

Abstract
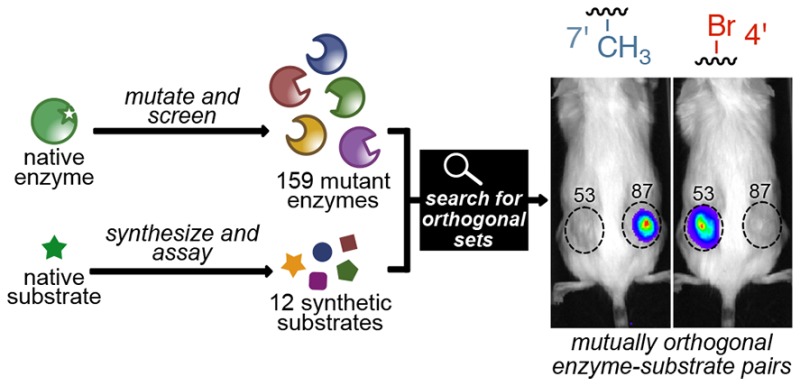
Bioluminescence imaging with luciferase enzymes and luciferin small molecules is a well-established technique for tracking cells and other biological features in rodent models. Despite its popularity, bioluminescence has long been hindered by a lack of distinguishable probes. Here we present a method to rapidly identify new substrate-selective luciferases for multicomponent imaging. Our strategy relies on parallel screening of luciferin analogues with panels of mutant enzymes. The compiled data set is then analyzed in silico to uncover mutually orthogonal sets. Using this approach, we screened 159 mutant enzymes with 12 luciferins. Thousands of orthogonal pairs were revealed with sufficient selectivity for use in biological environments. Over 100 pairs were validated in vitro, and three were applied in cell and animal models. The parallel screening method is both generalizable and scalable and will streamline the search for larger collections of orthogonal probes.
Short abstract
We screened sterically modified luciferins with libraries of mutant luciferases. Substrate-selective (orthogonal) pairs were identified using a computer script, and validated in biological assays.
Introduction
Our understanding of living systems is profoundly shaped by our ability to “see” biology in action. Central to these efforts are robust and translatable imaging tools.1,2 Decades of work to engineer and optimize fluorescent proteins have provided a palette of designer probes for cellular studies. Using combinations of these tools, it is now possible to trace the orchestrated behaviors of immune cells,3 nerve cell connections,4 and other interactions.5 Widespread application of fluorescent probes will continue to reveal unanticipated facets of biology. Concurrently, gaps in our knowledge will spur the development of innovative tools. Fluorescent probes endowed with novel functions (altered colors, photoswitches, etc.) are already enabling new pursuits.6,7
Biological discoveries will be further bolstered by advances in bioluminescent probe development. Bioluminescence relies on light generation via luciferase enzymes and luciferin small molecules.8,9 Since no excitation light is required, this modality is attractive for studies involving large length or time scales.10−12 Indeed, firefly luciferase (Fluc) and its cognate substrate (d-luciferin) are ubiquitously used in rodent models to interrogate molecular and cellular events.13,14 Fluc and its homologues have been further engineered to provide different colors of light.15,16 Such tools have been applied for multicomponent imaging in vitro. The wavelengths achieved, though, are unsatisfactory for routine use in vivo. Multicomponent imaging with even the most spectrally resolved probes remains challenging due to interference from surrounding tissue.10 As a consequence, bioluminescence has lagged behind fluorescence for multicellular studies in tissues and whole organisms.
To realize multicomponent imaging in vivo, we turned to a more tractable parameter: substrate selectivity. Enzymes exhibiting mutually exclusive (i.e., orthogonal) substrate preferences should be readily distinguished in a variety of biological models. Fluc is remarkably tolerant of a variety of luciferin modifications, including both electronic17−21 and steric22−26 derivatives. While dozens of luciferin analogues have been crafted, most result in reduced photon outputs relative to d-luciferin, the native substrate, at saturating doses.27 In some cases, boosts in light emission have been achieved using modified versions of the enzyme.23,25 These results set the stage for developing designer luciferase–luciferin pairs, but few methods to systematically generate orthogonal sets have been pursued.25,28 Substrate-selective luciferases are found in nature, and a handful have been coopted for dual imaging (e.g., combinations of d-luciferin- and coelenterazine-utilizing enzymes).29−32 However, most remain suboptimal for use in vivo. The characterization of other naturally occurring luciferases and luciferins has also not kept pace with the demand for new pairs. Consequently, bioluminescence imaging has been limited by a lack of mutually orthogonal enzymes and substrates.
We aimed to expedite the search for luciferases that exhibit unique preferences for distinct luciferin analogues. Accessing enzymes with alternative substrate use is well precedented in directed evolution.33−35 However, traditional applications of this technique have focused on optimizing one enzyme at a time. Selectivity for one molecule over another is often realized as a consequence, but is not typically the parameter being screened.36−41 Here, we present a general and rapid approach to achieve substrate selectivity and engineer orthogonal luciferase–luciferin pairs. This strategy relies on parallel screening of functional luciferases with collections of chemically diverse luciferins (Figure 1a). The large data sets are then mined for orthogonal combinations using a custom computer script. Enzyme–substrate pairs are deemed orthogonal if robust reactivity is observed when complementary partners interact, but minimal to no reactivity is observed in all other cases (Figure 1a). Collectively, we screened 159 mutants and 12 analogues, generating a candidate list of greater than 800,000 possible pairs. We evaluated the orthogonality of 175 pairs in vitro. A subset was successfully applied in cultured cell and animal models, highlighting the feasibility and translatability of the approach. We also analyzed principles governing selective substrate use and identified methods to search for expanded collections of orthogonal imaging agents. Overall, this work greatly expands the number of viable bioluminescence probes for multicomponent imaging and presents a strategy to accelerate the identification of new ones. The parallel screening method is also applicable to other areas where selective substrate use is required.
Figure 1.

Parallel screening of luciferase mutants and luciferin analogues to identify orthogonal pairs. (a) General strategy for identifying substrate-selective, mutually orthogonal enzymes. For bioluminescent probes, positive (matched) pairs are enzyme–substrate combinations that provide robust light emission. Negative (mismatched) pairs are combinations that exhibit reduced photon outputs. (b) Collections of 4′- and 7′-modified luciferins used for parallel screening. The molecules were synthesized from a common intermediate. (c) Fluc residues targeted for mutagenesis to accommodate 4′- and 7′-modified luciferins (shaded in red and blue hues, respectively; PDB structure: 4G36). A bound d-luciferin-AMP analogue (yellow) is shown for reference.
Results and Discussion
Expanding the Pool of Candidate Luciferins and Luciferases
As a starting point for substrate modification, we focused on d-luciferin derivatives with steric appendages at C4′ and C7′. These positions lie in close proximity to the Fluc backbone,42 and preliminary work revealed that modifications here do not quench or otherwise impede photon emission.25 We also previously identified a pair of luciferases that could discriminate between luciferins with modifications at these positions,25 suggesting that they were good starting points for new probe development. However, attempts to optimize this pair via traditional directed evolution (focusing on one enzyme at a time) did not result in improved substrate selectivity (Figure S1).
We reasoned that screening for selectivity at the outset would provide a more rapid route to new bioluminescent pairs. Engineering luciferases to discriminate among structurally similar compounds can be difficult.23,43 Thus, we initially focused on diversifying the enzyme and substrate inputs. Collections of both new and known24,25 luciferins were assembled (Figure 1b). These molecules covered a broad range of chemical space and comprised both hydrophilic and hydrophobic functional groups. The luciferins were benchmarked for light emission with Fluc (Figure S2). All compounds were functional light emitters, though they varied in terms of photon output. Some level of enzyme activity is necessary for successful evolution, but weak performers can be advantageous starting points for evolving new functions.33
In parallel with luciferin diversification, we targeted broad sectors of Fluc sequence space for mutagenesis. Twenty-three residues near the active site were selected, and the mutations were covered in 8 libraries (labeled in Figure 1c). The majority of the mutants would likely be nonfunctional, and thus not ideal starting points for probe development. We aimed to eliminate these luciferases early on and perform parallel screens with an enriched pool of viable mutants. Such an approach would save time and reagents as luciferases are not amenable to high-volume separations (e.g., FACS) or selections; rather, each mutant must be physically interrogated with a given substrate. We adapted a high-throughput method to traverse the luciferase libraries and cull nonfunctional members (Figure S3a).25 The libraries were transformed into bacteria, and the transformants were grown on agar containing one of four minimally perturbed luciferins: 4′/7′-BrLuc or 4′/7′-MeLuc (Figure 1b, Figure S3a). These analogues were selected for on-plate screens since they are among the “brightest” emitters and easy to access in bulk. Light-emitting colonies were picked and further assayed in lysate and by sequencing (Figure S3a). A variety of mutants were identified (Figure S4), including enzymes that were unique to each luciferin. Some hits were further diversified (1–3 generations) via random mutagenesis to enlarge the pool of luciferase mutants (Table S1 and Figure S3).
Screening for Orthogonal Luciferase–Luciferin Pairs in Silico
With enriched sets of functional luciferases, we aimed to screen the collection for orthogonal pairs. Testing each combination of two mutants and two substrates would have required 829,026 separate experiments (Figure 2a), an impractical number. Instead, we screened each analogue across the same panel of 159 luciferases, generating 1908 (12 substrates × 159 enzymes) individual data points (Figure S3b). An ideal orthogonal enzyme would be “positively” matched with a single substrate and “negatively” matched with all other luciferins. To identify such enzymes, we established a metric to quantify orthogonality and mine the data. We reasoned that perfect selectivity could be represented by an identity matrix (Figure S5, Supplementary Note). Orthogonality would be maximal if each enzyme was completely selective for its cognate substrate (represented by a “1” in the identity matrix) and nonfunctional with other luciferins (“0” in the identity matrix). An orthogonality score was determined by representing each set of two luciferases and two luciferins as a square matrix, with enzymes in rows and substrates in columns. These data were compared to the ideal case (identity matrix) via root-mean-square distance (RMSD). The RMSD values were then converted to numeric values (i.e., orthogonality scores) representing the fold resolution between the positive and negative pairings (see Supplementary Note for more details). We wrote a computer script to assemble each possible matrix from the screening data and calculate the orthogonality of each pairing. The pairs were sorted by increasing RMSD, with the smallest value (highest orthogonality score) representing the most orthogonal pair.
Figure 2.

Uncovering orthogonal pairs in silico. (a) Computational approach to identifying orthogonal sets. Parallel screens of mutant enzymes (Em) and substrate analogues (Sn, where n and m are integers) were performed and light emission values entered into a database. Data were analyzed with a custom computer script to identify orthogonal sets. (b) Sample orthogonal bioluminescent probes. Bacteria expressing mutant enzymes were expanded, lysed, and distributed evenly among replicate wells. Lysates were treated with luciferin analogues and imaged. Representative images are shown, along with quantified photon outputs. (c) Orthogonality scores correlated with computer script rank. Orthogonal sets predicted in silico were verified biochemically as in panel b. Each bar (beyond rank 11) represents >40 unique sets that were evaluated in head-to-head comparisons in vitro. (For interval 1-10, all ten orthogonal sets were examined). For panels b and c, error bars represent the standard error of the mean for n ≥ 3 experiments.
The algorithm provided a ranked list of the 829,026 possible orthogonal sets (Figure 2a). The top pair comprised analogues 2 and 11 (4′-MorphoLuc and 7′-MorPipLuc) with mutants 81 and 104 (Figure 2a). Selective light emission with these enzymes and substrates was verified in vitro (Figure 2b). We further validated the top ten unique pairings on the ranked list, along with a handful of others in the data set (every tenth rank among the top 100, every 100th rank among the top 1000, and every 1000th rank down to position 5000). In all cases, orthogonality scores were measured in bacterial lysate (Figure 2c). Among the top 1000 pairs, >10-fold photon outputs were observed with the positively paired luciferase–luciferin set compared to the negatively paired set (Figure 2c). Diminishments in selectivity were observed farther down the list. These results suggest that the in silico rank order is a good predictor of orthogonal substrate use. The method also culled 99.9% (∼828,000 of the total 829,026) of irrelevant enzyme–substrate pairings (Figure S6), enabling fast convergence on important hits. As more luciferases and luciferins are screened, the data set can be expanded and continually mined for new orthogonal pairs.
Imaging with Orthogonal Pairs in Cultured Cell and Animal Models
We aimed to transition lead pairs from the screening analyses to mammalian cell imaging. In these more complex environments, issues of enzyme stability, substrate biocompatibility, and compound transport are of paramount concern. Fortunately, our approach to enriching functional luciferases preselects for luciferases and luciferins that are well behaved. Three of the top pairs from the script were analyzed in cultured cell (Figure S7) and animal models (Figure 3): (1) 4′-MorphoLuc/enzyme 81 (R218A, F250M, S314T, G316T) with 7′-DMAMeLuc/enzyme 37 (R218K), (2) 7′-MeLuc/enzyme 87 (R218K, F250Y, S314T, G316T) with 4′-BrLuc/enzyme 53 (V240I, V241M, F243M, F247Y, S347G), and (3) 4′-BrLuc/enzyme 51 (F243M, S347G) with d-luciferin/enzyme 93 (R218K, M249L, S314T, G316S). These pairs were selected due to the ease of accessing the substrates, along with their relative brightness. The mutants were stably expressed in DB7 mouse mammary carcinoma cells. The cells were treated with relevant luciferins and imaged (Figure S7). Substrate specificity was maintained in all cases, highlighting the success of the parallel screening method.
Figure 3.

Noninvasive in vivo imaging with orthogonal pairs. DB7 cells expressing mutants 37 and 81 (a), 51 and 93 (b), or 53 and 87 (c) were inoculated in opposing flanks of FVB/NJ mice. The sites of implantation are indicated with dashed circles. Luciferin analogues were administered ip, and light emission was recorded. Representative bioluminescence images are shown for each set. For panel a, images were acquired 5 days post cell implantation. For panels b and c, images were acquired 3 days post cell implantation. Photon outputs were quantified and plotted. Black lines represent mean photon intensities for n = 3 mice in each set.
Selectivity was also maintained in vivo. DB7 cells expressing the relevant mutants were implanted in FVB/NJ mice. Subsequent administration of the complementary luciferin analogues resulted in light emission for positively paired compounds with minimal cross reactivity (Figure 3). These images mark an initial demonstration of dual imaging with purely engineered luciferase–luciferin pairs. It is also important to note that perfect resolution is not required for multicomponent imaging applications. Rather, patterns of substrate use can serve as diagnostic fingerprints.44 Photon outputs from the orthogonal pairs are in a useful range for monitoring bulk cell populations. The dimmest set (enzyme 37/7′-DMAMeLuc and enzyme 81/4′-MorphoLuc) emits enough photons to visualize ∼6 × 106 cells in subcutaneous models. The other orthogonal sets are substantially brighter and can enable more sensitive imaging. Collectively, these data show that parallel screens and in silico analyses can be used to identify and transition orthogonal sets to a variety of biological models.
Analyzing Trends in Orthogonal Substrate Use
To gain insight into principles governing orthogonality, we undertook a detailed analysis of the screening results. The highest-ranked pairs comprised a variety of enzymes and substrates. Seven unique luciferins (from both the 4′ and 7′ series) were found among the top 10 pairs, along with luciferases comprising mutations at 18 unique sites (Supplementary Data; Figure S8). The diversity in hits implies that there are a variety of paths to achieve substrate resolution. Among the pairs, orthogonality was primarily realized not by markedly enhanced turnover of a preferred substrate. Rather, selectivity arose from reduced photon production with other compounds. As shown in Figure 4a, matched enzymes and substrates (positive pairs) were on par with native Fluc in terms of photon output. The unmatched enzymes and substrates (negative pairs), by contrast, demonstrated reduced activities (∼10–1000-fold lower). Thus, in a given orthogonal pair, selectivity is mostly achieved by reducing light emission with the negatively paired compound versus selectively increasing light emission with the positively paired compound. For example, mutant 81 provides ∼4-fold enhanced light output with 4′-MorphoLuc compared to Fluc. With every other luciferin screened, including the negatively paired compound 7′-MorPipLuc, mutant 81 emits >10-fold fewer photons than the native enzyme. So while light output with 4′-MorphoLuc is slightly improved with mutant 81, the decrease in light emission observed with 7′-MorPipLuc (>100-fold) contributes more to orthogonality.
Figure 4.

Examining the origins of substrate selectivity. (a) In a given orthogonal pair, each luciferase retains activity with the matched luciferin (positive pairing, colored circle), while losing activity with the mismatched analogue (negative pairing, colored square). Mutant luciferases were also poorly reactive with all other mismatched analogues examined (open circles). These data suggest that orthogonality arises from selective retention of activity with a single compound. Light emission values are plotted relative to native Fluc and the indicated luciferin. (b) Frequency of luciferin analogue pairings predicted to be orthogonal. The majority of orthogonal sets from the top 5000 pairs (0.6%) comprise structurally divergent compounds (i.e., 4′-modified luciferins paired with 7′-modified luciferins).
Since compound selectivity appears to be achieved by destroying enzyme–substrate interactions, structurally related compounds would be expected to exhibit similar trends in orthogonality. Indeed, bulky 7′-modified compounds tend to positively pair with the same types of enzymes (Figure S9). Many of these mutants (e.g., mutant 104) likely harbor space in the active site to accommodate 7′ substituents (Figure S9a). Conversely, 4′-modified luciferins tend to produce less light with these same mutants and are thus negatively paired. Structurally divergent compounds were also more likely to comprise an orthogonal pair (Figure 4b). For example, substrates with a modification at the 4′ position were rarely orthogonal to other 4′ compounds. It is probably difficult to destroy activity with one 4′-modified compound without impacting others in the same series. When 4′- and 7′-modified substrates are paired, though, each substrate likely interacts differently with the enzyme, making it easier to achieve orthogonality. These results suggest a strategy for continued orthogonal bioluminescent probe development: incorporate more diverse analogues in parallel screens.
Some compounds appear uniquely suited for orthogonal probe development. For example, 4′-BrLuc shows up nearly twice as often in the top 1,000 hits compared to other compounds (Figure S10). The mechanistic basis for this preference is unclear. The bromine substituent is roughly the same size as the methyl group in 4′-MeLuc, negating a pure steric argument. 4′-MeOHLuc is predicted to form orthogonal pairs (albeit less frequently) with similar compounds as 4′-BrLuc (Figure 4), suggesting that polarizable substituents might be preferred. Heavy halogen atoms (e.g., Br) are also known to quench the fluorescence of some molecules via intersystem crossing.45 Thus, certain 4′-BrLuc conformations could result in internal quenching (and poor light emission) and thus pair negatively with several mutants. Additional compound screens and analyses will be necessary to discriminate among these possibilities and gain more insight.
We further analyzed the frequency of positive and negative pairings between luciferins and individual residues (Figure 5). Luciferases with mutations at residues 240, 247, or 347 seemed to prefer 4′-modified compounds. These residues are known to modulate the binding and light emission of the native substrate, d-luciferin.43,46−48 Docking studies corroborated these findings, suggesting that the mutations (e.g., S347G in mutants 51 and 53) likely create space for bulky substituents (e.g., 4′-BrLuc, Figure S11). These residues are also negatively paired with most of the 7′-modified compounds, suggesting that they are good candidates for future orthogonal probe design. Surprisingly few hot spot residues correlated with selective 7′ analogue use (Figure 5). Fluc residues near C7′ primarily comprise backbone amides.42,49 Thus, it is unclear how specificity for these analogues might arise.
Figure 5.

Orthogonal pair analysis. Heat map of mutation frequency (at a given residue) for enzymes positively paired (top) or negatively paired (bottom) with luciferin analogues. For each plot, the top 5000 pairs from in silico analyses were examined. Enzyme residues are organized by their relative proximity to C4′ or C7′ of luciferin (as in Figure 1b).
Added Diversity Improves Orthogonality
Multicomponent imaging requires not just pairs of orthogonal enzymes and substrates, but also triplets, quadruplets, and higher order sets. Identifying such expanded collections requires structurally diverse enzyme and substrate architectures. If only a few privileged luciferases or luciferins from our data set could provide the desired selectivities, it would be difficult to achieve larger collections of orthogonal probes. To assess whether we were approaching an upper limit on orthogonality, we performed simulations within the existing data set. Random subsets of various sizes were selected from the full pool of substrates and enzymes. The sets were analyzed using the algorithm from above, and orthogonality scores were generated (Figure 6a). Regardless of the identities of enzymes or substrates used, scores increased with greater numbers of both enzymes (from 2 to 159) and substrates (from 2 to 12). This result implies that we have not reached a plateau in identifying orthogonal pairs. Exploring more sequence space with mutant luciferases and chemical space with modified luciferins should also improve the orthogonality of the top pairings.
Figure 6.

Improving orthogonality via enzyme–substrate diversity. (a) Orthogonality scores increase as more enzymes and substrates are considered. Computational analyses were performed on random subsets of luciferin analogues and mutant enzymes (from the entire data set). Orthogonality scores for all inputs were calculated as before, and the top orthogonal hits were averaged. (b) Validating an orthogonal triplet set. Bacteria expressing mutant luciferases 95, 53, or 81 were lysed and incubated with their corresponding luciferin (250 μM). Sample images are shown. Photon outputs were quantified and error bars represent the standard error of the mean for n = 3 experiments.
As a next step, we modified the algorithm to search for not just two pairs of orthogonal probes but also triplets and multiple sets in general. A set of three adds significant complexity, as not only three positive pairings, but also six negative pairings, must be identified. From our current data set, this required sifting through >144 million combinations. We combed the original data set in search of three mutually orthogonal enzyme–substrate pairs. A total of 6171 potential sets were identified. The orthogonalities of the top ten were verified in bacterial lysate (Figure S11). The top triplet set is shown in Figure 6b and comprises two enzyme–substrate pairs previously validated in vivo. The overall orthogonality score for this set was lower than that of the individual pairs from above. However, this result represents key proof-of-concept and a starting point for the development of larger collections of mutually orthogonal luciferase–luciferin sets. Perfect selectivity is also not required for using the probes in biological environments. Rather, patterns in substrate use are most important and can be discerned using standard imaging equipment and rates of change in photon output.
Conclusions
We developed a general and rapid strategy to engineer orthogonal luciferase–luciferin pairs. The method relies on developing an initial pool of functional enzymes and screening the collection with chemically diverse luciferins. Using this approach, we generated >800,000 possible pairings and mined the data for orthogonal pairs with a custom computer algorithm. Dozens of candidates were identified and validated in vitro. A handful of hits were further translated into cultured cell and animal models, greatly expanding the number of bioluminescent probes for multicomponent imaging.
We further analyzed the principles governing orthogonal substrate use. Chemical and sequence diversity was key to eliciting high levels of selectivity. Thus, the addition of more luciferins and libraries to our “living” data set should improve orthogonality and lower the barrier to identifying higher-order sets. A fleet of sensitive, selective pairs will bolster imaging capabilities and push the boundaries of what we can “see” and learn about biological systems. The methods reported here are also applicable beyond the field of bioluminescence. Parallel screens and in silico analyses can expedite the search for other orthogonal enzyme–substrate or protein–ligand pairs relevant to optogenetics, cell signaling, and other disciplines.
Acknowledgments
This work was supported by the U.S. National Institutes of Health (R01 GM107630 to J.A.P.). W.B.P. was supported by the National Science Foundation via the BEST IGERT program (DGE-1144901) and an Allergan Graduate Fellowship. K.A.J. was supported by an institutional Chemical and Structural Biology Training Grant predoctoral fellowship (T32-GM10856). C.M.R. was supported by the National Science Foundation Graduate Research Fellowship under Grant No. DGE-1321846. We thank Brendan Zhang for assisting with luciferin docking studies, along with members of the Weiss and Martin laboratories for reagents. We also thank Prof. Aaron Leconte (Claremont-McKenna) and members of the Prescher laboratory for helpful discussions.
Supporting Information Available
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acscentsci.7b00394.
Experimental details on luciferin synthesis, library development, parallel screens, and bioluminescence imaging (PDF)
Supplementary Note giving details on evaluation of orthogonal enzyme–substrate pairs (PDF)
The full list of predicted orthogonal pairs is also provided and details on custom computer script (XLSX)
Author Contributions
‡ C.M.R., W.B.P., and K.A.J. contributed equally to this work.
The authors declare no competing financial interest.
Supplementary Material
References
- Specht E. A.; Braselmann E.; Palmer A. E. A critical and comparative review of fluorescent tools for live-cell imaging. Annu. Rev. Physiol. 2017, 79, 93–117. 10.1146/annurev-physiol-022516-034055. [DOI] [PubMed] [Google Scholar]
- Lavis L. D.; Raines R. T. Bright building blocks for chemical biology. ACS Chem. Biol. 2014, 9, 855–866. 10.1021/cb500078u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Germain R. N.; Robey E. A.; Cahalan M. D. A decade of imaging cellular motility and interaction dynamics in the immune system. Science 2012, 336, 1676–1681. 10.1126/science.1221063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai D.; Cohen K. B.; Luo T.; Lichtman J. W.; Sanes J. R. Improved tools for the Brainbow toolbox. Nat. Methods 2013, 10, 540–547. 10.1038/nmeth.2450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porterfield W. B.; Prescher J. A. Tools for visualizing cell-cell ’interactomes’. Curr. Opin. Chem. Biol. 2015, 24, 121–130. 10.1016/j.cbpa.2014.11.006. [DOI] [PubMed] [Google Scholar]
- Rodriguez E. A.; et al. The growing and glowing toolbox of fluorescent and photoactive proteins. Trends Biochem. Sci. 2017, 42, 111–129. 10.1016/j.tibs.2016.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Betzig E.; et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006, 313, 1642–1645. 10.1126/science.1127344. [DOI] [PubMed] [Google Scholar]
- Prescher J. A.; Contag C. H. Guided by the light: visualizing biomolecular processes in living animals with bioluminescence. Curr. Opin. Chem. Biol. 2010, 14, 80–89. 10.1016/j.cbpa.2009.11.001. [DOI] [PubMed] [Google Scholar]
- Branchini B. R.; et al. Experimental support for a single electron-transfer oxidation mechanism in firefly bioluminescence. J. Am. Chem. Soc. 2015, 137, 7592–7595. 10.1021/jacs.5b03820. [DOI] [PubMed] [Google Scholar]
- Zhao H.; et al. Emission spectra of bioluminescent reporters and interaction with mammalian tissue determine the sensitivity of detection in vivo. J. Biomed. Opt. 2005, 10, 041210. 10.1117/1.2032388. [DOI] [PubMed] [Google Scholar]
- Rumyantsev K. A.; Turoverov K. K.; Verkhusha V. V. Near-infrared bioluminescent proteins for two-color multimodal imaging. Sci. Rep. 2016, 6, 36588. 10.1038/srep36588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troy T.; Jekic-McMullen D.; Sambucetti L.; Rice B. Quantitative comparison of the sensitivity of detection of fluorescent and bioluminescent reporters in animal models. Mol. Imaging 2004, 3, 9–23. 10.1162/153535004773861688. [DOI] [PubMed] [Google Scholar]
- Adams S. T.; Miller S. C. Beyond D-luciferin: Expanding the scope of bioluminescence imaging in vivo. Curr. Opin. Chem. Biol. 2014, 21, 112–120. 10.1016/j.cbpa.2014.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu T. T.; et al. The expanding toolbox of in vivo bioluminescent imaging. Front. Oncol. 2016, 6 (150), 1–8. 10.3389/fonc.2016.00150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branchini B. R.; et al. Thermostable red and green light-producing firefly luciferase mutants for bioluminescent reporter applications. Anal. Biochem. 2007, 361, 253–262. 10.1016/j.ab.2006.10.043. [DOI] [PubMed] [Google Scholar]
- Nakatsu T.; et al. Structural basis for the spectral difference in luciferase bioluminescence. Nature 2006, 440, 372–376. 10.1038/nature04542. [DOI] [PubMed] [Google Scholar]
- Woodroofe C. C.; et al. Novel heterocyclic analogues of firefly luciferin. Biochemistry 2012, 51, 9807–9813. 10.1021/bi301411d. [DOI] [PubMed] [Google Scholar]
- McCutcheon D. C.; Paley M. A.; Steinhardt R. C.; Prescher J. A. Expedient synthesis of electronically modified luciferins for bioluminescence imaging. J. Am. Chem. Soc. 2012, 134, 7604–7607. 10.1021/ja301493d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conley N. R.; Dragulescu-Andrasi A.; Rao J. H.; Moerner W. E. A selenium analogue of firefly D-luciferin with red-shifted bioluminescence emission. Angew. Chem., Int. Ed. 2012, 51, 3350–3353. 10.1002/anie.201105653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuchimaru T.; et al. A luciferin analogue generating near-infrared bioluminescence achieves highly sensitive deep-tissue imaging. Nat. Commun. 2016, 7, 11856. 10.1038/ncomms11856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branchini B. R.; Hayward M. M.; Bamford S.; Brennan P. M.; Lajiness E. J. Naphthyl- and quinolylluciferin: green and red light emitting firefly luciferin analogs. Photochem. Photobiol. 1989, 49, 689–695. 10.1111/j.1751-1097.1989.tb08442.x. [DOI] [PubMed] [Google Scholar]
- Woodroofe C. C.; et al. N-Alkylated 6′-aminoluciferins are bioluminescent substrates for Ultra-Glo and QuantiLum luciferase: New potential scaffolds for bioluminescent assays. Biochemistry 2008, 47, 10383–10393. 10.1021/bi800505u. [DOI] [PubMed] [Google Scholar]
- Mofford D. M.; Reddy G. R.; Miller S. C. Aminoluciferins extend firefly luciferase bioluminescence into the near-infrared and can be preferred substrates over D-luciferin. J. Am. Chem. Soc. 2014, 136, 13277–13282. 10.1021/ja505795s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinhardt R. C.; et al. Brominated luciferns are versatile bioluminescent probes. ChemBioChem 2017, 18, 96–100. 10.1002/cbic.201600564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones K. A.; et al. Orthogonal luciferase-luciferin pairs for bioluminescence imaging. J. Am. Chem. Soc. 2017, 139, 2351–2358. 10.1021/jacs.6b11737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jathoul A. P.; Grounds H.; Anderson J. C.; Pule M. A. A dual-color far-red to near-infrared firefly luciferin analogue designed for multiparametric bioluminescence imaging. Angew. Chem., Int. Ed. 2014, 53, 13059–13063. 10.1002/anie.201405955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rathbun C. M.; Prescher J. A. Bioluminescent Probes for Imaging Biology beyond the Culture Dish. Biochemistry 2017, 56, 5178–5184. 10.1021/acs.biochem.7b00435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishihara R.; et al. Luciferase-specific coelenterazine analogues for optical contamination-free bioassays. Sci. Rep. 2017, 7 (908), 1–8. 10.1038/s41598-017-00955-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paley M. A.; Prescher J. A. Bioluminescence: a versatile technique for imaging cellular and molecular features. MedChemComm 2014, 5, 255–267. 10.1039/C3MD00288H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stacer A. C.; et al. NanoLuc reporter for dual luciferase imaging in living animals. Mol. Imaging 2013, 12, 457–469. [PMC free article] [PubMed] [Google Scholar]
- Petushkov V. N.; et al. A novel type of luciferin from the Siberian luminous earthworm Fridericia heliota: structure elucidation by spectral studies and total synthesis. Angew. Chem., Int. Ed. 2014, 53, 5566–5568. 10.1002/anie.201400529. [DOI] [PubMed] [Google Scholar]
- Kaskova Z. M.; et al. Mechanism and color modulation of fungal bioluminescence. Sci. Adv. 2017, 3, e1602847. 10.1126/sciadv.1602847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renata H.; Wang Z.J.; Arnold F. H. Expanding the Enzyme Universe: Accessing non-natural reactions by mechanism-guided directed evolution. Angew. Chem., Int. Ed. 2015, 54, 3351–3367. 10.1002/anie.201409470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldsmith M.; Tawfik D. S. Directed enzyme evolution: beyond the low-hanging fruit. Curr. Opin. Struct. Biol. 2012, 22, 406–412. 10.1016/j.sbi.2012.03.010. [DOI] [PubMed] [Google Scholar]
- Packer M. S.; Liu D. R. Methods for the directed evolution of proteins. Nat. Rev. Genet. 2015, 16, 379–394. 10.1038/nrg3927. [DOI] [PubMed] [Google Scholar]
- Taylor N. D.; et al. Engineering an allosteric transcription factor to respond to new ligands. Nat. Methods 2016, 13, 177–183. 10.1038/nmeth.3696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng J.; et al. A general strategy to construct small molecule biosensors in eukaryotes. eLife 2015, 4, e10606. 10.7554/eLife.10606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang K.; Chin J. W. Cellular Incorporation of unnatural amino acids and bioorthogonal labeling of proteins. Chem. Rev. 2014, 114, 4764–4806. 10.1021/cr400355w. [DOI] [PubMed] [Google Scholar]
- Neumann H.; Wang K.; Davis L.; Garcia-Alai M.; Chin J. W. Encoding multiple unnatural amino acids via evolution of a quadruplet-decoding ribosome. Nature 2010, 464, 441–444. 10.1038/nature08817. [DOI] [PubMed] [Google Scholar]
- Wan W.; et al. A facile system for genetic incorporation of two different noncanonical amino acids into one protein in Escherichia coli. Angew. Chem., Int. Ed. 2010, 49, 3211–3214. 10.1002/anie.201000465. [DOI] [PubMed] [Google Scholar]
- Owens A. E.; Grasso K. T.; Ziegler C. A.; Fasan R. Two-tier screening platform for directed evolution of aminoacyl-tRNA synthetases with enhanced stop codon suppression efficiency. ChemBioChem 2017, 18, 1109–1116. 10.1002/cbic.201700039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sundlov J. A.; Fontaine D. M.; Southworth T. L.; Branchini B. R.; Gulick A. M. Crystal structure of firefly luciferase in a second catalytic conformation supports a domain alternation mechanism. Biochemistry 2012, 51, 6493–6495. 10.1021/bi300934s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harwood K. R.; Mofford D. M.; Reddy G. R.; Miller S. C. Identification of mutant firefly luciferases that efficiently utilize aminoluciferins. Chem. Biol. 2011, 18, 1649–1657. 10.1016/j.chembiol.2011.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peacor B. C.; Ramsay C. M.; Waters M. L. Fluorogenic sensor platform for the histone code using receptors from dynamic combinatorial libraries. Chem. Sci. 2017, 8, 1422–1428. 10.1039/C6SC03003C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solov'ev K. N.; Borisevich E. A. Intramolecular heavy-atom effect in the photophysics of organic molecules. Phys.-Usp. 2005, 48, 231–253. 10.1070/PU2005v048n03ABEH001761. [DOI] [Google Scholar]
- Branchini B. R.; Southworth T. L.; Murtiashaw M. H.; Boije H.; Fleet S. E. A mutagenesis study of the putative luciferin binding site residues of firefly luciferase. Biochemistry 2003, 42, 10429–10436. 10.1021/bi030099x. [DOI] [PubMed] [Google Scholar]
- Branchini B. R.; et al. Synergistic mutations produce blue-shifted bioluminescence in firefly luciferase. Biochemistry 2007, 46, 13847–13855. 10.1021/bi7015052. [DOI] [PubMed] [Google Scholar]
- Viviani V. R.; Prado R. A.; Neves D. R.; Kato D.; Barbosa J. A. A route from darkness to light: emergence and evolution of luciferase activity in AMP-CoA-ligases inferred from a mealworm luciferase-like enzyme. Biochemistry 2013, 52, 3963–3973. 10.1021/bi400141u. [DOI] [PubMed] [Google Scholar]
- Viviani V. R.; Amaral D. T.; Neves D. R.; Simões A.; Arnoldi F. G. C. The luciferin binding site residues C/T311 (S314) influence the bioluminescence color of beetle luciferases through main-chain interaction with oxyluciferin phenolate. Biochemistry 2013, 52, 19–27. 10.1021/bi300740y. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.