

Abstract
Dysregulation in immune responses during pregnancy increase the risk of a having a child with an autism spectrum disorder (ASD). Asthma is one of the most common chronic diseases among pregnant women, and symptoms often worsen during pregnancy. We recently developed a mouse model of maternal allergic asthma (MAA) that induces changes in sociability, repetitive and perseverative behaviors in the offspring. Since epigenetic changes help a static genome adapt to the maternal environment, activation of the immune system may epigenetically alter fetal microglia, the brain’s resident immune cells. We therefore tested the hypothesis that epigenomic alterations to microglia may be involved in behavioral abnormalities observed in MAA offspring. We used the genome-wide approaches of whole genome bisulfite sequencing to examine DNA methylation and RNA sequencing to examine gene expression in microglia from juvenile MAA offspring. Differentially methylated regions (DMRs) were enriched for immune signaling pathways and important microglial developmental transcription factor binding motifs. Differential expression analysis identified genes involved in controlling microglial sensitivity to the environment and shaping neuronal connections in the developing brain. Differentially expressed associated genes significantly overlapped genes with altered expression in human ASD cortex, supporting a role for microglia in the pathogenesis of ASD.
Keywords: Maternal Immune Activation, Maternal Allergic Asthma, Autism Spectrum Disorders, DNA methylation, Microglia
Introduction
Epigenetic mechanisms fine tune gene expression during the development and maturation of the nervous and immune systems (Vogel Ciernia and LaSalle, 2016) resulting in long-lived phenotypic alterations without changing DNA sequence. Epigenetic mechanisms such as DNA methylation can be altered by environmental changes during a particular critical window in early life (Dolinoy et al., 2007; Jirtle and Skinner, 2007). The developing mammalian brain is particularly sensitive to epigenetic alterations, as mutations in epigenetic effectors can result in human neurodevelopmental disorders (Lasalle, 2013; Vogel Ciernia and LaSalle, 2016). Epigenetic changes are thought to adapt a static genome to a dynamic environment, providing an important interface between genetic and environmental risk factors in the complex etiology of autism spectrum disorders (ASD).
Epidemiological reports suggest a strong association between periods of maternal immune activation (MIA) during pregnancy and an increased risk of having a child with ASD (Atladottir et al., 2009, 2010, Zerbo et al., 2013, 2015). Early case reports and small comparative studies focused on infections during pregnancy associated with ASD risk (Chess, 1977; Sweeten et al., 2004; Libbey et al., 2005). Large population based studies in Denmark showed that mothers with severe infections during pregnancy that required hospitalization were at increased risk of having a child with ASD (Atladottir et al., 2010); a finding also confirmed in a Californian cohort (Zerbo et al., 2013). Moreover, taking antipyretic medication to reduce fever during pregnancy eliminated this increased risk for ASD (Zerbo et al., 2013), suggesting that treating the MIA attenuates risk. In addition to infections, associations of risk for ASD with autoimmune and immunological conditions such allergy, asthma, celiac disease, type-1 diabetes, autoimmune thyroid disease, psoriasis, rheumatoid arthritis, and rheumatic fever have been reported (Croen et al., 2005; Mouridsen et al., 2007; Atladottir et al., 2009; Mostafa and Kitchener, 2009; Keil et al., 2010), providing additional evidence for maternal immune dysfunction and increased risk of ASD.
Asthma is one of the most common chronic diseases among pregnant women. During pregnancy the median percent of women hospitalized for acute exacerbations of asthma is 6% (Murphy et al., 2005, 2006, Ali and Ulrik, 2013a; b); however, in a study of 330 women with asthma, symptoms worsened in as many as 35% during pregnancy (Schatz et al., 1988). A large epidemiological study demonstrated that mothers with new onset allergies and asthma during the time of pregnancy were at increased risk for having a child with ASD (Croen et al., 2005). A separate study that looked at maternal allergies and asthma, irrespective of whether they were new onset or existing conditions, also found increased risk with certain allergies and ASD (Lyall et al., 2014). Recently we identified a profile of elevated serum cytokines midgestation in women who gave birth to a child later diagnosed with ASD (Goines et al., 2011; Jones et al., 2017). This cytokine profile of elevated interleukins IL-4 and IL-5 was consistent with an allergic asthma clinical phenotype (Magnan et al., 2000; Cho et al., 2005; Tamasi et al., 2005; Kumar et al., 2006). Furthermore, a different midgestation maternal cytokine profile of elevated IL-6 and IL-2 was associated with having a child with other developmental disorders, but not ASD (Goines et al., 2011). Amniotic fluid taken from children later diagnosed with ASD also had elevated levels of IL-4 whereas those with developmental delay had increased levels of IL-6 (Abdallah et al., 2012). These reports implicate asthma/allergy and associated cytokine responses as a potential contributing factor to the development of ASD.
Previous work in animal models examining the impact of gestational MIA on ASD-like behaviors of the offspring have focused on viral and bacterial responses, using polyinosinic-polycytidylic acid (polyI:C) or lipopolysaccharide (LPS) as experimental models (Whitehead et al., 2003; Nials and Uddin, 2008). However, allergies and asthma represent an alternative inflammatory response mediated by T-helper type 2 cells (TH2) that produce a different cytokine profile that may uniquely alter brain development relevant to ASD. We recently developed the first preclinical model of maternal allergic asthma (MAA) that produces the hallmark immunological responses in the MAA treated dams analogous to those observed in humans, including allergic airway inflammation and elevated levels of IL-4 and IL-5 (Schwartzer et al., 2015). MAA produced a phenotype in the offspring of impaired social approach and increased repetitive behaviors at both juvenile and adult ages (Schwartzer et al., 2015, 2017) that resemble behavioral features of ASD (Thomas et al., 2009). MAA challenge in gestation also resulted in altered brain neurochemistry in offspring similar to those observed in ASD (Schwartzer et al., 2015).
In humans, fetal exposure to maternal asthma led to epigenetic alterations in DNA methylation patterns in peripheral blood samples of the offspring (Gunawardhana et al., 2014). Notably, hypo-methylation in immune signaling gene MAP8KIP3 in fetal blood correlated with blood eosinophils, nitric oxide levels, and total serum IgE in the mother (Gunawardhana et al., 2014), suggesting that human MAA responses may be related to additional epigenetic changes in the offspring. These findings are consistent with previous work which found histone acetylation (Tang et al., 2013) and DNA methylation (Basil et al., 2014; Richetto et al., 2016) changes in brain from juvenile and adult offspring in the polyI:C MIA mouse model. Together these results suggest that epigenetic changes within the immune cells of the brain may be responsible for the long-lived behavioral changes of offspring exposed to gestational MAA.
Within the brain microglia are brain-resident macrophages derived from erythro-myeloid precursors from the embryonic yolk sac that contribute to brain homeostasis and shaping synaptic and long-range neuronal connections (Tremblay et al., 2010; Paolicelli et al., 2011; Schafer et al., 2012; Hashimoto et al., 2013; Zhan et al., 2014). Microglia monitor and respond to their environment via specific cell surface receptors and secreted molecules, thereby acting as sentinel cells of tissue stress and injury (Murray and Wynn, 2011). The gestational timing of MAA may lead to alteration in microglial activity during key organizational phases of prenatal development producing long lasting changes in neural circuitry. Microglia arise from the yolk sac myeloid progenitors, migrate to the brain and colonize the neural folds during embryogenesis with minimal replacement from peripheral monocytes (Squarzoni, 2015), suggesting that any perturbations to microglia in early development may also persist through adulthood. To test the hypothesis that gestational MAA is associated with long-lived ASD-relevant epigenetic changes in microglia, we performed genome-wide analyses of the DNA methylome and transcriptome of microglia isolated from juvenile offspring. The results of this study provide genome-wide evidence that gestational MAA alters DNA methylation of key immune regulatory elements and transcription of genes dysregulated in human ASD brain, supporting the validity of the MAA mouse model, microglia as a relevant cell type, and maternal asthma as an ASD risk factor.
Materials and Methods
Animals
All experiments used C57Bl/6J mice from Jackson (Jax strain 000664). All experiments were conducted in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. All procedures were approved by the Institutional Animal Care and Use Committee of the University of California, Davis.
Maternal Allergic Asthma Paradigm
The MAA paradigm was conducted as previously described (Schwartzer et al., 2015, 2017). Briefly, on postnatal day (P) 42 and 49 sexually naive female C57Bl/6J mice were sensitized with a single intraperitoneal injection of 10 μg ovalbumin (OVA, Sigma, St. Louis, MO USA) in 1 mg (Al)OH3 (InvivoGen, San Diego, CA USA) dissolved in 200 μl phosphate buffered saline (PBS). One week following the second sensitization treatment, female mice were mated overnight and checked daily for the presence of a seminal plug, noted as gestational day 0.5 (G0.5). Pregnant mice were randomly assigned to either the allergic asthma or control group and exposed to either an aerosolized solution of 1% (wt/vl) OVA in PBS (MAA group) or vehicle control for three 45-minute induction on gestational days 9.5, 12.5, and 17.5, to correspond with early, middle, and late gestation. Following the final induction, mice were returned to their home cages, single housed, and left undisturbed until the birth of their litters. Pups remained with their mother until weaning on P21, at which time the offspring were group housed with same-sex littermates. For all experiments female offspring were chosen for analysis due to previously identified increased microglial reactivity at P30 in females compared to males (Schwarz et al., 2012). Female MAA offspring also show a similar behavioral phenotype to male MAA offspring with deficits in social interactions, increased marble burying and decreased grooming (Schwartzer et al., 2015, 2017).
Microglial Isolation P35
P35 female mice were deeply anaesthetized with CO2 and then quickly perfused intracardially with ice-cold PBS. Whole brains were removed and stored on ice in HBSS without Ca2+/Mg2+ until processed. Each brain was gently homogenized to a single cell solution using a dounce homogenizer, then added to 1.8 ml of isotonic Percoll to form a 30% isotonic Percoll solution. Isotonic Percoll gradients were then constructed with layers of 70% with 1× phenol red, 37% and 30%. The resulting gradients were centrifuged for 30 min at 500×g at room temperature. The top layer of myelin and non-microglial cells were discarded and the microglia at the interface between the 37% and 70% layers were collected. The resulting microglia were then washed in HBSS and counted. The collected cells were immediately flash frozen for RNA/DNA extraction. Given the low yield from individual animals, an additional animal was run in parallel and harvested microglia were used to assess purity by flow cytometry for CD45.2 and CD11b. Gating was first performed on live single cells (96%). Any highly granular myelin contamination was removed (92% remaining). From that >90% was microglia (CD45.2 and CD11b positive). The remaining ≈6% (CD45.2 positive, but CD11b negative) represent a minimal contamination of other mononuclear cells (Figure S1).
RNA sequencing library preparation and sequencing
RNA and DNA were isolated from the same isolated microglia samples using ZR-Duet RNA/DNA kit (Zymo, D7001) with on-column DNase digestion (Qiagen, 79254) for RNA extraction. RNA and DNA were quantified using Qubit High Sensitivity assays. RNA quality was assessed by Bioanalyzer and only samples with RNA Integrity Number (RIN) scores greater than 7 were used for library preparation. 60–100 ng of input RNA per sample were used for RiboGone rRNA removal (Clonetech, 634847). 2 ng of rRNA depleted RNA per sample was then used for library construction using the SMARTer Stranded RNA-Seq kit (Clontech, 634836) and 14 cycles of PCR amplification. Library quality was assessed using a bioanalyzer and quantified by Qubit high sensitivity assay. All samples were barcoded, pooled, and run on a single sequencing lane of a HiSeq3000 to generate 50 bp single end reads. RNA-seq reads were analyzed for both quality and adapter contamination using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). All reads were trimmed using Trimmomatic (Bolger et al., 2014) using Illuminaclip single end adapter trimming, a removal of the first three bp from the 5′ as recommended for SMARTer libraries, and a 4 bp sliding window trim for quality scores less than 15. All remaining reads greater than 15 bp were then reanalyzed using FASTQC to verify quality. Potential library contamination was examined by aligning 100,000 reads from each sample to genomic sequences from E. coli, mouse (mm10, including rRNA and mtDNA), human (hg38), and PhiX using FastQ Screen (http://www.bioinformatics.babraham.ac.uk/projects/fastq_screen/).
RNA sequencing PCA analysis
Principle Component Analysis (PCA) was conducted on read counts from published cell type specific RNA sequencing (Zhang et al., 2014; Mo et al., 2015) with normalization for library size. Raw Fastq files for each cell type were downloaded from GEO (GSE63137 and GSE52564) and processed as described except for paired end sequencing. PCA analysis was conducted on the top 500 most variably expressed genes across all cell types using custom R scripts. To further assess microglial purity subsampling of the aligned reads for both neuronal and microglial samples(Zhang et al., 2014) (GEO GSE52564) were performed to match the average 3×107 reads/sample (Table S2) obtained in this experiment. Specific proportions of randomly selected reads were then extracted from the neuron and microglial samples, combined, counted and processed in EdgeR with normalization for total library size. Proportions for each combined mixed sample are listed in Figure S1.
RNA sequencing differential expression analysis
RNA-seq reads were aligned to the current mouse genome mm10 using the splicing-aware aligner Tophat2 (v2.0.14/bowtie2.2.5, stranded, Single End)(Kim et al., 2013) and reads aligning to each gene were counted using FeatureCounts (Liao et al., 2014) with exclusion of multimapping reads. The resulting table of counts per gene and sample was analyzed using the EdgeR pipeline (Robinson et al., 2010). Lowly expressed genes (less than 1 count per million reads in at least three biological replicates from either MAA or PBS groups) were removed from the analysis and the remaining reads were normalized for library size and RNA composition using the calcNormFactors function with trimmed mean of M-size method (Robinson and Oshlack, 2010). The normalized libraries were then fed into a one factor model (MAA vs. PBS) with a zero intercept. Both common and tagwise dispersions were estimated using estimateDisp and the resulting model was used for differential expression using exactTest and topTags with a filter for false discovery rate < 0.05. All differential expression analysis was performed on normalized counts per million (CPM) mapped reads. For graphical comparison of selected genes of interest and heatmap generation Reads per kilobase of transcript per million mapped reads (RPKM) was calculated using the built-in rpkm function that normalized for gene length. Gene Ontology and Pathway enrichment for differentially expressed genes were conducted using Enrichr (http://amp.pharm.mssm.edu/Enrichr/enrich) (Chen et al., 2013; Kuleshov et al., 2016) and then trimmed using REVIGO (http://revigo.irb.hr/) (Supek et al., 2011).
Whole Genome Bisulifte sequencing (WGBS) analysis
WGBS libraries were constructed using the Illumina EpiGnome/TruSeq DNA Methylation kit (Illumina, EGMK81312) with indexing barcodes (Illumina, EGIDX81312). 100 ng of DNA from each sample was bisulfite converted using the EZ DNA Methylation-Lightning kit (Zymo, D5030). Each library was given a unique barcode identifier and 14 cycles of PCR amplification. Libraries were quantified by Qubit and quality was assessed by Bioanalyzer. Sequencing of barcoded and pooled libraries was performed across two lanes of Illumina HiSeq4000 to obtain 100 bp single end reads.
After adapter trimming and quality assessment, all WGBS reads were mapped to the mouse genome (mm10) using the Bisulfite Seeker2 program (v2.0.8 with bowtie1.1.1) (Guo et al., 2013) and subsequent analysis was done with custom Perl and R scripts (Dunaway et al., 2016b). Differentially methylated regions (DMR) between experimental conditions was examined using the R packages DSS and bsseq (Feng et al., 2014). DMRs were identified as sets of CpGs with a t-statistic greater than the critical value for alpha 0.05 and with a gap between CpGs of less than 300 bases. DMRs were then filtered for those with more than 3 CpGs, a mean methylation difference of greater than or equal to 10% between conditions, and an area stat of +/− 20. Further permutation testing was used to identify DMRs with a family wise error rate <0.05 based on 1,000 permutations of random shuffling of sample group assignment. The average difference in methylation was 29.94% for hyper-methylated and 31.57% for hypo-methylated DMRs. The average size of the hyper-methylated DMRs were 492 bases with an average of 10 CpGs and hypo-methylated DMRs were on average composed of 450 bases and 10.9 CpGs.
DMRs were assessed for Ontology and pathway enrichment using Genomic Regions Enrichment of Annotations Tool (GREAT) with default settings for regulatory regions (5 kb +/− 1 kb of TSS and basal regulatory domain extension from nearest gene of 1Mb) (McLean et al., 2010) with filters for both significant binomial and hypergeometric tests (FDR < 0.05 The resulting GO term lists were trimmed to remove redundancy using REVIGO. Motif enrichment within DMRs was assessed using HOMER (Heinz et al., 2010) (FDR < 0.05) and lists were filtered for motifs commonly identified in both hyper- and hypo-methylated DMRs, hypo-methylated DMRs only, or hyper-methylated DMRs only. Genes with transcription start sites within +/− 5kb of each DMR were identified using HOMER (Heinz et al., 2010).
In addition to global assessment of CpG and CpH methylation levels, CpG methylation over gene bodies, CpG islands, CpG shores (±2 kb from a CpG island), and tissue specific enhancer regions were also examined and tested for differential methylation using independent, two-tailed t-test with FDR correction and a minimum of 5 reads per region across all samples.
WGBS PCA analysis
PCA was conducted on percent mCG/CG from publish cell type specific WGBS (Lister et al., 2013; Mo et al., 2015). Raw Fastq files for each cell type were downloaded from GEO (GSE63137 and GSE47966) and processed as described for WGBS. Percent CG/CG methylation was extracted for 20 kb windows, gene bodies, and CpG islands for each sample. CpG Islands were masked from the analysis of the other two regions. PCA analysis was conducted on the top 500 most variable regions of each type for all cell types using custom R scripts.
Fluidigm Biomark HD Delta Gene Expression Assay
Gene expression for selected targets identified from RNAseq analysis were examined in an independent set of microglial samples isolated from PND35 female littermates of animals used for sequencing (Supplemental Table S3). Gene expression analysis was performed using the Fluidigm Biomark HD Delta gene assay on a 48.48 Integrated Fluidic Circuit. Delta gene assay design was performed by Fluidigm and primer sequences are listed in Supplemental Table S3. 20ng of total RNA was used as input for cDNA synthesis (Fluidigm, 100–6472 B1) followed by pre-amplification (13ng per sample) (Fluidigm, 100–5875 C1) using a mix of all forward and reverse primers. Samples were then diluted five-fold and analyzed on a 48.48 IFC and Biomark HD machine (1.4ng/sample) in duplicate (Fluidigm, 100–9791 B1). Cycle threshold values were averaged across replicates and then normalized to hprt expression (no difference in mean Ct values for hprt between conditions, t= 1.27, p = 0.239) using the delta Ct method (Ct gene – Ct hprt). These values were then used for group comparisons with a two-tailed t-test without an assumption of equal variance. For display purposes, the delta delta Ct values were then calculated relative to the average Ct value of the PBS treated control for each gene, and relative expression levels were then calculated for each group by 2^-delta delta Ct (Figure S4D).
Overlap of Gene lists
Gene list overlap was performed using the GeneOverlap (Shen, 2013) package in R with a Fisher’s exact test and a False Discovery Rate correction to p=0.05. Gene lists and references are listed in Table S7 (Voineagu et al., 2011a; Oskvig et al., 2012; Garbett et al., 2012; Sugathan et al., 2014; Gilissen et al., 2014; Gunawardhana et al., 2014; Gupta et al., 2014; Iossifov et al., 2014b; Cotney et al., 2015; Cronk et al., 2015; Sanders et al., 2015; Holtman et al., 2015; Matcovitch-Natan et al., 2016; Richetto et al., 2016; Rube et al., 2016; Dunaway et al., 2016b; Zhao et al., 2017; Lombardo et al., 2017). For comparisons to differentially expressed genes the background gene list was all genes identified as expressed in the RNAseq experiment (see above description) (14,301 genes). For comparisons to DMR associated genes, background genes were defined from a call to the DMR finder for mm10 with no filters or cutoffs. This produces a list of all potential locations in the mm10 genome that could be called as a significant DMR. Genes associated with (+/− 5 kb) this background DMR set were used as the background list (22,412 genes). For comparisons between differentially expressed and DMR associated genes, a common background gene set was used that included the intersection of both the differentially expressed and DMR associated background gene list (13,694 genes that could have both been differentially expressed and within 5 kb of a DMR). All gene lists used in each comparison were filtered to include only genes present on the respective background list for that comparison. Drug target analysis was performed using The Drug Gene Interaction Database (dgidb.genome.wustl.edu)(Dunaway et al., 2016b).
Results
Figure 1A shows the experimental design prior to microglial isolation. To induce a MAA response, female mice were pre-sensitized with OVA exposure at 6 and 7 weeks of age prior to breeding. Timed pregnant dams were then given three exposures to ovalbumin (OVA) during gestation (gestation day 9.5, 12.5, and 17.5) or three exposures to endotoxin free PBS to mimic immune challenge or control, respectively (Schwartzer et al., 2015).
Figure 1. Maternal Allergic Asthma Model.
A. Maternal allergic allergy/asthma (MAA) was induced by pre-sensitizing dams to ovalbumin (OVA) prior to mating. During subsequent pregnancy half of the dams were challenged with OVA (gestation day (GD) 9,12, and 17) and the other half received PBS. Microglia were then harvested from the resulting female pups on postnatal day 35 (P35). Male pups were used for behavioral assessment as part of a recently published study (Schwartzer et al., 2017).
While ASD has a pronounced sex-bias for males, both male and female MAA offspring show similar behavioral phenotypes deficits in social interactions, increased marble burying, and decreased grooming (Schwartzer et al., 2015, 2017). In rodents, adolescent females show increased microglial reactivity compared to males (Schwarz et al., 2012). Consequently, this study focused on characterizing juvenile microglia from female MAA offspring. In order to study the effects of in utero MAA on persistent microglial epigenomic signatures that coincide with the observance of ASD-like behaviors (Schwartzer et al., 2015, 2017), microglia were isolated from juvenile mice from multiple litters (n= 4 mice from a total of 4 litters per condition) on P35. There were no significant differences in the number of microglia isolated or the RNA/DNA yields between the two treatment groups (Table S1).
Microglial Methylome and Transcriptome Reveal Cell Type Specific Signatures
RNA sequencing was performed to examine differences in gene expression associated with MAA in juvenile microglia (n=4/group). There were on average 28.6 million uniquely mapped reads per sample and no differences in read depth or mapping between conditions (Table S2). All libraries passed FastQC assessment and none indicated significant sources of contamination during library preparation (Figure S1A & B).
We first sought to ensure that the gene expression patterns in both experimental conditions fit with expected microglial markers and gene expression patterns. To examine cell type specific gene expression patterns we compared the MAA and PBS microglia collected in this experiment to previously published RNA sequencing data on carefully sorted cell populations from mouse cerebral cortex (neurons, astrocytes, oligodendrocyte precursor cells, newly formed oligodendrocytes, myelinating oligodendrocytes, microglia, endothelial cells, and pericytes) (Zhang et al., 2014) and several specific types of neurons (excitatory, PV interneurons and VIP interneurons) (Mo et al., 2015). Both the PBS and MAA microglia transcriptomes cluster more closely with those of adult cortical microglia and are separated from those of other cortical cell types (Figure 2A). Further analysis of expression (reads per kilobase of transcript per million mapped reads, RPKM) values for cell type marker genes (Hickman et al., 2013; Zhang et al., 2014) revealed high expression of microglial markers and low expression of cell type specific markers from other cortical cell types (Figure 2B). In addition, subsampling microglial and neuronal reads from previously published RNAseq (Zhang et al., 2014) indicated that the percentage of neuronal RNA contamination in our microglial samples was minimal and between 0.2 to 4% (Figure S1C). Analysis by flow cytometry on samples prepared in parallel indicate a similar purity greater than 90% (Figure S1D).
Figure 2. Microglial Transcriptome and Methylome Reveal Cell Type Specific Signatures.
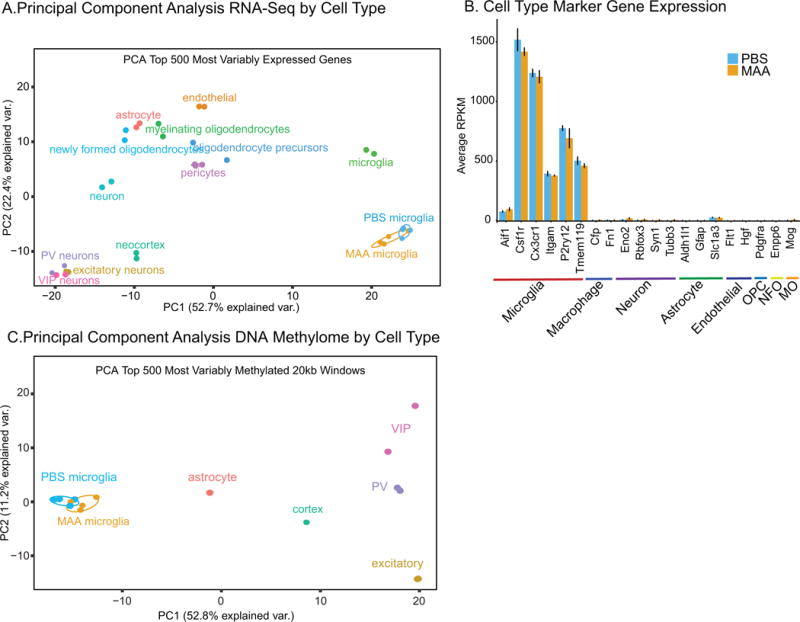
A. Principle component analysis of the gene counts from the top 500 most variably expressed genes across multiple brain cell types (neurons, astrocytes, oligodendrocyte precursor cells, newly formed oligodendrocytes, myelinating oligodendrocytes, microglia, endothelial cells, and pericytes (Zhang et al., 2014) and several specific types of neurons (excitatory, PV interneurons and VIP interneurons (Mo et al., 2015)). B. RPKM values for selected cell type marker genes (Hickman et al., 2013; Zhang et al., 2014) for MAA and PBS samples (n=4/group). Oligodendrocyte precursor cells (OPC), newly formed oligodendrocyte (NFO), myelinating oligodendrocyte (MO). Error bars +/− SEM C. Log2RPKM values for MAA compared to PBS microglial RNAseq samples (n=4/group). C. Principle component analysis of the %mCG/CG from the top 500 most variably methylated 20 kb windows across the genome of multiple brain cell types including several specific types of neurons (excitatory, PV interneurons and VIP interneurons (Mo et al., 2015)), astrocytes and total fetal cortex (P28) (Lister et al., 2013). CpG Islands were masked from the analysis.
To examine DNA methylation changes in P35 microglia between MAA and PBS treated samples using an unbiased genome-wide approach, we performed WGBS. There were no significant differences in sequencing coverage, bisulfite conversion, or in the global percentage of CG, CHG, or CHH methylation between treatments (Table S2). On average microglia showed similar levels of mCG and mCH as previously described in neurons (Lister et al., 2009). To examine cell type specific methylation patterns we compared 20 kb windows across the genome from the MAA and PBS microglia collected in this experiment to previously published WGBS data on several specific types of neurons (excitatory, PV interneurons and VIP interneurons) (Mo et al., 2015), astrocytes, and brain (P28) (Lister et al., 2013). Both the PBS and MAA P35 microglia samples cluster more closely with each other than with other cortical cell types, indicating microglia show a distinct methylation pattern from other brain cell types (Figure 2C). Similar patterns were observed over gene bodies and to a lesser extent over CpG islands (Figures S2A and B). There were no significant differences between MAA and PBS treatment groups in average percent CG methylation for a number of global genomic features, including gene bodies, 5 kb upstream promoter regions (with or without CpG islands), nor CpG islands (Figure S2C).
Differentially Methylated Regions in MAA Offspring Microglia
To assess the long-term impact of MAA on epigenetic regulation in offspring microglia we identified locus-specific differentially methylated regions (DMRs) between MAA and PBS (Dunaway et al., 2016a; b). 1,184 DMRs were identified, with nearly equivalent numbers of hyper-methylated (626 MAA>PBS) and hypo-methylated (558 PBS>MAA) regions (Figure 3A, Table S3). Three DMRs also passed family wise error rate (FWER) correction (Figure S2D). In general, MAA associated DMRs were largely localized within introns (40%) and intergenic sites (26%) (Figure 3B) indicating potential regulatory function.
Figure 3. Differential Methylation between MAA and PBS Microglia.
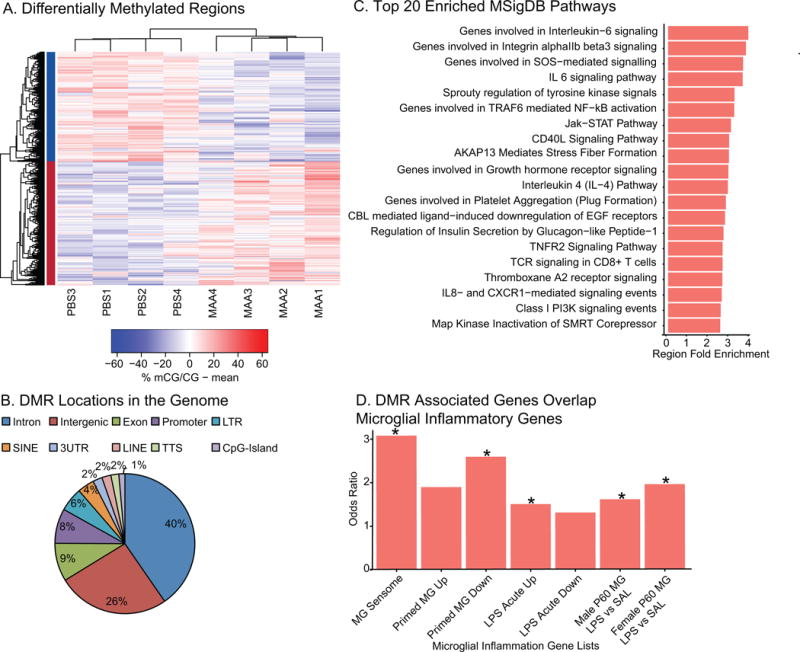
A. Differentially methylated regions (DMRs) with an area statistic >= +/−20 and a 10% or greater difference in average methylation between treatments. 1184 DMRs were identified in two clusters: hyper-methylation (MAA>PBS, 626 and hypomethylation (PBS>MAA, 558). Values shown in the heatmap are the difference from the mean for mCG methylation: % mCG/CG minus the mean of %mCG/CG for all samples. See Table S3. B. Distribution of DMRs relative to Transcription Start Sites within +/− 5kb of the DMR (HOMER (Heinz et al., 2010) annotation). C. Top 20 significantly enriched pathways from MSigDB using GREAT annotation of 1184 DMRs. Region Fold Enrichment: observed number of DMRs with the annotation/expected number of DMRs with the annotation (McLean et al., 2010). See Table S4 for full lists. D. Fisher’s Exact Test Odds Ratios for gene list overlap between genes with transcription start sites +/− 5kb from the MAA DMRs. DMR associated genes significantly overlapped genes from the microglial (MG) sensome (Hickman et al., 2013), primed microglia, genes up-regulated acutely in response to LPS (Holtman et al., 2015), and genes up-regulated in microglia two hours after LPS in males and females at postnatal day 60 (P60) (Hanamsagar et al., 2017). See Table S7 for full lists and statistics.
Functional annotation was assigned to each DMR using GREAT, which included biological processes such as regulation of cell number, embryo development, and response to cytokine stimulus. Pathway analysis (Panther and MsigDB) showed enrichment for a number of immune pathways including inflammation mediated by cytokine and chemokine signaling such as IL-6, IL-4 and IL-8 signaling, as well as Jak-STAT, NF-κB, TNF and mTOR signaling (Figure 3C and Table S3). When specific enrichment was examined for MSigDB immune signatures, DMRs were found to be enriched for gene sets associated with LPS treatment and other types of immune cell activators (Table S3). Furthermore, genes with transcription start sites +/−5kb of DMRs (424 DMRs, total with 209 hyper-methylated DMRs and 215 hypo-methylated DMRs) (Table S5) were significantly enriched for genes responsive to LPS, and other immune stimuli in microglia (Figure 3D) (Hickman et al., 2013; Holtman et al., 2015; Hanamsagar et al., 2017). Together these findings indicate that regions sensitive to MAA methylation differences may contain critical regulatory regions responsive to immune stimuli in microglia.
To evaluate potential transcription factor binding sites enriched within MAA-DMRs, we examined motif enrichment analysis using HOMER (Table S4). Motifs commonly enriched in hypo- and hyper-methylated DMRs include transcription factors related to a number of different functions including the SOX, SMAD, and TGFβ-induced families of transcription factors. Hyper-methylated DMRs were enriched for several interesting transcription factor motifs that are critical for early microglial development and immune activation including RUNX1, PU.1, IRF8, NF-kB, and MAFB. Hypo-methylated DMRs were enriched for several zinc finger and LIM homeobox proteins, together suggesting that regulation of inflammatory signaling and microglial developmental may be altered by DNA methylation differences in MAA microglia.
Differentially Expressed Genes in MAA Offspring Microglia
In order to understand how epigenomic differences in MAA offspring microglia were related to regulation of gene expression we examined differential gene expression patterns from the same microglial isolations that were profiled for DNA methylation. Differential expression between MAA and PBS samples was conducted using the standard EdgeR pipeline and revealed 162 differentially expressed genes at an FDR < 0.05 (Figure 4A & B and Table S5). Of these genes the vast majority showed greater expression in MAA samples compared to PBS (157 MAA>PBS). Gene Ontology enrichment analysis revealed significant enrichment of several categories of GO terms including gated and ion channel activity, synapse part, and regulation of cell projection organization (Figure S3A and Table S6). Pathway enrichment using KEGG, Panther and Reactome revealed significant enrichment for pathways involved in a number of processes critical for neurodevelopment including several neurotransmitter systems, axon guidance, and synapse function (Table S6). While many of these gene targets are typically examined in neurons, microglia express GABAB, serotonin and dopamine, NMDA, AMAP, and kainic acid and metabotropic glutamate receptors that can be utilized to sample the local environment and consequently regulate activation, chemotaxis, secretion, proliferation and phagocytosis (Craner et al., 2005; Li et al., 2008; Black and Waxman, 2012; Stebbing et al., 2015). Gene expression changes were further validated on an independent group of microglial samples using a Fluidigm Delta Gene Expression Assay on a selected subset of transcripts (Table S3B). With this independent method and cohort all transcripts validated the direction of expression change seen in the sequencing cohort (Table S5) with Hspalb levels reaching significance (p < 0.05) and Ina levels approaching significant (p < 0.07) (Figure S3B).
Figure 4. RNA Sequencing Analysis of P35 Microglia.
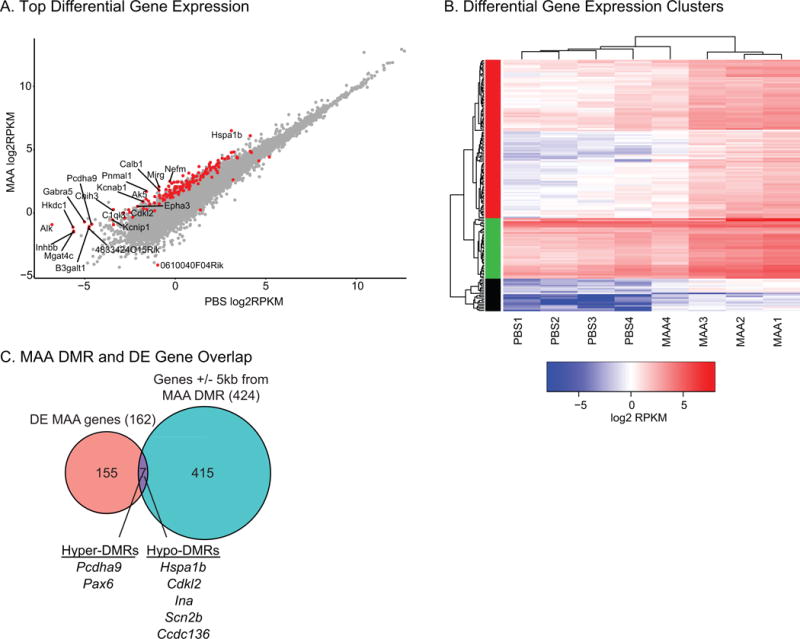
A. Significantly differentially expressed genes (162; FDR q <0.05) are highlighted in red. Genes with log fold change >= 2.5 are labeled. B. Heatmap of log2 RPKM values for the 162 differentially expressed genes between MAA and PBS conditions. Hierarchical clusters (k=3) were used to identify patterns of gene expression using hclust in R. See Table S5 for gene lists, RPKM values, and statistics. C. MAA differentially expressed and DMR associated (+/− 5kb from a DMR) share seven genes. See Table S7 for statistics.
To investigate the relationship between MAA-induced methylation and expression changes, DMR associated genes were compared with differentially expressed genes. There was not a significant overlap between all differentially expressed genes and all genes associated with DMRs (Fisher’s exact test, Odds Ratio = 1.96, p = 0.116) with only 7 genes shared between the two lists (Figure 4C). There was also not a significant overlap with the 70 genes identified as associated with single site CpG differential methylation from blood from human infants from asthmatic mothers (Fisher’s exact test, all p > 0.05) (Gunawardhana et al., 2014). For the seven genes that did overlap between MAA differentially expressed and DMR associated genes, Pcdha9, Pax6 and Ina are all critical genes for normal neurodevelopment and are SFARI ASD risk genes (SFARI.org), suggesting that alterations in MAA microglial gene expression and methylation may impact similar pathways disrupted by ASD genetic mutations.
MAA Microglial Impacted Genes are Enriched for Genes Dysregulated in ASD
In order to more fully evaluate the impact of MAA on ASD-related risk genes we overlapped the differentially expressed and differentially methylated gene lists with previously published risk gene lists for ASD, schizophrenia, and intellectual disability (ID) (Sanders et al., 2012, 2015; Gilissen et al., 2014; Iossifov et al., 2014a) (Figure 5A and Table S7). Genes that were differentially expressed and genes that were associated with DMRs in MAA microglia significantly overlapped a hand curated list of SFARI ASD risk genes and genes with Likely Gene Disrupting (LGD) mutations in exome-sequencing from ASD probands (Iossifov et al., 2014). However, the majority of known genetic risk factors were not found within the MAA microglial gene lists, suggesting that MAA is largely targeting gene pathways independently impacted by genetic mutations linked to ASD and ID. This is not surprising given that the majority of genetic risk factors for ASD and ID are not immune-related (Needleman and Mcallister, 2012; Sanders et al., 2015). However, differences in gene expression profiling from human ASD brain have revealed changes in expression of immune signaling pathways and microglial genes (Voineagu et al., 2011; Gupta et al., 2014; Parikshak et al., 2016). To test the hypothesis that MAA microglia may show similar alterations in gene expression as human ASD brain samples, we overlapped the MAA microglial dysregulated genes with genes that showed altered expression or differential epigenomic marks (DNA methylation or histone acetylation) in human ASD brain samples (Voineagu et al., 2011; Gupta et al., 2014; Dunaway et al., 2016b; Parikshak et al., 2016; Sun et al., 2016) (Figure 5B). Differentially expressed genes in MAA microglia significantly overlapped genes with lower expression in both idiopathic and genetic (15q11-13 duplication) forms of ASD as well as genes with increased acetylation. Furthermore, both MAA differentially expressed and DMR associated genes showed enrichment in a number of modules identified from network analysis of differential ASD gene expression from human ASD cortex (Gupta et al., 2014; Parikshak et al., 2016) (Figure 5C and Table S7), suggesting some common pathways are disrupted in human ASD brain and MAA microglia.
Figure 5.
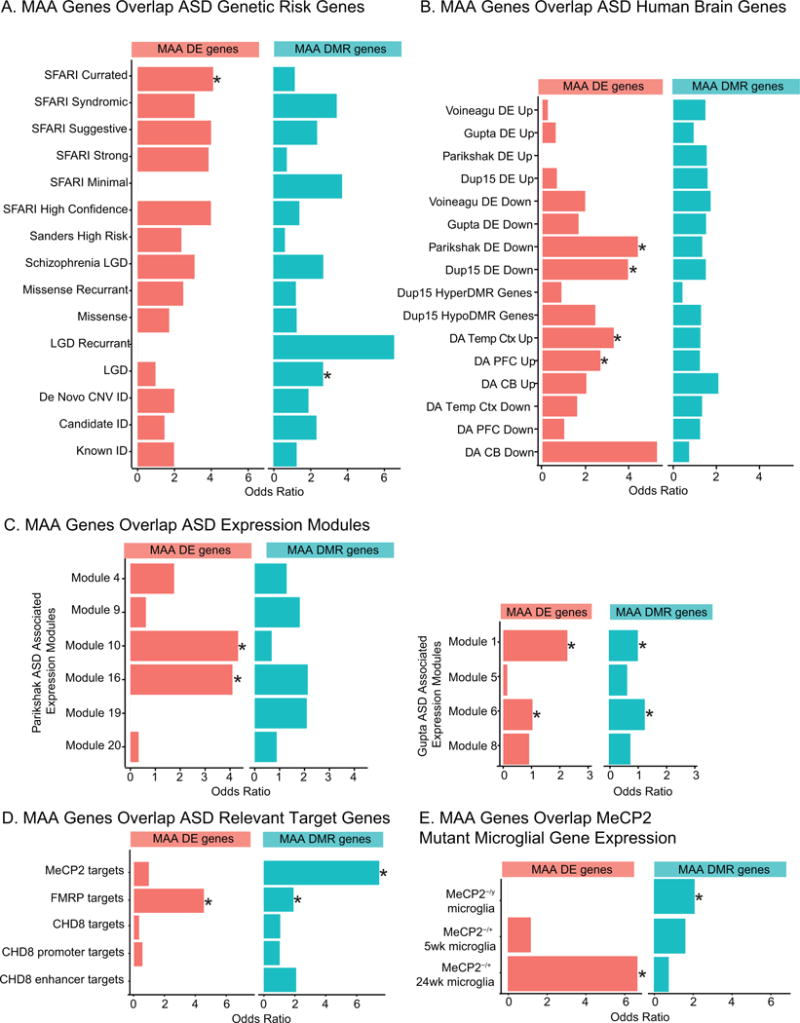
MAA Differentially Expressed and DMR Associated Genes Overlap Genes Dysregulated in ASD. A. Gene list overlaps between MAA differentially expressed genes (MAA DE genes), MAA DMR associated genes (MAA DMR genes) and ASD genetic risk genes (Sanders et al., 2012, 2015; Gilissen et al., 2014; Iossifov et al., 2014a). C. Gene list overlaps between MAA DE genes, MAA DMR associated genes and genes differentially expressed, methylated, or acetylated in human ASD brain samples (Voineagu et al., 2011; Gupta et al., 2014; Dunaway et al., 2016b; Parikshak et al., 2016; Sun et al., 2016). D. Gene list overlaps between MAA DE genes, MAA DMR associated genes and MeCP2, FMRP, or CHD8 target genes (Darnell et al., 2011; Sugathan et al., 2014; Cotney et al., 2015; Rube et al., 2016). E. Gene list overlaps between MAA DE genes, MAA DMR associated genes and genes differentially expressed in microglia from Rett syndrome mouse models (Cronk et al., 2015; Zhao et al., 2017). Lists are from MeCP2−/y null males (fully symptomatic), MeCP2−/+ heterozygous females at either 5 weeks (prior to symptom onset) or at 24 weeks (fully symptomatic). * Fisher’s exact test with FDR corrected p-value <0.05. Gene lists, citations and individual statistics are listed in Table S7.
The enrichment of dysregulated genes in MAA microglia with transcriptionally and epigenetically dysregulated genes in ASD brain suggests that MAA may alter similar pathways as those impacted by ASD. Consequently, we examined the overlap between MAA regulated genes and gene targets of three of the most commonly mutated regulatory factors implicated in ASD and ID: Fragile X Mental Retardation Protein (FMRP), Chromatin-Helicase DNA Binding Protein 8 (CHD8), and Methyl CpG Binding Protein 2 (MeCP2) (Figure 4D). FMRP targets from mouse brain (Darnell et al., 2011) overlapped with both MAA differentially expressed and DMR associated genes (Figure 5D and Table S7) suggesting that key gene targets downstream of FMRP may also be impacted by MAA. Similar overlap between FMRP targets and genes acutely regulated 4 hours following LPS induced MIA has also been previously observed (Lombardo et al., 2017), suggesting that FMRP target genes may be specifically vulnerable to maternal inflammation. There was no significant overlap between either MAA gene list and identified CHD8 target genes from multiple datasets (Sugathan et al., 2014; Cotney et al., 2015). MAA DMR associated genes were enriched in MeCP2 target genes identified from high resolution ChIP-sequencing from olfactory epithelium neurons (Rube et al., 2016), suggesting MeCP2 may be associated with sites of MAA differential methylation. MeCP2 has previously been implicated in microglial function in Rett syndrome (Derecki et al., 2012; Cronk et al., 2015; Schafer and Stevens, 2015), and microglia from Rett mouse models show altered gene expression patterns related to glucocorticoid signaling and stress response (Cronk et al., 2015; Zhao et al., 2017). Genes differentially expressed in Rett syndrome mouse model microglia overlapped both MAA differentially expressed and DMR associated genes (Figure 5E and Table S7), further supporting the role for MeCP2 as a reader of DNA methylation in microglia and indicating that MeCP2 function may be negatively impacted in MAA microglia.
MAA Offspring Microglia Compared to Other MIA Models
Maternal immune activation has been utilized as a model of ASD as well as schizophrenia and general neurodevelopmental impairment (Hagberg et al., 2012; Mattei et al., 2017). The MAA model was originally designed to examine allergic asthma as a risk factor for ASD, but the model may be applicable to other neurodevelopmental disorders that share maternal immune activation as a risk factor. To compare our MAA model to other MIA models we overlapped the MAA gene lists with published datasets from maternal exposures to LPS, PolyI:C, influenza, or direct cytokine injection. DNA methylation in MAA microglia and whole prefrontal cortex from PolyI:C induced MIA (Richetto et al., 2016) showed similar patterns of differential methylation (Figure S4A) suggesting the long-lasting DNA methylation signatures in the brain may be target similar pathways across MIA/MAA models. In comparison, our MAA differentially expressed and DMR associated microglial gene lists largely did not overlap with whole brain gene expression patterns induced four hours following different types of MIA (LPS, recombinant IL6, PolyI:C) (Figure S4B and Table S7). The one exception to this was a significant overlap with genes acutely downregulated in response to LPS MIA. There were mixed findings for gene overlap with microglia isolated from PolyI:C MIA models. MAA microglial gene lists did not overlap dysregulated genes identified in a PolyI:C MIA model from isolated microglia from whole brain on P0 or P28 (Matcovitch-Natan et al., 2016) (Figure S4B). However, MAA differentially expressed genes significantly overlapped with genes that were up-regulated in a PolyI:C MIA model from hippocampal microglia from adult animals (Mattei et al., 2017) (Figure S4C) suggesting that MAA may impact hippocampal microglia. Interestingly, both MAA differentially expressed and DMR associated gene lists overlapped genes that were responsive to minocycline treatment in adult MIA animals (Mattei et al., 2017) (Figure S4C), indicating that minocycline may serve as a potential therapeutic for reversing some of the MAA offspring phenotypes.
Discussion
In this study we identified epigenetic changes in microglia from juvenile offspring of MAA dams that advance the understanding of maternal immune activation and ASD risk in several important ways. We found differentially methylation regions enriched for transcription factor binding sites related to immune signaling, microglial inflammation, and regulation of microglial development. This immune related methylome signature in MAA offspring microglia implicates altered epigenomic regulation of microglial inflammatory responses as a result of MAA. Importantly for ASD relevance, transcriptome profiling of MAA microglia identified key developmental genes related to synaptic function that are also misregulated in human ASD brain.
The gestational timing of MAA may preferentially affect changes in microglial function in offspring, a perturbation that may be marked by changes in DNA methylation that persist as microglia develop. Microglia migration to the brain and colonization of the neural folds during embryogenesis with minimal turnover (Squarzoni, 2015), suggesting that perturbations to microglia in early development may be maintained throughout the lifetime of the animal. Previous work with PolyI:C MIA demonstrated long-lasting alterations in histone modifications (Tang et al., 2013) and DNA methylation (Basil et al., 2014; Richetto et al., 2014, 2016) in adult brain that stem from in utero exposures at mid gestation. While it is unclear if the methylation changes in juvenile MAA microglia are direct or secondary impacts from MAA, the enrichment for numerous immune related pathways including IL-6, IL-4, and IL-8 signaling, NF-kB activation and JAK/STAT signaling, suggests that DMRs may either serve as a signature of prior immune activation during gestation or as epigenetic mechanisms for priming future immune function. The enrichment of DMRs for IL-4 signaling is consistent with the cytokine profile of elevated IL-4 and IL-5 observed in asthma (Magnan et al., 2000; Cho et al., 2005; Tamasi et al., 2005; Kumar et al., 2006), the mid-gestation serum cytokines profiles observed in women who gave birth to a child later diagnosed with ASD (Goines et al., 2011; Jones et al., 2017), and in amniotic fluid taken from children later diagnosed with ASD (Abdallah et al., 2012). Similarly, the enrichment of DMRs for a number of transcription factor motifs critical for early development of microglia (RUNX1, PU.1, and IRF8) indicate that MAA may shift the developmental regulation of microglial function. For example, RUNX1 is expressed in microglial yolk sac progenitors and plays a critical role in regulating microglial proliferation in early brain development (Ginhoux et al., 2010) by ensuring that developing microglia transition from an amoeboid to mature, ramified state during differentiation (Zusso et al., 2012). Similarly, PU.1 is a pioneer transcription factor with DNA methylation sensitive binding (Stephens and Poon, 2016) critical for microglia progenitor development (Kierdorf et al., 2013; Goldmann et al., 2016), regulates microglia specific enhancers (Gosselin et al., 2014) and gene expression (Satoh et al., 2014). PU.1 and shows increased expression in human ASD brain(Gupta et al., 2014), and manipulations of PU.1 alter the expression of key microglial genes and phagocytosis function (Huang et al., 2017), indicating that changes in PU.1 binding could dramatically alter microglial function in ASD. Future studies will be needed to examine the impact of MAA on microglia from different stages of development and in altering the transcriptional response to secondary immune activation (i.e. LPS).
In human ASD brain samples, whole genome analysis has identified alterations in DNA methylation (Nardone et al., 2014; Dunaway et al., 2016b) and gene expression (Voineagu et al., 2011; Gupta et al., 2014; Werling et al., 2016) of a number of immune related genes including components of the complement cascade C1Q, C3, ITGB2 and the cytokine TNFα. Several studies have identified alterations in microglia morphology and density in post-mortem brain samples from ASD patients (Vargas et al., 2005; Morgan et al., 2010, 2012, 2014; Tetreault et al., 2012). In at least a subset of ASD cases, microglia adopt an immune activated morphology characterized by retraction and thickening of processes and enlargement of the soma in multiple brain regions including the dorsolateral prefrontal cortex (Morgan et al., 2010), amygdala (Morgan et al., 2014), and cerebellum (Vargas et al., 2005). Using positron emission tomography and a radiotracer for microglia, Suzuki and colleagues found evidence for increased microglia activation in multiple brain regions in young adults with ASD (Suzuki et al., 2013). Given that immune system related genes are not highly prevalent among the genetic candidates for ASD risk, the alterations in expression and methylation of immune genes in ASD may occur in reaction to abnormal neurodevelopment and/or in response to environmental factors such as maternal immune activation.
The significant overlap of MAA differentially expressed genes with genes differentially expressed or differentially acetylated in ASD brain provides support for the role of maternal immune dysregulation in the etiology of ASD. MAA may also impact some of the same downstream transcriptional pathways that are regulated by MeCP2 and FMRP. While FRMP was not a direct target of MAA in our data, Cytoplasmic FMRP Interaction Protein 2 (Cyfip2) had an intronic hyper-methylated DMR in the MAA microglia. CYIFP2 expression is altered in Fragile X-syndrome (Hoeffer et al., 2012) and defective CYFIP/FMRP interactions have been implicated in ASD and ID (Abekhoukh and Bardoni, 2014) and may mediate some of the overlapping genes targeted by FMRP and regulated by MAA. MeCP2 expression was also not altered by MAA, but genes targeted by MeCP2 binding and genes sensitive to loss of MeCP2 in microglia significantly overlapped MAA genes, indicating MeCP2 may play a role in regulating gene expression differences in MAA microglia.
The vast majority of differentially expressed genes in MAA microglia showed a significant increase in expression relative to PBS controls. While not expected, this increase in expression is consistent with other MIA experiments in whole brain examining acute responses to MIA treatments (Garbett et al., 2012), suggesting an increased sensitivity to environmental signals. For example, we found increased expression of voltage-gated calcium channel subunit Cacna1b, voltage-gated potassium channel subunits (Kcnab1 and Kcnd2), and voltage-gated sodium channel subunits (Scn1a, Scn2a, and Scn2b). Canca1b, Kcnd2 and Scn1a are also ASD candidate genes and increased expression of a number of ligand-gated ion channels upon MAA could lead to altered function (Eder, 1998; Craner et al., 2005; Schilling and Eder, 2007, 2015; Li et al., 2008; Black et al., 2009; Wu et al., 2009; Black and Waxman, 2012; Stebbing et al., 2015). Increased surface receptor expression in MAA microglia would be consistent with motif enrichment of DMRs for transcription factors related to adult homeostatic function in microglia (FOS, MEF2A, MAFB)(Matcovitch-Natan et al., 2016). For example, MAFB expression is induced during microglial maturation, its motif is enriched in macrophage-specific enhancers, and deletion of Mafb specifically in microglia increases gene expression for immune and viral response genes (Matcovitch-Natan et al., 2016). Together these findings suggest that MAA results in long-lasting changes in the ability of microglia to sense and respond to changes in the local brain environment. Altered sensitivity of MAA microglia could potentially dramatically alter the key role of microglia in sculpting neuronal connections and neuronal cell number in the developing brain (Tremblay et al., 2010; Paolicelli et al., 2011; Hagberg et al., 2012; Schafer et al., 2012; Zhan et al., 2014), as further suggested by enrichment of MAA differentially expressed genes for pathways involved in regulation of cell projection organization, neuronal differentiation, and neuronal apoptotic processes (Table S6). Experimentally removing or manipulating microglia in fetal life or during transient postnatal stages has been demonstrated to have profound effects on the number and strength of neuronal synapses, neuronal circuit development, and behavior (Paolicelli et al., 2011; Cunningham et al., 2013; Zhan et al., 2014; Kim et al., 2016). Previous work showed that early life perinatal inflammation (LPS GD15 and 16) augmented fetal microglia activity and decreased neuronal precursors in the developing rat cerebral cortex (Cunningham et al., 2013), leading to impaired corpus callosum fasciculation (Pont-Lezica et al., 2014), dopaminergic axon outgrowth (Squarzoni et al., 2014), and alterations in glutamatergic synapses in the adult offspring (Roumier et al., 2008). Our results show that MAA microglia have significant transcriptional and epigenetic alterations to genes important in axon guidance and receptor function that are consistent with altered sensitivity to the environment and neuronal circuit development.
One potential limitation of our study was a low level of neuronal contamination (0.2-2% in control versus 0.5-4% in MAA, Fig. S1C) in our microglial isolations due to the inherent microglial-neuronal interactions including phagocytosis that occur in brain. While our microglia isolations were above the purity of another published microglial transcriptome study (Zhao et al., 2017), we cannot be certain that the higher expression of some of the typically neuronal genes were cell intrinsic to microglia rather than a result of increased neuronal phagocytosis in the MAA brain (Solga et al., 2015), although either explanation is of interest for understanding the pathogenesis. While the function of many of these neurotransmitter receptors on microglia is largely unknown (Schafer et al., 2013), their overall increase in expression following MAA suggests that microglia have altered sensitivity in the juvenile MAA brain. This is consistent with recent work from a PolyI:C MIA model that showed altered gene expression of cell surface receptors and phagocytosis activity of adult hippocampal microglia (Mattei et al., 2017). Genes misregulated in this model overlapped the MAA differentially expressed genes indicating shared molecular pathways and functions may be disrupted. In addition, MAA impacted genes also overlapped genes responsive to adult minocycline treatment in this MIA model, suggesting minocycline may be able to similarly reverse the behavioral, gene expression and microglial functional deficits in the MAA offspring. Alternatively, many of the receptors showing increased expression in MAA offspring microglia are identified drug targets for a variety of pharmaceuticals (Table S6) that may provide novel options for therapeutics, as is being explored in other neurologic disorders (Eder, 2010).
This work provides a novel link between maternal allergic asthma and changes in the microglial epigenome, indicating that microglia may serve as a potential therapeutic target for normalizing fetal brain connectivity deficits observed in ASD. Removal of microglia from the adult mouse brain by pharmacological treatment did not negatively impact adult behavior (Elmore et al., 2014, 2015) and improved behavioral deficits in an Alzheimer’s disease model (Rice et al., 2015), suggesting microglial contributions to neuro-immune dysfunction can be reversed in the adult. Given the emerging importance of microglia to multiple neuropsychiatric disorders, uncovering the microglial epigenetic landscape is the first step towards understanding how in utero exposures may influence microglia function and dysfunction in later life. Together, our findings present the first whole genome evaluation of microglial DNA methylation and support a role for maternal immune activation in altering microglial function as part of the pathogenesis of ASD.
Supplementary Material
Main Points.
Maternal allergic asthma induces changes in DNA methylation and transcription in juvenile offspring microglia
Differentially methylated regions are enriched for functions and transcription factor binding motifs involved in inflammation and microglial development
Differentially expressed genes and differentially methylated regions are enriched for genes dysregulated in Autism Spectrum Disorders
Acknowledgments
National Institutes of Health T32MH073124-06, 3R01NS081913-11S1, R21HD086669, R21ES025560, IDDRC U54 HD079125 and 5R01NS081913-14, International Rett Syndrome Foundation, Autism Speaks Foundation (#7567), NARSAD foundation, Peter Emch Foundation and the Jane Botsford Johnson Foundation. This work used the Vincent J. Coates Genomics Sequencing Laboratory at UC Berkeley, supported by NIH S10 OD018174 Instrumentation Grant. The fluidigm gene expression experiment was supported by the University of California Davis Flow Cytometry Shared Resource Laboratory with funding from the NCI P30 CA093373, and NIH NCRR C06-RR12088, S10 RR12964 and S10 RR 026825 grants and with technical assistance from Ms. Bridget McLaughlin.
List of Abbreviations
- OVA
Ovalbumin
- MAA
Maternal Allergy/Asthma
- PBS
Phosphate buffered saline
- mCG
methylated CpG
- CHH
methylated CpA or CpC or CpT
Footnotes
Availability of Data and Material
The datasets supporting the conclusions of this article are available in the GEO repository under accession GSE94569. Processed gene lists and enrichments are given in supplemental tables for each experiment.
Authors’ Contributions
A.V.C. prepared libraries for sequencing and conducted bioinformatics analysis. M.C. conducted the maternal allergic asthma procedure, collected, and analyzed purity of microglia. A.V.C, J.L. and P.A. wrote the manuscript.
All authors read and approved the final manuscript.
Compliance with Ethical Standards
The authors declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. All procedures performed in studies involving animals were in accordance with the ethical standards of the University of California, Davis.
Contributor Information
Annie Vogel Ciernia, Medical Microbiology and Immunology, University of California, Davis, Davis, CA 95616.
Milo Careaga, MIND Institute, 2825 50th Street, Sacramento, CA 95817, University of California, Davis.
Paul Ashwood, MIND Institute, 2825 50th Street, Sacramento, CA 95817, University of California, Davis.
Janine LaSalle, Medical Microbiology and Immunology, University of California, Davis, Davis, CA 95616.
References
- Abdallah MW, Larsen N, Grove J, Norgaard-Pedersen B, Thorsen P, Mortensen EL, Hougaard DM. Amniotic fluid chemokines and autism spectrum disorders: an exploratory study utilizing a Danish Historic Birth Cohort. Brain Behav Immun [Internet] 2012;26:170–176. doi: 10.1016/j.bbi.2011.09.003. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21933705. [DOI] [PubMed] [Google Scholar]
- Abekhoukh S, Bardoni B. CYFIP family proteins between autism and intellectual disability: links with Fragile X syndrome. Front Cell Neurosci [Internet] 2014;8:81. doi: 10.3389/fncel.2014.00081. Available from: http://journal.frontiersin.org/article/10.3389/fncel.2014.00081/abstract. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali Z, Ulrik CS. Obesity and asthma: a coincidence or a causal relationship? A systematic review. Respir Med [Internet] 2013a;107:1287–1300. doi: 10.1016/j.rmed.2013.03.019. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23642708. [DOI] [PubMed] [Google Scholar]
- Ali Z, Ulrik CS. Incidence and risk factors for exacerbations of asthma during pregnancy. J Asthma Allergy [Internet] 2013b;6:53–60. doi: 10.2147/JAA.S43183. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23671393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anon. Genome sequencing identifies major causes of severe intellectual disability. Nature [Internet] 2014;511:344–347. doi: 10.1038/nature13394. Available from: http://www.nature.com/doifinder/10.1038/nature13394. [DOI] [PubMed] [Google Scholar]
- Atladottir HO, Pedersen MG, Thorsen P, Mortensen PB, Deleuran B, Eaton WW, Parner ET. Association of family history of autoimmune diseases and autism spectrum disorders. Pediatrics [Internet] 2009;124:687–694. doi: 10.1542/peds.2008-2445. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19581261. [DOI] [PubMed] [Google Scholar]
- Atladottir HO, Thorsen P, Ostergaard L, Schendel DE, Lemcke S, Abdallah M, Parner ET. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J Autism Dev Disord [Internet] 2010;40:1423–1430. doi: 10.1007/s10803-010-1006-y. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20414802. [DOI] [PubMed] [Google Scholar]
- Basil P, Li Q, Dempster EL, Mill J, Sham PC, Wong CC, McAlonan GM. Prenatal maternal immune activation causes epigenetic differences in adolescent mouse brain. Transl Psychiatry [Internet] 2014;4:e434. doi: 10.1038/tp.2014.80. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25180573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Black JA, Liu S, Waxman SG. Sodium channel activity modulates multiple functions in microglia. Glia [Internet] 2009;57:1072–1081. doi: 10.1002/glia.20830. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19115387. [DOI] [PubMed] [Google Scholar]
- Black JA, Waxman SG. Sodium channels and microglial function. Exp Neurol. 2012;234:302–315. doi: 10.1016/j.expneurol.2011.09.030. [DOI] [PubMed] [Google Scholar]
- Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics [Internet] 2014;30:2114–2120. doi: 10.1093/bioinformatics/btu170. Available from: https://academic.oup.com/bioinformatics/article-lookup/doi/10.1093/bioinformatics/btu170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen EY, Tan CM, Kou Y, Duan Q, Wang Z, Meirelles G, Clark NR, Ma’ayan A. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics [Internet] 2013;14:128. doi: 10.1186/1471-2105-14-128. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23586463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chess S. Follow-up report on autism in congenital rubella. J Autism Child Schizophr [Internet] 1977;7:69–81. doi: 10.1007/BF01531116. Available from: http://www.ncbi.nlm.nih.gov/pubmed/576606. [DOI] [PubMed] [Google Scholar]
- Cho SH, Stanciu LA, Holgate ST, Johnston SL. Increased interleukin-4, interleukin-5, and interferon-gamma in airway CD4+ and CD8+ T cells in atopic asthma. Am J Respir Crit Care Med [Internet] 2005;171:224–230. doi: 10.1164/rccm.200310-1416OC. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15502111. [DOI] [PubMed] [Google Scholar]
- Cotney J, Muhle RA, Sanders SJ, Liu L, Willsey AJ, Niu W, Liu W, Klei L, Lei J, Yin J, Reilly SK, Tebbenkamp AT, Bichsel C, Pletikos M, Sestan N, Roeder K, State MW, Devlin B, Noonan JP. The autism-associated chromatin modifier CHD8 regulates other autism risk genes during human neurodevelopment. Nat Commun [Internet] 2015;6:6404. doi: 10.1038/ncomms7404. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25752243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craner MJ, Damarjian TG, Liu S, Hains BC, Lo AC, Black JA, Newcombe J, Cuzner ML, Waxman SG. Sodium channels contribute to microglia/macrophage activation and function in EAE and MS. Glia [Internet] 2005;49:220–229. doi: 10.1002/glia.20112. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15390090. [DOI] [PubMed] [Google Scholar]
- Croen LA, Grether JK, Yoshida CK, Odouli R, Van de Water J. Maternal autoimmune diseases, asthma and allergies, and childhood autism spectrum disorders: a case-control study. Arch Pediatr Adolesc Med. 2005;159:151–157. doi: 10.1001/archpedi.159.2.151. [DOI] [PubMed] [Google Scholar]
- Cronk JC, Derecki NC, Ji E, Xu Y, Lampano AE, Smirnov I, Baker W, Norris GT, Marin I, Coddington N, Wolf Y, Turner SD, Aderem A, Klibanov AL, Harris TH, Jung S, Litvak V, Kipnis J. Methyl-CpG Binding Protein 2 Regulates Microglia and Macrophage Gene Expression in Response to Inflammatory Stimuli. Immunity [Internet] 2015;42:679–691. doi: 10.1016/j.immuni.2015.03.013. Available from: http://linkinghub.elsevier.com/retrieve/pii/S1074761315001272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham CL, Martinez-Cerdeno V, Noctor SC. Microglia Regulate the Number of Neural Precursor Cells in the Developing Cerebral Cortex. J Neurosci [Internet] 2013;33:4216–4233. doi: 10.1523/JNEUROSCI.3441-12.2013. Available from: http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.3441-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darnell JC, Van Driesche SJ, Zhang C, Hung KYS, Mele A, Fraser CE, Stone EF, Chen C, Fak JJ, Chi SW, Licatalosi DD, Richter JD, Darnell RB. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell [Internet] 2011;146:247–261. doi: 10.1016/j.cell.2011.06.013. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21784246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derecki NC, Cronk JC, Lu Z, Xu E, Abbott SBG, Guyenet PG, Kipnis J. Wild type microglia arrest pathology in a mouse model of Rett syndrome. Nature. 2012;484:105–109. doi: 10.1038/nature10907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolinoy DC, Weidman JR, Jirtle RL. Epigenetic gene regulation: linking early developmental environment to adult disease. Reprod Toxicol [Internet] 2007;23:297–307. doi: 10.1016/j.reprotox.2006.08.012. Available from: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17046196. [DOI] [PubMed] [Google Scholar]
- Dunaway K, Goorha S, Matelski L, Urraca N, Lein PJ, Korf I, Reiter LT, LaSalle JM. Dental Pulp Stem Cells Model Early Life and Imprinted DNA Methylation Patterns. Stem Cells [Internet] 2016a doi: 10.1002/stem.2563. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28032673. [DOI] [PMC free article] [PubMed]
- Dunaway KW, Islam MS, Coulson RL, Lopez SJ, Vogel Ciernia A, Chu RG, Yasui DH, Pessah IN, Lott P, Mordaunt C, Meguro-Horike M, Horike S-I, Korf I, LaSalle JM. Cumulative Impact of Polychlorinated Biphenyl and Large Chromosomal Duplications on DNA Methylation, Chromatin, and Expression of Autism Candidate Genes. Cell Rep [Internet] 2016b;17:3035–3048. doi: 10.1016/j.celrep.2016.11.058. Available from: http://dx.doi.org/10.1016/j.celrep.2016.11.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eder C. Ion channels in microglia (brain macrophages) AmJPhysiol [Internet] 1998;275:C327–C342. doi: 10.1152/ajpcell.1998.275.2.C327. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20554024. [DOI] [PubMed] [Google Scholar]
- Eder C. Ion channels in monocytes and microglia / brain macrophages: Promising therapeutic targets for neurological diseases. J Neuroimmunol. 2010;224:51–55. doi: 10.1016/j.jneuroim.2010.05.008. [DOI] [PubMed] [Google Scholar]
- Elmore MRP, Lee RJ, West BL, Green KN. Characterizing Newly Repopulated Microglia in the Adult Mouse: Impacts on Animal Behavior, Cell Morphology, and Neuroinflammation. PLoS One [Internet] 2015;10:e0122912. doi: 10.1371/journal.pone.0122912. Available from: http://dx.plos.org/10.1371/journal.pone.0122912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elmore MRP, Najafi AR, Koike MA, Dagher NN, Spangenberg EE, Rice RA, Kitazawa M, Matusow B, Nguyen H, West BL. Colony-stimulating factor 1 receptor signaling is necessary for microglia viability, unmasking a microglia progenitor cell in the adult brain. Neuron. 2014;82:380–397. doi: 10.1016/j.neuron.2014.02.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng H, Conneely KN, Wu H. A Bayesian hierarchical model to detect differentially methylated loci from single nucleotide resolution sequencing data. Nucleic Acids Res [Internet] 2014;42:e69–e69. doi: 10.1093/nar/gku154. Available from: https://academic.oup.com/nar/article-lookup/doi/10.1093/nar/gku154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garbett KA, Hsiao EY, Kalman S, Patterson PH, Mirnics K. Effects of maternal immune activation on gene expression patterns in the fetal brain. Transl Psychiatry [Internet] 2012;2:e98. doi: 10.1038/tp.2012.24. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22832908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, Mehler MF, Conway SJ, Ng LG, Stanley ER, Samokhvalov IM, Merad M. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science [Internet] 2010;330:841–5. doi: 10.1126/science.1194637. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20966214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goines PE, Croen LA, Braunschweig D, Yoshida CK, Grether J, Hansen R, Kharrazi M, Ashwood P, Van de Water J. Increased midgestational IFN-gamma, IL-4 and IL-5 in women bearing a child with autism: A case-control study. Mol Autism [Internet] 2011;2:13. doi: 10.1186/2040-2392-2-13. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21810230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldmann T, Wieghofer P, Jordão MJC, Prutek F, Hagemeyer N, Frenzel K, Amann L, Staszewski O, Kierdorf K, Krueger M, Locatelli G, Hochgerner H, Zeiser R, Epelman S, Geissmann F, Priller J, Rossi FMV, Bechmann I, Kerschensteiner M, Linnarsson S, Jung S, Prinz M. Origin, fate and dynamics of macrophages at central nervous system interfaces. Nat Immunol [Internet] 2016;17:797–805. doi: 10.1038/ni.3423. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27135602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gosselin D, Link VM, Romanoski CE, Fonseca GJ, Eichenfield DZ, Spann NJ, Stender JD, Chun HB, Garner H, Geissmann F, Glass CK. Environment drives selection and function of enhancers controlling tissue-specific macrophage identities. Cell [Internet] 2014;159:1327–1340. doi: 10.1016/j.cell.2014.11.023. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25480297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunawardhana LP, Baines KJ, Mattes J, Murphy VE, Simpson JL, Gibson PG. Differential DNA methylation profiles of infants exposed to maternal asthma during pregnancy. Pediatr Pulmonol [Internet] 2014;49:852–862. doi: 10.1002/ppul.22930. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24166889. [DOI] [PubMed] [Google Scholar]
- Guo W, Fiziev P, Yan W, Cokus S, Sun X, Zhang MQ, Chen P-Y, Pellegrini M, Cokus S, Feng S, Zhang X, Chen Z, Merriman B, Haudenschild C, Pradhan S, Nelson S, Pellegrini M, Jacobsen S, Lister R, Pelizzola M, Dowen R, Hawkins R, Hon G, Tonti-Filippini J, Nery J, Lee L, Ye Z, Ngo Q-M, Edsall L, Antosiewicz-Bourget J, Stewart R, Ruotti V, Millar A, Thomson J, Ren B, Ecker J, Meissner A, Mikkelsen T, Gu H, Wernig M, Hanna J, Sivachenko A, Zhang X, Bernstein B, Nusbaum C, Jaffe D, Gnirke A, Jaenisch R, Lander E, Wang J, Xia Y, Li L, Gong D, Yao Y, Luo H, Lu H, Yi N, Wu H, Zhang X, Tao Q, Gao F, Chen P-Y, Cokus S, Pellegrini M, Langmead B, Trapnell C, Pop M, Salzberg S, Krueger F, Andrews S, Harris E, Ponts N, Roch K Le, Lonardi S, Pedersen B, Hsieh T-F, Ibarra C, Fischer R, Xi Y, Li W, Smith A, Chung W-Y, Hodges E, Kendall J, Hannon G, Hicks J, Xuan Z, Zhang M, Wu T, Nacu S, Xi Y, Bock C, Müller F, Sun D, Meissner A, Li W, Langmead B, Salzberg S, et al. BS-Seeker2: a versatile aligning pipeline for bisulfite sequencing data. BMC Genomics [Internet] 2013;14:774. doi: 10.1186/1471-2164-14-774. Available from: http://bmcgenomics.biomedcentral.com/articles/10.1186/1471-2164-14-774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta S, Ellis SE, Ashar FN, Moes A, Bader JS, Zhan J, West AB, Arking DE. Transcriptome analysis reveals dysregulation of innate immune response genes and neuronal activity-dependent genes in autism. Nat Commun [Internet] 2014;5:5748. doi: 10.1038/ncomms6748. Available from: http://www.nature.com/doifinder/10.1038/ncomms6748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagberg H, Gressens P, Mallard C. Inflammation during fetal and neonatal life: Implications for neurologic and neuropsychiatric disease in children and adults. Ann Neurol [Internet] 2012;71:444–457. doi: 10.1002/ana.22620. Available from: http://doi.wiley.com/10.1002/ana.22620. [DOI] [PubMed] [Google Scholar]
- Hanamsagar R, Alter MD, Block CS, Sullivan H, Bolton JL, Bilbo SD. Generation of a microglial developmental index in mice and in humans reveals a sex difference in maturation and immune reactivity. Glia [Internet] 2017:1–17. doi: 10.1002/glia.23176. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28618077. [DOI] [PMC free article] [PubMed]
- Hashimoto D, Chow A, Noizat C, Teo P, Beasley MB, Leboeuf M, Becker CD, See P, Price J, Lucas D, Greter M, Mortha A, Boyer SW, Forsberg EC, Tanaka M, van Rooijen N, Garcia-Sastre A, Stanley ER, Ginhoux F, Frenette PS, Merad M. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity [Internet] 2013;38:792–804. doi: 10.1016/j.immuni.2013.04.004. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23601688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinz S, Benner C, Spann N, Bertolino E, Lin YC, Laslo P, Cheng JX, Murre C, Singh H, Glass CK, et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol Cell [Internet] 2010;38:576–89. doi: 10.1016/j.molcel.2010.05.004. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20513432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickman SE, Kingery ND, Ohsumi TK, Borowsky ML, Wang L, Means TK, El Khoury J. The microglial sensome revealed by direct RNA sequencing. Nat Neurosci [Internet] 2013;16:1896–1905. doi: 10.1038/nn.3554. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24162652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoeffer CA, Sanchez E, Hagerman RJ, Mu Y, Nguyen DV, Wong H, Whelan AM, Zukin RS, Klann E, Tassone F. Altered mTOR signaling and enhanced CYFIP2 expression levels in subjects with fragile X syndrome. Genes, Brain Behav [Internet] 2012;11:332–341. doi: 10.1111/j.1601-183X.2012.00768.x. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22268788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holtman IR, Raj DD, Miller JA, Schaafsma W, Yin Z, Brouwer N, Wes PD, Möller T, Orre M, Kamphuis W, Hol EM, Boddeke EWGM, Eggen BJL. Induction of a common microglia gene expression signature by aging and neurodegenerative conditions: a co-expression meta-analysis. Acta Neuropathol Commun [Internet] 2015;3:1–18. doi: 10.1186/s40478-015-0203-5. Available from: http://www.actaneurocomms.org/content/3/1/31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang K, Marcora E, Pimenova AA, Di Narzo AF, Kapoor M, Jin SC, Harari O, Bertelsen S, Fairfax BP, Czajkowski J, Chouraki V, Grenier-Boley B, Bellenguez C, Deming Y, McKenzie A, Raj T, Renton AE, Budde J, Smith A, Fitzpatrick A, Bis JC, DeStefano A, Adams HHH, Ikram MA, van der Lee S, Del-Aguila JL, Fernandez MV, Ibañez L, Sims R, Escott-Price V, Mayeux R, Haines JL, Farrer LA, Pericak-Vance MA, Lambert JC, van Duijn C, Launer L, Seshadri S, Williams J, Amouyel P, Schellenberg GD, Zhang B, Borecki I, Kauwe JSK, Cruchaga C, Hao K, Goate AM, Hao K, Goate AM. A common haplotype lowers PU.1 expression in myeloid cells and delays onset of Alzheimer’s disease. Nat Neurosci [Internet] 2017;20:1052–1061. doi: 10.1038/nn.4587. Available from: http://www.nature.com/doifinder/10.1038/nn.4587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iossifov I, O’roak BJ, Sanders SJ, Ronemus M, Krumm N, Levy D, Stessman HA, Witherspoon KT, Vives L, Patterson KE, Smith JD, Paeper B, Nickerson DA, Dea J, Dong S, Gonzalez LE, Mandell JDJD, Mane SM, Murtha MT, Sullivan CA, Walker MF, Waqar Z, Wei L, Willsey AJ, Yamrom B, Lee Y, Grabowska E, Dalkic E, Wang Z, Marks S, Andrews P, Leotta A, Kendall J, Hakker I, Rosenbaum J, Ma B, Rodgers L, Troge J, Narzisi G, Yoon S, Schatz MC, Ye K, Mccombie WR, Shendure J, Eichler EE, State MW, Wigler M, O’roak BJ, Sanders SJ, Ronemus M, Krumm N, Levy D, Stessman HA, Witherspoon KT, Vives L, Patterson KE, Smith JD, Paeper B, Nickerson DA, Dea J, Dong S, Gonzalez LE, Mandell JDJD, Mane SM, Murtha MT, Sullivan CA, Walker MF, Waqar Z, Wei L, Willsey AJ, Yamrom B, Lee Y, Grabowska E, Dalkic E, Wang Z, Marks S, Andrews P, Leotta A, Kendall J, Hakker I, Rosenbaum J, Ma B, Rodgers L, Troge J, Narzisi G, Yoon S, Schatz MC, Ye K, Mccombie WR, Shendure J, Eichler EE, State MW, Wigler M, Roak BJO, Sanders SJ, Ronemus M, Krumm N, Levy D, et al. The contribution of de novo coding mutations to autism spectrum disorder. Nature [Internet] 2014;0:216–221. doi: 10.1038/nature13908. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25363768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jirtle RL, Skinner MK. Environmental epigenomics and disease susceptibility. Nat Rev Genet [Internet] 2007;8:253–262. doi: 10.1038/nrg2045. Available from: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=17363974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones KL, Croen LA, Yoshida CK, Heuer L, Hansen R, Zerbo O, DeLorenze GN, Kharrazi M, Yolken R, Ashwood P, Van de Water J. Autism with intellectual disability is associated with increased levels of maternal cytokines and chemokines during gestation. Mol Psychiatry [Internet] 2017;22:273–279. doi: 10.1038/mp.2016.77. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27217154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keil A, Daniels JL, Forssen U, Hultman C, Cnattingius S, Soderberg KC, Feychting M, Sparen P. Parental autoimmune diseases associated with autism spectrum disorders in offspring. Epidemiology [Internet] 2010;21:805–808. doi: 10.1097/EDE.0b013e3181f26e3f. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20798635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kierdorf K, Erny D, Goldmann T, Sander V, Schulz C, Gomez Perdiguero E, Wieghofer P, Heinrich A, Riemke P, Hölscher C, Müller DN, Luckow B, Brocker T, Debowski K, Fritz G, Opdenakker G, Diefenbach A, Biber K, Heikenwalder M, Geissmann F, Rosenbauer F, Prinz M. Microglia emerge from erythromyeloid precursors via Pu.1- and Irf8-dependent pathways. Nat Neurosci [Internet] 2013;16:273–280. doi: 10.1038/nn.3318. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23334579. [DOI] [PubMed] [Google Scholar]
- Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol [Internet] 2013;14:R36. doi: 10.1186/gb-2013-14-4-r36. Available from: http://ccb.jhu.edu/software/tophat. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H-J, Cho M-H, Shim WH, Kim JK, Jeon E-Y, Kim D-H, Yoon SY. Deficient autophagy in microglia impairs synaptic pruning and causes social behavioral defects. Mol Psychiatry [Internet] 2016 doi: 10.1038/mp.2016.103. doi: 1–9 Available from: http://www.nature.com/doifinder/10.1038/mp.2016.103. [DOI] [PMC free article] [PubMed]
- Kuleshov MV, Jones MR, Rouillard AD, Fernandez NF, Duan Q, Wang Z, Koplev S, Jenkins SL, Jagodnik KM, Lachmann A, McDermott MG, Monteiro CD, Gundersen GW, Ma’ayan A. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res [Internet] 2016;44:W90–W97. doi: 10.1093/nar/gkw377. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27141961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar RK, Webb DC, Herbert C, Foster PS. Interferon-γ as a possible target in chronic asthma. Inflamm Allergy-Drug Targets (Formerly Curr Drug Targets-Inflammation Allergy) 2006;5:253–256. doi: 10.2174/187152806779010909. [DOI] [PubMed] [Google Scholar]
- Lasalle JM. Epigenomic strategies at the interface of genetic and environmental risk factors for autism. 2013;58:396–401. doi: 10.1038/jhg.2013.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Lu J, Wu CY, Kaur C, Sivakumar V, Sun J, Li S, Ling EA. Expression of Kv1.2 in microglia and its putative roles in modulating production of proinflammatory cytokines and reactive oxygen species. J Neurochem [Internet] 2008;106:2093–2105. doi: 10.1111/j.1471-4159.2008.05559.x. Available from: http://www.ncbi.nlm.nih.gov/pubmed/18627436. [DOI] [PubMed] [Google Scholar]
- Liao Y, Smyth GK, Shi W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics [Internet] 2014;30:923–930. doi: 10.1093/bioinformatics/btt656. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24227677. [DOI] [PubMed] [Google Scholar]
- Libbey JE, Sweeten TL, McMahon WM, Fujinami RS. Autistic disorder and viral infections. J Neurovirol [Internet] 2005;11:1–10. doi: 10.1080/13550280590900553. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15804954. [DOI] [PubMed] [Google Scholar]
- Lister R, Mukamel EA, Nery JR, Urich M, Puddifoot CA, Nicholas D, Lucero J, Huang Y, Dwork AJ, Schultz MD, Yu M, Heyn H, Hu S, Wu JC, Rao A, Esteller M, He C. Global Epigenomic Reconfiguration During Mammalian Brain Development. Science (80-) 2013;341:629–643. doi: 10.1126/science.1237905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lister R, Pelizzola M, Dowen RH, Hawkins RD, Hon G, Tonti-Filippini J, Nery JR, Lee L, Ye Z, Ngo Q-M, Edsall L, Antosiewicz-Bourget J, Stewart R, Ruotti V, Millar AH, Thomson JA, Ren B, Ecker JR. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature [Internet] 2009;462:315–322. doi: 10.1038/nature08514. Available from: http://dx.doi.org/10.1038/nature08514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombardo MV, Moon HM, Su J, Palmer TD, Courchesne E, Pramparo T. Maternal immune activation dysregulation of the fetal brain transcriptome and relevance to the pathophysiology of autism spectrum disorder. Nature Publishing Group; 2017. Available from: http://www.nature.com/doifinder/10.1038/mp.2017.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyall K, Ashwood P, Van de Water J, Hertz-Picciotto I. Maternal immune-mediated conditions, autism spectrum disorders, and developmental delay. J Autism Dev Disord [Internet] 2014;44:1546–1555. doi: 10.1007/s10803-013-2017-2. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24337796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magnan AO, Mely LG, Camilla CA, Badier MM, Montero-Julian FA, Guillot CM, Casano BB, Prato SJ, Fert V, Bongrand P. Assessment of the Th1/Th2 Paradigm in Whole Blood in Atopy and Asthma: Increased IFN-γ–producing CD8+ T Cells in Asthma. Am J Respir Crit Care Med. 2000;161:1790–1796. doi: 10.1164/ajrccm.161.6.9906130. [DOI] [PubMed] [Google Scholar]
- Matcovitch-Natan O, Winter DR, Giladi A, Aguilar SV, Spinrad A, Sarrazin S, Ben-Yehuda H, David E, Gonzalez FZ, Perrin P, Keren-shaul H, Gury M, Lara-Astaiso D, Thaiss CA, Cohen M, Halpern Bahar K, Baruch K, Deczkowska A, Lorenzo-Vivas E, Itzkovitz S, Elinav E, Sieweke MH, Schwartz M, Amit I. Microglia development follows a stepwise program to regulate brain homeostasis. Science (80-) 2016;8670 doi: 10.1126/science.aad8670. [DOI] [PubMed] [Google Scholar]
- Mattei D, Ivanov A, Ferrai C, Jordan P, Guneykaya D, Buonfiglioli A, Schaafsma W, Przanowski P, Deuther-Conrad W, Brust P, Hesse S, Patt M, Sabri O, Ross TL, Eggen BJL, Boddeke EWGM, Kaminska B, Beule D, Pombo A, Kettenmann H, Wolf SA. Maternal immune activation results in complex microglial transcriptome signature in the adult offspring that is reversed by minocycline treatment. Transl Psychiatry [Internet] 2017;7:e1120. doi: 10.1038/tp.2017.80. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28485733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLean CY, Bristor D, Hiller M, Clarke SL, Schaar BT, Lowe CB, Wenger AM, Bejerano G. GREAT improves functional interpretation of cis-regulatory regions. Nat Biotechnol [Internet] 2010;28:495–501. doi: 10.1038/nbt.1630. Available from: http://bejerano.stanford.edu/papers/GREAT.pdf. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mo A, Mukamel EA, Davis FP, Luo C, Henry GL, Picard S, Urich MA, Nery JR, Sejnowski TJ, Lister R, Eddy SR, Ecker JR, Nathans J. Epigenomic Signatures of Neuronal Diversity in the Mammalian Brain. Neuron [Internet] 2015;86:1369–1384. doi: 10.1016/j.neuron.2015.05.018. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0896627315004250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan JT, Barger N, Amaral DG, Schumann CM. Stereological Study of Amygdala Glial Populations in Adolescents and Adults with Autism Spectrum Disorder. PLoS One [Internet] 2014;9:e110356. doi: 10.1371/journal.pone.0110356. Available from: http://dx.plos.org/10.1371/journal.pone.0110356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan JT, Chana G, Abramson I, Semendeferi K, Courchesne E, Everall IP. Abnormal microglial-neuronal spatial organization in the dorsolateral prefrontal cortex in autism. Brain Res [Internet] 2012;1456:72–81. doi: 10.1016/j.brainres.2012.03.036. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22516109. [DOI] [PubMed] [Google Scholar]
- Morgan JT, Chana G, Pardo CA, Achim C, Semendeferi K, Buckwalter J, Courchesne E, Everall IP. Microglial activation and increased microglial density observed in the dorsolateral prefrontal cortex in autism. Biol Psychiatry [Internet] 2010;68:368–376. doi: 10.1016/j.biopsych.2010.05.024. Available from: http://www.ncbi.nlm.nih.gov/pubmed/20674603. [DOI] [PubMed] [Google Scholar]
- Mostafa GA, Kitchener N. Serum anti-nuclear antibodies as a marker of autoimmunity in Egyptian autistic children. Pediatr Neurol [Internet] 2009;40:107–112. doi: 10.1016/j.pediatrneurol.2008.10.017. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19135624. [DOI] [PubMed] [Google Scholar]
- Mouridsen SE, Rich B, Isager T, Nedergaard NJ. Autoimmune diseases in parents of children with infantile autism: a case-control study. Dev Med Child Neurol [Internet] 2007;49:429–432. doi: 10.1111/j.1469-8749.2007.00429.x. Available from: http://www.ncbi.nlm.nih.gov/pubmed/17518928. [DOI] [PubMed] [Google Scholar]
- Murphy VE, Clifton VL, Gibson PG. Asthma exacerbations during pregnancy: incidence and association with adverse pregnancy outcomes. Thorax [Internet] 2006;61:169–176. doi: 10.1136/thx.2005.049718. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16443708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy VE, Gibson P, Talbot PI, Clifton VL. Severe asthma exacerbations during pregnancy. Obs Gynecol [Internet] 2005;106:1046–1054. doi: 10.1097/01.AOG.0000185281.21716.02. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16260524. [DOI] [PubMed] [Google Scholar]
- Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol [Internet] 2011;11:723–737. doi: 10.1038/nri3073. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21997792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nardone S, Sams DS, Reuveni E, Getselter D, Oron O, Karpuj M, Elliott E. DNA methylation analysis of the autistic brain reveals multiple dysregulated biological pathways. Transl Psychiatry [Internet] 2014;4:e433–9. doi: 10.1038/tp.2014.70. Available from: http://dx.doi.org/10.1038/tp.2014.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Needleman LA, Mcallister AK. The major histocompatibility complex and autism spectrum disorder. Dev Neurobiol [Internet] 2012;72:1288–1301. doi: 10.1002/dneu.22046. Available from: http://doi.wiley.com/10.1002/dneu.22046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nials AT, Uddin S. Mouse models of allergic asthma: acute and chronic allergen challenge. Dis Model Mech [Internet] 2008;1:213–220. doi: 10.1242/dmm.000323. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19093027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oskvig DB, Elkahloun AG, Johnson KR, Phillips TM, Herkenham M. Maternal immune activation by LPS selectively alters specific gene expression profiles of interneuron migration and oxidative stress in the fetus without triggering a fetal immune response. Brain Behav Immun [Internet] 2012;26:623–634. doi: 10.1016/j.bbi.2012.01.015. Available from: http://www.sciencedirect.com/science/article/pii/S0889159112000177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P, Giustetto M, Ferreira TA, Guiducci E, Dumas L, Ragozzino D, Gross CT. Synaptic Pruning by Microglia Is Necessary for Normal Brain Development. Science (80-) 2011;333:1456–1458. doi: 10.1126/science.1202529. [DOI] [PubMed] [Google Scholar]
- Parikshak NN, Swarup V, Belgard TG, Irimia M, Ramaswami G, Gandal MJ, Hartl C, Leppa V, Ubieta L de la T, Huang J, Lowe JK, Blencowe BJ, Horvath S, Geschwind DH. Genome-wide changes in lncRNA, splicing, and regional gene expression patterns in autism. Nature [Internet] 2016;540:423–427. doi: 10.1038/nature20612. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27919067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pont-Lezica L, Beumer W, Colasse S, Drexhage H, Versnel M, Bessis A. Microglia shape corpus callosum axon tract fasciculation: functional impact of prenatal inflammation. Eur J Neurosci [Internet] 2014;39:1–7. doi: 10.1111/ejn.12508. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24593277. [DOI] [PubMed] [Google Scholar]
- Rice RA, Spangenberg EE, Yamate-Morgan H, Lee RJ, Arora RPS, Hernandez MX, Tenner AJ, West BL, Green KN. Elimination of Microglia Improves Functional Outcomes Following Extensive Neuronal Loss in the Hippocampus. J Neurosci [Internet] 2015;35:9977–9989. doi: 10.1523/JNEUROSCI.0336-15.2015. Available from: http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.0336-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richetto J, Calabrese F, Riva MA, Meyer U. Prenatal immune activation induces maturation-dependent alterations in the prefrontal GABAergic transcriptome. Schizophr Bull [Internet] 2014;40:351–361. doi: 10.1093/schbul/sbs195. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23328159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richetto J, Massart R, Weber-Stadlbauer U, Szyf M, Riva MA, Meyer U. Genome-Wide DNA Methylation Changes in a Mouse Model of Infection-Mediated Neurodevelopmental Disorders. Biol Psychiatry [Internet] 2016;81:1–12. doi: 10.1016/j.biopsych.2016.08.010. Available from: http://linkinghub.elsevier.com/retrieve/pii/S0006322316326713. [DOI] [PubMed] [Google Scholar]
- Robinson M, Oshlack A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol [Internet] 2010;11:R25. doi: 10.1186/gb-2010-11-3-r25. Available from: http://genomebiology.com/2010/11/3/R25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics [Internet] 2010;26:139–40. doi: 10.1093/bioinformatics/btp616. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19910308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roumier A, Pascual O, Béchade C, Wakselman S, Poncer J-C, Réal E, Triller A, Bessis A. Prenatal Activation of Microglia Induces Delayed Impairment of Glutamatergic Synaptic Function. PLoS One [Internet] 2008;3:e2595. doi: 10.1371/journal.pone.0002595. Available from: http://dx.plos.org/10.1371/journal.pone.0002595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rube HT, Lee W, Hejna M, Chen H, Yasui DH, Hess JF, LaSalle JM, Song JS, Gong Q. Sequence features accurately predict genome-wide MeCP2 binding in vivo. Nat Commun [Internet] 2016;7:11025. doi: 10.1038/ncomms11025. Available from: http://www.nature.com/ncomms/2016/160324/ncomms11025/full/ncomms11025.html#supplementary-information. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders SJ, He X, Willsey AJ, Devlin B, Roeder K, State MW, Sanders SJ, He X, Willsey AJ, Ercan-sencicek AG, Samocha KE. Insights into Autism Spectrum Disorder Genomic Architecture and Biology from 71 Risk Loci. Neuron [Internet] 2015;87:1215–1233. doi: 10.1016/j.neuron.2015.09.016. Available from: http://dx.doi.org/10.1016/j.neuron.2015.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders SJ, Murtha MT, Gupta AR, Murdoch JD, Raubeson MJ, Willsey AJ, Ercan-Sencicek AG, DiLullo NM, Parikshak NN, Stein JL, Walker MF, Ober GT, Teran NA, Song Y, El-Fishawy P, Murtha RC, Choi M, Overton JD, Bjornson RD, Carriero NJ, Meyer KA, Bilguvar K, Mane SM, Sestan N, Lifton RP, Günel M, Roeder K, Geschwind DH, Devlin B, State MW, Šestan N, Lifton RP, Günel M, Roeder K, Geschwind DH, Devlin B, State MW. De novo mutations revealed by whole-exome sequencing are strongly associated with autism. Nature [Internet] 2012;485:237–41. doi: 10.1038/nature10945. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3667984&tool=pmcentrez&rendertype=abstract. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satoh J, Asahina N, Kitano S, Kino Y. A Comprehensive Profile of ChIP-Seq-Based PU.1/Spi1 Target Genes in Microglia. Gene Regul Syst Bio [Internet] 2014;8:127. doi: 10.4137/GRSB.S19711. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25574134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mardinly AR, Yamasaki R, Ransohoff RM, Greenberg ME, Barres BA, Stevens B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron [Internet] 2012;74:691–705. doi: 10.1016/j.neuron.2012.03.026. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22632727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer DP, Lehrman EK, Stevens B. The “quad-partite” synapse: Microglia-synapse interactions in the developing and mature CNS. Glia [Internet] 2013;61:24–36. doi: 10.1002/glia.22389. Available from: http://doi.wiley.com/10.1002/glia.22389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schafer DP, Stevens B. Brains, Blood, and Guts: MeCP2 Regulates Microglia, Monocytes, and Peripheral Macrophages. Immunity [Internet] 2015;42:600–602. doi: 10.1016/j.immuni.2015.04.002. Available from: http://dx.doi.org/10.1016/j.immuni.2015.04.002. [DOI] [PubMed] [Google Scholar]
- Schatz M, Harden K, Forsythe A, Chilingar L, Hoffman C, Sperling W, Zeiger RS. The course of asthma during pregnancy, post partum, and with successive pregnancies: a prospective analysis. J Allergy Clin Immunol [Internet] 1988;81:509–517. Available from: http://www.ncbi.nlm.nih.gov/pubmed/3346481. [PubMed] [Google Scholar]
- Schilling T, Eder C. Ion channel expression in resting and activated microglia of hippocampal slices from juvenile mice. Brain Res. 2007;1186:21–28. doi: 10.1016/j.brainres.2007.10.027. [DOI] [PubMed] [Google Scholar]
- Schilling T, Eder C. Microglial K+ channel expression in young adult and aged mice. Glia [Internet] 2015;63:664–672. doi: 10.1002/glia.22776. Available from: http://doi.wiley.com/10.1002/glia.22776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartzer JJ, Careaga M, Chang C, Onore CE, Ashwood P. Allergic fetal priming leads to developmental, behavioral and neurobiological changes in mice. Transl Psychiatry [Internet] 2015;5:e543. doi: 10.1038/tp.2015.40. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25849982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartzer JJ, Careaga M, Coburn MA, Rose DR, Hughes HK, Ashwood P. Behavioral impact of maternal allergic-asthma in two genetically distinct mouse strains. Brain Behav Immun [Internet] 2017;36:99–107. doi: 10.1016/j.bbi.2016.09.007. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27622677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarz JM, Sholar PW, Bilbo SD. Sex differences in microglial colonization of the developing rat brain. J Neurochem. 2012;120:948–963. doi: 10.1111/j.1471-4159.2011.07630.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen L. GeneOverlap: An R package to test and visualize gene overlaps. 2013 Available from: http://shenlab-sinai.github.io/shenlab-sinai/
- Solga AC, Pong WW, Walker J, Wylie T, Magrini V, Apicelli AJ, Griffith M, Griffith OL, Kohsaka S, Wu GF, Brody DL, Mardis ER, Gutmann DH. RNA-sequencing reveals oligodendrocyte and neuronal transcripts in microglia relevant to central nervous system disease. Glia [Internet] 2015;63:531–548. doi: 10.1002/glia.22754. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25258010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Squarzoni P. Neuronal and microglial regulators of cortical wiring: usual and novel guideposts. Front Neurosci [Internet] 2015;9:1–16. doi: 10.3389/fnins.2015.00248. Available from: http://journal.frontiersin.org/Article/10.3389/fnins.2015.00248/abstract. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Squarzoni P, Oller G, Hoeffel G, Pont-Lezica L, Rostaing P, Low D, Bessis A, Ginhoux F, Garel S. Microglia Modulate Wiring of the Embryonic Forebrain. Cell Rep [Internet] 2014;8:1271–1279. doi: 10.1016/j.celrep.2014.07.042. Available from: http://linkinghub.elsevier.com/retrieve/pii/S2211124714006263. [DOI] [PubMed] [Google Scholar]
- Stebbing MJ, Cottee JM, Rana I. The role of ion channels in microglial activation and proliferation - a complex interplay between ligand-gated ion channels, K+ channels, and intracellular Ca2+ Front Immunol [Internet] 2015;6:497. doi: 10.3389/fimmu.2015.00497. Available from: http://journal.frontiersin.org/Article/10.3389/fimmu.2015.00497/abstract. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephens DC, Poon GMK. Differential sensitivity to methylated DNA by ETS-family transcription factors is intrinsically encoded in their DNA-binding domains. Nucleic Acids Res [Internet] 2016;44:8671–8681. doi: 10.1093/nar/gkw528. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27270080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugathan A, Biagioli M, Golzio C, Erdin S, Blumenthal I, Manavalan P, Ragavendran A, Brand H, Lucente D, Miles J, Sheridan SD, Stortchevoi A, Kellis M, Haggarty SJ, Katsanis N, Gusella JF, Talkowski ME. CHD8 regulates neurodevelopmental pathways associated with autism spectrum disorder in neural progenitors. Proc Natl Acad Sci [Internet] 2014;111:E4468–E4477. doi: 10.1073/pnas.1405266111. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25294932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun W, Poschmann J, Cruz-Herrera del Rosario R, Parikshak NN, Hajan HS, Kumar V, Ramasamy R, Belgard TG, Elanggovan B, Wong CCY, Mill J, Geschwind DH, Prabhakar S. Histone Acetylome-wide Association Study of Autism Spectrum Disorder. Cell [Internet] 2016;167:1385–1397. doi: 10.1016/j.cell.2016.10.031. e11. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27863250. [DOI] [PubMed] [Google Scholar]
- Supek F, Bošnjak M, Škunca N, Šmuc T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS One [Internet] 2011;6:e21800. doi: 10.1371/journal.pone.0021800. Available from: http://dx.plos.org/10.1371/journal.pone.0021800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki K, Sugihara G, Ouchi Y, Nakamura K, Futatsubashi M, Takebayashi K, Yoshihara Y, Omata K, Matsumoto K, Tsuchiya KJ, Iwata Y, Tsujii M, Sugiyama T, Mori N. Microglial activation in young adults with autism spectrum disorder. JAMA Psychiatry [Internet] 2013;70:49–58. doi: 10.1001/jamapsychiatry.2013.272. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23404112. [DOI] [PubMed] [Google Scholar]
- Sweeten TL, Posey DJ, McDougle CJ. Brief report: autistic disorder in three children with cytomegalovirus infection. J Autism Dev Disord [Internet] 2004;34:583–586. doi: 10.1007/s10803-004-2552-y. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15628611. [DOI] [PubMed] [Google Scholar]
- Tamasi L, Bohacs A, Pallinger E, Falus A, Rigo J, Jr, Muller V, Komlosi Z, Magyar P, Losonczy G. Increased interferon-gamma- and interleukin-4-synthesizing subsets of circulating T lymphocytes in pregnant asthmatics. Clin Exp Allergy [Internet] 2005;35:1197–1203. doi: 10.1111/j.1365-2222.2005.02322.x. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16164448. [DOI] [PubMed] [Google Scholar]
- Tang B, Jia H, Kast RJ, Thomas EA. Epigenetic changes at gene promoters in response to immune activation in utero. Brain Behav Immun [Internet] 2013;30:168–175. doi: 10.1016/j.bbi.2013.01.086. Available from: http://www.ncbi.nlm.nih.gov/pubmed/23402795. [DOI] [PubMed] [Google Scholar]
- Tetreault NA, Hakeem AY, Jiang S, Williams BA, Allman E, Wold BJ, Allman JM. Microglia in the Cerebral Cortex in Autism. J Autism Dev Disord [Internet] 2012;42:2569–2584. doi: 10.1007/s10803-012-1513-0. Available from: http://link.springer.com/10.1007/s10803-012-1513-0. [DOI] [PubMed] [Google Scholar]
- Thomas A, Burant A, Bui N, Graham D, Yuva-Paylor LA, Paylor R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacol [Internet] 2009;204:361–373. doi: 10.1007/s00213-009-1466-y. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19189082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tremblay M-È, Lowery RL, Majewska AK. Microglial Interactions with Synapses Are Modulated by Visual Experience. PLoS Biol [Internet] 2010;8:e1000527. doi: 10.1371/journal.pbio.1000527. Available from: http://dx.plos.org/10.1371/journal.pbio.1000527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann Neurol [Internet] 2005;57:67–81. doi: 10.1002/ana.20315. Available from: http://www.ncbi.nlm.nih.gov/pubmed/15546155. [DOI] [PubMed] [Google Scholar]
- Vogel Ciernia A, LaSalle J. The landscape of DNA methylation amid a perfect storm of autism aetiologies. Nat Rev Neurosci [Internet] 2016;6:411–423. doi: 10.1038/nrn.2016.41. Available from: http://www.nature.com/doifinder/10.1038/nrn.2016.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voineagu I, Wang X, Johnston P, Lowe JK, Tian Y, Horvath S, Mill J, Cantor RM, Blencowe BJ, Geschwind DH. Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature [Internet] 2011;474:380–384. doi: 10.1038/nature10110. Available from: http://dx.doi.org/10.1038/nature10110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werling DM, Parikshak NN, Geschwind DH. Gene expression in human brain implicates sexually dimorphic pathways in autism spectrum disorders. Nat Commun [Internet] 2016;7:10717. doi: 10.1038/ncomms10717. Available from: http://www.nature.com/doifinder/10.1038/ncomms10717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitehead GS, Walker JK, Berman KG, Foster WM, Schwartz DA. Allergen-induced airway disease is mouse strain dependent. Am J Physiol Lung Cell Mol Physiol [Internet] 2003;285:L32–42. doi: 10.1152/ajplung.00390.2002. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12626335. [DOI] [PubMed] [Google Scholar]
- Wu CY, Kaur C, Sivakumar V, Lu J, Ling EA. Kv1.1 expression in microglia regulates production and release of proinflammatory cytokines, endothelins and nitric oxide. Neuroscience [Internet] 2009;158:1500–1508. doi: 10.1016/j.neuroscience.2008.11.043. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19118603. [DOI] [PubMed] [Google Scholar]
- Zerbo O, Iosif AM, Walker C, Ozonoff S, Hansen RL, Hertz-Picciotto I. Is maternal influenza or fever during pregnancy associated with autism or developmental delays? Results from the CHARGE (CHildhood Autism Risks from Genetics and Environment) study. J Autism Dev Disord [Internet] 2013;43:25–33. doi: 10.1007/s10803-012-1540-x. Available from: http://www.ncbi.nlm.nih.gov/pubmed/22562209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zerbo O, Leong A, Barcellos L, Bernal P, Fireman B, Croen LA. Immune mediated conditions in autism spectrum disorders. Brain Behav Immun [Internet] 2015;46:232–6. doi: 10.1016/j.bbi.2015.02.001. Available from: http://www.sciencedirect.com/science/article/pii/S0889159115000197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhan Y, Paolicelli RC, Sforazzini F, Weinhard L, Bolasco G, Pagani F, Vyssotski AL, Bifone A, Gozzi A, Ragozzino D, Gross CT. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat Neurosci. 2014;17:400–406. doi: 10.1038/nn.3641. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O’Keeffe S, Phatnani HP, Guarnieri P, Caneda C, Ruderisch N, Deng S, Liddelow SA, Zhang C, Daneman R, Maniatis T, Barres BA, Wu JQ. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci [Internet] 2014;34:11929–11947. doi: 10.1523/JNEUROSCI.1860-14.2014. Available from: http://www.ncbi.nlm.nih.gov/pubmed/25186741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao D, Mokhtari R, Pedrosa E, Birnbaum R, Zheng D, Lachman HM. Transcriptome analysis of microglia in a mouse model of Rett syndrome: differential expression of genes associated with microglia/macrophage activation and cellular stress. Mol Autism [Internet] 2017;8:17. doi: 10.1186/s13229-017-0134-z. Available from: http://molecularautism.biomedcentral.com/articles/10.1186/s13229-017-0134-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zusso M, Methot L, Lo R, Greenhalgh AD, David S, Stifani S. Regulation of Postnatal Forebrain Amoeboid Microglial Cell Proliferation and Development by the Transcription Factor Runx1. J Neurosci [Internet] 2012;32:11285–11298. doi: 10.1523/JNEUROSCI.6182-11.2012. Available from: http://www.jneurosci.org/content/32/33/11285. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.