

Abstract
Exploring gene function is critical for understanding the complexity of life. DNA sequences and the three-dimensional organization of chromatin (chromosomal interactions) are considered enigmatic factors underlying gene function, and interactions between two distant fragments can regulate transactivation activity via mediator proteins. Thus, a series of chromosome conformation capture techniques have been developed, including chromosome conformation capture (3C), circular chromosome conformation capture (4C), chromosome conformation capture carbon copy (5C), and high-resolution chromosome conformation capture (Hi-C). The application of these techniques has expanded to various fields, but cancer remains one of the major topics. Interactions mediated by proteins or long noncoding RNAs (lncRNAs) are typically found using 4C-sequencing and chromatin interaction analysis by paired-end tag sequencing (ChIA-PET). Currently, Hi-C is used to identify chromatin loops between cancer risk-associated single-nucleotide polymorphisms (SNPs) found by genome-wide association studies (GWAS) and their target genes. Chromosomal conformations are responsible for altered gene regulation through several typical mechanisms and contribute to the biological behavior and malignancy of different tumors, particularly prostate cancer, breast cancer and hematologic neoplasms. Moreover, different subtypes may exhibit different 3D-chromosomal conformations. Thus, C-tech can be used to help diagnose cancer subtypes and alleviate cancer progression by destroying specific chromosomal conformations. Here, we review the fundamentals and improvements in chromosome conformation capture techniques and their clinical applications in cancer to provide insight for future research.
Keywords: Chromosomal conformations, Cancer, Diagnosis, Treatment, Chromosome conformation capture
Background
Gene expression in eukaryotic cells is regulated by many different complex mechanisms. In addition to epigenetic alterations (i.e., histone modifications, DNA methylation and noncoding RNA), chromosome conformation can influence gene function.
Chromosomes have unique, high-ordered structures that influence gene function. The higher-ordered genome structure often formed by hierarchical folding. First, the DNA double helix winds around an octamer of histone proteins to create the nucleosome. A beads-on-string fiber with a width of approximately 10 nm is then formed [1], followed by 30-nm fibers and global structures, and the length of the DNA chain folds to one-hundred-thousandth of the nonfolded size.
Proteins are involved in the formation of higher-ordered chromosome structures, such as chromosome loops. Some proteins, including special AT-rich sequence-binding protein-1 (SATB1), CCCTC-binding factor (CTCF) and cohesin, play key roles in disease development and recovery. Genes located at long distances from one another can interact through these known and unknown folding mechanisms to produce variable results.
Techniques to detect chromosome conformation
To better study chromosome conformations, the following two types of techniques have been developed: observation methods and C-techs. Observation methods include fluorescence in situ hybridization (FISH), microscopy (light microscopy or electron microscopy), and nuclear ligation [2]; C-tech methods are more novel and technical. Chromosome conformation capture (3C) was invented in 2002 and was the first member of the C-tech family [3]. Subsequently, 3C has rapidly advanced to circular chromosome conformation capture (4C), chromosome conformation capture carbon copy (5C), high-resolution chromosome conformation capture (Hi-C), chromatin interaction analysis by paired-end tag sequencing (ChIA-PET), targeted chromatin capture (T2C), Capture-C and many others to meet a wider variety of needs (Table 1).
Table 1.
Techniques used for chromosome conformation capture
| Technology | 3C | 4C | 5C | Hi-C | ChIA-PET |
|---|---|---|---|---|---|
| Data | One-one | One-all | Many-many | All-all | Many-many |
| Readout | PCR | Inverse PCR, microarrays, NGS | LMA, sequencing | Sequencing | Sequencing |
| Advantages | Simple, cheap | Relatively simple, goal-oriented | Explores interactions among many fragments, reduces bias, relatively targeted | Genome-wide interactions | Protein bonded interactions |
| Limitations | Laborious, only known combinations, poor resolution | Only chosen regions | Difficult to design hundreds of primers, analyzes only chosen fragments, some long-distance interactions are missing | Difficult to sequence and analyze, expensive | Analyzes only chosen protein |
| Ref. | [3, 10] | [12] | [17, 18, 142] | [19] | [24, 25] |
Abbreviations: PCR Polymerase Chain Reaction, NGS Next-Generation Sequencing, LMA Ligation-mediated Amplification
Most of above mentioned technologies are based on 3C technology. After proteins are fixed and crosslinked with formaldehyde, chromatin is cut into 4- to 10-kb pieces [3] by 6-base recognizing enzymes, such as HindIII [3], EcoRI [4], BglII [5], DdeI [6], DpnII [7] and BamHI [8]. The use of a 4-base recognizing enzyme may be ideal for analyzing a narrow area, but these enzymes create too many small pieces. DNase I has been recently used to enhance the resolution [9]. One megabase is often considered the limit of 3C technologies, but larger distances [10] even those between chromosomes [11] have been reported. Semi-quantitative polymerase chain reaction (PCR) is always performed to verify the ligation junctions of interest. 3C technology is often used to analyze enhancer or insulator activities using CTCF but is limited in its ability to identify unknown regions.
4C technology enables the discovery of longer distance co-associations. Unlike the 6-base recognizing enzymes used in 3C, 4-base recognizing enzymes are used to cut chromosomes into 256-bp fragments. First, a single chromosome region of interest is selected, and global chromosomes associated with that region are sequenced. The target genes are amplified and sequenced, and global genome-wide interactions with the chosen fragment can be identified. This process is always described as one-to-all [12]. 4C technology also includes chromosome conformation capture on chromatin immunoprecipitation (ChIP) [13], which is similar to enhanced the chromosome conformation capture on ChIP (e4C) [14] approach that adds an optional ChIP step to test fragment binding to a chosen protein and can be more sensitive. Using 4C-seq [15], we can quickly and comprehensively identify the sequences of interest.
5C can be used to map many interactions among several fragments in a region [16]. After the difficult step of designing hundreds of primers [17], ligation-mediated amplification (LMA) is commonly performed following 5C instead of PCR, which is commonly performed following 3C [18]. 5C can provide more detailed information and be used to construct a special network around genes of interest. Hi-C was designed in 2009 to recover ligated fragments using streptavidin to capture global genome-wide interactions, including long-distance interactions [19], and protocols and algorithms have demonstrated that it improves capture range and accuracy [20, 21]. Single-cell Hi-C can be used to analyze differences in chromosome conformation among cells and determine the uniqueness of cells [22]. In addition to chromosomes, ChIA-PET can be used to test global interactions with a chosen protein, such as CTCF [23] or polymerase II (Pol II) [24]. Moreover, new statistical patterns have been identified for ChIA-PET [25]. Additional new C-techs have been developed. Capture-C is a 3C-like technique with high resolution that can capture interactions in cis conformations using oligonucleotide probes [26]. T2C focuses on the target gene and explores its local interactions, thus reducing cost and labor [27], and is widely used to predict local loops [28]. Capture-Hi-C is a many-to-all Hi-C technique with higher resolution because it includes an additional step in which the sequences of interest (baits) are enriched [29]. RNA-guided chromatin conformation capture (R3C) is a new method that can be used to precisely study interactions between RNA and DNA [30, 31] (Fig. 1).
Fig. 1.
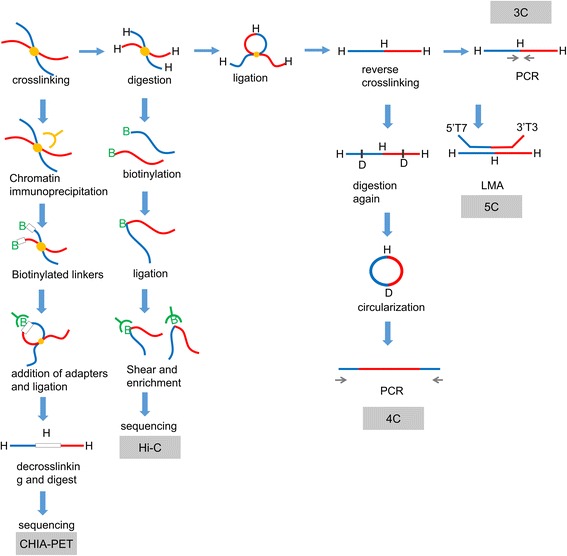
Overview of 3C-based C-techniques. After crosslinking, chromatin is digested into 4-10 kb pieces by restriction enzymes, followed by ligation and reverse crosslinking to change the crosses into lines. An additional digestion step is added in 4C. LMA is used instead of PCR, which is commonly used in 5C; biotinylation and streptavidin are used in Hi-C, and a chromatin immunoprecipitation (CHIP) step is added in ChIA-PET
Other techniques, mainly developed prior to 3C techniques, can help to identify chromosomal interactions. FISH, microscopy (light microscopy or electron microscopy), and nuclear ligation [2] are widely used to observe and verify the higher-ordered chromosome organization. ChIA-PET is another advanced technique used to test global interactions with a chosen protein, such as CTCF [23] or DNA Pol II [24]. New statistical tools for ChIA-PET analysis have also been developed [25]. The 4D Nucleome Project provides a coherent view of how chromosome conformations change, providing insight beyond the static state [32]. One of the newest tools, genome architecture mapping (GAM), shows chromosomal contacts between long-range fragments by sequencing thin nuclear sections [33]. Currently, dCas9 capture with a biotinylated dCas9 step before 3C, RNA-seq or proteomics shows chromosomal interactions at a single-copy genomic locus and clarifies trans-regulatory factors [34].
Characteristics and formation of chromosome loops
Because DNA is packaged and unpackaged in the nucleus, long-range intra- or inter-chromosomal interactions occur, and loops may be formed to stabilize these interactions. In fact, only 7% of loops occur between the two nearest genes. Of course, chromosome loops are not completely random. The specific DNA topology dictates the sequence [35], flexibility of the chromatin (the ring-closure probability) [36], and other dynamic factors, such as distance, energy [37] and negative DNA supercoiling [38]. A chromosome loop often contains at least two DNA sequences (for example, one sequence is an enhancer/silencer, and the other is a promoter) and some mediators. Although different cell types have different interactions, enhancers, CTCFs, and actively transcribed chromatin states rich in loops, 79% of loops span CTCF binding sites, which can prevent enhancer-promoter interactions without loops [39]. CTCF and cohesin are present in over 86% of loops. All loop domains are separated into 6 subcompartments by 8 histone markers, i.e., histone 3 lysine 36 trimethylation (H3K36me3), histone 3 lysine 27 trimethylation (H3K27me3), histone 3 lysine 4 methylation (H3K4me1), histone 3 lysine 4 dimethylation (H3K4me2), histone 3 lysine 4 trimethylation (H3K4me3), histone 3 lysine 9 trimethylation (H3K9me3), histone 3 lysine 79 dimethylation (H3K79me2), and histone 4 lysine 20 methylation (H4K20me1); the contact domains contain the same markers and change together [40]. In addition, these long-range interactions are inherited from sperm but not the ovum to regulate embryo development and growth [41, 42]. Loops are stable from the G1 to S and G2 phases of the cell cycle [43].
The mechanism by which chromosomal loops alter gene regulation remains unclear, but several mechanisms have been proposed, particularly in cancer (Fig. 2). The examination of topologically associated domains (TADs) is another avenue for studying the relationship between chromosome loops and cancers. CTCF is enriched in the boundaries of TADs and can guide interactions by forming loops with cohesin between distantly located sequences [44] to influence gene expression [45]. These loops bind Pol II factories, including Pol II, transcription factors (TFs) and co-TFs [46]. CTCF and cohesin have been widely reported as the most common and important protein mediators [39, 40, 47]. Estrogen receptor α (ERα) is an estradiol (E2)-dependent mediator that binds E2 in the cytoplasm and is then translocated to the nucleus to activate Pol II and recruit forkhead box A1 (FoxA1) and activator protein (AP)-2γ [48–50]. Pol II can mediate interactions between enhancers and promoters and between distinct promoters [51], indicating that promoters can act as enhancers [24]. New mediators and mechanisms have been reported. Mediator complex subunit 1 (MED1) forms loops only when it is phosphorylated; phosphorylated MED1 facilitates loop formation by FoxA1, Pol II and TATA-box binding protein (TBP) [52]. Cut-like homeobox 1 (CUX1) is another mediator that can recruit Pol II via a new mechanism; at normal concentrations, CUX1 mediates interactions between promoters and several enhancers, but the number of binding sites decreases when the dosage of CUX1 is halved [53]. Binding of lncRNA to the polycomb repressive complex may increase the chance of interactions [54] or recruit a mediator protein to form long-range interactions [30]. Most loops form between promoters and promoters or promoters and enhancers, whereas a minority between promoters and insulators [55]; some loops are even involved in chromosomal translocations [56].
Fig. 2.
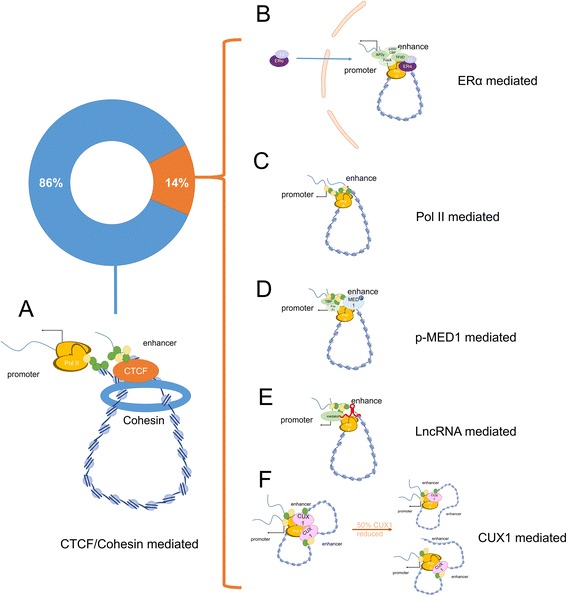
Several mechanisms for forming chromosomal loops in cancer. Pol II (orange), transcription factors (green) and co-transcription factors (yellow) are always recruited by long-range loop mediators in transcription factories. a Most loops are mediated by CTCF and cohesin, with or without other mediators. b ERα is another common mediator that functions after binding E2 and entering the nucleus. c Pol II can mediate long-range loops between promoters, suggesting that promoters sometimes act as enhancers. d MED1 acts as a chromosomal loop mediator only when it is phosphorylated. e Some lncRNAs can mediate chromosomal loops. f CUX1 is special in that it mediates the binding of two enhancers to one promoter, and when its expression is reduced to 50%, it mediates the binding of one enhancer to one promoter
Roles of chromosomal conformations in cancer
The rapid development of new techniques makes it possible to determine important roles of chromosomal conformations in many fields, especially in cancer. Specific proteins mediate the chromosome loops between different cancer-related genes (including their promoters) and an enhancer or a repressor to regulate gene transcription and influence the biological behaviors of carcinomas. In this review, we discuss the function of these loops and the possible clinical applications of these fatal chromosome conformations in diagnosis and treatment (Table 2).
Table 2.
Summary of chromosomal interactions in cancer
| Cancer | Typical interactions | Mediators | Therapeutic targets | Potential drugs | Ref | |
|---|---|---|---|---|---|---|
| Prostate cancer | CRPC-specific enhancers | UBE2C | FOXA1 and phosphorylated MED1 | PI3K/AKT/phosphorylated MED1 pathway | Carvacrol, ipatasertib, abiraterone acetate | [52] |
| PSA enhancer | PSA promoter | AR/PCGEM1 and PRNCR1 (PCAT8) | AR | Sigma1 inhibitor, ASC-J9, cisplatin, niclosamide, EPI-001, | [62] [66] | |
| TFF3 | ERG | ERG | ERG binding domain | ERG inhibitory peptides | [64] | |
| MOXD1 | FYN/SERPINB9/HEY2 | |||||
| 2p11.2 | CAPG | / | / | / | [90] | |
| 6q22.1 | RFX6/GPRC6A | |||||
| 1q32.1 | NFASC | |||||
| 8q24 | MYC | |||||
| Breast cancer | IGFBP3 | EGFR/MCF7/BCAS1-4 | / | / | / | [68] |
| GREB1 | TFF1 promoter | ERα | / | / | [48–50] | |
| ERα | EGR1/TGFBR2/DAXX/BCL2L1 | / | / | / | [74] | |
| ZEB2 promoter 1a | ZEB2 promoter 1b | AP-1 | AP-1, ZEB2 and ERK/Akt | AP1-C301-S, SR 11302, valproic acid, U0126, miR-132 | [77] | |
| Hematologic neoplasms | GATA2 enhancer | EVI1 | / | EVI1 | Pyrrole-Imidazole Polyamide 1 | [79] |
| BCR | ABL | / | BCR-ABL fusion gene, BCL2 | ZINC12891610 (hit2), venetoclax | [136] | |
| HoxA | Genes on chromosome 14 | TFBS | / | / | [81] | |
| IGF1R promoter | IGF1R enhancer | IRAIN | / | / | [82] | |
| RUNX1 promoters | RUNX1 enhancer | RUNXOR | / | / | [56] | |
| Unknown promoters | Unknown enhancers | CUX1 | CUX1 | / | [53] | |
| Colorectal cancer | MYC promoter | 335-kb enhancer | CCAT1-L CTCF |
/ | / | [84] |
| MYC promoter MECOM ETS1 |
rs6983267 (contains multiple enhancers) 3q26.2 11q23 |
CCAT1 | / | / | [95] | |
| Pancreatic cancer | rs386772267 | DIS3 promoter | An allele-specific TF | / | / | [96] |
| Melanoma | MITF | BRN2/CDKN1A/TBX3 | / | MITF | HIV1 protease inhibitor nelfinavir | [86] |
| Thyroid carcinoma | rs965513 mutant | FOXE1 and PTCSC2 promoter | / | / | / | [97] |
| Gliomas | FIPL1L1 enhancer | PDGFRA | CTCF altered by mutant IDH | / | / | [98] |
| rs73001406 | DDX6 promoter | / | / | / | [99] | |
| Neurofibromatosis | NF1 | 1-Mb fragment nearby | / | / | / | [87] |
| Adenoid cystic carcinoma | MYB promoter | MYB enhancer | ACCMYB-TGFBR3 translocation | Super-enhancer BRD4 | JQ1 | [88] |
| Testicular germ cell tumors | Predicted GATA4 promoter 1q22 11q14.1 |
20q13.2 15q25.2 15q22.31 |
/ | / | / | [100] |
PSA prostate-specific antigen
Chromosomal conformations in cancer
Two research strategies are used to identify new chromosomal conformations and their characteristics. C-techs are used to identify classical interactions that are mediated by typical proteins, and Hi-C is currently used to identify chromatin loops between cancer risk-associated single-nucleotide polymorphisms (SNPs) found by genome-wide association studies (GWAS) and their target genes.
Classical long-range interactions
ChIA-PET, CHIP-seq and 4C-seq are widely used to identify long-range interactions that are mediated by proteins or lncRNAs. In addition, most interactions are found in prostate cancer, breast cancer and leukemia, which will be discussed in detail in the following section.
The abnormal activation of the androgen receptor (AR) is a major cause of androgen-dependent prostate cancer (ADPC) and its advanced stage, i.e., castration-resistant prostate cancer (CRPC), in which the AR acts as a transcription factor. Some AR binding regions are far from the AR and associate with the AR through chromosome loops. In general, there are two ways for AR-driven chromosome looping to function in neoplasia [57]. First, AR can increase spatial association of the transmembrane protease, serine 2 (TMPRSS2) gene and ETS-related gene (ERG) and mediate TMPRSS2-ERG gene fusion [58], which is a clinical marker of prostate cancer. Moreover, the loops can associate long-range genes, which are often enhancers, with cancer-related gene promoters and change their expression [59]; this process is mediated by AR-collaborating TFs [60]. In addition, changes in certain active histone modifications, such as H3K4me1, H3K4me2 and histone H3 acetylation, are also important in forming chromosomal conformations. For example, ERG disrupts the AR by activating the H3K27 methyltransferase EZH2 [61]. Three CRPC-specific enhancers interact with the AR target gene ubiquitin-conjugating enzyme E2C (UBE2C) through chromosome loops mediated by FOXA1 and phosphorylated MED1, which leads to high expression of UBE2C and makes CRPC an incurable disease, unlike ADPC [52]. Similar loops can be observed between the AR binding sites and the promoters of prostate-specific antigen (PSA) [62], which is one of the most important genes in prostate cancer. Overexpression of ERG may also contribute to a change in chromosome topology [63]. In addition, the long-distance interactions among TFF3-ERG, MOXD1-FYN, MOXD1-SERPINB9, and MOXD1-HEY2 were specifically found in prostate cancer cells by a Hi-C analysis [64]. TADs are smaller and more plentiful in the prostate cancer cell genome, and the boundaries of these TADs attract more CTCF binding and H3K4me3 [65], providing more opportunities to form long-range interactions. In fact, these differences lead to cancer-related interactions between enhancers and promoters that occur in epigenetically deregulated domains [65]. lncRNAs are also involved; for example, PCGEM1 and PRNCR1 (PCAT8) mediate the enhancer-promoter chromosome loops and promote AR activation [66].
Chromosome conformations also influence tumorigenesis and breast cancer development. The long-range interactions of insulin-like growth factor-binding protein 3 (IGFBP3), a key gene in the development of breast cancer [67], are increased in breast cancer cells relative to those in normal cells [68]. The association between IGFBP3 with epidermal growth factor receptor (EGFR), another breast cancer-related gene, may contribute to tumorigenesis in a manner similar to the long-range interactions between IGFBP3 and breast carcinoma amplified sequence (BCAS1-4) in MCF7 cells [68]. ERα is another important mediator of long-range interactions in breast cancer. ERα was first reported to mediate long-distance interactions between GREB1 and the trefoil factor 1 (TFF1) gene promoter with the help of FOXA1 and AP-2γ [48–50]. Several other cofactors, including GATA3 [69], TLE1 [70], and PBX1 [71], also play roles in this dramatic process. The TF CTCF without cohesin binding sites is very rich in ERα binding sites and mediates enhancer-promoter loops [72]; a few sites have both cohesin and ERα but rarely CTCF [73]. A recent Hi-C experiment showed that many gene sites interacted with ERα, including cancer suppressor genes EGR1 and TGFBR2, apoptosis genes DAXX and BCL2L1, and densely mapped, distant estrogen-responsive elements located in 17q23 and 20q13 [74]. Another Hi-C study showed that runt-related transcription factor 1 (RUNX1) contributes to local interactions rather than long-range interactions in MCF7 cells [75]. Due to the development of ChIA-PET, more ERα-mediated interactions deeply involved in breast cancer have been identified [76].
Triple-negative breast cancer (TNBC) is a highly malignant breast cancer that is not associated with ERα. Activator protein 1 (AP1) is activated by tumor necrosis factor alpha (TNFα) and binds the long-range promoter of zinc finger E-box-binding homeobox 2 (ZEB2) to influence its expression, resulting in epithelial-mesenchymal transitions that are considered a potential novel therapeutic target for TNBC [77]. CTCF mediates the binding of the unmethylated imprinting control region to the IGF2 promoter region in the breast cancer MCF7 cell line [52]. An antisense noncoding RNA, i.e., IRAIN, is involved in the allele-switch pattern in breast cancer [31]. However, in many cases, more inter-chromosomal interactions appear in normal cells than in tumor cells [78].
The Philadelphia chromosome, which is an interchange between chromosomes 9 and 22 that produces a BCR-ABL fusion gene that can be found in almost all types of chronic myelogenous leukemia (CML), some types of acute lymphoblastic leukemia (ALL) and several types of acute myelocytic leukemia (AML), is a key example of the role of chromosome conformations in leukemia. Long-range interactions play an important role in this disease.
In AML with inv.(3)/t(3;3), an enhancer that interacts with GATA2 in normal cells, loses its connection to GATA2 and re-interacts with (active) EVI1, leading to cancer [79]. Translocation partner genes (TPGs) are located in transcriptional activity-related regions and mediate interactions among higher-order chromosome structures. This phenomenon was observed in both GM06990 and K562 cell lines [80]. In addition, it was determine the position of morbigenous translocations and revealed the global interactions of the HoxA gene on chromosome 7 [81]. lncRNAs, such as IRAIN, participate in the formation of intra-chromosomal loops; lncRNAs help the IGF1R promoter contact the enhancer, which is located 150 kb away [82]. RUNXOR can mediate the interaction between the RUNX1 promoters and its enhancer by forming a RUNX1 intra-chromosome loop and is involved in chromosomal translocation [56]. CUX1, a haploinsufficient TF, is a new mediator that contributes to long-range interactions between enhancers and sites close to the TSS in hematologic neoplasms and solid tumors; 32-49% of CUX1 co-localizes with RNA Pol II, and 34-70% of CUX1 co-localizes with EP300, a transcriptional co-activator. However, the cohesion complex appears more frequently, further demonstrating that CUX1 contributes to chromatin loops [53].
Chromosomal conformations play important roles in many malignant tumors. CTCF binding sites can change in colorectal cancer and are considered alterable chromosomal conformations [83]. In addition, lncRNA CCAT1-L together with CTCF can mediate a 335-kb interaction between the MYC promoter and its enhancer in colorectal cancer [84]. Microphthalmia-associated transcription factor (MITF) plays a major role in the development and metastasis of melanoma, another highly malignant tumor [85]. Three MITF-related chromosome loops (with BRN2, CDKN1A and TBX3) have been found in both cell lines and patient blood samples. This study was the first confirmation of long-range chromosome loops in melanoma and provided valuable prospects for diagnosis by examination of circulating immune cells [86]. A study investigating neurofibromatosis found a 1-Mb fragment containing the neurofibromatosis type I (NF1) gene that interacts with another nearby 1-Mb fragment recorded in the HindIII and NcoI-maps ([87]) using Hi-C data.
An interesting feedback loop exists in adenoid cystic carcinoma in which MYB protein-bound enhancers can interact long range to activate the promoter of MYB, and ACCMYB-TGFBR3 translocation also places the super-enhancer in contact with the promoter of MYB [88].
Loops involving cancer risk-associated SNPs
Cancer risk-associated SNPs are a leading area of study in oncomolecular biology, and long-range loops at these high-risk sites may be valuable. Some researchers combined Hi-C and occasionally 4C-seq with GWAS to identify additional novel and meaningful loops in which a new browser enables easy visual examination [89].
In prostate cancer, a capture Hi-C experiment revealed that some high-risk SNPs interact with their long-distance target genes, including CAPG, C2orf43, RFX6, NFASC, MYC and AGAP7P, through chromosome loops [90]. 4C-seq with GWAS was used to identify a risk locus for prostate cancers (LNCaP and C4-2B cells), and MYC and POU5F1B were ranked the highest, followed by CD96, PVT1, GSDMC, CXorf36, RRP12, USP14 and SMIN3, which may exhibit abnormal chromatin looping [91]. These techniques also revealed the following key pathways: the TFG-beta signaling pathway, p53 pathway and hypoxia response via HIF activation; which are considered highly important in the development of prostate cancer [91]. In breast cancer, Hi-C and GWAS have been combined to identify risk loci, including the protein-coding genes IGFBP5, KLF4, NSMCE2, and MYC, and the lncRNAs DIRC3, PVT1, and CCDC26, which are associated with CTCF [92, 93]. Capture Hi-C tech has also been applied to the study of colorectal cancer and used to identify long-range interactions between rs6983267 (later shown to contain multiple enhancers [94]) and a MYC promoter mediated by the MYC-related lncRNA CCAT1. Similar loops were identified between 3q26.2 and MECOM and between 11q23 and ETS1 [95]. The pancreatic cancer risk locus rs386772267 suppresses the expression of DIS3 via the loop between rs386772267 and the promoter of DIS3 with an allele-specific TF [96]. In papillary thyroid carcinoma (PTC), a combination of low-penetrance genes is critical, and one of these genes has SNP rs965513. Its mutants (i.e., SNPs rs7864322, rs12352658, rs7847449, and rs10759944) interact with a promoter shared by FOXE1 and PTCSC2 and act as enhancers [97]. Gliomas also display SNP-mediated interactions in which the oncogene PDGFRA is changed by CTCF-related chromosome folding [98]. Other interactions among enhancers, SNP rs73001406, and the DDX6 gene promoter affect cancer risk [99]. By combining GWAS and Hi-C, 19 new risk loci were identified in testicular germ cell tumors, including interactions between a predicted GATA4 promoter and 20q13.2, between 1q22 and 15q25.2, and between 11q14.1 and 15q22.31 [100].
Chromosomal conformations lead to malignancy in cancer
Interactions between promoters and enhancers (or promoters sometimes) can influence gene expression. Some interactions have been reported to specifically change tumor behavior, and these effects depend on the target genes. Some target genes are known to be key in cancer: some are involved in chromosomal translocation [56], and others may be novel diagnostic targets.
Some examples include loops mediated by phosphorylated MED1 [52], the lncRNAs PCGEM1 and PRNCR1 [66], and MYB [88] that strongly promote cell growth. When the loops are disrupted, cancer growth is robustly inhibited in mouse models. The lncRNA IRAIN changes tumor migration ability [31]. When the interactions between GATA2 and EVI1 are disrupted, cancer cells have higher apoptosis rates [79], and CUX1 alters the expression of a cluster of genes and the cell cycle to promote cell proliferation [53]. However, long-range loops enable expression of PSA [62] and overexpression of oncogenic TFs [64].
Potential diagnostic tools and therapeutic targets based on chromosomal conformations
Newly discovered loops could be used as novel diagnostic markers. These newly discovered interactions contribute to previously unknown pathways and may be potential therapeutic targets. There are many applications for the identification of chromosomal conformations in diagnosis and therapy.
Diagnostic targets
Hi-C is a new tool that can be used to detect chromosomal aberrations and copy number variations in human cancer samples (for example, glioblastoma and anaplastic astrocytoma) to identify oncogene amplification, tumor suppressor gene deletion, fusion genes, and balanced/unbalanced structural rearrangements [101].
In prostate cancer, new loops with specific histone modifications may serve as new diagnostic markers; the TMPRSS2-ERG fusion has been used as a marker in diagnosis and risk assessment for many years, even before chromatin loops were identified as the culprit [102]. Some clinical trials have treated the TMPRSS2-ERG fusion gene as a biomarker; this gene can correct most of the false-negative results of the prostate cancer antigen 3 (PCA3) test, can act as a supplement to the serum PSA test [102, 103] and is associated with a poor outcome [104]. But deeper studies are needed to determine whether newly identified lncRNAs could be used as markers of breast cancer and AML.
In CML, the BCR-ABL fusion gene is often indicative of a poor prognosis, while other mutations lead to varying sensitivity to chemotherapy drugs according to European LeukemiaNet recommendations 2013 [105]. In addition, due to the characteristics of 3′ and 5′ TPG, disease-causing fusion genes may be predictive and used in diagnosis [80]. Human leukemia can be classified according to the different chromosome conformations. By mapping the HOXA gene cluster using the 5C technique and analyzing the gene cluster using 3D DNA disease-signature predictor, the MLL-fusion protein can be classified as wild-type, and the subtypes of leukemia can be classified according to their different MLL-fusion partners with high accuracy; thus, the MLL-fusion protein can be used as a novel marker, but this has been verified only in cell lines, and a clinical trial is necessary [106]. Studies have also identified new risk loci, such as NCK1, NCAPH2 and L3MBTL4, in B-cell malignancies (BCMs), including chronic lymphocytic leukemia, Hodgkin lymphoma and multiple myeloma [107]. Moreover, as CUX1 is inactivated in more than half of high-risk myeloid leukemia cases [108], it can be used as a biomarker of poor prognosis.
In melanoma, MITF-related loops identified in circulating tumor cells can be used to noninvasively detect cancer [86] and predict a poor prognosis [109], as shown in humans. In gliomas, 11q23.3 was identified as a susceptibility locus that is closely related to abnormal chromosome conformations [99], and rs965513 was identified as a risk factor in papillary thyroid carcinoma [97].
Many characteristic features have been found in different cancers and other diseases, such as autoimmune disease [110] and obesity [111]. An increasing number of candidate genes have been proposed, but they have not yet been commonly used as clinical markers. Applying these techniques remains challenging.
Therapeutic targets
As previously discussed, many interactions and new functions of RNA and proteins have recently been found in many cancers, and these discoveries may present new therapeutic targets. In addition to small-molecule inhibitors of specific targets that may be predicted by drugs, plasmids carried by histidine-lysine peptide (HKP) enable the use of siRNA as a new agent for targeting specific sequences [112]. Here, we mention some therapeutic targets and drugs, and we describe completed clinical trials and studies with significant effects in vitro and in mice.
As androgen deprivation therapy for prostate cancer often leads to eventual resistance, there is an urgent need for new therapies. The chromosomal loops of UBE2C that are mediated by MED1 in both AR-positive and AR-negative CRPC, as discussed above, were reported to be a new therapeutic target in addition to the PI3K/AKT/phosphorylated MED1 pathway [52]. In clinical and experimental trials, many small-molecule inhibitors have been effective in targeting the AKT and P13K pathways in prostate cancer, including but not limited to carvacrol [113], ipatasertib [114], and abiraterone acetate [115]; phase III trials of abiraterone acetate have been completed and showed a benefit in CRPC patients pre- or post-chemotherapy [116]. Additionally, AR-targeted therapies have also had great success; the small molecule Sigma1 inhibitor suppresses AR function and further suppresses prostate cancer in vivo and in vitro [117]. Anti-AR drugs, such as cisplatin, niclosamide [118], ASC-J9 [119], EPI-001 [120], and even miRNA [121] or lncRNA [122, 123], also suppress prostate cancer [124]. Cisplatin was investigated in a multicenter phase II trial and was shown to be a safe, feasible and active therapy [125]. Gene fusions, such as TMPRSS2-ERG, are often the result of double-strand DNA breaks and error-prone DNA repair, mainly through the error-prone non-homologous end-joining pathway and we found that small-molecule inhibitors and siRNAs targeting this pathway decrease TMPRSS2-ERG gene fusion [126]. Other treatments with clinical potential include those that target the key enzyme EZH2 via its inhibitors DZNep [127] and GSK126 [128]; similarly, ERG inhibitory peptides (EIPs) specifically target the ERG binding domain [129].
In TNBC, C-techs have not only provided evidence that AP1 mediates the loop of the ZEB2 promoters but also identified AP-1, ZEB2, and ERK/Akt signaling as potential drug targets [77]. Therefore, the AP1 inhibitors AP1-C301-S and SR 11302 [130]; ERK/Akt inhibiting drugs, such as valproic acid and U0126 [131]; and miR-132, which targets ZEB2 [132], may be potential drugs; a phase I/II trial of valproic acid was completed and showed the ability of valproic acid to potentiate the effects of epirubicin [133].
Inhibitors that target key proteins in chromosome loops may be novel drugs for the treatment of hematologic neoplasms. For example, pyrrole-imidazole polyamide 1 can target and decrease the expression of EVI1 [134], while GZD856 [135], ZINC08764498 and ZINC12891610 are unique Bcr–Abl fusion gene inhibitors [136]. Venetoclax inhibits BCL2 (an overexpressed cancer-related gene in BCM) [107], and it has received global approval as an oral medicine while under study in two phase III trials [137].
In melanoma, drugs targeting MITF, such as the HIV1 protease inhibitor nelfinavir, can decrease the formation of loops [138]. BET bromodomain inhibitors, such as JQ1 [139], which can suppress the super-enhancer BRD4 [140], may be developed into treatments for ACC. Moreover, the cancer inhibiting role of DIS3 could be critical for the design of new drugs [96].
However, even with such promising targets and treatments, much work is needed to identify drugs that target each loop of interest.
Conclusions
Long-range interactions play an increasingly significant role in the development of malignant tumors. An increasing number of long-range interactions have been discovered in prostate cancer, breast cancer, hematologic neoplasms and other cancers and may serve as new diagnostic and therapeutic targets.
Currently, we can study interactions between only two fragments of DNA due to the limitations of available techniques. To determine the underlying mechanisms of cancer onset and progression, we must develop tools for broader analysis. The question of how chromosomes fold and loop is fascinating and has attracted the attention of scientists from many fields, including those studying cancer. As discussed above, we know that risk loci and certain key genes are “hot spots” in which chromosome conformation changes can be reliably found. In addition to these sites, many different mutations of oncogenes and tumor suppressor genes have been detected in cancers. We can surmise that typical cancer-related genes may encompass a new category of chromosome-conformation-related protein binding sites; the binding of these sites leads to chromosome conformation changes, subsequent gene function changes and possibly new pathogenic mechanisms. The use of C-techs may also help to identify more new cancer-related genes. We can then concentrate on the development of small molecule drugs that target these new genes or mechanisms because novel targeting drugs are at the forefront of cancer research and greatly contribute to medical development.
It is likely that there are more unknown proteins and RNAs (or known proteins and RNAs with unknown functions) that mediate these interactions; we call these proteins and RNAs new loop organizers. These undiscovered loop organizers might underlie more new mechanisms and pathways. New loop organizers and their foundations alike may yield new diagnostic and therapeutic targets. Moreover, there are many elements involved in forming long-range interactions, including lncRNAs and proteins, that cannot all be targeted simultaneously in new therapies. However, key elements that alter other elements when targeted must be identified to increase the efficacy of oncotherapy, similar to the idea of “one vehicle with multiple satellites”. There have been some attempts to target key lncRNAs in oncotherapy [141], and if we develop new therapies related to high-order chromatin conformations based on this idea, we may create a closer link between chromatin conformation and clinical applications.
Future studies should explore how chromosome conformation changes can become a tool for clinical diagnosis and improved therapies.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (31470757), the ShuGuang Project of Shanghai Municipal Education Commission and Shanghai Education Development Foundation (17SG19), the Outstanding Yong Medical Scholar of Shanghai Municipal Commission of Health and Family Planning(2017YQ067), the Program for Professor of Special Appointment (Eastern Scholar) at the Shanghai Institutions of Higher Learning (1410000159), the SMC-ChenXing Yong Scholar Program (2014, Class B), the Shanghai Municipal Education Commission-Gaofeng Clinical Medicine Grant (20161317), the Science and Technology Commission of Shanghai (17DZ2260100) and the Outstanding Yong Scholar Grant of Shanghai JiaoTong University School of Medicine (16XJ11002).
Availability of data and materials
Not applicable.
Abbreviations
- 3C
Chromosome conformation capture
- 4C
Circular chromosome conformation capture
- 5C
Chromosome conformation capture carbon copy
- ADPC
Androgen-dependent prostate cancer
- ALL
Acute lymphoblastic leukemia
- AML
Acute myelocytic leukemia
- AR
Androgen receptor
- BCAS1-4
Breast carcinoma amplified sequence
- BCM
B-cell malignancies
- ChIA-PET
Chromatin interaction analysis by paired-end tag sequencing
- CHIP
Chromatin immunoprecipitation
- CML
Chronic myelogenous leukemia
- CRPC
Castration-resistant prostate cancer
- CTCF
CCCTC-binding factor
- CUX1
Cut-like homeobox 1
- E2
Estradiol
- e4C
Enhanced chromosome conformation capture-on-ChIP
- EGFR
Epidermal growth factor receptor
- ERG
ETS-related gene
- ERα
Estrogen receptor α
- FISH
Fluorescence in situ hybridization
- FoxA1
Forkhead box A1
- GAM
Genome architecture mapping
- GATA1
GATA-binding factor 1
- GWAS
Genome-wide association studies
- H3K27me3
Histone 3 lysine 27 trimethylation
- H3K36me3
Histone 3 lysine 36 trimethylation
- H3K4me1
Histone 3 lysine 4 methylation
- H3K4me2
Histone 3 lysine 4 dimethylation
- H3K4me3
Histone 3 lysine 4 trimethylation
- H3K79me2
Histone 3 lysine 79 dimethylation
- H3K9me3
Histone 3 lysine 9 trimethylation
- H4K20me1
Histone 4 lysine 20 methylation
- Hi-C
High-resolution chromosome conformation capture
- HKP
Histidine-lysine peptide
- IGF2
Insulin-like growth factor 2
- IGFBP3
Insulin-like growth factor-binding protein 3
- LMA
Ligation-mediated amplification
- lncRNAs
Long noncoding RNA
- MED1
Mediator complex subunit 1
- MITF
Microphthalmia-associated transcription factor
- NGS
Next-generation sequencing
- PCR
Polymerase chain reaction
- Pol II
Polymerase II
- PSA
Prostate-specific antigen
- PTC
Papillary thyroid carcinoma
- R3C
RNA-guided chromatin conformation capture
- RUNX1
Runt-related transcription factor 1
- SATB1
Special AT-rich sequence-binding protein-1
- SNPs
Single-nucleotide polymorphisms
- T2C
Targeted chromatin capture
- TAD
Topologically associated domain
- TBP
TATA-box binding protein
- TF
Transcription factor
- TFF1
Trefoil factor 1
- TNBC
Triple-negative breast cancer
- TNFα
Tumor necrosis factor alpha
- UBE2C
Ubiquitin-conjugating enzyme E2 C
- ZEB2
Zinc finger E-box-binding homeobox 2
Authors’ contributions
HZ and XF provided direction and guidance throughout the preparation of this manuscript. RJ collected and interpreted studies and was a major contributor to the writing and editing of the manuscript. PC reviewed and made significant revisions to the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
He Zhang, Email: zhanghe@sjtu.edu.cn.
Xianqun Fan, Email: fanxq@sjtu.edu.cn.
References
- 1.Bascom G, Schlick T. Linking chromatin fibers to gene folding by hierarchical looping. Biophys J. 2017;112(3):434–445. doi: 10.1016/j.bpj.2017.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cullen KE, Kladde MP, Seyfred MA. Interaction between transcription regulatory regions of prolactin chromatin. Science. 1993;261(5118):203–206. doi: 10.1126/science.8327891. [DOI] [PubMed] [Google Scholar]
- 3.Dekker J, Rippe K, Dekker M, Kleckner N. Capturing chromosome conformation. Science. 2002;295(5558):1306–1311. doi: 10.1126/science.1067799. [DOI] [PubMed] [Google Scholar]
- 4.Palstra RJ, Tolhuis B, Splinter E, Nijmeijer R, Grosveld F, de Laat W. The beta-globin nuclear compartment in development and erythroid differentiation. Nat Genet. 2003;35(2):190–194. doi: 10.1038/ng1244. [DOI] [PubMed] [Google Scholar]
- 5.Tolhuis B, Palstra RJ, Splinter E, Grosveld F, de Laat W. Looping and interaction between hypersensitive sites in the active beta-globin locus. Mol Cell. 2002;10(6):1453–1465. doi: 10.1016/s1097-2765(02)00781-5. [DOI] [PubMed] [Google Scholar]
- 6.Salem T, Gomard T, Court F, Moquet-Torcy G, Brockly F, Forne T, et al. Chromatin loop organization of the junb locus in mouse dendritic cells. Nucleic Acids Res. 2013;41(19):8908–8925. doi: 10.1093/nar/gkt669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Comet I, Schuettengruber B, Sexton T, Cavalli G. A chromatin insulator driving three-dimensional Polycomb response element (PRE) contacts and Polycomb association with the chromatin fiber. Proc Natl Acad Sci U S A. 2011;108(6):2294–2299. doi: 10.1073/pnas.1002059108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tan-Wong SM, French JD, Proudfoot NJ, Brown MA. Dynamic interactions between the promoter and terminator regions of the mammalian BRCA1 gene. Proc Natl Acad Sci U S A. 2008;105(13):5160–5165. doi: 10.1073/pnas.0801048105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ramani V, Cusanovich DA, Hause RJ, Ma W, Qiu R, Deng X, et al. Mapping 3D genome architecture through in situ DNase hi-C. Nat Protoc. 2016;11(11):2104–2121. doi: 10.1038/nprot.2016.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Harada A, Mallappa C, Okada S, Butler JT, Baker SP, Lawrence JB, et al. Spatial re-organization of myogenic regulatory sequences temporally controls gene expression. Nucleic Acids Res. 2015;43(4):2008–2021. doi: 10.1093/nar/gkv046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Spilianakis CG, Lalioti MD, Town T, Lee GR, Flavell RA. Interchromosomal associations between alternatively expressed loci. Nature. 2005;435(7042):637–645. doi: 10.1038/nature03574. [DOI] [PubMed] [Google Scholar]
- 12.Zhao Z, Tavoosidana G, Sjolinder M, Gondor A, Mariano P, Wang S, et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nat Genet. 2006;38(11):1341–1347. doi: 10.1038/ng1891. [DOI] [PubMed] [Google Scholar]
- 13.Simonis M, Klous P, Splinter E, Moshkin Y, Willemsen R, de Wit E, et al. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C) Nat Genet. 2006;38(11):1348–1354. doi: 10.1038/ng1896. [DOI] [PubMed] [Google Scholar]
- 14.Sexton T, Kurukuti S, Mitchell JA, Umlauf D, Nagano T, Fraser P. Sensitive detection of chromatin coassociations using enhanced chromosome conformation capture on chip. Nat Protoc. 2012;7(7):1335–1350. doi: 10.1038/nprot.2012.071. [DOI] [PubMed] [Google Scholar]
- 15.Brouwer RW, van den Hout MC, van IWF, Soler E, Stadhouders R. Unbiased interrogation of 3D genome topology using chromosome conformation capture coupled to high-throughput sequencing (4C-Seq) Methods Mol Biol. 2017;1507:199–220. doi: 10.1007/978-1-4939-6518-2_15. [DOI] [PubMed] [Google Scholar]
- 16.Dixon JR, Selvaraj S, Yue F, Kim A, Li Y, Shen Y, et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 2012;485(7398):376–380. doi: 10.1038/nature11082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ferraiuolo MA, Sanyal A, Naumova N, Dekker J, Dostie J. From cells to chromatin: capturing snapshots of genome organization with 5C technology. Methods. 2012;58(3):255–267. doi: 10.1016/j.ymeth.2012.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dostie J, Dekker J. Mapping networks of physical interactions between genomic elements using 5C technology. Nat Protoc. 2007;2(4):988–1002. doi: 10.1038/nprot.2007.116. [DOI] [PubMed] [Google Scholar]
- 19.Lieberman-Aiden E, van Berkum NL, Williams L, Imakaev M, Ragoczy T, Telling A, et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 2009;326(5950):289–293. doi: 10.1126/science.1181369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Flyamer IM, Gassler J, Imakaev M, Brandao HB, Ulianov SV, Abdennur N, et al. Single-nucleus hi-C reveals unique chromatin reorganization at oocyte-to-zygote transition. Nature. 2017;544(7648):110–114. doi: 10.1038/nature21711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Belaghzal H, Dekker J, Gibcus JH. Hi-C 2.0: an optimized hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods. 2017;123:56–65. doi: 10.1016/j.ymeth.2017.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ramani V, Deng X, Qiu R, Gunderson KL, Steemers FJ, Disteche CM, et al. Massively multiplex single-cell hi-C. Nat Methods. 2017;14(3):263–266. doi: 10.1038/nmeth.4155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Handoko L, Xu H, Li G, Ngan CY, Chew E, Schnapp M, et al. CTCF-mediated functional chromatin interactome in pluripotent cells. Nat Genet. 2011;43(7):630–638. doi: 10.1038/ng.857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li G, Ruan X, Auerbach RK, Sandhu KS, Zheng M, Wang P, et al. Extensive promoter-centered chromatin interactions provide a topological basis for transcription regulation. Cell. 2012;148(1-2):84–98. doi: 10.1016/j.cell.2011.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li G, Chen Y, Snyder MP, Zhang MQ. ChIA-PET2: a versatile and flexible pipeline for ChIA-PET data analysis. Nucleic Acids Res. 2017;45(1):e45. doi: 10.1093/nar/gkw809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hughes JR, Roberts N, McGowan S, Hay D, Giannoulatou E, Lynch M, et al. Analysis of hundreds of cis-regulatory landscapes at high resolution in a single, high-throughput experiment. Nat Genet. 2014;46(2):205–212. doi: 10.1038/ng.2871. [DOI] [PubMed] [Google Scholar]
- 27.Kolovos P, van de Werken HJ, Kepper N, Zuin J, Brouwer RW, Kockx CE, et al. Targeted chromatin capture (T2C): a novel high resolution high throughput method to detect genomic interactions and regulatory elements. Epigenetics Chromatin. 2014;7:10. doi: 10.1186/1756-8935-7-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wachsmuth M, Knoch TA, Rippe K. Dynamic properties of independent chromatin domains measured by correlation spectroscopy in living cells. Epigenetics Chromatin. 2016;9:57. doi: 10.1186/s13072-016-0093-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cairns J, Freire-Pritchett P, Wingett SW, Varnai C, Dimond A, Plagnol V, et al. CHiCAGO: robust detection of DNA looping interactions in capture hi-C data. Genome Biol. 2016;17(1):127. doi: 10.1186/s13059-016-0992-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhang H, Zeitz MJ, Wang H, Niu B, Ge S, Li W, et al. Long noncoding RNA-mediated intrachromosomal interactions promote imprinting at the Kcnq1 locus. J Cell Biol. 2014;204(1):61–75. doi: 10.1083/jcb.201304152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kang L, Sun J, Wen X, Cui J, Wang G, Hoffman AR, et al. Aberrant allele-switch imprinting of a novel IGF1R intragenic antisense non-coding RNA in breast cancers. Eur J Cancer. 2015;51(2):260–270. doi: 10.1016/j.ejca.2014.10.031. [DOI] [PubMed] [Google Scholar]
- 32.Dekker J. Mapping the 3D genome: aiming for consilience. Nat Rev Mol Cell Biol. 2016;17(12):741–742. doi: 10.1038/nrm.2016.151. [DOI] [PubMed] [Google Scholar]
- 33.Beagrie RA, Scialdone A, Schueler M, Kraemer DC, Chotalia M, Xie SQ, et al. Complex multi-enhancer contacts captured by genome architecture mapping. Nature. 2017;543(7646):519–524. doi: 10.1038/nature21411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu X, Zhang Y, Chen Y, Li M, Zhou F, Li K, et al. In situ capture of chromatin interactions by Biotinylated dCas9. Cell. 2017;170(5):1028–1043. doi: 10.1016/j.cell.2017.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wei J, Czapla L, Grosner MA, Swigon D, Olson WK. DNA topology confers sequence specificity to nonspecific architectural proteins. Proc Natl Acad Sci U S A. 2014;111(47):16742–16747. doi: 10.1073/pnas.1405016111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Diesinger PM, Kunkel S, Langowski J, Heermann DW. Histone depletion facilitates chromatin loops on the kilobasepair scale. Biophys J. 2010;99(9):2995–3001. doi: 10.1016/j.bpj.2010.08.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shore D, Baldwin RL. Energetics of DNA twisting. I. Relation between twist and cyclization probability. J Mol Biol. 1983;170(4):957–981. doi: 10.1016/s0022-2836(83)80198-3. [DOI] [PubMed] [Google Scholar]
- 38.Polikanov YS, Bondarenko VA, Tchernaenko V, Jiang YI, Lutter LC, Vologodskii A, et al. Probability of the site juxtaposition determines the rate of protein-mediated DNA looping. Biophys J. 2007;93(8):2726–2731. doi: 10.1529/biophysj.107.111245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sanyal A, Lajoie BR, Jain G, Dekker J. The long-range interaction landscape of gene promoters. Nature. 2012;489(7414):109–113. doi: 10.1038/nature11279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rao SS, Huntley MH, Durand NC, Stamenova EK, Bochkov ID, Robinson JT, et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell. 2014;159(7):1665–1680. doi: 10.1016/j.cell.2014.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Du Z, Zheng H, Huang B, Ma R, Wu J, Zhang X, et al. Allelic reprogramming of 3D chromatin architecture during early mammalian development. Nature. 2017;547(7662):232–235. doi: 10.1038/nature23263. [DOI] [PubMed] [Google Scholar]
- 42.Ke Y, Xu Y, Chen X, Feng S, Liu Z, Sun Y, et al. 3D chromatin structures of mature gametes and structural reprogramming during mammalian embryogenesis. Cell. 2017;170(2):367–381. doi: 10.1016/j.cell.2017.06.029. [DOI] [PubMed] [Google Scholar]
- 43.Nagano T, Lubling Y, Várnai C, Dudley C, Leung W, Baran Y, et al. Cell-cycle dynamics of chromosomal organization at single-cell resolution. Nature. 2017;547(7661):61–67. doi: 10.1038/nature23001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Vietri Rudan M, Barrington C, Henderson S, Ernst C, Odom DT, Tanay A, et al. Comparative hi-C reveals that CTCF underlies evolution of chromosomal domain architecture. Cell Rep. 2015;10(8):1297–1309. doi: 10.1016/j.celrep.2015.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Guo Y, Xu Q, Canzio D, Shou J, Li J, Gorkin DU, et al. CRISPR inversion of CTCF sites alters genome topology and enhancer/promoter function. Cell. 2015;162(4):900–910. doi: 10.1016/j.cell.2015.07.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mora A, Sandve GK, Gabrielsen OS, Eskeland R. In the loop: promoter-enhancer interactions and bioinformatics. Brief Bioinform. 2016;17(6):980–995. doi: 10.1093/bib/bbv097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hansen AS, Pustova I, Cattoglio C, Tjian R, Darzacq X. CTCF and cohesin regulate chromatin loop stability with distinct dynamics. Elife. 2017;6. https://elifesciences.org/articles/25776. [DOI] [PMC free article] [PubMed]
- 48.Carroll JS, Liu XS, Brodsky AS, Li W, Meyer CA, Szary AJ, et al. Chromosome-wide mapping of estrogen receptor binding reveals long-range regulation requiring the forkhead protein FoxA1. Cell. 2005;122(1):33–43. doi: 10.1016/j.cell.2005.05.008. [DOI] [PubMed] [Google Scholar]
- 49.Hu Q, Kwon YS, Nunez E, Cardamone MD, Hutt KR, Ohgi KA, et al. Enhancing nuclear receptor-induced transcription requires nuclear motor and LSD1-dependent gene networking in interchromatin granules. Proc Natl Acad Sci U S A. 2008;105(49):19199–19204. doi: 10.1073/pnas.0810634105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Tan SK, Lin ZH, Chang CW, Varang V, Chng KR, Pan YF, et al. AP-2gamma regulates oestrogen receptor-mediated long-range chromatin interaction and gene transcription. EMBO J. 2011;30(13):2569–2581. doi: 10.1038/emboj.2011.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang Y, Wong CH, Birnbaum RY, Li G, Favaro R, Ngan CY, et al. Chromatin connectivity maps reveal dynamic promoter-enhancer long-range associations. Nature. 2013;504(7479):306–310. doi: 10.1038/nature12716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chen Z, Zhang C, Wu D, Chen H, Rorick A, Zhang X, et al. Phospho-MED1-enhanced UBE2C locus looping drives castration-resistant prostate cancer growth. EMBO J. 2011;30(12):2405–2419. doi: 10.1038/emboj.2011.154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Arthur RK, An N, Khan S, McNerney ME. The haploinsufficient tumor suppressor, CUX1, acts as an analog transcriptional regulator that controls target genes through distal enhancers that loop to target promoters. Nucleic Acids Res. 2017;45(11):6350–361. doi:10.1093/nar/gkx218. [DOI] [PMC free article] [PubMed]
- 54.Riley KJ, Steitz JA. The "observer effect" in genome-wide surveys of protein-RNA interactions. Mol Cell. 2013;49(4):601–604. doi: 10.1016/j.molcel.2013.01.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hadjur S, Williams LM, Ryan NK, Cobb BS, Sexton T, Fraser P, et al. Cohesins form chromosomal cis-interactions at the developmentally regulated IFNG locus. Nature. 2009;460(7253):410–413. doi: 10.1038/nature08079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang H, Li W, Guo R, Sun J, Cui J, Wang G, et al. An intragenic long noncoding RNA interacts epigenetically with the RUNX1 promoter and enhancer chromatin DNA in hematopoietic malignancies. Int J Cancer. 2014;135(12):2783–2794. doi: 10.1002/ijc.28922. [DOI] [PubMed] [Google Scholar]
- 57.Wu D, Zhang C, Shen Y, Nephew KP, Wang Q. Androgen receptor-driven chromatin looping in prostate cancer. Trends Endocrinol Metab. 2011;22(12):474–480. doi: 10.1016/j.tem.2011.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lin C, Yang L, Tanasa B, Hutt K, Ju BG, Ohgi K, et al. Nuclear receptor-induced chromosomal proximity and DNA breaks underlie specific translocations in cancer. Cell. 2009;139(6):1069–1083. doi: 10.1016/j.cell.2009.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Takayama K, Kaneshiro K, Tsutsumi S, Horie-Inoue K, Ikeda K, Urano T, et al. Identification of novel androgen response genes in prostate cancer cells by coupling chromatin immunoprecipitation and genomic microarray analysis. Oncogene. 2007;26(30):4453–4463. doi: 10.1038/sj.onc.1210229. [DOI] [PubMed] [Google Scholar]
- 60.Wang Q, Li W, Liu XS, Carroll JS, Janne OA, Keeton EK, et al. A hierarchical network of transcription factors governs androgen receptor-dependent prostate cancer growth. Mol Cell. 2007;27(3):380–392. doi: 10.1016/j.molcel.2007.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yu J, Yu J, Mani RS, Cao Q, Brenner CJ, Cao X, et al. An integrated network of androgen receptor, polycomb, and TMPRSS2-ERG gene fusions in prostate cancer progression. Cancer Cell. 2010;17(5):443–454. doi: 10.1016/j.ccr.2010.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wang Q, Carroll JS, Brown M. Spatial and temporal recruitment of androgen receptor and its coactivators involves chromosomal looping and polymerase tracking. Mol Cell. 2005;19(5):631–642. doi: 10.1016/j.molcel.2005.07.018. [DOI] [PubMed] [Google Scholar]
- 63.Elemento O, Rubin MA, Rickman DS. Oncogenic transcription factors as master regulators of chromatin topology: a new role for ERG in prostate cancer. Cell Cycle. 2012;11(18):3380–3383. doi: 10.4161/cc.21401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rickman DS, Soong TD, Moss B, Mosquera JM, Dlabal J, Terry S, et al. Oncogene-mediated alterations in chromatin conformation. Proc Natl Acad Sci U S A. 2012;109(23):9083–9088. doi: 10.1073/pnas.1112570109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Taberlay PC, Achinger-Kawecka J, Lun AT, Buske FA, Sabir K, Gould CM, et al. Three-dimensional disorganization of the cancer genome occurs coincident with long-range genetic and epigenetic alterations. Genome Res. 2016;26(6):719–731. doi: 10.1101/gr.201517.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yang L, Lin C, Jin C, Yang JC, Tanasa B. Li W, et al. lncRNA-dependent mechanisms of androgen-receptor-regulated gene activation programs. Nature. 2013;500(7464):598–602. doi: 10.1038/nature12451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Scully T, Firth SM, Scott CD, de Silva HC, Pintar JE, Chan-Ling T, et al. Insulin-like growth factor binding protein-3 links obesity and breast cancer progression. Oncotarget. 2016;7(34):55491–55505. doi: 10.18632/oncotarget.10675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zeitz MJ, Ay F, Heidmann JD, Lerner PL, Noble WS, Steelman BN, et al. Genomic interaction profiles in breast cancer reveal altered chromatin architecture. PLoS One. 2013;8(9):e73974. doi: 10.1371/journal.pone.0073974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Theodorou V, Stark R, Menon S, Carroll JS. GATA3 acts upstream of FOXA1 in mediating ESR1 binding by shaping enhancer accessibility. Genome Res. 2013;23(1):12–22. doi: 10.1101/gr.139469.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Holmes KA, Hurtado A, Brown GD, Launchbury R, Ross-Innes CS, Hadfield J, et al. Transducin-like enhancer protein 1 mediates estrogen receptor binding and transcriptional activity in breast cancer cells. Proc Natl Acad Sci U S A. 2012;109(8):2748–2753. doi: 10.1073/pnas.1018863108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Magnani L, Ballantyne EB, Zhang X, Lupien M. PBX1 genomic pioneer function drives ERalpha signaling underlying progression in breast cancer. PLoS Genet. 2011;7(11):e1002368. doi: 10.1371/journal.pgen.1002368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Phillips-Cremins JE, Sauria ME, Sanyal A, Gerasimova TI, Lajoie BR, Bell JS, et al. Architectural protein subclasses shape 3D organization of genomes during lineage commitment. Cell. 2013;153(6):1281–1295. doi: 10.1016/j.cell.2013.04.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schmidt D, Schwalie PC, Ross-Innes CS, Hurtado A, Brown GD, Carroll JS, et al. A CTCF-independent role for cohesin in tissue-specific transcription. Genome Res. 2010;20(5):578–588. doi: 10.1101/gr.100479.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Mourad R, Hsu PY, Juan L, Shen C, Koneru P, Lin H, et al. Estrogen induces global reorganization of chromatin structure in human breast cancer cells. PLoS One. 2014;9(12):e113354. doi: 10.1371/journal.pone.0113354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Barutcu AR, Hong D, Lajoie BR, McCord RP, van Wijnen AJ, Lian JB, et al. RUNX1 contributes to higher-order chromatin organization and gene regulation in breast cancer cells. Biochim Biophys Acta. 2016;1859(11):1389–1397. doi: 10.1016/j.bbagrm.2016.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fullwood MJ, Liu MH, Pan YF, Liu J, Xu H, Mohamed YB, et al. An oestrogen-receptor-alpha-bound human chromatin interactome. Nature. 2009;462(7269):58–64. doi: 10.1038/nature08497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Qiao Y, Shiue CN, Zhu J, Zhuang T, Jonsson P, Wright AP, et al. AP-1-mediated chromatin looping regulates ZEB2 transcription: new insights into TNFalpha-induced epithelial-mesenchymal transition in triple-negative breast cancer. Oncotarget. 2015;6(10):7804–7814. doi: 10.18632/oncotarget.3158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Espinal-Enriquez J, Fresno C, Anda-Jauregui G, Hernandez-Lemus E. RNA-Seq based genome-wide analysis reveals loss of inter-chromosomal regulation in breast cancer. Sci Rep. 2017;7(1):1760. doi: 10.1038/s41598-017-01314-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Groschel S, Sanders MA, Hoogenboezem R, de Wit E, Bouwman BA, Erpelinck C, et al. A single oncogenic enhancer rearrangement causes concomitant EVI1 and GATA2 deregulation in leukemia. Cell. 2014;157(2):369–381. doi: 10.1016/j.cell.2014.02.019. [DOI] [PubMed] [Google Scholar]
- 80.Shugay M, Ortiz de Mendibil I, Vizmanos JL, Novo FJ. Genomic hallmarks of genes involved in chromosomal translocations in hematological cancer. PLoS Comput Biol. 2012;8(12):e1002797. doi: 10.1371/journal.pcbi.1002797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wang Z, Cao R, Taylor K, Briley A, Caldwell C, Cheng J. The properties of genome conformation and spatial gene interaction and regulation networks of normal and malignant human cell types. PLoS One. 2013;8(3):e58793. doi: 10.1371/journal.pone.0058793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sun J, Li W, Sun Y, Yu D, Wen X, Wang H, et al. A novel antisense long noncoding RNA within the IGF1R gene locus is imprinted in hematopoietic malignancies. Nucleic Acids Res. 2014;42(15):9588–9601. doi: 10.1093/nar/gku549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Katainen R, Dave K, Pitkanen E, Palin K, Kivioja T, Valimaki N, et al. CTCF/cohesin-binding sites are frequently mutated in cancer. Nat Genet. 2015;47(7):818–821. doi: 10.1038/ng.3335. [DOI] [PubMed] [Google Scholar]
- 84.Xiang JF, Yin QF, Chen T, Zhang Y, Zhang XO, Wu Z, et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014;24(5):513–531. doi: 10.1038/cr.2014.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kim IS, Heilmann S, Kansler ER, Zhang Y, Zimmer M, Ratnakumar K, et al. Microenvironment-derived factors driving metastatic plasticity in melanoma. Nat Commun. 2017;8:14343. doi: 10.1038/ncomms14343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bastonini E, Jeznach M, Field M, Juszczyk K, Corfield E, Dezfouli M, et al. Chromatin barcodes as biomarkers for melanoma. Pigment Cell Melanoma Res. 2014;27(5):788–800. doi: 10.1111/pcmr.12258. [DOI] [PubMed] [Google Scholar]
- 87.Hamby SE, Reviriego P, Cooper DN, Upadhyaya M, Chuzhanova N. Screening in silico predicted remotely acting NF1 gene regulatory elements for mutations in patients with neurofibromatosis type 1. Hum Genomics. 2013;7:18. doi: 10.1186/1479-7364-7-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Drier Y, Cotton MJ, Williamson KE, Gillespie SM, Ryan RJ, Kluk MJ, et al. An oncogenic MYB feedback loop drives alternate cell fates in adenoid cystic carcinoma. Nat Genet. 2016;48(3):265–272. doi: 10.1038/ng.3502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Martin JS, Xu Z, Reiner AP, Mohlke KL, Sullivan P, Ren B, et al. HUGIn: Hi-C Unifying Genomic Interrogator. Bioinformatics. 2017. doi:10.1093/bioinformatics/btx359 [DOI] [PMC free article] [PubMed]
- 90.Du M, Tillmans L, Gao J, Gao P, Yuan T, Dittmar RL, et al. Chromatin interactions and candidate genes at ten prostate cancer risk loci. Sci Rep. 2016;6:23202. doi: 10.1038/srep23202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Cai M, Kim S, Wang K, Farnham PJ, Coetzee GA, Lu W. 4C-seq revealed long-range interactions of a functional enhancer at the 8q24 prostate cancer risk locus. Sci Rep. 2016;6:22462. doi: 10.1038/srep22462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Tordini F, Aldinucci M, Milanesi L, Lio P, Merelli I. The genome conformation as an integrator of multi-Omic data: the example of damage spreading in cancer. Front Genet. 2016;7:194. doi: 10.3389/fgene.2016.00194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Dryden NH, Broome LR, Dudbridge F, Johnson N, Orr N, Schoenfelder S, et al. Unbiased analysis of potential targets of breast cancer susceptibility loci by capture hi-C. Genome Res. 2014;24(11):1854–1868. doi: 10.1101/gr.175034.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sotelo J, Esposito D, Duhagon MA, Banfield K, Mehalko J, Liao H, et al. Long-range enhancers on 8q24 regulate c-Myc. Proc Natl Acad Sci U S A. 2010;107(7):3001–3005. doi: 10.1073/pnas.0906067107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Jager R, Migliorini G, Henrion M, Kandaswamy R, Speedy HE, Heindl A, et al. Capture hi-C identifies the chromatin interactome of colorectal cancer risk loci. Nat Commun. 2015;6:6178. doi: 10.1038/ncomms7178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Hoskins JW, Ibrahim A, Emmanuel MA, Manmiller SM, Wu Y, O'Neill M, et al. Functional characterization of a chr13q22.1 pancreatic cancer risk locus reveals long-range interaction and allele-specific effects on DIS3 expression. Hum Mol Genet. 2016;25(21):4726–4738. doi: 10.1093/hmg/ddw300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.He H, Li W, Liyanarachchi S, Srinivas M, Wang Y, Akagi K, et al. Multiple functional variants in long-range enhancer elements contribute to the risk of SNP rs965513 in thyroid cancer. Proc Natl Acad Sci U S A. 2015;112(19):6128–6133. doi: 10.1073/pnas.1506255112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ing-Simmons E, Merkenschlager M. Oncometabolite tinkers with genome folding, boosting Oncogene expression. Trends Mol Med. 2016;22(3):185–187. doi: 10.1016/j.molmed.2016.01.008. [DOI] [PubMed] [Google Scholar]
- 99.Baskin R, Woods NT, Mendoza-Fandino G, Forsyth P, Egan KM, Monteiro AN. Functional analysis of the 11q23.3 glioma susceptibility locus implicates PHLDB1 and DDX6 in glioma susceptibility. Sci Rep. 2015;5:17367. doi: 10.1038/srep17367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Litchfield K, Levy M, Orlando G, Loveday C, Law PJ, Migliorini G, et al. Identification of 19 new risk loci and potential regulatory mechanisms influencing susceptibility to testicular germ cell tumor. Nat Genet. 2017;49(7):1133–1140. doi: 10.1038/ng.3896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Harewood L, Kishore K, Eldridge MD, Wingett S, Pearson D, Schoenfelder S, et al. Hi-C as a tool for precise detection and characterisation of chromosomal rearrangements and copy number variation in human tumours. Genome Biol. 2017;18(1):125. doi: 10.1186/s13059-017-1253-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Tomlins SA, Day JR, Lonigro RJ, Hovelson DH, Siddiqui J, Kunju LP, et al. Urine TMPRSS2:ERG plus PCA3 for individualized prostate cancer risk assessment. Eur Urol. 2016;70(1):45–53. doi: 10.1016/j.eururo.2015.04.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Robert G, Jannink S, Smit F, Aalders T, Hessels D, Cremers R, et al. Rational basis for the combination of PCA3 and TMPRSS2:ERG gene fusion for prostate cancer diagnosis. Prostate. 2013;73(2):113–120. doi: 10.1002/pros.22546. [DOI] [PubMed] [Google Scholar]
- 104.Hagglof C, Hammarsten P, Stromvall K, Egevad L, Josefsson A, Stattin P, et al. TMPRSS2-ERG expression predicts prostate cancer survival and associates with stromal biomarkers. PLoS One. 2014;9(2):e86824. doi: 10.1371/journal.pone.0086824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Baccarani M, Deininger MW, Rosti G, Hochhaus A, Soverini S, Apperley JF, et al. European LeukemiaNet recommendations for the management of chronic myeloid leukemia: 2013. Blood. 2013;122(6):872–884. doi: 10.1182/blood-2013-05-501569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Rousseau M, Ferraiuolo MA, Crutchley JL, Wang XQ, Miura H, Blanchette M, et al. Classifying leukemia types with chromatin conformation data. Genome Biol. 2014;15(4):R60. doi: 10.1186/gb-2014-15-4-r60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Law PJ, Sud A, Mitchell JS, Henrion M, Orlando G, Lenive O, et al. Genome-wide association analysis of chronic lymphocytic leukaemia, Hodgkin lymphoma and multiple myeloma identifies pleiotropic risk loci. Sci Rep. 2017;7:41071. doi: 10.1038/srep41071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.McNerney ME, Brown CD, Wang X, Bartom ET, Karmakar S, Bandlamudi C, et al. CUX1 is a haploinsufficient tumor suppressor gene on chromosome 7 frequently inactivated in acute myeloid leukemia. Blood. 2013;121(6):975–983. doi: 10.1182/blood-2012-04-426965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kang Y, Pantel K. Tumor cell dissemination: emerging biological insights from animal models and cancer patients. Cancer Cell. 2013;23(5):573–581. doi: 10.1016/j.ccr.2013.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Davison LJ, Wallace C, Cooper JD, Cope NF, Wilson NK, Smyth DJ, et al. Long-range DNA looping and gene expression analyses identify DEXI as an autoimmune disease candidate gene. Hum Mol Genet. 2012;21(2):322–333. doi: 10.1093/hmg/ddr468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Smemo S, Tena JJ, Kim KH, Gamazon ER, Sakabe NJ, Gomez-Marin C, et al. Obesity-associated variants within FTO form long-range functional connections with IRX3. Nature. 2014;507(7492):371–375. doi: 10.1038/nature13138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Leng Q, Mixson AJ. The neuropilin-1 receptor mediates enhanced tumor delivery of H2K polyplexes. J Gene Med. 2016;18(7):134–144. doi: 10.1002/jgm.2886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Luo Y, Wu JY, Lu MH, Shi Z, Na N, Di JM. Carvacrol alleviates prostate cancer cell proliferation, migration, and invasion through regulation of PI3K/Akt and MAPK signaling pathways. Oxidative Med Cell Longev. 2016;2016:1469693. doi: 10.1155/2016/1469693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Saura C, Roda D, Rosello S, Oliveira M, Macarulla T, Perez-Fidalgo JA, et al. A first-in-human phase I study of the ATP-competitive AKT inhibitor Ipatasertib demonstrates robust and safe targeting of AKT in patients with solid tumors. Cancer Discovery. 2017;7(1):102–113. doi: 10.1158/2159-8290.CD-16-0512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Wei XX, Hsieh AC, Kim W, Friedlander T, Lin AM, Louttit M, et al. A phase I study of abiraterone acetate combined with BEZ235, a dual PI3K/mTOR inhibitor, in metastatic castration resistant prostate cancer. Oncologist. 2017;22(5):503–e43. doi: 10.1634/theoncologist.2016-0432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Fizazi K, Flaig TW, Stockle M, Scher HI, de Bono JS, Rathkopf DE, et al. Does Gleason score at initial diagnosis predict efficacy of abiraterone acetate therapy in patients with metastatic castration-resistant prostate cancer? An analysis of abiraterone acetate phase III trials. Ann Oncol. 2016;27(4):699–705. doi: 10.1093/annonc/mdv545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Thomas JD, Longen CG, Oyer HM, Chen N, Maher CM, Salvino JM, et al. Sigma1 targeting to suppress aberrant androgen receptor signaling in prostate cancer. Cancer Res. 2017;77(9):2439–2452. doi: 10.1158/0008-5472.CAN-16-1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Liu C, Armstrong C, Zhu Y, Lou W, Gao AC. Niclosamide enhances abiraterone treatment via inhibition of androgen receptor variants in castration resistant prostate cancer. Oncotarget. 2016;7(22):32210–32220. doi: 10.18632/oncotarget.8493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Lai KP, Huang CK, Chang YJ, Chung CY, Yamashita S, Li L, et al. New therapeutic approach to suppress castration-resistant prostate cancer using ASC-J9 via targeting androgen receptor in selective prostate cells. Am J Pathol. 2013;182(2):460–473. doi: 10.1016/j.ajpath.2012.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Myung JK, Banuelos CA, Fernandez JG, Mawji NR, Wang J, Tien AH, et al. An androgen receptor N-terminal domain antagonist for treating prostate cancer. J Clin Invest. 2013;123(7):2948–2960. doi: 10.1172/JCI66398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Gong AY, Eischeid AN, Xiao J, Zhao J, Chen D, Wang ZY, et al. miR-17-5p targets the p300/CBP-associated factor and modulates androgen receptor transcriptional activity in cultured prostate cancer cells. BMC Cancer. 2012;12:492. doi: 10.1186/1471-2407-12-492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Schmitt AM, Chang HY. Gene regulation: long RNAs wire up cancer growth. Nature. 2013;500(7464):536–537. doi: 10.1038/nature12548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Zhang A, Zhao JC, Kim J, Fong KW, Yang YA, Chakravarti D, et al. LncRNA HOTAIR enhances the androgen-receptor-mediated transcriptional program and drives castration-resistant prostate cancer. Cell Rep. 2015;13(1):209–221. doi: 10.1016/j.celrep.2015.08.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Guo C, Yeh S, Niu Y, Li G, Zheng J, Li L, et al. Targeting androgen receptor versus targeting androgens to suppress castration resistant prostate cancer. Cancer Lett. 2017;397:133–143. doi: 10.1016/j.canlet.2017.03.022. [DOI] [PubMed] [Google Scholar]
- 125.Grimison PS, Stockler MR, Chatfield M, Thomson DB, Gebski V, Friedlander M, et al. Accelerated BEP for metastatic germ cell tumours: a multicenter phase II trial by the Australian and New Zealand Urogenital and prostate cancer trials group (ANZUP) Ann Oncol. 2014;25(1):143–148. doi: 10.1093/annonc/mdt369. [DOI] [PubMed] [Google Scholar]
- 126.Haffner MC, Aryee MJ, Toubaji A, Esopi DM, Albadine R, Gurel B, et al. Androgen-induced TOP2B-mediated double-strand breaks and prostate cancer gene rearrangements. Nat Genet. 2010;42(8):668–675. doi: 10.1038/ng.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Tan J, Yang X, Zhuang L, Jiang X, Chen W, Lee PL, et al. Pharmacologic disruption of Polycomb-repressive complex 2-mediated gene repression selectively induces apoptosis in cancer cells. Genes Dev. 2007;21(9):1050–1063. doi: 10.1101/gad.1524107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.McCabe MT, Ott HM, Ganji G, Korenchuk S, Thompson C, Van Aller GS, et al. EZH2 inhibition as a therapeutic strategy for lymphoma with EZH2-activating mutations. Nature. 2012;492(7427):108–112. doi: 10.1038/nature11606. [DOI] [PubMed] [Google Scholar]
- 129.Wang X, Qiao Y, Asangani IA, Ateeq B, Poliakov A, Cieslik M, et al. Development of Peptidomimetic inhibitors of the ERG gene fusion product in prostate cancer. Cancer Cell. 2017;31(4):532–548. doi: 10.1016/j.ccell.2017.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Gonzalez-Rubio S, Linares CI, Aguilar-Melero P, Rodriguez-Peralvarez M, Montero-Alvarez JL, de la Mata M, et al. AP-1 inhibition by SR 11302 protects human Hepatoma HepG2 cells from bile acid-induced Cytotoxicity by restoring the NOS-3 expression. PLoS One. 2016;11(8):e0160525. doi: 10.1371/journal.pone.0160525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zhang C, Liu S, Yuan X, Hu Z, Li H, Wu M, et al. Valproic acid promotes human Glioma U87 cells apoptosis and inhibits glycogen Synthase Kinase-3beta through ERK/Akt signaling. Cell Physiol Biochem. 2016;39(6):2173–2185. doi: 10.1159/000447912. [DOI] [PubMed] [Google Scholar]
- 132.Zheng YB, Luo HP, Shi Q, Hao ZN, Ding Y, Wang QS, et al. miR-132 inhibits colorectal cancer invasion and metastasis via directly targeting ZEB2. World J Gastroenterol. 2014;20(21):6515–6522. doi: 10.3748/wjg.v20.i21.6515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Munster P, Marchion D, Bicaku E, Lacevic M, Kim J, Centeno B, et al. Clinical and biological effects of valproic acid as a histone deacetylase inhibitor on tumor and surrogate tissues: phase I/II trial of valproic acid and epirubicin/FEC. Clin Cancer Res. 2009;15(7):2488–2496. doi: 10.1158/1078-0432.CCR-08-1930. [DOI] [PubMed] [Google Scholar]
- 134.Syed J, Pandian GN, Sato S, Taniguchi J, Chandran A, Hashiya K, et al. Targeted suppression of EVI1 oncogene expression by sequence-specific pyrrole-imidazole polyamide. Chem Biol. 2014;21(10):1370–1380. doi: 10.1016/j.chembiol.2014.07.019. [DOI] [PubMed] [Google Scholar]
- 135.Lu X, Zhang Z, Ren X, Wang D, Ding K. Synthesis and identification of GZD856 as an orally bioavailable Bcr-AblT315I inhibitor overcoming acquired imatinib resistance. J Enzyme Inhib Med Chem. 2017;32(1):331–336. doi: 10.1080/14756366.2016.1250757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Krishna P, Sarvagalla S, Madhuri B, Pajaniradje S, Baskaran V, Coumar MS, et al. Identification of Natural inhibitors of Bcr-Abl for the treatment of Chronic Myeloid Leukemia. Chem Biol Drug Des. 2017;90(4):596–608. [DOI] [PubMed]
- 137.Deeks ED. Venetoclax: First Global Approval. Drugs. 2016;76(9):979–987. doi: 10.1007/s40265-016-0596-x. [DOI] [PubMed] [Google Scholar]
- 138.Kim H, Ronai ZA. HIV drug to aid melanoma therapies? Cancer Cell. 2016;29(3):245–246. doi: 10.1016/j.ccell.2016.02.014. [DOI] [PubMed] [Google Scholar]
- 139.Ember SW, Lambert QT, Berndt N, Gunawan S, Ayaz M, Tauro M, et al. Molecular cancer therapeutics. 2017. Potent dual BET bromodomain-kinase inhibitors as value added multi-targeted chemical probes and cancer therapeutics. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Loven J, Hoke HA, Lin CY, Lau A, Orlando DA, Vakoc CR, et al. Selective inhibition of tumor oncogenes by disruption of super-enhancers. Cell. 2013;153(2):320–334. doi: 10.1016/j.cell.2013.03.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Xu S, Wang H, Pan H, Shi Y, Li T, Ge S, et al. ANRIL lncRNA triggers efficient therapeutic efficacy by reprogramming the aberrant INK4-hub in melanoma. Cancer Lett. 2016;381(1):41–48. doi: 10.1016/j.canlet.2016.07.024. [DOI] [PubMed] [Google Scholar]
- 142.Davies JO, Oudelaar AM, Higgs DR, Hughes JR. How best to identify chromosomal interactions: a comparison of approaches. Nat Methods. 2017;14(2):125–134. doi: 10.1038/nmeth.4146. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.