs of up to 90% of the pre-treatment values has been observed after 12 weeks of treatment. Additionally, the ratio of the differentiating pool of sebocytes versus the undifferentiating cell pool changed from 2:1 to 1:7 [107]. Furthermore, the labelling index of sebocytes regressed significantly under isotretinoin therapy. Today, this dramatic SG involution can be explained by isotretinoin-mediated sebocyte apoptosis (programmed cell death). Sebocytes are able to isomerize 13-cis retinoic acid to all-trans retinoic acid (ATRA), which binds to and activates retinoic acid receptors (RARs) that modify gene expression [109] (Fig. 1). One most important ATRA-responsive gene is the transcription factor p53 [39]. Activated p53 induces the expression of the pro-apoptotic effector TRAIL (tumour necrosis factor-related apoptosis-inducing ligand) [110]. There are two p53 DNA-binding sites in the human TRAIL promoter region [110]. Furthermore, ATRA induces RAR-dependent transcriptional upregulation of the TRAIL receptor 1 (TRAIL-R1, also known as death receptor 4) [111], thus promotes apoptotic TRAIL signalling at the ligand- and receptor level. ATRA also activates the expression of the transcription factor FoxO3a [112, 113]. p53 directly binds and activates the expression of the FOXO3A gene [114, 115]. Many of the genes targeted by p53 were also targeted by FOXO transcription factors, indicating that p53 functions in a coordinate manner to suppress gene expression downstream of PI3K/AKT/mTORC1 signalling [116, 117]. Both ATRA-induced p53 and ATRA-induced FoxO3a synergistically promote TRAIL expression [118]. In isotretinoin treated acne patients, TdT-mediated dUTP-biotin nick end labelling (TUNEL), a marker of apoptotic cells, was strongest in the nuclei of sebocytes in the basal layer and in early differentiated sebocytes adjacent to the basal layer of SGs [119]. In accordance, upregulated TRAIL expression has been observed in the basal and suprabasal layers of SG during isotretinoin treatment of acne patients [120], which allows the conclusion that isotretinoin-ATRA-p53/FoxO3a-induced TRAIL signalling explains isotretinoin-induced sebocyte apoptosis resulting in the involution of SGs (Fig. 1). Kelhälä et al. [106] confirmed increased TRAIL mRNA expression in lesional skin of isotretinoin-treated acne patients. TRAIL-mediated activation of caspase 8 and caspase 3 inactivates p63 [121], a critical marker of seboblasts/progenitor cells located in the outermost layer of SGs [122]. Thus, isotretinoin via increased p53 signalling apparently depletes the number and survival of p63-regulated sebocyte progenitor cells.
Fig. 1.
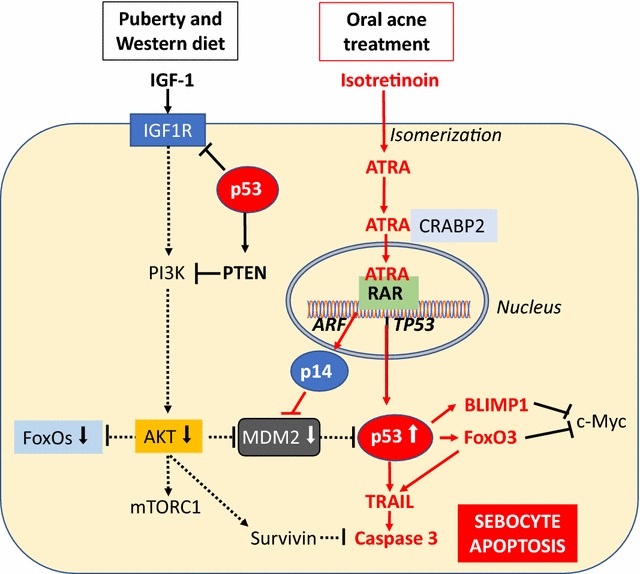
Isotretinoin-induced p53-mediated sebocyte apoptosis. In the sebocyte, isotretinoin is isomerized to all-trans-retinoic acid (ATRA), which is transported to the nucleus via cellular retinoic acid binding protein 2 (CRABP2). In the nucleus, ATRA binds to retinoic acid receptor (RAR) and activates RAR-responsive genes such as TP53, which promotes the expression of p53. ATRA-induced expression of ARF promotes the expression of p14, which is a negative regulator of mouse double minute 2 (MDM2), the key inhibitor of p53 via proteasomal degradation of p53. Increased IGF-1 signalling is attenuated by p53 and reduces the activity of the kinase AKT, that via phosphorylation inhibits the activity of FoxO1 and FoxO3 but stimulates MDM2. Thus, isotretinoin increases p53 activity via its direct transcriptional induction and posttranslational inhibition of its negative regulator MDM2. Subsequently, increased p53 activates several apoptosis-promoting proteins such as tumour necrosis factor-related apoptosis-inducing ligand (TRAIL). p53-attenuated IGF-1 signalling reduces the expression of survivin, a critical inhibitor of caspase 3. p53-induced expression of BLIMP1 and FoxO3 suppresses c-Myc, a key transcription factor of sebocyte differentiation. The final outcome is sebocyte apoptosis, the primary mechanism of isotretinoin-induced sebum suppression
The expression of IGF-1, the most important pro-survival stimulus and mitogen of SGs, was increased in the basal and suprabasal layers of SGs of acne patients [7]. In normal skin, lGF-1 receptor (IGF1R) mRNA expression was most intense in the basal cells of the SG in immature sebocytes. Some weaker staining was present in mature fully differentiated sebocytes [119]. Expression was also detected in all cells of the infundibulum [123]. IGF-1 may thus promote infundibular keratinocyte proliferation (comedogenesis) in acne [124]. The pattern of IGF-1 and IGF1R expression suggests a critical role for IGF-1 as a sebaceous mitogen and morphogen [123]. IGF-1-deficient patients with Laron syndrome do not develop acne and other mTORC1-driven diseases of civilization [124, 125]. The expression pattern of the IGF-1/IGF1R system thus perfectly fits to the hyperproliferative cell layers of SGs and infundibular keratinocytes observed in acne patients [126, 127]. Importantly, p53 has been identified as a negative regulator of the IGF1R gene [128], which mediates increased IGF-1/mTORC1 signalling of puberty and Western diet (Fig. 1) [6, 22, 129]. Recent evidence underlines that the IGF-1 signalling axis and p53 genome protection pathways are tightly interconnected [130]. IGF-1/AKT/mTORC1 signalling also increases the anti-apoptotic regulator survivin [24, 25], which is upregulated in the skin of acne patients [26]. Survivin’s anti-apoptotic effects are mediated via inhibition of caspase 3 [131], which is the downstream effector caspase activated by TRAIL signalling [132]. FoxO3a, which is suppressed via IGF-1/AKT signalling [133], is an inducer of TRAIL expression (Fig. 1) [131]. Thus, p53-mediated inhibition of IGF-1 signalling will reduce survivin expression and its anti-apoptotic action in the pilosebaceous follicle. Furthermore, p53 and p53-mediated FoxO3a signalling increase pro-apoptotic TRAIL signalling.
Isotretinoin treatment of SEB-1 sebocytes induced G1 cell cycle arrest via upregulation of the cell cycle inhibitor p21 [134]. It is known that p53 uses cell cycle checkpoints to induce G1/S and G2/M cell cycle arrest [135, 136]. p21 (WAF1) was among the first p53 target genes that have been identified [137, 138].
mTORC1 signalling, which is increased in SGs of acne patients [10, 17], is negatively regulated by p53 [42, 116]. Deletion of p53 enhances mTORC1 activity by altering lysosomal dynamics of TSC2 and Rheb [139]. mTORC1 orchestrates the expression of SREBP1c and PPARγ [13–15], which play a crucial role in sebaceous lipogenesis, sebocyte differentiation, and sebum production [18, 19, 140–142].
IGF binding protein-3 (IGFBP-3) is a nuclear regulator that binds to retinoid X receptor-α (RXRα) and several of its dimerization partners, including nuclear receptor Nur77 and PPARγ [143, 144]. RXRα-IGFBP3 interaction leads to modulation of the transcriptional activity of RXRα that is essential for mediating the effects of IGFBP3 on apoptosis [145]. In response to IGFBP3, the RXRα binding partner nuclear receptor Nur77 rapidly undergoes translocation from the nucleus to the mitochondria, initiating an apoptotic cascade resulting in caspase activation [146]. IGFBP3 attenuates the activation of PPARγ and inhibits adipocyte differentiation [147]. IGFBP3 interacted with PPARγ and inhibited PPARγ heterodimerization with RXRα [147]. Isotretinoin treatment of SEB-1 sebocytes resulted in a threefold over-expression of IGFBP3 [119]. Notably, IGFBP3 is a target gene of p53 [148]. Thus, p53-mediated induction of IGFBP3 gene expression inhibits mitogenic IGF-1 signalling (Fig. 2).
Fig. 2.
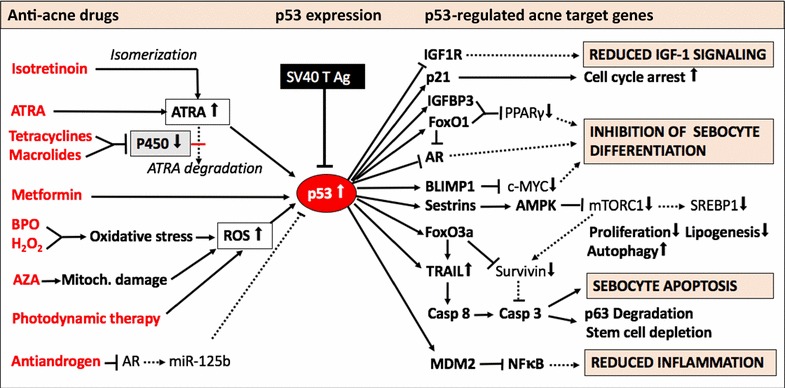
Synoptic illustration of p53-activating anti-acne therapies. Systemic isotretinoin (13-cis retinoic acid) via isomerization to all-trans retinoic acid (ATRA), tretinoin (ATRA), as well as cytochrome p450-inhibiting tetracyclines and macrolides all enhance ATRA-mediated upregulation of p53. Benzoyl peroxide (BPO) and hydrogen peroxide (H2O2) enhance p53 expression as well as a azelaic acid (AZA)-induced mitochondrial damage and photodynamic therapy, which generate reactive oxygen species (ROS). Activated p53 attenuates the expression of IGF-1 receptor (IGF1R) and of androgen receptor (AR). p53 activates expression of cell cycle inhibitor p21 and via upregulation of IGF binding protein-3 (IGFBP3) suppresses the transactivation of peroxisome proliferator-activated receptor-γ (PPARγ), which is important for sebocyte differentiation. Oxidative stress-responsive sestrins activate AMP kinase (AMPK), which inhibits mechanistic target of rapamycin complex 1 (mTORC1) downregulating anabolism, cell growth and sterol regulatory element binding protein 1c (SREBP1c)- and PPARγ-dependent lipogenesis. p53-mediated upregulation of FoxO1 expression inhibits AR, PPARγ, and SREBP1c, key transcription factors of sebaceous lipogenesis and sebocyte differentiation. p53-induced expression of FoxO3a and tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) activate pro-apoptotic signalling with upregulation of caspase 8 (Casp8) and caspase 3 (Casp3), which execute apoptosis and promote p63 degradation. p53 increases the expression of the ubiquitin E3 ligase MDM2, which inhibits nuclear factor κB (NFκB), the key transcription factor for inflammatory cytokine expression. Anti-androgens attenuate AR-mediated expression of miRNA-125b, a key negative regulator of p53. Thus, p53 upregulation balances all pathological deviations observed in the sebaceous follicle of patients with acne vulgaris: increased proliferation, exaggerated lipogenesis, and inflammation. Note, that p53 is suppressed in SV40 immortalized sebocytes, because SV40 large T antigen physically inhibits p53
Taken together, pro-apoptotic isotretinoin-ATRA-p53 signalling induces a complex regulatory network that counteracts exaggerated IGF-1-AKT-mTORC1-mediated pro-survival signalling in acne vulgaris. Whereas isotretinoin-induced p53-TRAIL signalling is the desired effect promoting sebum suppression via sebocyte apoptosis, all adverse effects of the drug including teratogenicity can be explained by p53-mediated apoptosis of vulnerable ATRA-sensitive cells such as neuronal crest cells (Table 1) [149]. Intriguingly, hyper-activated p53 induced neural crest cell apoptosis in mice and craniofacial abnormalities resembling retinoid embryopathy [150, 151].
Table 1.
p53-regulated target genes involved in isotretinoin’s mode of action
| p53 target genes | Desired and adverse drug effects |
|---|---|
| Tumor necrosis factor-related apoptosis-inducing ligand, TRAIL (TNFSF10) upregulation | Sebocyte apoptosis: sebum suppression Meibomian cell apoptosis: dry eyes Neural crest cell apoptosis: teratogenicity Hypothalamic cell apoptosis: depression Intestinal cell apoptosis: inflammatory bowel disease |
| Insulin-like growth factor-1 receptor (IGF1R) suppression | Attenuated pro-survival and mitogenic signaling of IGF-1 |
| Androgen receptor (AR) suppression | Reduced AR expression and miRNA-125b-mediated suppression of p53 |
| IGF binding protein-3 (IGFBP3) upregulation | Enhanced pro-apoptotic signalling and suppressed PPARγ signalling: attenuated lipogenesis |
| Cyclin-dependent kinase inhibitor 1A, p21 (CDKN1A) upregulation | G1/S cell cycle arrest: Suppression of comedogenesis and sebocyte proliferation |
| B lymphocyte-induced maturation protein 1 (BLIMP1) (PRDM1) upregulation | Increased BLIMP1-mediated c-Myc suppression reducing sebocyte differentiation |
| Sestrin 1 (SESN1) and sestrin 2 (SESN2) upregulation | Activation of AMPK resulting in mTORC1 and ACC inhibition: sebum suppression |
| Forkhead box O1 (FOXO1) upregulation | Suppression of AR, SREBP1c and PPARγ: suppression of lipogenesis |
| Forkhead box O3a (FOXO3A) upregulation | Enhanced upregulation of TRAIL: enhancement of apoptosis |
| AMP-activated protein kinase (PRKAA1) | Increased expression of AMPK and AMPK-mediated inhibition of mTORC1 |
| Aquaporin 3 (AQP3) upregulation | Increased aquaporin 3 expression: increased transepidermal water loss, dry skin, xerosis, |
| Aquaporin 4 (AQP4) upregulation | Increased aquaporin 4 expression increasing cerebrospinal fluid (risk of pseudotumor cerebri) |
| Apolipoprotein B100 (APOB) and apoB mRNA editing enzyme complex 1 (APOBEC1) | Increased hepatic synthesis of ApoB100: hypertriglyceridaemia with increased hepatic secretion of triglyceride-rich VLDL |
Anti-androgens
Antiandrogens play an important role in sebum suppression and acne therapy in female patients [152, 153]. Androgen receptor (AR)-mediated signalling contributes to sebocyte differentiation and maximization of sebaceous lipogenesis [154]. In hamster sebocytes, phosphorylation and thus activation of TOR was increased by the addition of testosterone in the presence of IGF-1 [154]. Furthermore, IGF-1 enhances adrenal and gonadal androgen synthesis and via activation of 5α-reductase promotes the conversion of testosterone to its ten times more potent AR-ligand dihydrotestosterone (DHT) [6]. Increased IGF-1 signalling in acne suppresses nuclear FoxO1 [8–10], which is a nuclear co-suppressor of AR [155], and thus increases AR-mediated target gene expression. Recently, p53 has been identified as transcriptional inducer of FOXO1 and PTEN [156], an important observation that confirms the role of p53 in regulating multiple signalling levels of IGF-1/IGF1R/PI3K/AKT/FoxO1 signalling. AR is regarded as a sensitive marker of sebaceous differentiation [157]. Androgens induce sebaceous differentiation in sebocytes expressing a stable functional AR. DHT up-regulated the expression of genes potentially related to sebocyte differentiation such as MUC1/EMA, AQP3, and FADS2 [158]. Remarkably, AR is a direct target of p53 and is negatively regulated by p53 [159, 160]. This allows the conclusion that all p53-activating anti-acne agents attenuate AR signalling and thus exert anti-androgenic activity, which is further suppressed via classical anti-androgens such as cyproterone acetate (CPA).
c-Myc is a further important transcription factor promoting sebocyte differentiation [161, 162]. Interestingly, a functional interaction between c-Myc and p53 has been reported [163]. Expression of c-Myc significantly attenuated apoptosis and impaired the transcriptional activity of p53 on p21 [163]. c-Myc overexpression may antagonize the pro-apoptotic function of p53 [163]. Recent evidence indicates that c-Myc-induced SG differentiation is controlled by an AR/p53 axis [163]. c-Myc-induced SG differentiation was reduced in mice lacking a functional AR. In contrast, testosterone treatment or p53 deletion activated AR signalling and restored c-Myc-induced differentiation [164]. Recent studies have revealed that FoxO3a acts as an antagonist of c-Myc (Fig. 1) [165]. Thus, increased IGF-1-AKT signalling in acne via FoxO3a suppression may favour c-Myc-driven SG differentiation.
Anti-androgens with proven effects in the treamtment of acne are CPA, spironolactone and flutamide [152, 153]. These three major anti-androgens used for acne therapy are AR ligands that antagonize the actions of testosterone and DHT by competing for AR binding sites. Testosterone and DHT-mediated activation of AR induces the expression of miRNA-125b [166, 167]. Importantly, miRNA-125b is a highly conserved key suppressor of p53 [168–170]. The MIR125B2 gene promoter exhibits four AR response elements pointing to close interaction between androgens and miRNA-125b expression [167]. Anti-androgens such as CPA or flutamide reduce AR-mediated expression of miRNA-125b [167], which increases p53 activity [167–170]. Remarkably, p53-dependent expression of the pro-apoptotic proteins TRAIL and death receptor 5 (DR5) increased by CPA treatment [171]. p53 suppresses the expression of AR, thus reduces AR signaling [159, 160]. Indeed, oral isotretinoin, which enhances p53 activity, has been demonstrated to reduce AR levels in the skin of isotretinoin-treated acne patients [172]. In this regard, isotretinoin and anti-androgens converge in p53-induced TRAIL-mediated sebocyte apoptosis and sebum suppression.
Androgen/AR-induced miRNA-125b not only targets p53 but also BLIMP1 [173]. p53 positively regulates BLIMP1 transcription [174]. BLIMP1 is a suppressor of c-Myc [175]. Anti-androgen treatment of acne via attenuation of miRNA-125b may thus increase the inhibitory effect of BLIMP1 on c-Myc thereby inhibiting sebocyte differentiation and sebaceous lipogenesis.
Immortalized sebocytes
A huge number of experimental acne research has been performed with immortalized sebocytes such as the SZ95 or SEB-1 sebocyte cell lines, which are derived from human sebocytes transfected with the SV40 large T antigen [176, 177]. Via transfection of the HPV16-E6/7 oncoproteins, the immortalized human sebocyte cell line SEBO662 has been established [178]. It is believed that immortalized sebocyte culture models provide valuable insights into the development and management of acne [179, 180]. However, immortalized cell lines are not a suitable model to study acne nor the in vivo pharmacological action of anti-acne agents as immortalization abolishes p53 activity [179, 180]. Immortalization by SV40 large T antigen and oncogenic HPV16 proteins inactivates p53, pRB and SEN6 [180, 181].
The large T antigen of simian virus 40 (SV40) forms a specific complex with p53 [182, 183] and inhibits p53-mediated transcription [184] (Fig. 2). During evolutionary viral adaptation to host organisms, viruses have developed strategies to manipulate host cell p53 dependent pathways to facilitate viral survival via inhibition of host cell apoptosis [185]. SV40 T antigen-mediated p53 suppression apparently impairs ATRA-p53-induced sebocyte apoptosis. In fact, isotretinoin (10−8 − 10−5 M) did not affect externalized phosphatidylserine levels, DNA fragmentation, and lactate dehydrogenase cell release, despite increased caspase 3 levels [186]. Only, after addition of a further apoptosis-inducing agent (staurosporine) DNA fragmentation in SZ95 sebocytes was induced [186]. In B16F-10 melanoma cells, isotretinoin alone induced apoptosis associated with upregulated p53 expression [187]. Despite a multitude of studies with immortalized sebocytes, no data on the expression and regulation of p53, the guardian of the genome, have yet been reported.
Conclusion
There is compelling evidence for the key role of p53 in sebocyte homeostasis. It can be concluded from translational evidence that currently available anti-acne agents have a common mode of action: the upregulation of p53 expression. p53 controls a web of critical genes related to acne pathogenesis such as AR, FoxO transcription factors, BLIMP1, and mTORC1 activity, that all play a key role in acne pathogenesis as well as pharmacological actions of anti-acne agents [188]. p53, the guardian of the genome, is a pivotal regulator for cell homeostasis. p53 controls most important cellular responses such as IGF-1 and AR signalling and via induction of MDM2 terminates p53-induced cellular responses via ubiquitination and proteasomal degradation of p53, FoxO1 and FoxO3a, respectively [189–191]. All these essential regulatory mechanisms are compromised in immortalized sebocytes via transfection with SV 40 large T antigen or HPV16-E6/7 oncoproteins. In contrast to the in vivo situation, p53 in acne patients is not artificially inactivated and still responsive to pharmacological targeting. We have to appreciate that acne is a pro-survival disease of the sebaceous follicle with increased IGF-1/AKT/mTORC1-survivin signalling [192]. Anti-acne agents such as retinoids, antibiotics, peroxides, azelaic acid, metformin and anti-androgens induce p53-mediated signalling and thus readjust the delicate p53-dependent balance between survival and death. Immortalized sebocytes with inactivated p53 transcription are thus a most critical and perhaps misleading model system to study p53-driven apoptotic signalling pathway in acne, which have recently excited the field of acne research [193].
Acknowledgements
The author thanks Gerd Plewig, Department of Dermatology, University of Munich; for fruitful discussions of the presented concept and Susan Broy for literature supply.
Competing interests
The author declares that he has no competing interests.
Availability of data and materials
Not applicable.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
There is no funding of this translational research study.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- BLIMP1
B-lymphocyte-induced nuclear maturation protein 1
- AKT
Akt kinase (protein kinase B)
- AMPK
AMP-responsive protein kinase
- AR
androgen receptor
- ATRA
all-trans retinoic acid
- AZA
azelaic acid
- BPO
benzoyl peroxide
- CPA
cyproterone acetate
- DHT
dihydrotestosterone
- FoxO
Forkhead box O
- HPV
human papilloma virus
- IGF-1
insulin-like growth factor-1
- IGFBP-3
IGF binding protein 3
- IGF1R
IGF-1 receptor
- MDM2
mouse double minute homolog 2
- mTORC1
mechanistic target of rapamycin complex 1
- NFκB
nuclear factor κB
- PDT
photodynamic therapy
- PI3K
phosphoinositide-3 kinase
- PPAR
peroxisome proliferator-activated receptor
- RB
retinoblastoma protein
- RAR
retinoic acid receptor
- ROS
reactive oxygen species
- RXR
retinoid receptor
- SG
sebaceous gland
- S6K1
S6 kinase 1
- SREBP
sterol regulatory element binding protein
- SV40
simian virus 40
- TRAIL
tumour necrosis factor-related apoptosis-inducing ligand
- TSC2
tuberin
References
- 1.Lynn DD, Umari T, Dunnick CA, Dellavalle RP. The epidemiology of acne vulgaris in late adolescence. Adolesc Health Med Ther. 2016;7:13–25. doi: 10.2147/AHMT.S55832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Moradi Tuchayi S, Makrantonaki E, Ganceviciene R, Dessinioti C, Feldman SR, Zouboulis CC. Acne vulgaris. Nat Rev Dis Primers. 2015;1:15029. doi: 10.1038/nrdp.2015.29. [DOI] [PubMed] [Google Scholar]
- 3.Fischer H, Fumicz J, Rossiter H, Napirei M, Buchberger M, Tschachler E, et al. Holocrine secretion of sebum is a unique DNase2-dependent mode of programmed cell death. J Invest Dermatol. 2017;137:587–594. doi: 10.1016/j.jid.2016.10.017. [DOI] [PubMed] [Google Scholar]
- 4.Cappel M, Mauger D, Thiboutot D. Correlation between serum levels of insulin-like growth factor 1, dehydroepiandrosterone sulfate, and dihydrotestosterone and acne lesion counts in adult women. Arch Dermatol. 2005;141:333–338. doi: 10.1001/archderm.141.3.333. [DOI] [PubMed] [Google Scholar]
- 5.Vora S, Ovhal A, Jerajani H, Nair N, Chakrabortty A. Correlation of facial sebum to serum insulin-like growth factor-1 in patients with acne. Br J Dermatol. 2008;159:990–991. doi: 10.1111/j.1365-2133.2008.08764.x. [DOI] [PubMed] [Google Scholar]
- 6.Melnik BC, Schmitz G. Role of insulin, insulin-like growth factor-1, hyperglycaemic food and milk consumption in the pathogenesis of acne vulgaris. Exp Dermatol. 2009;18:833–841. doi: 10.1111/j.1600-0625.2009.00924.x. [DOI] [PubMed] [Google Scholar]
- 7.Seleit I, Bakry OA, Abdou AG, Hashim A. Body mass index, selected dietary factors, and acne severity: are they related to in situ expression of insulin- like growth factor-1? Anal Quant Cytopathol Histpathol. 2014;36:267–278. [PubMed] [Google Scholar]
- 8.Melnik BC, Zouboulis CC. Potential role of FoxO1 and mTORC1 in the pathogenesis of Western diet-induced acne. Exp Dermatol. 2013;22:311–315. doi: 10.1111/exd.12142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mirdamadi Y, Thielitz A, Wiede A, Goihl A, Papakonstantinou E, Hartig R, et al. Insulin and insulin-like growth factor-1 can modulate the phosphoinositide- 3-kinase/Akt/FoxO1 pathway in SZ95 sebocytes in vitro. Mol Cell Endocrinol. 2015;415:32–44. doi: 10.1016/j.mce.2015.08.001. [DOI] [PubMed] [Google Scholar]
- 10.Agamia NF, Abdallah DM, Sorour O, Mourad B, Younan DN. Skin expression of mammalian target of rapamycin and forkhead box transcription factor O1, and serum insulin-like growth factor-1 in patients with acne vulgaris and their relationship with diet. Br J Dermatol. 2016;174:1299–1307. doi: 10.1111/bjd.14409. [DOI] [PubMed] [Google Scholar]
- 11.Tsitsipatis D, Klotz LO, Steinbrenner H. Multifaceted functions of the forkhead box transcription factors FoxO1 and FoxO3 in skin. Biochim Biophys Acta. 2017;1861:1057–1064. doi: 10.1016/j.bbagen.2017.02.027. [DOI] [PubMed] [Google Scholar]
- 12.Gross DN, van den Heuvel AP, Birnbaum MJ. The role of FoxO in the regulation of metabolism. Oncogene. 2008;27:2320–2336. doi: 10.1038/onc.2008.25. [DOI] [PubMed] [Google Scholar]
- 13.Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and disease. Cell. 2017;168:960–976. doi: 10.1016/j.cell.2017.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ben-Sahra I, Manning BD. mTORC1 signaling and the metabolic control of cell growth. Curr Opin Cell Biol. 2017;45:72–82. doi: 10.1016/j.ceb.2017.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Laplante M, Sabatini DM. Regulation of mTORC1 and its impact on gene expression at a glance. J Cell Sci. 2013;126:1713–1719. doi: 10.1242/jcs.125773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Melnik B. Dietary intervention in acne. Attenuation of increased mTORC1 signaling promoted by Western diet. Dermatoendocrinol. 2012;4:20–32. doi: 10.4161/derm.19828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Monfrecola G, Lembo S, Caiazzo G, De Vita V, Di Caprio R, Balato A, et al. Mechanistic target of rapamycin (mTOR) expression is increased in acne patients’ skin. Exp Dermatol. 2016;25:153–155. doi: 10.1111/exd.12885. [DOI] [PubMed] [Google Scholar]
- 18.Smith TM, Cong Z, Gilliland KL, Clawson GA, Thiboutot DM. Insulin-like growth factor-1 induces lipid production in human SEB-1 sebocytes via sterol response element-binding protein-1. J Invest Dermatol. 2006;126:1226–1232. doi: 10.1038/sj.jid.5700278. [DOI] [PubMed] [Google Scholar]
- 19.Smith TM, Gilliland K, Clawson GA, Thiboutot D. IGF-1 induces SREBP-1 expression and lipogenesis in SEB-1 sebocytes via activation of the phosphoinositide 3-kinase/Akt pathway. J Invest Dermatol. 2008;128:1286–1293. doi: 10.1038/sj.jid.5701155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ottaviani M, Camera E, Picardo M. Lipid mediators in acne. Mediators Inflamm. 2010;2010:858176. doi: 10.1155/2010/858176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zouboulis CC, Jourdan E, Picardo M. Acne is an inflammatory disease and alterations of sebum composition initiate acne lesions. J Eur Acad Dermatol Venereol. 2014;28:527–532. doi: 10.1111/jdv.12298. [DOI] [PubMed] [Google Scholar]
- 22.Melnik BC. Linking diet to acne metabolomics, inflammation, and comedogenesis: an update. Clin Cosmet Investig Dermatol. 2015;8:371–388. doi: 10.2147/CCID.S69135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Melnik BC. Acne vulgaris: an inflammasomopathy of the sebaceous follicle induced by deviated FoxO1/mTORC1 signalling. Br J Dermatol. 2016;174:1186–1188. doi: 10.1111/bjd.14564. [DOI] [PubMed] [Google Scholar]
- 24.Roca H, Varsos ZS, Pienta KJ. CCL2 is a negative regulator of AMP- activated protein kinase to sustain mTOR complex-1 activation, survivin expression, and cell survival in human prostate cancer PC3 cells. Neoplasia. 2009;11:1309–1317. doi: 10.1593/neo.09936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Song K, Shankar E, Yang J, Bane KL, Wahdan-Alaswad R, Danielpour D. Critical role of a survivin/TGF-β/mTORC1 axis in IGF-I-mediated growth of prostate epithelial cells. PLoS ONE. 2013;8:e61896. doi: 10.1371/journal.pone.0061896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Assaf HA, Abdel-Maged WM, Elsadek BE, Hassan MH, Adly MA, Ali SA. Survivin as a novel biomarker in the pathogenesis of acne vulgaris and its correlation to insulin-like growth factor-I. Dis Markers. 2016;2016:7040312. doi: 10.1155/2016/7040312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Melnik BC. Diet in acne: further evidence for the role of nutrient signalling in acne pathogenesis. Acta Derm Venereol. 2012;92:228–231. doi: 10.2340/00015555-1358. [DOI] [PubMed] [Google Scholar]
- 28.Melnik BC, John SM, Plewig G. Acne: risk indicator for increased body mass index and insulin resistance. Acta Derm Venereol. 2013;93:644–649. doi: 10.2340/00015555-1677. [DOI] [PubMed] [Google Scholar]
- 29.Lane DP. Cancer. p53, guardian of the genome. Nature. 1992;358:15–16. doi: 10.1038/358015a0. [DOI] [PubMed] [Google Scholar]
- 30.Botchkarev VA, Flores ER. p53/p63/p73 in the epidermis in health and disease. Cold Spring Harb Perspect Med. 2014;4:a015248. doi: 10.1101/cshperspect.a015248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kligman AM. How to use topical tretinoin in treating acne. Cutis. 1995;56:83–84. [PubMed] [Google Scholar]
- 32.Yeh L, Bonati LM, Silverberg NB. Topical retinoids for acne. Semin Cutan Med Surg. 2016;35:50–56. doi: 10.12788/j.sder.2016.024. [DOI] [PubMed] [Google Scholar]
- 33.Zheng P, Gendimenico GJ, Mezick JA, Kligman AM. Topical all-trans retinoic acid rapidly corrects the follicular abnormalities of the rhino mouse. An ultrastructural study. Acta Derm Venereol. 1993;73:97–101. doi: 10.2340/000155557397101. [DOI] [PubMed] [Google Scholar]
- 34.Um SJ, Kim EJ, Hwang ES, Kim SJ, Namkoong SE, Park JS. Antiproliferative effects of retinoic acid/interferon in cervical carcinoma cell lines: cooperative growth suppression of IRF-1 and p53. Int J Cancer. 2000;85:416–423. doi: 10.1002/(SICI)1097-0215(20000201)85:3<416::AID-IJC19>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- 35.Zheng A, Mäntymaa P, Säily M, Savolainen E, Vähäkangas K, Koistinen P. p53 pathway in apoptosis induced by all-trans-retinoic acid in acute myeloblastic leukaemia cells. Acta Haematol. 2000;103:135–143. doi: 10.1159/000041036. [DOI] [PubMed] [Google Scholar]
- 36.Curtin JC, Dragnev KH, Sekula D, Christie AJ, Dmitrovsky E, Spinella MJ. Retinoic acid activates p53 in human embryonal carcinoma through retinoid receptor-dependent stimulation of p53 transactivation function. Oncogene. 2001;20:2559–2569. doi: 10.1038/sj.onc.1204370. [DOI] [PubMed] [Google Scholar]
- 37.Lu J, Zhang F, Yuan Y, Ding C, Zhang L, Li Q. All-trans retinoic acid upregulates the expression of p53 via axin and inhibits the proliferation of glioma cells. Oncol Rep. 2013;29:2269–2274. doi: 10.3892/or.2013.2391. [DOI] [PubMed] [Google Scholar]
- 38.Jain AK, Allton K, Iacovino M, Mahen E, Milczarek RJ, Zwaka TP, et al. p53 regulates cell cycle and microRNAs to promote differentiation of human embryonic stem cells. PLoS Biol. 2012;10:e1001268. doi: 10.1371/journal.pbio.1001268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee DD, Stojadinovic O, Krzyzanowska A, Vouthounis C, Blumenberg M, Tomic-Canic M. Retinoid-responsive transcriptional changes in epidermal keratinocytes. J Cell Physiol. 2009;220:427–439. doi: 10.1002/jcp.21784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mrass P, Rendl M, Mildner M, Gruber F, Lengauer B, Ballaun C, et al. Retinoic acid increases the expression of p53 and proapoptotic caspases and sensitizes keratinocytes to apoptosis: a possible explanation for tumor preventive action of retinoids. Cancer Res. 2004;64:6542–6548. doi: 10.1158/0008-5472.CAN-04-1129. [DOI] [PubMed] [Google Scholar]
- 41.Kim J, Nakasaki M, Todorova D, Lake B, Yuan CY, Jamora C, et al. p53 induces skin aging by depleting Blimp1+ sebaceous gland cells. Cell Death Dis. 2014;5:e1141. doi: 10.1038/cddis.2014.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Feng Z, Zhang H, Levine AJ, Jin S. The coordinate regulation of the p53 and mTOR pathways in cells. Proc Natl Acad Sci USA. 2005;102:8204–8209. doi: 10.1073/pnas.0502857102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Budanov AV, Karin M. p53 target genes sestrin1 and sestrin2 connect genotoxic stress and mTOR signaling. Cell. 2008;134:451–460. doi: 10.1016/j.cell.2008.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cooks T, Harris CC, Oren M. Caught in the cross fire: p53 in inflammation. Carcinogenesis. 2014;35:1680–1690. doi: 10.1093/carcin/bgu134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Heyne K, Winter C, Gerten F, Schmidt C, Roemer K. A novel mechanism of crosstalk between the p53 and NFκB pathways: MDM2 binds and inhibits p65RelA. Cell Cycle. 2013;12:2479–2492. doi: 10.4161/cc.25495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mehta K. Retinoic acid—a player that rules the game of life and death in neutrophils. Indian J Exp Biol. 2002;40:874–881. [PubMed] [Google Scholar]
- 47.Channabasappa S, Stewart S, Caldwell S, Carr A, Singh B. Retinoic acid induces apoptosis in activated canine neutrophils. Vet Immunol Immunopathol. 2014;157:175–181. doi: 10.1016/j.vetimm.2013.11.003. [DOI] [PubMed] [Google Scholar]
- 48.Duquette SC, Fischer CD, Feener TD, Muench GP, Morck DW, Barreda DR, et al. Anti-inflammatory effects of retinoids and carotenoid derivatives on caspase-3-dependent apoptosis and efferocytosis of bovine neutrophils. Am J Vet Res. 2014;75:1064–1075. doi: 10.2460/ajvr.75.12.1064. [DOI] [PubMed] [Google Scholar]
- 49.Nawata H, Maeda Y, Sumimoto Y, Miyatake J, Kanamaru A. A mechanism of apoptosis induced by all-trans retinoic acid on adult T-cell leukemia cells: a possible involvement of the Tax/NF-kappaB signaling pathway. Leuk Res. 2001;25:323–331. doi: 10.1016/S0145-2126(00)00126-0. [DOI] [PubMed] [Google Scholar]
- 50.Fulton JE, Jr, Farzad-Bakshandeh A, Bradley S. Studies on the mechanism of action to topical benzoyl peroxide and vitamin A acid in acne vulgaris. J Cutan Pathol. 1974;1:191–200. doi: 10.1111/j.1600-0560.1974.tb00628.x. [DOI] [PubMed] [Google Scholar]
- 51.Oh CW, Myung KB. An ultrastructural study of the retention hyperkeratosis of experimentally induced comedones in rabbits: the effects of three comedolytics. J Dermatol. 1996;23:169–180. doi: 10.1111/j.1346-8138.1996.tb03993.x. [DOI] [PubMed] [Google Scholar]
- 52.Waller JM, Dreher F, Behnam S, Ford C, Lee C, Tiet T, et al. ‘Keratolytic’ properties of benzoyl peroxide and retinoic acid resemble salicylic acid in man. Skin Pharmacol Physiol. 2006;19:283–289. doi: 10.1159/000093984. [DOI] [PubMed] [Google Scholar]
- 53.Gloor M, Klump H, Wirth H. Cytokinetic studies on the sebo-suppressive effect of drugs using the example of benzoyl peroxide. Arch Dermatol Res. 1980;267:97–99. doi: 10.1007/BF00416928. [DOI] [PubMed] [Google Scholar]
- 54.Fanta D. Klinische und experimentelle Untersuchungen über die Wirkung von Benzoylperoxid in der Behandlung der Akne. Hautarzt. 1978;29:481–486. [PubMed] [Google Scholar]
- 55.Wirth H, Spürgel D, Gloor M. Untersuchungen zur Wirkung von Benzoylperoxid auf die Talgdrüsensekretion. Dermatol Monatsschr. 1983;169:289–293. [PubMed] [Google Scholar]
- 56.Puschmann M. Klinisch-experimentelle Untersuchungen zum Wirkungsmechanismus von Benzoylperoxid. Hautarzt. 1982;33:257–265. [PubMed] [Google Scholar]
- 57.Mezick JA, Thorne EG, Bhatia MC, Shea LM, Capetola JR. The rabbit ear microcomedo prevention assay. A new model to evaluate antiacne agents. In: Maibach HI, Lowe NJ, editors. Models in Dermatology, vol 3. Karger: Basel, München, Paris, London, New York, New Delhi, Singapore, Tokyo, Sydney; 1987. p. 68–73.
- 58.Valacchi G, Rimbach G, Saliou C, Weber SU, Packer L. Effect of benzoyl peroxide on antioxidant status, NF-kappaB activity and interleukin-1alpha gene expression in human keratinocytes. Toxicology. 2001;165:225–234. doi: 10.1016/S0300-483X(01)00430-9. [DOI] [PubMed] [Google Scholar]
- 59.Kennedy CH, Winston GW, Church DF, Pryor WA. Benzoyl peroxide interaction with mitochondria: inhibition of respiration and induction of rapid, large-amplitude swelling. Arch Biochem Biophys. 1989;271:456–470. doi: 10.1016/0003-9861(89)90296-8. [DOI] [PubMed] [Google Scholar]
- 60.Milani M, Bigardi A, Zavattarelli M. Efficacy and safety of stabilized hydrogen peroxide cream (Crystacide®) in mild-to-moderate acne vulgaris: a randomized, controlled trial versus benzoyl peroxide gel. Curr Med Res Opin. 2003;19:135–138. doi: 10.1185/030079902125001523. [DOI] [PubMed] [Google Scholar]
- 61.Muizzuddin N, Schnittger S, Maher W, Maes DH, Mammone T. Enzymatically generated hydrogen peroxide reduces the number of acne lesions in acne vulgaris. J Cosmet Sci. 2013;64:1–8. [PubMed] [Google Scholar]
- 62.Veraldi S, Micali G, Berardesca E, Dall’Oglio F, Sinagra JL, Guanziroli E. Results of a multicenter, randomized, controlled trial of a hydrogen peroxide-based kit versus a benzoyl peroxide-based kit in mild-to-moderate acne. J Clin Aesthet Dermatol. 2016;9:50–54. [PMC free article] [PubMed] [Google Scholar]
- 63.Pronsato L, Milanesi L. Effect of testosterone on the regulation of p53 and p66Shc during oxidative stress damage in C2C12 cells. Steroids. 2016;106:41–54. doi: 10.1016/j.steroids.2015.12.007. [DOI] [PubMed] [Google Scholar]
- 64.Park JH, Zhuang J, Li J, Hwang PM. p53 as guardian of the mitochondrial genome. FEBS Lett. 2016;590:924–934. doi: 10.1002/1873-3468.12061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Budanov AV, Sablina AA, Feinstein E, Koonin EV, Chumakov PM. Regeneration of peroxiredoxins by p53-regulated sestrins, homologs of bacterial AhpD. Science. 2004;304:596–600. doi: 10.1126/science.1095569. [DOI] [PubMed] [Google Scholar]
- 66.Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, et al. Sestrin as a feedback inhibitor of TOR that prevents age-related pathologies. Science. 2010;327:1223–1228. doi: 10.1126/science.1182228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Budanov AV. Stress-responsive sestrins link p53 with redox regulation and mammalian target of rapamycin signaling. Antioxid Redox Signal. 2011;15:1679–1690. doi: 10.1089/ars.2010.3530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Fabbrocini G, Izzo R, Faggiano A, Del Prete M, Donnarumma M, Marasca C, et al. Low glycaemic diet and metformin therapy: a new approach in male subjects with acne resistant to common treatments. Clin Exp Dermatol. 2016;41:38–42. doi: 10.1111/ced.12673. [DOI] [PubMed] [Google Scholar]
- 69.Melnik BC, Schmitz G. Metformin: an inhibitor of mTORC1 signaling. J Endocrinol Diabetes Obes. 2014;2:1029. [Google Scholar]
- 70.Shafiee MN, Malik DA, Yunos RI, Atiomo W, Omar MH, Ghani NA, et al. The effect of metformin on endometrial tumor-regulatory genes and systemic metabolic parameters in polycystic ovarian syndrome—a proof-of-concept study. Gynecol Endocrinol. 2015;31:286–290. doi: 10.3109/09513590.2014.989982. [DOI] [PubMed] [Google Scholar]
- 71.Boen M, Brownell J, Patel P, Tsoukas MM. The role of photodynamic therapy in acne: an evidence-based review. Am J Clin Dermatol. 2017;18:311–321. doi: 10.1007/s40257-017-0255-3. [DOI] [PubMed] [Google Scholar]
- 72.Fonda-Pascual P, Moreno-Arrones OM, Alegre-Sanchez A, Saceda-Corralo D, Buendia-Castaño D, Pindado-Ortega C, et al. In situ production of ROS in the skin by photodynamic therapy as a powerful tool in clinical dermatology. Methods. 2016;109:190–202. doi: 10.1016/j.ymeth.2016.07.008. [DOI] [PubMed] [Google Scholar]
- 73.Zuliani T, Khammari A, Chaussy H, Knol AC, Dréno B. Ex vivo demonstration of a synergistic effect of Adapalene and benzoyl peroxide on inflammatory acne lesions. Exp Dermatol. 2011;20:850–853. doi: 10.1111/j.1600-0625.2011.01339.x. [DOI] [PubMed] [Google Scholar]
- 74.Shemer A, Weiss G, Amichai B, Kaplan B, Trau H. Azelaic acid (20%) cream in the treatment of acne vulgaris. J Eur Acad Dermatol Venereol. 2002;16:178–179. doi: 10.1046/j.1468-3083.2002.00392_6.x. [DOI] [PubMed] [Google Scholar]
- 75.Schulte BC, Wu W, Rosen T. Azelaic acid: evidence-based update on mechanism of action and clinical application. J Drugs Dermatol. 2015;14:964–968. [PubMed] [Google Scholar]
- 76.Mayer-da Silva A, Gollnick H, Imcke E, Orfanos CE. Azelaic acid vs. placebo: effects on normal human keratinocytes and melanocytes. Electron microscopic evaluation after long-term application in vivo. Acta Derm Venereol. 1987;67:116–122. doi: 10.2340/0001555567116122. [DOI] [PubMed] [Google Scholar]
- 77.Detmar M, Mayer-da-Silva A, Stadler R, Orfanos CE. Effects of azelaic acid on proliferation and ultrastructure of mouse keratinocytes in vitro. J Invest Dermatol. 1989;93:70–74. doi: 10.1111/1523-1747.ep12277353. [DOI] [PubMed] [Google Scholar]
- 78.Mayer-da-Silva A, Gollnick H, Detmar M, Gassmüller J, Parry A, Müller R, et al. Effects of azelaic acid on sebaceous gland, sebum excretion rate and keratinization pattern in human skin. An in vivo and in vitro study. Acta Derm Venereol Suppl (Stockh) 1989;143:20–30. [PubMed] [Google Scholar]
- 79.Passi S, Picardo M, Nazzaro-Porro M, Breathnach A, Confaloni AM, Serlupi- Crescenzi G. Antimitochondrial effect of saturated medium chain length (C8–C13) dicarboxylic acids. Biochem Pharmacol. 1984;33:103–108. doi: 10.1016/0006-2952(84)90376-9. [DOI] [PubMed] [Google Scholar]
- 80.Chen R, Yang L, McIntyre TM. Cytotoxic phospholipid oxidation products. Cell death from mitochondrial damage and the intrinsic caspase cascade. J Biol Chem. 2007;282:24842–24850. doi: 10.1074/jbc.M702865200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Holley AK, St Clair DK. Watching the watcher: regulation of p53 by mitochondria. Future Oncol. 2009;5:117–130. doi: 10.2217/14796694.5.1.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Pan Y, Liu D, Wei Y, Su D, Lu C, Hu Y, et al. Azelaic acid exerts antileukemic activity in acute myeloid leukemia. Front Pharmacol. 2017;8:359. doi: 10.3389/fphar.2017.00359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Del Rosso JQ. Topical and oral antibiotics for acne vulgaris. Semin Cutan Med Surg. 2016;35:57–61. doi: 10.12788/j.sder.2016.025. [DOI] [PubMed] [Google Scholar]
- 84.Perret LJ, Tait CP. Non-antibiotic properties of tetracyclines and their clinical application in dermatology. Australas J Dermatol. 2014;55:111–118. doi: 10.1111/ajd.12075. [DOI] [PubMed] [Google Scholar]
- 85.Moore A, Ling M, Bucko A, Manna V, Rueda MJ. Efficacy and safety of subantimicrobial dose, modified-release doxycycline 40 mg versus doxycycline 100 mg versus placebo for the treatment of inflammatory lesions in moderate and severe acne: a randomized, double-blinded, controlled study. J Drugs Dermatol. 2015;14:581–586. [PubMed] [Google Scholar]
- 86.Mollan SP, Ali F, Hassan-Smith G, Botfield H, Friedman DI, Alexandra J, et al. Evolving evidence in adult idiopathic intracranial hypertension: pathophysiology and management. J Neurol Neurosurg Psychiatry. 2016;87:982–992. doi: 10.1136/jnnp-2015-311302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Regen F, Le Bret N, Hildebrand M, Herzog I, Heuser I, Hellmann-Regen J. Inhibition of brain retinoic acid catabolism: a mechanism for minocycline’s pleiotropic actions? World J Biol Psychiatry. 2016;17:634–640. doi: 10.3109/15622975.2015.1036116. [DOI] [PubMed] [Google Scholar]
- 88.Hellmann-Regen J, Herzog I, Fischer N, Heuser I, Regen F. Do tetracyclines and erythromycin exert anti-acne effects by inhibition of P450-mediated degradation of retinoic acid? Exp Dermatol. 2014;23:290–293. doi: 10.1111/exd.12358. [DOI] [PubMed] [Google Scholar]
- 89.Regen F, Hildebrand M, Le Bret N, Herzog I, Heuser I, Hellmann-Regen J. Inhibition of retinoic acid catabolism by minocycline: evidence for a novel mode of action? Exp Dermatol. 2015;24:473–476. doi: 10.1111/exd.12692. [DOI] [PubMed] [Google Scholar]
- 90.Zhu Z, Hotchkiss SA, Boobis AR, Edwards RJ. Expression of P450 enzymes in rat whole skin and cultured epidermal keratinocytes. Biochem Biophys Res Commun. 2002;297:65–70. doi: 10.1016/S0006-291X(02)02134-4. [DOI] [PubMed] [Google Scholar]
- 91.Burt HJ, Galetin A, Houston JB. IC50-based approaches as an alternative method for assessment of time-dependent inhibition of CYP3A4. Xenobiotica. 2010;40:331–343. doi: 10.3109/00498251003698555. [DOI] [PubMed] [Google Scholar]
- 92.Ross AC, Zolfaghari R. Cytochrome P450 s in the regulation of cellular retinoic acid metabolism. Annu Rev Nutr. 2011;31:65–87. doi: 10.1146/annurev-nutr-072610-145127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Melnik BC. The P450 system and mTORC1 signalling in acne. Exp Dermatol. 2014;23:318–319. doi: 10.1111/exd.12359. [DOI] [PubMed] [Google Scholar]
- 94.Ataie-Kachoie P, Pourgholami MH, Bahrami-B F, Badar S, Morris DL. Minocycline attenuates hypoxia-inducible factor-1α expression correlated with modulation of p53 and AKT/mTOR/p70S6K/4E-BP1 pathway in ovarian cancer: in vitro and in vivo studies. Am J Cancer Res. 2015;5:575–588. [PMC free article] [PubMed] [Google Scholar]
- 95.Lee MD, Ayanoglu E, Gong L. Drug-induced changes in P450 enzyme expression at the gene expression level: a new dimension to the analysis of drug-drug interactions. Xenobiotica. 2006;36:1013–1080. doi: 10.1080/00498250600861785. [DOI] [PubMed] [Google Scholar]
- 96.Lynch T, Price A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am Fam Phys. 2007;76:391–396. [PubMed] [Google Scholar]
- 97.Matoulková P, Pávek P, Malý J, Vlček J. Cytochrome P450 enzyme regulation by glucocorticoids and consequences in terms of drug interaction. Expert Opin Drug Metab Toxicol. 2014;10:425–435. doi: 10.1517/17425255.2014.878703. [DOI] [PubMed] [Google Scholar]
- 98.Momin SB, Peterson A, Del Rosso JQ. A status report on drug-associated acne and acneiform eruptions. J Drugs Dermatol. 2010;9:627–636. [PubMed] [Google Scholar]
- 99.Du-Thanh A, Kluger N, Bensalleh H, Guillot B. Drug-induced acneiform eruption. Am J Clin Dermatol. 2011;12:233–245. doi: 10.2165/11588900-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 100.Kazandjieva J, Tsankov N. Drug-induced acne. Clin Dermatol. 2017;35:156–162. doi: 10.1016/j.clindermatol.2016.10.007. [DOI] [PubMed] [Google Scholar]
- 101.Peck GL, Olsen TG, Yoder FW, Strauss JS, Downing DT, Pandya M, et al. Prolonged remissions of cystic and conglobate acne with 13-cis-retinoic acid. N Engl J Med. 1979;300:329–333. doi: 10.1056/NEJM197902153000701. [DOI] [PubMed] [Google Scholar]
- 102.Strauss JS, Stranieri AM. Changes in long-term sebum production from isotretinoin therapy. J Am Acad Dermatol. 1982;6(4 Pt 2 Suppl):751–756. doi: 10.1016/S0190-9622(82)80055-8. [DOI] [PubMed] [Google Scholar]
- 103.Melnik B, Kinner T, Plewig G. Influence of oral isotretinoin treatment on the composition of comedonal lipids. Implications for comedogenesis in acne vulgaris. Arch Dermatol Res. 1988;280:97–102. doi: 10.1007/BF00417712. [DOI] [PubMed] [Google Scholar]
- 104.Plewig G, Wagner A. Anti-inflammatory effects of 13-cis-retinoic acid. An in vivo study. Arch Dermatol Res. 1981;270:89–94. doi: 10.1007/BF00417154. [DOI] [PubMed] [Google Scholar]
- 105.Strauss JS, Stewart ME, Downing DT. The effect of 13-cis-retinoic acid on sebaceous glands. Arch Dermatol. 1987;123:1538a–1541a. doi: 10.1001/archderm.1987.01660350142031. [DOI] [PubMed] [Google Scholar]
- 106.Kelhälä HL, Fyhrquist N, Palatsi R, Lehtimäki S, Väyrynen JP, Kubin ME, et al. Isotretinoin treatment reduces acne lesions but not directly lesional acne inflammation. Exp Dermatol. 2016;25:477–478. doi: 10.1111/exd.12971. [DOI] [PubMed] [Google Scholar]
- 107.Landthaler M, Kummermehr J, Wagner A, Plewig G. Inhibitory effects of 13-cis-retinoic acid on human sebaceous glands. Arch Dermatol Res. 1980;269:297–309. doi: 10.1007/BF00406424. [DOI] [PubMed] [Google Scholar]
- 108.Nelson AM, Zhao W, Gilliland KL, Zaenglein AL, Liu W, Thiboutot DM. Temporal changes in gene expression in the skin of patients treated with isotretinoin provide insight into its mechanism of action. Dermatoendocrinology. 2009;1:177–187. doi: 10.4161/derm.1.3.8258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Tsukada M, Schröder M, Roos TC, Chandraratna RA, Reichert U, Merk HF, et al. 13-cis retinoic acid exerts its specific activity on human sebocytes through selective intracellular isomerization to all-trans retinoic acid and binding to retinoid acid receptors. J Invest Dermatol. 2000;115:321–327. doi: 10.1046/j.1523-1747.2000.00066.x. [DOI] [PubMed] [Google Scholar]
- 110.Kuribayashi K, Krigsfeld G, Wang W, Xu J, Mayes PA, Dicker DT, et al. TNFSF10 (TRAIL), a p53 target gene that mediates p53-dependent cell death. Cancer Biol Ther. 2008;7:2034–2038. doi: 10.4161/cbt.7.12.7460. [DOI] [PubMed] [Google Scholar]
- 111.Dhandapani L, Yue P, Ramalingam SS, Khuri FR, Sun SY. Retinoic acid enhances TRAIL-induced apoptosis in cancer cells by upregulating TRAIL receptor 1 expression. Cancer Res. 2011;71:5245–5254. doi: 10.1158/0008-5472.CAN-10-4180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Kim MJ, Ahn K, Park SH, Kang HJ, Jang BG, Oh SJ, et al. SIRT1 regulates tyrosine hydroxylase expression and differentiation of neuroblastoma cells via FOXO3a. FEBS Lett. 2009;583:1183–1188. doi: 10.1016/j.febslet.2009.03.007. [DOI] [PubMed] [Google Scholar]
- 113.Sakoe Y, Sakoe K, Kirito K, Ozawa K, Komatsu N. FOXO3A as a key molecule for all-trans retinoic acid-induced granulocytic differentiation and apoptosis in acute promyelocytic leukemia. Blood. 2010;115:3787–3795. doi: 10.1182/blood-2009-05-222976. [DOI] [PubMed] [Google Scholar]
- 114.Kurinna S, Stratton SA, Tsai WW, Akdemir KC, Gu W, Singh P, et al. Direct activation of forkhead box O3 by tumor suppressors p53 and p73 is disrupted during liver regeneration in mice. Hepatology. 2010;52:1023–1032. doi: 10.1002/hep.23746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Renault VM, Thekkat PU, Hoang KL, White JL, Brady CA, Kenzelmann Broz D, et al. The pro-longevity gene FoxO3 is a direct target of the p53 tumor suppressor. Oncogene. 2011;30:3207–3221. doi: 10.1038/onc.2011.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Nayak G, Cooper GM. p53 is a major component of the transcriptional and apoptotic program regulated by PI 3-kinase/Akt/GSK3 signaling. Cell Death Dis. 2012;3:e400. doi: 10.1038/cddis.2012.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Feng Z. p53 regulation of the IGF-1/AKT/mTOR pathways and the endosomal compartment. Cold Spring Harb Perspect Biol. 2010;2:a001057. doi: 10.1101/cshperspect.a001057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zhang X, Tang N, Hadden TJ, Rishi AK. Akt, FoxO and regulation of apoptosis. Biochim Biophys Acta. 2011;1813:1978–1986. doi: 10.1016/j.bbamcr.2011.03.010. [DOI] [PubMed] [Google Scholar]
- 119.Nelson AM, Zhao W, Gilliland KL, Zaenglein AL, Liu W, Thiboutot DM. Neutrophil gelatinase-associated lipocalin mediates 13-cis retinoic acid- induced apoptosis of human sebaceous gland cells. J Clin Invest. 2008;118:1468–1478. doi: 10.1172/JCI33869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Nelson AM, Cong Z, Gilliland KL, Thiboutot DM. TRAIL contributes to the apoptotic effect of 13-cis retinoic acid in human sebaceous gland cells. Br J Dermatol. 2011;165:526–533. doi: 10.1111/j.1365-2133.2011.10392.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Wu NL, Lee TA, Tsai TL, Lin WW. TRAIL-induced keratinocyte differentiation requires caspase activation and p63 expression. J Invest Dermatol. 2011;131:874–883. doi: 10.1038/jid.2010.402. [DOI] [PubMed] [Google Scholar]
- 122.Tsujita-Kyutoku M, Kiuchi K, Danbara N, Yuri T, Senzaki H, Tsubura A. p63 expression in normal human epidermis and epidermal appendages and their tumors. J Cutan Pathol. 2003;30:11–17. doi: 10.1034/j.1600-0560.2003.300102.x. [DOI] [PubMed] [Google Scholar]
- 123.Rudman SM, Philpott MP, Thomas GA, Kealey T. The role of IGF-I in human skin and its appendages: morphogen as well as mitogen? J Invest Dermatol. 1997;109:770–777. doi: 10.1111/1523-1747.ep12340934. [DOI] [PubMed] [Google Scholar]
- 124.Ben-Amitai D, Laron Z. Effect of insulin-like growth factor-1 deficiency or administration on the occurrence of acne. J Eur Acad Dermatol Venereol. 2011;25:950–954. doi: 10.1111/j.1468-3083.2010.03896.x. [DOI] [PubMed] [Google Scholar]
- 125.Melnik BC, John SM, Schmitz G. Over-stimulation of insulin/IGF-1 signaling by western diet may promote diseases of civilization: lessons learnt from Laron syndrome. Nutr Metab (Lond) 2011;8:41. doi: 10.1186/1743-7075-8-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Plewig G. Acne vulgaris: proliferative cells in sebaceous glands. Br J Dermatol. 1974;90:623–630. doi: 10.1111/j.1365-2133.1974.tb06690.x. [DOI] [PubMed] [Google Scholar]
- 127.Plewig G, Fulton JE, Kligman AM. Cellular dynamics of comedo formation in acne vulgaris. Arch Dermatol Forsch. 1971;242:12–29. doi: 10.1007/BF00595286. [DOI] [PubMed] [Google Scholar]
- 128.Werner H, Karnieli E, Rauscher FJ, LeRoith D. Wild-type and mutant p53 differentially regulate transcription of the insulin-like growth factor I receptor gene. Proc Natl Acad Sci USA. 1996;93:8318–8323. doi: 10.1073/pnas.93.16.8318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Melnik BC. Milk—A nutrient system of mammalian evolution promoting mTORC1-dependent translation. Int J Mol Sci. 2015;16:17048–17087. doi: 10.3390/ijms160817048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Werner H, Sarfstein R, LeRoith D, Bruchim I. Insulin-like growth factor 1 signaling axis meets p53 genome protection pathways. Front Oncol. 2016;6:159. doi: 10.3389/fonc.2016.00159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hilmi C, Larribere L, Deckert M, Rocchi S, Giuliano S, Bille K, et al. Involvement of FKHRL1 in melanoma cell survival and death. Pigment Cell Melanoma Res. 2008;21:139–146. doi: 10.1111/j.1755-148X.2008.00440.x. [DOI] [PubMed] [Google Scholar]
- 132.Kiraz Y, Adan A, Kartal Yandim M, Baran Y. Major apoptotic mechanisms and genes involved in apoptosis. Tumour Biol. 2016;37:8471–8486. doi: 10.1007/s13277-016-5035-9. [DOI] [PubMed] [Google Scholar]
- 133.Hay N. Interplay between FOXO, TOR, and Akt. Biochim Biophys Acta. 2011;1813:1965–1970. doi: 10.1016/j.bbamcr.2011.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Nelson AM, Gilliland KL, Cong Z, Thiboutot DM. 13-cis Retinoic acid induces apoptosis and cell cycle arrest in human SEB-1 sebocytes. J Invest Dermatol. 2006;126:2178–2189. doi: 10.1038/sj.jid.5700289. [DOI] [PubMed] [Google Scholar]
- 135.Boulaire J, Fotedar A, Fotedar R. The functions of the cdk-cyclin kinase inhibitor p21WAF1. Pathol Biol (Paris) 2000;48:190–202. [PubMed] [Google Scholar]
- 136.Fischer M. Census and evaluation of p53 target genes. Oncogene. 2017;36:3943–3956. doi: 10.1038/onc.2016.502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.el-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, et al. WAF1, a potential mediator of p53 tumor suppression. Cell. 1993;75:817–825. doi: 10.1016/0092-8674(93)90500-P. [DOI] [PubMed] [Google Scholar]
- 138.el-Deiry WS, Tokino T, Waldman T, Oliner JD, Velculescu VE, Burrell M, et al. Topological control of p21WAF1/CIP1 expression in normal and neoplastic tissues. Cancer Res. 1995;55:2910–2919. [PubMed] [Google Scholar]
- 139.Agarwal S, Bell CM, Taylor SM, Moran RG. p53 Deletion or hotspot mutations enhance mTORC1 activity by altering lysosomal dynamics of TSC2 and Rheb. Mol Cancer Res. 2016;14:66–77. doi: 10.1158/1541-7786.MCR-15-0159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Downie MM, Sanders DA, Maier LM, Stock DM, Kealey T. Peroxisome proliferator-activated receptor and farnesoid X receptor ligands differentially regulate sebaceous differentiation in human sebaceous gland organ cultures in vitro. Br J Dermatol. 2004;151:766–775. doi: 10.1111/j.1365-2133.2004.06171.x. [DOI] [PubMed] [Google Scholar]
- 141.Trivedi NR, Cong Z, Nelson AM, Albert AJ, Rosamilia LL, Sivarajah S, et al. Peroxisome proliferator-activated receptors increase human sebum production. J Invest Dermatol. 2006;126:2002–2009. doi: 10.1038/sj.jid.5700336. [DOI] [PubMed] [Google Scholar]
- 142.Dozsa A, Dezso B, Toth BI, Bacsi A, Poliska S, Camera E, et al. PPARγ- mediated and arachidonic acid-dependent signaling is involved in differentiation and lipid production of human sebocytes. J Invest Dermatol. 2014;134:910–920. doi: 10.1038/jid.2013.413. [DOI] [PubMed] [Google Scholar]
- 143.Schedlich LJ, Graham LD, O’Han MK, Muthukaruppan A, Yan X, Firth SM, et al. Molecular basis of the interaction between IGFBP-3 and retinoid X receptor: role in modulation of RAR-signaling. Arch Biochem Biophys. 2007;465:359–369. doi: 10.1016/j.abb.2007.06.013. [DOI] [PubMed] [Google Scholar]
- 144.Baxter RC. Nuclear actions of insulin-like growth factor binding protein-3. Gene. 2015;569:7–13. doi: 10.1016/j.gene.2015.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Liu B, Lee HY, Weinzimer SA, Powell DR, Clifford JL, Kurie JM, et al. Direct functional interactions between insulin-like growth factor-binding protein-3 and retinoid X receptor-alpha regulate transcriptional signaling and apoptosis. J Biol Chem. 2000;275:33607–33613. doi: 10.1074/jbc.M002547200. [DOI] [PubMed] [Google Scholar]
- 146.Lee KW, Ma L, Yan X, Liu B, Zhang XK, Cohen P. Rapid apoptosis induction by IGFBP-3 involves an insulin-like growth factor-independent nucleomitochondrial translocation of RXRalpha/Nur77. J Biol Chem. 2005;280:16942–16948. doi: 10.1074/jbc.M412757200. [DOI] [PubMed] [Google Scholar]
- 147.Chan SS, Schedlich LJ, Twigg SM, Baxter RC. Inhibition of adipocyte differentiation by insulin-like growth factor-binding protein-3. Am J Physiol Endocrinol Metab. 2009;296:E654–E663. doi: 10.1152/ajpendo.90846.2008. [DOI] [PubMed] [Google Scholar]
- 148.Buckbinder L, Talbott R, Velasco-Miguel S, Takenaka I, Faha B, Seizinger BR, et al. Induction of the growth inhibitor IGF-binding protein 3 by p53. Nature. 1995;377:646–649. doi: 10.1038/377646a0. [DOI] [PubMed] [Google Scholar]
- 149.Melnik BC. Apoptosis may explain the pharmacological mode of action and adverse effects of isotretinoin, including teratogenicity. Acta Derm Venereol. 2017;97:173–181. doi: 10.2340/00015555-2535. [DOI] [PubMed] [Google Scholar]
- 150.Van Nostrand JL, Brady CA, Jung H, Fuentes DR, Kozak MM, Johnson TM, et al. Inappropriate p53 activation during development induces features of CHARGE syndrome. Nature. 2014;514:228–232. doi: 10.1038/nature13585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Melnik BC. Over-expression of p53 explains isotretinoin’s teratogenicity. Exp Dermatol. 2017. doi: 10.1111/exd.13420. [Epub ahead of print]. [DOI] [PubMed]
- 152.Franks S, Layton A, Glasier A. Cyproterone acetate/ethinyl estradiol for acne and hirsutism: time to revise prescribing policy. Hum Reprod. 2008;23:231–232. doi: 10.1093/humrep/dem379. [DOI] [PubMed] [Google Scholar]
- 153.Hassoun LA, Chahal DS, Sivamani RK, Larsen LN. The use of hormonal agents in the treatment of acne. Semin Cutan Med Surg. 2016;35:68–73. doi: 10.12788/j.sder.2016.027. [DOI] [PubMed] [Google Scholar]
- 154.Inoue T, Miki Y, Kakuo S, Hachiya A, Kitahara T, Aiba S, et al. Expression of steroidogenic enzymes in human sebaceous glands. J Endocrinol. 2014;222:301–312. doi: 10.1530/JOE-14-0323. [DOI] [PubMed] [Google Scholar]
- 155.Fan W, Yanase T, Morinaga H, Okabe T, Nomura M, Daitoku H, et al. Insulin- like growth factor 1/insulin signaling activates androgen signaling through direct interactions of Foxo1 with androgen receptor. J Biol Chem. 2007;282:7329–7338. doi: 10.1074/jbc.M610447200. [DOI] [PubMed] [Google Scholar]
- 156.Pappas K, Xu J, Zairis S, Resnick-Silverman L, Abate F, Steinbach N, et al. p53 maintains baseline expression of multiple tumor suppressor genes. Mol Cancer Res. 2017;15:1051–1062. doi: 10.1158/1541-7786.MCR-17-0089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Azmahani A, Nakamura Y, McNamara KM, Sasano H. The role of androgen under normal and pathological conditions in sebaceous glands: the possibility of target therapy. Curr Mol Pharmacol. 2016;9:311–319. doi: 10.2174/1874467208666150710120217. [DOI] [PubMed] [Google Scholar]
- 158.Barrault C, Garnier J, Pedretti N, Cordier-Dirikoc S, Ratineau E, Deguercy A, et al. Androgens induce sebaceous differentiation in sebocyte cells expressing a stable functional androgen receptor. J Steroid Biochem Mol Biol. 2015;152:34–44. doi: 10.1016/j.jsbmb.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 159.Shenk JL, Fisher CJ, Chen SY, Zhou XF, Tillman K, Shemshedini L. p53 represses androgen-induced transactivation of prostate-specific antigen by disrupting hAR amino- to carboxyl-terminal interaction. J Biol Chem. 2001;276:38472–38479. doi: 10.1074/jbc.M103652200. [DOI] [PubMed] [Google Scholar]
- 160.Alimirah F, Panchanathan R, Chen J, Zhang X, Ho SM, Choubey D. Expression of androgen receptor is negatively regulated by p53. Neoplasia. 2007;9:1152–1159. doi: 10.1593/neo.07769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Arnold I, Watt FM. c-Myc activation in transgenic mouse epidermis results in mobilization of stem cells and differentiation of their progeny. Curr Biol. 2001;11:558–568. doi: 10.1016/S0960-9822(01)00154-3. [DOI] [PubMed] [Google Scholar]
- 162.Lo Celso C, Berta MA, Braun KM, Frye M, Lyle S, Zouboulis CC, et al. Characterization of bipotential epidermal progenitors derived from human sebaceous gland: contrasting roles of c-Myc and beta-catenin. Stem Cells. 2008;26:1241–1252. doi: 10.1634/stemcells.2007-0651. [DOI] [PubMed] [Google Scholar]
- 163.Ceballos E, Delgado MD, Gutierrez P, Richard C, Müller D, Eilers M, et al. c-Myc antagonizes the effect of p53 on apoptosis and p21WAF1 transactivation in K562 leukemia cells. Oncogene. 2000;19:2194–2204. doi: 10.1038/sj.onc.1203541. [DOI] [PubMed] [Google Scholar]
- 164.Cottle DL, Kretzschmar K, Schweiger PJ, Quist SR, Gollnick HP, Natsuga K, et al. c-MYC-induced sebaceous gland differentiation is controlled by an androgen receptor/p53 axis. Cell Rep. 2013;3:427–441. doi: 10.1016/j.celrep.2013.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Peck B, Ferber EC, Schulze A. Antagonism between FOXO and MYC regulates cellular powerhouse. Front Oncol. 2013;3:96. doi: 10.3389/fonc.2013.00096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Shi XB, Xue L, Yang J, Ma AH, Zhao J, Xu M, et al. An androgen-regulated miRNA suppresses Bak1 expression and induces androgen-independent growth of prostate cancer cells. Proc Natl Acad Sci USA. 2007;104:19983–19988. doi: 10.1073/pnas.0706641104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Sen A, Prizant H, Light A, Biswas A, Hayes E, Lee HJ, et al. Androgens regulate ovarian follicular development by increasing follicle stimulating hormone receptor and microRNA-125b expression. Proc Natl Acad Sci USA. 2014;111:3008–3013. doi: 10.1073/pnas.1318978111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Le MT, Teh C, Shyh-Chang N, Xie H, Zhou B, Korzh V, et al. MicroRNA-125b is a novel negative regulator of p53. Genes Dev. 2009;23:862–876. doi: 10.1101/gad.1767609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Le MT, Shyh-Chang N, Khaw SL, Chin L, Teh C, Tay J, et al. Conserved regulation of p53 network dosage by microRNA-125b occurs through evolving miRNA-target gene pairs. PLoS Genet. 2011;7:e1002242. doi: 10.1371/journal.pgen.1002242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Liu J, Zhang C, Zhao Y, Feng Z. MicroRNA control of p53. J Cell Biochem. 2017;118:7–14. doi: 10.1002/jcb.25609. [DOI] [PubMed] [Google Scholar]
- 171.Chen L, Wolff DW, Xie Y, Lin MF, Tu Y. Cyproterone acetate enhances TRAIL-induced androgen-independent prostate cancer cell apoptosis via up- regulation of death receptor 5. BMC Cancer. 2017;17:179. doi: 10.1186/s12885-017-3153-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Boudou P, Soliman H, Chivot M, Villette JM, Vexiau P, Belanger A, et al. Effect of oral isotretinoin treatment on skin androgen receptor levels in male acneic patients. J Clin Endocrinol Metab. 1995;80:1158–1161. doi: 10.1210/jcem.80.4.7714084. [DOI] [PubMed] [Google Scholar]
- 173.Kretzschmar K, Cottle DL, Donati G, Chiang MF, Quist SR, Gollnick HP, et al. BLIMP1 is required for postnatal epidermal homeostasis but does not define a sebaceous gland progenitor under steady-state conditions. Stem Cell Rep. 2014;3:620–633. doi: 10.1016/j.stemcr.2014.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Yan J, Jiang J, Lim CA, Wu Q, Ng HH, Chin KC. BLIMP1 regulates cell growth through repression of p53 transcription. Proc Natl Acad Sci USA. 2007;104:1841–1846. doi: 10.1073/pnas.0605562104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Horsley V, O’Carroll D, Tooze R, Ohinata Y, Saitou M, Obukhanych T, et al. Blimp1 defines a progenitor population that governs cellular input to the sebaceous gland. Cell. 2006;126:597–609. doi: 10.1016/j.cell.2006.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Zouboulis C, Seltmann H, Neitzel H, Orfanos C. Establishment and characterization of an immortalized human sebaceous gland cell line. J Invest Dermatol. 1999;113:1011–1020. doi: 10.1046/j.1523-1747.1999.00771.x. [DOI] [PubMed] [Google Scholar]
- 177.Thiboutot D, Jabara S, McAllister JM, Sivarajah A, Gilliland K, Cong Z, et al. Human skin is a steroidogenic tissue: steroidogenic enzymes and cofactors are expressed in epidermis, normal sebocytes, and an immortalized sebocyte cell line (SEB-1) J Invest Dermatol. 2003;120:905–914. doi: 10.1046/j.1523-1747.2003.12244.x. [DOI] [PubMed] [Google Scholar]
- 178.Barrault C, Dichamp I, Garnier J, Pedretti N, Juchaux F, Deguercy A, et al. Immortalized sebocytes can spontaneously differentiate into a sebaceous-like phenotype when cultured as a 3D epithelium. Exp Dermatol. 2012;21:314–316. doi: 10.1111/j.1600-0625.2012.01463.x. [DOI] [PubMed] [Google Scholar]
- 179.Zouboulis CC, Xia L, Akamatsu H, Seltmann H, Fritsch M, Hornemann S, et al. The human sebocyte culture model provides new insights into development and management of seborrhoea and acne. Dermatology. 1998;196:21–31. doi: 10.1159/000017861. [DOI] [PubMed] [Google Scholar]
- 180.Hubbard K, Ozer HL. Mechanism of immortalization. Age (Omaha). 1999;22:65–69. doi: 10.1007/s11357-999-0008-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Jha KK, Banga S, Palejwala V, Ozer HL. SV40-mediated immortalization. Exp Cell Res. 1998;245:1–7. doi: 10.1006/excr.1998.4272. [DOI] [PubMed] [Google Scholar]
- 182.McCormick F, Clark R, Harlow E, Tjian R. SV40 T antigen binds specifically to a cellular 53 K protein in vitro. Nature. 1981;292:63–65. doi: 10.1038/292063a0. [DOI] [PubMed] [Google Scholar]
- 183.Dobbelstein M, Roth J. The large T antigen of simian virus 40 binds and inactivates p53 but not p73. J Gen Virol. 1998;79:3079–3083. doi: 10.1099/0022-1317-79-12-3079. [DOI] [PubMed] [Google Scholar]
- 184.Jiang D, Srinivasan A, Lozano G, Robbins PD. SV40 T antigen abrogates p53-mediated transcriptional activity. Oncogene. 1993;8:2805–2812. [PubMed] [Google Scholar]
- 185.Lazo PA, Santos CR. Interference with p53 functions in human viral infections, a target for novel antiviral strategies? Rev Med Virol. 2011;21:285–300. doi: 10.1002/rmv.696. [DOI] [PubMed] [Google Scholar]
- 186.Wróbel A, Seltmann H, Fimmel S, Müller-Decker K, Tsukada M, Bogdanoff B, et al. Differentiation and apoptosis in human immortalized sebocytes. J Invest Dermatol. 2003;120:175–181. doi: 10.1046/j.1523-1747.2003.12029.x. [DOI] [PubMed] [Google Scholar]
- 187.Guruvayoorappan C, Pradeep CR, Kuttan G. 13-cis-retinoic acid induces apoptosis by modulating caspase-3, bcl-2, and p53 gene expression and regulates the activation of transcription factors in B16F-10 melanoma cells. J Environ Pathol Toxicol Oncol. 2008;27:197–207. doi: 10.1615/JEnvironPatholToxicolOncol.v27.i3.40. [DOI] [PubMed] [Google Scholar]
- 188.Melnik BC, Schmitz G. Are therapeutic effects of antiacne agents mediated by activation of FoxO1 and inhibition of mTORC1? Exp Dermatol. 2013;22:502–504. doi: 10.1111/exd.12172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Toledo F, Wahl GM. Regulating the p53 pathway: in vitro hypotheses, in vivo veritas. Nat Rev Cancer. 2006;6:909–923. doi: 10.1038/nrc2012. [DOI] [PubMed] [Google Scholar]
- 190.Lazo PA. Reverting p53 activation after recovery of cellular stress to resume with cell cycle progression. Cell Signal. 2017;33:49–58. doi: 10.1016/j.cellsig.2017.02.005. [DOI] [PubMed] [Google Scholar]
- 191.Fu W, Ma Q, Chen L, Li P, Zhang M, Ramamoorthy S, et al. MDM2 acts downstream of p53 as an E3 ligase to promote FOXO ubiquitination and degradation. J Biol Chem. 2009;284:13987–14000. doi: 10.1074/jbc.M901758200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Melnik BC. Pro-inflammatory sebocyte growth and survival signalling in acne vulgaris are reversed by pro-apoptotic isotretinoin signalling. Exp Dermatol. 2016;25:676–677. doi: 10.1111/exd.13032. [DOI] [PubMed] [Google Scholar]
- 193.Melnik BC. The TRAIL to acne pathogenesis: let’s focus on death pathways. Exp Dermatol. 2017;26:270–272. doi: 10.1111/exd.13169. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.