

Abstract
Background
Atherosclerosis is a chronic vascular inflammatory disease. Since even low-level endotoxemia constitutes a powerful and independent risk factor for the development of atherosclerosis, it is important to find therapies directed against the vascular effects of endotoxin to prevent atherosclerosis. Taraxacum officinale (TO) is used for medicinal purposes because of its choleretic, diuretic, antioxidative, anti-inflammatory, and anti-carcinogenic properties, but its anti-inflammatory effect on endothelial cells has not been established.
Methods
We evaluated the anti-inflammatory activity of TO filtered methanol extracts in LPS-stimulated human umbilical vein endothelial cells (HUVECs) by monocyte adhesion and western blot assays. HUVECs were pretreated with 100 μg/ml TO for 1 h and then incubated with 1 μg/ml LPS for 24 h. The mRNA and protein expression levels of the targets (pro-inflammatory cytokines and adhesion molecules) were analyzed by real-time PCR and western blot assays. We also preformed HPLC analysis to identify the components of the TO methanol extract.
Results
The TO filtered methanol extracts dramatically inhibited LPS-induced endothelial cell–monocyte interactions by reducing vascular cell adhesion molecule-1 and monocyte chemoattractant protein-1, and pro-inflammatory cytokine expression. TO suppressed the LPS-induced nuclear translocation of NF-κB, whereas it did not affect MAPK activation.
Conclusions
Our findings demonstrated that methanol extracts of TO could attenuate LPS-induced endothelial cell activation by inhibiting the NF-κB pathway. These results indicate the potential clinical benefits and applications of TO for the prevention of vascular inflammation and atherosclerosis.
Electronic supplementary material
The online version of this article (10.1186/s12906-017-2022-7) contains supplementary material, which is available to authorized users.
Keywords: Taraxacum officinale, Anti-inflammation, Endothelial cell, Endotoxin, NF-κb
Background
Insults to the vasculature can cause a wide range of life-threatening diseases, including stroke, myocardial infarction, hypertension, and chronic kidney disease. Inflammation is emerging as a key contributor to many vascular diseases, and furthermore plays a major role in autoimmune diseases, arthritis, allergic reactions, and cancer.
Besides autoimmune disorders related to vascular inflammation, a more common chronic vascular inflammatory disease is atherosclerosis [1, 2]. Clinical evidence indicates that atherosclerosis is accelerated in patients suffering from large and medium-sized vessel vasculitis [3].
Endotoxemia, the presence of endotoxin in the blood, constitutes a strong risk factor for early atherogenesis in subjects with chronic or recurrent bacterial infections [4]. Even low-level endotoxemia (as low as 50 pg/ml) can be a powerful and independent risk factor for the development of atherosclerosis [5] through multiple mechanisms, including increases in reactive oxygen species, chemotactic and pro-inflammatory cytokines, and adhesion molecules. Therefore, it is important to find therapies directed against the vascular effects of endotoxin to prevent atherosclerosis in humans.
Most therapy regimens for treating atherosclerosis aim at modulating hypertension and hyperlipidemia or controlling hemostasis in order to avoid thrombotic complications [6]. However, many of these modalities often neglect the role of inflammation in atherosclerosis [6]. As these therapies remain insufficient in reducing the burden of atherosclerosis-related mortality, other approaches like complementary and alternative medicine have recently started to gain more attention [7, 8].
Taraxacum officinale (TO), commonly known as dandelion, is used for medicinal purposes because of its choleretic, diuretic, antioxidative, anti-inflammatory, and anti-carcinogenic properties [9, 10]. The anti-inflammatory effects of extracts of TO or its single components have been reported in both in vitro and animal models. TO extracts (100 and 1000 μg/ml) were demonstrated to inhibit lipopolysaccharide (LPS)-induced tumor necrosis factor-alpha (TNF-α) production in rat astrocytes by inhibiting interleukin-1 (IL-1) production [11]. Luteolin and luteolin-7-O-glucoside, two active components from TO flower extracts, significantly suppressed the production of both inducible nitric oxide synthase and cyclooxygenase-2 in LPS-activated mouse macrophage RAW264.7 cells [12]. Pretreatment with TO extracts also protected against LPS-induced acute lung injury in mice [13]. However, the effect of TO on endothelial activation has not been established.
Our study shows that TO reduces the expression of vascular cell adhesion molecule-1 (VCAM-1) and pro-inflammatory cytokines in endothelial cells, and decreases mononuclear cell adhesion by suppressing nuclear factor-kappa B (NF-κB) signaling.
Methods
Reagents
LPS, dimethyl sulfoxide (DMSO), and Hoechst 33,258 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Calcein AM was obtained from Thermo Fisher Scientific (Waltham, MA, USA). Antibodies against VCAM-1, intercellular adhesion molecule-1 (ICAM-1), β-actin, NF-κB p65, inhibitor of NF-κB alpha (IκBα), and phospho-IκBα were obtained from Cell Signaling Technology (Danvers, MA, USA).
Methanol extracts of Taraxacum officinale
Methanol extracts (code numbers: PB5027.2) from whole plants of TO were purchased from the Plant Extract Bank at the Korea Research Institute of Bioscience and Biotechnology (Daejeon, Republic of Korea; http://extract.kribb.re.kr; E-mail: plantext@kribb.re.kr) [14, 15]. The extracts were dissolved in DMSO at a concentration of 100 mg/ml.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were obtained from ScienCell Research Laboratories (San Diego, CA, USA) [16]. The cells were cultured in endothelial cell medium (ScienCell Research Laboratories) containing 5% (v/v) fetal bovine serum, at 37 °C under an atmosphere with 5% (v/v) CO2 and 95% humidity.
Cytotoxicity assay
Cell viability was assessed by using the CellTilter 96 Aqueous One Solution Cell Proliferation Assay (Promega Corporation, Madison, WI, USA), according to the manufacturer’s instructions. Cells were seeded at a density of 1 × 104 cells/well into 96-well plates. After their subjection to different treatments, the cells were incubated with 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H–tetrazolium, inner salt (MTS) solution at a final concentration of 0.4 mg/ml for 4 h at 37 °C. One Solution Reagent was then added directly to the culture wells, and the plates were incubated for 4 h, following which the absorbance at 490 nm was recorded with a 96-well plate reader [17, 18]. The absorbance was measured at 490 nm with a Multiskan GO microplate spectrophotometer (Thermo Fisher Scientific).
Western blot analysis
Cells were washed with ice-cold PBS and lysed on ice in RIPA lysis buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Nonidet P-40, 0.1% sodium dodecyl sulfate, and 0.5% sodium deoxycholate) supplemented with protease and phosphatase inhibitors. Aliquots with equal amounts of protein were loaded and separated on a SDS-PAGE gel. The proteins on the gel were then transferred to a nitrocellulose membrane (Bio-Rad, Hercules, CA, USA) and probed using specific antibodies as indicated. The bands were detected by chemiluminescence on a ChemiDoc imaging system (Bio-Rad). To control for sample loading, the blots were subsequently stripped and re-probed for total IκBα or β-actin.
RNA extraction and real-time reverse-transcription polymerase chain reaction
Total RNA was isolated from the cells, using the NucleoSpin RNA Plus Kit (MACHEREY-NAGEL, Düren, Germany), and 1 μg was then used for cDNA synthesis with the iScript cDNA Synthesis Kit (Bio-Rad). The resulting cDNA was PCR amplified with the appropriate primer pairs: MCP-1, 5′-AGAATCACCAGCAGCAAGTGTCC-3′ (forward) and 5′-TCCTGAACCCACTTCTGCTTGG-3′ (reverse); TNF-α, 5′-CTCTTCTGCCTGCTGCACTTTG-3′ (forward) and 5′-ATGGGCTACAGGCTTGTCACTC-3′ (reverse); IL-1β, 5′- CCACAGACCTTCCAGGAGAATG-3′ (forward) and 5′-GTGCAGTTCAGTGATCGTACAGG-3′ (reverse); IL-6, 5′-AGACAGCCACTCACCTCTTCAG-3′ (forward) and 5′- TTCTGCCAGTGCCTCTTTGCTG-3′ (reverse); and 18S rRNA, 5′-AACCCGTTGAACCCCATT-3′ (forward) and 5′-CCATCCAATCGGTAGTAGCG-3′ (reverse) (Bioneer, Daejeon, Republic of Korea). The RT-qPCR was performed and analyzed with a CFX Connect Real-Time PCR detection system (Bio-Rad), and the gene expression levels were normalized to that of 18S rRNA as the housekeeping gene [19].
Analysis of monocyte adhesion to HUVECs
HUVECs were seeded and incubated in 6-well plates until they reached >85% confluence. Subsequently, the cells were pre-incubated with the TO extract (100 μg/ml) for 1 h prior to stimulation with LPS (1 μg/ml) for 24 h. Human monocytic THP-1 cells were labeled with 5 μM Calcein AM for 30 min in RPMI-1640 medium and then added to the HUVEC-containing 6-well plates and incubated for 1 h. Subsequently, unbound monocytes were removed by 3 washes with warm phosphate-buffered saline (PBS) [20]. Bound monocytes were determined using a fluorescence microscope (EVOS FL Cell Imaging System, Thermo Fisher Scientific).
Immunofluorescence analysis of NF-κB p65 nuclear translocation
HUVECs were pre-incubated with the TO extract (100 μg/ml) for 1 h, followed by LPS (1 μg/ml) for 1 h. The cells were then fixed with 4% paraformaldehyde in PBS (pH 7.4) for 20 min at room temperature, and permeabilized with 0.1% Triton X-100. Nonspecific binding was blocked with 5% normal goat serum for 1 h. Then, the cells were incubated overnight with NF-κB p65 antibody at 4 °C, followed by incubation with Alexa Fluor 594-conjugated goat anti-rabbit IgG (Thermo Fisher Scientific) for 1 h at room temperature. Nuclei were stained with Hoechst 33,258. The locations of NF-κB p65 and nuclei were determined using a fluorescence microscope (EVOS FL Cell Imaging System). Results were expressed as the percentage of NF-κB p65-positive cells in the total cells counted from 4 randomly chosen high-power (×20) fields in each well. Each assay was performed in triplicate.
High-performance liquid chromatography analysis
The TO methanol extracts were filtered (Millipore 0.45 μm) and 5 μl was injected into a high-performance liquid chromatography (HPLC) column (Supelcosil LC-18, 25 cm × 4.6 mm × 5 μm), at 25 °C. The extract separation was carried out on an HPLC device composed of Waters 510 pumps, 2489 UV/Vis detector, gradient controller, and Rheodyne injector, with a solvent A and B mixture as follows: methanol:acetic acid:water (10:2:88; solvent A) and methanol:acetic acid:water (90:3:7; solvent B) at a flow of 1 ml/min, with detection at 280 nm. The solvent A/B gradient applied was as follows: A from 100% to 85% (minutes 0–10), A from 85% to 50% (minutes 10–30), A from 50% to 15% (minutes 30–45), and A from 15% to 100% (minutes 45–55) [21]. The identification of peaks was made by comparison with HPLC chromatograms of individual pure phenolic acid (gallic, protocatechuic, chlorogenic, caffeic, p-coumaric, and ferulic acids) standards procured from Sigma-Aldrich.
Statistics
Data were analyzed by analysis of variance (Sigma Stat 12.0) and tested for use of parametric or nonparametric post hoc analysis. Multiple comparisons were performed using the least significant difference method. All data are presented as the mean ± standard error of at least 3 independent experiments. Results were considered statistically significant at the p < 0.05 level.
Results
Effect of TO on HUVEC viability
The cytotoxicity of the TO extract was examined by MTS assay. The viability of HUVECs treated with TO for 24 h was not affected at any of the extract concentrations (50, 100, and 200 μg/ml) tested (Fig. 1).
Fig. 1.
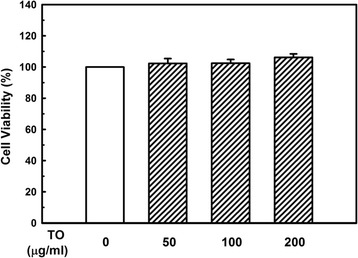
Effect of the Taraxacum officinale (TO) methanol extract on the viability of HUVECs. Cells were treated with different concentrations of the TO extract for 24 h, and viability was determined by the MTS assay. The results of independent experiments were averaged as the percentage cell viability compared with untreated control. Results shown are the mean ± SE of triple determinations
Effect of TO on monocyte adhesion to LPS-stimulated endothelial cells
To determine the effect of TO on endothelial cell–monocyte interactions, we performed adhesion assays. We used human monocytic THP-1 cells to mimic the interaction of monocytes with the endothelium. LPS treatment increased THP-1 cell adherence to the endothelial surface. Pretreatment with the TO extract (100 μg/ml, 1 h) decreased the LPS-induced THP-1 cell adhesion to a near-baseline level, indicating that TO can significantly decrease LPS-induced monocyte adhesion to endothelial cells (Fig. 2).
Fig. 2.
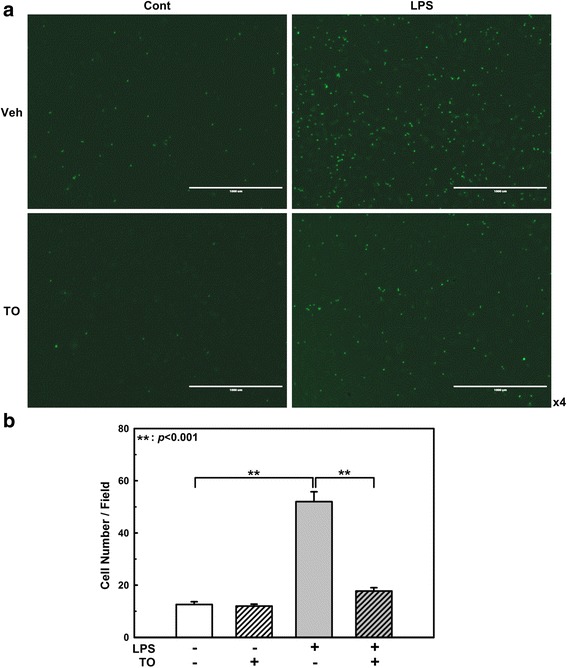
Effect of the Taraxacum officinale (TO) methanol extract on LPS-induced monocyte adhesion to HUVECs. HUVECs were pretreated with TO extract (100 μg/ml) for 1 h, and then stimulated with 1 μg/ml LPS for 24 h. Adhesion of fluorescent THP-1 monocytes was photographed by fluorescence microscopy (a) and analyzed (b). Scale bars: 1000 μm. Results shown are the mean ± SE of 3 independent experiments. **p < 0.01 vs LPS only treatment
Effect of TO on adhesion molecule and inflammatory cytokine expression in LPS-stimulated endothelial cells
Endothelial activation involves the coordinated induction of genes encoding for leukocyte adhesion molecules (e.g., VCAM-1 and ICAM-1), chemotactic factors (e.g., monocyte chemoattractant protein-1 [MCP-1]), and growth factors (e.g., macrophage colony-stimulating factor). Endothelial VCAM-1 is an important mediator of mononuclear cell adhesion, given that its cognate ligand, the integrin very late antigen-4, is selectively expressed on monocytes (and on some T lymphocytes) but not on neutrophils [22]. We therefore examined whether TO treatment decreases VCAM-1 induction in response to LPS stimulation in HUVECs. Pretreatment of TO decreased the LPS-induced VCAM-1 expression by 53% (Fig. 3a). The potency of 100 μg/ml TO was quite compatible with that of 1 mM acetylsalicylic acid (Additional file 1: Figure S1). We also measured the ICAM-1 protein levels but did not observe any changes with TO treatment (Additional file 1: Figure S2).
Fig. 3.
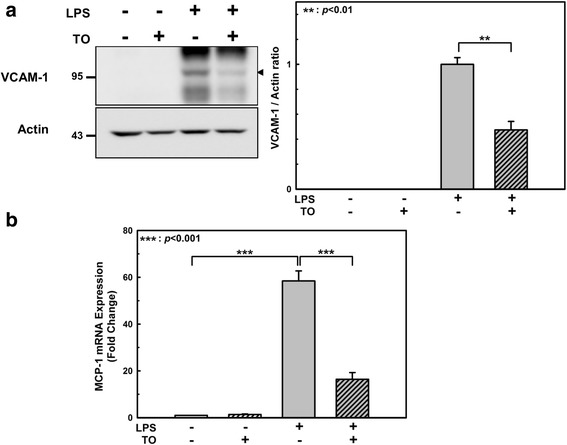
Effect of the Taraxacum officinale (TO) methanol extract on LPS-induced VCAM-1 and MCP-1 expression. HUVECs were pretreated with TO extract (100 μg/ml) for 1 h and then incubated with 1 μg/ml LPS for 24 h. a Total cell lysate was prepared, the proteins were separated by SDS-PAGE, and the VCAM-1 levels were assessed by western blot analysis. b The mRNA levels were normalized to that of a housekeeping gene (18S rRNA) as well as the vehicle-only-treated control, and the 2-ΔΔCt for each mRNA is reported. Results shown are the mean ± SE of 3 independent experiments. **p < 0.01, ***p < 0.001 vs LPS only treatment
Since MCP-1 is one of the key chemokines that regulate leukocyte trafficking and modulate interactions between leukocytes and endothelial cells [23, 24], we next examined whether TO inhibits MCP-1 expression in LPS-stimulated HUVECs. To this end, we used RT-qPCR to examine MCP-1 mRNA expression in HUVECs in response to LPS stimulation. MCP-1 mRNA expression was induced by LPS (58.5-fold), but this effect was significantly decreased (16.4-fold) in HUVECs pretreated with the TO extract (Fig. 3b).
We also measured pro-inflammatory cytokine mRNA expression levels by RT-qPCR. TNF-α, IL-1β, and IL-6 were induced by 9-, 157-, and 30-fold, respectively (Fig. 4). However, these pro-inflammatory cytokine mRNA expression levels were markedly reduced by TO pretreatment (Fig. 4).
Fig. 4.
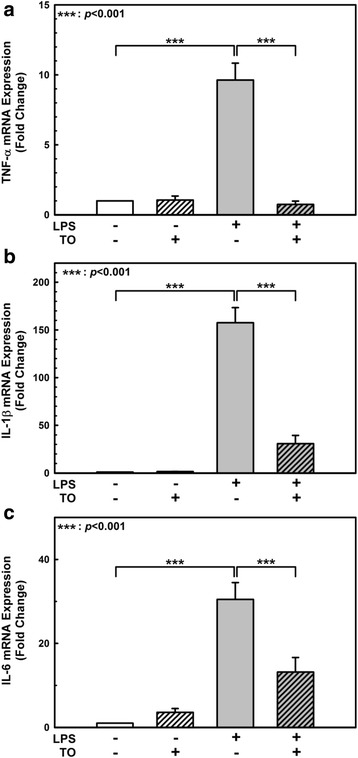
Effect of the Taraxacum officinale (TO) methanol extract on LPS-induced pro-inflammatory cytokine expression. The pro-inflammatory cytokine mRNA levels were normalized to that of a housekeeping gene (18S rRNA) as well as the vehicle-only-treated control, and the 2-ΔΔCt for each mRNA is reported. Results shown are the mean ± SE of 3 independent experiments (a, b, c). ***p < 0.001 vs LPS only treatment
Effect of TO on NF-κB activation in LPS-stimulated endothelial cells
LPS stimulation can lead to the activation of several intracellular signaling molecules. It has been reported that LPS can activate the mitogen-activated protein kinase (MAPK) [25, 26] and NF-κB pathways in endothelial cells [27]. These pathways are involved in controlling the expression of adhesion molecules and pro-inflammatory cytokines. Therefore, we first examined whether the TO extract inhibits MAPK (Erk1/2, p38MAPK, and JNK) activation in response to LPS stimulation. Western blot analysis indicated that TO did not markedly affect LPS-induced MAPK activation (Additional file 1: Figure S3).
To determine whether TO regulates VCAM-1 and pro-inflammatory cytokine expression by inhibiting NF-κB, we performed immunofluorescence staining of the NF-κB p65 subunit and showed that TO markedly suppresses LPS-induced NF-κB nuclear translocation (Fig. 5a and b). Since NF-κB nuclear translocation is regulated by IκBα, the degradation of which is triggered by phosphorylation, we studied whether TO treatment was associated with the inhibition of IκBα phosphorylation. As shown in Fig. 5c, TO pretreatment inhibited IκBα phosphorylation by 65%, and the degradation of IκBα induced by LPS in 30 min was also partially recovered by TO treatment. These results suggest that TO reduces the LPS-induced endothelial expression of VCAM-1 and pro-inflammatory cytokines by suppressing activation of the NF-κB pathways.
Fig. 5.
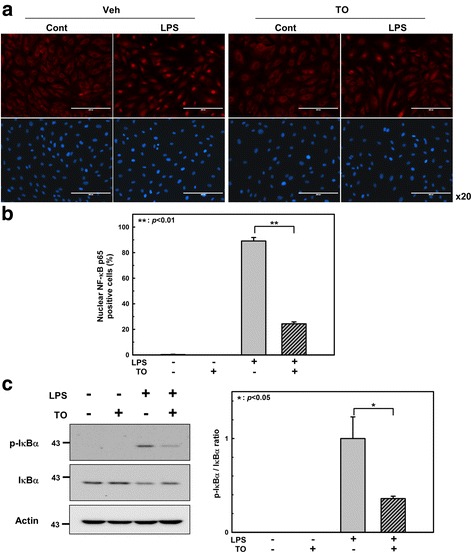
Effect of the Taraxacum officinale (TO) methanol extract on the LPS-induced nuclear translocation of NF-κB p65 in HUVECs. HUVECs were pretreated with TO extract (100 μg/ml) for 1 h and then stimulated with LPS (1 μg/ml) for 1 h. a Representative images of immunofluorescence staining showing NF-κB p65 (red) and cell nuclei stained with Hoechst 33,258 (blue). Scale bars: 200 μm. b Quantitation of NF-κB p65 nuclear translocation in the indicated groups. c IκBα phosphorylation assessed by western blot analysis. Results are shown as the mean ± SE (n = 3–4). *p < 0.05, **p < 0.01 vs LPS only treatment
HPLC analysis of the TO methanol extract
Several peaks were monitored in the HPLC profile of the TO extract (Fig. 6). By comparing the chromatographic peaks of TO with those of the reference standards, protocatechuic acid, chlorogenic acid, caffeic acid, p-coumaric acid, and ferulic acid were qualitatively identified in the extract. The other peaks of the extract could not be identified owing to a lack of authentic references to compare with.
Fig. 6.
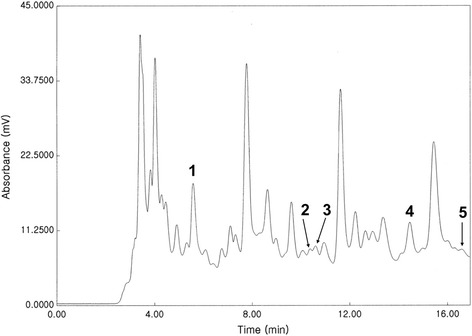
HPLC chromatogram of the Taraxacum officinale methanol extract. 1. Protocatechuic acid; 2. Chlorogenic acid; 3. Caffeic acid; 4. p-Coumaric acid; and 5. Ferulic acid. The other peaks were not identified owing to a lack of authentic compounds to compare with
Discussion
Inflammation is a set of interrelated processes in response to injuries caused by a variety of biological, chemical, and physical stimuli [28]. Vascular endothelial cells form an interface between blood flow and the vessel wall, and execute a number of important functions in the maintenance of the body’s homeostasis [29]. Not only does the endothelium provide a nonadhesive and highly selective physical barrier to control the vascular permeability, but it also secretes a large number of vasoactive substances to regulate the vascular tone and remodeling of the vessel wall [30]. Most importantly, as the key regulators and major targets of the inflammatory process, endothelial cells are indispensable components of inflammation. Endothelial cells are constantly exposed to various biological, chemical, and mechanical milieus, and maintain a quiescent state with antithrombotic, anti-inflammatory, and antiproliferative properties [31]. During inflammatory responses, endothelial cells are phenotypically converted into an “activated” state that is characterized by increased permeability, induced leukocyte adhesion, and gene expression of a variety of pro-inflammatory cytokines [32]. Endothelial cell activation leads to endothelial dysfunction, which can be caused by several conditions, including various infections, diabetes or the metabolic syndrome, hypertension, smoking, and physical inactivity [33, 34].
Atherosclerosis is a chronic inflammatory disease characterized by monocyte infiltration and macrophage accumulation in the vessel wall [35]. One potentially important source of inflammation is endotoxin (LPS), a unique glycolipid that comprises most of the outer leaflet of the outer wall of gram-negative bacteria [36, 37]. The Bruneck study provided the first epidemiological evidence that subclinical endotoxemia constitutes a strong risk factor for the development of carotid atherosclerosis, particularly among smokers [5]. A 5-year prospective study showed that in subjects without atherosclerosis at baseline, ~40% of newly developed carotid atherosclerosis was attributable to chronic infection, making it a leading atherogenic risk predictor [4]. Moreover, chronic infections caused by gram-negative bacteria conferred an increased risk of atherosclerosis development, even in low-risk subjects who lacked conventional vascular risk factors [4]. Even if there is no apparent infection source, a high-fat diet augments plasma LPS (“metabolic endotoxemia”) to a concentration sufficient to trigger inflammation and metabolic diseases, such as obesity and diabetes [38, 39], by increased intestinal permeability, favoring translocation of microbiome-derived LPS to the bloodstream [38]. In addition, the plasma LPS level is also markedly increased in diabetic patients compared with that in non-diabetic subjects [40, 41]. These observations support the hypothesis that chronic exposure to endotoxins may be pathogenically linked to atherosclerosis.
TO has long been used as a herbal remedy to treat medical problems, including inflammatory disease [10, 42–44]. The anti-inflammatory effects of TO extracts or its single components have been reported in both in vitro and animal models [11, 13, 45–47]. Among the identified components in our HPLC analysis, protocatechuic acid [48, 49], chlorogenic acid [50], caffeic acid [51, 52], and ferulic acid [53, 54] have showed anti-inflammatory activity in the endothelial system. Recently, Hu et al. reported that aqueous extracts of TO inhibited both TNF-α and ICAM-1 expression in LPS-stimulated rat mammary microvascular endothelial cells [55]. However, the authors did not identify any underlying molecular mechanisms of the TO extracts.
In this study, we have shown that the anti-inflammatory effect of the TO methanol extract on human endothelial cells is mediated through its reduction of VCAM-1 and pro-inflammatory cytokine expression. Since endothelial VCAM-1 is an important mediator of mononuclear cell (monocytes and some T lymphocytes) adhesion, our finding of the reduced VCAM-1 expression explains the significantly inhibited monocyte adhesion to LPS-stimulated endothelial cells. We also examined whether TO reduces ICAM-1 induction in LPS-stimulated endothelial cells, but could not repeat the findings of Hu et al. [55] in this study. This might be due to the difference in solvents used for the TO extraction, because the solvent type is one of the most common factors affecting bioactive compounds in extraction processes [56, 57]. Since acetylsalicylic acid, also known as Aspirin, is reported to suppress endothelial VCAM-1 induction [58], we compared the potency of TO with acetylsalicylic acid on VCAM-1 induction in LPS-stimulated HUVECs (Additional file 1: Figure S1). At 1 mM, acetylsalicylic acid was significantly more potent than 100 μg/ml of TO at suppressing LPS-stimulated VCAM-1 induction, although 500 μM of acetylsalicylic acid only inhibited VCAM-1 induction by 18% (data not shown).
To elucidate the underlying molecular mechanism of the TO effect, components of the MAPK signaling pathway and NF-κB and its upstream effectors were examined, because this pathway and transcription factor play an essential role in the modulation of LPS-induced inflammation and transcriptional regulation. As shown in Fig. 5, the phosphorylation of IκBα and the nuclear translocation of p65 were suppressed by TO pretreatment. However, TO had no apparent effect on the phosphorylation of Erk1/2, p38MAPK, and JNK (Additional file 1: Figure S3). These results suggest that inhibition of the LPS-induced transactivation of p65 consequently reduces the expression of inflammatory mediators in TO-pretreated HUVECs.
Conclusion
Our findings demonstrated that the methanol extract of TO could attenuate LPS-induced endothelial activation by inhibiting the NF-κB pathway. Although TO has been used in the treatment of various disease, recent studies suggest it can be used effectively in the treatment of endothelial inflammation and atherosclerosis. However, to confirm these claims, more systematic, well-designed animal and randomized clinical studies with sufficient sample sizes are essential to investigate the exact action mechanisms, safety, and pharmacokinetics of this plant.
Acknowledgements
The authors would like to thank Dr. Heon Joo Park for the valuable comments and suggestions to improve the quality of the paper.
Funding
This work was supported by grants to H.S.K.; the National Research Foundation of Korea (NRF) grant (NRF-2014R1A5A2009392 and NRF-2015R1D1A1A01058340) and Inha University Research Grant (INHA-51361).
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article.
Abbreviations
- DMSO
Dimethyl sulfoxide
- HPLC
High-performance liquid chromatography
- HUVEC
Human umbilical vein endothelial cell
- IκBα
Inhibitor of NF-κB alpha
- ICAM-1
Intercellular adhesion molecule-1
- IL-1
Interleukin-1
- LPS
Lipopolysaccharide
- MAPK
Mitogen-activated protein kinase
- MCP-1
Monocyte chemoattractant protein-1
- NF-κB
Nuclear factor-kappa B
- TNF-α
Tumor necrosis factor-alpha
- TO
Taraxacum officinale
- VCAM-1
Vascular cell adhesion molecule-1
Additional file
Comparison of TO efficacy with acetylsalicylic acid (ASA). HUVECs were pretreated with TO (100 μg/ml) or different concentrations of ASA for one hour, and then stimulated with 1 μg/ml LPS for 24 h. VCAM-1 and actin levels were assessed by Western blot analysis. Arrow head indicates VCAM-1. Figure S2. TO decreases VCAM-1 induction dose dependently but does not affect ICAM-1 levels in LPS-stimulated HUVECs. HUVECs were pretreated with different concentrations of TO for one hour, and then stimulated with 1 μg/ml LPS for 24 h. VCAM-1, ICAM-1, and actin levels were assessed by Western blot analysis. Arrow head indicates VCAM-1. Figure S3. TO does not affect MAPK activation induced by LPS stimulation in HUVECs. HUVECs were preincubated with TO (100 μg/ml) for one hour and then stimulated with LPS (1 μg/ml) for 30 min. MAPKs activation was assessed by Western blot analysis. (PDF 368 kb)
Authors’ contributions
H.S.K designed the study. D.J., S.J.K, and H.S.K performed the experiments. D.J., S.J.K., and H.S.K. analyzed the data. H.S.K. wrote the paper. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing financial interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s12906-017-2022-7) contains supplementary material, which is available to authorized users.
Contributor Information
Daun Jeon, Email: beautwin@naver.com.
Seok Joong Kim, Email: skim@dongduk.ac.kr.
Hong Seok Kim, Phone: 82-32-860-9834, Email: kimhs0622@inha.ac.kr.
References
- 1.Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med. 1999;340(2):115–126. doi: 10.1056/NEJM199901143400207. [DOI] [PubMed] [Google Scholar]
- 2.Libby P. Inflammation in atherosclerosis. Arterioscler Thromb Vasc Biol. 2012;32(9):2045–2051. doi: 10.1161/ATVBAHA.108.179705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cohen Tervaert JW. Cardiovascular disease due to accelerated atherosclerosis in systemic vasculitides. Best Pract Res Clin Rheumatol. 2013;27(1):33–44. doi: 10.1016/j.berh.2012.12.004. [DOI] [PubMed] [Google Scholar]
- 4.Kiechl S, Egger G, Mayr M, Wiedermann CJ, Bonora E, Oberhollenzer F, Muggeo M, Xu Q, Wick G, Poewe W, et al. Chronic infections and the risk of carotid atherosclerosis: prospective results from a large population study. Circulation. 2001;103(8):1064–1070. doi: 10.1161/01.CIR.103.8.1064. [DOI] [PubMed] [Google Scholar]
- 5.Wiedermann CJ, Kiechl S, Dunzendorfer S, Schratzberger P, Egger G, Oberhollenzer F, Willeit J. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: prospective results from the Bruneck study. J Am Coll Cardiol. 1999;34(7):1975–1981. doi: 10.1016/S0735-1097(99)00448-9. [DOI] [PubMed] [Google Scholar]
- 6.Weber C, Noels H. Atherosclerosis: current pathogenesis and therapeutic options. Nat Med. 2011;17(11):1410–1422. doi: 10.1038/nm.2538. [DOI] [PubMed] [Google Scholar]
- 7.Frishman WH, Beravol P, Carosella C. Alternative and complementary medicine for preventing and treating cardiovascular disease. Dis Mon. 2009;55(3):121–192. doi: 10.1016/j.disamonth.2008.12.002. [DOI] [PubMed] [Google Scholar]
- 8.Orekhov AN, Sobenin IA, Korneev NV, Kirichenko TV, Myasoedova VA, Melnichenko AA, Balcells M, Edelman ER, Bobryshev YV. Anti-atherosclerotic therapy based on botanicals. Recent Pat Cardiovasc Drug Discov. 2013;8(1):56–66. doi: 10.2174/18722083113079990008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Guarrera PM, Savo V. Perceived health properties of wild and cultivated food plants in local and popular traditions of Italy: a review. J Ethnopharmacol. 2013;146(3):659–680. doi: 10.1016/j.jep.2013.01.036. [DOI] [PubMed] [Google Scholar]
- 10.Schutz K, Carle R, Schieber A. Taraxacum--a review on its phytochemical and pharmacological profile. J Ethnopharmacol. 2006;107(3):313–323. doi: 10.1016/j.jep.2006.07.021. [DOI] [PubMed] [Google Scholar]
- 11.Kim HM, Shin HY, Lim KH, Ryu ST, Shin TY, Chae HJ, Kim HR, Lyu YS, An NH, Lim KS. Taraxacum Officinale inhibits tumor necrosis factor-alpha production from rat astrocytes. Immunopharmacol Immunotoxicol. 2000;22(3):519–530. doi: 10.3109/08923970009026009. [DOI] [PubMed] [Google Scholar]
- 12.Hu C, Kitts DD. Luteolin and luteolin-7-O-glucoside from dandelion flower suppress iNOS and COX-2 in RAW264.7 cells. Mol Cell Biochem. 2004;265(1–2):107–113. doi: 10.1023/B:MCBI.0000044364.73144.fe. [DOI] [PubMed] [Google Scholar]
- 13.Liu L, Xiong H, Ping J, Ju Y, Zhang X. Taraxacum Officinale protects against lipopolysaccharide-induced acute lung injury in mice. J Ethnopharmacol. 2010;130(2):392–397. doi: 10.1016/j.jep.2010.05.029. [DOI] [PubMed] [Google Scholar]
- 14.Kim IG, Kang SC, Kim KC, Choung ES, Zee OP. Screening of estrogenic and antiestrogenic activities from medicinal plants. Environ Toxicol Pharmacol. 2008;25(1):75–82. doi: 10.1016/j.etap.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 15.Hossen MJ, Jeon SH, Kim SC, Kim JH, Jeong D, Sung NY, Yang S, Baek KS, Kim JH, Yoon DH, et al. Vitro and in vivo anti-inflammatory activity of Phyllanthus Acidus methanolic extract. J Ethnopharmacol. 2015;168:217–228. doi: 10.1016/j.jep.2015.03.043. [DOI] [PubMed] [Google Scholar]
- 16.Wang Y, Wang B, Guerram M, Sun L, Shi W, Tian C, Zhu X, Jiang Z, Zhang L. Deoxypodophyllotoxin suppresses tumor vasculature in HUVECs by promoting cytoskeleton remodeling through LKB1-AMPK dependent rho a activatio. Oncotarget. 2015;6(30):29497–29512. doi: 10.18632/oncotarget.4985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goodwin CJ, Holt SJ, Downes S, Marshall NJ. Microculture tetrazolium assays: a comparison between two new tetrazolium salts, XTT and MTS. J Immunol Methods. 1995;179(1):95–103. doi: 10.1016/0022-1759(94)00277-4. [DOI] [PubMed] [Google Scholar]
- 18.Cho YY, Tang F, Yao K, Lu C, Zhu F, Zheng D, Pugliese A, Bode AM, Dong Z. Cyclin-dependent kinase-3-mediated c-Jun phosphorylation at Ser63 and Ser73 enhances cell transformation. Cancer Res. 2009;69(1):272–281. doi: 10.1158/0008-5472.CAN-08-3125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.ET O, Kim JW, Kim JM, Kim SJ, Lee JS, Hong SS, Goodwin J, Ruthenborg RJ, Jung MG, Lee HJ, et al. NQO1 inhibits proteasome-mediated degradation of HIF-1alpha. Nat Commun. 2016;7:13593. doi: 10.1038/ncomms13593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wang Y, Cao J, Fan Y, Xie Y, Xu Z, Yin Z, Gao L, Wang C. Artemisinin inhibits monocyte adhesion to HUVECs through the NF-kappaB and MAPK pathways in vitro. Int J Mol Med. 2016;37(6):1567–1575. doi: 10.3892/ijmm.2016.2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pop R, Csernatoni F, Ranga F, Fetea F, Socaciu C: HPLC-UV Analysis Coupled with Chemometry to Identify Phenolic Biomarkers from Medicinal Plants, used as Ingredients in Two Food Supplement Formulas 2013, v. 70.
- 22.Elices MJ, Osborn L, Takada Y, Crouse C, Luhowskyj S, Hemler ME, Lobb RR. VCAM-1 on activated endothelium interacts with the leukocyte integrin VLA-4 at a site distinct from the VLA-4/fibronectin binding site. Cell. 1990;60(4):577–584. doi: 10.1016/0092-8674(90)90661-W. [DOI] [PubMed] [Google Scholar]
- 23.Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an overview. J Interf Cytokine Res. 2009;29(6):313–326. doi: 10.1089/jir.2008.0027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Olson TS, Ley K. Chemokines and chemokine receptors in leukocyte trafficking. Am J Physiol Regul Integr Comp Physiol. 2002;283(1):R7–28. doi: 10.1152/ajpregu.00738.2001. [DOI] [PubMed] [Google Scholar]
- 25.Yan W, Zhao K, Jiang Y, Huang Q, Wang J, Kan W, Wang S. Role of p38 MAPK in ICAM-1 expression of vascular endothelial cells induced by lipopolysaccharide. Shock. 2002;17(5):433–438. doi: 10.1097/00024382-200205000-00016. [DOI] [PubMed] [Google Scholar]
- 26.Anand AR, Bradley R, Ganju RK. LPS-induced MCP-1 expression in human microvascular endothelial cells is mediated by the tyrosine kinase, Pyk2 via the p38 MAPK/NF-kappaB-dependent pathway. Mol Immunol. 2009;46(5):962–968. doi: 10.1016/j.molimm.2008.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ghosh S, Hayden MS. New regulators of NF-kappaB in inflammation. Nat Rev Immunol. 2008;8(11):837–848. doi: 10.1038/nri2423. [DOI] [PubMed] [Google Scholar]
- 28.Coggins M, Rosenzweig A. The fire within: cardiac inflammatory signaling in health and disease. Circ Res. 2012;110(1):116–125. doi: 10.1161/CIRCRESAHA.111.243196. [DOI] [PubMed] [Google Scholar]
- 29.Aird WC. Phenotypic heterogeneity of the endothelium: I. Structure, function, and mechanisms. Circ Res. 2007;100(2):158–173. doi: 10.1161/01.RES.0000255691.76142.4a. [DOI] [PubMed] [Google Scholar]
- 30.Tousoulis D, Kampoli AM, Papageorgiou N, Androulakis E, Antoniades C, Toutouzas K, Stefanadis C. Pathophysiology of atherosclerosis: the role of inflammation. Curr Pharm Des. 2011;17(37):4089–4110. doi: 10.2174/138161211798764843. [DOI] [PubMed] [Google Scholar]
- 31.Chiu JJ, Chien S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. Physiol Rev. 2011;91(1):327–387. doi: 10.1152/physrev.00047.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pober JS, Cotran RS. Cytokines and endothelial cell biology. Physiol Rev. 1990;70(2):427–451. doi: 10.1152/physrev.1990.70.2.427. [DOI] [PubMed] [Google Scholar]
- 33.Stenvinkel P. Endothelial dysfunction and inflammation-is there a link? Nephrol Dial Transplant. 2001;16(10):1968–1971. doi: 10.1093/ndt/16.10.1968. [DOI] [PubMed] [Google Scholar]
- 34.Hadi HA, Carr CS, Al Suwaidi J. Endothelial dysfunction: cardiovascular risk factors, therapy, and outcome. Vasc Health Risk Manag. 2005;1(3):183–198. [PMC free article] [PubMed] [Google Scholar]
- 35.Libby P. Inflammation in atherosclerosis. Nature. 2002;420(6917):868–874. doi: 10.1038/nature01323. [DOI] [PubMed] [Google Scholar]
- 36.Rietschel ET, Kirikae T, Schade FU, Mamat U, Schmidt G, Loppnow H, Ulmer AJ, Zahringer U, Seydel U, Di Padova F, et al. Bacterial endotoxin: molecular relationships of structure to activity and function. FASEB J. 1994;8(2):217–225. doi: 10.1096/fasebj.8.2.8119492. [DOI] [PubMed] [Google Scholar]
- 37.Raetz CR, Whitfield C. Lipopolysaccharide endotoxins. Annu Rev Biochem. 2002;71:635–700. doi: 10.1146/annurev.biochem.71.110601.135414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Saito T, Hayashida H, Furugen R. Comment on: Cani et al. (2007) Metabolic endotoxemia initiates obesity and insulin resistance: Diabetes 56:1761–1772. Diabetes. 2007;56(12):e20. doi: 10.2337/db07-1181. [DOI] [PubMed] [Google Scholar]
- 39.Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008;57(6):1470–1481. doi: 10.2337/db07-1403. [DOI] [PubMed] [Google Scholar]
- 40.Jayashree B, Bibin YS, Prabhu D, Shanthirani CS, Gokulakrishnan K, Lakshmi BS, Mohan V, Balasubramanyam M. Increased circulatory levels of lipopolysaccharide (LPS) and zonulin signify novel biomarkers of proinflammation in patients with type 2 diabetes. Mol Cell Biochem. 2014;388(1–2):203–210. doi: 10.1007/s11010-013-1911-4. [DOI] [PubMed] [Google Scholar]
- 41.Gomes JM, Costa JA, Alfenas RC. Metabolic endotoxemia and diabetes mellitus: a systematic review. Metabolism. 2017;68:133–144. doi: 10.1016/j.metabol.2016.12.009. [DOI] [PubMed] [Google Scholar]
- 42.Ahmad VU, Yasmeen S, Ali Z, Khan MA, Choudhary MI, Akhtar F, Miana GA, Zahid M. Taraxacin, a new guaianolide from Taraxacum Wallichii. J Nat Prod. 2000;63(7):1010–1011. doi: 10.1021/np990495+. [DOI] [PubMed] [Google Scholar]
- 43.Sweeney B, Vora M, Ulbricht C, Basch E. Evidence-based systematic review of dandelion (Taraxacum Officinale) by natural standard research collaboration. J Herb Pharmacother. 2005;5(1):79–93. doi: 10.1080/J157v05n01_09. [DOI] [PubMed] [Google Scholar]
- 44.Jeon HJ, Kang HJ, Jung HJ, Kang YS, Lim CJ, Kim YM, Park EH. Anti-inflammatory activity of Taraxacum Officinale. J Ethnopharmacol. 2008;115(1):82–88. doi: 10.1016/j.jep.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 45.Koh YJ, Cha DS, Ko JS, Park HJ, Choi HD. Anti-inflammatory effect of Taraxacum Officinale leaves on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J Med Food. 2010;13(4):870–878. doi: 10.1089/jmf.2009.1249. [DOI] [PubMed] [Google Scholar]
- 46.Park CM, Jin KS, Lee YW, Song YS. Luteolin and chicoric acid synergistically inhibited inflammatory responses via inactivation of PI3K-Akt pathway and impairment of NF-kappaB translocation in LPS stimulated RAW 264.7 cells. Eur J Pharmacol. 2011;660(2–3):454–459. doi: 10.1016/j.ejphar.2011.04.007. [DOI] [PubMed] [Google Scholar]
- 47.Zhang X, Xiong H, Liu L. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J Ethnopharmacol. 2012;141(1):206–211. doi: 10.1016/j.jep.2012.02.020. [DOI] [PubMed] [Google Scholar]
- 48.Stumpf C, Fan Q, Hintermann C, Raaz D, Kurfurst I, Losert S, Pflederer W, Achenbach S, Daniel WG, Garlichs CD. Anti-inflammatory effects of danshen on human vascular endothelial cells in culture. Am J Chin Med. 2013;41(5):1065–1077. doi: 10.1142/S0192415X13500729. [DOI] [PubMed] [Google Scholar]
- 49.Jiang X, Lv B, Li P, Ma X, Wang T, Zhou Q, Wang X, Gao X. Bioactivity-integrated UPLC/Q-TOF-MS of Danhong injection to identify NF-kappaB inhibitors and anti-inflammatory targets based on endothelial cell culture and network pharmacology. J Ethnopharmacol. 2015;174:270–276. doi: 10.1016/j.jep.2015.08.026. [DOI] [PubMed] [Google Scholar]
- 50.Hebeda CB, Bolonheis SM, Nakasato A, Belinati K, Souza PD, Gouvea DR, Lopes NP, Farsky SH. Effects of chlorogenic acid on neutrophil locomotion functions in response to inflammatory stimulus. J Ethnopharmacol. 2011;135(2):261–269. doi: 10.1016/j.jep.2011.02.033. [DOI] [PubMed] [Google Scholar]
- 51.Kim SR, Jung YR, Kim DH, An HJ, Kim MK, Kim ND, Chung HY. Caffeic acid regulates LPS-induced NF-kappaB activation through NIK/IKK and c-Src/ERK signaling pathways in endothelial cells. Arch Pharm Res. 2014;37(4):539–547. doi: 10.1007/s12272-013-0211-6. [DOI] [PubMed] [Google Scholar]
- 52.Moon MK, Lee YJ, Kim JS, Kang DG, Lee HS. Effect of caffeic acid on tumor necrosis factor-alpha-induced vascular inflammation in human umbilical vein endothelial cells. Biol Pharm Bull. 2009;32(8):1371–1377. doi: 10.1248/bpb.32.1371. [DOI] [PubMed] [Google Scholar]
- 53.Zhao J, Suyama A, Tanaka M, Matsui T. Ferulic acid enhances the vasorelaxant effect of epigallocatechin gallate in tumor necrosis factor-alpha-induced inflammatory rat aorta. J Nutr Biochem. 2014;25(7):807–814. doi: 10.1016/j.jnutbio.2014.03.013. [DOI] [PubMed] [Google Scholar]
- 54.Ma ZC, Hong Q, Wang YG, Tan HL, Xiao CR, Liang QD, Cai SH, Gao Y. Ferulic acid attenuates adhesion molecule expression in gamma-radiated human umbilical vascular endothelial cells. Biol Pharm Bull. 2010;33(5):752–758. doi: 10.1248/bpb.33.752. [DOI] [PubMed] [Google Scholar]
- 55.Hu G, Wang J, Hong D, Zhang T, Duan H, Mu X, Yang Z. Effects of aqueous extracts of Taraxacum Officinale on expression of tumor necrosis factor-alpha and intracellular adhesion molecule 1 in LPS-stimulated RMMVECs. BMC Complement Altern Med. 2017;17(1):38. doi: 10.1186/s12906-016-1520-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Basri AM, Taha H, Ahmad N, Review A. On the pharmacological activities and phytochemicals of Alpinia Officinarum (galangal) extracts derived from bioassay-guided fractionation and isolation. Pharmacogn Rev. 2017;11(21):43–56. doi: 10.4103/phrev.phrev_55_16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.DP X, Li Y, Meng X, Zhou T, Zhou Y, Zheng J, Zhang JJ, Li HB. Natural antioxidants in foods and medicinal plants: extraction, assessment and resources. Int J Mol Sci. 2017;18(1):E96. doi: 10.3390/ijms18010096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Iademarco MF, McQuillan JJ, Rosen GD, Dean DC. Characterization of the promoter for vascular cell adhesion molecule-1 (VCAM-1) J Biol Chem. 1992;267(23):16323–16329. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article.