

Summary
Nanobodies (Nbs) are the smallest functional antibody fragments known in nature and have multiple applications in biomedicine or environmental monitoring. Nbs are derived from the variable segment of camelid heavy chain‐only antibodies, known as VHH. For selection, libraries of VHH gene segments from naïve, immunized animals or of synthetic origin have been traditionally cloned in E. coli phage display or yeast display systems, and clones binding the target antigen recovered, usually from plastic surfaces with the immobilized antigen (phage display) or using fluorescence‐activated cell sorting (FACS; yeast display). This review briefly describes these conventional approaches and focuses on the distinct properties of an E. coli display system developed in our laboratory, which combines the benefits of both phage display and yeast display systems. We demonstrate that E. coli display using an N‐terminal domain of intimin is an effective platform for the surface display of VHH libraries enabling selection of high‐affinity Nbs by magnetic cell sorting and direct selection on live mammalian cells displaying the target antigen on their surface. Flow cytometry analysis of E. coli bacteria displaying the Nbs on their surface allows monitoring of the selection process, facilitates screening, characterization of antigen‐binding clones, specificity, ligand competition and estimation of the equilibrium dissociation constant (KD).
Introduction
Nanobodies (Nbs) are the smallest, intact antigen‐binding fragments derived from a functional immunoglobulin (Ig). Nbs are recombinant single domain antibody (Ab) fragments with a molecular weight of ~14 kDa and ~2–4 nm in size. They comprise the variable domain of a heavy chain‐only antibody (HCAb; Fig. 1A), which was discovered in the serum of camelids in the early 1990s (Hamers‐Casterman et al., 1993), and are likely an outcome of adaptive changes occurring in conventional Abs within the Camelidae lineage, playing a role in the immune response of these animals (Nguyen et al., 2002; Flajnik et al., 2011; Muyldermans and Smider, 2016). HCAbs lack light chains, and thus, antigen recognition is possible solely through the variable domain of the heavy chain, referred to as VHH for VH of HCAbs. Although Nbs and VHHS are often used interchangeably, it is generally accepted that VHH refers to the gene, or to the polypeptide with unknown specificity, and Nb to the recombinant, purified antigen‐specific polypeptide.
Figure 1.
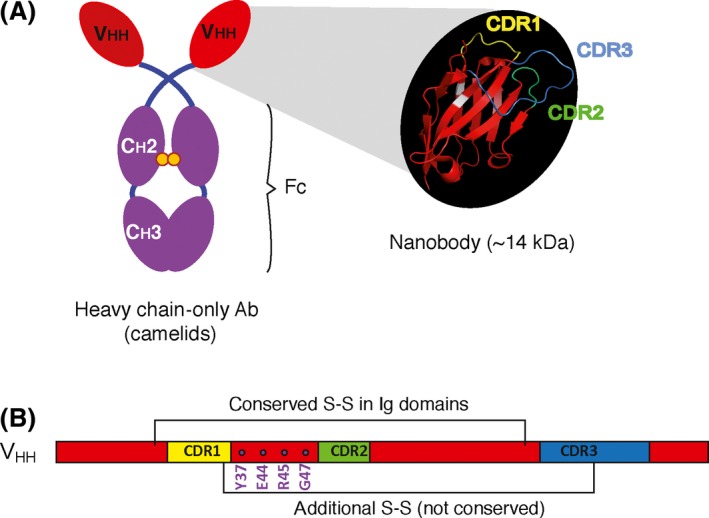
Structure of camelid heavy chain‐only antibodies and nanobodies. (A) Schematic representation of a heavy chain‐only Ab with the structure of the VHH domain (Nb) highlighted. The complementarity‐determining regions (CDRs) are labelled in different colours: CDR1 in yellow, CDR2 in green, CDR3 in blue and the framework regions (FR) are indicated in red. (B) The linear structural diagram of a VHH domain, indicating CDRs, disulfide bonds (S‐S) in the molecule and positions of framework region 2 of VHHs that contain more hydrophilic amino acids (e.g. Y37, E44, R45, G47) compared with conventional VHs (e.g. V37, G44, L45, W47). The conserved canonical S‐S in Ig domains and the additional non‐conserved S‐S found in many Nbs are also illustrated.
Nbs have naturally acquired important adaptations to remain soluble and functional in the absence of the associated light chain variable domain. They have evolved long complementarity‐determining regions (CDRs, Fig. 1B) capable of adopting novel conformations for antigen recognition, subtle amino acid adaptations including substitutions of conserved hydrophobic residues in classical VHs (V37, G44, L45 and F47/W47) for more hydrophilic amino acids (Y37/F37; E44/Q44; R45/C45; G47/R47/L47/S47; Muyldermans, 2013; Muyldermans and Smider, 2016), which confer them strict monomeric behaviour, reversible folding properties, resistance to proteolysis and thermal degradation when compared with the VH from conventional antibodies (van der Linden et al., 1999; Muyldermans et al., 2001; Dumoulin et al., 2002). Furthermore, Nbs often contain, besides the canonical disulfide bond of Ig domains, an extra disulfide bond connecting CDR3 and CDR1 (in camels) or CDR3 with CDR2 (in llamas; Fig. 1B) that assist in stabilizing the conformation of these CDRs and the overall stability of the domain (Govaert et al., 2012). The small size and long CDRs of Nbs allow them to successfully target enzymatic clefts located within concave epitopes of enzymes and proteases (Lauwereys et al., 1998; Conrath et al., 2001; Kromann‐Hansen et al., 2016), and conserved inner regions of surface proteins from pathogens, frequently less accessible to the larger conventional Abs (Stijlemans et al., 2004, 2011; Rossey et al., 2017). Nbs have been used as stabilization agents in crystallographic studies (Ring et al., 2013; Hassaine et al., 2014; Pardon et al., 2014) and as molecular probes in cell biology for visualization of proteins in living cells (Helma et al., 2015). In addition, they share high sequence identity with human VH sequences of family 3, which opens the possibility of their use in human therapy and in in vivo diagnosis (Vaneycken et al., 2011; Chakravarty et al., 2014; Muyldermans and Smider, 2016; Steeland et al., 2016). The short serum half‐life of Nbs due to renal clearance may limit the efficacy of monomeric Nbs in therapeutic applications, but this has been largely overcome by generation of bispecific Nbs recognizing long‐lived serum proteins (e.g. albumin) and the therapeutic target, thus resulting in increased serum half‐lives (Tijink et al., 2008; Vosjan et al., 2012). In addition, the stability and small size of Nbs make them ideal candidates for targeting intracellular proteins and show great potential as intrabodies (Lobato and Rabbitts, 2003; Blanco‐Toribio et al., 2010; Bouchet et al., 2011; Van Impe et al., 2013; Bethuyne et al., 2014; Fulcher et al., 2016; Moutel et al., 2016). Other applications of Nbs have been recently reviewed (Bruce et al., 2016).
Besides these advantages, Nbs can be easily expressed in E. coli, where they are produced as soluble proteins with incorporated affinity tags for purification (Arbabi‐Ghahroudi et al., 2005; Salema and Fernández, 2013). Higher production levels can be obtained from high‐density yeast cultures, like Pichia pastoris and Saccharomyces cerevisiae (Frenken et al., 2000; Holliger, 2002), or in plants (Rajabi‐Memari et al., 2006; De Meyer et al., 2015).
Libraries of VHHs are commonly generated through camel immunization, often dromedaries (Camelus dromedarius) or llamas (Lama glama; Arbabi Ghahroudi et al., 1997; Conrath et al., 2001; Koch‐Nolte et al., 2007; Hassanzadeh‐Ghassabeh et al., 2011), but these may also be obtained from naïve (non‐immune) animals or be from synthetic origin by in vitro CDR randomization (Monegal et al., 2009; Olichon and de Marco, 2012; Moutel et al., 2016). In most cases, the VHHs are cloned into a vector that allows their expression on the surface of a biological entity, usually a bacteriophage or a microbial cell (i.e. yeast, bacteria). Similar to other Ab fragments, the library of bacteriophage or microbial cells displaying the Nbs is then incubated with the target antigen either immobilized, in the soluble biotinylated form, or expressed on cells, for selection (Hoogenboom, 2005). Phages or microbial cells binding the target antigen are isolated and amplified, resulting in the enrichment of high‐affinity binders expressing the selected Nbs.
Phage display
Phage display is a powerful and robust technology that allows display of Nbs (and other Ab fragments) on the surface of filamentous bacteriophages infecting E. coli through the F pilus, such as M13 or f1 (Rakonjac et al., 2011; Verheesen and Laeremans, 2012; Frenzel et al., 2016). In the most common configuration, the VHHs are cloned in a phagemid vector as fusions to the minor phage coat protein 3 (pIII; Fig. 2A; Qi et al., 2012). The cloned VHH is also in frame with an N‐terminal signal peptide driving insertion of the protein fusion into the bacterial inner membrane (IM) of E. coli (Thie et al., 2008). Upon IM insertion, the Nb domain of the Nb‐pIII fusion faces the bacterial periplasm, where protein chaperones and specialized disulfide bond (Dsb) catalysts assist on the folding of the Ig V domains (Fig. 2A; Kadokura et al., 2003; Heras et al., 2009; Schlapschy and Skerra, 2011; Bodelón et al., 2013). The Nb‐pIII fusions are then assembled into phage particles (carrying the phagemid DNA) by infection of bacteria with a ‘helper’ phage that provides all proteins needed for phage replication but has a defective origin of packaging (Rakonjac et al., 2011). A scheme illustrating the display of the Nb into the final phage particle is shown in Fig. 2B. The high phage titres that are obtained from infected E. coli cultures (≥1011 phages ml−1) ensure a good representation of VHHs found in the library, even when large libraries with > 109 clones are utilized (e.g. from naïve and synthetic origins). Nonetheless, representation of VHHs in large phage libraries is also limited by intrinsic E. coli host factors like the capacity of the helper phage to initially infect a sufficient number of bacteria (> 109) in the culture that will produce the phage particles. In addition, phage particles display a maximum of one copy of the Nb‐pIII fusion, with the large majority of phage particles not displaying any fusion unless a helper phage mutant △pIII is used (Rondot et al., 2001; Soltes et al., 2003). In all cases, phagemid particles produced from these cultures are concentrated by precipitation, titred with fresh E. coli cultures and maintained in high‐titre stocks > 1013 colony‐forming units (CFU) ml−1. These phage stocks are subjected to ‘panning’ to enrich for antigen‐binding clones. Most often, this is achieved by incubating phages with the purified antigen, either immobilized onto a plastic surface or in a soluble form covalently labelled with biotin. Due to the ‘sticky’ nature of filamentous phages, extensive and stringent washing conditions need to be used to remove unbound phage particles. Lastly, the bound phages are recovered by elution using acid or alkaline buffers, limited proteolysis, or by competition with soluble antigen, ligand or specific mAb binding an epitope of interest on the antigen (Hoogenboom, 2002). Isolated phage particles are then used to infect E. coli for amplification and eventual analysis of antigen binding by ‘phage ELISA’ using secondary antibodies against the major phage coat protein. Multiple examples of high‐affinity Nbs selected by phage display using purified antigens can be found in the literature; just a few recent examples are cited here (Bethuyne et al., 2014; Lo et al., 2014; Kazemi‐Lomedasht et al., 2015; Nguyen et al., 2015; Zhou et al., 2016; Rossey et al., 2017). However, despite its widespread use, phage display suffers some limitations. These include the need of extensive manipulation for the production of phage stocks (i.e. infection of bacteria with helper phage, amplification by re‐infection), the low levels of Nb display and the high background binding of phage particles, which makes selections against complex non‐purified antigens (e.g. intact cells and tissues) more challenging (Siva et al., 2008; Even‐Desrumeaux et al., 2014; Pavoni et al., 2014).
Figure 2.
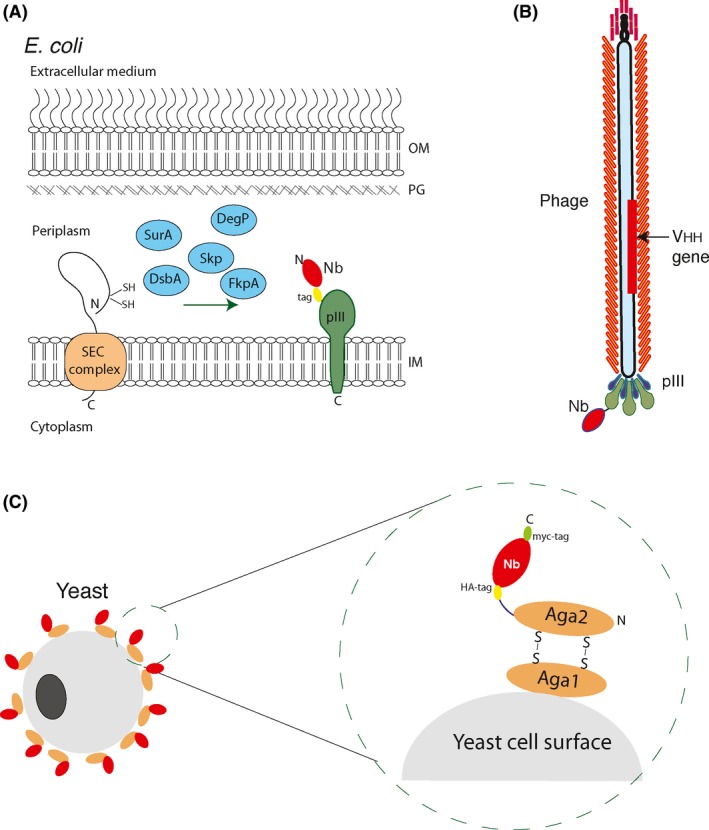
Phage display and yeast display of nanobodies. (A) Expression of Nbs fused to the minor coat protein III (pIII) of filamentous phages in the periplasm of E. coli. The fusion protein is exported to the periplasm with a N‐terminal signal peptide (e.g. PelB) through the Sec complex. The Nb domain is exposed to the periplasm and is tethered to the inner membrane (IM) of E. coli by pIII. An epitope tag (e.g. HA, myc) is usually included between Nb and pIII moieties. Periplasmic chaperones (e.g. SurA, Skp, FkpA, DegP) and disulfide bond catalysts (e.g. DsbA) assist folding of the Nb domain in the periplasm. The E. coli inner membrane (IM), peptidoglycan (PG) and outer membrane (OM) are also indicated. (B) Scheme of a filamentous phage–Nb particle assembled upon infection of E. coli cells expressing Nb‐pIII fusions with a helper phage. The Nb‐pIII fusion is displayed at the tip of the filamentous phage capsid (usually one copy per phage), which contains the phagemid DNA with the encoding VHH gene. (C) Scheme of the display of Nbs on a yeast cell fused to the C‐terminus of the yeast agglutinin receptor (Aga2) present on the yeast cell surface (enlarged). The Aga2‐Nb fusion follows the secretory pathway of yeast cells from the endoplasmic reticulum (ER) to the plasma membrane. Epitope tags flanking the Nb domain are also indicated (e.g. HA, myc).
Yeast display
Cell surface display systems are alternative methods of protein display and are effective tools for Ab or Nb selection and engineering by directed evolution (e.g. affinity maturation; Pardon et al., 2014; Galan et al., 2016). The basic principle is analogous to phage display, providing a physical linkage between the displayed polypeptide anchored in the cell surface and its encoding gene inside the cell. Cell surface display systems have been developed for different host microorganisms, including bacteria and yeast cells. General benefits of these systems versus phage display are their reduced background and culture manipulation, higher surface display level and the possibility of utilizing high‐throughput methods such as fluorescence flow cytometry for the selection of binders and characterization of their antigen affinity and specificity (Gai and Wittrup, 2007). Yeast display system, originally described in Saccharomyces cerevisiae (Boder and Wittrup, 1997, 2000; Feldhaus and Siegel, 2004; Chao et al., 2006) and also developed in the methylotrophic yeast Pichia pastoris (De Schutter and Callewaert, 2012), has been applied in Ab engineering. This system utilizes the α‐agglutinin adhesion receptor found on the surface of yeast cells for protein display (Fig. 2C). The receptor consists of two secreted proteins, Aga1 and Aga2, which are covalently bound to the yeast cell wall. Aga1 attaches to β‐glucans, whereas Aga2 is bound to Aga1 by two disulfide bonds. The Ab fragment to be displayed is fused in frame to Aga2 C‐terminus in a plasmid vector with a regulated promoter (GAL1). The protein fusion also includes two epitope tags (e.g. HA‐ and myc‐tag) flanking the Ab fragment, which enable its detection and surface display using specific monoclonal Abs (mAbs) by fluorescence flow cytometry. The use of a eukaryotic secretory pathway, coupled with protein folding and quality control systems of yeasts, is thought to contribute to the correct folding of more complex mammalian Ab or Ab fragments on yeast cells. The display level on the yeast cell is variable but has been estimated about 3–5 × 104 fusions per cell for single‐chain variable fragments (scFv; Chao et al., 2006; Gai and Wittrup, 2007; Pepper et al., 2008). The potential intrinsic avidity of these elevated display levels is counteracted by the power of fluorescence‐activated cell sorting (FACS). By staining with both soluble biotinylated antigen and the epitope‐tag mAb, and subsequent incubation with secondary fluorescence‐labelled reagents (e.g. streptavidin, anti‐mouse IgG), the yeast cells can be sorted by FACS according to the level of antigen binding and Ab fusion displayed on the surface. Avidity effects are limited using monomeric antigen in solution, but can be increased with multimeric antigens or by preloading tetrameric streptavidin with the biotinylated antigen prior to its incubation with yeast cells (Chao et al., 2006). A clear advantage of yeast display is the possibility to characterize the equilibrium dissociation constant (KD) of selected Ab clones with a titration curve of the fluorescence signals obtained by flow cytometry of yeast cells with different concentrations of the labelled antigen (Gai and Wittrup, 2007), thus avoiding the need of expression and purification of the soluble Ab fragments to have an estimation of their affinity constants.
Yeast display selections often utilize relatively small libraries of ~106–107 clones, either obtained after antigen immunization or by mutagenesis of a pre‐existing clone for affinity maturation procedures (van den Beucken et al., 2003; Colby et al., 2004; Siegel, 2009; Koide and Koide, 2012). Larger Ab libraries (e.g. ~109 clones from naïve origin) have also been constructed and screened for antigen‐binding clones (Yeung and Wittrup, 2002; Feldhaus et al., 2003; Gai and Wittrup, 2007). However, large libraries are more difficult to generate in yeast than in E. coli due to the lower transformation efficiency of yeast cells. Also, yeast cultures require longer times for growth, and reach cellular densities of c. 2 × 107 CFU ml−1 at late exponential phase, which are much lower than those obtained with E. coli cultures (c. 1 × 109 CFU ml−1). Hence, representation of naïve libraries requires growth of large yeast culture volumes and the concentration of yeast cells (Chao et al., 2006). The bigger size of yeast cells than bacteria makes difficult the handling of liquid suspensions concentrated to above 109 CFU ml−1, whereas E. coli is easily concentrated to ~1011 CFU ml−1 (e.g. for electroporation). Therefore, the use of an E. coli display system that could combine the major advantages of the phage and yeast display systems would be highly beneficial.
E. coli display
The outer membrane barrier
Escherichia coli is the most commonly used bacterial host for expression and engineering of Ab fragments, although other bacteria, especially some Gram‐positive species, have been used for display of Ab fragments and Nbs (Krüger et al., 2002; Pant et al., 2006; Kronqvist et al., 2008; Fleetwood et al., 2013). For instance, an immune library of VHHs of c. 107 clones was displayed on Staphylococcus carnosus cell surface fused to the anchoring domain of protein A, and screened by FACS for Nbs binding GFP (Fleetwood et al., 2013). One likely advantage with the use of Gram‐positive bacteria is their rather simple cell envelope with a single cell membrane, which simplifies translocation of the Nbs to the surface. Also, Gram‐positive bacteria have comparative higher tolerance to mechanical stress due to a thicker cell wall, which may result in higher viability during selection and cell sorting. Nonetheless, E. coli is a more suitable microorganism for the cloning, amplification and maintenance of large Nb repertoires owing to its high transformation efficiency, the stability of the cloned DNA in laboratory E. coli strains, its fast and simple growth and the availability of multiple well‐validated protein expression and secretion systems. The larger size of E. coli bacteria than phages allows the direct use of flow cytometry‐based methods for selection, screening and characterization. Also, E. coli bacteria enable higher display levels and are less sticky than phages, which results in lower background levels. Lastly, E. coli bacteria can be grown and concentrated to sufficiently high densities (see above) to represent large naïve and synthetic libraries.
Despite the apparent beneficial properties of E. coli, the development of effective E. coli display systems for Ab/Nb fragments has not been straightforward. This is because E. coli, being a Gram‐negative bacterium, has a cell envelope with two biological membranes: the cytoplasmic inner membrane (IM) and the outer membrane (OM; Bos et al., 2007; Delcour, 2009). These membranes are separated by the periplasmic space, containing a peptidoglycan layer and a variety of transport proteins, enzymes and protein‐folding chaperones that participate, among other processes, in peptidyl‐prolyl cis/trans‐isomerization (e.g. FkpA, SurA) and catalysis of disulfide bridges and their isomerization (e.g. DsbA, DsbC; Kadokura et al., 2003; Merdanovic et al., 2011). The presence of the OM has been the major obstacle hindering the development of effective display methods for Ab selection in E. coli. In order to be displayed on the surface, the polypeptides produced in the cytoplasm have to translocate through the IM, traverse the periplasm, be inserted into the OM and expose the displayed polypeptide region of interest to the extracellular milieu (Dalbey and Kuhn, 2012). Translocation of Nb/Ab fragments to the periplasm using N‐terminal signal peptides of the Sec pathway was solved in phage display vectors (Thie et al., 2008; Tsirigotaki et al., 2017). Once in the periplasm, chaperones such as DsbA, DsbC, FkpA, SurA and Skp assist correct folding, disulfide bond formation and prolyl cis/trans‐isomerization of the Ig domains (Bodelón et al., 2013). It is known that overexpression of these chaperones increases the production of functional Ab fragments and Nbs in the periplasm of E. coli (Schlapschy et al., 2006; Friedrich et al., 2010; Schlapschy and Skerra, 2011; Shriver‐Lake et al., 2017). Hence, the surface display of functional Nb/Ab fragments on E. coli requires an OM protein anchor allowing translocation of folded Ig V domains across the OM from the periplasm.
As an alternative to OM display, a periplasmic display method was developed for the selection of Ab libraries on E. coli, named ‘anchored periplasmic expression’ (APEx; Harvey et al., 2004; Mazor et al., 2007, 2008). In APEx, Ab fragments and full‐length IgGs are expressed in the periplasm of E. coli tethered to the IM with a chimeric lipoprotein anchor. As the Abs in APEx are not actually displayed on the bacterial surface, access of the labelled antigen for FACS requires permeabilization of the OM to generate spheroplasts of bacteria. But spheroplasts are highly sensitive to lysis, thus restricting the selection conditions that can be used, impeding their direct growth on plates after cell sorting and forcing the amplification, subcloning of the selected Ab genes after every selection cycle, all of which lead to increased manipulation.
E. coli display with outer membrane proteins and surface appendages
Some earlier work reported the surface display of scFv fragments on E. coli cells fused to the C‐terminus of the chimeric lipoprotein Lpp‐OmpA’ (Francisco et al., 1993), which consists of the N‐terminal signal peptide and the first nine amino acids of the mature lipoprotein Lpp fused to residues 46–159 of OmpA. This truncated fragment of OmpA comprises the first six β‐strands and a portion of the 7th β‐strand of its eight‐stranded β‐barrel (Pautsch and Schulz, 2000) and therefore lacks the characteristic stability of native β‐barrel outer membrane proteins (OMPs; Koebnik et al., 2000). This fact may have contributed to the reported OM leakage and cellular toxicity of Lpp‐OmpA’ fusions (Georgiou et al., 1996; Stathopoulos et al., 1996), which could induce a significant growth bias of clones in libraries. In fact, the Lpp‐OmpA’ fusion was only used in combination with FACS for affinity maturation of preselected scFv clones displayed on E. coli (Daugherty et al., 1998, 1999), but not for the selection of clones with novel specificities from diverse libraries (either immune or naïve). Native β‐barrel OMPs from E. coli and other Gram‐negative bacteria were also analysed for OM anchoring. Interestingly, periplasmic transport of β‐barrel OMPs uses the same set of periplasmic protein chaperones that participate in the folding of Ig domains (Bos et al., 2007; Bodelón et al., 2013). Porins (e.g. OmpA, OmpC, OmpX, LamB) and protein subunits of fimbrial or flagellar appendages (e.g. FimA, FimH, FliC, FliD) were used as fusion partners for the surface display of peptides and small polypeptides (Lu et al., 1995; Sousa et al., 1996; Georgiou et al., 1997; Klemm and Schembri, 2000; Lee et al., 2003; Bessette et al., 2004; Majander et al., 2005; Rice et al., 2006; Munera et al., 2007), but none of these systems tolerate the transport of folded Ig domains.
E. coli display with autotransporters and the intimin–invasin proteins
The limitations of the above systems led us to explore alternative E. coli display systems for the selection of Nbs (and perhaps other Ab fragments). An important clue came from the study of a large family of proteins secreted by Gram‐negative bacteria called autotransporters (ATs; Henderson et al., 2004; Nicolay et al., 2015). Classical ATs contain within their sequence a C‐terminal β‐barrel, comprising of 12 antiparallel β‐strands and an internal α‐helix (Oomen et al., 2004; van den Berg, 2010; Gawarzewski et al., 2014), which acts as a ‘helper’ domain assisting the translocation of the N‐terminal portion of the polypeptide across the OM. The ‘passenger’ N‐domain is the actual part secreted to the medium and usually fold as long β‐helix rod (Leo et al., 2012; van Ulsen et al., 2014). ATs were originally thought to have self‐translocation capacity across the OM (Pohlner et al., 1987; Henderson et al., 1998), having an internal pore in the β‐barrel for passenger translocation (Klauser et al., 1992; Oomen et al., 2004), or forming a protein translocation channel by oligomerization of β‐barrels (Veiga et al., 2002). However, compelling evidence supports that the β‐barrel of ATs acts as a signal tethering the polypeptide to the conserved β‐barrel assembly machinery (BAM), or to a related translocation and assembly module (TAM) complex (Ieva and Bernstein, 2009; Sauri et al., 2009; Rossiter et al., 2011; Gruss et al., 2013).
The BAM is a large protein complex in the OM that assists the insertion of β‐barrel proteins in the OM (Knowles et al., 2009; Hagan et al., 2011; Han et al., 2016). The BAM complex associates with the amphipathic β‐strands of unfolded OMPs in the periplasm, assisting the folding of the β‐barrel and its simultaneous insertion in the lipid bilayer of the OM (Noinaj et al., 2017). In the case of ATs, the passenger domain appears to be secreted concomitantly with the folding of the β‐barrel (Ieva and Bernstein, 2009), using a translocation pore transiently formed between the β‐barrels of the AT and that of BamA/Omp85 subunit of BAM/TAM complexes (Gruss et al., 2013; Noinaj et al., 2017). Although the large native β‐helix rod passengers of ATs need to have a partially unfolded conformation for their translocation (Jong et al., 2007), we observed that folded Ig domains fused to the β‐barrel of various ATs were efficiently translocated to the surface of E. coli cells (Veiga et al., 2004; Marín et al., 2010). Importantly, an AT‐related family of adhesins, the intimin and invasin family, originally found in enteropathogenic E. coli (EPEC), enterohemorrhagic E. coli (EHEC) and Yersinia pseudotuberculosis and Y. enterocolitica strains, naturally have passenger domains based on repeats of Ig‐like domains that are translocated to the bacterial surface (Tsai et al., 2010; Leo et al., 2015; Sadana et al., 2017). Intimin–invasin proteins are similar to ATs but have an opposite topology, with the N‐terminal signal peptide followed by a 12‐stranded β‐barrel with an internal peptide segment (Fairman et al., 2012) that connects to a surface‐exposed passenger C‐terminal region comprising repeats of Ig‐like and lectin domains (Hamburger, 1999; Kelly et al., 1999; Luo et al., 2000; Sadana et al., 2017). Work from our laboratory demonstrated that the native Ig‐like domains of intimin fold and form a disulfide bridge catalysed by DsbA in the periplasm prior to their translocation (Bodelón et al., 2009). This study also showed that the intimin β‐barrel domain requires the BAM complex for OM insertion, clearly suggesting that they share a similar mechanism of secretion with ATs (Bodelón et al., 2009). The capacity to translocate folded Ig domains by intimin and ATs prompted us to investigate and compare these β‐barrel domains for the surface display and selection of Nb libraries in E. coli (Salema et al., 2013).
Intimin fusions are optimal for E. coli display of Nbs
Heterologous peptides and small polypeptides had been previously displayed on E. coli and other Gram‐negative bacteria by replacing the native passenger domains of ATs and intimin (Klauser et al., 1990; Valls et al., 2000; Wentzel et al., 2001; Jose and Meyer, 2007). For Nb display, we constructed a plasmid vector for cloning the VHHs in frame with the N‐terminal fragment of EHEC intimin (Neae; residues 1–654) comprising the β‐barrel domain and the first Ig‐like domain called D0 (Salema et al., 2013). This vector, called pNeae2, carries the lac promoter (Plac) for induction of the fusion protein with isopropylthio‐β‐D‐galactoside (IPTG), includes restriction sites commonly used in phagemids for the cloning of VHHs (SfiI and NotI), and epitope tags (E‐tag and myc‐tag) flanking the VHH to detect expression of the fusion protein on E. coli surface using commercial mAbs (Fig. 3A and B). We used a similar vector, called pHEA, for fusion of VHHs to the C‐terminal β‐barrel domain of EHEC AT EhaA (C‐EhaA; residues 989–1327), which is well expressed in E. coli (Wells et al., 2008; Marín et al., 2010). Although both types of protein fusions were displayed on E. coli surface, higher display levels of Nbs were obtained with intimin fusions, both when individual clones and an immune library of VHHs were compared (Salema et al., 2013). Similarly, antigen‐binding signals determined by fluorescence flow cytometry of bacteria displaying Nbs of defined specificity were higher with intimin Neae fusions than with AT C‐EhaA fusions. Upon plasmid induction, E. coli bacteria carrying pNeae2‐VHH derivatives displayed ~8000 Nb molecules on their surface. In addition, selection of antigen‐binding clones from immune libraries was more efficient in bacteria displaying intimin Neae fusions than in bacteria displaying C‐EhaA fusions, requiring less number of selection cycles (i.e. two versus four cycles; Salema et al., 2013). Therefore, the Neae fragment of intimin proved to be an effective system for display and selection of Nbs in E. coli, showing higher expression levels and antigen‐binding signals.
Figure 3.
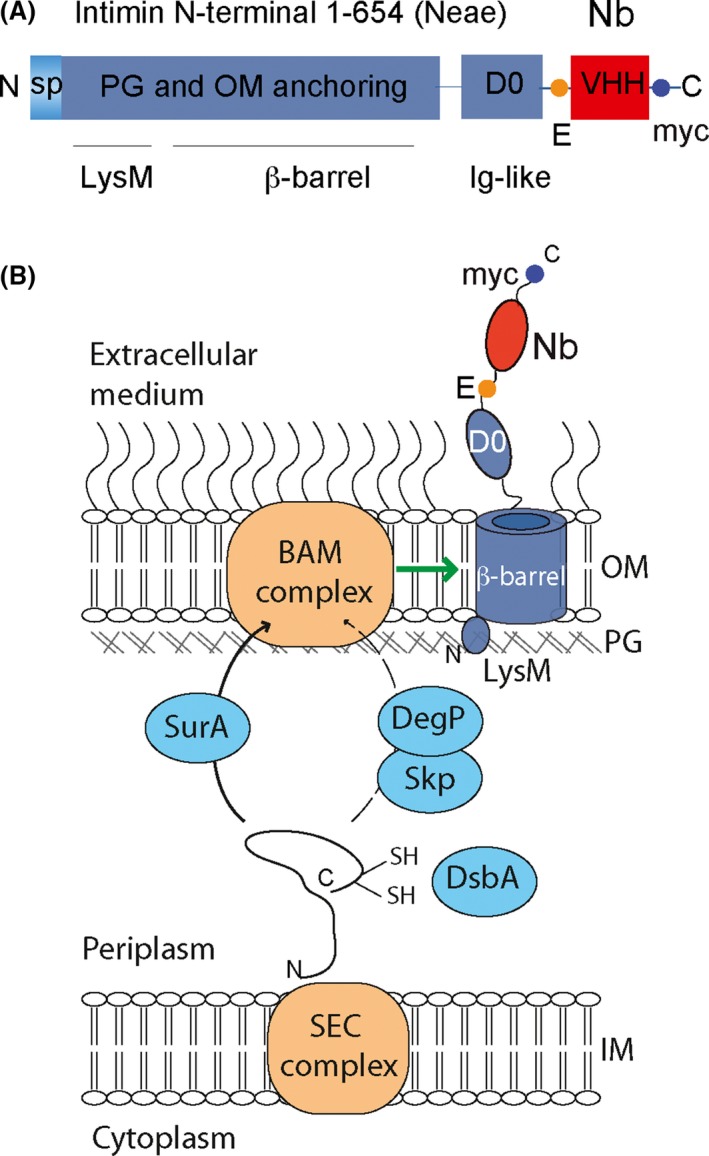
Structure of Neae‐VHH fusions and their secretion mechanism in E. coli display of nanobodies. (A) Linear representation of the polypeptide encoded by Neae‐VHH fusions for E. coli display. The intimin fragment Neae (residues 1–654) comprises the N‐terminal signal peptide (sp), LysM domain for PG binding, β‐barrel for OM‐anchoring and exposed D0 Ig‐like domain. The VHH encodes the Nb. The Neae‐VHH fusion also contains the E‐tag and myc‐tag epitopes flanking the VHH. (B) The fusion polypeptide expressed in the bacterial cytoplasm is translocated across the IM via the Sec complex. In the periplasm, chaperones assist folding of the Nb and transport of the intimin β‐barrel domain to the BAM complex in the OM. The BAM complex is responsible for the folding and insertion of the intimin β‐barrel in the OM and participates in the translocation of the D0 and Nb domains to the bacterial surface. The epitopes E‐ and myc‐tags are used for the immunodetection of Nb displayed on E. coli surface by flow cytometry.
Albeit most E. coli laboratory strains could potentially be used with Neae display, we chose two E. coli K‐12 strains with deletions in the fim operon encoding type 1 fimbriae to avoid the possible interference of these expressed proteinaceous filaments on E. coli surface with the binding of antigens to the OM (Hasman et al., 1999). We have successfully employed the reference K‐12 strain MG1655 (Blattner et al., 1997) with deletion of fimA‐fimH (called AAEC072 or EcM1; Blomfield et al., 1991; Salema et al., 2013), and the high‐efficiency cloning strain DH10B‐T1R that naturally lacks the fim operon (Durfee et al., 2008). Both strains showed similar levels of Nb display on their surface, without any deleterious effects on bacterial growth and viability upon induction.
Selection of Nbs by E. coli display using magnetic cell sorting (MACS)
We have tested the intimin E. coli display system with three different VHH immune libraries, two from dromedaries and one from llamas. Each library was obtained after immunization with antigens of different origin: a purified recombinant bacterial polypeptide (TirM; Salema et al., 2013), purified human fibrinogen (Fib; Salema et al., 2016a) and cells from the human tumour cell line A431 overexpressing the epidermal growth factor receptor (EGFR; Salema et al., 2016a, b). In all cases, the VHH gene segments were amplified from lymphocytes isolated from peripheral blood samples and cloned in frame with the Neae fragment in pNeae2 vector to obtain ~107 independent clones (Fig. 4A).
Figure 4.
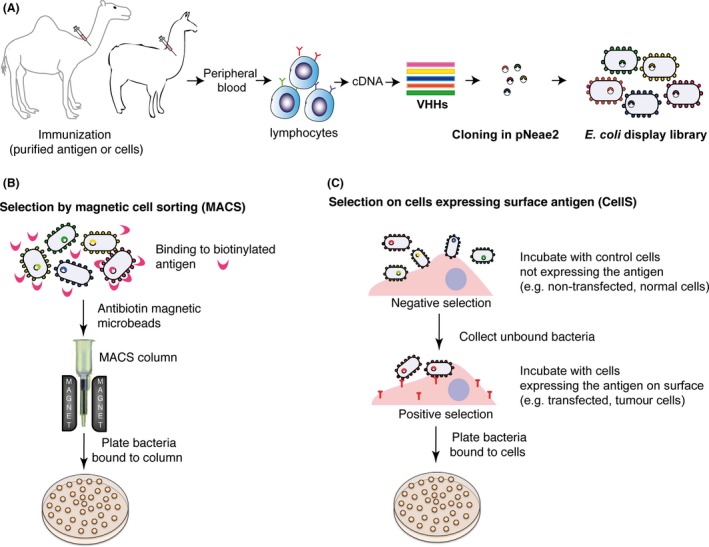
Generation of E. coli display immune libraries and their selection by magnetic cell sorting (MACS) and on mammalian cells (CellS). (A) Scheme of the generation of an E. coli display immune library of Nbs. Animals (e.g. dromedaries or llamas) are immunized with either purified antigen or cells expressing the target antigen. After immunization, the VHH gene segments are amplified from peripheral blood lymphocytes by RT‐PCR, cloned into plasmid vector pNeae2 and induced on E. coli for surface display. (B) Selection of an E. coli display library using MACS. Escherichia coli bacteria are incubated with the biotinylated antigen and antibiotin magnetic beads. Bacteria with bound antigen are captured in an iron column held in a magnet. Elution of bound bacteria is carried out with fresh LB media upon column removal from the magnet and grown by plating. (C) Selection of an E. coli display library on mammalian cells (CellS). Escherichia coli bacteria are initially incubated with control cells lacking expression of the target antigen (negative selection). Bacteria that do not bind to control cells are incubated with cells expressing the target antigen on their surface for positive selection. Bacteria bound to antigen‐positive cells after washing (e.g. PBS) are recovered by lysis of the mammalian cells and grown by plating.
For the selection of antigen‐specific Nbs, we used magnetic cell sorting (MACS) of E. coli bacteria incubated with the corresponding purified antigens labelled with biotin: TirM, Fib and eEGFR‐Fc (the extracellular domain of EGFR fused to human Fc). The MACS procedure for selection is illustrated in Fig. 4B. Induced bacteria are incubated with the biotinylated antigen, washed (e.g. with PBS) to remove the unbound antigen and incubated with a suspension of antibiotin paramagnetic microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). Next, the mixture is passed through a small ferromagnetic column (mini‐MACS MS; Miltenyi Biotec) held in a magnetic holder, which retains E. coli bacteria coated with bound biotinylated antigen and antibiotin microbeads, whereas bacteria with no antigen on their surface are washed out of the column. Elution of bound bacteria is carried out with fresh culture media (e.g. LB) upon removal of the column from the magnet. All steps were carried out at room temperature using mild buffers like PBS and LB, which were sufficient to wash out most non‐specific binders and isolate antigen‐specific clones. Nonetheless, more stringent washing conditions and buffers containing detergents (e.g. Triton X‐100, SDS or deoxycholate) could also be used as E. coli with an intact OM can grow in the presence of these detergents (Delcour, 2009).
We chose MACS versus FACS to isolate E. coli bacteria bound to the antigen for practical reasons. First, MACS does not require expensive laboratory equipment like the fluorescent cell sorter, facilitating its implementation in most laboratories. Also, processing of libraries is faster in MACS than in FACS, and multiple samples can be run in parallel (e.g. selection of a single library with various antigens, including negative control antigens). In our experiments, we employed a manual system holding up to 8 columns (OctoMACS; Miltenyi Biotec) each with a capacity of at least ~2 × 108 bacteria. Given the size of immune libraries, it was sufficient to load in ~2 × 108 CFU of E. coli incubated with the biotinylated antigen (at concentrations from 50 to 200 nM), equivalent to ~0.2 ml of an induced culture at OD600 ~1, to have > 10× representation of the immune libraries. Nonetheless, MACS can be automated and is scalable to columns with higher loading capacities (e.g. 2 × 109 bacteria for MACS LS columns; Miltenyi Biotec), thus making screening of large Nb libraries (e.g. naïve) faster and more efficient than FACS. The use of MACS for selections of large naïve scFv libraries by yeast display has also been described (Chao et al., 2006).
The number of bacteria eluted from the MACS column in the washed and antigen‐bound fractions can be determined by plating on LB agar as CFU, which provide a quantitative estimation of the enrichment in antigen‐binding clones in every selection cycle. Bacterial colonies grown from the antigen‐bound fraction can be harvested and be frozen for long‐term storage, grown for fluorescence flow cytometry analysis to determine antigen‐binding (e.g. for biotin‐labelled antigens using streptavidin–phycoerythrin, PE) and Nb display levels (e.g. with anti‐myc mAb and fluorophore‐labelled anti‐IgG mouse antibodies) and, if needed, for further cycles of MACS selection. In our experience, the number of cycles required for the isolation of high‐affinity Nb clones varies depending on the library, but usually two rounds of MACS biopanning are sufficient to observe a significant enrichment of antigen‐specific binders from immune libraries allowing isolation of high‐affinity Nb clones with KDs in the low nanomolar and subnanomolar range (i.e. 0.2–20 nM; Salema et al., 2013, 2016a,b). Upon MACS biopannings, isolation of E. coli clones binding the antigen is performed by flow cytometry screening of individual colonies (e.g. 96) randomly picked from the antigen‐bound fraction of the last MACS selection round. The use of flow cytometry for the characterization of Nbs displayed on E. coli is discussed later in the text. Lastly, although we have not observed significant differences in growth or loss of viability of E. coli bacteria carrying pNeae2 derivatives, we used mild induction conditions (i.e. 0.05 mM IPTG at 30 °C) to minimize potential toxicity effects of Neae‐VHH fusion overexpression from this multicopy vector, which may cause a growth bias of specific clones within the libraries. Similarly, to reduce the potential outgrowth of clones with lower levels of Neae‐VHH expression after each induction–selection cycle, we made preparations of the plasmid pool from antigen‐bound bacteria and used it to transform fresh E. coli bacteria for the next round of induction–selection cycle.
Selection of Nbs by E. coli display using selection on live cells (CellS)
Given the potential of Nbs for tumour therapy and in vivo tumour imaging, we evaluated E. coli display for the direct selection on live cells of Nbs against tumour‐associated cell surface antigens (Scott et al., 2012; Tumeh et al., 2014; Postow et al., 2015; Redman et al., 2015). In particular, we performed selection on live cells of Nbs binding human EGFR as a model tumour cell surface antigen (Salema et al., 2016a, b). Although MACS is a convenient method to select high‐affinity binders, it relies on the availability of purified antigen, which is not always practical or feasible. Some antigens (e.g. membrane proteins exposed on the surface of tumour cells) may be produced at low yields, have low solubility or may not maintain their native conformation upon purification, all of which limits the construction of immune libraries and the selection strategy. Also, in vitro conditions with purified proteins may alter conformational epitopes that could be relevant in vivo, leading to selection of Nbs that may not recognize the native antigen on the tumour cell surface. In these situations, it is advantageous to screen Nb libraries directly on live intact tumour cells expressing the antigen, either endogenously or upon transfection. Cell selection (CellS) is also an attractive approach to identify Nbs against novel antigens expressed on the surface of target cells. In the case of phage display, CellS requires experimental optimization to minimize the background binding of phage particles to cells. The more simple phage biopanning procedure involves a depletion step on cells lacking expression of the target antigen, to remove non‐specific phage binders, followed by incubation of the unbound particles with target cells expressing the antigen of interest (Hoogenboom et al., 1999; Liu et al., 2004; Siva et al., 2008). Nonetheless, this simple depletion strategy is usually inefficient with phages and further steps are needed, such as competitive elution with a ligand or mAbs (Figini et al., 1998; Beiboer et al., 2000; Klimka et al., 2000; Veggiani et al., 2011), centrifugation of cells through an organic phase (Lipes et al., 2008) or masking non‐relevant cell epitopes with soluble Nb fragments (Even‐Desrumeaux et al., 2014).
We employed a whole cell‐based experimental strategy (Fig. 4C), from tumour cell immunization to on‐cell selection taking advantage of the excellent specificity of E. coli bacteria displaying Nbs to target cells with surface antigens (Piñero‐Lambea et al., 2015). We used a library of VHHs obtained by immunizing llamas with A431 human tumour cells that overexpress EGFR (Roovers et al., 2007). The VHHs were cloned into pNeae2, induced in E. coli bacteria and clones binding EGFR were selected directly on cells using a simple depletion strategy, by combination of a negative selection step on a mouse cell line that does not express human EGFR (named NIH‐3T3 2.2), followed by a positive selection step on cells of the same cell line but transfected to express EGFR (named HER14; Fig. 4C). To screen for binders, we examined by bright‐field light microscopy the adhesion of 96 clones, picked from the second round of CellS, to Her14 (EGFR+) and NIH‐3T3 2.2 (EGFR‐) cells. Clones binding specifically Her14 cells were selected, identifying high‐affinity Nbs that bind specifically EGFR‐Fc by flow cytometry (Salema et al., 2016a, b). As high‐affinity binders were predominantly selected during the screening, this indicates that the high levels of display of Nbs on E. coli surface and of the target EGFR antigen on the plasma cell membrane of the transfected cell line had no significant avidity effect, which could have resulted in the frequent isolation of low‐affinity binders. The efficiency of selection by E. coli display using a simple subtractive CellS suggests that E. coli bacteria may have less ‘non‐specific’ binding to cells than phage particles.
Fluorescence flow cytometry for characterization of Nbs
The use of fluorescence flow cytometry for the determination of expression levels, antigen binding and estimation of the affinity of selected Nbs is a major advantage of the E. coli display system (Fig. 5). Libraries, bacterial pools isolated after selection and individual clones can all be analysed by fluorescence flow cytometry with the anti‐myc‐tag mAb and secondary anti‐mouse IgG antibodies labelled with a fluorophore (e.g. Alexa 488) for the determination of expression/display levels of the Nb on E. coli bacteria. Similarly, bacteria can be incubated with different concentrations of the biotinylated antigen used in the selections, or a negative control antigen (e.g. biotinylated BSA), and secondary‐labelled streptavidin (e.g. streptavidin–PE) for the determination of antigen binding and specificity (Fig. 5; Top panel; Salema et al., 2013, 2016a,b). Nb display and antigen binding can be analysed simultaneously (double staining) or independently (single staining). Specific binders are identified by flow cytometry screening of individual colonies grown after MACS or CellS. Remarkably, flow cytometry can also be used to analyse binding competition of Nbs with ligands (e.g. EGF ligand EGFR), allowing identification of Nbs that compete (or do not compete) with a ligand (Fig. 5; central panel; Salema et al., 2016a, b).
Figure 5.
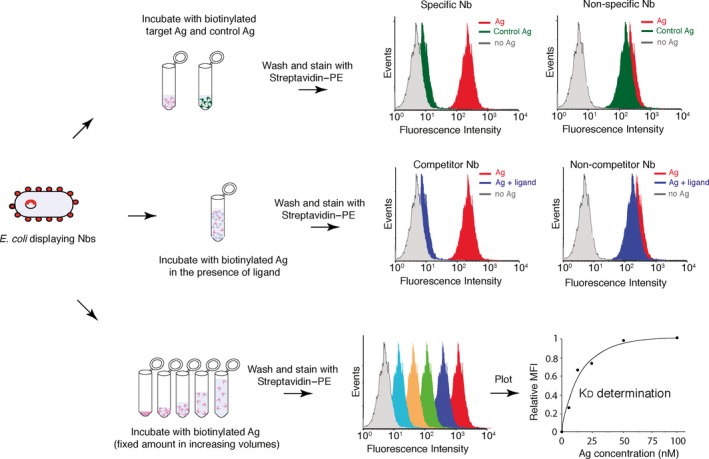
Fluorescence flow cytometry analysis for the characterization of the antigen‐binding properties of nanobodies displayed on E. coli. Scheme showing different applications of fluorescence flow cytometry for the characterization of the antigen (Ag)‐binding properties of Nbs displayed on E. coli. Top: incubation of bacteria with target and control Ags (e.g. labelled with biotin) to determine binding specificity. Central: incubation of bacteria with labelled Ag in the presence or absence of a ligand (i.e. a molecule binding the Ag) for the identification of Nbs competing or not competing with ligand binding. Bottom: incubation of bacteria with different concentrations of the labelled Ag for the estimation of equilibrium dissociation constant (KD) by plotting Ag concentrations versus mean fluorescence intensity (MFI) values. All examples shown use Streptavidin–phycoerythrin (PE) for fluorescent staining of bacteria with bound biotinylated Ag.
Lastly, this methodology can also be applied to estimate the apparent KD of the Nbs prior to their purification using antigen binding under equilibrium conditions (Fig. 5; bottom panel). This approach has been used to estimate the apparent KD of scFvs on the surface of yeast and demonstrated to be similar to KD values obtained with purified scFv by surface plasmon resonance (SPR; Boder and Wittrup, 2000; Feldhaus et al., 2003). We have demonstrated that flow cytometry analysis of E. coli bacteria displaying Neae‐VHH fusions gives a good estimation of the apparent KD of Nbs, with values similar to those obtained by SPR with purified Nbs (Salema et al., 2013, 2016a,b). A fixed amount of bacteria displaying NVHH fusions (~3 × 107 CFU; ~2 × 1011 Nb molecules) are incubated with a fixed amount of biotinylated antigen (~2 pmols; ~1 × 1012 antigen molecules) in twofold increasing volumes (e.g. from 0.1 to 10 ml of PBS to reach a final concentration ranging from 20 to 0.2 nM). After 90‐min incubation, bacteria are washed, labelled with Streptavidin–PE and analysed in a flow cytometer. The relative mean fluorescence intensities (MFI) obtained at the different concentrations of antigen are plotted in a curve and the antigen concentration giving 50% MFI is extrapolated to estimate the apparent KD of the Nb.
Conclusions and future directions
E. coli display based on the N‐terminal domain of intimin (Neae) is an effective platform for the surface display of VHH libraries on E. coli, allowing selection of high‐affinity Nbs using purified biotinylated antigen with MACS, and on cell selection (CellS) with live mammalian cells displaying the target antigen on their surface. In some circumstances, it might be useful to use FACS in combination with MACS and/or CellS to improve recovery of high‐affinity binders with high level of expression at later stages of the selection, when library diversity is reduced. So far, we have demonstrated that E. coli display is suitable for use with Nb immune libraries containing ~107 clones, but larger naïve and synthetic libraries of ~109 clones could also be used as MACS and CellS can be easily scaled up to these bacterial numbers, and suspensions of E. coli bacteria at densities of ~1010–1011 CFU/ml are handled easily. Escherichia coli bacteria with intact OMs can be washed with most common buffers as well as tolerate significant concentrations of detergents (e.g. Triton X‐100, SDS), a marked improvement over the washing conditions tolerated by E. coli spheroplasts. The multivalent display and less sticky properties of E. coli bacteria compared with bacteriophages facilitates their selection on complex antigenic surfaces, e.g. mammalian cells, tissues and organs with reduced background. Flow cytometry analysis of bacteria is also a major advantage of E. coli display, allowing monitoring of the selection process, identification of binders and characterization of their antigen‐binding properties, such as specificity, ligand competition and determination of KD. Escherichia coli display is likely to be effective not only with camelid VHHs, but also other single domain Abs with similar structural properties, like VNARs from sharks (Dooley et al., 2003) and those based on the engineering of human VHs and VLs (van den Beucken et al., 2001; Holt et al., 2003, 2008; Ignatovich et al., 2012). The system might be also suitable for use with larger Ab fragments based on a single polypeptide, like scFvs (Bruce et al., 2016), although the tendency of some scFvs to aggregate may hinder their translocation across the OM (Veiga et al., 2004). Nonetheless, this property could be advantageous to ‘filter out’ more stable and soluble scFv clones from scFv libraries. Future work will be aimed to demonstrate these potential benefits and applications of E. coli display.
Conflict of Interest
The authors declare that they have no conflict of interest.
Acknowledgements
We are grateful to the ‘La Caixa Foundation’ for the support of VS PhD contract.
Microbial Biotechnology (2017) 10(6), 1468–1484
Funding Information
Work in the laboratory of LAF is supported by research grants from the Spanish Ministerio de Economía y Competitividad (BIO2014‐60305R) and the European Research Council (ERC‐2012‐ADG_20120314).
References
- Arbabi Ghahroudi, M. , Desmyter, A. , Wyns, L. , Hamers, R. , and Muyldermans, S. (1997) Selection and identification of single domain antibody fragments from camel heavy‐chain antibodies. FEBS Lett 414: 521–526. [DOI] [PubMed] [Google Scholar]
- Arbabi‐Ghahroudi, M. , Tanha, J. , and MacKenzie, R. (2005) Prokaryotic expression of antibodies. Cancer Metastasis Rev 24: 501–519. [DOI] [PubMed] [Google Scholar]
- Beiboer, S.H. , Reurs, A. , Roovers, R.C. , Arends, J.W. , Whitelegg, N.R. , Rees, A.R. , and Hoogenboom, H.R. (2000) Guided selection of a pan carcinoma specific antibody reveals similar binding characteristics yet structural divergence between the original murine antibody and its human equivalent. J Mol Biol 296: 833–849. [DOI] [PubMed] [Google Scholar]
- van den Berg, B. (2010) Crystal structure of a full‐length autotransporter. J Mol Biol 396: 627–633. [DOI] [PubMed] [Google Scholar]
- Bessette, P.H. , Rice, J.J. , and Daugherty, P.S. (2004) Rapid isolation of high‐affinity protein binding peptides using bacterial display. Protein Eng Des Sel 17: 731–739. [DOI] [PubMed] [Google Scholar]
- Bethuyne, J. , De Gieter, S. , Zwaenepoel, O. , Garcia‐Pino, A. , Durinck, K. , Verhelle, A. , et al (2014) A nanobody modulates the p53 transcriptional program without perturbing its functional architecture. Nucleic Acids Res 42: 12928–12938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Beucken, T. , van Neer, N. , Sablon, E. , Desmet, J. , Celis, L. , Hoogenboom, H.R. , and Hufton, S.E. (2001) Building novel binding ligands to B7.1 and B7.2 based on human antibody single variable light chain domains. J Mol Biol 310: 591–601. [DOI] [PubMed] [Google Scholar]
- van den Beucken, T. , Pieters, H. , Steukers, M. , van der Vaart, M. , Ladner, R.C. , Hoogenboom, H.R. , and Hufton, S.E. (2003) Affinity maturation of Fab antibody fragments by fluorescent‐activated cell sorting of yeast‐displayed libraries. FEBS Lett 546: 288–294. [DOI] [PubMed] [Google Scholar]
- Blanco‐Toribio, A. , Muyldermans, S. , Frankel, G. , and Fernández, L.A. (2010) Direct injection of functional single‐domain antibodies from E. coli into human cells. PLoS One 5: e15227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blattner, F.R. , Plunkett, G. 3rd , Bloch, C.A. , Perna, N.T. , Burland, V. , Riley, M. , et al (1997) The complete genome sequence of Escherichia coli K‐12. Science 277: 1453–1474. [DOI] [PubMed] [Google Scholar]
- Blomfield, I.C. , McClain, M.S. , and Eisenstein, B.I. (1991) Type 1 fimbriae mutants of Escherichia coli K12: characterization of recognized afimbriate strains and construction of new fim deletion mutants. Mol Microbiol 5: 1439–1445. [DOI] [PubMed] [Google Scholar]
- Bodelón, G. , Marín, E. , and Fernández, L.A. (2009) Role of periplasmic chaperones and BamA (YaeT/Omp85) in folding and secretion of intimin from enteropathogenic Escherichia coli strains. J Bacteriol 191: 5169–5179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodelón, G. , Palomino, C. , and Fernández, L.A. (2013) Immunoglobulin domains in Escherichia coli and other enterobacteria: from pathogenesis to applications in antibody technologies. FEMS Microbiol Rev 37: 204–250. [DOI] [PubMed] [Google Scholar]
- Boder, E.T. , and Wittrup, K.D. (1997) Yeast surface display for screening combinatorial polypeptide libraries. Nat Biotechnol 15: 553–557. [DOI] [PubMed] [Google Scholar]
- Boder, E.T. , and Wittrup, K.D. (2000) Yeast surface display for directed evolution of protein expression, affinity, and stability. Methods Enzymol 328: 430–444. [DOI] [PubMed] [Google Scholar]
- Bos, M.P. , Robert, V. , and Tommassen, J. (2007) Biogenesis of the gram‐negative bacterial outer membrane. Annu Rev Microbiol 61: 191–214. [DOI] [PubMed] [Google Scholar]
- Bouchet, J. , Basmaciogullari, S.E. , Chrobak, P. , Stolp, B. , Bouchard, N. , Fackler, O.T. , et al (2011) Inhibition of the Nef regulatory protein of HIV‐1 by a single‐domain antibody. Blood 117: 3559–3568. [DOI] [PubMed] [Google Scholar]
- Bruce, V.J. , Ta, A.N. , and McNaughton, B.R. (2016) Minimalist antibodies and mimetics: an update and recent applications. ChemBioChem 17: 1892–1899. [DOI] [PubMed] [Google Scholar]
- Chakravarty, R. , Goel, S. , and Cai, W. (2014) Nanobody: the “magic bullet” for molecular imaging? Theranostics 4: 386–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao, G. , Lau, W.L. , Hackel, B.J. , Sazinsky, S.L. , Lippow, S.M. , and Wittrup, K.D. (2006) Isolating and engineering human antibodies using yeast surface display. Nat Protoc 1: 755–768. [DOI] [PubMed] [Google Scholar]
- Colby, D.W. , Kellogg, B.A. , Graff, C.P. , Yeung, Y.A. , Swers, J.S. , and Wittrup, K.D. (2004) Engineering antibody affinity by yeast surface display. Methods Enzymol 388: 348–358. [DOI] [PubMed] [Google Scholar]
- Conrath, K.E. , Lauwereys, M. , Galleni, M. , Matagne, A. , Frere, J.M. , Kinne, J. , et al (2001) Beta‐lactamase inhibitors derived from single‐domain antibody fragments elicited in the Camelidae. Antimicrob Agents Chemother 45: 2807–2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalbey, R.E. , and Kuhn, A. (2012) Protein Traffic in Gram‐negative bacteria – how exported and secreted proteins find their way. FEMS Microbiol Rev 36: 1023–1045. [DOI] [PubMed] [Google Scholar]
- Daugherty, P.S. , Chen, G. , Olsen, M.J. , Iverson, B.L. , and Georgiou, G. (1998) Antibody affinity maturation using bacterial surface display. Protein Eng 11: 825–832. [DOI] [PubMed] [Google Scholar]
- Daugherty, P.S. , Olsen, M.J. , Iverson, B.L. , and Georgiou, G. (1999) Development of an optimized expression system for the screening of antibody libraries displayed on the Escherichia coli surface. Protein Eng 12: 613–621. [DOI] [PubMed] [Google Scholar]
- De Meyer, T. , Laukens, B. , Nolf, J. , Van Lerberge, E. , De Rycke, R. , De Beuckelaer, A. , et al (2015) Comparison of VHH‐Fc antibody production in Arabidopsis thaliana, Nicotiana benthamiana and Pichia pastoris . Plant Biotechnol J 13: 938–947. [DOI] [PubMed] [Google Scholar]
- De Schutter, K. , and Callewaert, N. (2012) Pichia surface display: a tool for screening single domain antibodies. Methods Mol Biol 911: 125–134. [DOI] [PubMed] [Google Scholar]
- Delcour, A.H. (2009) Outer membrane permeability and antibiotic resistance. Biochem Biophys Acta 1794: 808–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dooley, H. , Flajnik, M.F. , and Porter, A.J. (2003) Selection and characterization of naturally occurring single‐domain (IgNAR) antibody fragments from immunized sharks by phage display. Mol Immunol 40: 25–33. [DOI] [PubMed] [Google Scholar]
- Dumoulin, M. , Conrath, K. , Van Meirhaeghe, A. , Meersman, F. , Heremans, K. , Frenken, L.G. , et al (2002) Single‐domain antibody fragments with high conformational stability. Protein Sci 11: 500–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durfee, T. , Nelson, R. , Baldwin, S. , Plunkett, G. 3rd , Burland, V. , Mau, B. , et al (2008) The complete genome sequence of Escherichia coli DH10B: insights into the biology of a laboratory workhorse. J Bacteriol 190: 2597–2606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Even‐Desrumeaux, K. , Nevoltris, D. , Lavaut, M.N. , Alim, K. , Borg, J.P. , Audebert, S. , et al (2014) Masked selection: a straightforward and flexible approach for the selection of binders against specific epitopes and differentially expressed proteins by phage display. Mol Cell Proteomics 13: 653–665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairman, J.W. , Dautin, N. , Wojtowicz, D. , Liu, W. , Noinaj, N. , Barnard, T.J. , et al (2012) Crystal structures of the outer membrane domain of intimin and invasin from enterohemorrhagic E. coli and enteropathogenic Y. pseudotuberculosis . Structure 20: 1233–1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldhaus, M.J. , and Siegel, R.W. (2004) Yeast display of antibody fragments: a discovery and characterization platform. J Immunol Methods 290: 69–80. [DOI] [PubMed] [Google Scholar]
- Feldhaus, M.J. , Siegel, R.W. , Opresko, L.K. , Coleman, J.R. , Feldhaus, J.M. , Yeung, Y.A. , et al (2003) Flow‐cytometric isolation of human antibodies from a nonimmune Saccharomyces cerevisiae surface display library. Nat Biotechnol 21: 163–170. [DOI] [PubMed] [Google Scholar]
- Figini, M. , Obici, L. , Mezzanzanica, D. , Griffiths, A. , Colnaghi, M.I. , Winter, G. , and Canevari, S. (1998) Panning phage antibody libraries on cells: isolation of human Fab fragments against ovarian carcinoma using guided selection. Cancer Res 58: 991–996. [PubMed] [Google Scholar]
- Flajnik, M.F. , Deschacht, N. , and Muyldermans, S. (2011) A case of convergence: why did a simple alternative to canonical antibodies arise in sharks and camels? PLoS Biol 9: e1001120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleetwood, F. , Devoogdt, N. , Pellis, M. , Wernery, U. , Muyldermans, S. , Ståhl, S. , and Löfblom, J. (2013) Surface display of a single‐domain antibody library on Gram‐positive bacteria. Cell Mol Life Sci 70: 1081–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francisco, J.A. , Campbell, R. , Iverson, B.L. , and Georgiou, G. (1993) Production and fluorescence‐activated cell sorting of Escherichia coli expressing a functional antibody fragment on the external surface. Proc Natl Acad Sci USA 90: 10444–10448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frenken, L.G.J. , van der Linden, R.H.J. , Hermans, P.W.J.J. , Bos, J.W. , Ruuls, R.C. , de Geus, B. , and Verrips, C.T. (2000) Isolation of antigen specific llama VHH antibody fragments and their high level secretion by Saccharomyces cerevisiae . J Biotechnol 78: 11–21. [DOI] [PubMed] [Google Scholar]
- Frenzel, A. , Schirrmann, T. , and Hust, M. (2016) Phage display‐derived human antibodies in clinical development and therapy. MAbs 8: 1177–1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedrich, L. , Stangl, S. , Hahne, H. , Kuster, B. , Kohler, P. , Multhoff, G. , and Skerra, A. (2010) Bacterial production and functional characterization of the Fab fragment of the murine IgG1/lambda monoclonal antibody cmHsp70.1, a reagent for tumour diagnostics. Protein Eng Des Sel 23: 161–168. [DOI] [PubMed] [Google Scholar]
- Fulcher, L.J. , Macartney, T. , Bozatzi, P. , Hornberger, A. , Rojas‐Fernandez, A. , and Sapkota, G.P. (2016) An affinity‐directed protein missile system for targeted proteolysis. Open Biol 6: 160255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gai, S.A. , and Wittrup, K.D. (2007) Yeast surface display for protein engineering and characterization. Curr Opin Struct Biol 17: 467–473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galan, A. , Comor, L. , Horvatic, A. , Kules, J. , Guillemin, N. , Mrljak, V. , and Bhide, M. (2016) Library‐based display technologies: where do we stand? Mol BioSyst 12: 2342–2358. [DOI] [PubMed] [Google Scholar]
- Gawarzewski, I. , DiMaio, F. , Winterer, E. , Tschapek, B. , Smits, S.H. , Jose, J. , and Schmitt, L. (2014) Crystal structure of the transport unit of the autotransporter adhesin involved in diffuse adherence from Escherichia coli . J Struct Biol 187: 20–29. [DOI] [PubMed] [Google Scholar]
- Georgiou, G. , Stephens, D.L. , Stathopoulos, C. , Poetschke, H.L. , Mendenhall, J. , and Earhart, C.F. (1996) Display of beta‐lactamase on the Escherichia coli surface: outer membrane phenotypes conferred by Lpp’‐OmpA’‐beta‐lactamase fusions. Protein Eng 9: 239–247. [DOI] [PubMed] [Google Scholar]
- Georgiou, G. , Stathopoulos, C. , Daugherty, P.S. , Nayak, A.R. , Iverson, B.L. , and Curtiss, R. 3rd (1997) Display of heterologous proteins on the surface of microorganisms: from the screening of combinatorial libraries to live recombinant vaccines. Nat Biotechnol 15: 29–34. [DOI] [PubMed] [Google Scholar]
- Govaert, J. , Pellis, M. , Deschacht, N. , Vincke, C. , Conrath, K. , Muyldermans, S. , and Saerens, D. (2012) Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. J Biol Chem 287: 1970–1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gruss, F. , Zähringer, F. , Jakob, R.P. , Burmann, B.M. , Hiller, S. , and Maier, T. (2013) The structural basis of autotransporter translocation by TamA. Nat Struct Mol Biol 20: 1318–1320. [DOI] [PubMed] [Google Scholar]
- Hagan, C.L. , Silhavy, T.J. , and Kahne, D.E. (2011) b‐Barrel membrane protein assembly by the Bam complex. Annu Rev Biochem 80: 189–210. [DOI] [PubMed] [Google Scholar]
- Hamburger, Z.A. (1999) Crystal structure of invasin: a bacterial integrin‐binding protein. Science 286: 291–295. [DOI] [PubMed] [Google Scholar]
- Hamers‐Casterman, C. , Atarhouch, T. , Muyldermans, S. , Robinson, G. , Hamers, C. , Songa, E.B. , et al (1993) Naturally occurring antibodies devoid of light chains. Nature 363: 446–448. [DOI] [PubMed] [Google Scholar]
- Han, L. , Zheng, J. , Wang, Y. , Yang, X. , Liu, Y. , Sun, C. , et al (2016) Structure of the BAM complex and its implications for biogenesis of outer‐membrane proteins. Nat Struct Mol Biol 23: 192–196. [DOI] [PubMed] [Google Scholar]
- Harvey, B.R. , Georgiou, G. , Hayhurst, A. , Jeong, K.J. , Iverson, B.L. , and Rogers, G.K. (2004) Anchored periplasmic expression, a versatile technology for the isolation of high‐affinity antibodies from Escherichia coli‐expressed libraries. Proc Natl Acad Sci USA 101: 9193–9198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasman, H. , Chakraborty, T. , and Klemm, P. (1999) Antigen‐43‐mediated autoaggregation of Escherichia coli is blocked by fimbriation. J Bacteriol 181: 4834–4841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassaine, G. , Deluz, C. , Grasso, L. , Wyss, R. , Tol, M.B. , Hovius, R. , et al (2014) X‐ray structure of the mouse serotonin 5‐HT3 receptor. Nature 512: 276–281. [DOI] [PubMed] [Google Scholar]
- Hassanzadeh‐Ghassabeh, G. , Saerens, D. , and Muyldermans, S. (2011) Generation of anti‐infectome/anti‐proteome nanobodies. Methods Mol Biol 790: 239–259. [DOI] [PubMed] [Google Scholar]
- Helma, J. , Cardoso, M.C. , Muyldermans, S. , and Leonhardt, H. (2015) Nanobodies and recombinant binders in cell biology. J Cell Biol 209: 633–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson, I.R. , Navarro‐Garcia, F. , and Nataro, J.P. (1998) The great escape: structure and function of the autotransporter proteins. Trends Microbiol 6: 370–378. [DOI] [PubMed] [Google Scholar]
- Henderson, I.R. , Navarro‐Garcia, F. , Desvaux, M. , Fernandez, R.C. and Ala'Aldeen, D. (2004) Type V protein secretion pathway: the autotransporter story. Microbiol Mol Biol Rev 68, 692–744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heras, B. , Shouldice, S.R. , Totsika, M. , Scanlon, M.J. , Schembri, M.A. , and Martin, J.L. (2009) DSB proteins and bacterial pathogenicity. Nat Rev Microbiol 7: 215–225. [DOI] [PubMed] [Google Scholar]
- Holliger, P. (2002) Expression of antibody fragments in Pichia pastoris . Methods Mol Biol 178: 349–357. [DOI] [PubMed] [Google Scholar]
- Holt, L.J. , Herring, C. , Jespers, L.S. , Woolven, B.P. , and Tomlinson, I.M. (2003) Domain antibodies: proteins for therapy. Trends Biotechnol 21: 484–490. [DOI] [PubMed] [Google Scholar]
- Holt, L.J. , Basran, A. , Jones, K. , Chorlton, J. , Jespers, L.S. , Brewis, N.D. , and Tomlinson, I.M. (2008) Anti‐serum albumin domain antibodies for extending the half‐lives of short lived drugs. Protein Eng Des Sel 21: 283–288. [DOI] [PubMed] [Google Scholar]
- Hoogenboom, H.R. (2002) Overview of antibody phage‐display technology and its applications. Methods Mol Biol 178: 1–37. [DOI] [PubMed] [Google Scholar]
- Hoogenboom, H.R. (2005) Selecting and screening recombinant antibody libraries. Nat Biotechnol 23: 1105–1116. [DOI] [PubMed] [Google Scholar]
- Hoogenboom, H.R. , Lutgerink, J.T. , Pelsers, M.M. , Rousch, M.J. , Coote, J. , Van Neer, N. , et al (1999) Selection‐dominant and nonaccessible epitopes on cell‐surface receptors revealed by cell‐panning with a large phage antibody library. Eur J Biochem 260: 774–784. [DOI] [PubMed] [Google Scholar]
- Ieva, R. , and Bernstein, H.D. (2009) Interaction of an autotransporter passenger domain with BamA during its translocation across the bacterial outer membrane. Proc Natl Acad Sci USA 106: 19120–19125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ignatovich, O. , Jespers, L. , Tomlinson, I.M. , and de Wildt, R.M. (2012) Creation of the large and highly functional synthetic repertoire of human VH and Vkappa domain antibodies. Methods Mol Biol 911: 39–63. [DOI] [PubMed] [Google Scholar]
- Jong, W.S.P. , ten Hagen‐Jongman, C.M. , den Blaauwen, T. , Jan Slotboom, D. , Tame, J.R.H. , Wickstrom, D. , et al (2007) Limited tolerance towards folded elements during secretion of the autotransporter Hbp. Mol Microbiol 63: 1524–1536. [DOI] [PubMed] [Google Scholar]
- Jose, J. , and Meyer, T.F. (2007) The autodisplay story, from discovery to biotechnical and biomedical applications. Microbiol Mol Biol Rev 71: 600–619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadokura, H. , Katzen, F. , and Beckwith, J. (2003) Protein disulfide bond formation in prokaryotes. Annu Rev Biochem 72: 111–135. [DOI] [PubMed] [Google Scholar]
- Kazemi‐Lomedasht, F. , Behdani, M. , Bagheri, K.P. , Habibi‐Anbouhi, M. , Abolhassani, M. , Arezumand, R. , et al (2015) Inhibition of angiogenesis in human endothelial cell using VEGF specific nanobody. Mol Immunol 65: 58–67. [DOI] [PubMed] [Google Scholar]
- Kelly, G. , Prasannan, S. , Daniell, S. , Fleming, K. , Frankel, G. , Dougan, G. , et al (1999) Structure of the cell‐adhesion fragment of intimin from enteropathogenic Escherichia coli . Nat Struct Biol 6: 313–318. [DOI] [PubMed] [Google Scholar]
- Klauser, T. , Pohlner, J. , and Meyer, T.F. (1990) Extracellular transport of cholera toxin B subunit using Neisseria IgA protease b‐domain: conformation‐dependent outer membrane translocation. EMBO J 9: 1991–1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klauser, T. , Pohlner, J. , and Meyer, T.F. (1992) Selective extracellular release of cholera toxin B subunit by Escherichia coli: dissection of Neisseria Igab‐mediated outer membrane transport. EMBO J 11: 2327–2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klemm, P. , and Schembri, M.A. (2000) Fimbrial surface display systems in bacteria: from vaccines to random libraries. Microbiology 146(Pt 12): 3025–3032. [DOI] [PubMed] [Google Scholar]
- Klimka, A. , Matthey, B. , Roovers, R.C. , Barth, S. , Arends, J.W. , Engert, A. , and Hoogenboom, H.R. (2000) Human anti‐CD30 recombinant antibodies by guided phage antibody selection using cell panning. Br J Cancer 83: 252–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowles, T.J. , Scott‐Tucker, A. , Overduin, M. , and Henderson, I.R. (2009) Membrane protein architects: the role of the BAM complex in outer membrane protein assembly. Nat Rev Microbiol 7: 206–214. [DOI] [PubMed] [Google Scholar]
- Koch‐Nolte, F. , Reyelt, J. , Schossow, B. , Schwarz, N. , Scheuplein, F. , Rothenburg, S. , et al (2007) Single domain antibodies from llama effectively and specifically block T cell ecto‐ADP‐ribosyltransferase ART2.2 in vivo. Faseb J 21: 3490–3498. [DOI] [PubMed] [Google Scholar]
- Koebnik, R. , Locher, K.P. , and Van Gelder, P. (2000) Structure and function of bacterial outer membrane proteins: barrels in a nutshell. Mol Microbiol 37: 239–253. [DOI] [PubMed] [Google Scholar]
- Koide, A. , and Koide, S. (2012) Affinity maturation of single‐domain antibodies by yeast surface display. Methods Mol Biol 911: 431–443. [DOI] [PubMed] [Google Scholar]
- Kromann‐Hansen, T. , Oldenburg, E. , Yung, K.W. , Ghassabeh, G.H. , Muyldermans, S. , Declerck, P.J. , et al (2016) A camelid‐derived antibody fragment targeting the active site of a serine protease balances between inhibitor and substrate behavior. J Biol Chem 291: 15156–15168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kronqvist, N. , Lofblom, J. , Jonsson, A. , Wernerus, H. , and Stahl, S. (2008) A novel affinity protein selection system based on staphylococcal cell surface display and flow cytometry. Protein Eng Des Sel 21: 247–255. [DOI] [PubMed] [Google Scholar]
- Krüger, C. , Hu, Y. , Pan, Q. , Marcotte, H. , Hultberg, A. , Delwar, D. , et al (2002) In situ delivery of passive immunity by lactobacilli producing single‐chain antibodies. Nat Biotechnol 20: 702–706. [DOI] [PubMed] [Google Scholar]
- Lauwereys, M. , Arbabi Ghahroudi, M. , Desmyter, A. , Kinne, J. , Holzer, W. , De Genst, E. , et al (1998) Potent enzyme inhibitors derived from dromedary heavy‐chain antibodies. EMBO J 17: 3512–3520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, S.Y. , Choi, J.H. , and Xu, Z. (2003) Microbial cell‐surface display. Trends Biotechnol 21: 45–52. [DOI] [PubMed] [Google Scholar]
- Leo, J.C. , Grin, I. , and Linke, D. (2012) Type V secretion: mechanism(s) of autotransport through the bacterial outer membrane. Philos Trans R Soc Lond B Biol Sci 367: 1088–1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leo, J.C. , Oberhettinger, P. , Schutz, M. , and Linke, D. (2015) The inverse autotransporter family: intimin, invasin and related proteins. Int J Med Microbiol 305: 276–282. [DOI] [PubMed] [Google Scholar]
- van der Linden, R.H. , Frenken, L.G. , de Geus, B. , Harmsen, M.M. , Ruuls, R.C. , Stok, W. , et al (1999) Comparison of physical chemical properties of llama VHH antibody fragments and mouse monoclonal antibodies. Biochim Biophys Acta 1431: 37–46. [DOI] [PubMed] [Google Scholar]
- Lipes, B.D. , Chen, Y.H. , Ma, H. , Staats, H.F. , Kenan, D.J. , and Gunn, M.D. (2008) An entirely cell‐based system to generate single‐chain antibodies against cell surface receptors. J Mol Biol 379: 261–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, B. , Conrad, F. , Cooperberg, M.R. , Kirpotin, D.B. , and Marks, J.D. (2004) Mapping tumor epitope space by direct selection of single‐chain Fv antibody libraries on prostate cancer cells. Cancer Res 64: 704–710. [DOI] [PubMed] [Google Scholar]
- Lo, A.W. , Moonens, K. , De Kerpel, M. , Brys, L. , Pardon, E. , Remaut, H. , and De Greve, H. (2014) The molecular mechanism of Shiga toxin Stx2e neutralization by a single‐domain antibody targeting the cell receptor‐binding domain. J Biol Chem 289: 25374–25381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lobato, M.N. , and Rabbitts, T.H. (2003) Intracellular antibodies and challenges facing their use as therapeutic agents. Trends Mol Med 9: 390–396. [DOI] [PubMed] [Google Scholar]
- Lu, Z. , Murray, K.S. , Cleave, V.V. , LaVallie, E.R. , Stahl, M.L. , and McCoy, J.M. (1995) Expression of thioredoxin random peptide libraries on the Escherichia coli cell surface as functional fusions to flagellin: a system designed for exploring protein‐protein interactions. Nat Biotechnol 13: 366–372. [DOI] [PubMed] [Google Scholar]
- Luo, Y. , Frey, E.A. , Pfuetzner, R.A. , Creagh, A.L. , Knoechel, D.G. , Haynes, C.A. , et al (2000) Crystal structure of enteropathogenic Escherichia coli intimin‐receptor complex. Nature 405: 1073–1077. [DOI] [PubMed] [Google Scholar]
- Majander, K. , Korhonen, T.K. , and Westerlund‐Wikstrom, B. (2005) Simultaneous display of multiple foreign peptides in the FliD capping and FliC filament proteins of the Escherichia coli flagellum. Appl Environ Microbiol 71: 4263–4268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marín, E. , Bodelón, G. , and Fernández, L.A. (2010) Comparative analysis of the biochemical and functional properties of C‐terminal domains of autotransporters. J Bacteriol 192: 5588–5602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazor, Y. , Van Blarcom, T. , Mabry, R. , Iverson, B.L. , and Georgiou, G. (2007) Isolation of engineered, full‐length antibodies from libraries expressed in Escherichia coli . Nat Biotechnol 25: 563–565. [DOI] [PubMed] [Google Scholar]
- Mazor, Y. , Van Blarcom, T. , Iverson, B.L. , and Georgiou, G. (2008) E‐clonal antibodies: selection of full‐length IgG antibodies using bacterial periplasmic display. Nat Protoc 3: 1766–1777. [DOI] [PubMed] [Google Scholar]
- Merdanovic, M. , Clausen, T. , Kaiser, M. , Huber, R. , and Ehrmann, M. (2011) Protein quality control in the bacterial periplasm. Annu Rev Microbiol 65: 149–168. [DOI] [PubMed] [Google Scholar]
- Monegal, A. , Ami, D. , Martinelli, C. , Huang, H. , Aliprandi, M. , Capasso, P. , et al (2009) Immunological applications of single‐domain llama recombinant antibodies isolated from a naive library. Protein Eng Des Sel 22: 273–280. [DOI] [PubMed] [Google Scholar]
- Moutel, S. , Bery, N. , Bernard, V. , Keller, L. , Lemesre, E. , de Marco, A. , et al (2016) NaLi‐H1: a universal synthetic library of humanized nanobodies providing highly functional antibodies and intrabodies. eLife 5, e16228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munera, D. , Hultgren, S. , and Fernández, L.A. (2007) Recognition of the N‐terminal lectin domain of FimH adhesin by the usher FimD is required for type 1 pilus biogenesis. Mol Microbiol 64: 333–346. [DOI] [PubMed] [Google Scholar]
- Muyldermans, S. (2013) Nanobodies: natural single‐domain antibodies. Annu Rev Biochem 82: 775–797. [DOI] [PubMed] [Google Scholar]
- Muyldermans, S. , and Smider, V.V. (2016) Distinct antibody species: structural differences creating therapeutic opportunities. Curr Opin Immunol 40: 7–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muyldermans, S. , Cambillau, C. , and Wyns, L. (2001) Recognition of antigens by single‐domain antibody fragments: the superfluous luxury of paired domains. Trends Biochem Sci 26: 230–235.11295555 [Google Scholar]
- Nguyen, V.K. , Su, C. , Muyldermans, S. , and van der Loo, W. (2002) Heavy‐chain antibodies in Camelidae; a case of evolutionary innovation. Immunogenetics 54: 39–47. [DOI] [PubMed] [Google Scholar]
- Nguyen, V.S. , Logger, L. , Spinelli, S. , Desmyter, A. , Le, T.T.H. , Kellenberger, C. , et al (2015) Inhibition of type VI secretion by an anti‐TssM llama nanobody. PLoS One 10: e0122187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolay, T. , Vanderleyden, J. , and Spaepen, S. (2015) Autotransporter‐based cell surface display in Gram‐negative bacteria. Crit Rev Microbiol 41: 109–123. [DOI] [PubMed] [Google Scholar]
- Noinaj, N. , Gumbart, J.C. , and Buchanan, S.K. (2017) The [beta]‐barrel assembly machinery in motion. Nat Rev Microbiol 15: 197–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olichon, A. , and de Marco, A. (2012) Preparation of a naive library of camelid single domain antibodies. Methods Mol Biol 911: 65–78. [DOI] [PubMed] [Google Scholar]
- Oomen, C.J. , Van Ulsen, P. , Van Gelder, P. , Feijen, M. , Tommassen, J. , and Gros, P. (2004) Structure of the translocator domain of a bacterial autotransporter. EMBO J 23: 1257–1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pant, N. , Hultberg, A. , Zhao, Y. , Svensson, L. , Pan‐Hammarström, Q. , Johansen, K. , et al (2006) Lactobacilli expressing variable domain of llama heavy‐chain antibody fragments (lactobodies) confer protection against rotavirus‐induced diarrhea. J Infect Dis 194: 1580–1588. [DOI] [PubMed] [Google Scholar]
- Pardon, E. , Laeremans, T. , Triest, S. , Rasmussen, S.G. , Wohlkonig, A. , Ruf, A. , et al (2014) A general protocol for the generation of Nanobodies for structural biology. Nat Protoc 9: 674–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pautsch, A. , and Schulz, G.E. (2000) High‐resolution structure of the OmpA membrane domain. J Mol Biol 298: 273–282. [DOI] [PubMed] [Google Scholar]
- Pavoni, E. , Vaccaro, P. , Anastasi, A.M. , and Minenkova, O. (2014) Optimized selection of anti‐tumor recombinant antibodies from phage libraries on intact cells. Mol Immunol 57: 317–322. [DOI] [PubMed] [Google Scholar]
- Pepper, L.R. , Cho, Y.K. , Boder, E.T. , and Shusta, E.V. (2008) A decade of yeast surface display technology: where are we now? Comb Chem High Throughput Screening 11: 127–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piñero‐Lambea, C. , Bodelón, G. , Fernández‐Periáñez, R. , Cuesta, A.M. , Álvarez‐Vallina, L. , and Fernández, L.A. (2015) Programming controlled adhesion of E. coli to target surfaces, cells, and tumors with synthetic adhesins. ACS Synth Biol 4: 463–473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pohlner, J. , Halter, R. , Beyreuther, K. , and Meyer, T.F. (1987) Gene structure and extracellular secretion of Neisseria gonorrhoea IgA protease. Nature 325: 458–462. [DOI] [PubMed] [Google Scholar]
- Postow, M.A. , Callahan, M.K. , and Wolchok, J.D. (2015) Immune checkpoint blockade in cancer therapy. J Clin Oncol 33: 1974–1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi, H. , Lu, H. , Qiu, H.‐J. , Petrenko, V. , and Liu, A. (2012) Phagemid vectors for phage display: properties, characteristics and construction. J Mol Biol 417: 129–143. [DOI] [PubMed] [Google Scholar]
- Rajabi‐Memari, H. , Jalali‐Javaran, M. , Rasaee, M.J. , Rahbarizadeh, F. , Forouzandeh‐Moghadam, M. , and Esmaili, A. (2006) Expression and characterization of a recombinant single‐domain monoclonal antibody against MUC1 mucin in tobacco plants. Hybridoma (Larchmt) 25: 209–215. [DOI] [PubMed] [Google Scholar]
- Rakonjac, J. , Bennett, N.J. , Spagnuolo, J. , Gagic, D. , and Russel, M. (2011) Filamentous bacteriophage: biology, phage display and nanotechnology applications. Curr Issues Mol Biol 13: 51–76. [PubMed] [Google Scholar]
- Redman, J.M. , Hill, E.M. , AlDeghaither, D. , and Weiner, L.M. (2015) Mechanisms of action of therapeutic antibodies for cancer. Mol Immunol 67: 28–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rice, J.J. , Schohn, A. , Bessette, P.H. , Boulware, K.T. , and Daugherty, P.S. (2006) Bacterial display using circularly permuted outer membrane protein OmpX yields high affinity peptide ligands. Protein Sci 15: 825–836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ring, A.M. , Manglik, A. , Kruse, A.C. , Enos, M.D. , Weis, W.I. , Garcia, K.C. , and Kobilka, B.K. (2013) Adrenaline‐activated structure of beta2‐adrenoceptor stabilized by an engineered nanobody. Nature 502: 575–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rondot, S. , Koch, J. , Breitling, F. , and Dubel, S. (2001) A helper phage to improve single‐chain antibody presentation in phage display. Nat Biotechnol 19: 75–78. [DOI] [PubMed] [Google Scholar]
- Roovers, R.C. , Laeremans, T. , Huang, L. , De Taeye, S. , Verkleij, A.J. , Revets, H. , et al (2007) Efficient inhibition of EGFR signaling and of tumour growth by antagonistic anti‐EFGR Nanobodies. Cancer Immunol Immunother 56: 303–317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossey, I. , Gilman, M.S. , Kabeche, S.C. , Sedeyn, K. , Wrapp, D. , Kanekiyo, M. , et al (2017) Potent single‐domain antibodies that arrest respiratory syncytial virus fusion protein in its prefusion state. Nat Commun 8: 14158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossiter, A.E. , Leyton, D.L. , Tveen‐Jensen, K. , Browning, D.F. , Sevastsyanovich, Y. , Knowles, T.J. , et al (2011) The essential beta‐barrel assembly machinery complex components BamD and BamA are required for autotransporter biogenesis. J Bacteriol 193: 4250–4253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sadana, P. , Monnich, M. , Unverzagt, C. and Scrima, A. (2017) Structure of the Y. pseudotuberculosis adhesin InvasinE. Protein Sci 26: 1182–1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salema, V. , and Fernández, L.Á. (2013) High yield purification of nanobodies from the periplasm of E. coli as fusions with the maltose binding protein. Protein Expr Purif 91: 42–48. [DOI] [PubMed] [Google Scholar]
- Salema, V. , Marín, E. , Martínez‐Arteaga, R. , Ruano‐Gallego, D. , Fraile, S. , Margolles, Y. , et al (2013) Selection of single domain antibodies from immune libraries displayed on the surface of E. coli cells with two β‐domains of opposite topologies. PLoS One 8: e75126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salema, V. , López‐Guajardo, A. , Gutiérrez, C. , Mencía, M. , and Fernández, L.Á. (2016a) Characterization of nanobodies binding human fibrinogen selected by E. coli display. J Biotechnol 234: 58–65. [DOI] [PubMed] [Google Scholar]
- Salema, V. , Mañas, C. , Cerdán, L. , Piñero‐Lambea, C. , Marín, E. , Roovers, R.C. , et al (2016b) High affinity nanobodies against human epidermal growth factor receptor selected on cells by E. coli display. MAbs 8: 1286–1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauri, A. , Soprova, Z. , Wickstrom, D. , de Gier, J.W. , Van der Schors, R.C. , Smit, A.B. , et al (2009) The Bam (Omp85) complex is involved in secretion of the autotransporter haemoglobin protease. Microbiology 155: 3982–3991. [DOI] [PubMed] [Google Scholar]
- Schlapschy, M. , and Skerra, A. (2011) Periplasmic chaperones used to enhance functional secretion of proteins in E. coli . Methods Mol Biol 705: 211–224. [DOI] [PubMed] [Google Scholar]
- Schlapschy, M. , Grimm, S. , and Skerra, A. (2006) A system for concomitant overexpression of four periplasmic folding catalysts to improve secretory protein production in Escherichia coli . Protein Eng Des Sel 19: 385–390. [DOI] [PubMed] [Google Scholar]
- Scott, A.M. , Wolchok, J.D. , and Old, L.J. (2012) Antibody therapy of cancer. Nat Rev Cancer 12: 278–287. [DOI] [PubMed] [Google Scholar]
- Shriver‐Lake, L.C. , Goldman, E.R. , Zabetakis, D. , and Anderson, G.P. (2017) Improved production of single domain antibodies with two disulfide bonds by co‐expression of chaperone proteins in the Escherichia coli periplasm. J Immunol Methods 443: 64–67. [DOI] [PubMed] [Google Scholar]
- Siegel, R.W. (2009) Antibody affinity optimization using yeast cell surface display. Methods Mol Biol 504: 351–383. [DOI] [PubMed] [Google Scholar]
- Siva, A.C. , Kirkland, R.E. , Lin, B. , Maruyama, T. , McWhirter, J. , Yantiri‐Wernimont, F. , et al (2008) Selection of anti‐cancer antibodies from combinatorial libraries by whole‐cell panning and stringent subtraction with human blood cells. J Immunol Methods 330: 109–119. [DOI] [PubMed] [Google Scholar]
- Soltes, G. , Barker, H. , Marmai, K. , Pun, E. , Yuen, A. , and Wiersma, E.J. (2003) A new helper phage and phagemid vector system improves viral display of antibody Fab fragments and avoids propagation of insert‐less virions. J Immunol Methods 274: 233–244. [DOI] [PubMed] [Google Scholar]
- Sousa, C. , Cebolla, A. , and de Lorenzo, V. (1996) Enhanced metalloadsorption of bacterial cells displaying poly‐His peptides. Nat Biotechnol 14: 1017–1020. [DOI] [PubMed] [Google Scholar]
- Stathopoulos, C. , Georgiou, G. , and Earhart, C.F. (1996) Characterization of Escherichia coli expressing an Lpp'OmpA(46‐159)‐PhoA fusion protein localized in the outer membrane. Appl Microbiol Biotechnol 45: 112–119. [DOI] [PubMed] [Google Scholar]
- Steeland, S. , Vandenbroucke, R.E. and Libert, C. (2016) Nanobodies as therapeutics: big opportunities for small antibodies. Drug Discov Today 21: 1076–1113. [DOI] [PubMed] [Google Scholar]
- Stijlemans, B. , Conrath, K. , Cortez‐Retamozo, V. , Van Xong, H. , Wyns, L. , Senter, P. , et al (2004) Efficient targeting of conserved cryptic epitopes of infectious agents by single domain antibodies. African trypanosomes as paradigm. J Biol Chem 279: 1256–1261. [DOI] [PubMed] [Google Scholar]
- Stijlemans, B. , Caljon, G. , Natesan, S.K. , Saerens, D. , Conrath, K. , Perez‐Morga, D. , et al (2011) High affinity nanobodies against the Trypanosome brucei VSG are potent trypanolytic agents that block endocytosis. PLoS Pathog 7: e1002072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thie, H. , Schirrmann, T. , Paschke, M. , Dubel, S. , and Hust, M. (2008) SRP and Sec pathway leader peptides for antibody phage display and antibody fragment production in E. coli . New Biotechnol 25: 49–54. [DOI] [PubMed] [Google Scholar]
- Tijink, B.M. , Laeremans, T. , Budde, M. , Stigter‐van Walsum, M. , Dreier, T. , de Haard, H.J. , et al (2008) Improved tumor targeting of anti‐epidermal growth factor receptor Nanobodies through albumin binding: taking advantage of modular Nanobody technology. Mol Cancer Ther 7: 2288–2297. [DOI] [PubMed] [Google Scholar]
- Tsai, J.C. , Yen, M.‐R. , Castillo, R. , Leyton, D.L. , Henderson, I.R. , and Saier, M.H. Jr (2010) The bacterial intimins and invasins: a large and novel family of secreted proteins. PLoS One 5: e14403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsirigotaki, A. , De Geyter, J. , Sostaric, N. , Economou, A. , and Karamanou, S. (2017) Protein export through the bacterial Sec pathway. Nat Rev Microbiol 15: 21–36. [DOI] [PubMed] [Google Scholar]
- Tumeh, P.C. , Harview, C.L. , Yearley, J.H. , Shintaku, I.P. , Taylor, E.J.M. , Robert, L. , et al (2014) PD‐1 blockade induces responses by inhibiting adaptive immune resistance. Nature 515: 568–571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Ulsen, P. , Rahman, S. , Jong, W.S. , Daleke‐Schermerhorn, M.H. , and Luirink, J. (2014) Type V secretion: from biogenesis to biotechnology. Biochim Biophys Acta 1843: 1592–1611. [DOI] [PubMed] [Google Scholar]
- Valls, M. , Atrian, S. , de Lorenzo, V. , and Fernández, L.A. (2000) Engineering a mouse metallothionein on the cell surface of Ralstonia eutropha CH34 for immobilization of heavy metals in soil. Nat Biotechnol 18: 661–665. [DOI] [PubMed] [Google Scholar]
- Van Impe, K. , Bethuyne, J. , Cool, S. , Impens, F. , Ruano‐Gallego, D. , De Wever, O. , et al (2013) A nanobody targeting the F‐actin capping protein CapG restrains breast cancer metastasis. Breast Cancer Res 15: R116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaneycken, I. , D'huyvetter, M. , Hernot, S. , De Vos, J. , Xavier, C. , Devoogdt, N. , et al (2011) Immuno‐imaging using nanobodies. Curr Opin Biotechnol 22, 877–881. [DOI] [PubMed] [Google Scholar]
- Veggiani, G. , Ossolengo, G. , Aliprandi, M. , Cavallaro, U. , and de Marco, A. (2011) Single‐domain antibodies that compete with the natural ligand fibroblast growth factor block the internalization of the fibroblast growth factor receptor 1. Biochem Biophys Res Commun 408: 692–696. [DOI] [PubMed] [Google Scholar]
- Veiga, E. , Sugawara, E. , Nikaido, H. , de Lorenzo, V. , and Fernández, L.A. (2002) Export of autotransported proteins proceeds through an oligomeric ring shaped by C‐terminal domains. EMBO J 21: 2122–2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veiga, E. , De Lorenzo, V. , and Fernández, L.A. (2004) Structural tolerance of bacterial autotransporters for folded passenger protein domains. Mol Microbiol 52: 1069–1080. [DOI] [PubMed] [Google Scholar]
- Verheesen, P. , and Laeremans, T. (2012) Selection by phage display of single domain antibodies specific to antigens in their native conformation. Methods Mol Biol 911: 81–104. [DOI] [PubMed] [Google Scholar]
- Vosjan, M.J. , Vercammen, J. , Kolkman, J.A. , Stigter‐van Walsum, M. , Revets, H. , and van Dongen, G.A. (2012) Nanobodies targeting the hepatocyte growth factor: potential new drugs for molecular cancer therapy. Mol Cancer Ther 11: 1017–1025. [DOI] [PubMed] [Google Scholar]
- Wells, T.J. , Sherlock, O. , Rivas, L. , Mahajan, A. , Beatson, S.A. , Torpdahl, M. , et al (2008) EhaA is a novel autotransporter protein of enterohemorrhagic Escherichia coli O157:H7 that contributes to adhesion and biofilm formation. Environ Microbiol 10: 589–604. [DOI] [PubMed] [Google Scholar]
- Wentzel, A. , Christmann, A. , Adams, T. , and Kolmar, H. (2001) Display of passenger proteins on the surface of Escherichia coli K‐12 by the enterohemorrhagic E. coli intimin EaeA. J Bacteriol 183: 7273–7284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung, Y.A. , and Wittrup, K.D. (2002) Quantitative screening of yeast surface‐displayed polypeptide libraries by magnetic bead capture. Biotechnol Prog 18: 212–220. [DOI] [PubMed] [Google Scholar]
- Zhou, X. , Hendrickx, M.L. , Hassanzadeh‐Ghassabeh, G. , Muyldermans, S. , and Declerck, P.J. (2016) Generation and in vitro characterisation of inhibitory nanobodies towards plasminogen activator inhibitor 1. Thromb Haemost 116: 1032–1040. [DOI] [PubMed] [Google Scholar]