

Abstract
The nuclear receptor gene family includes 18 members that are broadly conserved among multiple disparate animal phyla, indicating that they trace their evolutionary origins to the time at which animal life arose. Typical nuclear receptors contain two major domains: a DNA-binding domain and a C-terminal domain that may bind a lipophilic hormone. Many of these nuclear receptors play varied roles in animal development, including coordination of life cycle events and cellular differentiation. The well-studied genetic model systems of Drosophila, C. elegans, and mouse permit an evaluation of the extent to which nuclear receptor function in development is conserved or exapted (repurposed) over animal evolution. While there are some specific examples of conserved functions and pathways, there are many clear examples of exaptation. Overall, the evolutionary theme of exaptation appears to be favored over strict functional conservation. Despite strong conservation of DNA-binding domain sequences and activity, the nuclear receptors prove to be highly-flexible regulators of animal development.
Keywords: development, C. elegans, Drosophila, mouse, functional conservation, exaptation
Among the earliest nuclear receptors (NRs) to be defined by mutation are androgen receptor in mice in 1970 [1], unc-55/COUP-TF in C. elegans in 1973 [2], knirps in Drosophila in 1980 [3], and daf-12 in C. elegans in 1981 [4]. While their molecular identity and characterization as NRs largely required the development of positional cloning strategies of the 1980’s, there is a long record of genetic analysis of NRs in model systems. Most of these early mutations were recovered from unbiased forward genetic screens for strong developmental phenotypes, highlighting the importance of NRs as regulators of ontogenetic mechanisms.
The typical nuclear receptor includes two major domains: a DNA-binding domain (DBD) that allows sequence-specific binding to DNA, and a ligand-binding domain (LBD) that binds a lipophilic hormone in some cases. Both ligand-dependent and ligand-independent NRs function as important regulators of transcription. They arose over 500 million years ago, apparently concomitant with the genesis of animal life. NRs are represented in all animal phyla, including Porifera, but are not found in plants or prokaryotes [5-7]. Thus the NRs are included in the basic genetic “toolkit” that animals adapted and refined over the course of evolution [8]. Several lines of evidence argue that lipophilic hormone binding activity—which makes them so interesting from the perspective of endocrinology and human health—is a derived property [6,9,10]. Indeed, some of the earliest NR genes defined by mutation produce proteins that are believed to be ligand-independent, such as unc-55 and knirps [2,3]. This suggests that the ancestral function of NRs was DNA-binding and transcriptional regulation.
In various phyla, NRs have proved to be a nimble and adaptable gene family. There are multiple examples of gene loss in specific animal clades, as well as massive gene proliferation and divergence in specific clades, such as the radiation of HNF4 NRs in C. elegans [6,11]. Many species have evolved atypical nuclear receptors, including some that lack a DBD or LBD, or have two DBD’s [5,12]. An example of this includes the 2DBD-NRs in the flatworm Schistosoma mansoni, which have two DBD’s in tandem with one LBD [12]. Some of the first genetically-defined NRs were discovered in the classic Drosophila patterning screens [13]. However, NRs show less functional conservation in development than some of the homeobox genes, which include conserved arbiters of axis positional information in vertebrates and invertebrates [13]. Despite the long history of research on NRs as regulators of developmental processes, the majority of published work has focused on vertebrate-specific steroid receptors [7]. In this review, we take a broad biological view of the genetic function of conserved NRs in development, with an eye towards patterns of exaptation (repurposing; [14]) and functional conservation across phylogeny. Because an understanding of developmental mechanisms benefits from analysis of organisms with robust genetic technologies, we will focus primarily on orthologs from the animal model systems Drosophila, C. elegans, and mouse.
Genome sequencing projects have allowed a fairly comprehensive analysis of the conservation and divergence of nuclear receptors among different phyla. The recent completion and annotation of mollusk, annelid, and platyhelminth genomes has provided additional clarity on patterns of conservation and orthology [12, 15,16]. Including lophotrochozoan NR proteins produces a phylogenetic analysis that is largely consistent with standard nomenclature for nuclear receptors (Figure 1). Each systematically-numbered NR class (e.g. NR2B, NR4A [6]) generally assembles nicely into a well-supported clade with orthologs in various species. Bootstrap replicates, as a test of the reliability of each clade, are strong for well-conserved nuclear receptors such as the various NR2 members (ranging from 58% support for NR2E1 to 93% support for the NR2F clade). In contrast, some NR1 clades assemble relatively poorly, with lower bootstrap support of reliability, consistent with the previously-noted sequence divergence of these NRs [6]. Short branch lengths illustrate the strong sequence conservation of DBD sequences over long evolutionary periods.
Figure 1. Molecular Phylogenetic Analysis of Nuclear Receptors.
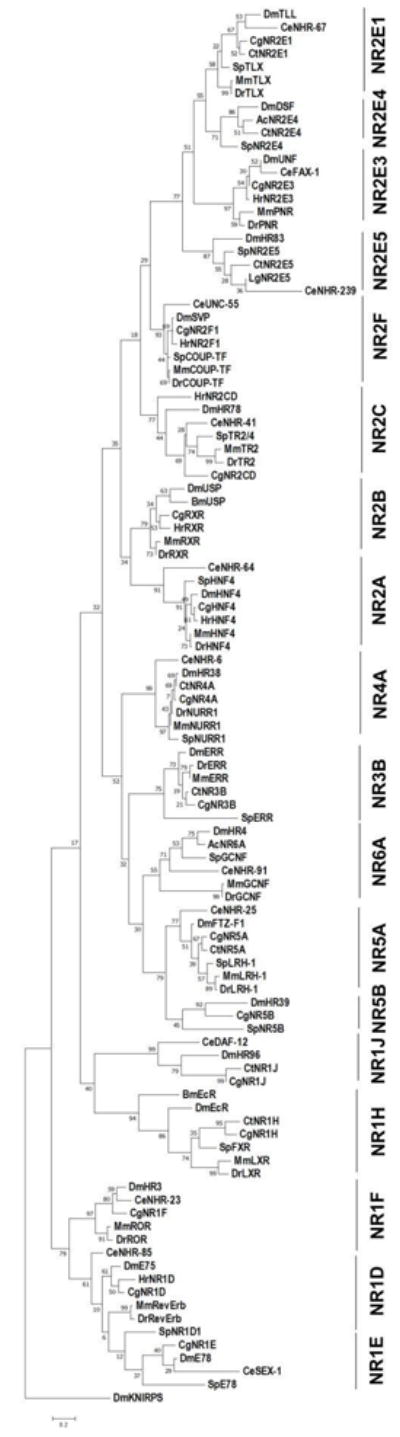
We generated a phylogenetic tree using the Maximum Likelihood method based on the Dayhoff matrix based model [266] from amino acid sequences of selected conserved NRs. Sequences were obtained from the NCBI database, aligned using Clustal W, and trimmed to include only the extended DBD for each sequence record (this is helpful when including C. elegans NRs, which have highly-diverged LBDs [11,185]). The tree with the highest log likelihood (-10026.43) is shown. The small number placed at each branch indicates the percentage of trees in which the associated taxa clustered together in 100 bootstrap replicates. Initial tree for the heuristic search was obtained by applying Neighbor-Joining and BioNJ algorithms to a matrix of pairwise distances estimated using a JTT model, and then selecting the topology with superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The tree was rooted to the atypical (NR0) nuclear receptor knirps as an outgroup. Analysis was conducted using MEGA7 [267]. For each class, a C. elegans (or Brugia malayi), Drosophila, Mus musculus, and Danio rerio ortholog was included (when present), organizing the analysis around two ecydysozoans and two vertebrates. Additional NRs were added from the complete genome of the echinoderm S. purpuratus, and two lophotrochozoan species. The lophotrochozoan species included varied due to the differences in completeness and annotation of these genomes, as well as a few possible cases of gene loss. Whenever possible, a representative annelid and mollusk was included. Species abbreviations: Dm Drosophila melanogaster, Ce Caenorhabditis elegans, Dr Danio rerio, Mm Mus musculus, Bm Brugia malayi, Sp Strongylocentrotus purparatis, Ac Aplysia californica, Cg Crassostrea gigas, Ct Capitella teleta, Hr Helobdella robusta, and Lg Lottia gigantea. This phylogenetic analysis establishes relationships among the major conserved nuclear receptor that is largely consistent with previously-published studies (e.g., [6]), but includes new data from the more recently-completed invertebrate genomes.
We identified 18 nuclear receptor genes that are represented in all three major phyletic groups (Ecdysozoa, Lophotrochozoa, and Deuterostomata) and therefore show clear ancient evolutionary origins: one each from NR class 1D, 1E, 1F, 1H, 1J, 2A, 2B, 2C, 2F, 3B, 4A, 5A, 5B, and 6A, and four from NR class 2E (Table 1; Figure 1). Eleven of these also have apparent orthologs in the platyhelminth S. mansoni [12], providing further evidence for their Urbilaterian origin. Four have been lost from Chordata, but are represented in Ecdysozoa and Echinodermata (NR1E/E78, NR2E4/DSF, NR2E5/HR83, and NR5B/HR39). Four additional nuclear receptors show more complicated patterns of conservation, being absent from Ecdysozoa, but present in Lophotrochozoa and Deuterostomata (NR1A/TRa, NR1B/RAR, NR1C/PPAR, and NR3A/ER). This could result from gene loss in early ecdysozoan evolution or the generation of novel NR genes in a common ancestor to lophotrochozoans and deuterostomes. The presence of an apparent NR1A/TR ortholog in S. mansoni argues for the former interpretation for that gene, although it is possible that this represents an example of horizontal gene transfer from chordates. Because these four nuclear receptor subfamilies are not represented in Ecdysozoa, they are not considered further. For the purposes of this review, we focus our analysis on the 18 nuclear receptors (shaded gray in Table 1) that display broad polyphyletic conservation and are thus represented in the classic developmental model system of Drosophila.
TABLE 1.
Conserved nuclear receptors
| Platyhelminthes | Ecdysozoa | Lophotrochozoa | Echinodermata | Chordata | ||
|---|---|---|---|---|---|---|
|
| ||||||
| Class | S. mansoni | D. melanogaster | C. elegans | C. gigas | S. purpuratus | H. sapiens |
|
| ||||||
| 1A | TRa/TRb | NR1A | THR | TRα/β1 | ||
|
| ||||||
| 1B | NR1B | RAR | RAR α/β/γ1 | |||
|
| ||||||
| 1C | NR1C/CDa/CDb | PPAR 1/2 | PPAR α/β/γ1 | |||
|
| ||||||
| 1D | E75 | NHR-85 | NR1D | RevErb | Rev-Erb α/β1 | |
|
| ||||||
| 1E | E78 | E78 | SEX-1 | NR1E | E78 a/b | |
|
| ||||||
| 1F | HR31 | NHR-23 | NR1F | ROR α/β/γ1 | ||
|
| ||||||
| 1H | EcR1 | (Brugia EcR) | NR1H | FXR | LXR α/β1 | |
| FXR1 | ||||||
|
| ||||||
| 1J | HR96 | HR96 | DAF-121 | NR1J a/b/g | PXR1 | |
| NHR-8/-48 | VDR1 | |||||
| CAR1 | ||||||
|
| ||||||
| 2A | HNF4 | HNF4 | superfamily | NR2A | HNF4 | HNF4 α/γ1 |
|
| ||||||
| 2B | RXR1 | USP | (Brugia USP) | NR2B | RXR | RXR α/β/γ1 |
| RXR2 | ||||||
|
| ||||||
| 2C | TR2/4 | HR78 | NHR-41 | NR2CD | NR2C | TR2 |
| TR2/4 | TR4 | |||||
|
| ||||||
| 2E | TLL | TLL | NHR-67 | NR2E1 | TLX | TLX |
|
| ||||||
| 2E | DSF | DSF | NR2E4 | NR2E4 | ||
|
| ||||||
| 2E | PNR | UNF | FAX-1 | NR2E3 | PNR | PNR |
|
| ||||||
| 2E | HR83 | NHR-239 | NR2E5 | NR2E5 | ||
|
| ||||||
| 2F | COUP-TFI | SVP | UNC-55 | NR2F | COUP-TF | COUP-TF α/β |
| COUP-TFII | EAR2 | |||||
|
| ||||||
| 3A | NR3A | ER α/β1 | ||||
|
| ||||||
| 3B | ERR | NR3B | ERR | ERR α/β/γ | ||
|
| ||||||
| 4A | NR4A5 | HR38 | NHR-6 | NR4A | NURR | Nur77 |
| Nurr1 | ||||||
| NOR1 | ||||||
|
| ||||||
| 5A | FTZ-F1a | FTZ-F11 | NHR-25/-97/-119 | NR5A | LRH-1 | SF-11 |
| LRH-11 | ||||||
|
| ||||||
| 5B | FTZ-F1b | HR391 | NR5B | NR5B | ||
|
| ||||||
| 6A | HR41 | NHR-91 | (Aplysia NR6A) | GCNF | GCNF | |
| GRF | ||||||
NR with well-defined ligand.
Shaded rows identify the 18 NRs with broad phylogenetic conservation discussed in this review.
E75/NHR-85/Rev-Erb (NR1D)
The Drosophila E75 gene functions in molting, metamorphosis, oogenesis, and circadian rhythms [17,18]. It is essential for larval development, responding to ecdysone regulatory pathways at the onset of metamorphosis [19]. E75 is required for ecdysteroid biosynthesis in the prothoracic gland, and it counteracts DHR3 to avoid the premature repression of steroidogenesis [20]. Germline clones of E75 mutations cause germline arrest prior to meiosis entry, and degeneration of oocytes with follicle cell migration defects [21,22]. In Drosophila males, E75 promotes stem cell maintenance in the testis, and knockdown causes a loss of germ stem cells and cyst stem cells [23]. The final known role of E75 in Drosophila is similar to that of its mammalian ortholog, Rev-Erb : E75 functions in the fly molecular clock and circadian rhythm in clock and pacemaker neurons [24]. E75 and unf (see below) act together to enhance transcription of per, which finishes the transcriptional feedback loop needed for rhythmic cycles of gene expression within cells [19].
The nhr-85 gene in C. elegans is less well-characterized. RNAi knockdown experiments indicate that nhr-85 plays a role in the development of the egg laying system, vulval development, and formation of a proper dauer larva cuticle [25]. Like its Drosophila homolog E75, nhr-85 functions in both cuticular development and female organogenesis. The gene appears to be upregulated at larval molts. C. elegans molts punctuate periods of quiescent inactivity that represent a sleep-like state [26]. Therefore, upregulation of nhr-85 in anticipation of quiescent periods may be functionally analogous to circadian function in flies.
The vertebrate homologs, Rev-Erbα (NR1D1) and Rev-Erbβ (NR1D2), play a significant role in circadian rhythms and metabolism [27,28]. Rev-Erb paralogs are expressed in neurons, the thymus, skeletal muscle, and kidney [29]. Mutant mice lacking both Rev-Erb paralogs in the liver have arrhythmic expression of the clock genes Bmal1 and Cry1 [30], reminiscent of E75 function in the fly. In the zebrafish Danio rerio, Rev-Erbα is similarly important for circadian control of autophagy rhythms [31,32]. A double knockout of both Rev-Erb genes in mice yields animals that are fairly normal and viable, suggesting that the genes do not play a major role in development. However, cerebellar defects have been described in Rev-Erbα knockout mice, leaving open the possibility of a developmental function [33]. Neither Rev-Erb gene is known to play a role in reproductive development.
In summary, the NR1D gene family appears to have a broadly conserved function in controlling cellular rhythms across animal phyla, particularly in control of arousal. Developmental functions are evident in both flies and worms; however conservation of the specific functions is ambiguous at best. These functions may represent separate examples of exaptation.
E78/SEX-1 (NR1E)
In Drosophila, E78 functions in female reproductive development and is ecdysone-induced. Loss of E78 results in decreased numbers of cap cells, germline stem cells, and follicle cells [34,35]. As a consequence E78 mutants lay few eggs and display early onset female sterility [34]. Embryos from E78 mothers fail to complete gastrulation. Finally, like E75, Drosophila E78 is required for ecdysone regulatory pathways that control the onset of metamorphosis [19].
The C. elegans homolog of Drosophila E78 is sex-1, which plays a role in the regulation of nematode sex [36,37]. The xol-1 gene specifies male development, and sex-1 is responsible for repressing xol-1 transcription, leading to hermaphrodite sexual development. Mutations in sex-1 cause death from multiple developmental defects due to inappropriate dosage compensation [36].
While both nuclear receptor genes function in sex-specific development, the roles of these genes differ across species. E78 functions in Drosophila female reproductive development, whereas sex-1 plays an early key role in global sex determination of C. elegans in multiple tissues. E78 also has an early embryonic function unrelated to sex-specific development, which has not been described for sex-1. Thus even though there are hints at some common, conserved function, this class also illustrates the theme of exaptation. Although bona fide E78 orthologs appear to be present in the platyhelminth S. mansoni, lophotrochozoans, and echinoderms, the gene appears to have been lost in vertebrate lineages (Table 1).
HR3/NHR-23/ROR (NR1F)
The Drosophila HR3 gene was identified by molecular strategies on the basis of sequence relationship to E75 [38]. It is ecdysone-responsive and initially serves to repress other ecdysone-responsive genes that trigger metamorphosis and then induces expression of FTZ-F1 as part of the prepupal response to ecdysone [39,40]. Deletion of HR3 results in embryonic lethality with severe ventral nerve cord and tracheal defects, indicating that it plays a central role in development [41,42]. Mutants that are permitted to reach larval stages expire at morphogenesis with multiple defects in wing, bristle, and cuticle formation [39]. Therefore, HR3 is an important developmental control gene with various roles in patterning embryos as well as temporal coordination of life cycle transitions.
The C. elegans HR3 ortholog, nhr-23, plays an apparently similar role in coordinating molting and developmental transitions. While embryonic functions appear likely, they have not been evaluated in detail. Mutations in nhr-23 cause mid-larval arrest with cuticle defects, demonstrating a critical function in hypodermal development and cuticle formation [43,44]. Expression of the gene is sharply up-regulated with larval molts [25], and it is a target of the heterochronic miRNA let-7 that regulates transitions between larval stages [45].
The three RAR-related orphan receptors (ROR; NR1F1, NR1F2, NR1F3) of vertebrates were first identified by molecular strategies in screens for new nuclear receptors [46-49]. Mutations in RORα were found to be the cause of the staggerer mutation in mice [50]. These mice present with severe cerebellar dysfunction and altered gait, due to an underlying defect in proliferation and differentiation of Purkinje cells [50, 51]. Like its invertebrate orthologs, RORα is a critical regulator of multiple developmental events [52-54]. Metabolic and immunological functions have been described [53,54], as well as a role in circadian gene regulatory patterns [55]. The RORβ gene is expressed in a circadian pattern in the eye and pineal gland. Deletion of RORβ in mice results in altered gait, suggestive of underlying cerebellar dysfunction, disorganized retinas with defective rod photoreceptors, and progressive blindness [56-57]. Recent studies have suggested a broader role in cerebral neuron development, with loss of RORβ associated with epilepsy and intellectual disability in humans [58]. Deletion of the third paralog, RORγ, results in defects in the production of thymic cells and lymph nodes, due to a critical role for this nuclear receptor in maintaining these cells and preventing apoptosis [59]. Mutations in the human RORγ gene cause immunodeficiency [60].
The common function in life cycle transition in both Drosophila and C. elegans illustrates some level of functional conservation for the NR1F class. While not exactly the same, the vertebrate functions in circadian rhythms are at least reminiscent of the broad, temporal functions of the gene class in Ecdysozoa. The repurposing of vertebrate genes to immune system development is a clear case of exaptation.
EcR/LXR/FXR (NR1H)
The steroid hormone 20-hydroxyecdysone is produced in the prothoracic glands and serves as the primary endocrine signaling system that coordinates life cycle transitions in the fly [61]. Ecydsone receptor (EcR) mediates the hormonal response in various tissues [62] via heterodimers with the coordinating nuclear receptor usp (NR2B; described below). EcR functions in germ band retraction and head involution during morphogenesis of the embryo [63]. EcR and usp play a central role in late third instar larvae in responding to the ecdysone pulse that triggers pupation and metamorphosis. In mutant larvae, imaginal discs fail to expand and differentiate, the larval gut and salivary glands fail to undergo apoptosis, and the adult gut does not form. In the developing wing, sensory neurons of usp and EcR mutant larvae undergo precocious differentiation [64]. EcR and usp also function in the development of male-specific neuronal architecture and male-mating behaviors [65]. More recently, usp and EcR have been shown to be required for the survival of prothoracic gland cells during metamorphosis via an ecdysone-independent pathway [66].
Surprisingly, C. elegans lacks both an EcR and an usp ortholog [25]. Despite ecdysis as a central property of Ecdysozoa, Caenorhabditis appears to have dispatched this pathway entirely. Nonetheless, several nuclear receptors, in particular nhr-23 and nhr-25, perform molting regulatory functions in C. elegans, providing an example of the flexibility of nuclear receptors, even in broad physiological and developmental control. It is important to note that this is not a feature of nematodes in general, since the parasitic nematode Brugia malayi and free-living nematode Pristionchus pacificus both retain EcR, RXR, and ecdysteroid-based regulation [67,68]. Instead, this unexpected gene loss appears to be a peculiarity of an evolutionary lineage within the Rhabdita order.
Two vertebrate orthologs of EcR, LXRα and LXRβ (Liver X Receptor; NR1H3 and NR1H2, respectively), were initially identified in several labs using molecular techniques [69,70]. Just like EcR and USP, both LXRs heterodimerize with RXR partners (NR2B; see below) and are responsive to oxysterol ligands [71]. Mice deleted for LXRα display defects in cholesterol metabolism and inflammatory responses, but were born viable and healthy, suggesting that developmental functions are few or redundant [69]. LXRβ mutations lead to adult-onset motor neuron degeneration in male mice [72], inflammatory response defects, and hyperplasia of T-cells [73]. These effects appear to be secondary to cholesterol metabolism defects rather than direct functions in development. Loss of both LXRα and LXRβ results in susceptibility to bacterial infection due to apoptosis of macrophages, indicating that LXR function is required for macrophage survival, but not generation [74]. A third vertebrate-specific paralog of the LXR’s is FXR (Farnesoid X Receptor; NR1H4). This gene was identified by molecular strategies from a rat liver cDNA library and shown to bind bile acids [75]. Knockout of the gene in mice results in animals that are apparently developmentally normal, but display elevated bile acid levels and metabolic defects [76]. Therefore, the broad developmental regulatory functions evident in Drosophila appear to have been dropped in vertebrate lineages.
HR96/DAF-12/VDR/PXR/CAR (NR1IJ)
The HR96 gene of Drosophila was first identified by molecular strategies [19]. Mutations in the HR96 gene cause defects in triacyl glycerol and cholesterol metabolism, as well as increased sensitivity to xenobiotics such as phenobarbital and the insecticide DDT [77-81]. Research on HR96 has served as a model to understand cholesterol metabolism and detoxification [82,83]. No significant role in regulating developmental events is evident.
The daf-12 nuclear receptor is the most extensively-studied NR in C. elegans due to its pivotal role in regulating response to TGFβ and insulin signaling in the dauer developmental pathway and regulation of lifespan [84]. The dauer is a long-lived, highly-resistant dispersal larva form of the nematode that is an alternative to the normal L3 larval stage [85]. Mutations in daf-12 were first identified in screens for defective formation of dauer larvae [4]. Subsequent research showed that daf-12 encoded a Vitamin D Receptor- related gene that binds steroid hormones called dafachronic acids [86,87]. The gene broadly regulates temporal developmental and metabolic processes in the worm [84]. In the presence of food and lower populations of worms, the enzymatic product of the daf-9 gene produces dafachronic acids that activate the DAF-12 nuclear receptor, prompting it to activate transcription of target genes that favor normal reproductive development. When food becomes scarce and population conditions more crowded, DAF-12 binds the co-repressor DIN-1, shutting off its normal target genes and promoting a developmental switch to dauer larvae. The daf-12 gene also regulates heterochronic miRNA genes that are responsible for the timing of earlier developmental transitions [88], thus serving multiple roles in coordinating development in multiple tissues. Interestingly, the functions of daf-12 appear to be more conserved with EcR than its apparent fly and vertebrate orthologs, all of which function in lipid metabolism. In both flies and insects, insulin signaling and TGFβ signaling converge with the production of a lipophilic hormone that binds EcR (in the fly) or DAF-12 (in the worm), both of which regulate the let-7 miRNA that in turn mediates major life cycle transitions [89,90]. Interestingly, this pathway may also have a vertebrate analog, whereby estrogen signaling activates the transition to puberty via Estrogen Receptor. Another daf-12-like paralog in C. elegans, nhr-8, is expressed in the gut where it regulates cholesterol balance, fatty acid desaturation, apolipoprotein production, and bile acid metabolism, similar to the function of HR96 in flies, suggesting that duplication of this gene class in nematodes may have allowed segregation of functions [91]. An additional paralog, nhr-48, has no obvious phenotype when deleted and has not been extensively characterized.
Vertebrates possess three genes with apparent relationship to the HR96/daf-12 class: Vitamin D Receptor (VDR; NR1I1), Pregnane X Receptor (PXR; NR1I2), and Constitutive Androstane Receptor (CAR; NR1I3/4). All three were initially identified using molecular approaches [92-94]. Mouse knockouts of VDR are born overtly normal, but die within 15 weeks of birth with significant deficits associated with Vitamin-D deficiency, such as rickets [95]. In addition, female animals suffer from uterine and follicle developmental defects, which are not observed with Vitamin-D deficiency, raising the possibility for non-Vitamin-D dependent developmental functions for VDR. Subsequent work showed that some bone defects were caused by a failure of apoptosis in proliferating chondrocytes [96], and that natural killer T cells also depend on VDR for their proper development [97].
The PXR gene (aka SXR in humans) activates cytochrome P450-3A expression in response to xenobiotics and plays a critical role in mediating drug responses [98-100]. Similarly, the CAR gene functions in xenobiotic response by upregulating the cytochrome P-450 Cyp2b10 gene [101-103]. Mice lacking the CAR gene are developmentally normal and fertile [102].
The functions of PXR and CAR appear to conserve the function of Drosophila HR96 in at least some respects, and the NR1I and NR1J subfamilies seem to play broadly-conserved roles in metabolic regulation. The developmental function of daf-12 in C. elegans appears to be a distinct exaptation, although it is worth noting that dauer formation is a developmental event that is intimately linked to metabolic regulation.
HNF4 (NR2A)
The HNF4 gene is well-conserved across animal evolution. The Drosophila HNF4 gene was identified by molecular techniques on the basis of its similarity to the vertebrate gene [104]. Early analysis of expression patterns and a chromosomal deletion suggested that HNF4 might play a role in fly gastrulation and gut development as in vertebrates [104]. However, HNF4 null mutants hatch normally, indicating that the gene is not essential for gastrulation. Mutant larvae display metabolic defects in utilizing stored fat during larval life before arresting development either just before or after pupal eclosion [105]. Thus fly HNF4 appears to have a major role in regulating metabolism, but a less well-defined role in development.
The HNF4 class of nuclear receptor has undergone a remarkable expansion in C. elegans and related nematodes to generate hundreds of distinct family members (269 distinct paralogs in C. elegans [106]). While most of these nuclear receptors have not yet been functionally-characterized, of the dozen or so that have been studied most play some role in regulating metabolism, similar to flies and vertebrates [90]. This includes the well-characterized nuclear receptors nhr-13, -49, -63, -64, -66, -69, -76, and -80. In particular, the function of nhr-49 in regulating fat metabolism has striking parallels to the function of Drosophila HNF4 in the same process [107,108]. One possibility is that multiple metabolic functions of fly and vertebrate HNF4 genes have been distributed to various individual C. elegans HNF4 family members [109]. Some C. elegans HNF4 nuclear receptors have taken on key functions in development as well. The nhr-31 gene functions in the growth of the excretory tube [110], nhr-114 functions in somatic tissue to regulate germ-line proliferation in response to diet [111], nhr-40 functions in embryonic elongation and muscle development [112], and nhr-60 functions in hypodermal cell differentiation [113]. The HNF4 class has apparently expanded functionally to incorporate a range of physiological and developmental functions.
The first HNF4 ortholog was identified in vertebrates from liver extracts using molecular approaches [114]. Mice and humans have two paralogs, HNF4α and HNF4γ (NR2A1; NR2A3). Deletion of the HNF4α results in failure of late stages in gastrulation, indicating that the gene plays a central role in the development of the gut [115-117]. Analysis of gene function identified an important role in insulin production in the pancreas [118], and in hepatocyte differentiation [119]. Therefore, metabolic defects in HNF4 mutants may reflect secondary consequences of underlying developmental defects.
A recent study has also implicated HNF4α in the development of neural stem cells, hinting at broader developmental functions for the gene class in vertebrates [120]. The HNF4γ paralog has not been genetically-characterized, but expression analysis indicates that it could play roles in multiple organs [121]. A third paralog (HNF4β/NR2A2) has been identified in Xenopus [122].
Overall, functions of the HNF4 class have concentrated largely on metabolic regulation in Ecdysozoa, but this is not a conserved function in vertebrates. Given the large number of HNF4-related genes in C. elegans, it is difficult to make direct comparisons, but at least some have functions in the differentiation of cells and organogenesis. However, none of the functions identified so far align well with known vertebrate functions in development.
USP/RXR (NR2B)
The ultraspiracle gene of Drosophila was first identified as a recessive embryonic lethal mutation [123]. Subsequent molecular work identified it as an RXR ortholog that served as a heterodimerization partner for EcR, and thus usp is a critical mediator of ecdysone signaling in flies [124,125]. The major functions of usp are largely covered above under the functions for its partner, EcR. However, usp and EcR mutants have somewhat different embryonic phenotypes, suggesting that EcR has additional functions that do not depend on usp [123]. Nonetheless, as is the case for EcR, usp function is required in multiple tissues for cell survival and differentiation, consistent with its role as the cofactor for a global integrated hormonal response. As described above, C. elegans lacks an NR2B ortholog, just as it lacks an EcR ortholog.
The retinoid-binding RXR receptors were first identified in 1989 on the basis of molecular properties in vertebrates [126-128]. The three paralogs (RXRα, RXRβ, RXRγ; aka NR2B1, NR2B2, NR2B3) serve as key heterodimer partners for various NR1 nuclear receptors, including retinoic acid receptors, thyroid hormone receptors, PPARs, and LXRs, the vertebrate ortholog of fly usp partner EcR, demonstrating both conservation and flexible elaboration of their functional properties over evolution [7]. Knockout of RXRα in mice results in embryonic arrest with severe hypoplasia of the heart and eye malformations, identifying this receptor as a key regulator of retinoid signaling in cardiac and ocular organogenesis [129,130]. Knockout of RXRβ results in some embryonic lethality without a clear cause. Surviving animals are mostly normal, but males suffer from defects in spermatogenesis due to abnormalities in Sertoli cells [131]. Similarly, mice lacking the RXRγ gene are viable, normal in appearance, and fertile [132]. This is not a simple matter of redundancy among the RXR paralogs, since RXRα+/- RXRβ −/− RXRγ −/− triple mutants are also viable and crudely normal. More detailed analysis has revealed that RXRβ and RXRγ do play a role in neuron development or physiology, since knockout mice suffer from memory and affective behavior deficits [133,134]. Studies have also identified roles for RXR receptors in cyclic metabolic functions in the liver and vasculature [135,136], suggesting some conservation of global temporal regulation reminiscent to usp function in the fly. Nonetheless, despite the apparent conservation of heterodimerization properties, this subfamily also demonstrates significant functional elaboration across animal phyla.
HR78/NHR-41/TR2/TR4 (NR2CD)
The Drosophila HR78 gene was identified by molecular approaches as a nuclear receptor expressed during the onset of metamorphosis [19]. Its expression is induced by ecdysone and oscillates with the molting cycle, but it functions as a negative regulator of EcR/USP activity. Loss of HR78 results in arrest as undersized third instar larvae that fail to execute ecdysteroid-triggered events and have tracheal defects [137]. HR78 coordinates with the SAM domain co-repressor Moses to help regulate cell proliferation, with HR78 activity favoring and Moses activity inhibiting cell division [138]. The HR78 pro-proliferation function in the fly is somewhat reminiscent of vertebrate TR2/TR4 function in promoting cell proliferation in the mouse.
The C. elegans genome project identified nhr-41 as the nematode NR2CD ortholog [25]. Like HR78, nhr-41 is broadly expressed and oscillates with the molting cycle. Knockdown of nhr-41 results in SDS-sensitive dauer larvae, suggesting a defect in the dauer cuticle [25]. A probable null deletion of nhr-41 is viable, suggesting that nhr-41 plays a modest or partially-redundant developmental function in C. elegans.
The TR2 gene (NR2C1) of vertebrates was the founding member of this gene class, discovered by low-stringency hybridization from a human testis cDNA library [139]. Deletion of TR2 does not result in testicular or spermatogenesis defects— homozygous null mutant mice are viable and fertile [140]. A recent analysis has demonstrated that it regulates early transcription factors that coordinate the development of retinal progenitor cells and knockout mice have severe vision deficits [141]. In contrast, knockout of the TR4 paralog (NR2C2) has dramatic effects on mouse viability, with homozygotes dying either during fetal development or postnatally, with significant compromises in growth and reproductive behavior [142]. Mutant mice also display severe motor deficits associated with aberrant cerebellar development, establishing a major role for TR4 in neuronal development [143]. TR2 and TR4 appear to play partially-redundant functions in stem cell proliferation. Mice with both genes knocked out die during embryogenesis with increased apoptosis [144]. Expression of Oct-3/4 and Nanog depend on TR2/TR4, suggesting that TR2 and TR4 function in embryonic stem cell self-renewal and differentiation. Because of their potential as targets for anti-cancer therapies, interest in TR2 and TR4 remains high [145].
The NR2C subfamily illustrates some apparent conservation of NR function. In both flies and vertebrates, family members regulate cell proliferation events and play critical roles in development. In both flies and nematodes, they have regulatory patterns and functions associated with the molting cycle. However, the function in regulation of cell proliferation does not seem to have been maintained in C. elegans.
TLL/NHR-67/TLX (NR2E1)
The tailless (tll) gene was originally identified by a traditional forward genetic screen as a terminal/gap gene in Drosophila [146,147]. Deficiency in tll results in a failure to develop posterior body segments as well as parts of the brain, thus resulting in embryonic lethality. tailless has been shown to interact dynamically with a number of other gap genes including Krüpple, knirps, and hunchback [148]. Loss of function in tll decreases neuroblast formation in the protocerebrum and no boundary is formed between the protocerebrum and the deutocerebrum [148,149]. The tll gene also plays a role in eye development, in both the eye-precursor cells and the embryonic optic lobe [148]. These important roles in neural identity and boundary formation appear to be well-conserved in other animals. Loss of tll in the adult fly pars intercerebralis leads to a loss of neuropeptide expression and increased aggression, although the specific developmental effect in this part of the brain has not been described [150].
Identified by the C. elegans genome sequencing project, nhr-67 is involved in a number of developmental processes including neural development, vulval and uterine development, and cell migration. RNAi knockdown of nhr-67 demonstrated a function in cuticle shedding, egg-laying and vulval development [25]. nhr-67 is a component of a transcription factor circuit in neural differentiation in the ASEL and ASER sensory neurons [151]. During the L2 and L3 stages, nhr-67 functions in the differentiation of pre-VU and anchor cells of the ventral uterus [152]. Defective anchor cells fail to invade the underlying vulval tissue and remain in the cell cycle, generating additional cells [153]. Later in larval development, it plays a role in vulval morphogenesis [154] and in the male it regulates the timing of the migration of the linker cell, which is the male equivalent of the anchor cell [155]. Loss of nhr-67 results in embryonic and L1 lethality [152]. In the embryo, nhr-67 is expressed in the posterior-most daughters of multiple lineages—reminiscent of the function of tll in Drosophila embryonic termini— and appears to be responsive to Wnt regulation [156].
The vertebrate ortholog Tlx (NR2E1) was first cloned from a chick cDNA library screen [157]. Tlx is expressed in the developing central nervous system— the telencephalon, diencephalon, eye and nasal placode [157,158). Unlike its Drosophila and C. elegans orthologs, mutations in mouse Tlx are not lethal. Mutant mice display varying levels of anatomical and behavioral abnormalities, including patterning defects in the brain, particularly at the pallio-subpallio boundary [159]. Due to the limbic system’s development from these early structures, adult knockout mice show significant behavioral changes, most notably high levels of aggression and poor maternal instincts [160]. This may reflect a conserved function, since the fly pars intercerebralis is thought to be analogous to the vertebrate hypothalamus, and the same cofactor, Atrophin, participates in aggression in both flies and mice [150,161]. A variety of retinopathies also result from Tlx mutation, indicating that it plays a critical role in the development and maintenance of retinal cells [162]. Tlx plays a role in neural stem cell self-renewal and adult neurogenesis [163], where it functions upstream of a Wnt signaling pathway [164]. In mutant Tlx mice, neural stem cells undergo precocious maturation to neurons and are therefore quickly depleted, resulting in mice with smaller gyrus and forebrain regions [165,166]. This premature cell cycle exit phenotype appears opposite to the cell cycle phenotype in the C. elegans anchor cell, but suggests that Tlx might play a conserved role in regulating cell cycle decisions associated with differentiation.
In summary, functions in neuron development, optic development, and cell cycle control appear to be conserved among NR2E1 gene family members. Interestingly, the namesake phenotype “tailless” in flies appears to reflect an exapted function, since studies in C. elegans and vertebrates have failed to demonstrate a similar broad patterning function.
UNF/FAX-1/PNR (NR2E3)
The unfulfilled (HR51) gene was named on the basis of the mutant phenotype in which eclosed adults fail to unfold their wings and exhibit reduced fertility [167,168]. In mushroom bodies, it functions in specifying neuron identities and regulating axon pathfinding [169]. More recently, unf has been implicated in circadian rhythms in Drosophila by functioning in the identity of small ventral Lateral Neurons as the master pacemaker [170]. In late stage pupae, unf accumulates in the Mushroom Body of the brain and in some lateral neurons near the optic lobes. The MB γ-neurons normally undergo pruning and axon regrowth to form adult-specific neural connections via the TOR pathway, but axons fail to regrow following pruning in unf mutants [171]. This function depends on unf/E75 heterodimers and nitric oxide signaling [172].
The C. elegans ortholog, fax-1, functions in axon pathfinding in the nerve ring and ventral nerve cord [173]. Mutations in fax-1 cause loss of expression of neuropeptides, neurotransmitter receptors, and other markers of interneuron identity [174,175]. Similar to the defective axon regrowth phenotype noted in unf mutants, fax-1 mutants display reduced axon regeneration of an interneuron following laser damage [171]. Thus a conserved function in specifying neuronal identity and axonal growth appears to be a feature of this gene.
The photoreceptor-specific nuclear receptor (PNR; NR2E3) was identified by molecular techniques for genes related to Tlx [176]. PNR is an extreme example of a spatially restricted nuclear receptor, being expressed exclusively in the retina and pineal gland. In rod photoreceptors, PNR plays an important role in cell fate by repressing cone-specific genes [177,178]. Further, deletions in PNR in mice result in retinal degeneration consistent with flecked retinal degeneration in humans [179]. PNR has been shown to interact with Rev-Erb, probably as a heterodimer [172]. Interestingly, PNR has been shown to exhibit circadian regulatory patterns in zebrafish, suggesting an evolutionarily-conserved PNR/Rev-Erb regulatory module associated with neuron differentiation and rhythmic patterns of gene expression [180]. Mutations in human PNR result in inherited retinal degeneration diseases [181].
NR2E3 family members all seem to play roles in neuronal differentiation—and the common function in axonal arborization points to some more specific conservation of function between Drosophila and C. elegans. The function in photoreceptor development in vertebrates appears to be another example of exaptation, although in this case perhaps applying a conserved function in neuron differentiation to a new neuron type.
DSF (NR2E4) and HR83/NHR-239 (NR2E5)
There exist two additional gene classes in the NR2E family, but both appear to have been lost from vertebrates. dissatisfaction was identified as a mutation that affects sex-specific courtship and neural differentiation in both sexes of Drosophila, independent of the doublesex pathway [182,183]. Homozygous dsf females show resistance to copulation and fail to lay eggs when they do mate [183]. These phenotypes may be explained by the lack of synapses present in the circumferential muscles of the uterus in mutant dsf females. Males also display striking courtship behavior phenotypes, including increased male-male courting, delayed copulation, and defects in abdominal curling during the final stage of courtship. These phenotypes may be explained by the irregular innervation of the ventral longitudinal muscles of abdominal segment 5, particularly the presence of a few large, spherical boutons on each fiber. Thus dsf plays important roles in nervous system development. Once thought to be an arthropod innovation, dsf orthologs are now apparent in the genomes of lophotrochozoans and echinoderms.
Still another NR2E gene family member, HR83/nhr-239, has orthologs in some insects (HR83), C. elegans (nhr-239), lophotrochozoans, an echinoderm, and a hemichordate, but has not been identified in vertebrates [184]. There is considerable divergence in the LBD of the nhr-239 clade, and some species-specific orthologs have truncated LBDs. In C. elegans, nhr-239 is expressed at low levels in a few neurons. Mutations in HR83 are homozygous viable in the fly, but expression and function have not yet been described.
SVP/UNC-55/COUP-TF (NR2F)
The NR2F class of nuclear receptors is well conserved and has been extensively studied. Drosophila seven-up (svp) functions in a diverse set of cell specification and transcriptional regulation roles. svp was identified in an enhancer trap screen [185]. Loss of svp results in transformation of R1, R3, R4, and R6 photoreceptors of the compound eye into R7 photoreceptors. Over expression results in a loss of R7 and R8, indicating that it plays a sufficient role in sensory neuron specification [186]. The gene is also required for specification and development of fat body cells [187], cell proliferation in Malpighian tubule development [188], and proper cardioblast diversification by repressing tinman in a subset of cells [189]. svp also inhibits ecdysone mediated pathways by forming inactive heterodimers with EcR, thus inhibiting transcription mediated by usp/EcR dimers [190].
The C. elegans ortholog, unc-55, plays a role in synaptic patterning and male mating circuits [191,192]. The VD motor neurons of unc-55 mutants are transformed to resemble the afferent and efferent synaptic patterning of DD motor neurons. Therefore, unc-55 functions in specifying neuronal identities, similar to the role of its Drosophila ortholog.
The vertebrate orthologs, chicken ovalbumin upstream promoter-transcription-factors (COUP-TFI and COUP-TFII; NR2F1, NR2F2), play a significant role in epithelial differentiation in embryogenesis. COUP-TFI was discovered as a transcription factor that regulates expression of ovalbumin [193,194]. Human COUP-TFII was identified by its homology to COUP-TFI [195]. The COUP-TFs have overlapping but unique expression patterns. COUP-TFI has high expression in the nervous system and many developing tissues [195]. In particular, COUP-TFI is highly expressed in organs that develop by epithelial proliferation and differentiation, including those that develop by mesenchymal differentiation to epithelium. Mouse COUP-TFI mutants show dose-dependent embryonic lethality or lethality shortly after weaning, reduced growth of the head and heart, severe hemorrhage, edema, enlarged blood vessels, defective angiogenesis, ocular defects, bone defects, and vascular remodeling [194]. COUP-TFI mutants are also missing cortical layer IV in the cerebral cortex lamina, due to a failure of axons to extend to their cortical targets, and resulting excessive cell death [196]—thus a role in neuronal development appears to be evolutionarily-conserved. COUP-TFII mutants have low Angiopietin-1 (Ang-1) levels, which may account for heart and vasculature defects. Defects in the glossopharyngeal nerve result in poor sensation and motor function of the tongue and pharynx, contributing to malnutrition and perinatal death.
The comparison of functions of NR2F family members suggests themes of both exaptation and conservation. A general role in nervous system development is evident among species investigated in detail, but various different roles in organogenesis are apparent in flies and vertebrates.
ERR (NR3B)
The estrogen-related receptor (ERR) gene of Drosophila, although evolutionarily-related to steroid receptors of great interest to human health, has not been fully characterized. This gene class has been lost in nematodes, but is retained in vertebrates (Fig. 1, Table 1). Mutations in ERR are lethal in larval development, indicating that the gene plays an essential function. A recent study showed that ERR is required for normal spermatogenesis in the fly [197], but the essential function remains undescribed. ERR orthologs are present in Lophotrochozoa and Echinodermata, but were apparently lost from nematode lineages.
Vertebrates have three ERR orthologs (α/β/γ; NR3B1,NR3B2,NR3B3), none of which are known to bind a ligand. Knockouts of ERRα in mice are viable and fertile, displaying only modest metabolic defects [198]. The ERRβ paralog is required for normal placental development and function; gene knockout results in embryonic lethality [199]. Knockouts of the ERRγ ortholog present with significant physiological and development defects and result in death in a few weeks [200]. These defects included abnormal heart and spinal cord function [200,201] and developmental defects in early branching events of the ureteric duct of the kidney [202]. A recent study also described defects in hippocampal neuron physiology and function, but without clear developmental involvement [203]. Further evidence may clarify a role for ERR orthologs in development, but current studies have revealed scant evidence for conserved functions in ontogeny.
HR38/NHR-6/NUR77/NURR1/NOR1 (NR4A)
Drosophila HR38 was identified by molecular cloning as a nuclear receptor with expression associated with morphogenesis [19]. It is broadly expressed, dimerizes with USP/RXR, and functions downstream of ecdysteroids in a pathway that does not depend on EcR [204,205]. HR38 does not appear to have a critical role in embryogenesis, but is required for adult cuticle formation during later pupal morphogenesis [206,207]. Recent evidence has also indicated a regulatory role for HR38 in oscillatory patterns associated with circadian rhythms [208].
The C. elegans NR4A member, nhr-6, functions in the development of the spermatheca. In conjunction with the transcription factor Jun, it is required for cell cycle progression in spermathecal cells, thus leading to a defect in cell proliferation and a significant reduction in fertility [209-211]. Null alleles do not display molting defects, suggesting that this class of nuclear receptor has been repurposed to unrelated functions among Ecydsozoa, yet the role (however circumscribed) in cell proliferation does allow some comparison to vertebrate NR4A gene function.
Vertebrates boast three NR4A paralogs: Nur77 (aka NGF1B), Nurr1, and Nor-1 (NR4A1, NR4A2, NR4A3). They were first identified molecularly as immediate-early response genes [212]. Much of the work on this gene class has focused on cell biological studies of their role in metabolism, cell cycle control, and apoptosis [213-215]. Nur77 interacts with RXR and p53 to regulate genes associated the cellular proliferation and participates with RXR and Bcl-2 in regulating apoptosis via a non-transcriptional pathway [214]. While Nur77 has both pro- and anti-proliferative functions, loss of Nurr1 and Nor-1 both lead to net proliferation of cells, suggesting a tumor suppressor role for the latter two. Mice carrying mutations in both genes suffer from rapid-onset acute myeloid leukemia, dying after a few weeks with liver and hematopoietic defects [216]. Thus vertebrate NR4A genes play a major role in regulating hematopoietic stem cells. Although various NR4A knockouts in mice are viable at birth, the Nurr1 gene has been implicated in neuronal differentiation [217]. Other studies have pointed to a role for Nur77 in the differentiation of testicular Leydig cells in mice [218], reminiscent of the defined function of nhr-6 in the C. elegans spermatheca. Therefore, there is some hint of conserved functions between nematodes and mice, but overall the theme for NR4A subfamily members is one of exaptation.
FTZ-F1/NHR-25/SF-1/LRH-1 (NR5A) and HR39 (NR5B)
The NR5 family consists of one broadly-conserved gene family member (NR5A) and a second gene (HR39/NR5B) that appears to have been lost in nematodes and vertebrates (Table 1). The ftz-F1 gene of Drosophila was first identified on the basis of its biochemical activity as a factor that bound the “zebra element” controlling segmental patterns of ftz homeobox gene expression in the embryo [219,220]. During embryogenesis, maternally-produced Ftz-F1 protein interacts with Ftz protein to regulate downstream gene expression [221-223]. A second isoform functions in larval and pupal development [224]. Here, ftz-F1 is required for molting and morphogenesis; mutants arrest at larval molts and exhibit defects in adult head eversion, leg elongation, and salivary gland development during the transition to pupa [225,226]. In the nervous system, Ftz-F1 represses the other NR5 family member, HR39/NR5B, which in turn represses EcR (NR1H) transcription in mushroom body gamma neurons during morphogenesis [227, 228]. In these cells, ftz-F1 functions in pruning and remodeling of excess neurites.
As in Drosophila, the C. elegans ortholog nhr-25 is required for a wide variety of developmental processes, including embryogenesis, vulval and gonadal development, and timing of life cycle events [229-231]. Embryos deficient for nhr-25 arrest during elongation in mid-embryogenesis, with defects in hypodermal cell fusion [232, 233]. During larval development, NHR-25 acts in combination with the LIN-39/Dfd homeodomain in vulval precursor cells [233]. While this is reminiscent of the Ftz/Ftz-F1 interaction in Drosophila, the coordinating homeobox genes are not orthologs. It also functions in gonad morphogenesis and regulation of germ-line cell division [229,232]. nhr-25 is required for the larva-adult transition and interacts in a complex pathway with heterochronic genes [231], analogous to the morphogenesis requirement for Drosophila. Therefore, a role in the coordinated timing of events related to molting and developmental transition appears to be conserved in Ecdysozoa. In other developmental pathways, nhr-25 may synergize with or antagonize Wnt signaling [230]. This is particularly interesting since Drosophila Ftz-F1 regulates various patterning genes including engrailed, but has not been directly implicated in wingless signaling [234].
Two vertebrate members of NR5A subfamily have been identified. Steroidogenic Factor-1 (SF-1; NR5A1) was first identified by molecular techniques as a transcription factor associated with regulation of steroid hydroxylase genes [235,236]. It is expressed in gonads, adrenals, and the pituitary [237], where it plays a critical developmental role in the development of steroid-producing and responsive tissues. Loss of SF-1 results in absence of adrenal glands and gonads, indicating a critical role in organogenesis [238]. In the gonads of mice, SF-1 function is required for a range of masculinization events [239]. It plays a similar role in the juvenile-to-adult switch of gonadogenesis in zebrafish [240], reminiscent of its function in early metamorphosis in the fly. In zebrafish, it appears to function upstream of Wnt signaling, similar to nhr-25. Specific knockout of SF-1 in the pituitary results in loss of follicle-stimulating hormone and luteinizing hormone-producing cells, with concomitant under-development of gonads [241]. SF-1 is required for differentiation of the VMN ventromedial hypothalamic nucleus cells, where it also mediates the normal circuitry between the hypothalamus and limbic portions of the telencephalon [242]. While not precisely the same, these functions are reminiscent of neuronal functions described in Drosophila.
LRH-1 (NR5A2) is expressed in endodermal tissues, including the intestine, liver and pancreas, as well as the ovary and brain [243]. It is essential in mice due to multiple requirements in development of organs, perhaps due to its role as a key regulator of pluripotency in early stem cells [244-246]. It binds to β-catenin and thus is an apparent target of Wnt regulation, like several other NR5 nuclear receptors [247]. In the pancreas, LRH-1 functions in the formation of the multipotent progenitor cell, which eventually gives rise to a number of different pancreatic cell types [248]. In zebrafish, LRH-1 also functions in gut, liver, and pancreas development and synergizes with Wnt signaling pathways [249,250]. Both SF-1 and LRH-1 bind and respond to phospholipids [251].
Less is known about Drosophila HR39, which is absent from C. elegans and vertebrates, but present in lophotrochozoans and echinoderms. In addition to the aforementioned role in nervous system development, HR39 also functions in the development of female reproductive glands [252], once again emphasizing a role in gonadogenesis for this gene class. Because HR39 mutants are viable, the gene does not appear to play an essential role in life cycle transitions.
In summary, the role for NR5A subfamily members in gonadogenesis appears to be fairly well-conserved. Furthermore, subfamily members have broad functions in organogenesis and life cycle transition. Specific examples of exaptation are also evident, such as the role of Ftz-F1 in segmental patterning in Drosophila.
HR4/NHR-91/GCNF (NR6A)
The Drosophila HR4 gene was identified as a nuclear receptor that is ecdysone-responsive. Like its vertebrate ortholog GCNF, HR4 is essential for embryogenesis—although embryos arrest late with cuticular defects [224]. Disruption of HR4 during larval development results in premature pupariation and arrest in early metamorphosis [61, 224]. It is required for proper upregulation of FTZ-F1 and operates in conjunction with HR3 to execute an early temporal switch in the progression from larva to pupae. Some of this function is mediated by the protein’s movement from cytoplasm to nucleus in the prothoracic gland, where it inhibits the expression of ecdysone [253]. Thus HR4 functions both downstream and upstream of the major temporal regulatory hormone in Drosophila. Roles for HR4 in stem cell development or gametogenesis have not been described.
The nhr-91 gene of C. elegans was first identified based on its orthology to GCNF [25]. It is broadly expressed in epithelial cells including the vulva, the spermatheca, excretory cell and a few neurons. The function of nhr-91 has not been well characterized—attempts to reduce function by RNAi do not cause the severe arrest phenotype observed in mice and flies (although a lethal allele has been reported). nhr-91 is a target of miR-235 miRNA, where it plays a role in the transition from L1 to L2 larval stages [254]. Expression of nhr-91 is sharply up-regulated in response to activation of the larval feeding. These observations hint at a possible broad temporal regulatory role like that seen in Drosophila.
The Germ Cell Nuclear Factor (GCNF; NR6A1) gene was identified in the early 1990’s as a nuclear receptor expressed in mouse sperm [255]. Its function in gametogenesis and embryogenesis has been extensively studied in mice, frogs, and vertebrate tissue culture systems. Mouse knockouts cause homozygous embryonic arrest due to cardiovascular defects, anterior-posterior patterning disruptions, and defects in the development of somites [255]. It also plays a key role in nervous system development—including the normal differentiation and proliferation of neurons in the isthmic organizer and neural stem cells in the early neuroectoderm [256,257]. In multiple cells, GCNF is a potent transcriptional repressor of the POU homeodomain protein Oct4 [255,257]. In embryogenesis, expression of Oct4 is normally limited to the germ-line after gastrulation. Loss of GCNF causes ectopic expression of Oct4, thus broadly inhibiting the differentiation of various cells [255]. GCNF also plays multiple roles in both spermatogenesis and oogenesis [258,259]. A role for GCNF in broad temporal developmental events has not been described. Therefore, the overall theme for the NR6A subfamily is one of exaptation of function over evolutionary time.
Themes of conservation and exaptation in nuclear receptor evolution and development
Studies of homeodomain and HLH transcription factors have revealed broad conservation of functions in development across animal phyla. For example, the homeobox genes play similar roles in patterning anterior-posterior embryonic development in flies, worms, and mice [13], and can even substitute for each other functionally [260]. Likewise, HLH transcription factors play conserved roles in muscle development and control of cell division [261,262]. In most cases, mechanistic details differ from one species to another, so patterns of conserved functions may be general or extend to details of cellular pathways [263]. Given the early evolution and strong conservation of nuclear receptors, one might expect to discover similar themes of conserved function in animal development.
Many nuclear receptors play central roles in coordinating broad developmental responses to life cycle transitions such as larval molting and morphogenesis (Table 2; Figure 2). This includes 1D (E75), 1E (E78), 1F (HR3/nhr-23), 1H (EcR), 1J (daf-12), 2B (usp), 2C (HR78), 4A (HR38/nhr-6), 5A (Ftz-F1/nhr-25/SF-1), and 6A (HR4/nhr-91). The nuclear receptors seem particularly well-suited to such developmental events, since they may combine both ligand-responsiveness and transcriptional regulation into a single protein—an arrangement that should be effective for a coordinated whole-animal event. However, it is worth noting that several of these life cycle regulators do not have known ligands (Table 1). Moreover, aspects of these regulatory circuits differ somewhat even among insects [224]. In vertebrates, where life cycle transitions are less dramatic and molting is not required, these nuclear receptors regulate a range of different functions in cell differentiation and organogenesis. Therefore, the lack of sharp, dramatic life cycle transitions, such as molting and morphogenesis, can be seen as an opportunity for vertebrate repurposing of nuclear receptors to new functions. At the same time, some of these same nuclear receptors play roles in embryogenesis, nervous system development, and control of biological rhythms that are more broadly conserved (e.g. E75 and Rev-Erb in rhythms), suggesting that these may represent ancestral functions.
TABLE 2.
Functions of Conserved Nuclear Receptors in Development
| Class | D. melanogaster | C. elegans | M. musculus |
|---|---|---|---|
|
| |||
| 1D | E75 | NHR-85 | Rev-Erb α/β |
| molting, rhythms, metamorphosis, oogenesis | molting, life cycle, female organogenesis | rhythms | |
|
| |||
| 1E | E78 | SEX-1 | (ortholog absent) |
| female organogenesis, oogenesis, gastrulation, metamorphosis | female sex determination, female organogenesis | ||
|
| |||
| 1F | HR3 | NHR-23 | ROR α/β/γ |
| embryogenesis, metamorphosis, nervous system, trachea, cuticle, wings | molting, life cycle, cuticle | nervous system, immune system, rhythms, eyes | |
|
| |||
| 1H | EcR | (ortholog absent) | LXR α/β |
| embryogenesis, metamorphosis, gut, male nervous system | nervous system, immune system | ||
|
| |||
| 1J | HR96 | DAF-12/NHR-8/-48 | VDR/PXR/CAR |
| (none known) | life cycle, longevity | female organogenesis, immune system, bones | |
|
| |||
| 2A | HNF4 | superfamily | HNF4 α/γ |
| (none known) | excretory tube, germ-line, embryogenesis, hypodermis, muscle | gastrulation, liver, nervous system | |
|
| |||
| 2B | USP | (ortholog absent) | RXR α/β/γ |
| embryogenesis, metamorphosis, gut, male nervous system | embryogenesis, heart, male spermatogenesis, neurogenesis, eyes, rhythms | ||
|
| |||
| 2C | HR78 | NHR-41 | TR2/TR4 |
| molting, trachea, larval growth | cuticle | eyes, fetal growth, nervous system, embryogenesis, stem cells | |
|
| |||
| 2E | TLL | NHR-67 | TLX |
| embryonic patterning, nervous system, eyes | embryogenesis, nervous system, female organogenesis, cell migration, cell division | nervous system, stem cells, eyes | |
|
| |||
| 2E | DSF | (ortholog absent) | (ortholog absent) |
| nervous system | |||
|
| |||
| 2E | UNF | FAX-1 | PNR |
| nervous system, rhythms | nervous system | eyes | |
|
| |||
| 2E | HR83 | NHR-239 | (ortholog absent) |
| (none known) | (none known) | ||
|
| |||
| 2F | SVP | UNC-55 | COUP-TF α/β/EAR2 |
| eyes, fat cells, heart | nervous system | embryogenesis, heart, vascular system, nervous system, bone, eyes | |
|
| |||
| 3B | ERR | (ortholog absent) | ERR α/β/γ |
| spermatogenesis | embryogenesis, heart, nervous system, kidney | ||
|
| |||
| 4A | HR38 | NHR-6 | NUR77/NURR1/NOR1 |
| cuticle, morphogenesis, rhythms | male organogenesis, cell division | cell division, apoptosis, hematopoiesis, nervous system, liver, male organogenesis | |
|
| |||
| 5A | FTZ-F1 | NHR-25/-97/-119 | SF-1/LRH-1 |
| embryonic patterning, molting, morphogenesis, nervous system | embryogenesis, female organogenesis, hypodermis, life cycle | gonads, life cycle, adrenals, pituitary, nervous system, stem cells, gut, liver, pancreas | |
|
| |||
| 5B | HR39 | (ortholog absent) | (ortholog absent) |
| female organogenesis | |||
|
| |||
| 6A | HR4 | NHR-91 | GCNF |
| embryogenesis, cuticle, metamorphosis | life cycle | embryogenesis, gametogenesis, nervous system, heart | |
Figure 2. Developmental sites of action in flies, worms, and mice.

Selected key organ systems and broad patterning functions for nuclear receptors in development are shown. Color scheme at right identifies specific NRs by overall class. Some functions are indicated at the temporal stage at which they are observed, but may function earlier. Genes assigned to the “pattern” class may have roles in specifying embryonic region (e.g. tll), major embryonic events such as gastrulation (e.g. E78), or early cell divisions (e.g. LRH-1, TR2). Illustrations are not to scale. Artwork is by S. Bodofsky.
Some of the NRs with the strongest conservation and representation in different animal phyla are those of the NR2 group. This includes nuclear receptors such as tailless and COUP-TF that have extensively-studied functions in regulating development. The suggestion that a conserved regulatory pathway involving fly tll and vertebrate Tlx in the development of neuronal systems for aggressive behavior is particularly intriguing [161]. Evidence from worms, flies, and mice also suggests that nhr-67 and Tlx play similar roles in regulating cell division and neuron development [148]. The NR2E3 family (unf/fax-1) plays a conserved role in neuronal specification and regulation of neurite arborization. However, even with these more conserved NRs there are still clear examples of exaptation. For example, the functions of tll in embryonic patterning in Drosophila and nhr-67 in uterine development in C. elegans do not seem to be functions for which there are exact correlates in other phyla. Overall, the NR2 receptors are heavily utilized for developmental functions, particularly in embryogenesis and nervous system development (Figure 2).
Eye development is a major theme in vertebrate nuclear receptor developmental function. RORβ, RXRα, TR2, TLX, PNR and COUP-TFα all play critical roles in the patterning, differentiation, or survival of retinal cells (Table 2; Figure 2). This may reflect a specialized role for retinoid signaling in the developing vertebrate eye, however tll and svp both function in Drosophila eye development, which is not a homologous process and is not dependent on retinoids. Nematodes do not have eyes and it is not clear what cells have an equivalent ontogenetic origin. Nonetheless, the common function of tll/Tlx and svp/COUP-TFα suggests an evolutionarily ancient function in eye development.
The NR1 group appears to be more plastic in terms of sequence and, in at least one case, function. The NR1D-E-F and H-I-J groups are difficult to organize into well-supported clades due to lower overall sequence conservation [Figure 1]. Our evaluation of representative members of each orthologous gene represents a parsimonious interpretation of their relationships. However, common regulatory relationships and functions among EcR in Drosophila (NR1H), daf-12 in C. elegans (NR1J), and estrogen receptor in vertebrates (NR3A) [89, 90] suggest that functional pathways mediated by NR1 genes can be swapped to other NRs over evolutionary time. That is, individual NRs can repurpose to take on functions performed by another NR. Therefore, in some cases conserved functions may not be accomplished by strict orthologs in different species.
Conservation at the level of sequence is greatest in the DBD, consistent with observations that protein-DNA binding relationships have been maintained over large evolutionary distances [264,265]. By contrast, the LBD is considerably less conserved, especially in C. elegans as compared to other animals [106, 184]. This provides some flexibility in terms of potential ligand control as well as co-activators and co-repressors. Furthermore, exaptation may be accomplished at the level of regulation of the NR itself or in variation of the downstream targets that possess a conserved DNA binding site for a given NR. Knowledge of the precise targets of NR regulation in multiple species should help provide an understanding of these modes of evolutionary change.
Overall, the theme of nuclear receptor function in development emphasizes exaptation over functional conservation. A few examples, such as the Drosophila E75 and vertebrate Rev-Erb conserved function in circadian rhythms and possible conserved functions in neuronal developmental pathways between Drosophila tll and vertebrate Tlx, indicate that some functions have been maintained over a long history of animal evolution. But there are many examples of exaptation. In particular, the vertebrate immune system has recruited several nuclear receptors, including ROR, LXR and VDR, for regulating key steps in cellular differentiation or survival. Functional relationships among conserved nuclear receptors will benefit from studies that focus on functions in additional taxa such as Lophotrochozoa.
Acknowledgments
BW is supported by grant 1R15GM107799 from the National Institutes of Health.
Footnotes
Competing interests
The authors have no competing interests.
References
- 1.Lyon MF, Hawkes SG. X-linked gene for testicular feminization in the mouse. Nature. 1970 Sep 19;227(5264):1217–9. doi: 10.1038/2271217a0. [DOI] [PubMed] [Google Scholar]
- 2.Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974 May;77(1):71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980 Oct 30;287(5785):795–801. doi: 10.1038/287795a0. [DOI] [PubMed] [Google Scholar]
- 4.Riddle DL, Swanson MM, Albert PS. Interacting genes in nematode dauer larva formation. Nature. 1981 Apr 23;290(5808):668–71. doi: 10.1038/290668a0. [DOI] [PubMed] [Google Scholar]
- 5.Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schütz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, Evans RM. The nuclear receptor superfamily: the second decade. Cell. 1995 Dec 15;83(6):835–9. doi: 10.1016/0092-8674(95)90199-x. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bertrand S, Brunet FG, Escriva H, Parmentier G, Laudet V, Robinson-Rechavi M. Evolutionary genomics of nuclear receptors: from twenty-five ancestral genes to derived endocrine systems. Mol Biol Evol. 2004 Oct;21(10):1923–37. doi: 10.1093/molbev/msh200. Epub 2004 Jun 30. [DOI] [PubMed] [Google Scholar]
- 7.Evans RM, Mangelsdorf DJ. Nuclear Receptors, RXR, and the Big Bang. Cell. 2014 Mar 27;157(1):255–66. doi: 10.1016/j.cell.2014.03.012. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Carroll SB, Grenier JK, Weatherbee SD. From DNA to diversity: Molecular genetics and the evolution of animal design. 2. Blackwell Publishing; 2005. [Google Scholar]
- 9.Bridgham JT, Carroll SM, Thornton JW. Evolution of hormone-receptor complexity by molecular exploitation. Science. 2006 Apr 7;312(5770):97–101. doi: 10.1126/science.1123348. [DOI] [PubMed] [Google Scholar]
- 10.Baker ME. Trichoplax, the simplest known animal, contains an estrogen-related receptor but no estrogen receptor: Implications for estrogen receptor evolution. Biochem Biophys Res Commun. 2008 Oct 31;375(4):623–7. doi: 10.1016/j.bbrc.2008.08.047. Epub 2008 Aug 21. [DOI] [PubMed] [Google Scholar]
- 11.Sluder AE, Mathews SW, Hough D, Yin VP, Maina CV. The nuclear receptor superfamily has undergone extensive proliferation and diversification in nematodes. Genome Res. 1999 Feb;9(2):103–20. [PubMed] [Google Scholar]
- 12.Wu W, Niles EG, Hirai H, LoVerde PT. Evolution of a novel subfamily of nuclear receptors with members that each contain two DNA binding domains. BMC Evol Biol. 2007 Feb 23;7:27. doi: 10.1186/1471-2148-7-27. Two DBD NRs. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.McGinnis W, Krumlauf R. Homeobox genes and axial patterning. Cell. 1992 Jan 24;68(2):283–302. doi: 10.1016/0092-8674(92)90471-n. Review. [DOI] [PubMed] [Google Scholar]
- 14.Gould SJ, Vrba E. Exaptation—A missing term in the science of form. Paleobiology. 1982;8:4–15. [Google Scholar]
- 15.Vogeler S, Galloway TS, Lyons BP, Bean TP. The nuclear receptor gene family in the Pacific oyster, Crassostrea gigas, contains a novel subfamily group. BMC Genomics. 2014 May 15;15:369. doi: 10.1186/1471-2164-15-369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Seaver EC. Annelid models I: Capitella teleta. Curr Opin Genet Dev. 2016 Aug;39:35–41. doi: 10.1016/j.gde.2016.05.025. Epub 2016 Jun 16. Review. [DOI] [PubMed] [Google Scholar]
- 17.Crossgrove K, Maina CV, Robinson-Rechavi M, Lochner MC. Orthologues of the Drosophila melanogaster E75 molting control gene in the filarial parasites Brugia malayi and Dirofilaria immitis. Mol Biochem Parasitol. 2008 Jan;157(1):92–7. doi: 10.1016/j.molbiopara.2007.08.010. Epub 2007 Sep 2. [DOI] [PubMed] [Google Scholar]
- 18.Jaumouillé E, Machado Almeida P, Stähli P, Koch R, Nagoshi E. Transcriptional regulation via nuclear receptor crosstalk required for the Drosophila circadian clock. Curr Biol. 2015 Jun 1;25(11):1502–8. doi: 10.1016/j.cub.2015.04.017. Epub 2015 May 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fisk GJ, Thummel CS. Isolation, regulation, and DNA-binding properties of three Drosophila nuclear hormone receptor superfamily members. Proc Natl Acad Sci U S A. 1995 Nov 7;92(23):10604–8. doi: 10.1073/pnas.92.23.10604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Parvy JP, Wang P, Garrido D, Maria A, Blais C, Poidevin M, Montagne J. Forward and feedback regulation of cyclic steroid production in Drosophila melanogaster. Development. 2014 Oct;141(20):3955–65. doi: 10.1242/dev.102020. Epub 2014 Sep 24. [DOI] [PubMed] [Google Scholar]
- 21.Morris LX, Spradling AC. Steroid signaling within Drosophila ovarian epithelial cells sex-specifically modulates early germ cell development and meiotic entry. PLoS One. 2012;7(10):e46109. doi: 10.1371/journal.pone.0046109. Epub 2012 Oct 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Buszczak M, Freeman MR, Carlson JR, Bender M, Cooley L, Segraves WA. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development. 1999 Oct;126(20):4581–9. doi: 10.1242/dev.126.20.4581. [DOI] [PubMed] [Google Scholar]
- 23.Li Y, Ma Q, Cherry CM, Matunis EL. Steroid signaling promotes stem cell maintenance in the Drosophila testis. Dev Biol. 2014 Oct 1;394(1):129–41. doi: 10.1016/j.ydbio.2014.07.016. Epub 2014 Aug 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kumar S, Chen D, Jang C, Nall A, Zheng X, Sehgal A. An ecdysone-responsive nuclear receptor regulates circadian rhythms in Drosophila. Nat Commun. 2014 Dec 16;5:5697. doi: 10.1038/ncomms6697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gissendanner CR, Crossgrove K, Kraus KA, Maina CV, Sluder AE. Expression and function of conserved nuclear receptor genes in Caenorhabditis elegans. Dev Biol. 2004 Feb 15;266(2):399–416. doi: 10.1016/j.ydbio.2003.10.014. [DOI] [PubMed] [Google Scholar]
- 26.Raizen DM, Zimmerman JE, Maycock MH, Ta UD, You YJ, Sundaram MV, Pack AI. Lethargus is a Caenorhabditis elegans sleep-like state. Nature. 2008 Jan 31;451(7178):569–72. doi: 10.1038/nature06535. Epub 2008 Jan 9. Erratum in: Nature. 2008 Jun 12;453(7197):952. [DOI] [PubMed] [Google Scholar]
- 27.Preitner N, Damiola F, Lopez-Molina L, Zakany J, Duboule D, Albrecht U, Schibler U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002 Jul 26;110(2):251–60. doi: 10.1016/s0092-8674(02)00825-5. [DOI] [PubMed] [Google Scholar]
- 28.Cho H, Zhao X, Hatori M, Yu RT, Barish GD, Lam MT, Chong LW, DiTacchio L, Atkins AR, Glass CK, Liddle C, Auwerx J, Downes M, Panda S, Evans RM. Regulation of circadian behaviour and metabolism by REV-ERB-α and REV-ERB-β. Nature. 2012 Mar 29;485(7396):123–7. doi: 10.1038/nature11048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Guillaumond F, Dardente H, Giguère V, Cermakian N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J Biol Rhythms. 2005 Oct;20(5):391–403. doi: 10.1177/0748730405277232. [DOI] [PubMed] [Google Scholar]
- 30.Bugge A, Feng D, Everett LJ, Briggs ER, Mullican SE, Wang F, Jager J, Lazar MA. Rev-erbα and Rev-erbβ coordinately protect the circadian clock and normal metabolic function. Genes Dev. 2012 Apr 1;26(7):657–67. doi: 10.1101/gad.186858.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huang G, Zhang F, Ye Q, Wang H. The circadian clock regulates autophagy directly through the nuclear hormone receptor Nr1d1/Rev-erbα and indirectly via Cebpb/(C/ebpβ) in zebrafish. Autophagy. 2016 Aug 2;12(8):1292–309. doi: 10.1080/15548627.2016.1183843. Epub 2016 May 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Purushothaman S, Saxena S, Meghah V, Meena Lakshmi MG, Singh SK, Brahmendra Swamy CV, Idris MM. Proteomic and gene expression analysis of zebrafish brain undergoing continuous light/dark stress. J Sleep Res. 2015 Aug;24(4):458–65. doi: 10.1111/jsr.12287. Epub 2015 Feb 27. [DOI] [PubMed] [Google Scholar]
- 33.Chomez P, Neveu I, Mansén A, Kiesler E, Larsson L, Vennström B, Arenas E. Increased cell death and delayed development in the cerebellum of mice lacking the rev-erbA(alpha) orphan receptor. Development. 2000 Apr;127(7):1489–98. doi: 10.1242/dev.127.7.1489. [DOI] [PubMed] [Google Scholar]
- 34.Ables ET, Bois KE, Garcia CA, Drummond-Barbosa D. Ecdysone response gene E78 controls ovarian germline stem cell niche formation and follicle survival in Drosophila. Dev Biol. 2015 Apr 1;400(1):33–42. doi: 10.1016/j.ydbio.2015.01.013. Epub 2015 Jan 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Uryu O, Ameku T, Niwa R. Recent progress in understanding the role of ecdysteroids in adult insects: Germline development and circadian clock in the fruit fly Drosophila melanogaster. Zoological Lett. 2015 Nov 2;1:32. doi: 10.1186/s40851-015-0031-2. eCollection 2015. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Carmi I, Kopczynski JB, Meyer BJ. The nuclear hormone receptor SEX-1 is an X-chromosome signal that determines nematode sex. Nature. 1998 Nov 12;396(6707):168–73. doi: 10.1038/24164. [DOI] [PubMed] [Google Scholar]
- 37.Magner DB, Antebi A. Caenorhabditis elegans nuclear receptors: insights into life traits. Trends Endocrinol Metab. 2008 Jul;19(5):153–60. doi: 10.1016/j.tem.2008.02.005. Epub 2008 Apr 10. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Koelle MR, Segraves WA, Hogness DS. DHR3: a Drosophila steroid receptor homolog. Proc Natl Acad Sci U S A. 1992 Jul 1;89(13):6167–71. doi: 10.1073/pnas.89.13.6167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lam GT, Jiang C, Thummel CS. Coordination of larval and prepupal gene expression by the DHR3 orphan receptor during Drosophila metamorphosis. Development. 1997 May;124(9):1757–69. doi: 10.1242/dev.124.9.1757. [DOI] [PubMed] [Google Scholar]
- 40.White KP, Hurban P, Watanabe T, Hogness DS. Coordination of Drosophila metamorphosis by two ecdysone-induced nuclear receptors. Science. 1997 Apr 4;276(5309):114–7. doi: 10.1126/science.276.5309.114. [DOI] [PubMed] [Google Scholar]
- 41.Carney GE, Wade AA, Sapra R, Goldstein ES, Bender M. DHR3, an ecdysone-inducible early-late gene encoding a Drosophila nuclear receptor, is required for embryogenesis. Proc Natl Acad Sci U S A. 1997 Oct 28;94(22):12024–9. doi: 10.1073/pnas.94.22.12024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ruaud AF, Lam G, Thummel CS. The Drosophila nuclear receptors DHR3 and betaFTZ-F1 control overlapping developmental responses in late embryos. Development. 2010 Jan;137(1):123–31. doi: 10.1242/dev.042036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kostrouchova M, Krause M, Kostrouch Z, Rall JE. Nuclear hormone receptor CHR3 is a critical regulator of all four larval molts of the nematode Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2001 Jun 19;98(13):7360–5. doi: 10.1073/pnas.131171898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kouns NA, Nakielna J, Behensky F, Krause MW, Kostrouch Z, Kostrouchova M. NHR-23 dependent collagen and hedgehog-related genes required for molting. Biochem Biophys Res Commun. 2011 Oct 7;413(4):515–20. doi: 10.1016/j.bbrc.2011.08.124. Epub 2011 Sep 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hayes GD, Frand AR, Ruvkun G. The mir-84 and let-7 paralogous microRNA genes of Caenorhabditis elegans direct the cessation of molting via the conserved nuclear hormone receptors NHR-23 and NHR-25. Development. 2006 Dec;133(23):4631–41. doi: 10.1242/dev.02655. Epub 2006 Oct 25. [DOI] [PubMed] [Google Scholar]
- 46.Becker-André M, André E, DeLamarter JF. Identification of nuclear receptor mRNAs by RT-PCR amplification of conserved zinc-finger motif sequences. Biochem Biophys Res Commun. 1993 Aug 16;194(3):1371–9. doi: 10.1006/bbrc.1993.1976. [DOI] [PubMed] [Google Scholar]
- 47.Giguère V, Tini M, Flock G, Ong E, Evans RM, Otulakowski G. Isoform-specific amino-terminal domains dictate DNA-binding properties of ROR alpha, a novel family of orphan hormone nuclear receptors. Genes Dev. 1994 Mar 1;8(5):538–53. doi: 10.1101/gad.8.5.538. [DOI] [PubMed] [Google Scholar]
- 48.Carlberg C, Hooft van Huijsduijnen R, Staple JK, DeLamarter JF, Becker-André M. RZRs, a new family of retinoid-related orphan receptors that function as both monomers and homodimers. Mol Endocrinol. 1994 Jun;8(6):757–70. doi: 10.1210/mend.8.6.7935491. [DOI] [PubMed] [Google Scholar]
- 49.Hirose T, Smith RJ, Jetten AM. ROR gamma: the third member of ROR/RZR orphan receptor subfamily that is highly expressed in skeletal muscle. Biochem Biophys Res Commun. 1994 Dec 30;205(3):1976–83. doi: 10.1006/bbrc.1994.2902. [DOI] [PubMed] [Google Scholar]
- 50.Hamilton BA, Frankel WN, Kerrebrock AW, Hawkins TL, FitzHugh W, Kusumi K, Russell LB, Mueller KL, van Berkel V, Birren BW, Kruglyak L, Lander ES. Disruption of the nuclear hormone receptor RORalpha in staggerer mice. Nature. 1996 Feb 22;379(6567):736–9. doi: 10.1038/379736a0. Erratum in: Nature 1996 May 23;381(6580):346. [DOI] [PubMed] [Google Scholar]
- 51.Chen XR, Heck N, Lohof AM, Rochefort C, Morel MP, Wehrlé R, Doulazmi M, Marty S, Cannaya V, Avci HX, Mariani J, Rondi-Reig L, Vodjdani G, Sherrard RM, Sotelo C, Dusart I. Mature Purkinje cells require the retinoic acid-related orphan receptor-α (RORα) to maintain climbing fiber mono-innervation and other adult characteristics. J Neurosci. 2013 May 29;33(22):9546–62. doi: 10.1523/JNEUROSCI.2977-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lau P, Fitzsimmons RL, Raichur S, Wang SC, Lechtken A, Muscat GE. The orphan nuclear receptor, RORalpha, regulates gene expression that controls lipid metabolism: staggerer (SG/SG) mice are resistant to diet-induced obesity. J Biol Chem. 2008 Jun 27;283(26):18411–21. doi: 10.1074/jbc.M710526200. Epub 2008 Apr 25. [DOI] [PubMed] [Google Scholar]
- 53.Halim TY, MacLaren A, Romanish MT, Gold MJ, McNagny KM, Takei F. Retinoic-acid-receptor-related orphan nuclear receptor alpha is required for natural helper cell development and allergic inflammation. Immunity. 2012 Sep 21;37(3):463–74. doi: 10.1016/j.immuni.2012.06.012. Epub 2012 Sep 13. [DOI] [PubMed] [Google Scholar]
- 54.Wong SH, Walker JA, Jolin HE, Drynan LF, Hams E, Camelo A, Barlow JL, Neill DR, Panova V, Koch U, Radtke F, Hardman CS, Hwang YY, Fallon PG, McKenzie AN. Transcription factor RORα is critical for nuocyte development. Nat Immunol. 2012 Jan 22;13(3):229–36. doi: 10.1038/ni.2208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Akashi M, Takumi T. The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat Struct Mol Biol. 2005 May;12(5):441–8. doi: 10.1038/nsmb925. Epub 2005 Apr 10. [DOI] [PubMed] [Google Scholar]
- 56.André E, Conquet F, Steinmayr M, Stratton SC, Porciatti V, Becker-André M. Disruption of retinoid-related orphan receptor beta changes circadian behavior, causes retinal degeneration and leads to vacillans phenotype in mice. EMBO J. 1998 Jul 15;17(14):3867–77. doi: 10.1093/emboj/17.14.3867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jia L, Oh EC, Ng L, Srinivas M, Brooks M, Swaroop A, Forrest D. Retinoid-related orphan nuclear receptor RORbeta is an early-acting factor in rod photoreceptor development. Proc Natl Acad Sci U S A. 2009 Oct 13;106(41):17534–9. doi: 10.1073/pnas.0902425106. Epub 2009 Sep 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Liu H, Aramaki M, Fu Y, Forrest D. Retinoid-Related Orphan Receptor β and Transcriptional Control of Neuronal Differentiation. Curr Top Dev Biol. 2017;125:227–255. doi: 10.1016/bs.ctdb.2016.11.009. Epub 2016 Dec 27. [DOI] [PubMed] [Google Scholar]
- 59.Kurebayashi S, Ueda E, Sakaue M, Patel DD, Medvedev A, Zhang F, Jetten AM. Retinoid-related orphan receptor gamma (RORgamma) is essential for lymphoid organogenesis and controls apoptosis during thymopoiesis. Proc Natl Acad Sci U S A. 2000 Aug 29;97(18):10132–7. doi: 10.1073/pnas.97.18.10132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Okada S, Markle JG, Deenick EK, Mele F, Averbuch D, Lagos M, Alzahrani M, Al-Muhsen S, Halwani R, Ma CS, Wong N, Soudais C, Henderson LA, Marzouqa H, Shamma J, Gonzalez M, Martinez-Barricarte R, Okada C, Avery DT, Latorre D, Deswarte C, Jabot-Hanin F, Torrado E, Fountain J, Belkadi A, Itan Y, Boisson B, Migaud M, Arlehamn CS, Sette A, Breton S, McCluskey J, Rossjohn J, de Villartay JP, Moshous D, Hambleton S, Latour S, Arkwright PD, Picard C, Lantz O, Engelhard D, Kobayashi M, Abel L, Cooper AM, Notarangelo LD, Boisson-Dupuis S, Puel A, Sallusto F, Bustamante J, Tangye SG, Casanova JL. IMMUNODEFICIENCIES. Impairment of immunity to Candida and Mycobacterium in humans with bi-allelic RORC mutations. Science. 2015 Aug 7;349(6248):606–13. doi: 10.1126/science.aaa4282. Epub 2015 Jul 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.King-Jones K, Thummel CS. Nuclear receptors--a perspective from Drosophila. Nat Rev Genet. 2005 Apr;6(4):311–23. doi: 10.1038/nrg1581. Review. [DOI] [PubMed] [Google Scholar]
- 62.Koelle MR, Talbot WS, Segraves WA, Bender MT, Cherbas P, Hogness DS. The Drosophila EcR gene encodes an ecdysone receptor, a new member of the steroid receptor superfamily. Cell. 1991 Oct 4;67(1):59–77. doi: 10.1016/0092-8674(91)90572-g. [DOI] [PubMed] [Google Scholar]
- 63.Kozlova T, Thummel CS. Essential roles for ecdysone signaling during Drosophila mid-embryonic development. Science. 2003 Sep 26;301(5641):1911–4. doi: 10.1126/science.1087419. Epub 2003 Sep 4. [DOI] [PubMed] [Google Scholar]
- 64.Schubiger M, Carré C, Antoniewski C, Truman JW. Ligand-dependent de-repression via EcR/USP acts as a gate to coordinate the differentiation of sensory neurons in the Drosophila wing. Development. 2005 Dec;132(23):5239–48. doi: 10.1242/dev.02093. Epub 2005 Nov 2. [DOI] [PubMed] [Google Scholar]
- 65.Dalton JE, Lebo MS, Sanders LE, Sun F, Arbeitman MN. Ecdysone receptor acts in fruitless- expressing neurons to mediate drosophila courtship behaviors. Curr Biol. 2009 Sep 15;19(17):1447–52. doi: 10.1016/j.cub.2009.06.063. Epub 2009 Jul 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Mansilla A, Martín FA, Martín D, Ferrús A. Ligand-independent requirements of steroid receptors EcR and USP for cell survival. Cell Death Differ. 2016 Mar;23(3):405–16. doi: 10.1038/cdd.2015.108. Epub 2015 Aug 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Tzertzinis G, Egaña AL, Palli SR, Robinson-Rechavi M, Gissendanner CR, Liu C, Unnasch TR, Maina CV. Molecular evidence for a functional ecdysone signaling system in Brugia malayi. PLoS Negl Trop Dis. 2010 Mar 9;4(3):e625. doi: 10.1371/journal.pntd.0000625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Parihar M, Minton RL, Flowers S, Holloway A, Morehead BE, Paille J, Gissendanner CR. The genome of the nematode Pristionchus pacificus encodes putative homologs of RXR/Usp and EcR. Gen Comp Endocrinol. 2010 May 15;167(1):11–7. doi: 10.1016/j.ygcen.2010.02.005. Epub 2010 Feb 10. [DOI] [PubMed] [Google Scholar]
- 69.Peet DJ, Janowski BA, Mangelsdorf DJ. The LXRs: a new class of oxysterol receptors. Curr Opin Genet Dev. 1998 Oct;8(5):571–5. doi: 10.1016/s0959-437x(98)80013-0. Review. [DOI] [PubMed] [Google Scholar]
- 70.Willy PJ, Umesono K, Ong ES, Evans RM, Heyman RA, Mangelsdorf DJ. LXR, a nuclear receptor that defines a distinct retinoid response pathway. Genes Dev. 1995 May 1;9(9):1033–45. doi: 10.1101/gad.9.9.1033. [DOI] [PubMed] [Google Scholar]
- 71.Janowski BA, Willy PJ, Devi TR, Falck JR, Mangelsdorf DJ. An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature. 1996 Oct 24;383(6602):728–31. doi: 10.1038/383728a0. [DOI] [PubMed] [Google Scholar]
- 72.Andersson S, Gustafsson N, Warner M, Gustafsson JA. Inactivation of liver X receptor beta leads to adult-onset motor neuron degeneration in male mice. Proc Natl Acad Sci U S A. 2005 Mar 8;102(10):3857–62. doi: 10.1073/pnas.0500634102. Epub 2005 Feb 28. Erratum in: Proc Natl Acad Sci U S A. 2006 May 23;103(21):8298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bensinger SJ, Bradley MN, Joseph SB, Zelcer N, Janssen EM, Hausner MA, Shih R, Parks JS, Edwards PA, Jamieson BD, Tontonoz P. LXR signaling couples sterol metabolism to proliferation in the acquired immune response. Cell. 2008 Jul 11;134(1):97–111. doi: 10.1016/j.cell.2008.04.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Joseph SB, Bradley MN, Castrillo A, Bruhn KW, Mak PA, Pei L, Hogenesch J, O’connell RM, Cheng G, Saez E, Miller JF, Tontonoz P. LXR-dependent gene expression is important for macrophage survival and the innate immune response. Cell. 2004 Oct 15;119(2):299–309. doi: 10.1016/j.cell.2004.09.032. [DOI] [PubMed] [Google Scholar]
- 75.Forman BM, Goode E, Chen J, Oro AE, Bradley DJ, Perlmann T, Noonan DJ, Burka LT, McMorris T, Lamph WW, Evans RM, Weinberger C. Identification of a nuclear receptor that is activated by farnesol metabolites. Cell. 1995 Jun 2;81(5):687–93. doi: 10.1016/0092-8674(95)90530-8. [DOI] [PubMed] [Google Scholar]
- 76.Sinal CJ, Tohkin M, Miyata M, Ward JM, Lambert G, Gonzalez FJ. Targeted disruption of the nuclear receptor FXR/BAR impairs bile acid and lipid homeostasis. Cell. 2000 Sep 15;102(6):731–44. doi: 10.1016/s0092-8674(00)00062-3. [DOI] [PubMed] [Google Scholar]
- 77.King-Jones K, Horner MA, Lam G, Thummel CS. The DHR96 nuclear receptor regulates xenobiotic responses in Drosophila. Cell Metab. 2006 Jul;4(1):37–48. doi: 10.1016/j.cmet.2006.06.006. [DOI] [PubMed] [Google Scholar]
- 78.Sieber MH, Thummel CS. The DHR96 nuclear receptor controls triacylglycerol homeostasis in Drosophila. Cell Metab. 2009 Dec;10(6):481–90. doi: 10.1016/j.cmet.2009.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Horner MA, Pardee K, Liu S, King-Jones K, Lajoie G, Edwards A, Krause HM, Thummel CS. The Drosophila DHR96 nuclear receptor binds cholesterol and regulates cholesterol homeostasis. Genes Dev. 2009 Dec 1;23(23):2711–6. doi: 10.1101/gad.1833609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Bujold M, Gopalakrishnan A, Nally E, King-Jones K. Nuclear receptor DHR96 acts as a sentinel for low cholesterol concentrations in Drosophila melanogaster. Mol Cell Biol. 2010 Feb;30(3):793–805. doi: 10.1128/MCB.01327-09. Epub 2009 Nov 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Afschar S, Toivonen JM, Hoffmann JM, Tain LS, Wieser D, Finlayson AJ, Driege Y, Alic N, Emran S, Stinn J, Froehlich J, Piper MD, Partridge L. Nuclear hormone receptor DHR96 mediates the resistance to xenobiotics but not the increased lifespan of insulin-mutant Drosophila. Proc Natl Acad Sci U S A. 2016 Feb 2;113(5):1321–6. doi: 10.1073/pnas.1515137113. Epub 2016 Jan 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Niwa R, Niwa YS. The Fruit Fly Drosophila melanogaster as a Model System to Study Cholesterol Metabolism and Homeostasis. Cholesterol. 2011;2011:176802. doi: 10.1155/2011/176802. Epub 2011 Jan 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Hoffmann JM, Partridge L. Nuclear hormone receptors: Roles of xenobiotic detoxification and sterol homeostasis in healthy aging. Crit Rev Biochem Mol Biol. 2015;50(5):380–92. doi: 10.3109/10409238.2015.1067186. Epub 2015 Sep 18. Review. [DOI] [PubMed] [Google Scholar]
- 84.Antebi A. Steroid regulation of C. elegans diapause, developmental timing, and longevity. Curr Top Dev Biol. 2013;105:181–212. doi: 10.1016/B978-0-12-396968-2.00007-5. Review. [DOI] [PubMed] [Google Scholar]
- 85.Hu PJ. Dauer (August 08, 2007), WormBook. The C. elegans Research Community, WormBook; http://www.wormbook.org. [DOI] [Google Scholar]
- 86.Antebi A, Yeh WH, Tait D, Hedgecock EM, Riddle DL. daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 2000 Jun 15;14(12):1512–27. [PMC free article] [PubMed] [Google Scholar]
- 87.Motola DL, Cummins CL, Rottiers V, Sharma KK, Li T, Li Y, Suino-Powell K, Xu HE, Auchus RJ, Antebi A, Mangelsdorf DJ. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell. 2006 Mar 24;124(6):1209–23. doi: 10.1016/j.cell.2006.01.037. Epub 2006 Mar 9. [DOI] [PubMed] [Google Scholar]
- 88.Bethke A, Fielenbach N, Wang Z, Mangelsdorf DJ, Antebi A. Nuclear hormone receptor regulation of microRNAs controls developmental progression. Science. 2009 Apr 3;324(5923):95–8. doi: 10.1126/science.1164899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tennessen JM, Thummel CS. Coordinating growth and maturation - insights from Drosophila. Curr Biol. 2011 Sep 27;21(18):R750–7. doi: 10.1016/j.cub.2011.06.033. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Antebi A. WormBook. The C. elegans Research Community, WormBook; Jun 9, 2015. Nuclear receptor signal transduction in C. elegans. http://www.wormbook.org. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Magner DB, Wollam J, Shen Y, Hoppe C, Li D, Latza C, Rottiers V, Hutter H, Antebi A. The NHR-8 nuclear receptor regulates cholesterol and bile acid homeostasis in C. elegans. Cell Metab. 2013 Aug 6;18(2):212–24. doi: 10.1016/j.cmet.2013.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.McDonnell DP, Mangelsdorf DJ, Pike JW, Haussler MR, O’Malley BW. Molecular cloning of complementary DNA encoding the avian receptor for vitamin D. Science. 1987 Mar 6;235(4793):1214–7. doi: 10.1126/science.3029866. [DOI] [PubMed] [Google Scholar]
- 93.Smith DP, Mason CS, Jones EA, Old RW. A novel nuclear receptor superfamily member in Xenopus that associates with RXR, and shares extensive sequence similarity to the mammalian vitamin D3 receptor. Nucleic Acids Res. 1994 Jan 11;22(1):66–71. doi: 10.1093/nar/22.1.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Baes M, Gulick T, Choi HS, Martinoli MG, Simha D, Moore DD. A new orphan member of the nuclear hormone receptor superfamily that interacts with a subset of retinoic acid response elements. Mol Cell Biol. 1994 Mar;14(3):1544–52. doi: 10.1128/mcb.14.3.1544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Yoshizawa T, Handa Y, Uematsu Y, Takeda S, Sekine K, Yoshihara Y, Kawakami T, Arioka K, Sato H, Uchiyama Y, Masushige S, Fukamizu A, Matsumoto T, Kato S. Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine hypoplasia and growth retardation after weaning. Nat Genet. 1997 Aug;16(4):391–6. doi: 10.1038/ng0897-391. [DOI] [PubMed] [Google Scholar]
- 96.Donohue MM, Demay MB. Rickets in VDR null mice is secondary to decreased apoptosis of hypertrophic chondrocytes. Endocrinology. 2002 Sep;143(9):3691–4. doi: 10.1210/en.2002-220454. [DOI] [PubMed] [Google Scholar]
- 97.Yu S, Cantorna MT. The vitamin D receptor is required for iNKT cell development. Proc Natl Acad Sci U S A. 2008 Apr 1;105(13):5207–12. doi: 10.1073/pnas.0711558105. Epub 2008 Mar 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Xie W, Barwick JL, Downes M, Blumberg B, Simon CM, Nelson MC, Neuschwander-Tetri BA, Brunt EM, Guzelian PS, Evans RM. Humanized xenobiotic response in mice expressing nuclear receptor SXR. Nature. 2000 Jul 27;406(6794):435–9. doi: 10.1038/35019116. [DOI] [PubMed] [Google Scholar]
- 99.Bertilsson G, Heidrich J, Svensson K, Asman M, Jendeberg L, Sydow-Bäckman M, Ohlsson R, Postlind H, Blomquist P, Berkenstam A. Identification of a human nuclear receptor defines a new signaling pathway for CYP3A induction. Proc Natl Acad Sci U S A. 1998 Oct 13;95(21):12208–13. doi: 10.1073/pnas.95.21.12208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Watkins RE, Wisely GB, Moore LB, Collins JL, Lambert MH, Williams SP, Willson TM, Kliewer SA, Redinbo MR. The human nuclear xenobiotic receptor PXR: structural determinants of directed promiscuity. Science. 2001 Jun 22;292(5525):2329–33. doi: 10.1126/science.1060762. Epub 2001 Jun 14. [DOI] [PubMed] [Google Scholar]
- 101.Forman BM, Tzameli I, Choi HS, Chen J, Simha D, Seol W, Evans RM, Moore DD. Androstane metabolites bind to and deactivate the nuclear receptor CAR-beta. Nature. 1998 Oct 8;395(6702):612–5. doi: 10.1038/26996. [DOI] [PubMed] [Google Scholar]
- 102.Wei P, Zhang J, Egan-Hafley M, Liang S, Moore DD. The nuclear receptor CAR mediates specific xenobiotic induction of drug metabolism. Nature. 2000 Oct 19;407(6806):920–3. doi: 10.1038/35038112. [DOI] [PubMed] [Google Scholar]
- 103.Honkakoski P, Zelko I, Sueyoshi T, Negishi M. The nuclear orphan receptor CAR-retinoid X receptor heterodimer activates the phenobarbital-responsive enhancer module of the CYP2B gene. Mol Cell Biol. 1998 Oct;18(10):5652–8. doi: 10.1128/mcb.18.10.5652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Zhong W, Sladek FM, Darnell JE., Jr The expression pattern of a Drosophila homolog to the mouse transcription factor HNF-4 suggests a determinative role in gut formation. EMBO J. 1993;12:537–544. doi: 10.1002/j.1460-2075.1993.tb05685.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Palanker L, Tennessen JM, Lam G, Thummel CS. Drosophila HNF4 regulates lipid mobilization and beta-oxidation. Cell Metab. 2009 Mar;9(3):228–39. doi: 10.1016/j.cmet.2009.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Robinson-Rechavi M, Maina CV, Gissendanner CR, Laudet V, Sluder A. Explosive lineage-specific expansion of the orphan nuclear receptor HNF4 in nematodes. J Mol Evol. 2005;60:577–586. doi: 10.1007/s00239-004-0175-8. [DOI] [PubMed] [Google Scholar]
- 107.Van Gilst MR, Hadjivassiliou H, Yamamoto KR. A Caenorhabditis elegans nutrient response system partially dependent on nuclear receptor NHR-49. Proc Natl Acad Sci U S A. 2005 Sep 20;102(38):13496–501. doi: 10.1073/pnas.0506234102. Epub 2005 Sep 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Van Gilst MR, Hadjivassiliou H, Jolly A, Yamamoto KR. Nuclear hormone receptor NHR-49 controls fat consumption and fatty acid composition in C. elegans. PLoS Biol. 2005 Feb;3(2):e53. doi: 10.1371/journal.pbio.0030053. Epub 2005 Feb 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Taubert S, Ward JD, Yamamoto KR. Nuclear hormone receptors in nematodes: evolution and function. Mol Cell Endocrinol. 2011 Mar 1;334(1-2):49–55. doi: 10.1016/j.mce.2010.04.021. Epub 2010 May 10. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Hahn-Windgassen A, Van Gilst MR. The Caenorhabditis elegans HNF4alpha Homolog, NHR-31, mediates excretory tube growth and function through coordinate regulation of the vacuolar ATPase. PLoS Genet. 2009 Jul;5(7):e1000553. doi: 10.1371/journal.pgen.1000553. Epub 2009 Jul 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Gracida X, Eckmann CR. Fertility and germline stem cell maintenance under different diets requires nhr-114/HNF4 in C. elegans. Curr Biol. 2013 Apr 8;23(7):607–13. doi: 10.1016/j.cub.2013.02.034. Epub 2013 Mar 14. [DOI] [PubMed] [Google Scholar]
- 112.Brozová E, Simecková K, Kostrouch Z, Rall JE, Kostrouchová M. NHR-40, a Caenorhabditis elegans supplementary nuclear receptor, regulates embryonic and early larval development. Mech Dev. 2006 Sep;123(9):689–701. doi: 10.1016/j.mod.2006.06.006. Epub 2006 Jun 28. [DOI] [PubMed] [Google Scholar]
- 113.Simecková K, Brozová E, Vohánka J, Pohludka M, Kostrouch Z, Krause MW, Rall JE, Kostrouchová M. Supplementary nuclear receptor NHR-60 is required for normal embryonic and early larval development of Caenorhabditis elegans. Folia Biol (Praha) 2007;53(3):85–96. doi: 10.14712/fb2007053030085. [DOI] [PubMed] [Google Scholar]
- 114.Sladek FM, Zhong W, Lai E, Darnell JE., Jr Liver enriched transcription factor HNF-4 is a novel member of the steroid hormone receptor superfamily. Genes Dev. 1990;4:2353–2365. doi: 10.1101/gad.4.12b.2353. [DOI] [PubMed] [Google Scholar]
- 115.Duncan SA, Nagy A, Chan W. Murine gastrulation requires HNF-4 regulated gene expression in the visceral endoderm: Tetraploid rescue of Hnf-4(/) embryos. Development. 1997;124:279–287. doi: 10.1242/dev.124.2.279. [DOI] [PubMed] [Google Scholar]
- 116.Chen WS, Manova K, Weinstein DC, Duncan SA, Plump AS, Prezioso VR, Bachvarova RF, Darnell JE., Jr Disruption of the HNF-4 gene, expressed in visceral endoderm, leads to cell death in embryonic ectoderm and impaired gastrulation of mouse embryos. Genes Dev. 1994;8:2466–2477. doi: 10.1101/gad.8.20.2466. [DOI] [PubMed] [Google Scholar]
- 117.Morrisey EE, Tang Z, Sigrist K, Lu MM, Jiang F, Ip HS, Parmacek MS. GATA6 regulates HNF4 and is required for differentiation of visceral endoderm in the mouse embryo. Genes Dev. 1998 Nov 15;12(22):3579–90. doi: 10.1101/gad.12.22.3579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Pearson ER, Boj SF, Steele AM, Barrett T, Stals K, Shield JP, Ellard S, Ferrer J, Hattersley AT. Macrosomia and hyperinsulinaemic hypoglycaemia in patients with heterozygous mutations in the HNF4A gene. PLoS Med. 2007 Apr;4(4):e118. doi: 10.1371/journal.pmed.0040118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Li J, Ning G, Duncan SA. Mammalian hepatocyte differentiation requires the transcription factor HNF-4alpha. Genes Dev. 2000 Feb 15;14(4):464–74. [PMC free article] [PubMed] [Google Scholar]
- 120.Wang J, Cheng H, Li X, Lu W, Wang K, Wen T. Regulation of neural stem cell differentiation by transcription factors HNF4-1 and MAZ-1. Mol Neurobiol. 2013 Feb;47(1):228–40. doi: 10.1007/s12035-012-8335. Epub 2012 Sep 4. [DOI] [PubMed] [Google Scholar]
- 121.Drewes T, Senkel S, Holewa B, Ryffel GU. Human hepatocyte nuclear factor 4 isoforms are encoded by distinct and differentially expressed genes. Mol Cell Biol. 1996 Mar;16(3):925–31. doi: 10.1128/mcb.16.3.925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Holewa B, Zapp D, Drewes T, Senkel S, Ryffel GU. HNF4beta, a new gene of the HNF4 family with distinct activation and expression profiles in oogenesis and embryogenesis of Xenopus laevis. Mol Cell Biol. 1997 Feb;17(2):687–94. doi: 10.1128/mcb.17.2.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Perrimon N, Engstrom L, Mahowald AP. Developmental genetics of the 2C-D region of the Drosophila X chromosome. Genetics. 1985;111:23–41. doi: 10.1093/genetics/111.1.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Oro AE, McKeown M, Evans RM. Relationship between the product of the Drosophila ultraspiracle locus and the vertebrate retinoid X receptor. Nature. 1990 Sep 20;347(6290):298–301. doi: 10.1038/347298a0. [DOI] [PubMed] [Google Scholar]
- 125.Hall BL, Thummel CS. The RXR homolog ultraspiracle is an essential component of the Drosophila ecdysone receptor. Development. 1998 Dec;125(23):4709–17. doi: 10.1242/dev.125.23.4709. [DOI] [PubMed] [Google Scholar]
- 126.Hamada K, Gleason SL, Levi BZ, Hirschfeld S, Appella E, Ozato K. H-2RIIBP, a member of the nuclear hormone receptor superfamily that binds to both the regulatory element of major histocompatibility class I genes and the estrogen response element. Proc Natl Acad Sci U S A. 1989;86(21):8289–93. doi: 10.1073/pnas.86.21.8289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mangelsdorf DJ, Ong ES, Dyck JA, Evans RM. Nuclear receptor that identifies a novel retinoic acid response pathway. Nature. 1990 May 17;345(6272):224–9. doi: 10.1038/345224a0. [DOI] [PubMed] [Google Scholar]
- 128.Mangelsdorf DJ, Evans RM. The RXR heterodimers and orphan receptors. Cell. 1995 Dec 15;83(6):841–50. doi: 10.1016/0092-8674(95)90200-7. Review. [DOI] [PubMed] [Google Scholar]
- 129.Sucov HM, Dyson E, Gumeringer CL, Price J, Chien KR, Evans RM. RXR alpha mutant mice establish a genetic basis for vitamin A signaling in heart morphogenesis. Genes Dev. 1994 May 1;8(9):1007–18. doi: 10.1101/gad.8.9.1007. [DOI] [PubMed] [Google Scholar]
- 130.Kastner P, Grondona JM, Mark M, Gansmuller A, LeMeur M, Decimo D, Vonesch JL, Dollé P, Chambon P. Genetic analysis of RXR alpha developmental function: convergence of RXR and RAR signaling pathways in heart and eye morphogenesis. Cell. 1994 Sep 23;78(6):987–1003. doi: 10.1016/0092-8674(94)90274-7. [DOI] [PubMed] [Google Scholar]
- 131.Kastner P, Mark M, Leid M, Gansmuller A, Chin W, Grondona JM, Décimo D, Krezel W, Dierich A, Chambon P. Abnormal spermatogenesis in RXR beta mutant mice. Genes Dev. 1996 Jan 1;10(1):80–92. doi: 10.1101/gad.10.1.80. [DOI] [PubMed] [Google Scholar]
- 132.Krezel W, Dupé V, Mark M, Dierich A, Kastner P, Chambon P. RXR gamma null mice are apparently normal and compound RXR alpha +/-/RXR beta -/-/RXR gamma -/- mutant mice are viable. Proc Natl Acad Sci U S A. 1996 Aug 20;93(17):9010–4. doi: 10.1073/pnas.93.17.9010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Chiang MY, Misner D, Kempermann G, Schikorski T, Giguère V, Sucov HM, Gage FH, Stevens CF, Evans RM. An essential role for retinoid receptors RARbeta and RXRgamma in long-term potentiation and depression. Neuron. 1998 Dec;21(6):1353–61. doi: 10.1016/s0896-6273(00)80654-6. [DOI] [PubMed] [Google Scholar]
- 134.Krzyzosiak A, Szyszka-Niagolov M, Wietrzych M, Gobaille S, Muramatsu S, Krezel W. Retinoid x receptor gamma control of affective behaviors involves dopaminergic signaling in mice. Neuron. 2010 Jun 24;66(6):908–20. doi: 10.1016/j.neuron.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 135.McNamara P, Seo SB, Rudic RD, Sehgal A, Chakravarti D, FitzGerald GA. Regulation of CLOCK and MOP4 by nuclear hormone receptors in the vasculature: a humoral mechanism to reset a peripheral clock. Cell. 2001 Jun 29;105(7):877–89. doi: 10.1016/s0092-8674(01)00401-9. [DOI] [PubMed] [Google Scholar]
- 136.Yang X, Guo M, Wan YJ. Deregulation of growth factor, circadian clock, and cell cycle signaling in regenerating hepatocyte RXRalpha-deficient mouse livers. Am J Pathol. 2010 Feb;176(2):733–43. doi: 10.2353/ajpath.2010.090524. Epub 2009 Dec 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Fisk GJ, Thummel CS. The DHR78 nuclear receptor is required for ecdysteroid signaling during the onset of Drosophila metamorphosis. Cell. 1998 May 15;93(4):543–55. doi: 10.1016/s0092-8674(00)81184-8. [DOI] [PubMed] [Google Scholar]
- 138.Baker KD, Beckstead RB, Mangelsdorf DJ, Thummel CS. Functional interactions between the Moses corepressor and DHR78 nuclear receptor regulate growth in Drosophila. Genes Dev. 2007 Feb 15;21(4):450–64. doi: 10.1101/gad.1519007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Chang C, Kokontis J. Identification of a new member of the steroid receptor super-family by cloning and sequence analysis. Biochem Biophys Res Commun. 1988 Sep 15;155(2):971–7. doi: 10.1016/s0006-291x(88)80591-6. [DOI] [PubMed] [Google Scholar]
- 140.Shyr CR, Collins LL, Mu XM, Platt KA, Chang C. Spermatogenesis and testis development are normal in mice lacking testicular orphan nuclear receptor 2. Mol Cell Biol. 2002 Jul;22(13):4661–6. doi: 10.1128/MCB.22.13.4661-4666.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Olivares AM, Han Y, Soto D, Flattery K, Marini J, Molemma N, Haider A, Escher P, DeAngelis MM, Haider NB. The nuclear hormone receptor gene Nr2c1 (Tr2) is a critical regulator of early retina cell patterning. Dev Biol. 2017 May 25; doi: 10.1016/j.ydbio.2017.05.021. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 142.Collins LL, Lee YF, Heinlein CA, Liu NC, Chen YT, Shyr CR, Meshul CK, Uno H, Platt KA, Chang C. Growth retardation and abnormal maternal behavior in mice lacking testicular orphan nuclear receptor 4. Proc Natl Acad Sci USA. 2004;101:15058–15063. doi: 10.1073/pnas.0405700101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Chen YT, Collins LL, Uno H, Chang C. Deficits in motor coordination with aberrant cerebellar development in mice lacking testicular orphan nuclear receptor 4. Mol Cell Biol. 2005 Apr;25(7):2722–32. doi: 10.1128/MCB.25.7.2722-2732.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Shyr CR, Kang HY, Tsai MY, Liu NC, Ku PY, Huang KE, Chang C. Roles of testicular orphan nuclear receptors 2 and 4 in early embryonic development and embryonic stem cells. Endocrinology. 2009 May;150(5):2454–62. doi: 10.1210/en.2008-1165. Epub 2009 Jan 8. [DOI] [PubMed] [Google Scholar]
- 145.Lin SJ, Yang DR, Yang G, Lin CY, Chang HC, Li G, Chang C. TR2 and TR4 OrphanNuclear Receptors: An Overview. Curr Top Dev Biol. 2017;125:357–373. doi: 10.1016/bs.ctdb.2017.02.002. Epub 2017 Apr 5. [DOI] [PubMed] [Google Scholar]
- 146.Nüsslein-Volhard C, Wieschaus E, Kluding H. Mutations affecting the pattern of the larval cuticle inDrosophila melanogaster : I. Zygotic loci on the second chromosome. Wilehm Roux Arch Dev Biol. 1984 Sep;193(5):267–282. doi: 10.1007/BF00848156. [DOI] [PubMed] [Google Scholar]
- 147.Mahoney PA, Lengyel JA. The zygotic segmentation mutant tailless alters the blastoderm fate map of the Drosophila embryo. Dev Biol. 1987 Aug;122(2):464–70. doi: 10.1016/0012-1606(87)90310-1. [DOI] [PubMed] [Google Scholar]
- 148.Gui H, Li ML, Tsai CC. A tale of tailless. Dev Neurosci. 2011;33(1):1–13. doi: 10.1159/000321585. Epub 2010 Dec 2. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Younossi-Hartenstein A, Green P, Liaw GJ, Rudolph K, Lengyel J, Hartenstein V. Control of early neurogenesis of the Drosophila brain by the head gap genes tll, otd, ems, and btd. Dev Biol. 1997 Feb 15;182(2):270–83. doi: 10.1006/dbio.1996.8475. [DOI] [PubMed] [Google Scholar]
- 150.Davis SM, Thomas AL, Nomie KJ, Huang L, Dierick HA. Tailless and Atrophin control Drosophila aggression by regulating neuropeptide signalling in the pars intercerebralis. Nat Commun. 2014;5:3177. doi: 10.1038/ncomms4177. [DOI] [PubMed] [Google Scholar]
- 151.Sarin S, Antonio C, Tursun B, Hobert O. The C. elegans Tailless/TLX transcription factor nhr-67 controls neuronal identity and left/right asymmetric fate diversification. Development. 2009 Sep;136(17):2933–44. doi: 10.1242/dev.040204. Epub 2009 Jul 29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Verghese E, Schocken J, Jacob S, Wimer AM, Royce R, Nesmith JE, Baer GM, Clever S, McCain E, Lakowski B, Wightman B. The tailless ortholog nhr-67 functions in the development of the C. elegans ventral uterus. Dev Biol. 2011 Aug 15;356(2):516–28. doi: 10.1016/j.ydbio.2011.06.007. Epub 2011 Jun 28. [DOI] [PubMed] [Google Scholar]
- 153.Matus DQ, Lohmer LL, Kelley LC, Schindler AJ, Kohrman AQ, Barkoulas M, Zhang W, Chi Q, Sherwood DR. Invasive Cell Fate Requires G1 Cell-Cycle Arrest and Histone Deacetylase-Mediated Changes in Gene Expression. Dev Cell. 2015 Oct 26;35(2):162–74. doi: 10.1016/j.devcel.2015.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Fernandes JS, Sternberg PW. The tailless ortholog nhr-67 regulates patterning of gene expression and morphogenesis in the C. elegans vulva. PLoS Genet. 2007 Apr 27;3(4):e69. doi: 10.1371/journal.pgen.0030069. Epub 2007 Mar 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Kato M, Sternberg PW. The C. elegans tailless/Tlx homolog nhr-67 regulates a stage-specific program of linker cell migration in male gonadogenesis. Development. 2009 Dec;136(23):3907–15. doi: 10.1242/dev.035477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Zacharias AL, Walton T, Preston E, Murray JI. Quantitative Differences in Nuclear β-catenin and TCF Pattern Embryonic Cells in C. elegans. PLoS Genet. 2015 Oct 21;11(10):e1005585. doi: 10.1371/journal.pgen.1005585. eCollection 2015 Oct. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Yu RT, McKeown M, Evans RM, Umesono K. Relationship between Drosophila gap gene tailless and a vertebrate nuclear receptor Tlx. Nature. 1994 Aug 4;370(6488):375–9. doi: 10.1038/370375a0. [DOI] [PubMed] [Google Scholar]
- 158.Monaghan AP, Grau E, Bock D, Schütz G. The mouse homolog of the orphan nuclear receptor tailless is expressed in the developing forebrain. Development. 1995 Mar;121(3):839–53. doi: 10.1242/dev.121.3.839. [DOI] [PubMed] [Google Scholar]
- 159.Stenman JM, Wang B, Campbell K. Tlx controls proliferation and patterning of lateral telencephalic progenitor domains. J Neurosci. 2003 Nov 19;23(33):10568–76. doi: 10.1523/JNEUROSCI.23-33-10568.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Monaghan AP, Bock D, Gass P, Schwäger A, Wolfer DP, Lipp HP, Schütz G. Defective limbic system in mice lacking the tailless gene. Nature. 1997 Dec 4;390(6659):515–7. doi: 10.1038/37364. [DOI] [PubMed] [Google Scholar]
- 161.Thomas AL, Davis SM, Dierick HA. Of Fighting Flies, Mice, and Men: Are Some of the Molecular and Neuronal Mechanisms of Aggression Universal in the Animal Kingdom? PLoS Genet. 2015 Aug 27;11(8):e1005416. doi: 10.1371/journal.pgen.1005416. eCollection 2015 Aug. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Zhang CL, Zou Y, Yu RT, Gage FH, Evans RM. Nuclear receptor TLX prevents retinal dystrophy and recruits the corepressor atrophin1. Genes Dev. 2006 May 15;20(10):1308–20. doi: 10.1101/gad.1413606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Islam MM, Zhang CL. TLX: A master regulator for neural stem cell maintenance and neurogenesis. Biochim Biophys Acta. 2015 Feb;1849(2):210–6. doi: 10.1016/j.bbagrm.2014.06.001. Epub 2014 Jun 13. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Qu Q, Sun G, Li W, Yang S, Ye P, Zhao C, Yu RT, Gage FH, Evans RM, Shi Y. Orphan nuclear receptor TLX activates Wnt/beta-catenin signalling to stimulate neural stem cell proliferation and self-renewal. Nat Cell Biol. 2010 Jan;12(1):31–40. doi: 10.1038/ncb2001. sup pp 1-9. Epub 2009 Dec 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Roy K, Kuznicki K, Wu Q, Sun Z, Bock D, Schutz G, Vranich N, Monaghan AP. The Tlx gene regulates the timing of neurogenesis in the cortex. J Neurosci. 2004 Sep 22;24(38):8333–45. doi: 10.1523/JNEUROSCI.1148-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Land PW, Monaghan AP. Abnormal development of zinc-containing cortical circuits in the absence of the transcription factor Tailless. Brain Res Dev Brain Res. 2005 Aug 8;158(1-2):97–101. doi: 10.1016/j.devbrainres.2005.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Sung C, Wong LE, Chang Sen LQ, Nguyen E, Lazaga N, Ganzer G, McNabb SL, Robinow S. The unfulfilled/DHR51 gene of Drosophila melanogaster modulates wing expansion and fertility. Dev Dyn. 2009 Jan;238(1):171–82. doi: 10.1002/dvdy.21817. [DOI] [PubMed] [Google Scholar]
- 168.Bates KE, Molnar J, Robinow S. The unfulfilled gene and nervous system development in Drosophila. Biochim Biophys Acta. 2015 Feb;1849(2):217–23. doi: 10.1016/j.bbagrm.2014.06.013. Epub 2014 Jun 19. Review. [DOI] [PubMed] [Google Scholar]
- 169.Lin S, Huang Y, Lee T. Nuclear receptor unfulfilled regulates axonal guidance and cell identity of Drosophila mushroom body neurons. PLoS One. 2009 Dec 22;4(12):e8392. doi: 10.1371/journal.pone.0008392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Beuchle D, Jaumouillé E, Nagoshi E. The nuclear receptor unfulfilled is required for free-running clocks in Drosophila pacemaker neurons. Curr Biol. 2012 Jul 10;22(13):1221–7. doi: 10.1016/j.cub.2012.04.052. Epub 2012 May 31. [DOI] [PubMed] [Google Scholar]
- 171.Yaniv SP, Issman-Zecharya N, Oren-Suissa M, Podbilewicz B, Schuldiner O. Axon regrowth during development and regeneration following injury share molecular mechanisms. Curr Biol. 2012 Oct 9;22(19):1774–82. doi: 10.1016/j.cub.2012.07.044. Epub 2012 Aug 23. [DOI] [PubMed] [Google Scholar]
- 172.Rabinovich D, Yaniv SP, Alyagor I, Schuldiner O. Nitric Oxide as a Switching Mechanism between Axon Degeneration and Regrowth during Developmental Remodeling. Cell. 2016 Jan 14;164(1-2):170–182. doi: 10.1016/j.cell.2015.11.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Wightman B, Baran R, Garriga G. Genes that guide growth cones along the C. elegans ventral nerve cord. Development. 1997 Jul;124(13):2571–80. doi: 10.1242/dev.124.13.2571. [DOI] [PubMed] [Google Scholar]
- 174.Much JW, Slade DJ, Klampert K, Garriga G, Wightman B. The fax-1 nuclear hormone receptor regulates axon pathfinding and neurotransmitter expression. Development. 2000 Feb;127(4):703–12. doi: 10.1242/dev.127.4.703. [DOI] [PubMed] [Google Scholar]
- 175.Wightman B, Ebert B, Carmean N, Weber K, Clever S. The C. elegans nuclear receptor gene fax-1 and homeobox gene unc-42 coordinate interneuron identity by regulating the expression of glutamate receptor subunits and other neuron-specific genes. Dev Biol. 2005 Nov 1;287(1):74–85. doi: 10.1016/j.ydbio.2005.08.032. Epub 2005 Sep 23. [DOI] [PubMed] [Google Scholar]
- 176.Kobayashi M, Takezawa S, Hara K, Yu RT, Umesono Y, Agata K, Taniwaki M, Yasuda K, Umesono K. Identification of a photoreceptor cell-specific nuclear receptor. Proc Natl Acad Sci U S A. 1999 Apr 27;96(9):4814–9. doi: 10.1073/pnas.96.9.4814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Haider NB, Demarco P, Nystuen AM, Huang X, Smith RS, McCall MA, Naggert JK, Nishina PM. The transcription factor Nr2e3 functions in retinal progenitors to suppress cone cell generation. Vis Neurosci. 2006 Nov-Dec;23(6):917–29. doi: 10.1017/S095252380623027X. [DOI] [PubMed] [Google Scholar]
- 178.Chen J, Rattner A, Nathans J. The rod photoreceptor-specific nuclear receptor Nr2e3 represses transcription of multiple cone-specific genes. J Neurosci. 2005 Jan 5;25(1):118–29. doi: 10.1523/JNEUROSCI.3571-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Akhmedov NB, Piriev NI, Chang B, Rapoport AL, Hawes NL, Nishina PM, Nusinowitz S, Heckenlively JR, Roderick TH, Kozak CA, Danciger M, Davisson MT, Farber DB. A deletion in a photoreceptor-specific nuclear receptor mRNA causes retinal degeneration in the rd7 mouse. Proc Natl Acad Sci U S A. 2000 May 9;97(10):5551–6. doi: 10.1073/pnas.97.10.5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Laranjeiro R, Whitmore D. Transcription factors involved in retinogenesis are co-opted by the circadian clock following photoreceptor differentiation. Development. 2014 Jul;141(13):2644–56. doi: 10.1242/dev.104380. Epub 2014 Jun 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Haider NB, Jacobson SG, Cideciyan AV, Swiderski R, Streb LM, Searby C, Beck G, Hockey R, Hanna DB, Gorman S, Duhl D, Carmi R, Bennett J, Weleber RG, Fishman GA, Wright AF, Stone EM, Sheffield VC. Mutation of a nuclear receptor gene, NR2E3, causes enhanced S cone syndrome, a disorder of retinal cell fate. Nat Genet. 2000 Feb;24(2):127–31. doi: 10.1038/72777. [DOI] [PubMed] [Google Scholar]
- 182.Schüpbach T, Wieschaus E. Female sterile mutations on the second chromosome of Drosophila melanogaster. I. Maternal effect mutations. Genetics. 1989 Jan;121(1):101–17. doi: 10.1093/genetics/121.1.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Finley KD, Edeen PT, Foss M, Gross E, Ghbeish N, Palmer RH, Taylor BJ, McKeown M. Dissatisfaction encodes a tailless-like nuclear receptor expressed in a subset of CNS neurons controlling Drosophila sexual behavior. Neuron. 1998 Dec;21(6):1363–74. doi: 10.1016/s0896-6273(00)80655-8. [DOI] [PubMed] [Google Scholar]
- 184.Weber KP, Alvaro CG, Baer GM, Reinert K, Cheng G, Clever S, Wightman B. Analysis of C. elegans NR2E nuclear receptors defines three conserved clades and ligand-independent functions. BMC Evol Biol. 2012 Jun 12;12:81. doi: 10.1186/1471-2148-12-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Mlodzik M, Hiromi Y, Weber U, Goodman CS, Rubin GM. The Drosophila seven-up gene, a member of the steroid receptor gene superfamily, controls photoreceptor cell fates. Cell. 1990 Jan 26;60(2):211–24. doi: 10.1016/0092-8674(90)90737-y. [DOI] [PubMed] [Google Scholar]
- 186.Hiromi Y, Mlodzik M, West SR, Rubin GM, Goodman CS. Ectopic expression of seven-up causes cell fate changes during ommatidial assembly. Development. 1993 Aug;118(4):1123–35. doi: 10.1242/dev.118.4.1123. [DOI] [PubMed] [Google Scholar]
- 187.Hoshizaki DK, Blackburn T, Price C, Ghosh M, Miles K, Ragucci M, Sweis R. Embryonic fat-cell lineage in Drosophila melanogaster. Development. 1994 Sep;120(9):2489–99. doi: 10.1242/dev.120.9.2489. [DOI] [PubMed] [Google Scholar]
- 188.Kerber B, Fellert S, Hoch M. Seven-up, the Drosophila homolog of the COUP-TF orphan receptors, controls cell proliferation in the insect kidney. Genes Dev. 1998 Jun 15;12(12):1781–6. doi: 10.1101/gad.12.12.1781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Lo PC, Frasch M. A role for the COUP-TF-related gene seven-up in the diversification of cardioblast identities in the dorsal vessel of Drosophila. Mech Dev. 2001 Jun;104(1-2):49–60. doi: 10.1016/s0925-4773(01)00361-6. [DOI] [PubMed] [Google Scholar]
- 190.Zelhof AC, Yao TP, Chen JD, Evans RM, McKeown M. Seven-up inhibits ultraspiracle-based signaling pathways in vitro and in vivo. Mol Cell Biol. 1995 Dec;15(12):6736–45. doi: 10.1128/mcb.15.12.6736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Zhou HM, Walthall WW. UNC-55, an orphan nuclear hormone receptor, orchestrates synaptic specificity among two classes of motor neurons in Caenorhabditis elegans. J Neurosci. 1998 Dec 15;18(24):10438–44. doi: 10.1523/JNEUROSCI.18-24-10438.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Shan G, Walthall WW. Copulation in C. elegans males requires a nuclear hormone receptor. Dev Biol. 2008 Oct 1;322(1):11–20. doi: 10.1016/j.ydbio.2008.06.034. Epub 2008 Jul 4. [DOI] [PubMed] [Google Scholar]
- 193.Pastorcic M, Bagchi MK, Tsai SY, Tsai MJ, O’Malley BW. Multiple protein binding sites within the ovalbumin gene 5’-flanking region: isolation and characterization of sequence-specific binding proteins. Nucleic Acids Res. 1989 Aug 25;17(16):6693–711. doi: 10.1093/nar/17.16.6693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Pereira FA, Tsai MJ, Tsai SY. COUP-TF orphan nuclear receptors in development and differentiation. Cell Mol Life Sci. 2000 Sep;57(10):1388–98. doi: 10.1007/PL00000624. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Wang LH, Ing NH, Tsai SY, O’Malley BW, Tsai MJ. The COUP-TFs compose a family of functionally related transcription factors. Gene Expr. 1991;1(3):207–16. [PMC free article] [PubMed] [Google Scholar]
- 196.Zhou C, Qiu Y, Pereira FA, Crair MC, Tsai SY, Tsai MJ. The nuclear orphan receptor COUP-TFI is required for differentiation of subplate neurons and guidance of thalamocortical axons. Neuron. 1999 Dec;24(4):847–59. doi: 10.1016/s0896-6273(00)81032-6. [DOI] [PubMed] [Google Scholar]
- 197.Misra S, Pandey AK, Gupta S, Kumar A, Khanna P, Shankar J, Ravi Ram K. Estrogen related receptor is required for the testicular development and for the normal sperm axoneme/mitochondrial derivatives in Drosophila males. Sci Rep. 2017 Jan 17;7:40372. doi: 10.1038/srep40372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Luo J, Sladek R, Carrier J, Bader J-A, Richard D, Giguere V. Reduced fat mass in mice lacking orphan nuclear receptor estrogen-related receptor-alpha. Molec Cell Biol. 2003;23:7947–7956. doi: 10.1128/MCB.23.22.7947-7956.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Luo J, Sladek R, Bader JA, Matthyssen A, Rossant J, Giguère V. Placental abnormalities in mouse embryos lacking the orphan nuclear receptor ERR-beta. Nature. 1997 Aug 21;388(6644):778–82. doi: 10.1038/42022. [DOI] [PubMed] [Google Scholar]
- 200.Alaynick WA, Kondo RP, Xie W, He W, Dufour CR, Downes M, Jonker JW, Giles W, Naviaux RK, Giguere V, Evans RM. ERR-gamma directs and maintains the transition to oxidative metabolism in the postnatal heart. Cell Metab. 2007;6:13–24. doi: 10.1016/j.cmet.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 201.Wang T, McDonald C, Petrenko NB, Leblanc M, Wang T, Giguere V, Evans RM, Patel VV, Pei L. Estrogen-related receptor α (ERRα) and ERRγ are essential coordinators of cardiac metabolism and function. Mol Cell Biol. 2015 Apr;35(7):1281–98. doi: 10.1128/MCB.01156-14. Epub 2015 Jan 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Berry R, Harewood L, Pei L, Fisher M, Brownstein D, Ross A, Alaynick WA, Moss J, Hastie ND, Hohenstein P, Davies JA, Evans RM, FitzPatrick DR. Esrrg functions in early branch generation of the ureteric bud and is essential for normal development of the renal papilla. Hum Molec Genet. 2011;20:917–926. doi: 10.1093/hmg/ddq530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Pei L, Mu Y, Leblanc M, Alaynick W, Barish GD, Pankratz M, Tseng TW, Kaufman S, Liddle C, Yu RT, Downes M, Pfaff SL, Auwerx J, Gage FH, Evans RM. Dependence of hippocampal function on ERRγ-regulated mitochondrial metabolism. Cell Metab. 2015 Apr 7;21(4):628–36. doi: 10.1016/j.cmet.2015.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Baker KD, Shewchuk LM, Kozlova T, Makishima M, Hassell A, Wisely B, Caravella JA, Lambert MH, Reinking JL, Krause H, Thummel CS, Willson TM, Mangelsdorf DJ. The Drosophila orphan nuclear receptor DHR38 mediates an atypical ecdysteroid signaling pathway. Cell. 2003 Jun 13;113(6):731–42. doi: 10.1016/s0092-8674(03)00420-3. [DOI] [PubMed] [Google Scholar]
- 205.Sutherland JD, Kozlova T, Tzertzinis G, Kafatos FC. Drosophila hormone receptor 38: a second partner for Drosophila USP suggests an unexpected role for nuclear receptors of the nerve growth factor-induced protein B type. Proc Natl Acad Sci U S A. 1995 Aug 15;92(17):7966–70. doi: 10.1073/pnas.92.17.7966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Kozlova T, Pokholkova GV, Tzertzinis G, Sutherland JD, Zhimulev IF, Kafatos FC. Drosophila hormone receptor 38 functions in metamorphosis: a role in adult cuticle formation. Genetics. 1998 Jul;149(3):1465–75. doi: 10.1093/genetics/149.3.1465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Kozlova T, Lam G, Thummel CS. Drosophila DHR38 nuclear receptor is required for adult cuticle integrity at eclosion. Dev Dyn. 2009 Mar;238(3):701–7. doi: 10.1002/dvdy.21860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Mezan S, Feuz JD, Deplancke B, Kadener S. PDF Signaling Is an Integral Part of the Drosophila Circadian Molecular Oscillator. Cell Rep. 2016 Oct 11;17(3):708–719. doi: 10.1016/j.celrep.2016.09.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Gissendanner CR, Kelley K, Nguyen TQ, Hoener MC, Sluder AE, Maina CV. The Caenorhabditis elegans NR4A nuclear receptor is required for spermatheca morphogenesis. Dev Biol. 2008 Jan 15;313(2):767–86. doi: 10.1016/j.ydbio.2007.11.014. Epub 2007 Nov 22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Gissendanner CR, Cardin D, Dubose CJ, El Sayed M, Harmson JS, Praslicka B, Rowan BG. C. elegans nuclear receptor NHR-6 functionally interacts with the jun-1 transcription factor during spermatheca development. Genesis. 2014 Jan;52(1):29–38. doi: 10.1002/dvg.22723. Epub 2013 Nov 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Praslicka B, Gissendanner CR. The C. elegans NR4A nuclear receptor gene nhr-6 promotes cell cycle progression in the spermatheca lineage. Dev Dyn. 2015 Mar;244(3):417–30. doi: 10.1002/dvdy.24244. Epub 2015 Jan 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Maxwell MA, Muscat GE. The NR4A subgroup: immediate early response genes with pleiotropic physiological roles. Nucl Recept Signal. 2006;4:e002. doi: 10.1621/nrs.04002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Ranhotra HS. The NR4A orphan nuclear receptors: mediators in metabolism and diseases. J Recept Signal Transduct Res. 2015 Apr;35(2):184–8. doi: 10.3109/10799893.2014.948555. Epub 2014 Aug 4. Review. [DOI] [PubMed] [Google Scholar]
- 214.Beard JA, Tenga A, Chen T. The interplay of NR4A receptors and the oncogene-tumor suppressor networks in cancer. Cell Signal. 2015 Feb;27(2):257–66. doi: 10.1016/j.cellsig.2014.11.009. Epub 2014 Nov 15. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Safe S, Jin UH, Hedrick E, Reeder A, Lee SO. Minireview: role of orphan nuclear receptors in cancer and potential as drug targets. Mol Endocrinol. 2014 Feb;28(2):157–72. doi: 10.1210/me.2013-1291. Epub 2013 Dec 2. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Mullican SE, Zhang S, Konopleva M, Ruvolo V, Andreeff M, Milbrandt J, Conneely OM. Abrogation of nuclear receptors Nr4a3 and Nr4a1 leads to development of acute myeloid leukemia. Nat Med. 2007 Jun;13(6):730–5. doi: 10.1038/nm1579. Epub 2007 May 21. [DOI] [PubMed] [Google Scholar]
- 217.Jiang H, Xiang M. Subtype specification of GABAergic amacrine cells by the orphan nuclear receptor Nr4a2/Nurr1. J Neurosci. 2009 Aug 19;29(33):10449–59. doi: 10.1523/JNEUROSCI.3048-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Martin LJ, Tremblay JJ. Nuclear receptors in Leydig cell gene expression and function. Biol Reprod. 2010 Jul;83(1):3–14. doi: 10.1095/biolreprod.110.083824. Epub 2010 Apr 7. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Ueda H, Sonoda S, Brown JL, Scott MP, Wu C. A sequence-specific DNA-binding protein that activates fushi tarazu segmentation gene expression. Genes Dev. 1990 Apr;4(4):624–35. doi: 10.1101/gad.4.4.624. [DOI] [PubMed] [Google Scholar]
- 220.Lavorgna G, Ueda H, Clos J, Wu C. FTZ-F1, a steroid hormone receptor-like protein implicated in the activation of fushi tarazu. Science. 1991 May 10;252(5007):848–51. doi: 10.1126/science.1709303. [DOI] [PubMed] [Google Scholar]
- 221.Florence B, Guichet A, Ephrussi A, Laughon A. Ftz-F1 is a cofactor in Ftz activation of the Drosophila engrailed gene. Development. 1997 Feb;124(4):839–47. doi: 10.1242/dev.124.4.839. [DOI] [PubMed] [Google Scholar]
- 222.Bowler T, Kosman D, Licht JD, Pick L. Computational identification of Ftz/Ftz-F1 downstream target genes. Dev Biol. 2006 Nov 1;299(1):78–90. doi: 10.1016/j.ydbio.2006.07.007. Epub 2006 Jul 12. [DOI] [PubMed] [Google Scholar]
- 223.Yussa M, Löhr U, Su K, Pick L. The nuclear receptor Ftz-F1 and homeodomain protein Ftz interact through evolutionarily conserved protein domains. Mech Dev. 2001 Sep;107(1-2):39–53. doi: 10.1016/s0925-4773(01)00448-8. [DOI] [PubMed] [Google Scholar]
- 224.Martin D. Functions of nuclear receptors in insect development. In: Bunce CM, Campbell MJ, editors. Nuclear Receptors. Springer; [Google Scholar]
- 225.Yamada M, Murata T, Hirose S, Lavorgna G, Suzuki E, Ueda H. Temporally restricted expression of transcription factor betaFTZ-F1: Significance for embryogenesis, molting and metamorphosis in Drosophila melanogaster. Development. 2000;127:5083–5092. doi: 10.1242/dev.127.23.5083. [DOI] [PubMed] [Google Scholar]
- 226.Broadus J, McCabe JR, Endrizzi B, Thummel CS, Woodard CT. The Drosophila beta FTZ-F1 orphan nuclear receptor provides competence for stage-specific responses to the steroid hormone ecdysone. Mol Cell. 1999;3:143–149. doi: 10.1016/s1097-2765(00)80305-6. [DOI] [PubMed] [Google Scholar]
- 227.Awasaki T, Lee T. Orphan nuclear receptors control neuronal remodeling during fly metamorphosis. Nat Neurosci. 2011 Jan;14(1):6–7. doi: 10.1038/nn0111-6. [DOI] [PubMed] [Google Scholar]
- 228.Boulanger A, Clouet-Redt C, Farge M, Flandre A, Guignard T, Fernando C, Juge F, Dura JM. ftz-f1 and Hr39 opposing roles on EcR expression during Drosophila mushroom body neuron remodeling. Nat Neurosci. 2011 Jan;14(1):37–44. doi: 10.1038/nn.2700. Epub 2010 Dec 5. [DOI] [PubMed] [Google Scholar]
- 229.Gissendanner CR, Sluder AE. nhr-25, the Caenorhabditis elegans ortholog of ftz-f1, is required for epidermal and somatic gonad development. Dev Biol. 2000 May 1;221(1):259–72. doi: 10.1006/dbio.2000.9679. [DOI] [PubMed] [Google Scholar]
- 230.Asahina M, Valenta T, Silhankova M, Korinek V, Jindra M. Crosstalk between a nuclear receptor and beta-catenin signaling decides cell fates in the C. elegans somatic gonad. Dev Cell. 2006 Aug;11(2):203–11. doi: 10.1016/j.devcel.2006.06.003. [DOI] [PubMed] [Google Scholar]
- 231.Hada K, Asahina M, Hasegawa H, Kanaho Y, Slack FJ, Niwa R. The nuclear receptor gene nhr-25 plays multiple roles in the Caenorhabditis elegans heterochronic gene network to control the larva-to-adult transition. Dev Biol. 2010 Aug 15;344(2):1100–9. doi: 10.1016/j.ydbio.2010.05.508. Epub 2010 Jun 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Asahina M, Ishihara T, Jindra M, Kohara Y, Katsura I, Hirose S. The conserved nuclear receptor Ftz-F1 is required for embryogenesis, moulting and reproduction in Caenorhabditis elegans. Genes Cells. 2000 Sep;5(9):711–23. doi: 10.1046/j.1365-2443.2000.00361.x. [DOI] [PubMed] [Google Scholar]
- 233.Chen Z, Eastburn DJ, Han M. The Caenorhabditis elegans nuclear receptor gene nhr-25 regulates epidermal cell development. Mol Cell Biol. 2004 Sep;24(17):7345–58. doi: 10.1128/MCB.24.17.7345-7358.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Bowler T, Kosman D, Licht JD, Pick L. Computational identification of Ftz/Ftz-F1 downstream target genes. Dev Biol. 2006 Nov 1;299(1):78–90. doi: 10.1016/j.ydbio.2006.07.007. Epub 2006 Jul 12. [DOI] [PubMed] [Google Scholar]
- 235.Morohashi K, Honda S, Inomata Y, Handa H, Omura T. A common transacting factor, Ad4-binding protein, to the promoters of steroidogenic P-450s. J Biol Chem. 1992;267:17913–17919. [PubMed] [Google Scholar]
- 236.Lala DS, Rice DA, Parker KL. Steroidogenic factor I, a key regulator of steroidogenic enzyme expression, is the mouse homolog of fushi tarazu-factor I. Mol Endocrinol. 1992;6:1249–1258. doi: 10.1210/mend.6.8.1406703. [DOI] [PubMed] [Google Scholar]
- 237.Hoivik EA, Lewis AE, Aumo L, Bakke M. Molecular aspects of steroidogenic factor 1 (SF-1) Mol Cell Endocrinol. 2010 Feb 5;315(1-2):27–39. doi: 10.1016/j.mce.2009.07.003. Epub 2009 Jul 16. Review. [DOI] [PubMed] [Google Scholar]
- 238.Luo X, Ikeda Y, Parker KL. A cell-specific nuclear receptor is essential for adrenal and gonadal development and sexual differentiation. Cell. 1994;77:481–490. doi: 10.1016/0092-8674(94)90211-9. [DOI] [PubMed] [Google Scholar]
- 239.Jeyasuria P, Ikeda Y, Jamin SP, Zhao L, De Rooij DG, Themmen AP, Behringer RR, Parker KL. Cell-specific knockout of steroidogenic factor 1 reveals its essential roles in gonadal function. Mol Endocrinol. 2004;18:1610–1619. doi: 10.1210/me.2003-0404. [DOI] [PubMed] [Google Scholar]
- 240.Sreenivasan R, Jiang J, Wang X, Bártfai R, Kwan HY, Christoffels A, Orbán L. Gonad differentiation in zebrafish is regulated by the canonical Wnt signaling pathway. Biol Reprod. 2014 Feb 27;90(2):45. doi: 10.1095/biolreprod.113.110874. Print 2014 Feb. [DOI] [PubMed] [Google Scholar]
- 241.Zhao L, Bakke M, Parker KL. Pituitary-specific knockout of steroidogenic factor 1. Mol Cell Endocrinol. 2001 Dec 20;185(1-2):27–32. doi: 10.1016/s0303-7207(01)00621-9. Review. [DOI] [PubMed] [Google Scholar]
- 242.Tran PV, Lee MB, Marín O, Xu B, Jones KR, Reichardt LF, Rubenstein JR, Ingraham HA. Requirement of the orphan nuclear receptor SF-1 in terminal differentiation of ventromedial hypothalamic neurons. Mol Cell Neurosci. 2003 Apr;22(4):441–53. doi: 10.1016/S1044-7431(03)00027-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Fayard E, Auwerx J, Schoonjans K. LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis. Trends Cell Biol. 2004 May;14(5):250–60. doi: 10.1016/j.tcb.2004.03.008. Review. [DOI] [PubMed] [Google Scholar]
- 244.Gu P, Goodwin B, Chung AC, Xu X, Wheeler DA, Price RR, Galardi C, Peng L, Latour AM, Koller BH, Gossen J, Kliewer SA, Cooney AJ. Orphan nuclear receptor LRH-1 is required to maintain Oct4 expression at the epiblast stage of embryonic development. Mol Cell Biol. 2005 May;25(9):3492–505. doi: 10.1128/MCB.25.9.3492-3505.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Heng JC, Feng B, Han J, Jiang J, Kraus P, Ng JH, Orlov YL, Huss M, Yang L, Lufkin T, Lim B, Ng HH. The nuclear receptor Nr5a2 can replace Oct4 in the reprogramming of murine somatic cells to pluripotent cells. Cell Stem Cell. 2010 Feb 5;6(2):167–74. doi: 10.1016/j.stem.2009.12.009. Epub 2010 Jan 21. [DOI] [PubMed] [Google Scholar]
- 246.Wagner RT, Xu X, Yi F, Merrill BJ, Cooney AJ. Canonical Wnt/β-catenin regulation of liver receptor homolog-1 mediates pluripotency gene expression. Stem Cells. 2010 Oct;28(10):1794–804. doi: 10.1002/stem.502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Yumoto F, Nguyen P, Sablin EP, Baxter JD, Webb P, Fletterick RJ. Structural basis of coactivation of liver receptor homolog-1 by β-catenin. Proc Natl Acad Sci U S A. 2012 Jan 3;109(1):143–8. doi: 10.1073/pnas.1117036108. Epub 2011 Dec 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Hale MA, Swift GH, Hoang CQ, Deering TG, Masui T, Lee YK, Xue J, MacDonald RJ. The nuclear hormone receptor family member NR5A2 controls aspects of multipotent progenitor cell formation and acinar differentiation during pancreatic organogenesis. Development. 2014 Aug;141(16):3123–33. doi: 10.1242/dev.109405. Epub 2014 Jul 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Nissim S, Weeks O, Talbot JC, Hedgepeth JW, Wucherpfennig J, Schatzman-Bone S, Swinburne I, Cortes M, Alexa K, Megason S, North TE, Amacher SL, Goessling W. Iterative use of nuclear receptor Nr5a2 regulates multiple stages of liver and pancreas development. Dev Biol. 2016 Oct 1;418(1):108–23. doi: 10.1016/j.ydbio.2016.07.019. Epub 2016 Jul 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Zhai G, Song J, Shu T, Yan J, Jin X, He J, Yin Z. LRH-1 senses signaling from phosphatidylcholine to regulate the expansion growth of digestive organs via synergy with Wnt/β-catenin signaling in zebrafish. J Genet Genomics. 2017 Apr 27; doi: 10.1016/j.jgg.2017.03.006. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 251.Forman BM. Are those phospholipids in your pocket? Cell Metab. 2005 Mar;1(3):153–5. doi: 10.1016/j.cmet.2005.02.006. Review. [DOI] [PubMed] [Google Scholar]
- 252.Sun J, Spradling AC. NR5A nuclear receptor Hr39 controls three-cell secretory unit formation in Drosophila female reproductive glands. Curr Biol. 2012 May 22;22(10):862–71. doi: 10.1016/j.cub.2012.03.059. Epub 2012 May 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Ou Q, Magico A, King-Jones K. Nuclear receptor DHR4 controls the timing of steroid hormone pulses during Drosophila development. PLoS Biol. 2011 Sep;9(9):e1001160. doi: 10.1371/journal.pbio.1001160. Epub 2011 Sep 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Kasuga H, Fukuyama M, Kitazawa A, Kontani K, Katada T. The microRNA miR-235 couples blast-cell quiescence to the nutritional state. Nature. 2013 May 23;497(7450):503–6. doi: 10.1038/nature12117. Epub 2013 May 5. [DOI] [PubMed] [Google Scholar]
- 255.Fuhrmann G, Chung AC, Jackson KJ, Hummelke G, Baniahmad A, Sutter J, Sylvester I, Schöler HR, Cooney AJ. Mouse germline restriction of Oct4 expression by germ cell nuclear factor. Dev Cell. 2001 Sep;1(3):377–87. doi: 10.1016/s1534-5807(01)00038-7. [DOI] [PubMed] [Google Scholar]
- 256.Chung AC, Xu X, Niederreither KA, Cooney AJ. Loss of orphan nuclear receptor GCNF function disrupts forebrain development and the establishment of the isthmic organizer. Dev Biol. 2006 May 1;293(1):13–24. doi: 10.1016/j.ydbio.2005.12.017. Epub 2006 Mar 10. [DOI] [PubMed] [Google Scholar]
- 257.Akamatsu W, DeVeale B, Okano H, Cooney AJ, van der Kooy D. Suppression of Oct4 by germ cell nuclear factor restricts pluripotency and promotes neural stem cell development in the early neural lineage. J Neurosci. 2009 Feb 18;29(7):2113–24. doi: 10.1523/JNEUROSCI.4527-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Sabour D, Xu X, Chung AC, Le Menuet D, Ko K, Tapia N, Araúzo-Bravo MJ, Gentile L, Greber B, Hübner K, Sebastiano V, Wu G, Schöler HR, Cooney AJ. Germ cell nuclear factor regulates gametogenesis in developing gonads. PLoS One. 2014 Aug 20;9(8):e103985. doi: 10.1371/journal.pone.0103985. eCollection 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Zechel C. The germ cell nuclear factor (GCNF) Mol Reprod Dev. 2005 Dec;72(4):550–6. doi: 10.1002/mrd.20377. Review. [DOI] [PubMed] [Google Scholar]
- 260.Hunter CP, Kenyon C. Specification of anteroposterior cell fates in Caenorhabditis elegans by Drosophila Hox proteins. Nature. 1995;377:229–32. doi: 10.1038/377229a0. [DOI] [PubMed] [Google Scholar]
- 261.Benassayag C, Montero L, Colombie N, Gallant P, Cribbs D, Morello D. Human c-Myc isoforms differentially regulate cell growth and apoptosis in Drosophila melanogaster. Mol Cell Biol. 2005;25:9897–909. doi: 10.1128/MCB.25.22.9897-9909.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Krause M. MyoD and myogenesis in C. elegans. Bioessays. 1995 Mar;17(3):219–28. doi: 10.1002/bies.950170308. Review. [DOI] [PubMed] [Google Scholar]
- 263.Baylies MK, Michelson AM. Invertebrate myogenesis: looking back to the future of muscle development. Curr Opin Genet Dev. 2001 Aug;11(4):431–9. doi: 10.1016/s0959-437x(00)00214-8. Review. [DOI] [PubMed] [Google Scholar]
- 264.Laudet V, Gronemeyer H. The nuclear receptor facts book. Academic Press; 2002. [Google Scholar]
- 265.Helsen C, Kerkhofs S, Clinckemalie L, Spans L, Laurent M, Boonen S, Vanderschueren D, Claessens F. Structural basis for nuclear hormone receptor DNA binding. Mol Cell Endocrinol. 2012 Jan 30;348(2):411–7. doi: 10.1016/j.mce.2011.07.025. Epub 2011 Jul 27. Review. [DOI] [PubMed] [Google Scholar]
- 266.Schwartz RM, Dayhoff MO. Protein and nucleic Acid sequence data and phylogeny. Science. 1979 Sep 7;205(4410):1038–9. doi: 10.1126/science.205.4410.1038. [DOI] [PubMed] [Google Scholar]
- 267.Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol. 2016 Jul;33(7):1870–4. doi: 10.1093/molbev/msw054. Epub 2016 Mar 22. [DOI] [PMC free article] [PubMed] [Google Scholar]