

Abstract
Nus factors are broadly conserved across bacterial species, and are often essential for viability. A complex of five Nus factors (NusB, NusE, NusA, NusG and SuhB) is considered to be a dedicated regulator of ribosomal RNA folding, and has been shown to prevent Rho-dependent transcription termination. Here, we identify an additional cellular function for the Nus factor complex in Escherichia coli: repression of the Nus factor-encoding gene, suhB. This repression occurs primarily by translation inhibition, followed by Rho-dependent transcription termination. Thus, the Nus factor complex can prevent or promote Rho activity depending on the gene context. Conservation of putative NusB/E binding sites upstream of Nus factor genes suggests that Nus factor autoregulation occurs in many bacterial species. Additionally, many putative NusB/E binding sites are also found upstream of other genes in diverse species, and we demonstrate Nus factor regulation of one such gene in Citrobacter koseri. We conclude that Nus factors have an evolutionarily widespread regulatory function beyond ribosomal RNA, and that they are often autoregulatory.
The Nus factor complex regulates rRNA folding and prevents Rho-dependent transcription termination in bacteria. Here, Baniulyte et al. show that it also inhibits translation of one of the Nus factor-encoding genes, suhB, and probably regulates the expression of other genes in diverse bacterial species.
Introduction
Nus factors are widely conserved in bacteria and play a variety of important roles in transcription and translation1. The Nus factor complex comprises the four classical Nus factors, NusA, NusB, NusE (ribosomal protein S10), NusG, and a recently discovered member, SuhB. As a complex, Nus factors serve an important role in promoting expression of ribosomal RNA (rRNA)2,3. A NusB/E complex binds BoxA sequence elements in nascent rRNA, upstream of the 16S and 23S genes4,5. Once bound to BoxA, NusB/E has been proposed to interact with elongating RNAP via the NusE−NusG interaction6. The role of NusA in Nus complex function is unclear, but may involve binding of NusA to RNA flanking the BoxA7. NusA has also been proposed to be a general Rho antagonist by competing with Rho for RNA sites8. Early studies of Nus factors focused on their role in preventing both Rho-dependent and intrinsic termination of λ bacteriophage RNAs (‘antitermination’)9, which is completely dependent on the bacteriophage protein N. Nus factors can prevent Rho-dependent termination in the absence of N10,11, and for many years, Nus factors were believed to prevent Rho-dependent termination of rRNA9. However, it was recently shown that rRNA is intrinsically resistant to Rho termination, and that the primary role of Nus factors at rRNA is to promote proper RNA folding during ribosome assembly3,12.
The most recently discovered Nus factor, SuhB, has been proposed to stabilize interactions between the NusB/E-bound BoxA and elongating RNAP, thus contributing to proper folding of rRNA12. Genome-wide approaches revealed that suhB is upregulated in the presence of the Rho inhibitor bicyclomycin, suggesting that suhB is subject to premature Rho-dependent transcription termination13,14. Surprisingly, suhB is also one of the most upregulated genes in ΔnusB cells12, suggesting a possible autoregulatory function for Nus factors. Moreover, autoregulation of suhB has been suggested previously15, although the mechanism for this regulation is unclear.
Here, we show that suhB is translationally repressed by Nus factors, which in turn leads to premature Rho-dependent transcription termination. This represents a novel mechanism for control of premature Rho-dependent termination, and is the first described cellular function for Nus factors beyond regulation of rRNA. Moreover, the role of Nus factors at suhB is to promote Rho-dependent termination of suhB, in contrast to their established function in antagonizing Rho. Bioinformatic analysis suggests that regulation by Nus factors is widespread, and that autoregulation of genes encoding Nus factors, in particular suhB, is a common phenomenon. We confirm Nus factor association with suhB mRNA in Salmonella enterica, and we demonstrate Nus factor regulation of an unrelated gene in Citrobacter koseri. Thus, our data show that Nus factors are important regulators with diverse targets and diverse regulatory mechanisms.
Results
Rho-dependent termination within the suhB gene
Genome-wide analysis of Rho termination events suggested Rho-dependent termination within the Escherichia coli suhB gene13,14. To confirm this, we used Chromatin Immunoprecipitation (ChIP) coupled with quantitative PCR (ChIP-qPCR) to determine RNAP association across the suhB gene in wild-type cells and cells expressing a mutant Rho (R66S) that has impaired termination activity, likely due to a defect in RNA loading16. In wild-type cells, we observed a large decrease in RNAP association at the 3′ end of suhB relative to the 5′ end. This decrease was substantially reduced in rho mutant cells (Fig. 1). Thus, our ChIP data independently support the observation of Rho termination within suhB 13,14.
Fig. 1.
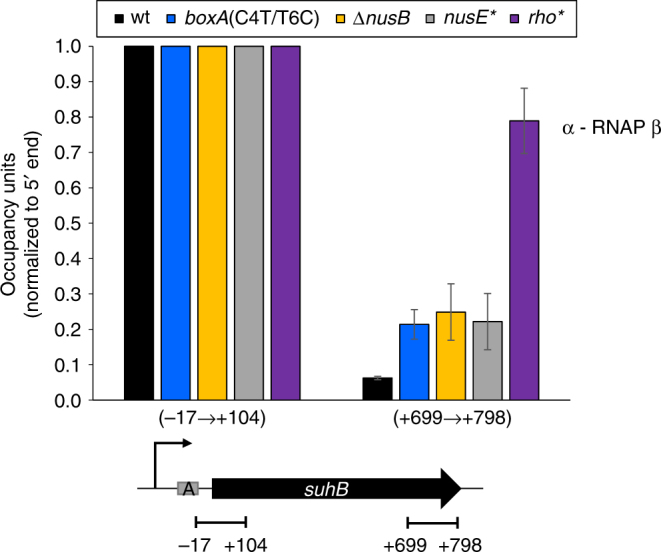
Transcription termination within suhB is dependent on Rho and Nus factors. RNAP (β) enrichment at suhB 5′ and 3′ regions was measured using ChIP-qPCR in wild-type MG1655, boxA(C4T/T6C), ΔnusB, nusE(A12E) or rho(R66S) mutant strains. Values are normalized to signal at the 5′ end of suhB. x-axis labels indicate qPCR amplicon position relative to suhB. Error bars represent ±1 standard deviation from the mean (n = 3). A schematic depicting the suhB gene, the transcription start site (bent arrow) and boxA (grey rectangle) is shown below the graph. Horizontal black lines indicate the position of PCR amplicons
Nus factors are trans-acting regulators of suhB
Based on an approach used to identify modulators of Rho-dependent termination within S. enterica chiP 17, we used a genetic selection to isolate 30 independent mutants defective in Rho-dependent termination within suhB (see Methods). All 30 strains isolated had a mutation in one of three genes: nusB (14 mutants), nusE (13 mutants) or nusG (3 mutants) (Supplementary Table 1). We then measured RNAP association across the suhB gene in wild-type, ΔnusB and nusE mutant cells (nusE A12E mutant isolated from the genetic selection). Mutation of nusB or nusE increased RNAP binding at the suhB 3′ end ~4-fold compared to wild-type cells (Fig. 1 and Supplementary Fig. 1). We conclude that Nus factors promote Rho-dependent termination within the suhB gene. However, RNAP occupancy at the 3′ end of suhB in nusB and nusE mutants was substantially lower than in the rho mutant (Fig. 1 and Supplementary Fig. 1). This difference may be due to spurious, non-coding transcripts arising from nearby intragenic promoters, which are widespread in E. coli 18 and are often terminated by Rho13,14.
A functional BoxA in the suhB 5′ UTR
We identified a sequence in the suhB 5′ UTR with striking similarity to boxA sequences from rRNA loci (Supplementary Table 2). Moreover, this boxA-like sequence is broadly conserved across Enterobacteriaceae species (Fig. 2a and Supplementary Fig. 2), suggesting that it is a genuine binding site for NusB/E. We generated a library of randomly mutated suhB-lacZ transcriptional fusions (see Methods), and identified fusions that had higher expression of lacZ. All identified mutants carried a single nucleotide change at one of five different positions within the putative boxA (Fig. 2b). We then constructed a strain carrying two chromosomal point mutations in the putative suhB boxA (C4T/T6C; numbers corresponding to the position in the consensus boxA; Supplementary Table 2). We used ChIP-qPCR to measure association of FLAG-tagged SuhB at the 5′ end of the suhB gene in wild-type cells, or cells containing the boxA mutation. We detected robust association of SuhB-FLAG in wild-type cells, but not in the boxA mutant strain (Fig. 2c). We conclude that the putative BoxA in the 5′ UTR of suhB is genuine, and recruits Nus factors. To test whether the BoxA controls Rho-dependent termination within suhB, we measured RNAP occupancy across suhB in the boxA mutant strain. We detected a ~4-fold increase in RNAP occupancy in the downstream portion of suhB in the boxA mutant strain relative to wild-type cells, mirroring the effect of mutating nusB or nusE (Fig. 1). Our data support a model in which Nus factor recruitment by the suhB BoxA leads to Rho-dependent termination within the gene.
Fig. 2.
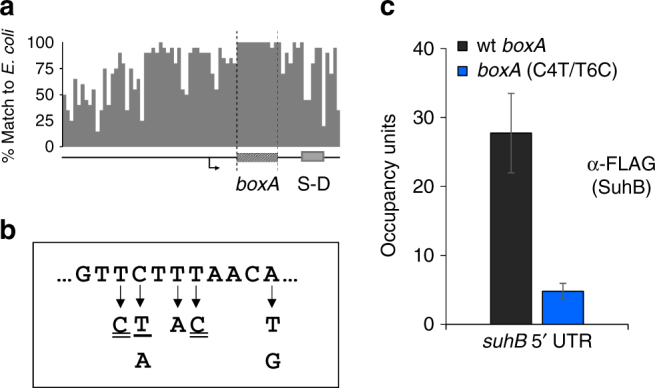
A functional BoxA in the 5′ UTR of suhB. a Sequence conservation of the 100 bp upstream of suhB and its homologues across 20 Enterobacteriaceae species. The transcription start site is indicated by a bent arrow, and the BoxA and S-D sequences are indicated. b List of boxA mutations that are associated with increased suhB expression. All single nucleotide changes are indicated by an arrow. Single underline indicates a mutation that was isolated in the absence of mutations anywhere else in the cloned region; other mutants included additional mutations outside the boxA. Double underline indicates that the boxA mutation was isolated in two or more independent clones. Critical position ‘−4’ is indicated (see Supplementary Table 2). c SuhB association with the 5′ end of suhB in wild-type (‘wt’) and boxA mutant (‘boxA C4T/T6C’) strains. SuhB-FLAG occupancy was measured by ChIP-qPCR using α-FLAG antibody. Error bars represent ±1 standard deviation from the mean (n = 3)
Nus factors mediate translational repression of suhB
The suhB BoxA is separated by only 6 nt from the Shine-Dalgarno (S-D) sequence (Fig. 2a). Rho cannot terminate transcription of translated RNA, likely because RNAP-bound NusG interacts with ribosome-associated NusE (S10)6. Hence, we hypothesized that NusB/E association with BoxA sterically blocks association of the 30S ribosome with the mRNA, repressing translation initiation, uncoupling transcription and translation, and thereby promoting Rho-dependent termination. To test this hypothesis, we used the suhB-lacZ transcriptional fusion (Fig. 3a), as well as an equivalent translational fusion (Fig. 3b). We reasoned that mutation of nusB, nusE or boxA would result in increased expression from both reporter fusions, since these mutations would relieve translational repression (reported by the translational fusion), which in turn would reduce Rho-dependent termination (reported by the transcriptional fusion). In contrast, we reasoned that mutation of rho would result in increased expression only from the transcriptional fusion reporter, since the SuhB-LacZ fusion protein (from the translational fusion construct) would still be translationally repressed. We measured expression of lacZ from each of these reporter fusions in wild-type cells, and cells with ΔnusB, nusE A12E or rho R66S mutations. We also measured expression of lacZ in these strains using reporter fusions carrying the C4T/T6C boxA mutation. Consistent with our model, we detected increased expression of both reporter fusion types in mutants of nusB, nusE or boxA, whereas mutation of rho resulted in increased expression of the transcriptional fusion but not the translational fusion reporter (Fig. 3a, b). Note that mutation of nusB, nusE or boxA does not lead to the same level of increase in expression of the reporter fusions (Fig. 3a, b). This is likely due to the fact that mutations in Nus factors have extensive pleiotropic effects, presumably due to the importance of Nus factors in ribosome assembly3. Moreover, mutation of boxA in a nusE mutant leads to a further increase in reporter expression, whereas mutation of boxA in a nusB mutant does not (Fig. 3a, b). This is likely due to the mutant NusE retaining partial function, whereas deletion of nusB completely abolishes Nus factor function.
Fig. 3.

Nus factors repress translation of suhB, leading to Rho-dependent termination within the gene. β-galactosidase activity of (a) transcriptional and (b) translational fusions of suhB to lacZ in wild-type cells, ΔnusB, nusE(A12E), or rho(R66S) mutants. The suhB-lacZ fusion had either a wild-type (‘wt’) or mutant boxA (‘C4T/T6C’). Data are normalized to levels in wild-type cells. Error bars represent ±1 standard deviation from the mean (n = 3). Schematics of constructs used in these experiments are depicted above the graphs. (c) and (d) Western blots showing SuhB-FLAG protein levels in wild-type cells, nusE(A12E), rho(R66S) c, and boxA(C4T/T6C) mutants d. SuhB-FLAG was probed with α-FLAG antibody; RNAP β′ was probed as a loading control. Representative blots from at least three independent experiments are shown. Images were cropped for clarity; unprocessed western blot images are in Supplementary Fig. 7
To confirm the effects of mutating nusB, nusE, rho and boxA on expression of suhB in the native context, we measured SuhB protein levels by western blotting using strains expressing a C-terminally FLAG-tagged derivative of SuhB. We compared SuhB protein levels in cells with nusE A12E, rho R66S or boxA C4T/T6C mutations; we have previously shown that SuhB protein levels are increased in a ΔnusB mutant12. SuhB protein levels in the mutant strains correlated well with the translational suhB-lacZ fusion reporter gene assay: mutation of nusE or boxA caused a modest increase in SuhB-FLAG levels, whereas mutation of rho had no discernible effect (Fig. 3c, d; Supplementary Fig. 7).
Rho-dependent transcription termination occurs early in the suhB gene
Rho-dependent termination requires a Rho loading sequence known as a Rut that typically occurs >60 nt upstream of the termination site(s), is pyrimidine-rich and G-poor19. To localize the Rut and the downstream termination site(s), we constructed a short transcriptional suhB-lacZ fusion that includes only the first 57 bp of the suhB gene. Expression of this reporter fusion was substantially higher in rho mutant cells than in wild-type cells (Fig. 4a). In contrast, expression was only marginally higher in rho mutant cells than in wild-type cells when the boxA sequence was mutated (Fig. 4a). Thus, the short suhB-lacZ reporter fusion behaves similarly to the fusion that includes the entire suhB gene (Fig. 3a), indicating that the rut and termination sequences must be upstream of position 57 within suhB.
Fig. 4.

The suhB BoxA overlaps the first of multiple Rut elements. β–galactosidase activity of short (a) and full-length (b) suhB transcriptional fusions to lacZ. Constructs included either a wild-type sequence (‘wt’), boxA mutation (‘C4T, T6C’) and/or suhB start codon mutation (‘ATG → CAG’), as indicated on the x-axis. 200 bp of sequence upstream of the suhB gene was included in all constructs. Error bars represent ±1 standard deviation from the mean (n = 3). Schematics of the constructs used for these experiments are depicted below the graphs
Given that the short suhB-lacZ fusion includes only 94 bp of transcribed sequence from suhB and its 5′ UTR, and that Rut sequences are typically found >60 nt from the site(s) of termination19, we reasoned that the Rut is likely located close to the 5′ end of the suhB 5′ UTR. Consistent with this, positions 2–22 of the 5′ UTR include 17 pyrimidines and only one G. This sequence completely encompasses the boxA, suggesting that the boxA and rut sequences overlap. To determine whether mutation of the boxA affects Rho-dependent termination independent of Nus factor-mediated translational repression, we constructed short suhB-lacZ fusions in which the suhB start codon was mutated, either alone or in conjunction with a mutated boxA. We reasoned that mutation of the suhB start codon would bypass the need for BoxA-mediated translational repression to cause Rho-dependent termination. As expected, expression of the fusion with the mutated start codon but wild-type boxA was substantially higher in a rho mutant than in wild-type cells (Fig. 4a), consistent with this construct being Rho-terminated. However, expression of the fusion with the mutated start codon and mutated boxA was only marginally higher in a rho mutant than in wild-type cells, indicating that Rho-dependent termination is disrupted by mutation of the boxA, even in the absence of suhB translation. We conclude that mutation of the boxA reduces Rho-dependent termination by disrupting the rut. This likely occurs due to the boxA and rut sequences overlapping, in which case mutating the boxA would also alter the rut. However, mutation of the boxA might also alter RNA secondary structure of the Rut.
Mutation of the boxA results in greatly decreased Rho-dependent termination of a fusion of the entire suhB gene to lacZ (Fig. 3a). Although this effect could be due to disruption of a rut overlapping the boxA, we reasoned that there are likely to be additional rut sequences within the suhB ORF. To test this hypothesis, we constructed transcriptional fusions of the entire suhB gene and 5′ UTR to lacZ with a mutation in the suhB start codon, either alone or in conjunction with a mutation in the boxA. For both constructs, expression was substantially higher in a rho mutant than in wild-type cells (Fig. 4b), indicating robust Rho-dependent termination within the suhB gene, even with a mutated boxA. We conclude that the suhB gene includes at least one additional rut, and that the effect of mutating the boxA on Rho-dependent termination with a long transcriptional fusion (Fig. 3) is due to loss of Nus factor binding rather than a direct effect on Rho loading.
BoxA-mediated occlusion of the S-D sequence is not due to steric occlusion
The data described above are consistent with a steric occlusion model in which NusB/E binding to the BoxA directly prevent 30S ribosome association with the S-D sequence. However, other mechanisms of translational repression are also possible. The steric occlusion model predicts that increasing the distance between the boxA and S-D elements would relieve translational repression, and consequently Rho-dependent termination. We constructed suhB-lacZ transcriptional fusions that carried insertions of sizes from 2 to 100 bp between the boxA and S-D sequences (see Methods for details). We constructed equivalent fusions carrying a boxA mutation (C4A; Supplementary Table 2). Surprisingly, separating the BoxA and S-D sequences with up to 100 nt intervening RNA did not abolish BoxA-mediated repression (Fig. 5). Note that differences in absolute expression levels for the different constructs are likely due to variability in secondary structure around the ribosome binding site. Additionally, we are confident that none of the insertions inadvertently introduces a new promoter, since a similar construct lacking an active upstream promoter was only weakly expressed (Supplementary Fig. 3). We conclude that the steric occlusion model is insufficient to explain BoxA-mediated translational repression of suhB, although the proximity of the BoxA and S-D sequences suggests that simple occlusion would prevent ribosome binding.
Fig. 5.
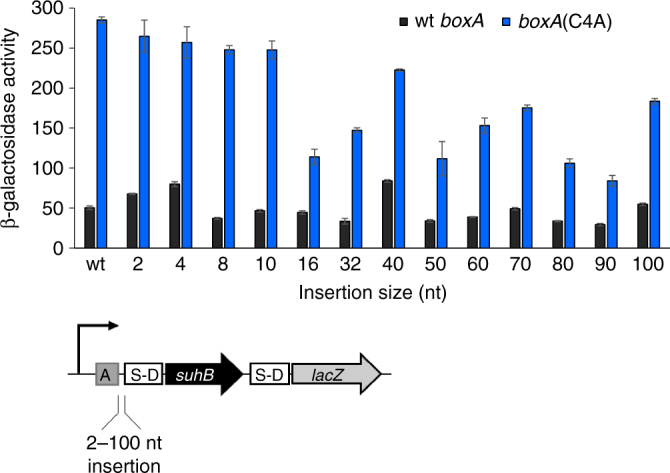
The effect on suhB-lacZ transcription levels of altering the distance between boxA and the S-D sequence. β–galactosidase activity of wild-type (‘wt boxA’; dark grey bars) and boxA mutant (‘C4A’; blue bars) transcriptional fusions of suhB to lacZ, with increasing lengths of non-coding DNA inserted between the boxA and S-D sequences. The length of inserted sequence (nt) is indicated on the x-axis. Constructs include 200 bp of upstream sequence and a full-length suhB fused to lacZ in the pAMD-BA-lacZ plasmid. Note that the sequence of inserted non-coding DNA differs for constructs with insertion sizes of ≤32 bp and ≥40 bp (see Methods and Supplementary Fig. 6 for details)
We reasoned that if steric occlusion of ribosomes by NusB/E binding is sufficient for repression of suhB, it would not require assembly of a complete Nus factor complex, since NusB/E alone has a high affinity for BoxA RNA4. Hence, we constructed suhB-lacZ translational fusions where the native promoter is replaced by a T7 promoter. Previous studies showed that gene regulation involving λ N or NusG is lost when E. coli RNAP is substituted with bacteriophage T7 RNAP20–22, suggesting that T7 RNAP does not interact with Nus factors; hence, transcription of this suhB-lacZ fusion by T7 RNAP would not be associated with the formation of a complete Nus factor complex. We grew cells at 37 °C, 30 °C or room temperature (23 °C), since the transcription elongation rate of T7 RNAP is similar to that of E. coli RNAP at room temperature, but considerably higher at 37 °C23,24. At all temperatures, we detected robust expression that was dependent upon expression of T7 RNAP in the same cells. However, we observed no effect on expression of mutating the boxA (Supplementary Fig. 4). We conclude that efficient BoxA-dependent repression of suhB requires assembly of a complete Nus factor complex.
Salmonella enterica suhB has a functional BoxA
Phylogenetic analysis of the region upstream of the suhB gene indicates that the boxA sequence is widely conserved among members of the family Enterobacteriaciae (Fig. 2a; Supplementary Table 2; Supplementary Fig. 2), suggesting that BoxA-mediated regulation of suhB occurs in these species. To investigate this possibility, we used ChIP of FLAG-tagged SuhB to measure association of SuhB with the suhB upstream region in S. enterica subspecies enterica serovar Typhimurium. We detected robust association of both RNAP (β subunit) and SuhB with the suhB upstream region (Supplementary Fig. 5), indicating that the suhB mRNA contains a functional BoxA. We also failed to detect association with a previously reported cryptic BoxA within the hisG gene (Supplementary Fig. 5), consistent with the sequence of this element differing at a critical position from the BoxA consensus (Supplementary Table 2).
BoxA-mediated regulation and Nus factor autoregulation are phylogenetically widespread
Aside from their role in lambdoid phage, Nus factors have historically been considered dedicated regulators of rRNA expression. Our discovery of suhB as a novel regulatory target of Nus factors suggests that BoxA-mediated regulation may be more extensive. BoxA sequences in rRNA are known to be highly conserved2. Based on the boxA sequences from E. coli rRNA and suhB loci, and a previous analysis of sequences required for BoxA function in E. coli 5, we derived a consensus sequence (GYTCTTTAANA) that is likely to be applicable to almost all γ-proteobacteria2. We searched for perfect matches to this sequence in 940 sequenced γ-proteobacterial genomes. We then selected sequence matches that are positioned within 50 bp of a downstream start codon for an annotated gene. Thus, we identified 407 putative BoxA sequences from 314 genomes, with between 0 and 7 instances per genome (Supplementary Data 1). We determined whether any gene functions were identified from multiple genomes. To minimize biases from the uneven distribution of genome sequences across different genera, we analysed gene functions at the genus rather than species level. Across all the species analysed, we identified 36 different gene functions with at least one representative from one genus. Strikingly, we identified 34 of 55 genera in which at least one species has a putative boxA sequence within 50 bp of the start of an annotated suhB homologue. We identified three additional genera in which at least one species has a putative boxA within 50 bp of the start of an unannotated suhB homologue, and one genus with a species in which the suhB homologue has a putative boxA 82 bp from the gene start. Thus, our analysis reinforces the notion that BoxA-mediated regulation of suhB is highly conserved (Fig. 2a and Supplementary Fig. 2). Three other gene functions were represented in multiple genera: prsA (encodes ribose-phosphate pyrophosphokinase) and rpsJ (encodes NusE) were each found in three genera, and genes encoding ParE-like toxins were found in two genera. We also identified two genera with species in which rpsJ is predicted to be a downstream gene in an operon where the first gene in the operon has a putative boxA < 50 bp from the gene start.
BoxA-mediated regulation of a toxin-antitoxin system in Citrobacter koseri
Bioinformatic analysis strongly suggested that BoxA-mediated regulation is evolutionarily widespread and extends to genes other than suhB. To determine whether Nus factors regulate genes other than rRNA and suhB in other species, we selected one putative BoxA-regulated gene identified by the bioinformatic search for boxA-like sequences: CKO_00699 from C. koseri (Supplementary Table 2). CKO_00699 is predicted to encode a ParE-like toxin, part of a putative toxin−antitoxin pair. A putative boxA was observed upstream of a homologous gene in Pasteurella multocida, suggesting conserved BoxA-mediated regulation. We reasoned that if CKO_00699 is a genuine target of Nus factors, it would likely retain this regulation in E. coli, since Nus factors are highly conserved between C. koseri and E. coli (e.g. the amino acid sequence of NusB is 97% identical and 100% similar between the two species). Hence, we constructed a transcriptional fusion of CKO_00699 to lacZ and measured expression in E. coli. Note that we included a mutation in CKO_00699 (R82A) to inactivate the predicted toxin activity to prevent growth inhibition. The lacZ fusion included a strong, constitutive promoter25, and the sequence from C. koseri began at the predicted transcription start site, based on manual analysis of likely promoter sequences (Fig. 6). We measured expression of fusions with wild-type and mutant boxA (C4A) sequences (Supplementary Table 2), in wild-type and ΔnusB strains. Mutation of the putative boxA, or deletion of nusB resulted in a substantial increase in expression, whereas mutation of the boxA did not affect expression in the ΔnusB strain (Fig. 6). We conclude that CKO_00699 is directly repressed by a BoxA and Nus factors.
Fig. 6.
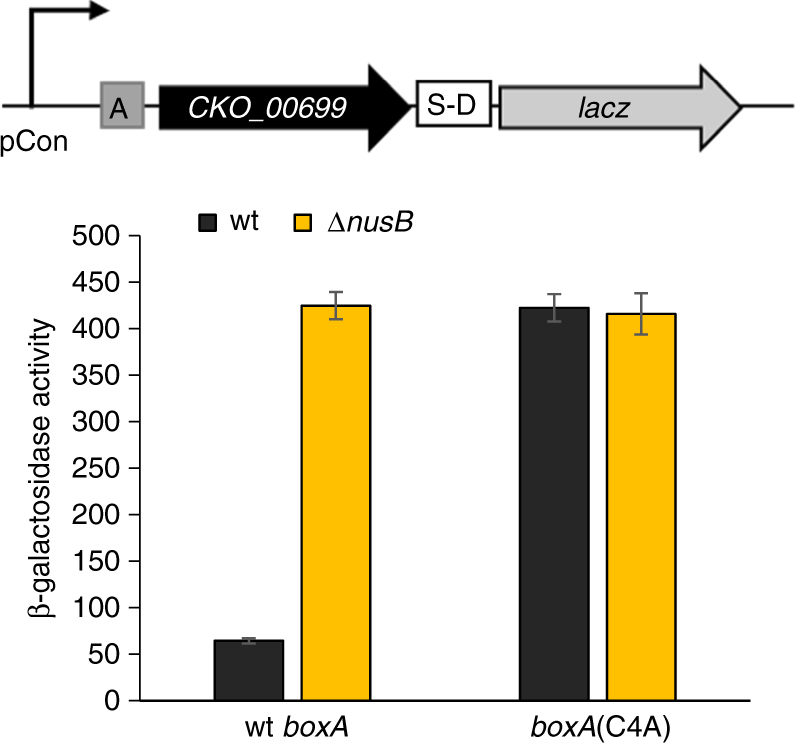
Identification of a BoxA element in Citrobacter koseri. β-galactosidase activity of wild-type (‘wt boxA’) and boxA mutant (‘C4A’) transcriptional fusions of CKO_00699 (R82A mutant, to avoid potential toxicity to E. coli in the absence of the anti-toxin) to lacZ in E. coli wild-type (‘wt’; dark grey bars) or nusB deletion (‘ΔnusB”; yellow bars) strains. CKO_00699-lacZ transcription was driven by a constitutive promoter25. Error bars represent ±1 standard deviation from the mean (n = 3)
Discussion
We have shown that premature Rho-dependent termination within the suhB gene is controlled by a BoxA and Nus factors. This likely serves as a mechanism for autoregulation of Nus factors, since SuhB is a critical component of the Nus machinery12. Premature Rho-dependent termination of mRNAs has been recently recognized to be a widespread regulatory mechanism26,27. Most regulation of this type occurs by alteration of mRNA accessibility around Rut sites. In the case of suhB, Rho-dependent termination occurs as a result of translational repression.
A function for Nus factors in promoting Rho-dependent termination is particularly striking because of their long association with antitermination9. The contrasting effects of Nus factors on Rho-dependent termination in different contexts, and their role in promoting ribosomal assembly, highlight the flexibility in the function of these proteins. Our data indicate that translational repression of suhB by Nus factors is not due to occlusion of the S-D. Previous studies of Nus factors suggest that they form a loop between the BoxA in the RNA and the elongating RNAP3,12. We propose that this loop prevents the 30S ribosome from accessing the S-D. Alternatively, association of NusG with NusE in the context of the Nus factor complex may prevent translation by blocking association of NusG with ribosome-associated NusE (S10).
Autoregulation of SuhB is strikingly similar to autoregulation of λ N. λ nutL is positioned ~200 bp upstream of the N gene. Binding of Nus factors and N to NutL results in translational repression of N28. The distance between NutL and the S-D sequence is such that a simple steric occlusion model is insufficient to explain translational repression by N and Nus factors; the RNA loop formed between NutL and the elongating RNAP provides a straightforward explanation of repression. Although the gap between NutL and the S-D sequence for the N gene is considerably longer than the longest distance we tested for suhB (Fig. 5), the intervening sequence is highly structured29, which may impact the compactness of the loop.
Although we have shown previously that Nus factors are not required to prevent Rho-dependent termination at rRNA loci12, Nus factors have been shown to prevent Rho-dependent termination in artificial reporter constructs10,11,30,31. Our finding that Nus factors promote Rho-dependent termination in suhB further indicates that context determines the precise function of Nus factors. Hence, it is likely that there are additional sequence elements in suhB that promote Rho-dependent termination, or that there are additional sequence elements in the artificial reporter constructs that prevent Rho-dependent termination.
Our data support a widespread regulatory role for Nus factors, implicating them in regulation in both a wide range of species, and of a diverse set of genes, although within any given species there are likely only a few regulatory targets. Strikingly, ~25% of the gene functions associated with an upstream boxA are known to be directly connected to translation. This is consistent with the established connection between Nus factors and ribosomal assembly3, and suggests that the impact of Nus factors on translation occurs by regulation of a variety of genes. Moreover, our data suggest that NusE is autoregulated in phylogenetically diverse species. Although we did not identify any genomes where genes encoding other Nus factors have putative upstream boxA sequences, we did identify a putative boxA sequence upstream of ribH in six different species of Pseudomonas. In all cases, nusB is the gene immediately downstream of ribH, suggesting that nusB is autoregulated in pseudomonads. Overall, we identified no species with a putative boxA upstream of more than one Nus factor-encoding gene, and only 11 genera had no putative boxA associated with any Nus factor-encoding gene. However, for five of these latter genera we were unable to identify a boxA sequence upstream of the rRNA genes, suggesting that the BoxA consensus is different to that in E. coli. Thus, our data strongly suggest that Nus factor autoregulation occurs in ~90% of gamma-proteobacterial species, and that typically, just one Nus factor is autoregulated. The evidence for autoregulation of SuhB, NusE and NusB suggests that the levels of these proteins contribute to feedback loops that control the primary function of Nus factors: promoting ribosomal assembly. Our observation of BoxA-mediated regulation of a ParE-like toxin in C. koseri demonstrates that Nus factors regulate genes other than their own. Indeed, our bioinformatic analysis suggests that genes of many functions may be regulated by Nus factors, with 36 gene functions represented in at least one genus. Our list is conservative because (i) it does not consider the possibility of regulation by BoxA sequences located >50 nt upstream of the gene start, which we know is possible (Fig. 5), (ii) it does not consider non-coding RNAs, (iii) the BoxA consensus may be different in some of the species analysed, and (iv) gene starts predicted by bioinformatic annotation pipelines may be incorrect32.
Our data indicate that regulation by Nus factors extends to many genes beyond rRNA, and that Nus factor autoregulation is an evolutionarily widespread phenomenon. Moreover, we have shown that Nus factors can provide contrasting forms of regulation, depending on the context of the target; despite their long-established function in antitermination9, Nus factors promote Rho-dependent termination within suhB. Key questions about the function of Nus factors remain to be addressed. What is the molecular architecture of the Nus factor machinery? What are the specific RNA sequences that determine whether Nus factors prevent Rho-dependent termination? How do Nus factors modulate the function of elongating RNAP? Our identification of novel Nus factor target genes with novel regulatory mechanisms provides an excellent opportunity to address these questions.
Methods
Strains and plasmids
All strains, plasmids and oligonucleotides used in this study are listed in Supplementary Tables 3 and 4. Mutations in rpsJ and rho were P1 transduced into MG165533 MG1655 ΔlacZ (AMD054)34 and MG1655suhB-FLAG3 (VS066)12. E. coli MG1655suhB(boxA(C4T/T6C)), MG1655suhB(boxA(C4T/T6C))-FLAG3 and S. Typhimurium hisGΔ+3::thyA, hisGΔ+100::thyA suhB-FLAG3 strains were constructed using the ‘Flexible Recombineering Using Integration of thyA’ (FRUIT) method35.
Plasmids pGB1-pGB36, pGB67–68 were constructed by cloning the suhB gene and 200 bp of upstream sequence into the pAMD-BA-lacZ plasmid34, creating transcriptional or translational fusions to lacZ. Plasmids pGB192-pGB193 included 200 bp of upstream sequence and 57 nt of suhB coding sequence followed by a stop codon. Fusions carrying boxA mutations were made by amplifying a suhB fragment from GB023 (boxA(C4T/T6C)) or by site-directed mutagenesis (boxA(C4A)); suhB start codon mutations (ATG → CAG) were made using site-directed mutagenesis. Insertions between the boxA and S-D sequences were generated by cloning fragments of random non-coding sequence (‘GAACTACCCATCTGGTCGCAGATAGTATGAAC′), modified from ref. 36, for insertions of up to 32 bp; 40–100 bp insertions carried a non-coding sequence from the 16S RNA gene in the reverse orientation (region from +1281 to +1380). The 5′ end of the insert remained the same, and inserted sequence was extended towards the S-D sequence (see Supplementary Fig. 6 for details). Plasmid pGB116 was made by cloning the T7 RNAP gene with a S-D sequence into pBAD1837. Plasmids pGB83–95 carried the suhB gene and 36 nt of the 5′UTR with wt or mutant boxA, and a 100 nt insertion between the BoxA and S-D elements, where indicated. suhB was under the control of pT7 promoter and was translationally fused to lacZ reporter on pAMD-BA-lacZ plasmid34. Plasmids pGB109–110 were made by cloning CKO_00699(R82A) gene with wt or mutant boxA (C4A) and a constitutive promoter25; the toxin gene was transcriptionally fused to lacZ reporter on pAMD-BA-lacZ plasmid.
Isolation and identification of trans- and cis-acting mutants
The trans-acting mutant genetic selection was performed using pJTW067 plasmid carrying a suhB-lacZ transcriptional fusion in MG1655 ΔlacZ. pJTW067 was isolated as an inactive suhB-lacZ clone from a cis-acting mutant screen (see below) and contains a single nucleotide mutation (T607C) that does not affect the expression of suhB. It is otherwise identical to pJTW067. Bacterial cultures were grown at 37 °C in LB medium. One hundred microlitres of an overnight culture was washed and plated on M9+0.2% lactose agar. Spontaneous survivors were first tested for increased plasmid copy number using qPCR, comparing the Ct values of plasmid and chromosomal amplicons. Strains with increased copy number were discarded. To eliminate plasmid mutants, plasmids were isolated and transformed into a clean MG1655 ΔlacZ background and plated on MacConkey agar indicator plates; mutants forming red colonies (upregulated suhB-lacZ) were discarded. Chromosomal mutations were identified either by PCR amplification and sequencing of nusB, nusE and nusG, or by whole genome sequencing. Specifically, genomic DNA was purified using a DNeasy blood and tissue kit (Qiagen), DNA libraries were prepared using a Nextera kit (Illumina), sequencing was performed on an Illumina MiSeq instrument, and sequence variants were identified using the CLC genomic workbench (default parameters). The cis-acting mutant genetic screen was performed by cloning a mutant suhB DNA library, generated by an error-prone DNA polymerase Taq (NEB) with oligonucleotides JW3605 and JW3607, into the pAMD-BA-lacZ vector, which was transformed into EPI300 background (lac -; Epicentre). The mutant library included the entire promoter, 5′ UTR and gene. We selected mutants that were visibly upregulated on MacConkey agar plates and sequenced the insert to identify mutations.
ChIP-qPCR
Bacteria were grown at 37 °C in LB medium until OD600 = 0.5–0.6. ChIP-qPCR was performed as described previously34, using monoclonal mouse anti-RpoB (Neoclone #W0002) and M2 monoclonal anti-FLAG (Sigma) antibodies. Occupancy units represent background-subtracted enrichment scores relative to transcriptionally silent regions within the bglB or ynbB genes in E. coli, and the sbcC gene in S. Typhimurium.
β-galactosidase assays
Bacterial cultures were grown at 37 °C in LB medium to an OD600 of 0.5–0.6. LB medium was supplemented with 0.2% arabinose when pBAD18 or its derivatives were used. A volume of 100–200 μL of culture was pelleted and resuspended in 800 μL of Z buffer (0.06 M Na2HPO4, 0.04 M NaH2PO4, 0.01 M KCl, 0.001 M MgSO4) supplemented with β-mercaptoethanol (50 mm final concentration), sodium dodecyl sulfate (0.001% final concentration), and 20 μL chloroform. Assays were initiated by adding 160 μL of 2-nitrophenyl β-d-galactopyranoside (4 mg/mL) and stopped by adding 400 μL of 1 M Na2CO3. The duration of the reaction and OD420 readings were recorded and β-galactosidase activity units were calculated as 1000× (A420/(A600)(timemin)).
Western blotting
Bacteria were grown at 37 °C in LB to an OD600 of 0.5–0.6. Cell pellets were boiled in gel loading dye, separated on gradient polyacrylamide gels (Bio-Rad), and transferred to a polyvinylidene fluoride membrane (Thermo Scientific). The membrane was cut in half at the 50 kDa molecular marker. The upper part of the membrane was probed with control mouse monoclonal anti-RpoC (BioLegend) antibody at 1:4000 dilution, and the lower part was probed with mouse monoclonal M2 anti-FLAG (Sigma) antibody at 1:10,000 dilution. Goat anti-mouse horseradish peroxidase-conjugated antibody was used for secondary probing at 1:20,000 dilution. Both parts of the membrane were always processed in parallel. Blots were developed with Clarity™ Western ECL Substrate (Bio-Rad).
Sequence alignment of suhB upstream regions
We extracted 100 bp of upstream sequence for suhB homologues in 19 species of the family Enterobacteriaciae, and aligned the sequences using MUSCLE38 (Supplementary Fig. 2). To determine the % match to E. coli at each position, we added 1 to the number of perfect matches (to account for the E. coli sequence), divided by 20 (to account for the 20 species in the alignment), and converted to a percentage.
Identification of putative boxA sequences in γ-proteobacterial genomes
We searched all sequenced γ-proteobacterial genomes for annotated protein-coding genes with the sequence GYTCTTTAANA within the 50 nt upstream of the annotated gene start. We compared gene functions using COG annotations39.
Data availability
All relevant data supporting the findings of the study are available in this article and its Supplementary Information files. All computer codes are available from the corresponding author upon request.
Electronic supplementary material
Description of Additional Supplementary Files
Acknowledgements
We thank Dave Grainger, Don Court, Todd Gray and Keith Derbyshire for helpful discussions and comments on the manuscript. We thank the Wadsworth Center Bioinformatics and Statistics Core Facility, and the Wadsworth Center Media and Tissue Culture Core Facility for technical assistance. This work was supported by the NIH Director’s New Innovator Award Program, 1DP2OD007188 (J.T.W.).
Author contributions
J.T.W., G.B. and N.S. designed the study. J.T.W. and G.B. wrote the manuscript. G.B., N.S., C.B., R.J., R.F., M.P., A.M.S. and A.S. generated experimental data. P.L. performed bioinformatic analysis. All authors contributed to data analysis and interpretation.
Competing interests
The authors declare no competing financial interests.
Footnotes
Electronic supplementary material
Supplementary Information accompanies this paper at 10.1038/s41467-017-02124-9.
References
- 1.Peters JM, Vangeloff AD, Landick R. Bacterial transcription terminators: the RNA 3′-end chronicles. J. Mol. Biol. 2011;412:793–813. doi: 10.1016/j.jmb.2011.03.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Arnvig KB, et al. Evolutionary comparison of ribosomal operon antitermination function. J. Bacteriol. 2008;190:7251–7257. doi: 10.1128/JB.00760-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bubunenko M, et al. Nus transcription elongation factors and RNase III modulate small ribosome subunit biogenesis in Escherichia coli. Mol. Microbiol. 2012;87:382–393. doi: 10.1111/mmi.12105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Luo X, et al. Structural and functional analysis of the E. coli NusB-S10 transcription antitermination complex. Mol. Cell. 2008;32:791–802. doi: 10.1016/j.molcel.2008.10.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nodwell JR, Greenblatt J. Recognition of boxA antiterminator RNA by the E. coli antitermination factors NusB and ribosomal protein S10. Cell. 1993;72:261–268. doi: 10.1016/0092-8674(93)90665-D. [DOI] [PubMed] [Google Scholar]
- 6.Burmann BM, et al. A NusE:NusG complex links transcription and translation. Science. 2010;328:501–504. doi: 10.1126/science.1184953. [DOI] [PubMed] [Google Scholar]
- 7.Prasch S, et al. RNA-binding specificity of E. coli NusA. Nucleic Acids Res. 2009;37:4736–4742. doi: 10.1093/nar/gkp452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Qayyum MZ, Dey D, Sen R. Transcription elongation factor NusA is a general antagonist of rho-dependent termination in Escherichia coli. J. Biol. Chem. 2016;291:8090–8108. doi: 10.1074/jbc.M115.701268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sen R, Chalissery J, Muteeb G. Nus factors of Escherichia coli. EcoSalPlus. 2008;3:4.5.3.1. doi: 10.1128/ecosalplus.4.5.3.1. [DOI] [PubMed] [Google Scholar]
- 10.Li SC, Squires CL, Squires C. Antitermination of E. coli rRNA transcription is caused by a control region segment containing lambda nut-like sequences. Cell. 1984;38:851–860. doi: 10.1016/0092-8674(84)90280-0. [DOI] [PubMed] [Google Scholar]
- 11.Torres M, Balada JM, Zellars M, Squires C, Squires CL. In vivo effect of NusB and NusG on rRNA transcription antitermination. J. Bacteriol. 2004;186:1304–1310. doi: 10.1128/JB.186.5.1304-1310.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Singh N, et al. SuhB associates with Nus factors to facilitate 30S ribosome biogenesis in Escherichia coli. mBio. 2016;7:e00114–e00116. doi: 10.1128/mBio.00114-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Peters JM, et al. Rho directs widespread termination of intragenic and stable RNA transcription. Proc. Natl. Acad. Sci. USA. 2009;106:15406–15411. doi: 10.1073/pnas.0903846106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Peters JM, et al. Rho and NusG suppress pervasive antisense transcription in Escherichia coli. Genes Dev. 2012;26:2621–2633. doi: 10.1101/gad.196741.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Inada T, Nakamura Y. Autogenous control of the suhB gene expression of Escherichia coli. Biochimie. 1996;78:209–212. doi: 10.1016/0300-9084(96)89508-3. [DOI] [PubMed] [Google Scholar]
- 16.Martinez A, Opperman T, Richardson JP. Mutational analysis and secondary structure model of the RNP1-like sequence motif of transcription termination factor Rho. J. Mol. Biol. 1996;257:895–908. doi: 10.1006/jmbi.1996.0210. [DOI] [PubMed] [Google Scholar]
- 17.Bossi L, Schwartz A, Guillemardet B, Boudvillain M, Figueroa-Bossi N. A role for Rho-dependent polarity in gene regulation by a noncoding small RNA. Genes Dev. 2012;26:1864–1873. doi: 10.1101/gad.195412.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wade JT, Grainger DC. Pervasive transcription: illuminating the dark matter of bacterial transcriptomes. Nat. Rev. Microbiol. 2014;12:647–653. doi: 10.1038/nrmicro3316. [DOI] [PubMed] [Google Scholar]
- 19.Mitra, P., Ghosh, G., Hafeezunisa, M. & Sen, R. Rho proteins: role and mechanisms. Annu. Rev. Microbiol.https://doi.org/10.1146/annurev-micro-030117-020432 (2017). [DOI] [PubMed]
- 20.Barik S, Ghosh B, Whalen W, Lazinski D, Das A. An antitermination protein engages the elongating transcription apparatus at a promoter-proximal recognition site. Cell. 1987;50:885–899. doi: 10.1016/0092-8674(87)90515-0. [DOI] [PubMed] [Google Scholar]
- 21.Pasman Z, von Hippel PH. Regulation of rho-dependent transcription termination by NusG is specific to the Escherichia coli elongation complex. Biochem. (Mosc.). 2000;39:5573–5585. doi: 10.1021/bi992658z. [DOI] [PubMed] [Google Scholar]
- 22.Wilson HR, Zhou JG, Yu D, Court DL. Translation repression by an RNA polymerase elongation complex. Mol. Microbiol. 2004;53:821–828. doi: 10.1111/j.1365-2958.2004.04170.x. [DOI] [PubMed] [Google Scholar]
- 23.Golomb M, Chamberlin M. Characterization of T7-specific ribonucleic acid polymerase. IV. Resolution of the major in vitro transcripts by gel electrophoresis. J. Biol. Chem. 1974;249:2858–2863. [PubMed] [Google Scholar]
- 24.Skinner GM, Baumann CG, Quinn DM, Molloy JE, Hoggett JG. Promoter binding, initiation, and elongation by bacteriophage T7 RNA polymerase. A single-molecule view of the transcription cycle. J. Biol. Chem. 2004;279:3239–3244. doi: 10.1074/jbc.M310471200. [DOI] [PubMed] [Google Scholar]
- 25.Burr T, Mitchell J, Kolb A, Minchin S, Busby S. DNA sequence elements located immediately upstream of the -10 hexamer in Escherichia coli promoters: a systematic study. Nucleic Acids Res. 2000;28:1864–1870. doi: 10.1093/nar/28.9.1864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kriner MA, Sevostyanova A, Groisman EA. Learning from the leaders: gene regulation by the transcription termination factor rho. Trends Biochem. Sci. 2016;41:690–699. doi: 10.1016/j.tibs.2016.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sedlyarova N, et al. sRNA-mediated control of transcription termination in E. coli. Cell. 2016;167:111–121. doi: 10.1016/j.cell.2016.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wilson HR, Kameyama L, Zhou JG, Guarneros G, Court DL. Translational repression by a transcriptional elongation factor. Genes Dev. 1997;11:2204–2213. doi: 10.1101/gad.11.17.2204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wilson HR, Yu D, Peters HK, III, Zhou JG, Court DL. The global regulator RNase III modulates translation repression by the transcription elongation factor N. EMBO J. 2002;21:4154–4161. doi: 10.1093/emboj/cdf395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Aksoy S, Squires CL, Squires C, J. B. 1984. Evidence for antitermination in Escherichia coli rRNA transcription. J. Bacteriol. 1984;159:260–264. doi: 10.1128/jb.159.1.260-264.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Squires CL, Greenblatt J, Li J, Condon C, Squires CL. Ribosomal RNA antitermination in vitro: requirement for Nus factors and one or more unidentified cellular components. Proc. Natl. Acad. Sci. USA. 1993;90:970–974. doi: 10.1073/pnas.90.3.970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Richardson EJ, Watson M. The automatic annotation of bacterial genomes. Brief. Bioinform. 2013;14:1–12. doi: 10.1093/bib/bbs007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Blattner FR, et al. The complete genome sequence of Escherichia coli K-12. Science. 1997;277:1453–1462. doi: 10.1126/science.277.5331.1453. [DOI] [PubMed] [Google Scholar]
- 34.Stringer AM, et al. Genome-scale analyses of Escherichia coli and Salmonella enterica AraC reveal non-canonical targets and an expanded core regulon. J. Bacteriol. 2014;196:660–671. doi: 10.1128/JB.01007-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Stringer AM, et al. FRUIT, a scar-free system for targeted chromosomal mutagenesis, epitope tagging, and promoter replacement in Escherichia coli and Salmonella enterica. PLoS ONE. 2012;7:e44841. doi: 10.1371/journal.pone.0044841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen Q, Kadner RJ. Effect of altered spacing between uhpT promoter elements on transcription activation. J. Bacteriol. 2000;182:4430–4436. doi: 10.1128/JB.182.16.4430-4436.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995;177:4121–4130. doi: 10.1128/jb.177.14.4121-4130.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–1797. doi: 10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Galperin MY, Makarova KS, Wolf YI, Koonin EV. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2015;43:D261–D269. doi: 10.1093/nar/gku1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Description of Additional Supplementary Files
Data Availability Statement
All relevant data supporting the findings of the study are available in this article and its Supplementary Information files. All computer codes are available from the corresponding author upon request.