

Abstract
Background
The circadian clock, which operates within an approximately 24-h period, is closely linked to the survival and fitness of almost all living organisms. The circadian clock is generated through a negative transcription-translation feedback loop. microRNAs (miRNAs) are small non-coding RNAs comprised of approximately 22 nucleotides that post-transcriptionally regulate target mRNA by either inducing mRNA degradation or inhibiting translation.
Results
In recent years, miRNAs have been found to play important roles in the regulation of the circadian clock, especially in Drosophila. In this review, we will use fruit flies as an example, and summarize the progress achieved in the study of miRNA-mediated clock regulation. Three main aspects of the circadian clock, namely, the free-running period, locomotion phase, and circadian amplitude, are discussed in detail in the context of how miRNAs are involved in these regulations. In addition, approaches regarding the discovery of circadian-related miRNAs and their targets are also discussed.
Conclusions
Research in the last decade suggests that miRNA-mediated post-transcriptional regulation is crucial to the generation and maintenance of a robust circadian clock in animals. In flies, miRNAs are known to modulate circadian rhythmicity and the free-running period, as well as circadian outputs. Further characterization of miRNAs, especially in the circadian input, will be a vital step toward a more comprehensive understanding of the functions underlying miRNA-control of the circadian clock.
Keywords: Circadian clock, Drosophila, miRNA, Post-transcriptional regulation
Introduction
Circadian rhythms with approximately 24-h periods are prevalent in all living organisms, from unicellular cyanobacteria to vertebrates. This endogenous timekeeping system endows organisms with the enormous advantage of anticipating and adjusting to environmental changes (e.g., light, food, temperature, etc.) [1]. Many physiological and behavioral processes, including hormone secretion, thermoregulation, metabolism, immune responses, and sleep–wake cycles, are modulated by the circadian clock [2–5]. Therefore, it is not surprising that disruption of the circadian clock is associated with pathogenesis, such as obesity, cardiovascular diseases, and even cancer [6–8]. Since the circadian clock plays a fundamental role in the maintenance of circadian-dependent processes, understanding the regulatory mechanism of the circadian clock is vitally important.
The clock’s machinery consists of exogenous, stimulus-sensitive input pathways, a central clock pacemaker capable of coordinating various endogenous pacemakers, and output pathways that produce overt rhythms [9]. The molecular components of the endogenous clock comprise a transcriptional-translational feedback loop (TTFL) that is fairly well-understood in mammals and flies. In the TTFL, the generation of rhythmic mRNA and protein abundance arises from repression- and activation-mediated temporal and spatial delays that eventually control downstream rhythmic behavioral patterns and physiological functions within an approximate 24-h period (Fig. 1).
Fig. 1.
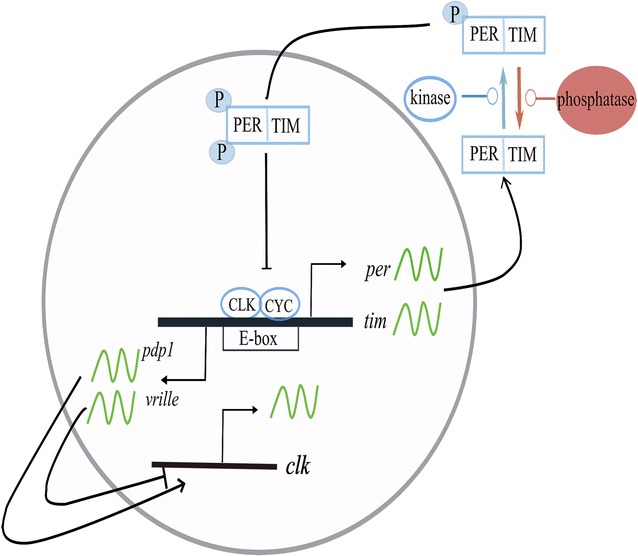
Circadian feedback regulatory loop in Drosophila melanogaster. In the per/tim loop, the CLK/CYC heterodimer binds to the E-box region of per and tim to activate their transcription. PER and TIM accumulate in the cytoplasm and form another complex at night. Phosphorylation of PER/TIM by kinases promotes their nuclear entry, which is counteracted by the dephosphorylation performed by phosphatases. The PER/TIM complex moves into the nucleus later in the night, and hyperphosphorylated PER/TIM represses CLK/CYC activity by promoting the phosphorylation of CLK. In the clk/cyc loop, CLK/CYC promotes the transcription of pdp1 and vri, which encode PDP1 and VRI transcription factors to activate and repress clk transcription, respectively. VRI inhibition precedes PDP1 activation in a way that causes clk mRNA to oscillate during the day. CLK—CLOCK, CYC—CYCLE, PER—PERIOD, TIM—TIMELESS, pdp1—PAR domain protein 1, VRI—VRILLE
Drosophila has been instrumental in the study of circadian rhythms because of powerful genetic tools and their relatively simple circadian neuronal circuitry. After more than four decades of studies in Drosophila, we now have a deep understanding of the molecular components of the circadian clock that play a pivotal role in coordinating circadian readouts to environmental inputs. Thus, in this review we will focus on the circadian rhythms of Drosophila.
In flies, approximately 150 circadian neurons make up the clock network in the brain. This network can be categorized into seven subclasses based on anatomical location, cellular size, and neuropeptide expression: the small and large ventral lateral neurons (LNvs) expressing pigment dispersing factor (PDF), the fifth small LNv, the dorsal lateral neurons (LNds), and three groups of dorsal neurons (DN1, DN2, and DN3) [10]. The core proteins of the TTFL, encoded by clock (clk), cycle (cyc), period (per), and timeless (tim), are expressed in all clock neuron groups, but transcriptome studies of individual circadian neurons have identified that most cycling transcripts are expressed only in a specific subgroup [11], which is likely due to different regulatory mechanisms. Moreover, next-generation sequencing (NGS) results have demonstrated that approximately 50% of rhythmic primary transcripts do not give rise to rhythmic mRNA, and rhythmic proteins do not necessarily arise from rhythmic mRNA transcripts [12, 13]. For these reasons, we sought to explore when, where and how post-transcriptional modification is integrated into the circadian control system. This review will focus on miRNA-dependent post-transcriptional regulation, due to accumulating evidence demonstrating the involvement of miRNAs in the regulation of circadian timekeeping.
miRNAs are small non-coding RNAs of approximately 22 nucleotides (nt) that are generated by consecutive cleavage by the endoribonucleases, Drosha and Dicer. Drosha, an RNase III enzyme, recognizes primary miRNA (pri-mRNA) and cleaves it into an approximately 70-nt precursor miRNA (pre-miRNA) in the nucleus. Subsequently, pre-miRNA is exported into the cytoplasm and is subjected to Dicer-mediated cleavage that results in a dual miRNA products. One of the miRNA strands is then degraded and the other is loaded into the RNA-induced silencing complex (RISC). miRNA in RISC promotes degradation and attenuates translation by imperfect complementary binding to the non-coding 3′ untranslated region (UTR) of the target mRNA [14]. The most important determinant of mRNA expression in miRNA depends on the “seed sequence”, located at nucleotides 2–7 at the 5′ end of miRNA, which is used for the perfect complementary binding [15]. miRNAs regulate most physiological and biological pathways and processes [16], and circadian rhythms are no exception. In this review, we highlight the current knowledge of miRNA-mediated regulation of rhythmicity, the free-running period, and diurnal phase (Fig. 2). Furthermore, we also discuss approaches regarding the identification of circadian-relevant miRNAs.
Fig. 2.
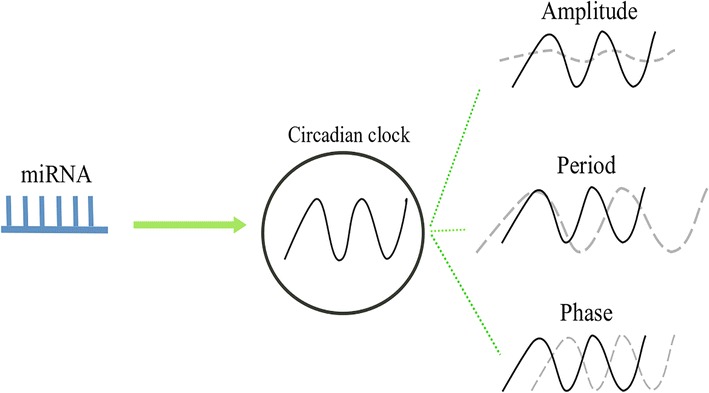
miRNA-mediated post-transcriptional regulation in different aspects of the circadian clock. Rhythmicity, period, and phase are important parameters of the circadian clock. Rhythmicity reflects the capability of flies to maintain the robust circadian clock; period reflects the speed of the circadian clock; and phase reflects the synchronization of the clock to the local environment. Solid black lines represent the oscillation of the clock. Dotted gray lines represent the miRNA-mediated corresponding changes to the clock
Transcriptional-translational feedback loops
The endogenous circadian clock maintains a free-running (in the absence of rhythmic environmental stimulus) period of approximately 24 h by employing interconnected feedback loops that typically include a primary loop and an ancillary loop. In flies, the primary negative feedback loop is composed of transcriptional activators (CLK/CYC) and repressors (PER/TIM) in which heterodimeric transactivators bind to promotor regions to induce repressor expression. Following 6–8 h of cytosolic repressor accumulation and modification, heterodimerization occurs, and the repressors translocate to the nucleus, inhibiting their own transcriptional activity by interacting with activators. Consequently, the levels of repressor complexes drop, and a new cycle of transactivation starts. The ancillary feedback loop shares core transactivators with the primary loop but differ in that Vrille (VRI) and Par domain protein 1 (PDP1) repress and activate clk transcription, respectively (Fig. 1). The ancillary feedback loop is believed to stabilize the molecular clock by regulating the phase and amplitude of core pacemakers and rhythmic outputs [17, 18].
In addition to the transcriptional control of the molecular clock, post-translational regulation is built into the TTFL to regulate protein stability, activity, and subcellular localization. Among different post-translational regulations, phosphorylation is one of the most interesting, as phosphorylation levels of principal clock proteins oscillate in a temporal pattern. For example, PER protein phosphorylation peaks in the early morning/late evening, and dPER is hypophosphorylated in the early evening [19]. The kinases involved in the phosphorylation of the PER/TIM complex are glycogen synthase kinase-3 [GSK3; also known as Shaggy (SGG)], casein kinase II (CK2), and DOUBLETIME (DBT; Drosophila homolog of mammalian CK1δ/ε) [20–23]. There are also several important phosphatases that dephosphorylate PER and TIM, such as Protein Phosphatase 1 (PP1) and Protein Phosphatase 2A (PP2A) [24, 25] (Fig. 1).
Post-transcriptional regulation
The temporal delay between mRNA and protein oscillation of core clock genes indicates the existence of a layer of post-transcriptional regulation. When RNA sequencing and nascent RNA sequencing were applied to assess the whole genome circadian gene expression in fly heads, it was observed that ~ 77% strong, rhythmic, and mature mRNAs were derived from weak, non-oscillating nascent transcripts, suggesting that post-transcriptional modification has a major role in circadian mRNA expression12. This builds upon prior research that showed that only approximately 22% of cycling mRNAs exhibit phases identical to those of their de novo transcripts, and certain circadian-related genes were even found to display constant mRNA levels over the course of a day [26]. Together, these data strongly suggest that post-transcriptional regulation is essential for circadian gene expression.
Although post-transcriptional mechanisms, including alternative splicing, mRNA modification, poly(A) tail length, and alternative polyadenylation [27–30], are known to regulate cyclic gene expression, mRNA stability has been the main focus in the study of post-transcriptional regulation of circadian rhythms. Not surprisingly, mRNAs with short half-lives are more likely to display significant oscillations in expression than mRNAs with long half-lives are. For example, a handful of core clock genes in the TTFL, namely, per, tim, and clk, show mRNA oscillations because of variable half-lives, which are likely to be mediated by post-transcriptional regulation. Many studies have demonstrated that mRNA stability depends upon environment and on developmental stage [31], but the most crucial regulatory factors include cis elements in mRNA structure and trans-acting factors (RNA-binding proteins, RBP), such as LARK, for example, which is an RBP that is a key regulator of translation of dbt [32].
A recent study showed that deletion of the 3′UTR of clk mRNA in Drosophila led to ectopic expression of CLK, generation of additional PDF-expressing neurons, and the induction of variable circadian behavior [33]. In addition, deletion of binding sites in clk for the miRNA bantam resulted in a similar phenotype characterized by ectopic expression of VRI and overgrowth of PDF-positive neurons, suggesting that miRNAs regulate the development of circadian neurons [33].
Roles of miRNA biogenesis in circadian regulation
Microarray-based experiments were used to identify miRNAs that show circadian oscillations in the fly head. Several conserved miRNAs, such as miR-263a/b, for instance, show strong daily oscillations [34]. Further analysis demonstrated that, in clock mutants, miR-263a/b have attenuated phases but elevated levels [34].
Later, studies demonstrated correlations between miRNA biogenesis and circadian rhythms. To monitor circadian behavior, flies are usually entrained by several 12-h light/12-h dark (LD) cycles and are then released into constant darkness (DD) for a few days. In DD, circadian rhythms are only dependent on the endogenous clock, as there are no environmental perturbations, and the entrained behaviors persist under DD. Downregulation of GW182, a protein that interacts with Argonaute (Ago) protein to modulate miRNA-mediated translational repression or mRNA degradation, in circadian neurons results in arrhythmicity under DD and an advanced evening phase with absent morning anticipation under LD [35]. The underlying mechanism for GW182 regulation of circadian rhythms appears to be downstream of the PDF-receptor signaling pathway [35].
Dicer1, which encodes an important ribonuclease gene for miRNA biogenesis, has also been implicated in the modulation of the circadian clock. Flies with downregulated Dicer1 in circadian neurons exhibited a lower amplitude of rest to activity rhythms [36]. This relatively weak effect on circadian rhythms might be due to the increased activity of repressors under these conditions, which likely buffers the translation from miRNA deficiencies [36]. The reduced amplitude is likely due to the downregulation of some circadian-relevant miRNAs. Interestingly, another study showed that the period of Dicer-deficient mice was shortened by approximately 2 h due to faster translation of two core clock proteins, namely, PER1 and PER2 [37].
miRNA-mediated circadian amplitude
In the following sections, we will discuss several circadian-related miRNAs to illustrate the effects of specific miRNA regulatory mechanisms on circadian rhythms. The free-running circadian amplitude in DD reflects the self-sustainability of the circadian clock and serves as an important parameter for the evaluation of behavioral robustness. It has been shown that when some miRNAs are overexpressed, they severely impact the self-sustainability of the circadian clock. These miRNAs include miR-279 and miR-276a [38, 39] (Fig. 2, Table 1).
Table 1.
Characteristics of circadian-related miRNAs
| miRNA | Target | Orcadian phenotype | Oscillation | Conservation |
|---|---|---|---|---|
| bantam | clk | Period | N | C. elegans and insects |
| let-7 | cwo | Period and phase | Peak at ZT8 Trough at ZT16 |
C. elegans, Drosophila, Mouse, Humans |
| miR-279 | upd | Rhythmicity | N | Insects |
| miR-276a | timeless | Rhythmicity | Peak at ZT10 Trough at ZT18 | Insects |
| miR-124 | – | Phase | Peak at ZT19 Trough at ZT7 | C. elegans, Drosophila, Mouse, Humans |
| miR959-964 | – | Immunity feeding | Peak at ZT12 Trough at ZT0 | Drosophila |
| miR263a/b | – | – | Peak at ZT19 miR263a trough at ZT1 miR263b trough at ZT7 |
C. elegans, Drosophila, Mouse, Humans |
Overexpression of miR-279 in circadian neurons was found to cause arrhythmia in most flies [38]. Luo et al. [38] reported that miR-279 regulated circadian rhythm output pathways by targeting the key component, unpaired (upd), in the JAK/STAT pathway. Though UPD is absent in the main pacemaker neurons (sLNv), it is prevalent in PER-positive neurons, DNs and laterally located neurons (LLNs), all of which are in close proximity to PDF-containing dorsal projections [38]. This further indicated that miR-279 functions in circadian output. Notably, both overexpression and knockout of miR-279 caused a high degree of arrhythmicity, possibly due to the multiple target effects or its essential role in the output pathway. Additional data have demonstrated that miR-996 can rescue the behavioral phenotype in miR-279 mutants [40]. miR-996 is a circadian-relevant miRNA located downstream of miR-279, which shares a similar seed region. Overexpression of miR-996 in circadian neurons was found to result in a phenotype similar to that of miR-279 overexpression [40]. It seems that miR-996 is redundant compared to miR-279, based on the regulatory capacities and similar seed region, as well as abundance, but the presence of miR-996 may add to the robustness of miR-279 regulation of circadian rhythms or may participate in unique functions.
Another miRNA known to influence the circadian clock is miR-276a [39]. miR-276a is positively regulated by the transcription factor Chorion factor 2 (CF2) and has a cyclic expression pattern under LD. Interestingly, flies overexpressing CF2 have a phenotype similar to that seen in flies with an overexpression of miR-276a. miR-276a appears to target the central clock gene, tim [39]. Analyses of flies with spatial and temporal misregulation of miR-276a showed that this miRNA is necessary for circadian behavior robustness in PDF-positive neurons (LNvs). TIM and PER levels were reduced following miR-276a overexpression. Reduced PER levels in PDF-positive neurons was deemed to be a secondary effect of direct targeting of the tim 3′UTR by miR-276a. Deletions of miR-276a binding sites in the tim 3′UTR phenocopied the deletion of miR-276a at both molecular and behavioral levels [39]. Interestingly, TIM overexpression did not fully rescue the arrhythmic phenotype. Two possible explanations for this observation are that the circadian clock may be extremely sensitivity to TIM dosage or that miR-276a targets additional circadian-relevant mRNAs.
miRNA-mediated circadian period
The free running period is one of the most important characteristics of circadian rhythms as it reflects the speed of the circadian clock. To date, two miRNAs, namely, bantam and let-7, have been found to be involved in the regulation of the free-running period. These two miRNAs target two central clock genes, clk and cwo, respectively [36, 41] (Fig. 2, Table 1).
It is worth noting that bantam was the first miRNA found to affect the circadian rhythms of flies. The miRNA, bantam, was first shown to play vital roles in germline maintenance, peripheral nervous system dendrite growth, and eye development [42–44]. Through tiling arrays, Kadener et al. [36] found that disruption of miRNA processing in the nucleus resulted in the accumulation of bantam within fly heads, and overexpression of bantam in the main pacemaker neurons lengthens the circadian period. The molecular mechanism behind the circadian-period regulatory function of bantam involves interactions with three conserved target sites of the clk 3′UTR. An interaction between clk mRNA and RISC has also been shown by AGO1 immunoprecipitation [36]. It appears that bantam has other functions, as restoration of clk levels by expression of clk with a 3′UTR with no bantam binding sites did not rescue the arrhythmicity phenotype of clkAR.
Like bantam, let-7 also modulates the circadian period in Drosophila. Along with two other non-redundant miRNAs, miR-100 and miR-125, let-7 is a co-transcriptional product from a single polycistronic locus, known as the let-7 complex (let-7-C) [45]. Previous studies of let-7-C miRNA were focused on its developmental functions. These miRNAs are important for the remodeling of the abdominal neuromusculature, ovary morphogenesis, nervous system formation, and lifespan control [45–48]. Expression of let-7-C is localized in the α/β lobes of the mushroom bodies [47], brain structures known to be essential for learning and memory regulation, as well as for sleep and locomotor activity [49, 50]. A recent study showed that let-7 is also expressed in LNv neurons, further indicating the role of let-7 in circadian rhythm regulation [41]. Abnormal expression of let-7 in the main pacemaker neurons disrupts normal circadian rhythms: overexpression of let-7 lengthens the period, whereas knockout of let-7 eliminates morning anticipation [41]. Developmental defects were also excluded, as restricted overexpression in adulthood led to a long period of rest-activity rhythm [41]. Notably, cwo, which negatively regulates clk expression, was identified as the target of let-7 both in vitro and in vivo [41]. Other phenotypes resulting from let-7-C knockout, such as abnormal PDF expression in PDF neuron projections and faster PER accumulation in LNv [41], may be due to secondary effects.
miRNA-mediated phase determination in locomotion rhythms
Fruit flies exhibit a bimodal behavioral profile during each LD cycle, consisting of increased activity before dawn and dusk (termed morning anticipation and evening anticipation, respectively), both of which are controlled by dedicated circadian neurons [51]. Additionally, flies also display sharp and transient increases of activity during light–dark transitions, known as the “startle” response, which is a non-circadian response that persists even without a functional clock [51] or the main pacemakers [52]. miR-124 is conserved across the animal kingdom and is abundantly expressed in the central nervous system [53] (Fig. 2, Table 1). In the fly head, miR-124 expression is under circadian regulation, with trough and peak expression levels at mid-day and midnight, respectively [34]. Interestingly, miR-124-knockout flies show decreased morning anticipation under LD and an advanced evening phase under DD, but the circadian period is normal [54, 55]. In flies that do not express miR-124, circadian oscillators are functional with unaffected PER/TIM oscillation in all circadian neurons, and the photoreceptors show normal light response, suggesting that neither input or central clock functions are affected [54, 55]. Thus, miR-124 may function in circadian output. Several potential targets of miR-124 encode factors involved in the retrograde BMP signaling pathway, including Mef2, MMP1, and other positive components. However, expression of Mef2 and MMP1 and activation of BMP signaling regulate the endogenous period or rhythmicity under constant darkness [56–58], which is not consistent with the observed phenotype of miR-124 mutants. A double heterozygous mutation of sax and mad was found to partially correct the phase shift in the miR-124 mutant background [55], indicating the multifunction of miR-124 in regulating circadian rhythms.
Unlike miR-124-mediated regulation of the locomotion phase, the miR959-964 cluster mainly regulates the phase of feeding and immune function [59] (Table 1). The miR959-964 cluster, which includes six miRNAs, is encoded by two introns proximal to the protein-coding gene, CG31646 [59]. These six miRNAs show a robust circadian oscillation, which peaks at dusk (ZT12) [59]. The pri-miRNA cluster of miR959-964 displays an even stronger oscillation than individual mature miRNAs do. This oscillation is abolished, and levels accumulate in the arrhythmic per01 mutant, and, oppositely, mature miR959-964 levels are reduced in the per01 mutant. Surprisingly, expression pattern analysis did not localize expression of the miR959-964 to circadian-relevant neurons, despite the fact that elimination of four of six miRNAs (miR959-962) displayed mild effects on the circadian period (approximately 0.5 h shorter), and overexpression of the cluster in TIM-positive neurons extended the period for more than 1 h [59]. In fact, this cluster is expressed in the peri-cerebral fat body (adult head fat body), an organ involved in metabolism and immunity [60], a finding that is consistent with the predication that most mRNA targets of miR959-964 are implicated in metabolic and immune functions. The miR959 ~ 964 cluster negatively modulates immune response, as the knockdown line exhibited dramatically lowered survival of the pathogen Pseudomonas aeruginosa [59]. The circadian-modulated feeding behavior was positively regulated by these six miRNAs, and there is a bidirectional regulation between feeding and levels of this cluster [59]. The circadian clock controls feeding time, and miRNAs in this cluster are likely to downregulate factors involved in responses to stress and starvation that regulate multiple downstream physiological processes, such as metabolism, feeding, and foraging time, until the stress is offset.
Approaches to study circadian-relevant miRNAs
One direct approach that aims to identify circadian-relevant miRNAs involves monitoring circadian behaviors after performing a forward genetic screen, which entails manipulation of miRNA expression via knockout, overexpression, or downregulation. Recently, a large collection of miRNA mutants generated by targeted homologous recombination has become available [61]. These stocks enable genetic screening for circadian-relevant miRNAs. In addition, the Gal4/UAS system is a powerful tool for manipulating gene expression due to the availability and diversity of transgenic flies with UAS-miRNA or UAS-miRNA-sponge constructs [62]. Thus, by using tissue- and cell-specific drivers, it is feasible to manipulate functions of specific miRNAs only in certain organs. In addition, targeted spatial and temporal knockout of specific genes has become readily available with the emergence of the CRISPR/Cas9 system [63].
One efficient way to identify circadian-relevant miRNAs is to do RNA-sequencing (RNA-seq) in either all circadian or specific circadian neurons [64]. miRNAs can be extracted from specific circadian neurons after FACS isolation or by manual sorting [64]. The comprehensive and high-throughput nature of RNA-seq enables identification of relatively short, non-coding transcripts. Moreover, this approach provides great insights into the oscillation of mature miRNAs and the abundance of pri-miRNAs through intron mapping.
Characterization of miRNA targets is essential for understanding the mechanisms of how miRNA functions. Bioinformatic tools, such as TargetScan [65] and PicTar [66], have been widely used for predicting potential miRNA targets. In the future, in vitro studies can be performed by co-transfecting the miRNA and its predicted target gene 3′UTR with a reporter. If the miRNA binds to its target, a reduced reporter signal will be observed. Additionally, manipulation of target gene expression should phenocopy the effect of the miRNA in vivo. Finally, if a mutant mRNA that does not have miRNA binding sites in the target resembles the miRNA knockout phenotype, this would provide strong evidence supporting miRNA-target interactions.
Concluding remarks
Emerging evidence has shown that miRNA-mediated post-transcriptional regulation is crucial to the generation and maintenance of a robust circadian clock in flies and mammals [9, 13, 26]. In this review, we discussed miRNAs known to modulate circadian rhythmicity and the free-running period, as well as locomotion and feeding phases in flies. Why is miRNA-mediated post-transcriptional regulation necessary in regulating the different attributes of circadian clock? One likely role is to contribute to the generation of the 24-h period. Although the established molecular mechanism of the circadian clock is based on the TTFL, it takes less than 24 h to close up the TTFL. Thus, additional regulatory levels, such as post-translational regulation and post-transcriptional regulation, for example, are required to generate the 24-h circadian period. A second likely role is to confer robustness to the running of the circadian clock. Multiple regulatory levels ensure that the circadian clock is less sensitive to changing environmental conditions. The third likely role is to enhance the mRNA oscillation of circadian-related genes as rhythmic mRNA expression is an important molecular output pathway of the circadian clock. Taken together, miRNA-mediated post-transcriptional regulation imposes benefits on the operation of the circadian clock.
As of yet, uncharacterized miRNAs may also play important roles in maintaining circadian rhythms. For example, miRNAs are found to be important for circadian photoresponses and seasonal adaptation in mammals [67], but no miRNA has been implicated in the circadian input pathways in flies yet. To date, 466 miRNAs have been identified in the fly genome (www.mirbase.org), but only a handful of miRNAs have been reported to be relevant to circadian clock function. Therefore, further characterization of miRNAs will be a vital step toward a more comprehensive understanding of the regulatory mechanisms underlying miRNA-mediated post-transcriptional control of the circadian clock.
Authors’ contributions
YZ and YX formed the idea for this review. YX and YZ wrote the manuscript. YX prepared the figures and table. Both authors read and approved the final manuscript.
Acknowledgements
We are grateful to the members of Zhang lab, especially Wesley Leigh and Eric Liu for their advice and discussions. Yong Zhang’s lab was supported by the National Institutes of Health COBRE Grant P20 GM103650.
Competing interests
The authors declare that they have no competing interests. Yong Zhang is an Associate Editor for BMC Neuroscience.
Availability of data and materials
Not applicable.
Consent to publish
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
Yong Zhang’s lab was supported by the National Institutes of Health COBRE Grant P20 GM103650.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- AGO
Argonaute
- CK2
casein kinase II
- CLK
CLOCK
- CWO
clockwork orange
- CYC
CYCLE
- DBT
doubletime
- DD
dark: dark cycle
- DN
dorsal neurons
- GSK3
glycogen synthase kinase 3
- LD
light: dark cycle
- LNd
dorsal lateral neurons
- LNv
ventral lateral neurons
- miRNA
microRNA
pigment dispersing factor
- PDP1
par domain protein 1
- PER
PERIOD
- PP1
protein phosphatase 1
- PP2A
protein phosphatase 2A
- RBP
RNA-binding proteins
- RISC
RNA-induced silencing complex
- TTFL
transcriptional-translational feedback loop
- TIM
TIMELESS
- UTR
untranslated region
- VRI
VRILLE
Contributor Information
Yongbo Xue, Email: yongbo@nevada.unr.edu.
Yong Zhang, Phone: (775) 682 7896, Email: yongzhang@unr.edu.
References
- 1.Johnson CH, Elliott JA, Foster R. Entrainment of circadian programs. Chronobiol Int. 2003;20(5):741–774. doi: 10.1081/CBI-120024211. [DOI] [PubMed] [Google Scholar]
- 2.Allada R, Chung BY. Circadian organization of behavior and physiology in Drosophila. Annu Rev Physiol. 2010;72:605–624. doi: 10.1146/annurev-physiol-021909-135815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Richards J, Gumz ML. Mechanism of the circadian clock in physiology. Am J Physiol Reg I. 2013;304(12):R1053–R1064. doi: 10.1152/ajpregu.00066.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Donelson NC, Sanyal S. Use of Drosophila in the investigation of sleep disorders. Exp Neurol. 2015;274:72–79. doi: 10.1016/j.expneurol.2015.06.024. [DOI] [PubMed] [Google Scholar]
- 5.Di Cara F, King-Jones K. The circadian clock is a key driver of steroid hormone production in Drosophila. Curr Biol. 2016;26(18):2469–2477. doi: 10.1016/j.cub.2016.07.004. [DOI] [PubMed] [Google Scholar]
- 6.Yu EA, Weaver DR. Disrupting the circadian clock: gene-specific effects on aging, cancer, and other phenotypes. Aging. 2011;3(5):479–493. doi: 10.18632/aging.100323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Roenneberg T, Merrow M. The circadian clock and human health. Curr Biol CB. 2016;26(10):R432–R443. doi: 10.1016/j.cub.2016.04.011. [DOI] [PubMed] [Google Scholar]
- 8.Karatsoreos IN, Bhagat S, Bloss EB, Morrison JH, McEwen BS. Disruption of circadian clocks has ramifications for metabolism, brain, and behavior. Proc Natl Acad Sci USA. 2011;108(4):1657–1662. doi: 10.1073/pnas.1018375108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mehta N, Cheng HY. Micro-managing the circadian clock: the role of microRNAs in biological timekeeping. J Mol Biol. 2013;425(19):3609–3624. doi: 10.1016/j.jmb.2012.10.022. [DOI] [PubMed] [Google Scholar]
- 10.Nitabach MN, Taghert PH. Organization of the Drosophila circadian control circuit. Curr Biol. 2008;18(2):R84–R93. doi: 10.1016/j.cub.2007.11.061. [DOI] [PubMed] [Google Scholar]
- 11.Abruzzi KC, Zadina A, Luo WF, Wiyanto E, Rahman R, Guo F, Shafer O, Rosbash M. RNA-seq analysis of Drosophila clock and non-clock neurons reveals neuron-specific cycling and novel candidate neuropeptides. PLoS Genet. 2017;13(2):e1006613. doi: 10.1371/journal.pgen.1006613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kojima S, Green CB. Circadian genomics reveal a role for post-transcriptional regulation in mammals. Biochemistry. 2015;54(2):124–133. doi: 10.1021/bi500707c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rodriguez J, Tang CH, Khodor YL, Vodala S, Menet JS, Rosbash M. Nascent-Seq analysis of Drosophila cycling gene expression. Proc Natl Acad Sci USA. 2013;110(4):E275–E284. doi: 10.1073/pnas.1219969110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.He L, Hannon GJ. Micrornas: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531. doi: 10.1038/nrg1379. [DOI] [PubMed] [Google Scholar]
- 15.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Carthew RW, Agbu P, Giri R. MicroRNA function in Drosophila melanogaster. Semin Cell Dev Biol. 2017;65:29–37. doi: 10.1016/j.semcdb.2016.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Blau J, Blanchard F, Collins B, Dahdal D, Knowles A, Mizrak D, Ruben M. What is there left to learn about the Drosophila clock? Cold Spring Harb Symp Quant Biol. 2007;72:243–250. doi: 10.1101/sqb.2007.72.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yu W, Hardin PE. Circadian oscillators of Drosophila and mammals. J Cell Sci. 2006;119(Pt 23):4793–4795. doi: 10.1242/jcs.03174. [DOI] [PubMed] [Google Scholar]
- 19.Edery I, Zwiebel LJ, Dembinska ME, Rosbash M. Temporal phosphorylation of the Drosophila period protein. Proc Natl Acad Sci USA. 1994;91(6):2260–2264. doi: 10.1073/pnas.91.6.2260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Martinek S, Inonog S, Manoukian AS, Young MW. A role for the segment polarity gene shaggy/GSK-3 in the Drosophila circadian clock. Cell. 2001;105(6):769–779. doi: 10.1016/S0092-8674(01)00383-X. [DOI] [PubMed] [Google Scholar]
- 21.Price JL, Blau J, Rothenfluh A, Abodeely M, Kloss B, Young MW. double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell. 1998;94(1):83–95. doi: 10.1016/S0092-8674(00)81224-6. [DOI] [PubMed] [Google Scholar]
- 22.Lin JM, Kilman VL, Keegan K, Paddock B, Emery-Le M, Rosbash M, Allada R. A role for casein kinase 2alpha in the Drosophila circadian clock. Nature. 2002;420(6917):816–820. doi: 10.1038/nature01235. [DOI] [PubMed] [Google Scholar]
- 23.Top D, Harms E, Syed S, Adams EL, Saez L. GSK-3 and CK2 kinases converge on timeless to regulate the master clock. Cell Rep. 2016;16(2):357–367. doi: 10.1016/j.celrep.2016.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fang Y, Sathyanarayanan S, Sehgal A. Post-translational regulation of the Drosophila circadian clock requires protein phosphatase 1 (PP1) Genes Dev. 2007;21(12):1506–1518. doi: 10.1101/gad.1541607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sathyanarayanan S, Zheng X, Xiao R, Sehgal A. Posttranslational regulation of Drosophila PERIOD protein by protein phosphatase 2A. Cell. 2004;116(4):603–615. doi: 10.1016/S0092-8674(04)00128-X. [DOI] [PubMed] [Google Scholar]
- 26.Koike N, Yoo SH, Huang HC, Kumar V, Lee C, Kim TK, Takahashi JS. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science (New York, NY) 2012;338(6105):349–354. doi: 10.1126/science.1226339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gerstner JR, Vanderheyden WM, LaVaute T, Westmark CJ, Rouhana L, Pack AI, Wickens M, Landry CF. Time of day regulates subcellular trafficking, tripartite synaptic localization, and polyadenylation of the astrocytic Fabp7 mRNA. J Neurosci Off J Soc Neurosci. 2012;32(4):1383–1394. doi: 10.1523/JNEUROSCI.3228-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Liu Y, Hu W, Murakawa Y, Yin J, Wang G, Landthaler M, Yan J. Cold-induced RNA-binding proteins regulate circadian gene expression by controlling alternative polyadenylation. Sci Rep. 2013;3:2054. doi: 10.1038/srep02054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tataroglu O, Emery P. The molecular ticks of the Drosophila circadian clock. Curr Opin Insect Sci. 2015;7:51–57. doi: 10.1016/j.cois.2015.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bartok O, Kyriacou CP, Levine J, Sehgal A, Kadener S. Adaptation of molecular circadian clockwork to environmental changes: a role for alternative splicing and miRNAs. Proc Biol Sci. 2013;280(1765):20130011. doi: 10.1098/rspb.2013.0011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wu X, Brewer G. The regulation of mRNA stability in mammalian cells. Gene. 2012;500(1):10–21. doi: 10.1016/j.gene.2012.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Huang Y, McNeil GP, Jackson FR. Translational regulation of the DOUBLETIME/CKIδ/ε kinase by LARK contributes to circadian period modulation. PLoS Genet. 2014;10(9):e1004536. doi: 10.1371/journal.pgen.1004536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lerner I, Bartok O, Wolfson V, Menet JS, Weissbein U, Afik S, Haimovich D, Gafni C, Friedman N, Rosbash M, et al. Clk post-transcriptional control denoises circadian transcription both temporally and spatially. Nat Commun. 2015;6:7056. doi: 10.1038/ncomms8056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yang M, Lee JE, Padgett RW, Edery I. Circadian regulation of a limited set of conserved microRNAs in Drosophila. BMC Genom. 2008;9:83. doi: 10.1186/1471-2164-9-83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang Y, Emery P. GW182 controls Drosophila circadian behavior and PDF-receptor signaling. Neuron. 2013;78(1):152–165. doi: 10.1016/j.neuron.2013.01.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kadener S, Menet JS, Sugino K, Horwich MD, Weissbein U, Nawathean P, Vagin VV, Zamore PD, Nelson SB, Rosbash M. A role for microRNAs in the Drosophila circadian clock. Genes Dev. 2009;23(18):2179–2191. doi: 10.1101/gad.1819509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chen R, D’Alessandro M, Lee C. miRNAs are required for generating a time delay critical for the circadian oscillator. Curr Biol CB. 2013;23(20):1959–1968. doi: 10.1016/j.cub.2013.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Luo W, Sehgal A. microRNA-279 acts through the JAK/STAT pathway to regulate circadian behavioral output in Drosophila. Cell. 2012;148(4):765–779. doi: 10.1016/j.cell.2011.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen X, Rosbash M. mir-276a strengthens Drosophila circadian rhythms by regulating timeless expression. Proc Natl Acad Sci USA. 2016;113(21):E2965–E2972. doi: 10.1073/pnas.1605837113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sun K, Jee D, de Navas LF, Duan H, Lai EC. Multiple in vivo biological processes are mediated by functionally redundant activities of Drosophila mir-279 and mir-996. PLoS Genet. 2015;11(6):e1005245. doi: 10.1371/journal.pgen.1005245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chen W, Liu Z, Li T, Zhang R, Xue Y, Zhong Y, Bai W, Zhou D, Zhao Z. Regulation of Drosophila circadian rhythms by miRNA let-7 is mediated by a regulatory cycle. Nat Commun. 2014;5:5549. doi: 10.1038/ncomms6549. [DOI] [PubMed] [Google Scholar]
- 42.Li Y, Padgett RW. bantam is required for optic lobe development and glial cell proliferation. PLoS ONE. 2012;7(3):e32910. doi: 10.1371/journal.pone.0032910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Parrish JZ, Xu P, Kim CC, Jan LY, Jan YN. The microRNA bantam functions in epithelial cells to regulate scaling growth of dendrite arbors in drosophila sensory neurons. Neuron. 2009;63(6):788–802. doi: 10.1016/j.neuron.2009.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shcherbata HR, Ward EJ, Fischer KA, Yu JY, Reynolds SH, Chen CH, Xu P, Hay BA, Ruohola-Baker H. Stage-specific differences in the requirements for germline stem cell maintenance in the Drosophila ovary. Cell Stem Cell. 2007;1(6):698–709. doi: 10.1016/j.stem.2007.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sokol NS, Xu P, Jan YN, Ambros V. Drosophila let-7 microRNA is required for remodeling of the neuromusculature during metamorphosis. Genes Dev. 2008;22(12):1591–1596. doi: 10.1101/gad.1671708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chawla G, Deosthale P, Childress S, Wu YC, Sokol NS. A let-7-to-miR-125 MicroRNA switch regulates neuronal integrity and lifespan in Drosophila. PLoS Genet. 2016;12(8):e1006247. doi: 10.1371/journal.pgen.1006247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kucherenko MM, Barth J, Fiala A, Shcherbata HR. Steroid-induced microRNA let-7 acts as a spatio-temporal code for neuronal cell fate in the developing Drosophila brain. EMBO J. 2012;31(24):4511–4523. doi: 10.1038/emboj.2012.298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yang H, Li M, Hu X, Xin T, Zhang S, Zhao G, Xuan T, Li M. MicroRNA-dependent roles of Drosha and Pasha in the Drosophila larval ovary morphogenesis. Dev Biol. 2016;416(2):312–323. doi: 10.1016/j.ydbio.2016.06.026. [DOI] [PubMed] [Google Scholar]
- 49.Joiner WJ, Crocker A, White BH, Sehgal A. Sleep in Drosophila is regulated by adult mushroom bodies. Nature. 2006;441(7094):757–760. doi: 10.1038/nature04811. [DOI] [PubMed] [Google Scholar]
- 50.Mabuchi I, Shimada N, Sato S, Ienaga K, Inami S, Sakai T. Mushroom body signaling is required for locomotor activity rhythms in Drosophila. Neurosci Res. 2016;111:25–33. doi: 10.1016/j.neures.2016.04.005. [DOI] [PubMed] [Google Scholar]
- 51.Stoleru D, Peng Y, Agosto J, Rosbash M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature. 2004;431(7010):862–868. doi: 10.1038/nature02926. [DOI] [PubMed] [Google Scholar]
- 52.Sheeba V, Fogle KJ, Holmes TC. Persistence of morning anticipation behavior and high amplitude morning startle response following functional loss of small ventral lateral neurons in Drosophila. PLoS ONE. 2010;5(7):e11628. doi: 10.1371/journal.pone.0011628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Weng R, Cohen SM. Drosophila miR-124 regulates neuroblast proliferation through its target anachronism. Development (Cambridge, England) 2012;139(8):1427–1434. doi: 10.1242/dev.075143. [DOI] [PubMed] [Google Scholar]
- 54.Zhang Y, Lamba P, Guo P, Emery P. miR-124 regulates the phase of Drosophila circadian locomotor behavior. J Neurosci Off J Soc Neurosci. 2016;36(6):2007–2013. doi: 10.1523/JNEUROSCI.3286-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Garaulet DL, Sun K, Li W, Wen J, Panzarino AM, O’Neil JL, Hiesinger PR, Young MW, Lai EC. miR-124 regulates diverse aspects of rhythmic behavior in Drosophila. J Neurosci Off J Soc Neurosci. 2016;36(12):3414–3421. doi: 10.1523/JNEUROSCI.3287-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Blanchard FJ, Collins B, Cyran SA, Hancock DH, Taylor MV, Blau J. The transcription factor Mef2 is required for normal circadian behavior in Drosophila. J Neurosci Off J Soc Neurosci. 2010;30(17):5855–5865. doi: 10.1523/JNEUROSCI.2688-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Depetris-Chauvin A, Fernandez-Gamba A, Gorostiza EA, Herrero A, Castano EM, Ceriani MF. Mmp1 processing of the PDF neuropeptide regulates circadian structural plasticity of pacemaker neurons. PLoS Genet. 2014;10(10):e1004700. doi: 10.1371/journal.pgen.1004700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Beckwith EJ, Gorostiza EA, Berni J, Rezaval C, Perez-Santangelo A, Nadra AD, Ceriani MF. Circadian period integrates network information through activation of the BMP signaling pathway. PLoS Biol. 2013;11(12):e1001733. doi: 10.1371/journal.pbio.1001733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Vodala S, Pescatore S, Rodriguez J, Buescher M, Chen YW, Weng R, Cohen SM, Rosbash M. The oscillating miRNA 959-964 cluster impacts Drosophila feeding time and other circadian outputs. Cell Metab. 2012;16(5):601–612. doi: 10.1016/j.cmet.2012.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Hwangbo DS, Gershman B, Tu MP, Palmer M, Tatar M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature. 2004;429(6991):562–566. doi: 10.1038/nature02549. [DOI] [PubMed] [Google Scholar]
- 61.Chen YW, Song S, Weng R, Verma P, Kugler JM, Buescher M, Rouam S, Cohen SM. Systematic study of Drosophila microRNA functions using a collection of targeted knockout mutations. Dev Cell. 2014;31(6):784–800. doi: 10.1016/j.devcel.2014.11.029. [DOI] [PubMed] [Google Scholar]
- 62.Ebert MS, Sharp PA. MicroRNA sponges: progress and possibilities. RNA (New York, NY) 2010;16(11):2043–2050. doi: 10.1261/rna.2414110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bassett AR, Liu JL. CRISPR/Cas9 and genome editing in Drosophila. J Genet Genomics. 2014;41(1):7–19. doi: 10.1016/j.jgg.2013.12.004. [DOI] [PubMed] [Google Scholar]
- 64.Abruzzi K, Chen X, Nagoshi E, Zadina A, Rosbash M. RNA-seq profiling of small numbers of Drosophila neurons. Methods Enzymol. 2015;551:369–386. doi: 10.1016/bs.mie.2014.10.025. [DOI] [PubMed] [Google Scholar]
- 65.Ruby JG, Stark A, Johnston WK, Kellis M, Bartel DP, Lai EC. Evolution, biogenesis, expression, and target predictions of a substantially expanded set of Drosophila microRNAs. Genome Res. 2007;17(12):1850–1864. doi: 10.1101/gr.6597907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Grun D, Wang YL, Langenberger D, Gunsalus KC, Rajewsky N. microRNA target predictions across seven Drosophila species and comparison to mammalian targets. PLoS Comput Biol. 2005;1(1):e13. doi: 10.1371/journal.pcbi.0010013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Cheng HY, Papp JW, Varlamova O, Dziema H, Russell B, Curfman JP, Nakazawa T, Shimizu K, Okamura H, Impey S, et al. microRNA modulation of circadian-clock period and entrainment. Neuron. 2007;54(5):813–829. doi: 10.1016/j.neuron.2007.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.