

Abstract
A growing body of nutritional science highlights the complex mechanisms and pleiotropic pathways of cardiometabolic effects of different foods. Among these, some of the most exciting advances are occurring in the area of flavonoids, bioactive phytochemicals found in plant foods; and in the area of dairy, including milk, yogurt, and cheese. Many of the relevant ingredients and mechanistic pathways are now being clarified, shedding new light on both the ingredients and the pathways for how diet influences health and well-being. Flavonoids, for example, have effects on skeletal muscle, adipocytes, liver and pancreas, and myocardial, renal, and immune cells, for instance related to AMPK phosphorylation, eNOS activation, and suppression of NF-κB and TLR4. Effects of dairy are similarly complex and may be mediated by specific amino acids, medium-chain and odd-chain saturated fats, unsaturated fats, branched-chain fats, natural trans fats, probiotics, vitamin K1/K2, and calcium, as well as by processing such as fermentation and homogenization. These characteristics of dairy foods influence diverse pathways including related to mammalian target of rapamycin, silent information regulator transcript-1, angiotensin-converting enzyme, peroxisome proliferator-activated receptors, osteocalcin, matrix glutamate protein, hepatic de novo lipogenesis, hepatic and adipose fatty acid oxidation and inflammation, and gut microbiome interactions such as intestinal integrity and endotoxemia. The complexity of these emerging pathways and corresponding biologic responses highlights the rapid advances in nutritional science and the continued need to generate robust empiric evidence on the mechanistic and clinical effects of specific foods.
Keywords: nutrition, flavonoids, dairy, cardiovascular disease, diabetes
Introduction
Dietary habits are a major determinant of cardiometabolic diseases including heart disease, stroke, and type 2 diabetes.1 In the US, for example, poor diet contributes to nearly half of all cardiometabolic deaths, causing an estimated 1,000 deaths each day.2 The resulting global health and economic burdens are staggering.3
In recent years, nutritional science has been transformed by an explosion of evidence, shedding new light on key compounds and pathways for how diet influences health and well-being. Among these, some of the most exciting advances are occurring in the areas of flavonoids, bioactive phytochemicals found in a range of plant foods; and dairy foods, including milk, yogurt, and cheese.
For these factors, the emerging evidence on the relevant ingredients and biologic mechanisms highlights the importance of investigating the pleiotropic pathways of effects of foods. We performed a narrative review of the emerging science and innovative discoveries in the understanding of how flavonoids and dairy foods influence cardiometabolic health, with a focus on experimental studies and molecular mechanisms, as well as supportive clinical evidence.
Flavonoids
Flavonoids are polyphenolic phytochemicals that include flavonols (in onions, broccoli, tea, and various fruits), flavones (in parsley, celery, and chamomile tea), flavanones (in citrus fruits), flavanols (flavan-3-ols) such as catechins and procyanidins (in cocoa, apples, grapes, red wine, and tea), anthocyanidins (in colored berries and red wine), and isoflavones (in soy) (Table 1).4–7
Table 1.
| Flavonoids
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anthocyanidins | Flavanols (flavan-3-ols) |
Flavanones | Flavones | Flavonols | Proanthocyanidins | Isoflavones | |||||||
|
| |||||||||||||
| Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
Major flavonoids |
Food sources |
| Cyanidin Delphinidin Malvidin Pelargonidin Peonidin Petunidin |
Berries, red wine | Catechins epicatachins | Apricots, cocoa, chocolates, red grapes, red wine, tea | Eriodictyol Hesperetin naringenin |
Citrus fruit, mint, tomatoes | Apigenin Luteolin |
Celery, parsley | Isohamnetin Kaempferol Myricetin Quercetin |
Apples, blueberries, broccoli, curly kale, leeks, onions, red wine, tea | Theaflavins Thearubigins |
Apples, chocolates, grapes | Daidzein Genistein Glycitein |
Soybeans, soy foods |
The structural diversity of flavonoids contribute to differences in their ability to modulate specific molecular pathways. Differences in absorption, distribution, metabolism, and elimination following consumption further modify their bioavailability, site of action, and formation of bioactive metabolites.8 Whereas some flavonoids are well absorbed and distributed to multiple tissues, others have limited absorption – although such flavonoids could still have systemic effects via interaction with the microbiota.9 Isoflavones and catechins have in particular been extensively studied (and covered by recent reviews) with regards to their cardiometabolic effects.10–14 As reviewed below, compelling experimental evidence suggests that flavonoids influence multiple physiologic pathways related to cardiometabolic diseases (Figure 1).
Figure 1. Selected cardiometabolic benefits of flavonoids and potential underlying molecular mechanisms.
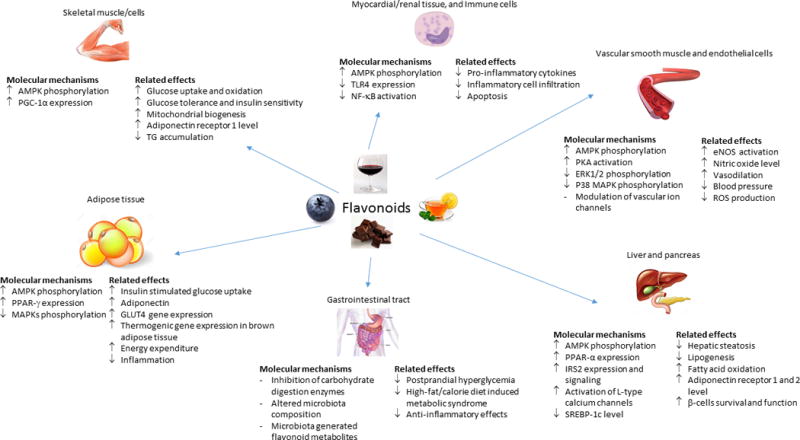
In vitro and animal studies support bioactivity of purified flavonoids or flavonoids-rich plant extracts across multiple tissues. Relevant molecular pathways appear to include: 1) Modulation of gene expression and signaling pathways. Enhancement of AMPK phosphorylation and activation appears to be a common mechanism affected by several types of flavonoids. Modulation of other signaling pathways have also been observed including increased expression of PPAR-γ and inhibition of NF- κB activation; 2) Interaction with gut-microbiota. Dietary flavonoids may alter gut-microbial composition due to probiotic-like properties and stimulate growth of specific bacteria (e.g. Akkermansia muciniphila) that may confer metabolic benefits. Conversely, metabolism of dietary flavonoids by gut bacteria generates downstream metabolites (e.g. phenolic acids) that may possess unique properties and/or reach higher circulating and tissue concentrations compared to parent flavonoids, thus enhance biologic activity of flavonoids; 3) Direct flavonoid-protein interactions. Growing evidence suggest flavonoids may stimulate and inhibit protein function, including ion channels in the vasculature and liver, and carbohydrate digestive enzymes (α-amylase and α-glucosidase) in the gastrointestinal tract. Such effects may partly contribute to regulation of vascular tone and glucose metabolism.
Abbreviations: AMPK, 5’-monophosphate-activated protein kinase; ERK1/2, extracellular signal-regulated kinases 1 and 2; GLUT4, glucose transporter type 4; IRS2, insulin receptor substrate-2; MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor-κB; PGC-1α, peroxisome proliferator-activated receptor-gamma coactivator-1α; PKA; protein kinase-A; PPAR, peroxisome proliferator-activated receptors; SREBP-1c, sterol regulatory element binding protein-1c; TG, triglycerides; TLR4, toll-like receptor 4,
Microbial-Generated Flavonoid Metabolites
Colonic microbiota can enzymatically convert flavonoids into small phenolic acids and aromatic metabolites.15, 16 Feeding studies that trace metabolic conversion suggest that such flavonoid catabolites are readily absorbed in the colon and often possess longer half-lives and reach substantially higher systemic concentrations than parent compounds.17–19 Such observations have increased interest in these microbiota-generated metabolites, including whether they mediate cardiometabolic effects of dietary flavonoids. In vitro studies provide preliminary concordant evidence. At physiologically relevant doses, several microbiome-derived phenolic metabolites suppressed production of pro-inflammatory cytokines and vascular adhesion molecules, compared with their parent flavonoids.20–22 Several microbial-derived flavonoid metabolites also protected against pancreatic beta-cell dysfunction and death.23
Dietary flavonoids may also alter gut microbial composition, for example due to probiotic-like properties and stimulation of growth of specific bacteria.24, 25 In animal models of obesity, feeding of flavonoids altered gut microbial community structure, including increased levels of Akkermansia muciniphila26–28 which appear to confer metabolic benefits.29, 30 Flavonoids may also influence the gut microbiota production of short-chain fatty acids (SCFA, up to 6 carbons in length).31 SCFA, predominantly acetic (2:0), propionic (3:0), and butyric (4:0) acids, are produced by large intestinal bacteria mainly from fermentation of non-digestible or poorly digestible carbohydrates (e.g., dietary fiber).32 In addition to being an energy source, experimental studies suggest that microbial-produced SCFA act as signaling molecules and can influence host energy metabolism, glucose-insulin homeostasis, production of endocrine hormones (e.g. GLP-1), and inflammatory pathways. In some studies in mice and rats, dietary SCFA protected against weight gain, improved glucose tolerance, and increased insulin sensitivity.33–36 However, conflicting results have also been observed: in mice fed a high-fat/calorie diet, oral or intravenous acetate reduced food intake and weight gain,36, 37 while intra-gastric infusion in rats fed a high-fat/calorie diet had the opposite effect.38 The reasons for these differences remain unclear, highlighting the need for further mechanistic studies including in humans. Experimental evidence suggests that physiologic effects of SCFA are partly mediated by specific G-protein coupled receptors (GPR) present in multiple cells and tissue types including colon, adipose, and the sympathetic nervous system.39 Specific SCFA may also act via different pathways: for example, in rats, metabolic benefits of dietary propionic acid required GPR activation whereas butyrate did not.34 GPR signaling40 or other mechanisms such as epigenetic modification41 may account for anti-hypertensive and anti-inflammatory effects of SCFA in some cellular and animal studies.42, 43 It remains unclear how much the variability of gut-produced SCFAs depends on flavonoids, and the clinical relevance of microbiota-generated SCFA in humans is being elucidated.44 Yet, the overall emerging evidence supports bi-directional interactions between flavonoid consumption and gut microbiota composition and function that alter physiologic pathways relevant to cardiometabolic health.
Glucose-insulin homeostasis
A large number of animal-experimental studies have tested the effects of purified flavonoid compounds or flavonoid-rich plant extracts on insulin-glucose homeostasis, with a substantial number suggesting possible benefits. Flavonoids may influence glucose metabolism in the small intestine, muscle, adipose, liver, and pancreas via a number of molecular mechanisms. In vitro studies suggest a variety of flavonoids inhibit key enzymes involved in the digestion and absorption of dietary carbohydrates including α-amylase, α-glucosidase, and sodium-dependent glucose transporter, which may contribute to reduced post-prandial glycaemia.45 Flavonoids could also improve glucose-insulin homeostasis via multiple signaling pathways. Cell culture and animal studies have identified adenosine 5′-adenosine monophosphate-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor-γ (PPAR-γ) as two of the key pathways via which some flavonoids enhance muscle glucose uptake and improve adipocyte function.46–50. Flavonoid treatment in animal models also led to reduced liver fat accumulation and improved hepatic insulin sensitivity, which were related to reductions in de novo lipogenesis and increase in fatty acid β-oxidation.51–54 Finally, cellular and animal studies suggest several types of flavonoids protected pancreatic β-cells against glucotoxicity and inflammation, and enhanced insulin secretion.55–57 Activation of AMPK has again been implicated in mediating the effects of flavonoids on insulin secretion, but other mechanisms including modulation of intra-cellular calcium through activation of membrane ion channels have also been identified for specific flavonoids.58, 59
Nitric oxide bioavailability, redox status, and vasoregulation
In animal experiments, administration of flavonoids exerted vasorelaxation effects and lowered blood pressure.60 There is strong evidence that a key pathway via which flavonoids regulate vascular health is through altered nitric oxide (NO) metabolism. Influence of flavonoids on NO could be further divided into direct and indirect mechanisms.7 Several flavonoids can directly increase endothelial nitric oxide synthase (eNOS) expression and activity (the main source of NO in the vasculature), leading to enhanced production of NO.61–63 Effects on eNOS level could be mediated through activation of AMPK.64 Flavonoids could also indirectly enhance NO bioavailability through lowering the production or enhance removal of reactive oxygen species that are known to breakdown NO. Treatment with different subgroups of flavonoids increased the activity of endogenous anti-oxidant enzymes including sodium oxide dismutase and catalase, reduced superoxide radical generation by NADPH oxidase, and lowered protein and lipid biomarkers of oxidative stress.65–67 In addition to regulating vascular function through NO, other NO independent mechanisms have also been observed in in vitro studies for flavonoids such as direct stimulation or inhibition of vascular calcium ion channels.68
Weight maintenance
Supplementation with flavonoids prevented diet-induced weight gain in several animal models of obesity. In these investigations, flavonoids did not appear to influence energy intake,69–72 suggesting they may contribute to weight regulation by increasing energy expenditure. For example, luteolin (a flavonoid abundant in pepper, apple skins, and carrots) up-regulated AMPK and PPAR-γ coactivator 1-α (PGC-1α) signaling cascades, leading to elevated thermogenic gene expression in brown and subcutaneous adipose tissues, and enhanced energy expenditure in C57BL/6 mice fed low or high fat diets.72 Other flavonoids have also demonstrated an ability to induce brown fat-specific genes and proteins in cultured adipocytes.73 Additional mechanisms via which flavonoids could increase energy expenditure have been observed in animal feeding studies, including stimulation of the sympathetic nerve system,74 and increased skeletal muscle mitochondrial biogenesis and function.71, 75 Several types of flavonoids may also prevent fat accumulation via reduced lipogenesis, and increased β-oxidation of fatty acids as demonstrated in cultured adipocytes and mice.76–78
Anti-inflammatory effects
Some flavonoids have demonstrated anti-inflammatory properties in adipose and myocardial tissues in animal studies following varied inflammatory stimuli including ischemia-reperfusion, diabetes, medication use, and high-fat diet.79–85 In these models, oral supplementation with flavonoids led to reduced inflammatory cell infiltration, lowered levels of pro-inflammatory cytokines and tissue fibrosis, and improved cell survival and function. A central pathway that appeared to mediate the anti-inflammatory effect of several flavonoids was inhibition of signaling via nuclear factor-κB (NF-κB).82–84 However, other mechanisms are likely involved and have been identified for specific flavonoids – for example, hexameric procyanidins (present in high concentrations in cocoa, tea, and apples) inhibited the binding of tumor necrosis factor-α to its receptor and subsequent pro-inflammatory activation in cultured cells.86
Clinical effects
A growing body of laboratory studies and randomized trials support cardiometabolic benefits of flavonoid-rich foods such as cocoa, tea, and berries. Flavonoid-rich cocoa produces small but measurable benefits on blood pressure (BP), endothelial function, insulin resistance, and blood lipids.87–89 In a systematic review and meta-analysis of 42 RCTs,88 chocolate, cocoa, and flavan-3-ol significantly reduced mean arterial pressure (−1.64mm Hg, 95% CI: −3.27, −0.01 mm Hg), improved flow-mediated dilatation (1.34%; 95% CI: 1%, 1.68%), lowered HOMA-IR (−0.67, 95% CI, −0.98, −0.36), and marginally improved LDL-C and HDL-C (−0.07 and +0.03mmol/L, respectively). BP-lowering has been seen with as little as 6.3 g/day of dark chocolate (~30 kcal/day, i.e. about 1.5% of total daily energy added to habitual diets without recommendations for other dietary calories).90 These benefits appear related to improved endothelial production of nitric oxide,90 a fundamental pathway for vascular and metabolic health that suggests the potential for benefits beyond lowering of BP alone. Accumulating data suggest green or black tea can also modestly reduce BP in pre-hypertensive and hypertensive individuals – a meta-analysis of 10 trials suggests reduction of SBP and DBP by 2.36 and 1.77 mm Hg, respectively – although risk of bias in most of these trials could not be fully evaluated due to insufficient reported information.91 A systematic review and meta-analysis of 22 RCTs of berries found moderate improvements in SBP (−2.72mm Hg; 95% CI, −5.32, −0.12), as well as small improvements in glycemic control (HbA1C, −0.20%), body mass index (−0.36kg/m2), LDL-C (−0.21mmol/L), and inflammatory biomarkers (tumor necrosis factor-α, −1pg/mL).92 Yet, most of these RCTs were small and of limited duration (<6 months).
Complementary to interventional evidence, observational studies evaluating dietary flavonoids or flavonoid-rich foods have observed lower risk of cardiometabolic events.93–99 For example, among >90,000 middle-aged nurses followed for 18 years, those with in the highest vs. lowest quintile of estimated dietary anthocyanin intake had 32% lower risk of incident myocardial infarction (95% CI, 4%, 51%, P=0.03); however, other major subclasses of flavonoids (flavanones, flavan-3-ol, flavonols, flavones, and flavonoid polymers) and total flavonoids were not associated with myocardial infarction.94 Sub-class specific associations were also observed in other population-based cohort studies, including for flavanones and ischemic stroke,95, 96 and flavonols and type 2 diabetes.93 These findings suggest potential heterogeneous effects of different types of flavonoids in relation to specific cardiometabolic outcomes. Estimation of dietary flavonoid intake has important limitations including errors in self-recall and inaccuracies in food composition databases.100 Assessment of urine or blood flavonoid biomarkers is therefore a complementary approach to examining exposure, but has only been utilized in a handful of long-term studies. These have predominantly focused on isoflavones and type 2 diabetes, and suggest moderate inverse associations for daidzein and genistein, major isoflavones in soy foods.101 Data for other flavonoid biomarkers and risk of cardiometabolic diseases are scarce and inconsistent.102–105 Additional studies with larger sample sizes across population groups with diverse demographic and dietary habits are needed.
There is also evidence for cardiometabolic benefits of nuts and extra-virgin olive oil, rich in other types of phenolic compounds (e.g. phenolic acids and lignans). In the PREvencion con DIeta MEDiterranea (PREDIMED) trial, participants at high risk of cardiovascular disease were randomized to a Mediterranean dietary pattern and provided with daily extra-virgin olive oil or mixed nuts (walnuts, hazelnuts, and almonds). Compared to the control diet (advice to reduce dietary fat), the intervention diets significantly improved cardiovascular disease (CVD) risk factor profiles including LDL-cholesterol, blood pressure, and inflammatory biomarkers;106–108 and resulted in ~30% lower risk of death, myocardial infarction, or stroke.109 Participants in the intervention groups also demonstrated less gain in central adiposity and decreases in body weight after ~5 years of follow-up.110 Meta-analysis of prospective cohort studies provide further support of cardiometabolic benefits of higher nuts consumption: for each 1 oz (28 g) per day, about 30% lower risk of coronary heart disease (CHD) (n=11 studies, RR=0.71, 95% CI, 0.63, 0.80) and 39% lower risk of diabetes (n=4 studies, RR=0.61, 95% CI, 0.43, 0.88).111
Overall, growing evidence supports meaningful cardiometabolic benefits of foods rich in flavonoids and other phenolics. These findings support recommendations to increase dietary consumption of these foods and provide clear impetus for additional mechanistic trials, prospective cohorts, and clinical trials to better characterize the specific compounds of interest and their dose-response effects.
Dairy Foods
Dairy products contribute about 10% of all calories in the US diet.112 Yet, for such a major share of the food supply, relatively little research has evaluated the direct health impact of consuming dairy foods. Traditional dietary recommendations on dairy derive mostly from theoretical considerations about isolated nutrients (e.g., eat three daily servings to obtain calcium or vitamin D for bone health; eat low-fat products to reduce calories for weight gain and saturated fat for heart disease),113 rather than empiric evidence on actual mechanistic and clinical effects of milk, cheese, yogurt, butter, or other dairy foods. Growing evidence suggests that different dairy foods have complex cardiometabolic effects based on potential interrelated influences of a range of nutrients and other characteristics such as probiotics, fermentation, and possibly homogenization (Figure 2). We do not discuss the potential role of vitamin D here, which has been extensively reviewed elsewhere.114, 115
Figure 2. Relevant characteristics of dairy foods and selected molecular pathways potentially linked to cardiometabolic disease risk.
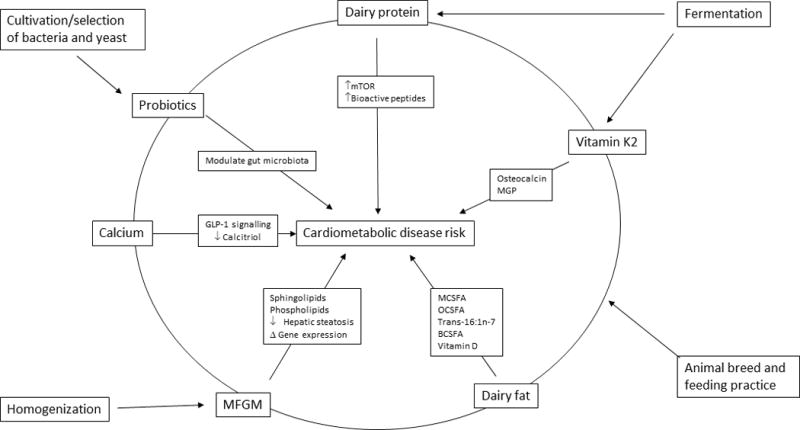
Dairy foods are characterized by a complex mixture of nutrients and processing methods that may influence cardiovascular and metabolic pathways. Relevant constituents include specific fatty acids, calcium, and probiotics. Relevant processing methods may include animal breeding and feeding, fermentation, selection and cultivation of bacterial and yeast strains (e.g., as fermentation starters), and homogenization. Such modifications can alter the food’s composition (e.g., fermentation leads to production of vitamin K2 from vitamin K1) as well as its lipid structures (e.g., homogenization damages MFGM), each of which can affect downstream molecular and signaling pathways.
Abbreviations: BCSFA, branched-chain saturated fats; GLP-1, glucagon-like peptide 1; MCSFA, medium-chain saturated fats; MFGM, milk-fat globule membranes; MGP, matrix glutamate protein; mTOR, mammalian target of rapamycin; OCSFA, odd chain saturated fats.
Calcium
Cell culture and animal experiments have assessed calcium and cardiometabolic risk, alone or in conjunction with other dairy components. In several animal models of obesity, calcium supplementation inhibited weight gain, attenuated hepatic steatosis, and reduced hyperglycemia and insulin resistance.116–119 These effects were potentially mediated by correction of leptin and glucagon-like peptide 1 (GLP-1) signaling,118, 120 reduced levels of calcitriol (1,25-dihydroxyvitamin D3), suppression of hepatic and adipose lipogenesis,121, 122 and alterations in gut microbiota composition.123, 124 However, other animal models have not demonstrated such benefits.125–127 For example, in a mouse model of diet-induced obesity, calcium supplementation caused weight gain relative to control.127 In a meta-analysis of 20 trials including 2711 participants, calcium supplementation did not significantly lower body weight (−0.17 kg, 95%CI: −0.70, 0.37) or body fat (−0.19 kg, 95%CI: −0.51, 0.13).128 In comparison, dairy foods increase lean mass and reduce body fat, compared to control, in the presence of energy restriction for weight loss (see Clinical Effects, below); suggesting that other components beyond calcium may be relevant.
In short-term trials in humans, calcium supplements modestly lower BP, with mean difference (95% CI) for SBP, −1.43 mmHg (−2.15, −0.72 mmHg, I2=0%); and for DBP, −0.98mmHg (−1.46, −0.50 mmHg, I2=49%).129 In some animal models of hypertension, reduction in BP following calcium supplementation were linked to improvement in both endothelial dependent and independent arterial relaxation, enhanced hyperpolarization of vascular smooth muscle, increased sodium excretion, and downregulation of renal angiotensin-converting enzyme.130–133 However, whether calcium intake has similar effects on these pathways in humans is not clear. Meta-analysis of long-term randomized trials found that calcium supplementation resulted in trends towards moderately elevated risk of myocardial infarction.134, 135 For example, in the study by Mao et al, the odds ratio (95% CI) for the calcium supplemented compared to the placebo group was 1.28 (0.97−1.68, P=0.08, I2=0%).135 Genetic variants related to higher serum calcium level also relates to elevated risk of myocardial infarction and coronary artery disease in Mendelian randomization studies.136 The potential for increased risk has been hypothesized to relate to postprandial hypercalcemia that occurs with supplements, in comparison to intake from foods, that may contribute to vascular calcification. Overall, calcium is not a convincing driver of cardiometabolic benefits of dairy foods, although effects could also depend on supplement vs. dietary sources.
Dairy protein
Bovine milk contains around 32–34g/l protein, largely casein (used to make curds during milk processing; ~80% of dairy protein) and also whey protein (~20%).137 Both casein and whey protein include several smaller protein fractions and differ in amino acid composition.137 In some animal studies, enriching diets with casein, whey protein, or complete milk protein improved glucose-insulin and cardiometabolic risk factors.138–140 Such benefits might relate to specific dairy amino acids. For example, whey protein is rich in the branched-chain amino acids (BCAA) leucine, isoleucine, and valine, that activates important signaling pathways including mammalian target of rapamycin (mTOR), and silent information regulator transcript 1;141, 142 which could contribute to enhanced thermogenesis and insulin secretion.143 However, BCAA supplementation in animal studies has shown mixed results related to metabolic outcomes.144–147 Relatively few controlled trials of intact milk protein isolates have been performed in humans.148 Several focused on casein-derived lactotripeptides, which significantly lowered systolic (men difference, −2.95mmHg, 95% CI, −4.17, −1.73mmHg) and diastolic BP (men difference, −1.51mmHg, 95% CI, −2.21, −0.8mmHg) based on pooled results across studies; although these findings should be interpreted cautiously due to substantial heterogeneity and potential for publication bias.149 Other short-term clinical studies (up to 12 weeks) evaluated effects of milk protein on glucose-insulin homeostasis: overall favorable effects were observed, but long-term studies remain limited.137
Bioactive peptides derived from dairy protein may also contribute, generated during fermentation (e.g. in the production of cheese or kefir, sour milk) via action of bacterial proteolytic enzymes, or during gastrointestinal (including microbiota-related) digestion.150 Several short peptides (3–4 amino acids in length) from casein and whey protein demonstrated inhibitory activity towards angiotensin-converting enzyme in vitro.151 Other dairy derived peptides have also been shown to moderately inhibit dipeptidyl peptidase-4,152, 153 which may contribute to increased half-live of incretin hormones (gastric inhibitory peptide and glucagon-like peptide-1) and improved glycemic control.143 On the other hand, the relevance of such dairy-derived bioactive peptides has been challenged based on their low bioavailability, which produces circulating levels in the pM to nM range.154
Overall, experimental and short-term human metabolic studies support potential cardiometabolic benefits of dairy protein, but the relative efficacy of casein vs. whey protein, effects of individual amino acids vs. peptide metabolites, and corresponding molecular mechanisms and relevant pathways remain understudied.
Dairy fats
Dietary guidelines generally recommend low/non-fat dairy based on LDL-raising effects of myristic (14:0) and stearic (16:0) saturated fatty acids, underemphasizing positive effects of these fatty acids on VLDL, chylomicron remnants, and HDL-cholesterol155 and paying even less attention to potential health effects of the many other fatty acids which comprise the majority of dairy fat (e.g. 14:0 plus 16:0 comprise ≤40% of total fatty acids in cow, sheep and goat’s milk).156 These include medium-chain saturated fats (MCSFA) (between 6 to 12 carbons, i.e. 6:0 to 12:0), odd-chain saturated fats (OCSFA) (15:0, 17:0), monounsaturated and polyunsaturated fatty acids (18:1n-9, 18:2n-6 and 18:3n-3), branched-chain saturated fats, and trace amounts of natural (ruminant) trans fats (e.g., trans-palmitoleic acid, trans-16:1n-7).156–158 Dairy fat is also a source of phospholipids (milk fat globule membrane) and fat-soluble vitamins including D, K, and K2 (produced during fermentation; see below).
MCSFA, representing ~6-17% of dairy milk fatty acids, have different molecular and metabolic activities than longer chain fatty acids. For example, whereas longer chain SFA (16:0 and 18:0) activated NF-κB and decreased insulin sensitivity in cultured skeletal muscle cells, the MCSFA 8:0 and 12:0 did not.159 MCSFA also enhanced mitochondrial oxidative capacity and reduced lipid accumulation in cultured muscle cells relative to 16:0.160 These effects may account for observed reductions in body fat accumulation and insulin resistance in animals fed high MCSFA vs. longer chain SFA.160, 161 On the other hand, relative to a low-fat control diet, high fat feeding with MCSFA enhanced hepatic de novo lipogenesis and triglyceride accumulation, and reduced hepatic insulin sensitivity, in animal models.161, 162 Induction of hepatic lipogenesis could be due to MCSFA activation and signaling via liver X Receptor-α.163 Notably, many of the prior animal experiments examining MCSFA were obesity models and also focused on fruit (coconut) sources, and thus the metabolic effects of dairy-derived MCSFA under eucaloric conditions (e.g., substituting for other types of dietary fatty acids) remain unclear.
The biologic effects of trans-16:1n-7, branched-chain saturated fats, and OCSFA have received relatively little attention. It has been hypothesized164 that dietary trans-16:1n-7 could exert similar effects as dietary cis-16:1n-7, which when consumed in the diet or produced outside the liver appears to act in a negative feedback loop to inhibit hepatic de novo lipogenesis, improve insulin sensitivity, and reduce inflammation;165–169 with corresponding risk factor improvements in one human trial.170 In cultured INS-1 β cells, treatment with trans-16:1n-7 activated PPAR-γ and the transcription factor pancreatic duodenal homeobox (PDX)-1.171 Yet, relevance of such effects on glucose-insulin homeostasis and other molecular effects of trans-16:1n-7 remain unknown. Potential mechanisms of branched chain saturated fats also remain little explored. A branched chain FA (15-methyl-hexadecanoic acid) exhibited similar effects on PPAR-γ and PDX-1 as trans-16:1n-7 in cultured INS-1 β cells under basal conditions, and additionally countered high glucose mediated suppression of PDX-1.171 Intake of branched-chain saturated fats is not insubstantial – with estimated average at ~500mg/day in the US (primarily from dairy and beef products),158 compared to between 125-160mg/day for seafood derived long-chain n-3 polyunsaturated fats.172 These findings highlight the potential quantitative importance of dietary intake of branched-chain saturated fats and the need to further assess their biologic functions. OCSFA from dairy fat are incorporated into a range of tissues including blood, liver, and adipose.173, 174 In addition to serving as an energy source via β-oxidation, other metabolic functions have been proposed such as enabling replenishment of the citric acid cycle and improving mitochondrial function,174 but such hypotheses remain to be tested in rigorous experimental investigations.
Milk fat globule membrane
Milk fat is naturally bound by milk fat globule membranes (MFGM), a tri-layered membrane rich in polar lipids (phospholipids and sphingolipids) and proteins, enclosing a triglyceride core (globule) of fatty acids.175 These polar lipids and proteins in MFGM appears to be bioactive. In mice, supplementation with sphingolipids and bovine milk phospholipids reduced serum cholesterol and hepatic lipid accumulation, attributed to reduced intestinal cholesterol uptake and changes in hepatic gene expression.176–178 Possible anti-inflammatory properties have also been reported – mice fed a MFGM-enriched diet exhibited decreased inflammatory responses to a systemic lipopolysaccharide (LPS) challenge, possibly due to reduced gut permeability.179 Processing of dairy products can change the content and structure of MFGM – for instance, homogenization may destroy MFGM.180 A recent randomized trial among 57 overweight adults compared the effects on blood lipids and genetic expression of consuming ~15% of calories from whipping cream (intact MFGM) vs. butter (little MFGM due to homogenization), otherwise equivalent in contents of dairy fat and saturated fat. After 8 weeks, those consuming butter had predictable increases in LDL-C and apolipoprotein B:A-I ratio, while those consuming whipping cream showed no changes in their lipid profile.180 The whipped cream group demonstrated significantly lower expression of 19 genes in peripheral blood mononuclear cells, including USP45, MDM2, SNRPN, and CAPZA1, supporting effects of MFGM on genetic expression. Similar blunted effects on total and LDL-C have been seen in cross-over trials comparing cheese to butter or non-dairy saturated fat (see Clinical Effects, below)181, 182. These findings suggest that MFGM and corresponding processing methods that preserve or destroy it may have important implications for cardiometabolic effects of dairy fat.
Probiotics
A growing body of evidence supports health effects of probiotics in foods, live microorganisms which can alter foods’ characteristics as well as host responses following consumption.183 Both yogurt and kefir (a fermented milk drink) often contain live bacteria (kefir can also contain yeasts). In several animal models of obesity and diabetes, dairy products with probiotics demonstrated cardiometabolic benefits compared to those without probiotics. For example, in C57BL/6 mice fed high-calorie/fat diets, animals given kefir had reduced weight gain, hepatic steatosis, LDL-C, and IL-6 levels compared to mice given unfermented milk.184 Such changes were accompanied by altered expression of hepatic and adipose tissues genes related to fatty acid oxidation (AOX, PPAR-α) and inflammation (MCP-1). Other studies suggest that efficacy is probiotic specific: e.g., compared to unfermented milk, milk fermented with different strains of Lactobacillus rhamnosus improved glucose tolerance and fasting glucose to varying extents in a diabetic rat model.185 The molecular mechanisms for probiotics’ health effects appear to involve changes in both composition and function of host gut microbiota.186 For instance, microbiota composition in animals was altered by probiotic dairy products such as yoghurt and kefir.184, 187, 188 Such compositional changes may enhance intestinal epithelial integrity and reduce low grade inflammation due to endotoxemia (leakage of gut-microbiota derived LPS into systemic circulation), a putative contributor to obesity-related diseases.189 Probiotics also appear to influence host microbiota function, e.g. altering production of functional mediators such as SCFA that may exert local and systemic effects on host metabolism.190, 191 In sum, animal-experimental studies and human trials support a role for probiotics and probiotic-microbiome interactions in protective effects of yogurt for weight gain, obesity, and related metabolic conditions such as gestational diabetes.192–203
Cheese, fermentation, and vitamin K
There are two major forms of vitamin K: K1 (phylloquinone, rich in green-leafy vegetables and certain vegetable oils) and K2 (menaquinone, MK, differentiated by the number of isoprene residues, MKn). Several vitamin K2-producing bacteria species are commonly used in industrial dairy fermentation, and cheese is a major source of vitamin K2 (especially MK7, -8, and -9) in Europe and North America.204, 205
All forms of vitamin K act as cofactors for post-translational carboxylation of protein glutamate residues into gamma-carboxy glutamate, required for vitamin K-dependent proteins (VKDP) to become active. While coagulation factors such as factors VII, IX, and X are well known VKDP, growing evidence suggests that additional VKDP influence cardiometabolic health.204 This includes osteocalcin (made in bone cells) and matrix glutamate protein (MGP, primarily made in vascular smooth muscle cells and cartilage). Animal studies support a role of osteocalcin in improving beta-cell proliferation, insulin expression, and upregulation of adiponectin in adipocytes.206 In several metabolic studies, vitamin K supplementation increased carboxylated osteocalcin concentrations and reduced insulin resistance.207, 208 However, results were not always consistent in human studies,209 and opposing directions of associations between carboxylated/undercarboxylated forms of osteocalcin with insulin sensitivity have been observed in mice vs. human, suggesting possible species differences.210 Levels of dietary vitamin K and proportions of osteocalcin that must be γ-carboxylated to improve glucose-insulin homeostasis also remain unclear.211 Similarly, while it has been hypothesized that vitamin K may reduce CVD risk by augmenting MGP, an inhibitor of vascular calcification, this has not yet been convincingly established. In several rats and mice studies, supplementation with vitamin K reduced arterial calcification, but whether such effects were mediated by MGP carboxylation or other mechanisms remains unclear.212–214
Human metabolic studies demonstrate that specific types of vitamin K2 have longer half-lives and reach higher circulating levels than vitamin K1. For instance, compared to a half-life of 1-2 hours for vitamin K1, MK-7 and MK-9 have estimated half-lives of 2–3 days,.215, 216 These differences in bioavailability may have functional consequences – in one study among healthy adults, supplementation with MK-7 induced more complete carboxylation of osteocalcin.216 Such findings suggest that vitamin K2 moieties (representing ~15–20% of total dietary vitamin K in Western diets, with the rest as vitamin K1), may disproportionally contribute to vitamin K activity in vivo.205 Furthermore, recent cohort studies suggest that K2, but not K1, is linked to lower CVD risk.217–219 For example, in a prospective cohort study among 16,057 women aged 49–70 years, the hazard ratio for the risk of CHD per 10μg/day (equivalent to ~1SD) of K2 intake was 0.91 (95% CI, 0.81, 1.00, P=0.04), but K1 intake was not related to CHD risk.219 Given these findings as well as the specific links of cheese and fermented milk to clinical outcomes (see below), the potential role of fermentation and vitamin K2 in cardiometabolic risk represents a new area of promise for further research.
Clinical Effects
In short-term randomized trials, consumption of milk or overall dairy products increases lean mass and reduces body fat, especially in the setting of energy-restricted weight loss diets.128, 220, 221 Long-term effects are less clear and may vary by type of dairy. Observationally, several studies suggest that children who drink more low-fat milk gain more weight over time, while those who drink more whole-fat milk gain less weight.222–226 Few long-term trials have been performed in children, other than multi-component dietary interventions that preclude inference on dairy per se.227, 228
In longitudinal studies among adults, relationships between dairy intake and weight, CVD, and diabetes endpoints vary more by food type (e.g., cheese, yogurt, milk, butter) than fat content.229–235 For example, neither low-fat nor whole milk are appreciably related to long-term weight gain,196, 236, 237 perhaps related to subtle caloric compensation: when people eat more low-fat dairy, they on average increase their consumption of carbohydrates, while people who eat more full-fat dairy on average decrease their carbohydrate intake.237 Cheese consumption similarly appears relatively neutral for long-term weight gain; although this might be modified by carbohydrate intake: weight gain is seen when cheese is accompanied by refined carbohydrates, and relative weight loss is seen when cheese replaces refined carbohydrates.237 Yogurt appears consistently protective against long-term weight gain,196, 236, 237 even if sugar-sweetened (although in this case, only about half the benefit is seen, compared with unsweetened yogurt237).
While increased intake of saturated fat from dairy products would be expected to increase LDL-C,238 recent randomized controlled trials support heterogeneity in such effects depending on the type of dairy foods consumed. For instance, in a randomized cross-over trial among 49 men and women, consuming equivalent amounts of fat and saturated fat from cheese, as compared to butter, lowered total, LDL, and HDL cholesterol concentrations.181 Similar blunted effects on total and LDL-C were seen in a randomized controlled cross-over trial comparing saturated fat from milk or cheese with saturated fat from non-dairy sources;182 as well as comparing whipping cream to butter.180 Such heterogeneous responses may be explained by other components in dairy (e.g. calcium) or by specific processing methods (e.g., presence or absence of MFGM, see above).239 Such counterbalancing effects, as well as beneficial effects of saturated fat on levels of triglyceride-rich VLDL-C,238 could explain why meta-analyses of long-term cohort studies demonstrate no significant associations of total dairy consumption with CHD events and actually lower risk of stroke, without consistent differences comparing reduced vs. regular fat products.240
Associations of dairy foods with risk of type 2 diabetes also vary by food type: yogurt, but not milk, is consistently associated with lower risk; while consumption of cheese, which has highest calorie, fat, and saturated fat content, also associates with lower risk in several although not all studies.232, 234, 235, 241, 242 These differences may be partly elucidated by the divergent associations of total milk (generally unassociated with diabetes) vs. fermented milk (linked to lower risk),234, 241, 243 suggesting a potential role for metabolic benefits of fermented products such as cheese (see above).
Interestingly, dairy fat itself may promote cardiometabolic health. In cohorts utilizing objective biomarkers, higher blood levels of dairy fatty acids consistently associate with lower incidence of diabetes244–248 and perhaps CHD,249–251 with mixed findings for stroke.252 As described above, mechanistic explanations for these observations remain unclear, which could include metabolic effects of fermented foods (especially cheese, a major source of dairy fat), links of such biomarkers to MFGM, specific fatty acids (e.g., branched-chain fatty acids, MCSFA, specific ruminant trans fats), other lipid-soluble factors, or unknown endogenous (non-dietary) determinants of these blood biomarkers.253
Future Directions and Conclusions
Modern nutritional science is elucidating the diversity of ingredients and mechanisms by which foods influence health. Numerous in vitro and animal studies support pleiotropic effects of flavonoids on multiple risk factors and pathways relevant to cardiometabolic diseases. While molecular mechanisms continue to be clarified, identified signaling pathways include AMPK, PPAR-γ, PGC-1α, and NF-κB. Existing experimental studies also have methodologic limitations and the potential for publication bias, and the relevance of their findings to humans remain unclear. In addition, with more than 5,000 naturally occurring flavonoids identified to date,254 observed effects on molecular pathways for some flavonoids are unlikely to be generalizable to others. Many mechanistic studies to-date have focused on parent aglycone forms of flavonoids, and frequently at supra-physiological concentrations (e.g., 25 to 100μM, whereas systemic circulating concentrations in vivo are unlikely to reach ≥ 10μM).21, 255 While findings based on supra-physiologic doses may be relevant for the development of flavonoids as pharmacologic agents, they are less generalizable to cardiometabolic effects of flavonoids at usual dietary levels of intake. Further, prior experimental studies have generally not accounted for complexities in flavonoid bioavailability and metabolism. For instance, most dietary flavonoids (except for flavan-3-ol) are found as glycosides, bound to one or more sugar moieties,7 which generally require hydrolysis prior to intestinal absorption.8 Following absorption, flavonoids undergo phase-I and phase-II metabolism and are transformed into diverse glucuronidated, sulphated, and methylated metabolites.64 Unabsorbed flavonoids are also catabolized by colonic bacteria into a number of phenolic acids. Compared with their parent compounds, many flavonoid metabolites have longer half-lives and achieve much higher concentrations in circulation.22 Cardiometabolic effects of flavonoids observed in animal studies may therefore be largely attributable to their metabolites, rather than the pre-metabolized flavonoids. Yet, relatively few investigations have evaluated potential biologic effects of flavonoid metabolites, partly limited by lack of available synthetic standards.20, 22 Based on the promise of these compounds for physiologic health, future mechanistic, experimental, and clinical studies are needed that take into account the diversity of types, bioavailability, and metabolism of flavonoids and their metabolites to better understand the most appropriate form and pathways for clinical benefits.
Similarly, for dairy foods, a variety of ingredients and processing methods appear to influence cardiometabolic health. Potentially relevant ingredients include specific amino acids, medium-chain and odd-chain saturated fats, unsaturated fats, branched-chain fats, natural trans fats, probiotics, vitamin K1/K2, and calcium, as well as by processing techniques such as fermentation and homogenization. Corresponding pathways of effects include those related to mTOR, silent information regulator transcript-1, angiotensin-converting enzyme, peroxisome proliferator-activated receptors, osteocalcin, matrix glutamate protein, hepatic de novo lipogenesis, hepatic and adipose fatty acid oxidation and inflammation, and gut microbiome interactions such as intestinal integrity and endotoxemia.
For both flavonoids and dairy foods, the complexity of the emerging mechanistic pathways and responses is remarkable. This new evidence highlights the tremendous growth in knowledge, as well as the extent of what remains to be learned, on how different dietary factors influence health. Given the prime importance of nutrition for cardiovascular and metabolic health, these results support the need for vigorous further investigation on the relevant components, biologic pathways, and clinical effects of these and other foods.
Acknowledgments
Funding Support: The research reported in this article was supported by The National Heart, Lung, and Blood Institute, National Institutes of Health (2R01-HL085710, PI Mozaffarian).
Abbreviations
- AMPK
5’-monophosphate-activated protein kinase
- BCAA
branched-chain amino acids
- BCSFA
branched-chain saturated fats
- BP
blood pressure
- CHD
coronary heart disease
- CVD
cardiovascular disease
- DBP
diastolic blood pressure
- eNOS
endothelial nitric oxide synthase
- ERK1/2
extracellular signal-regulated kinases 1 and 2
- GLP-1
glucagon-like peptide 1
- GPR
G-protein coupled receptors
- GLUT4
glucose transporter type 4
- IRS2
insulin receptor substrate-2
- LPS
lipopolysaccharide
- MAPK
mitogen-activated protein kinase
- MCSFA
medium-chain saturated fatty acids
- MFGM
milk-fat globule membranes
- MGP
matrix glutamate protein
- MK
menaquinone
- mTOR
mammalian target of rapamycin
- NF-κB
nuclear factor-κB
- NO
nitric oxide
- OCSFA
odd chain saturated fatty acids
- PDX
pancreatic duodenal homeobox
- PGC-1α
peroxisome proliferator-activated receptor-gamma coactivator-1α
- PKA
protein kinase-A
- PPAR
peroxisome proliferator-activated receptors
- PREDIMED
PREvencion con DIeta MEDiterranea trial
- RCT
randomized controlled trial
- SBP
systolic blood pressure
- SCFA
short chain fatty acids
- SREBP-1c
sterol regulatory element binding protein-1c
- TG
triglycerides
- TLR4
toll-like receptor 4
- VKDP
vitamin K-dependent proteins
Footnotes
COI Disclosures: Dr. Mozaffarian reports ad hoc honoraria or consulting from Life Sciences Research Organization, Astra Zeneca, Acasti Pharma, Boston Heart Diagnostics, GOED, DSM, Nutrition Impact, Haas Avocado Board, and Pollock Communications; scientific advisory board, Omada Health; and chapter royalties from UpToDate. Dr. Wu reports research support from Unilever.
References
- 1.Mozaffarian D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: A Comprehensive Review. Circulation. 2016;133:187–225. doi: 10.1161/CIRCULATIONAHA.115.018585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Micha R, Peñalvo J, Cudhea F, Imamura F, Rehm C, Mozaffarian D. Association between dietary factors and mortality from heart disease, stroke, and diabetes in the United States. JAMA. 2017 doi: 10.1001/jama.2017.0947. epub March 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bloom D, Cafiero E, Jané-Llopis E, Abrahams-Gessel S, Bloom L, Fathima S, Feigl A, Gaziano T, Mowafi M, Pandya A, Prettner K, Rosenberg L, Seligman B, Stein A, Weinstein C. The Global Economic Burden of Noncommunicable Diseases. Geneva: World Economic Forum; 2011. [Google Scholar]
- 4.Cassidy A, O’Reilly EJ, Kay C, Sampson L, Franz M, Forman JP, Curhan G, Rimm EB. Habitual intake of flavonoid subclasses and incident hypertension in adults. The American journal of clinical nutrition. 2011;93:338–47. doi: 10.3945/ajcn.110.006783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ding M, Pan A, Manson JE, Willett WC, Malik V, Rosner B, Giovannucci E, Hu FB, Sun Q. Consumption of soy foods and isoflavones and risk of type 2 diabetes: a pooled analysis of three US cohorts. European journal of clinical nutrition. 2016;70:1381–1387. doi: 10.1038/ejcn.2016.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gu L, Kelm MA, Hammerstone JF, Beecher G, Holden J, Haytowitz D, Gebhardt S, Prior RL. Concentrations of proanthocyanidins in common foods and estimations of normal consumption. The Journal of nutrition. 2004;134:613–7. doi: 10.1093/jn/134.3.613. [DOI] [PubMed] [Google Scholar]
- 7.Bondonno CP, Croft KD, Ward N, Considine MJ, Hodgson JM. Dietary flavonoids and nitrate: effects on nitric oxide and vascular function. Nutrition reviews. 2015;73:216–35. doi: 10.1093/nutrit/nuu014. [DOI] [PubMed] [Google Scholar]
- 8.Cassidy A, Minihane AM. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. The American journal of clinical nutrition. 2017;105:10–22. doi: 10.3945/ajcn.116.136051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Goya L, Martin MA, Sarria B, Ramos S, Mateos R, Bravo L. Effect of Cocoa and Its Flavonoids on Biomarkers of Inflammation: Studies of Cell Culture, Animals and Humans. Nutrients. 2016;8:212. doi: 10.3390/nu8040212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Legeay S, Rodier M, Fillon L, Faure S, Clere N. Epigallocatechin Gallate: A Review of Its Beneficial Properties to Prevent Metabolic Syndrome. Nutrients. 2015;7:5443–68. doi: 10.3390/nu7075230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mangels DR, Mohler ER., 3rd Catechins as Potential Mediators of Cardiovascular Health. Arteriosclerosis, thrombosis, and vascular biology. 2017;37:757–763. doi: 10.1161/ATVBAHA.117.309048. [DOI] [PubMed] [Google Scholar]
- 12.Ramdath DD, Padhi EM, Sarfaraz S, Renwick S, Duncan AM. Beyond the Cholesterol-Lowering Effect of Soy Protein: A Review of the Effects of Dietary Soy and Its Constituents on Risk Factors for Cardiovascular Disease. Nutrients. 2017;9 doi: 10.3390/nu9040324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Talaei M, Pan A. Role of phytoestrogens in prevention and management of type 2 diabetes. World journal of diabetes. 2015;6:271–83. doi: 10.4239/wjd.v6.i2.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vitale DC, Piazza C, Melilli B, Drago F, Salomone S. Isoflavones: estrogenic activity, biological effect and bioavailability. European journal of drug metabolism and pharmacokinetics. 2013;38:15–25. doi: 10.1007/s13318-012-0112-y. [DOI] [PubMed] [Google Scholar]
- 15.Williamson G, Clifford MN. Colonic metabolites of berry polyphenols: the missing link to biological activity? The British journal of nutrition. 2010;104s(Suppl 3):S48–66. doi: 10.1017/S0007114510003946. [DOI] [PubMed] [Google Scholar]
- 16.van Duynhoven J, Vaughan EE, Jacobs DM, Kemperman RA, van Velzen EJ, Gross G, Roger LC, Possemiers S, Smilde AK, Dore J, Westerhuis JA, Van de Wiele T. Metabolic fate of polyphenols in the human superorganism. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(Suppl 1):4531–8. doi: 10.1073/pnas.1000098107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Czank C, Cassidy A, Zhang Q, Morrison DJ, Preston T, Kroon PA, Botting NP, Kay CD. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: a (13)C-tracer study. The American journal of clinical nutrition. 2013;97:995–1003. doi: 10.3945/ajcn.112.049247. [DOI] [PubMed] [Google Scholar]
- 18.de Ferrars RM, Czank C, Zhang Q, Botting NP, Kroon PA, Cassidy A, Kay CD. The pharmacokinetics of anthocyanins and their metabolites in humans. British journal of pharmacology. 2014;171:3268–82. doi: 10.1111/bph.12676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.van Duynhoven J, van der Hooft JJ, van Dorsten FA, Peters S, Foltz M, Gomez-Roldan V, Vervoort J, de Vos RC, Jacobs DM. Rapid and sustained systemic circulation of conjugated gut microbial catabolites after single-dose black tea extract consumption. Journal of proteome research. 2014;13:2668–78. doi: 10.1021/pr5001253. [DOI] [PubMed] [Google Scholar]
- 20.Amin HP, Czank C, Raheem S, Zhang Q, Botting NP, Cassidy A, Kay CD. Anthocyanins and their physiologically relevant metabolites alter the expression of IL-6 and VCAM-1 in CD40L and oxidized LDL challenged vascular endothelial cells. Molecular nutrition & food research. 2015;59:1095–106. doi: 10.1002/mnfr.201400803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.di Gesso JL, Kerr JS, Zhang Q, Raheem S, Yalamanchili SK, O’Hagan D, Kay CD, O’Connell MA. Flavonoid metabolites reduce tumor necrosis factor-alpha secretion to a greater extent than their precursor compounds in human THP-1 monocytes. Molecular nutrition & food research. 2015;59:1143–54. doi: 10.1002/mnfr.201400799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Warner EF, Zhang Q, Raheem KS, O’Hagan D, O’Connell MA, Kay CD. Common Phenolic Metabolites of Flavonoids, but Not Their Unmetabolized Precursors, Reduce the Secretion of Vascular Cellular Adhesion Molecules by Human Endothelial Cells. The Journal of nutrition. 2016;146:465–73. doi: 10.3945/jn.115.217943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fernandez-Millan E, Ramos S, Alvarez C, Bravo L, Goya L, Martin MA. Microbial phenolic metabolites improve glucose-stimulated insulin secretion and protect pancreatic beta cells against tert-butyl hydroperoxide-induced toxicity via ERKs and PKC pathways. Food and chemical toxicology: an international journal published for the British Industrial Biological Research Association. 2014;66:245–53. doi: 10.1016/j.fct.2014.01.044. [DOI] [PubMed] [Google Scholar]
- 24.Kemperman RA, Gross G, Mondot S, Possemiers S, Marzorati M, de Wiele TV, Dore J, Vaughan EE. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Research International. 2013;53:659–669. [Google Scholar]
- 25.Ruan JQ, Li S, Li YP, Wu WJ, Lee SM, Yan R. The Presystemic Interplay between Gut Microbiota and Orally Administered Calycosin-7-O-beta-D-Glucoside. Drug metabolism and disposition: the biological fate of chemicals. 2015;43:1601–11. doi: 10.1124/dmd.115.065094. [DOI] [PubMed] [Google Scholar]
- 26.Anhe FF, Roy D, Pilon G, Dudonne S, Matamoros S, Varin TV, Garofalo C, Moine Q, Desjardins Y, Levy E, Marette A. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut. 2015;64:872–83. doi: 10.1136/gutjnl-2014-307142. [DOI] [PubMed] [Google Scholar]
- 27.Roopchand DE, Carmody RN, Kuhn P, Moskal K, Rojas-Silva P, Turnbaugh PJ, Raskin I. Dietary Polyphenols Promote Growth of the Gut Bacterium Akkermansia muciniphila and Attenuate High-Fat Diet-Induced Metabolic Syndrome. Diabetes. 2015;64:2847–58. doi: 10.2337/db14-1916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Collins B, Hoffman J, Martinez K, Grace M, Lila MA, Cockrell C, Nadimpalli A, Chang E, Chuang CC, Zhong W, Mackert J, Shen W, Cooney P, Hopkins R, McIntosh M. A polyphenol-rich fraction obtained from table grapes decreases adiposity, insulin resistance and markers of inflammation and impacts gut microbiota in high-fat-fed mice. The Journal of nutritional biochemistry. 2016;31:150–65. doi: 10.1016/j.jnutbio.2015.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne NM, de Vos WM, Cani PD. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:9066–71. doi: 10.1073/pnas.1219451110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Plovier H, Everard A, Druart C, Depommier C, Van Hul M, Geurts L, Chilloux J, Ottman N, Duparc T, Lichtenstein L, Myridakis A, Delzenne NM, Klievink J, Bhattacharjee A, van der Ark KC, Aalvink S, Martinez LO, Dumas ME, Maiter D, Loumaye A, Hermans MP, Thissen JP, Belzer C, de Vos WM, Cani PD. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nature medicine. 2017;23:107–113. doi: 10.1038/nm.4236. [DOI] [PubMed] [Google Scholar]
- 31.Mosele JI, Macia A, Motilva MJ. Metabolic and Microbial Modulation of the Large Intestine Ecosystem by Non-Absorbed Diet Phenolic Compounds: A Review. Molecules (Basel, Switzerland) 2015;20:17429–68. doi: 10.3390/molecules200917429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. Journal of lipid research. 2013;54:2325–40. doi: 10.1194/jlr.R036012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes. 2009;58:1509–17. doi: 10.2337/db08-1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.De Vadder F, Kovatcheva-Datchary P, Goncalves D, Vinera J, Zitoun C, Duchampt A, Backhed F, Mithieux G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014;156:84–96. doi: 10.1016/j.cell.2013.12.016. [DOI] [PubMed] [Google Scholar]
- 35.den Besten G, Bleeker A, Gerding A, van Eunen K, Havinga R, van Dijk TH, Oosterveer MH, Jonker JW, Groen AK, Reijngoud DJ, Bakker BM. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARgamma-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes. 2015;64:2398–408. doi: 10.2337/db14-1213. [DOI] [PubMed] [Google Scholar]
- 36.Kimura I, Ozawa K, Inoue D, Imamura T, Kimura K, Maeda T, Terasawa K, Kashihara D, Hirano K, Tani T, Takahashi T, Miyauchi S, Shioi G, Inoue H, Tsujimoto G. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nature communications. 2013;4:1829. doi: 10.1038/ncomms2852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Frost G, Sleeth ML, Sahuri-Arisoylu M, Lizarbe B, Cerdan S, Brody L, Anastasovska J, Ghourab S, Hankir M, Zhang S, Carling D, Swann JR, Gibson G, Viardot A, Morrison D, Louise Thomas E, Bell JD. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nature communications. 2014;5:3611. doi: 10.1038/ncomms4611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Perry RJ, Peng L, Barry NA, Cline GW, Zhang D, Cardone RL, Petersen KF, Kibbey RG, Goodman AL, Shulman GI. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature. 2016;534:213–7. doi: 10.1038/nature18309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Natarajan N, Pluznick JL. From microbe to man: the role of microbial short chain fatty acid metabolites in host cell biology. American journal of physiology Cell physiology. 2014;307:C979–85. doi: 10.1152/ajpcell.00228.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, Schilter HC, Rolph MS, Mackay F, Artis D, Xavier RJ, Teixeira MM, Mackay CR. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009;461:1282–6. doi: 10.1038/nature08530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Andrade-Oliveira V, Amano MT, Correa-Costa M, Castoldi A, Felizardo RJ, de Almeida DC, Bassi EJ, Moraes-Vieira PM, Hiyane MI, Rodas AC, Peron JP, Aguiar CF, Reis MA, Ribeiro WR, Valduga CJ, Curi R, Vinolo MA, Ferreira CM, Camara NO. Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion. Journal of the American Society of Nephrology: JASN. 2015;26:1877–88. doi: 10.1681/ASN.2014030288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Natarajan N, Hori D, Flavahan S, Steppan J, Flavahan NA, Berkowitz DE, Pluznick JL. Microbial short chain fatty acid metabolites lower blood pressure via endothelial G-protein coupled receptor 41. Physiological genomics. 2016 doi: 10.1152/physiolgenomics.00089.2016. 2016:physiolgenomics.00089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Vinolo MA, Rodrigues HG, Hatanaka E, Sato FT, Sampaio SC, Curi R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. The Journal of nutritional biochemistry. 2011;22:849–55. doi: 10.1016/j.jnutbio.2010.07.009. [DOI] [PubMed] [Google Scholar]
- 44.Morrison DJ, Preston T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut microbes. 2016;7:189–200. doi: 10.1080/19490976.2015.1134082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hanhineva K, Torronen R, Bondia-Pons I, Pekkinen J, Kolehmainen M, Mykkanen H, Poutanen K. Impact of dietary polyphenols on carbohydrate metabolism. International journal of molecular sciences. 2010;11:1365–402. doi: 10.3390/ijms11041365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Eid HM, Martineau LC, Saleem A, Muhammad A, Vallerand D, Benhaddou-Andaloussi A, Nistor L, Afshar A, Arnason JT, Haddad PS. Stimulation of AMP-activated protein kinase and enhancement of basal glucose uptake in muscle cells by quercetin and quercetin glycosides, active principles of the antidiabetic medicinal plant Vaccinium vitis-idaea. Molecular nutrition & food research. 2010;54:991–1003. doi: 10.1002/mnfr.200900218. [DOI] [PubMed] [Google Scholar]
- 47.Kim MS, Hur HJ, Kwon DY, Hwang JT. Tangeretin stimulates glucose uptake via regulation of AMPK signaling pathways in C2C12 myotubes and improves glucose tolerance in high-fat diet-induced obese mice. Molecular and cellular endocrinology. 2012;358:127–34. doi: 10.1016/j.mce.2012.03.013. [DOI] [PubMed] [Google Scholar]
- 48.Ding L, Jin D, Chen X. Luteolin enhances insulin sensitivity via activation of PPARgamma transcriptional activity in adipocytes. The Journal of nutritional biochemistry. 2010;21:941–7. doi: 10.1016/j.jnutbio.2009.07.009. [DOI] [PubMed] [Google Scholar]
- 49.Vazquez Prieto MA, Bettaieb A, Rodriguez Lanzi C, Soto VC, Perdicaro DJ, Galmarini CR, Haj FG, Miatello RM, Oteiza PI. Catechin and quercetin attenuate adipose inflammation in fructose-fed rats and 3T3-L1 adipocytes. Molecular nutrition & food research. 2015;59:622–33. doi: 10.1002/mnfr.201400631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Goto T, Teraminami A, Lee JY, Ohyama K, Funakoshi K, Kim YI, Hirai S, Uemura T, Yu R, Takahashi N, Kawada T. Tiliroside, a glycosidic flavonoid, ameliorates obesity-induced metabolic disorders via activation of adiponectin signaling followed by enhancement of fatty acid oxidation in liver and skeletal muscle in obese-diabetic mice. The Journal of nutritional biochemistry. 2012;23:768–76. doi: 10.1016/j.jnutbio.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 51.Li Z, Xu J, Zheng P, Xing L, Shen H, Yang L, Zhang L, Ji G. Hawthorn leaf flavonoids alleviate nonalcoholic fatty liver disease by enhancing the adiponectin/AMPK pathway. International journal of clinical and experimental medicine. 2015;8:17295–307. [PMC free article] [PubMed] [Google Scholar]
- 52.Lin CL, Huang HC, Lin JK. Theaflavins attenuate hepatic lipid accumulation through activating AMPK in human HepG2 cells. Journal of lipid research. 2007;48:2334–43. doi: 10.1194/jlr.M700128-JLR200. [DOI] [PubMed] [Google Scholar]
- 53.Rodriguez-Ramiro I, Vauzour D, Minihane AM. Polyphenols and non-alcoholic fatty liver disease: impact and mechanisms. The Proceedings of the Nutrition Society. 2016;75:47–60. doi: 10.1017/S0029665115004218. [DOI] [PubMed] [Google Scholar]
- 54.Assini JM, Mulvihill EE, Sutherland BG, Telford DE, Sawyez CG, Felder SL, Chhoker S, Edwards JY, Gros R, Huff MW. Naringenin prevents cholesterol-induced systemic inflammation, metabolic dysregulation, and atherosclerosis in Ldlr(−)/(−) mice. Journal of lipid research. 2013;54:711–24. doi: 10.1194/jlr.M032631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cai EP, Lin JK. Epigallocatechin gallate (EGCG) and rutin suppress the glucotoxicity through activating IRS2 and AMPK signaling in rat pancreatic beta cells. Journal of agricultural and food chemistry. 2009;57:9817–27. doi: 10.1021/jf902618v. [DOI] [PubMed] [Google Scholar]
- 56.Dai X, Ding Y, Zhang Z, Cai X, Li Y. Quercetin and quercitrin protect against cytokineinduced injuries in RINm5F beta-cells via the mitochondrial pathway and NF-kappaB signaling. International journal of molecular medicine. 2013;31:265–71. doi: 10.3892/ijmm.2012.1177. [DOI] [PubMed] [Google Scholar]
- 57.Fu Z, Zhang W, Zhen W, Lum H, Nadler J, Bassaganya-Riera J, Jia Z, Wang Y, Misra H, Liu D. Genistein induces pancreatic beta-cell proliferation through activation of multiple signaling pathways and prevents insulin-deficient diabetes in mice. Endocrinology. 2010;151:3026–37. doi: 10.1210/en.2009-1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Bardy G, Virsolvy A, Quignard JF, Ravier MA, Bertrand G, Dalle S, Cros G, Magous R, Richard S, Oiry C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. British journal of pharmacology. 2013;169:1102–13. doi: 10.1111/bph.12194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kappel VD, Frederico MJ, Postal BG, Mendes CP, Cazarolli LH, Silva FR. The role of calcium in intracellular pathways of rutin in rat pancreatic islets: potential insulin secretagogue effect. European journal of pharmacology. 2013;702:264–8. doi: 10.1016/j.ejphar.2013.01.055. [DOI] [PubMed] [Google Scholar]
- 60.Clark JL, Zahradka P, Taylor CG. Efficacy of flavonoids in the management of high blood pressure. Nutrition reviews. 2015;73:799–822. doi: 10.1093/nutrit/nuv048. [DOI] [PubMed] [Google Scholar]
- 61.Li PG, Sun L, Han X, Ling S, Gan WT, Xu JW. Quercetin induces rapid eNOS phosphorylation and vasodilation by an Akt-independent and PKA-dependent mechanism. Pharmacology. 2012;89:220–8. doi: 10.1159/000337182. [DOI] [PubMed] [Google Scholar]
- 62.Olaleye MT, Crown OO, Akinmoladun AC, Akindahunsi AA. Rutin and quercetin show greater efficacy than nifedipin in ameliorating hemodynamic, redox, and metabolite imbalances in sodium chloride-induced hypertensive rats. Human & experimental toxicology. 2014;33:602–8. doi: 10.1177/0960327113504790. [DOI] [PubMed] [Google Scholar]
- 63.Rivera L, Moron R, Sanchez M, Zarzuelo A, Galisteo M. Quercetin ameliorates metabolic syndrome and improves the inflammatory status in obese Zucker rats. Obesity (Silver Spring, Md) 2008;16:2081–7. doi: 10.1038/oby.2008.315. [DOI] [PubMed] [Google Scholar]
- 64.Croft KD. Dietary polyphenols: Antioxidants or not? Archives of biochemistry and biophysics. 2016;595:120–4. doi: 10.1016/j.abb.2015.11.014. [DOI] [PubMed] [Google Scholar]
- 65.Kukongviriyapan U, Sompamit K, Pannangpetch P, Kukongviriyapan V, Donpunha W. Preventive and therapeutic effects of quercetin on lipopolysaccharide-induced oxidative stress and vascular dysfunction in mice. Canadian journal of physiology and pharmacology. 2012;90:1345–53. doi: 10.1139/y2012-101. [DOI] [PubMed] [Google Scholar]
- 66.Liu PL, Liu JT, Kuo HF, Chong IW, Hsieh CC. Epigallocatechin gallate attenuates proliferation and oxidative stress in human vascular smooth muscle cells induced by interleukin-1beta via heme oxygenase-1. Mediators of inflammation. 2014;2014:523684. doi: 10.1155/2014/523684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Xu C, Chen J, Zhang J, Hu X, Zhou X, Lu Z, Jiang H. Naringenin inhibits angiotensin II-induced vascular smooth muscle cells proliferation and migration and decreases neointimal hyperplasia in balloon injured rat carotid arteries through suppressing oxidative stress. Biological & pharmaceutical bulletin. 2013;36:1549–55. doi: 10.1248/bpb.b13-00247. [DOI] [PubMed] [Google Scholar]
- 68.Fusi F, Spiga O, Trezza A, Sgaragli G, Saponara S. The surge of flavonoids as novel, fine regulators of cardiovascular Cav channels. European journal of pharmacology. 2017;796:158–174. doi: 10.1016/j.ejphar.2016.12.033. [DOI] [PubMed] [Google Scholar]
- 69.Mulvihill EE, Allister EM, Sutherland BG, Telford DE, Sawyez CG, Edwards JY, Markle JM, Hegele RA, Huff MW. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor-null mice with diet-induced insulin resistance. Diabetes. 2009;58:2198–210. doi: 10.2337/db09-0634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hoek-van den Hil EF, van Schothorst EM, van der Stelt I, Swarts HJ, Venema D, Sailer M, Vervoort JJ, Hollman PC, Rietjens IM, Keijer J. Quercetin decreases high-fat diet induced body weight gain and accumulation of hepatic and circulating lipids in mice. Genes & nutrition. 2014;9:418. doi: 10.1007/s12263-014-0418-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Tsutsumi R, Yoshida T, Nii Y, Okahisa N, Iwata S, Tsukayama M, Hashimoto R, Taniguchi Y, Sakaue H, Hosaka T, Shuto E, Sakai T. Sudachitin, a polymethoxylated flavone, improves glucose and lipid metabolism by increasing mitochondrial biogenesis in skeletal muscle. Nutrition & metabolism. 2014;11:32. doi: 10.1186/1743-7075-11-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhang X, Zhang QX, Wang X, Zhang L, Qu W, Bao B, Liu CA, Liu J. Dietary luteolin activates browning and thermogenesis in mice through an AMPK/PGC1alpha pathway-mediated mechanism. International journal of obesity (2005) 2016;40:1841–1849. doi: 10.1038/ijo.2016.108. [DOI] [PubMed] [Google Scholar]
- 73.Choi JH, Yun JW. Chrysin induces brown fat-like phenotype and enhances lipid metabolism in 3T3-L1 adipocytes. Nutrition (Burbank, Los Angeles County, Calif) 2016;32:1002–10. doi: 10.1016/j.nut.2016.02.007. [DOI] [PubMed] [Google Scholar]
- 74.Kamio N, Suzuki T, Watanabe Y, Suhara Y, Osakabe N. A single oral dose of flavan-3-ols enhances energy expenditure by sympathetic nerve stimulation in mice. Free radical biology & medicine. 2016;91:256–63. doi: 10.1016/j.freeradbiomed.2015.12.030. [DOI] [PubMed] [Google Scholar]
- 75.Watanabe N, Inagawa K, Shibata M, Osakabe N. Flavan-3-ol fraction from cocoa powder promotes mitochondrial biogenesis in skeletal muscle in mice. Lipids in health and disease. 2014;13:64. doi: 10.1186/1476-511X-13-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Okla M, Kang I, Kim DM, Gourineni V, Shay N, Gu L, Chung S. Ellagic acid modulates lipid accumulation in primary human adipocytes and human hepatoma Huh7 cells via discrete mechanisms. The Journal of nutritional biochemistry. 2015;26:82–90. doi: 10.1016/j.jnutbio.2014.09.010. [DOI] [PubMed] [Google Scholar]
- 77.Murase T, Misawa K, Minegishi Y, Aoki M, Ominami H, Suzuki Y, Shibuya Y, Hase T. Coffee polyphenols suppress diet-induced body fat accumulation by downregulating SREBP-1c and related molecules in C57BL/6J mice. American journal of physiology Endocrinology and metabolism. 2011;300:E122–33. doi: 10.1152/ajpendo.00441.2010. [DOI] [PubMed] [Google Scholar]
- 78.Tung YC, Li S, Huang Q, Hung WL, Ho CT, Wei GJ, Pan MH. 5-Demethylnobiletin and 5-Acetoxy-6,7,8,3′,4′-pentamethoxyflavone Suppress Lipid Accumulation by Activating the LKB1-AMPK Pathway in 3T3-L1 Preadipocytes and High Fat Diet-Fed C57BL/6 Mice. Journal of agricultural and food chemistry. 2016;64:3196–205. doi: 10.1021/acs.jafc.6b00706. [DOI] [PubMed] [Google Scholar]
- 79.Dong J, Zhang X, Zhang L, Bian HX, Xu N, Bao B, Liu J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: a mechanism including AMPKalpha1/SIRT1. Journal of lipid research. 2014;55:363–74. doi: 10.1194/jlr.M038786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lv D, Cheng X, Tang L, Jiang M. The cardioprotective effect of total flavonoids on myocardial ischemia/reperfusion in rats. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie. 2017;88:277–284. doi: 10.1016/j.biopha.2017.01.060. [DOI] [PubMed] [Google Scholar]
- 81.Mantawy EM, El-Bakly WM, Esmat A, Badr AM, El-Demerdash E. Chrysin alleviates acute doxorubicin cardiotoxicity in rats via suppression of oxidative stress, inflammation and apoptosis. European journal of pharmacology. 2014;728:107–18. doi: 10.1016/j.ejphar.2014.01.065. [DOI] [PubMed] [Google Scholar]
- 82.Prince PD, Fischerman L, Toblli JE, Fraga CG, Galleano M. LPS-induced renal inflammation is prevented by (−)-epicatechin in rats. Redox biology. 2016;11:342–349. doi: 10.1016/j.redox.2016.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Rani N, Bharti S, Bhatia J, Nag TC, Ray R, Arya DS. Chrysin, a PPAR-gamma agonist improves myocardial injury in diabetic rats through inhibiting AGE-RAGE mediated oxidative stress and inflammation. Chemico-biological interactions. 2016;250:59–67. doi: 10.1016/j.cbi.2016.03.015. [DOI] [PubMed] [Google Scholar]
- 84.Yuan X, Niu HT, Wang PL, Lu J, Zhao H, Liu SH, Zheng QS, Li CG. Cardioprotective Effect of Licochalcone D against Myocardial Ischemia/Reperfusion Injury in Langendorff-Perfused Rat Hearts. PloS one. 2015;10:e0128375. doi: 10.1371/journal.pone.0128375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zempo H, Suzuki J, Watanabe R, Wakayama K, Kumagai H, Ikeda Y, Akazawa H, Komuro I, Isobe M. Cacao polyphenols ameliorate autoimmune myocarditis in mice. Hypertension research: official journal of the Japanese Society of Hypertension. 2016;39:203–9. doi: 10.1038/hr.2015.136. [DOI] [PubMed] [Google Scholar]
- 86.Erlejman AG, Jaggers G, Fraga CG, Oteiza PI. TNFalpha-induced NF-kappaB activation and cell oxidant production are modulated by hexameric procyanidins in Caco-2 cells. Archives of biochemistry and biophysics. 2008;476:186–95. doi: 10.1016/j.abb.2008.01.024. [DOI] [PubMed] [Google Scholar]
- 87.Corti R, Flammer AJ, Hollenberg NK, Luscher TF. Cocoa and cardiovascular health. Circulation. 2009;119:1433–41. doi: 10.1161/CIRCULATIONAHA.108.827022. [DOI] [PubMed] [Google Scholar]
- 88.Hooper L, Kay C, Abdelhamid A, Kroon PA, Cohn JS, Rimm EB, Cassidy A. Effects of chocolate, cocoa, and flavan-3-ols on cardiovascular health: a systematic review and meta-analysis of randomized trials. The American journal of clinical nutrition. 2012;95:740–51. doi: 10.3945/ajcn.111.023457. [DOI] [PubMed] [Google Scholar]
- 89.Shrime MG, Bauer SR, McDonald AC, Chowdhury NH, Coltart CE, Ding EL. Flavonoid-rich cocoa consumption affects multiple cardiovascular risk factors in a meta-analysis of short-term studies. The Journal of nutrition. 2011;141:1982–8. doi: 10.3945/jn.111.145482. [DOI] [PubMed] [Google Scholar]
- 90.Taubert D, Roesen R, Lehmann C, Jung N, Schomig E. Effects of low habitual cocoa intake on blood pressure and bioactive nitric oxide: a randomized controlled trial. JAMA. 2007;298:49–60. doi: 10.1001/jama.298.1.49. [DOI] [PubMed] [Google Scholar]
- 91.Yarmolinsky J, Gon G, Edwards P. Effect of tea on blood pressure for secondary prevention of cardiovascular disease: a systematic review and meta-analysis of randomized controlled trials. Nutrition reviews. 2015;73:236–46. doi: 10.1093/nutrit/nuv001. [DOI] [PubMed] [Google Scholar]
- 92.Huang H, Chen G, Liao D, Zhu Y, Xue X. Effects of Berries Consumption on Cardiovascular Risk Factors: A Meta-analysis with Trial Sequential Analysis of Randomized Controlled Trials. Scientific reports. 2016;6:23625. doi: 10.1038/srep23625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Jacques PF, Cassidy A, Rogers G, Peterson JJ, Meigs JB, Dwyer JT. Higher dietary flavonol intake is associated with lower incidence of type 2 diabetes. The Journal of nutrition. 2013;143:1474–80. doi: 10.3945/jn.113.177212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Cassidy A, Mukamal KJ, Liu L, Franz M, Eliassen AH, Rimm EB. High anthocyanin intake is associated with a reduced risk of myocardial infarction in young and middle-aged women. Circulation. 2013;127:188–96. doi: 10.1161/CIRCULATIONAHA.112.122408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Cassidy A, Bertoia M, Chiuve S, Flint A, Forman J, Rimm EB. Habitual intake of anthocyanins and flavanones and risk of cardiovascular disease in men. The American journal of clinical nutrition. 2016;104:587–94. doi: 10.3945/ajcn.116.133132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Goetz ME, Judd SE, Hartman TJ, McClellan W, Anderson A, Vaccarino V. Flavanone Intake Is Inversely Associated with Risk of Incident Ischemic Stroke in the REasons for Geographic and Racial Differences in Stroke (REGARDS) Study. The Journal of nutrition. 2016;146:2233–2243. doi: 10.3945/jn.116.230185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Goetz ME, Judd SE, Safford MM, Hartman TJ, McClellan WM, Vaccarino V. Dietary flavonoid intake and incident coronary heart disease: the REasons for Geographic and Racial Differences in Stroke (REGARDS) study. The American journal of clinical nutrition. 2016;104:1236–1244. doi: 10.3945/ajcn.115.129452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Buitrago-Lopez A, Sanderson J, Johnson L, Warnakula S, Wood A, Di Angelantonio E, Franco OH. Chocolate consumption and cardiometabolic disorders: systematic review and meta-analysis. BMJ (Clinical research ed) 2011;343:d4488. doi: 10.1136/bmj.d4488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Guo X, Yang B, Tan J, Jiang J, Li D. Associations of dietary intakes of anthocyanins and berry fruits with risk of type 2 diabetes mellitus: a systematic review and meta-analysis of prospective cohort studies. European journal of clinical nutrition. 2016;70:1360–1367. doi: 10.1038/ejcn.2016.142. [DOI] [PubMed] [Google Scholar]
- 100.Zamora-Ros R, Touillaud M, Rothwell JA, Romieu I, Scalbert A. Measuring exposure to the polyphenol metabolome in observational epidemiologic studies: current tools and applications and their limits. The American journal of clinical nutrition. 2014;100:11–26. doi: 10.3945/ajcn.113.077743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Rienks J, Barbaresko J, Nothlings U. Association of isoflavone biomarkers with risk of chronic disease and mortality: a systematic review and meta-analysis of observational studies. Nutrition reviews. 2017;75:616–641. doi: 10.1093/nutrit/nux021. [DOI] [PubMed] [Google Scholar]
- 102.Bredsdorff L, Obel T, Dethlefsen C, Tjonneland A, Schmidt EB, Rasmussen SE, Overvad K. Urinary flavonoid excretion and risk of acute coronary syndrome in a nested case-control study. The American journal of clinical nutrition. 2013;98:209–16. doi: 10.3945/ajcn.112.046169. [DOI] [PubMed] [Google Scholar]
- 103.Sun Q, Wedick NM, Tworoger SS, Pan A, Townsend MK, Cassidy A, Franke AA, Rimm EB, Hu FB, van Dam RM. Urinary Excretion of Select Dietary Polyphenol Metabolites Is Associated with a Lower Risk of Type 2 Diabetes in Proximate but Not Remote Follow-Up in a Prospective Investigation in 2 Cohorts of US Women. The Journal of nutrition. 2015;145:1280–8. doi: 10.3945/jn.114.208736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Yu D, Shu XO, Li H, Yang G, Cai Q, Xiang YB, Ji BT, Franke AA, Gao YT, Zheng W, Zhang X. Dietary isoflavones, urinary isoflavonoids, and risk of ischemic stroke in women. The American journal of clinical nutrition. 2015;102:680–6. doi: 10.3945/ajcn.115.111591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhang X, Gao YT, Yang G, Li H, Cai Q, Xiang YB, Ji BT, Franke AA, Zheng W, Shu XO. Urinary isoflavonoids and risk of coronary heart disease. International journal of epidemiology. 2012;41:1367–75. doi: 10.1093/ije/dys130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Casas R, Sacanella E, Urpi-Sarda M, Chiva-Blanch G, Ros E, Martinez-Gonzalez MA, Covas MI, Salas-Salvado J, Fiol M, Aros F, Estruch R. The effects of the mediterranean diet on biomarkers of vascular wall inflammation and plaque vulnerability in subjects with high risk for cardiovascular disease. A randomized trial. PloS one. 2014;9:e100084. doi: 10.1371/journal.pone.0100084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Casas R, Sacanella E, Urpi-Sarda M, Corella D, Castaner O, Lamuela-Raventos RM, Salas-Salvado J, Martinez-Gonzalez MA, Ros E, Estruch R. Long-Term Immunomodulatory Effects of a Mediterranean Diet in Adults at High Risk of Cardiovascular Disease in the PREvencion con DIeta MEDiterranea (PREDIMED) Randomized Controlled Trial. The Journal of nutrition. 2016;146:1684–93. doi: 10.3945/jn.115.229476. [DOI] [PubMed] [Google Scholar]
- 108.Hernaez A, Castaner O, Goday A, Ros E, Pinto X, Estruch R, Salas-Salvado J, Corella D, Aros F, Serra-Majem L, Martinez-Gonzalez MA, Fiol M, Lapetra J, de la Torre R, Lopez-Sabater MC, Fito M. The Mediterranean Diet decreases LDL atherogenicity in high cardiovascular risk individuals: a randomized controlled trial. Molecular nutrition & food research. 2017:61. doi: 10.1002/mnfr.201601015. [DOI] [PubMed] [Google Scholar]
- 109.Estruch R, Ros E, Salas-Salvado J, Covas MI, Corella D, Aros F, Gomez-Gracia E, Ruiz-Gutierrez V, Fiol M, Lapetra J, Lamuela-Raventos RM, Serra-Majem L, Pinto X, Basora J, Munoz MA, Sorli JV, Martinez JA, Martinez-Gonzalez MA. Primary prevention of cardiovascular disease with a Mediterranean diet. The New England journal of medicine. 2013;368:1279–90. doi: 10.1056/NEJMc1806491. [DOI] [PubMed] [Google Scholar]
- 110.Estruch R, Martinez-Gonzalez MA, Corella D, Salas-Salvado J, Fito M, Chiva-Blanch G, Fiol M, Gomez-Gracia E, Aros F, Lapetra J, Serra-Majem L, Pinto X, Buil-Cosiales P, Sorli JV, Munoz MA, Basora-Gallisa J, Lamuela-Raventos RM, Serra-Mir M, Ros E. Effect of a high-fat Mediterranean diet on bodyweight and waist circumference: a prespecified secondary outcomes analysis of the PREDIMED randomised controlled trial. The lancet Diabetes & endocrinology. 2016;4:666–76. doi: 10.1016/S2213-8587(16)30085-7. [DOI] [PubMed] [Google Scholar]
- 111.Aune D, Keum N, Giovannucci E, Fadnes LT, Boffetta P, Greenwood DC, Tonstad S, Vatten LJ, Riboli E, Norat T. Nut consumption and risk of cardiovascular disease, total cancer, all-cause and cause-specific mortality: a systematic review and dose-response meta-analysis of prospective studies. BMC medicine. 2016;14:207. doi: 10.1186/s12916-016-0730-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Products NRCUCoTOtItNAoA. Designing Foods: Animal Product Options in the Marketplace. Washington (DC): National Academies Press (US); 1988. Current Trends in Consumption of Animal Products. [PubMed] [Google Scholar]
- 113.Dietary Guidelines Advisory Committee. Scientific Report of the 2015 Dietary Guidelines Advisory Committee. 2015;2015 [Google Scholar]
- 114.Leung PS. The Potential Protective Action of Vitamin D in Hepatic Insulin Resistance and Pancreatic Islet Dysfunction in Type 2 Diabetes Mellitus. Nutrients. 2016;8:147. doi: 10.3390/nu8030147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Muscogiuri G, Annweiler C, Duval G, Karras S, Tirabassi G, Salvio G, Balercia G, Kimball S, Kotsa K, Mascitelli L, Bhattoa HP, Colao A. Vitamin D and cardiovascular disease: From atherosclerosis to myocardial infarction and stroke. International journal of cardiology. 2017;230:577–584. doi: 10.1016/j.ijcard.2016.12.053. [DOI] [PubMed] [Google Scholar]
- 116.Conceicao EP, Moura EG, Soares PN, Ai XX, Figueiredo MS, Oliveira E, Lisboa PC. High calcium diet improves the liver oxidative stress and microsteatosis in adult obese rats that were overfed during lactation. Food and chemical toxicology: an international journal published for the British Industrial Biological Research Association. 2016;92:245–55. doi: 10.1016/j.fct.2016.04.015. [DOI] [PubMed] [Google Scholar]
- 117.Sun X, Zemel MB. Dietary calcium regulates ROS production in aP2-agouti transgenic mice on high-fat/high-sucrose diets. International journal of obesity (2005) 2006;30:1341–6. doi: 10.1038/sj.ijo.0803294. [DOI] [PubMed] [Google Scholar]
- 118.Nobre JL, Lisboa PC, Lima Nda S, Franco JG, Nogueira Neto JF, de Moura EG, de Oliveira E. Calcium supplementation prevents obesity, hyperleptinaemia and hyperglycaemia in adult rats programmed by early weaning. The British journal of nutrition. 2012;107:979–88. doi: 10.1017/S0007114511003928. [DOI] [PubMed] [Google Scholar]
- 119.Conceicao EP, Moura EG, Oliveira E, Guarda DS, Figueiredo MS, Quitete FT, Calvino C, Miranda RA, Mathias PC, Manhaes AC, Lisboa PC. Dietary calcium supplementation in adult rats reverts brown adipose tissue dysfunction programmed by postnatal early overfeeding. The Journal of nutritional biochemistry. 2017;39:117–125. doi: 10.1016/j.jnutbio.2016.09.013. [DOI] [PubMed] [Google Scholar]
- 120.Quitete FT, Nobre JL, Peixoto-Silva N, de Moura EG, Lisboa PC, de Oliveira E. Anti-obesogenic effects of calcium prevent changes in the GLP-1 profile in adult rats primed by early weaning. Molecular nutrition & food research. 2015;59:773–83. doi: 10.1002/mnfr.201400666. [DOI] [PubMed] [Google Scholar]
- 121.Sun X, Zemel MB. Calcium and 1,25-dihydroxyvitamin D3 regulation of adipokine expression. Obesity (Silver Spring, Md) 2007;15:340–8. doi: 10.1038/oby.2007.540. [DOI] [PubMed] [Google Scholar]
- 122.Sun X, Zemel MB. Calcitriol and calcium regulate cytokine production and adipocyte-macrophage cross-talk. The Journal of nutritional biochemistry. 2008;19:392–9. doi: 10.1016/j.jnutbio.2007.05.013. [DOI] [PubMed] [Google Scholar]
- 123.Chaplin A, Parra P, Laraichi S, Serra F, Palou A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Molecular nutrition & food research. 2016;60:468–80. doi: 10.1002/mnfr.201500480. [DOI] [PubMed] [Google Scholar]
- 124.Gomes JM, Costa JA, Alfenas RC. Could the beneficial effects of dietary calcium on obesity and diabetes control be mediated by changes in intestinal microbiota and integrity? The British journal of nutrition. 2015;114:1756–65. doi: 10.1017/S0007114515003608. [DOI] [PubMed] [Google Scholar]
- 125.de Wit NJ, Bosch-Vermeulen H, Oosterink E, Muller M, van der Meer R. Supplementary dietary calcium stimulates faecal fat and bile acid excretion, but does not protect against obesity and insulin resistance in C57BL/6J mice. The British journal of nutrition. 2011;105:1005–11. doi: 10.1017/S0007114510004654. [DOI] [PubMed] [Google Scholar]
- 126.Thomas AP, Dunn TN, Drayton JB, Oort PJ, Adams SH. A high calcium diet containing nonfat dry milk reduces weight gain and associated adipose tissue inflammation in diet-induced obese mice when compared to high calcium alone. Nutrition & metabolism. 2012;9:3. doi: 10.1186/1743-7075-9-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Thomas AP, Dunn TN, Drayton JB, Oort PJ, Adams SH. A dairy-based high calcium diet improves glucose homeostasis and reduces steatosis in the context of preexisting obesity. Obesity (Silver Spring, Md) 2013;21:E229–35. doi: 10.1002/oby.20039. [DOI] [PubMed] [Google Scholar]
- 128.Booth AO, Huggins CE, Wattanapenpaiboon N, Nowson CA. Effect of increasing dietary calcium through supplements and dairy food on body weight and body composition: a meta-analysis of randomised controlled trials. Br J Nutr. 2015;114:1013–25. doi: 10.1017/S0007114515001518. [DOI] [PubMed] [Google Scholar]
- 129.Cormick G, Ciapponi A, Cafferata ML, Belizan JM. Calcium supplementation for prevention of primary hypertension. The Cochrane database of systematic reviews. 2015:CD010037. doi: 10.1002/14651858.CD010037.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Jolma P, Kalliovalkama J, Tolvanen JP, Koobi P, Kahonen M, Hutri-Kahonen N, Wu X, Porsti I. High-calcium diet enhances vasorelaxation in nitric oxide-deficient hypertension. American journal of physiology Heart and circulatory physiology. 2000;279:H1036–43. doi: 10.1152/ajpheart.2000.279.3.H1036. [DOI] [PubMed] [Google Scholar]
- 131.Makynen H, Kahonen M, Wu X, Arvola P, Porsti I. Endothelial function in deoxycorticosterone-NaCl hypertension: effect of calcium supplementation. Circulation. 1996;93:1000–8. doi: 10.1161/01.cir.93.5.1000. [DOI] [PubMed] [Google Scholar]
- 132.McCarron DA, Lucas PA, Shneidman RJ, LaCour B, Drueke T. Blood pressure development of the spontaneously hypertensive rat after concurrent manipulations of dietary Ca2+ and Na+. Relation to intestinal Ca2+ fluxes. The Journal of clinical investigation. 1985;76:1147–54. doi: 10.1172/JCI112070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Porsti I, Fan M, Koobi P, Jolma P, Kalliovalkama J, Vehmas TI, Helin H, Holthofer H, Mervaala E, Nyman T, Tikkanen I. High calcium diet down-regulates kidney angiotensin-converting enzyme in experimental renal failure. Kidney international. 2004;66:2155–66. doi: 10.1111/j.1523-1755.2004.66006.x. [DOI] [PubMed] [Google Scholar]
- 134.Bolland MJ, Grey A, Avenell A, Gamble GD, Reid IR. Calcium supplements with or without vitamin D and risk of cardiovascular events: reanalysis of the Women’s Health Initiative limited access dataset and meta-analysis. BMJ (Clinical research ed) 2011;342:d2040. doi: 10.1136/bmj.d2040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Mao PJ, Zhang C, Tang L, Xian YQ, Li YS, Wang WD, Zhu XH, Qiu HL, He J, Zhou YH. Effect of calcium or vitamin D supplementation on vascular outcomes: a meta-analysis of randomized controlled trials. International journal of cardiology. 2013;169:106–11. doi: 10.1016/j.ijcard.2013.08.055. [DOI] [PubMed] [Google Scholar]
- 136.Larsson SC, Burgess S, Michaelsson K. Association of Genetic Variants Related to Serum Calcium Levels With Coronary Artery Disease and Myocardial Infarction. Jama. 2017;318:371–380. doi: 10.1001/jama.2017.8981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Fekete AA, Givens DI, Lovegrove JA. Can milk proteins be a useful tool in the management of cardiometabolic health? An updated review of human intervention trials. The Proceedings of the Nutrition Society. 2016;75:328–41. doi: 10.1017/S0029665116000264. [DOI] [PubMed] [Google Scholar]
- 138.Healy NP, Kirwan AM, McArdle MA, Holohan K, Nongonierma AB, Keane D, Kelly S, Celkova L, Lyons CL, McGillicuddy FC, Finucane OM, Murray BA, Kelly PM, Brennan L, FitzGerald RJ, Roche HM. A casein hydrolysate protects mice against high fat diet induced hyperglycemia by attenuating NLRP3 inflammasome-mediated inflammation and improving insulin signaling. Molecular nutrition & food research. 2016;60:2421–2432. doi: 10.1002/mnfr.201501054. [DOI] [PubMed] [Google Scholar]
- 139.Pezeshki A, Fahim A, Chelikani PK. Dietary Whey and Casein Differentially Affect Energy Balance, Gut Hormones, Glucose Metabolism, and Taste Preference in Diet-Induced Obese Rats. The Journal of nutrition. 2015;145:2236–44. doi: 10.3945/jn.115.213843. [DOI] [PubMed] [Google Scholar]
- 140.Singh A, Pezeshki A, Zapata RC, Yee NJ, Knight CG, Tuor UI, Chelikani PK. Diets enriched in whey or casein improve energy balance and prevent morbidity and renal damage in salt-loaded and high-fat-fed spontaneously hypertensive stroke-prone rats. The Journal of nutritional biochemistry. 2016;37:47–59. doi: 10.1016/j.jnutbio.2016.07.011. [DOI] [PubMed] [Google Scholar]
- 141.Li H, Xu M, Lee J, He C, Xie Z. Leucine supplementation increases SIRT1 expression and prevents mitochondrial dysfunction and metabolic disorders in high-fat diet-induced obese mice. American journal of physiology Endocrinology and metabolism. 2012;303:E1234–44. doi: 10.1152/ajpendo.00198.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Ren M, Zhang S, Liu X, Li S, Mao X, Zeng X, Qiao S. Different Lipopolysaccharide Branched-Chain Amino Acids Modulate Porcine Intestinal Endogenous beta-Defensin Expression through the Sirt1/ERK/90RSK Pathway. Journal of agricultural and food chemistry. 2016;64:3371–9. doi: 10.1021/acs.jafc.6b00968. [DOI] [PubMed] [Google Scholar]
- 143.Jakubowicz D, Froy O. Biochemical and metabolic mechanisms by which dietary whey protein may combat obesity and Type 2 diabetes. The Journal of nutritional biochemistry. 2013;24:1–5. doi: 10.1016/j.jnutbio.2012.07.008. [DOI] [PubMed] [Google Scholar]
- 144.Tong X, Li W, Xu JY, Han S, Qin LQ. Effects of whey protein and leucine supplementation on insulin resistance in non-obese insulin-resistant model rats. Nutrition (Burbank, Los Angeles County, Calif) 2014;30:1076–80. doi: 10.1016/j.nut.2014.01.013. [DOI] [PubMed] [Google Scholar]
- 145.Newgard CB, An J, Bain JR, Muehlbauer MJ, Stevens RD, Lien LF, Haqq AM, Shah SH, Arlotto M, Slentz CA, Rochon J, Gallup D, Ilkayeva O, Wenner BR, Yancy WS, Jr, Eisenson H, Musante G, Surwit RS, Millington DS, Butler MD, Svetkey LP. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell metabolism. 2009;9:311–26. doi: 10.1016/j.cmet.2009.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Zhenyukh O, Civantos E, Ruiz-Ortega M, Sanchez MS, Vazquez C, Peiro C, Egido J, Mas S. High concentration of branched-chain amino acids promotes oxidative stress, inflammation and migration of human peripheral blood mononuclear cells via mTORC1 activation. Free radical biology & medicine. 2017;104:165–177. doi: 10.1016/j.freeradbiomed.2017.01.009. [DOI] [PubMed] [Google Scholar]
- 147.Pedroso JA, Zampieri TT, Donato J., Jr Reviewing the Effects of L-Leucine Supplementation in the Regulation of Food Intake, Energy Balance, and Glucose Homeostasis. Nutrients. 2015;7:3914–37. doi: 10.3390/nu7053914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Fekete AA, Givens DI, Lovegrove JA. The impact of milk proteins and peptides on blood pressure and vascular function: a review of evidence from human intervention studies. Nutrition research reviews. 2013;26:177–90. doi: 10.1017/S0954422413000139. [DOI] [PubMed] [Google Scholar]
- 149.Fekete AA, Givens DI, Lovegrove JA. Casein-derived lactotripeptides reduce systolic and diastolic blood pressure in a meta-analysis of randomised clinical trials. Nutrients. 2015;7:659–81. doi: 10.3390/nu7010659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Capriotti AL, Cavaliere C, Piovesana S, Samperi R, Lagana A. Recent trends in the analysis of bioactive peptides in milk and dairy products. Analytical and bioanalytical chemistry. 2016;408:2677–85. doi: 10.1007/s00216-016-9303-8. [DOI] [PubMed] [Google Scholar]
- 151.Jakala P, Vapaatalo H. Antihypertensive Peptides from Milk Proteins. Pharmaceuticals (Basel, Switzerland) 2010;3:251–272. doi: 10.3390/ph3010251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Zhang Y, Chen R, Ma H, Chen S. Isolation and Identification of Dipeptidyl Peptidase IV-Inhibitory Peptides from Trypsin/Chymotrypsin-Treated Goat Milk Casein Hydrolysates by 2D-TLC and LC-MS/MS. Journal of agricultural and food chemistry. 2015;63:8819–28. doi: 10.1021/acs.jafc.5b03062. [DOI] [PubMed] [Google Scholar]
- 153.Tulipano G, Sibilia V, Caroli AM, Cocchi D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides. 2011;32:835–8. doi: 10.1016/j.peptides.2011.01.002. [DOI] [PubMed] [Google Scholar]
- 154.Foltz M, van der Pijl PC, Duchateau GS. Current in vitro testing of bioactive peptides is not valuable. The Journal of nutrition. 2010;140:117–8. doi: 10.3945/jn.109.116228. [DOI] [PubMed] [Google Scholar]
- 155.Micha R, Mozaffarian D. Saturated fat and cardiometabolic risk factors, coronary heart disease, stroke, and diabetes: a fresh look at the evidence. Lipids. 2010;45:893–905. doi: 10.1007/s11745-010-3393-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Markiewicz-Keszycka M, Czyzak-Runowska G, Lipinska P, Wojtowski J. Fatty Acid Profile of Milk - A Review. Bulletin of the Veterinary Institute in Pulawy. 2013;57:135. [Google Scholar]
- 157.Mansson HL. Fatty acids in bovine milk fat. Food & nutrition research. 2008;52 doi: 10.3402/fnr.v52i0.1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Ran-Ressler RR, Bae S, Lawrence P, Wang DH, Brenna JT. Branched-chain fatty acid content of foods and estimated intake in the USA. The British journal of nutrition. 2014;112:565–72. doi: 10.1017/S0007114514001081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Hommelberg PP, Plat J, Langen RC, Schols AM, Mensink RP. Fatty acid-induced NF-kappaB activation and insulin resistance in skeletal muscle are chain length dependent. American journal of physiology Endocrinology and metabolism. 2009;296:E114–20. doi: 10.1152/ajpendo.00436.2007. [DOI] [PubMed] [Google Scholar]
- 160.Montgomery MK, Osborne B, Brown SH, Small L, Mitchell TW, Cooney GJ, Turner N. Contrasting metabolic effects of medium- versus long-chain fatty acids in skeletal muscle. Journal of lipid research. 2013;54:3322–33. doi: 10.1194/jlr.M040451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Turner N, Hariharan K, TidAng J, Frangioudakis G, Beale SM, Wright LE, Zeng XY, Leslie SJ, Li JY, Kraegen EW, Cooney GJ, Ye JM. Enhancement of muscle mitochondrial oxidative capacity and alterations in insulin action are lipid species dependent: potent tissue-specific effects of medium-chain fatty acids. Diabetes. 2009;58:2547–54. doi: 10.2337/db09-0784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Chamma CM, Bargut TC, Mandarim-de-Lacerda CA, Aguila MB. A rich medium-chain triacylglycerol diet benefits adiposity but has adverse effects on the markers of hepatic lipogenesis and beta-oxidation. Food & function. 2017;8:778–787. doi: 10.1039/c6fo01663d. [DOI] [PubMed] [Google Scholar]
- 163.Bedi S, Hines GV, Lozada-Fernandez VV, de Jesus Piva C, Kaliappan A, Rider SD, Jr, Hostetler HA. Fatty acid binding profile of the liver X receptor alpha. Journal of lipid research. 2017;58:393–402. doi: 10.1194/jlr.M072447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Mozaffarian D. Natural trans fat, dairy fat, partially hydrogenated oils, and cardiometabolic health: the Ludwigshafen Risk and Cardiovascular Health Study. European heart journal. 2016;37:1079–81. doi: 10.1093/eurheartj/ehv595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Chan KL, Pillon NJ, Sivaloganathan DM, Costford SR, Liu Z, Theret M, Chazaud B, Klip A. Palmitoleate Reverses High Fat-induced Proinflammatory Macrophage Polarization via AMP-activated Protein Kinase (AMPK) The Journal of biological chemistry. 2015;290:16979–88. doi: 10.1074/jbc.M115.646992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Duckett SK, Volpi-Lagreca G, Alende M, Long NM. Palmitoleic acid reduces intramuscular lipid and restores insulin sensitivity in obese sheep. Diabetes, metabolic syndrome and obesity: targets and therapy. 2014;7:553–63. doi: 10.2147/DMSO.S72695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Souza CO, Teixeira AA, Biondo LA, Lima EA, Jr, Batatinha HA, Rosa Neto JC. Palmitoleic Acid Improves Metabolic Functions in Fatty Liver by PPARalpha-Dependent AMPK Activation. Journal of cellular physiology. 2016 doi: 10.1002/jcp.25715. [DOI] [PubMed] [Google Scholar]
- 168.Souza CO, Teixeira AA, Lima EA, Batatinha HA, Gomes LM, Carvalho-Silva M, Mota IT, Streck EL, Hirabara SM, Rosa Neto JC. Palmitoleic acid (n-7) attenuates the immunometabolic disturbances caused by a high-fat diet independently of PPARalpha. Mediators of inflammation. 2014;2014:582197. doi: 10.1155/2014/582197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Talbot NA, Wheeler-Jones CP, Cleasby ME. Palmitoleic acid prevents palmitic acid-induced macrophage activation and consequent p38 MAPK-mediated skeletal muscle insulin resistance. Molecular and cellular endocrinology. 2014;393:129–42. doi: 10.1016/j.mce.2014.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Bernstein AM, Roizen MF, Martinez L. Purified palmitoleic acid for the reduction of high-sensitivity C-reactive protein and serum lipids: a double-blinded, randomized, placebo controlled study. Journal of clinical lipidology. 2014;8:612–7. doi: 10.1016/j.jacl.2014.08.001. [DOI] [PubMed] [Google Scholar]
- 171.Kraft J, Jetton T, Satish B, G D. Dairy-derived bioactive fatty acids improve pancreatic ß-cell function. The FASEB Journal. 2015;29:608.25. [Google Scholar]
- 172.Mozaffarian D, Wu JH. Omega-3 Fatty Acids and Cardiovascular Disease – Effects on Risk Factors, Molecular Pathways, and Clinical Events. J Am Coll Cardiol. 2011;58:2047–67. doi: 10.1016/j.jacc.2011.06.063. [DOI] [PubMed] [Google Scholar]
- 173.Hodson L, Eyles HC, McLachlan KJ, Bell ML, Green TJ, Skeaff CM. Plasma and erythrocyte fatty acids reflect intakes of saturated and n-6 PUFA within a similar time frame. The Journal of nutrition. 2014;144:33–41. doi: 10.3945/jn.113.183749. [DOI] [PubMed] [Google Scholar]
- 174.Pfeuffer M, Jaudszus A. Pentadecanoic and Heptadecanoic Acids: Multifaceted Odd-Chain Fatty Acids. Advances in nutrition (Bethesda, Md) 2016;7:730–4. doi: 10.3945/an.115.011387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Arranz E, Corredig M. Invited review: Milk phospholipid vesicles, their colloidal properties, and potential as delivery vehicles for bioactive molecules. Journal of dairy science. 2017 doi: 10.3168/jds.2016-12236. [DOI] [PubMed] [Google Scholar]
- 176.Chung RW, Kamili A, Tandy S, Weir JM, Gaire R, Wong G, Meikle PJ, Cohn JS, Rye KA. Dietary sphingomyelin lowers hepatic lipid levels and inhibits intestinal cholesterol absorption in high-fat-fed mice. PloS one. 2013;8:e55949. doi: 10.1371/journal.pone.0055949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Kamili A, Wat E, Chung RW, Tandy S, Weir JM, Meikle PJ, Cohn JS. Hepatic accumulation of intestinal cholesterol is decreased and fecal cholesterol excretion is increased in mice fed a high-fat diet supplemented with milk phospholipids. Nutrition & metabolism. 2010;7:90. doi: 10.1186/1743-7075-7-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Wat E, Tandy S, Kapera E, Kamili A, Chung RW, Brown A, Rowney M, Cohn JS. Dietary phospholipid-rich dairy milk extract reduces hepatomegaly, hepatic steatosis and hyperlipidemia in mice fed a high-fat diet. Atherosclerosis. 2009;205:144–50. doi: 10.1016/j.atherosclerosis.2008.12.004. [DOI] [PubMed] [Google Scholar]
- 179.Snow DR, Ward RE, Olsen A, Jimenez-Flores R, Hintze KJ. Membrane-rich milk fat diet provides protection against gastrointestinal leakiness in mice treated with lipopolysaccharide. Journal of dairy science. 2011;94:2201–12. doi: 10.3168/jds.2010-3886. [DOI] [PubMed] [Google Scholar]
- 180.Rosqvist F, Smedman A, Lindmark-Mansson H, Paulsson M, Petrus P, Straniero S, Rudling M, Dahlman I, Riserus U. Potential role of milk fat globule membrane in modulating plasma lipoproteins, gene expression, and cholesterol metabolism in humans: a randomized study. The American journal of clinical nutrition. 2015;102:20–30. doi: 10.3945/ajcn.115.107045. [DOI] [PubMed] [Google Scholar]
- 181.Hjerpsted J, Leedo E, Tholstrup T. Cheese intake in large amounts lowers LDL-cholesterol concentrations compared with butter intake of equal fat content. The American journal of clinical nutrition. 2011;94:1479–84. doi: 10.3945/ajcn.111.022426. [DOI] [PubMed] [Google Scholar]
- 182.Soerensen KV, Thorning TK, Astrup A, Kristensen M, Lorenzen JK. Effect of dairy calcium from cheese and milk on fecal fat excretion, blood lipids, and appetite in young men. The American journal of clinical nutrition. 2014;99:984–91. doi: 10.3945/ajcn.113.077735. [DOI] [PubMed] [Google Scholar]
- 183.Guidelines for the Evaluation of Probiotics in Food. Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food. London Ontario: 2002. http://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf. [Google Scholar]
- 184.Kim DH, Kim H, Jeong D, Kang IB, Chon JW, Kim HS, Song KY, Seo KH. Kefir alleviates obesity and hepatic steatosis in high-fat diet-fed mice by modulation of gut microbiota and mycobiota: targeted and untargeted community analysis with correlation of biomarkers. The Journal of nutritional biochemistry. 2017;44:35–43. doi: 10.1016/j.jnutbio.2017.02.014. [DOI] [PubMed] [Google Scholar]
- 185.Singh S, Sharma RK, Malhotra S, Pothuraju R, Shandilya UK. Lactobacillus rhamnosus NCDC17 ameliorates type-2 diabetes by improving gut function, oxidative stress and inflammation in high-fat-diet fed and streptozotocintreated rats. Beneficial microbes. 2016:1–14. doi: 10.3920/BM2016.0090. [DOI] [PubMed] [Google Scholar]
- 186.Gerritsen J, Smidt H, Rijkers GT, de Vos WM. Intestinal microbiota in human health and disease: the impact of probiotics. Genes & nutrition. 2011;6:209–40. doi: 10.1007/s12263-011-0229-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.de Moreno de Leblanc A, Perdigon G. The application of probiotic fermented milks in cancer and intestinal inflammation. The Proceedings of the Nutrition Society. 2010;69:421–8. doi: 10.1017/S002966511000159X. [DOI] [PubMed] [Google Scholar]
- 188.Veiga P, Gallini CA, Beal C, Michaud M, Delaney ML, DuBois A, Khlebnikov A, van Hylckama Vlieg JE, Punit S, Glickman JN, Onderdonk A, Glimcher LH, Garrett WS. Bifidobacterium animalis subsp. lactis fermented milk product reduces inflammation by altering a niche for colitogenic microbes. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:18132–7. doi: 10.1073/pnas.1011737107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Bron PA, Kleerebezem M, Brummer RJ, Cani PD, Mercenier A, MacDonald TT, Garcia-Rodenas CL, Wells JM. Can probiotics modulate human disease by impacting intestinal barrier function? The British journal of nutrition. 2017;117:93–107. doi: 10.1017/S0007114516004037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Shirouchi B, Nagao K, Umegatani M, Shiraishi A, Morita Y, Kai S, Yanagita T, Ogawa A, Kadooka Y, Sato M. Probiotic Lactobacillus gasseri SBT2055 improves glucose tolerance and reduces body weight gain in rats by stimulating energy expenditure. The British journal of nutrition. 2016;116:451–8. doi: 10.1017/S0007114516002245. [DOI] [PubMed] [Google Scholar]
- 191.Yadav H, Lee JH, Lloyd J, Walter P, Rane SG. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. The Journal of biological chemistry. 2013;288:25088–97. doi: 10.1074/jbc.M113.452516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Barrett HL, Dekker Nitert M, Conwell LS, Callaway LK. Probiotics for preventing gestational diabetes. The Cochrane database of systematic reviews. 2014:Cd009951. doi: 10.1002/14651858.CD009951.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Jung SP, Lee KM, Kang JH, Yun SI, Park HO, Moon Y, Kim JY. Effect of Lactobacillus gasseri BNR17 on Overweight and Obese Adults: A Randomized, Double-Blind Clinical Trial. Korean journal of family medicine. 2013;34:80–9. doi: 10.4082/kjfm.2013.34.2.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Kadooka Y, Sato M, Imaizumi K, Ogawa A, Ikuyama K, Akai Y, Okano M, Kagoshima M, Tsuchida T. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. European journal of clinical nutrition. 2010;64:636–43. doi: 10.1038/ejcn.2010.19. [DOI] [PubMed] [Google Scholar]
- 195.Kadooka Y, Sato M, Ogawa A, Miyoshi M, Uenishi H, Ogawa H, Ikuyama K, Kagoshima M, Tsuchida T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. The British journal of nutrition. 2013;110:1696–703. doi: 10.1017/S0007114513001037. [DOI] [PubMed] [Google Scholar]
- 196.Mozaffarian D, Hao T, Rimm EB, Willett WC, Hu FB. Changes in diet and lifestyle and long-term weight gain in women and men. The New England journal of medicine. 2011;364:2392–404. doi: 10.1056/NEJMoa1014296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Park DY, Ahn YT, Park SH, Huh CS, Yoo SR, Yu R, Sung MK, McGregor RA, Choi MS. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PloS one. 2013;8:e59470. doi: 10.1371/journal.pone.0059470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Poutahidis T, Kleinewietfeld M, Smillie C, Levkovich T, Perrotta A, Bhela S, Varian BJ, Ibrahim YM, Lakritz JR, Kearney SM, Chatzigiagkos A, Hafler DA, Alm EJ, Erdman SE. Microbial reprogramming inhibits Western diet-associated obesity. PloS one. 2013;8:e68596. doi: 10.1371/journal.pone.0068596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Power SE, O’Toole PW, Stanton C, Ross RP, Fitzgerald GF. Intestinal microbiota, diet and health. The British journal of nutrition. 2014;111:387–402. doi: 10.1017/S0007114513002560. [DOI] [PubMed] [Google Scholar]
- 200.Sanchez M, Darimont C, Drapeau V, Emady-Azar S, Lepage M, Rezzonico E, Ngom-Bru C, Berger B, Philippe L, Ammon-Zuffrey C, Leone P, Chevrier G, St-Amand E, Marette A, Dore J, Tremblay A. Effect of Lactobacillus rhamnosus CGMCC1.3724 supplementation on weight loss and maintenance in obese men and women. The British journal of nutrition. 2014;111:1507–19. doi: 10.1017/S0007114513003875. [DOI] [PubMed] [Google Scholar]
- 201.Sharafedtinov KK, Plotnikova OA, Alexeeva RI, Sentsova TB, Songisepp E, Stsepetova J, Smidt I, Mikelsaar M. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients–a randomized double-blind placebo-controlled pilot study. Nutrition journal. 2013;12:138. doi: 10.1186/1475-2891-12-138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Smith JD, Hou T, Ludwig DS, Rimm EB, Willett W, Hu FB, Mozaffarian D. Changes in intake of protein foods, carbohydrate amount and quality, and long-term weight change: results from 3 prospective cohorts. The American journal of clinical nutrition. 2015;101:1216–24. doi: 10.3945/ajcn.114.100867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Wang H, Troy LM, Rogers GT, Fox CS, McKeown NM, Meigs JB, Jacques PF. Longitudinal association between dairy consumption and changes of body weight and waist circumference: the Framingham Heart Study. International journal of obesity (2005) 2014;38:299–305. doi: 10.1038/ijo.2013.78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Vermeer C. Vitamin K: the effect on health beyond coagulation - an overview. Food & nutrition research. 2012;56 doi: 10.3402/fnr.v56i0.5329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Walther B, Karl JP, Booth SL, Boyaval P. Menaquinones, bacteria, and the food supply: the relevance of dairy and fermented food products to vitamin K requirements. Advances in nutrition (Bethesda, Md) 2013;4:463–73. doi: 10.3945/an.113.003855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Kanazawa I. Osteocalcin as a hormone regulating glucose metabolism. World journal of diabetes. 2015;6:1345–54. doi: 10.4239/wjd.v6.i18.1345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Choi HJ, Yu J, Choi H, An JH, Kim SW, Park KS, Jang HC, Kim SY, Shin CS. Vitamin K2 supplementation improves insulin sensitivity via osteocalcin metabolism: a placebo-controlled trial. Diabetes care. 2011;34:e147. doi: 10.2337/dc11-0551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Rasekhi H, Karandish M, Jalali MT, Mohammad-Shahi M, Zarei M, Saki A, Shahbazian H. The effect of vitamin K1 supplementation on sensitivity and insulin resistance via osteocalcin in prediabetic women: a double-blind randomized controlled clinical trial. European journal of clinical nutrition. 2015;69:891–5. doi: 10.1038/ejcn.2015.17. [DOI] [PubMed] [Google Scholar]
- 209.Kumar R, Binkley N, Vella A. Effect of phylloquinone supplementation on glucose homeostasis in humans. The American journal of clinical nutrition. 2010;92:1528–32. doi: 10.3945/ajcn.2010.30108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Manna P, Kalita J. Beneficial role of vitamin K supplementation on insulin sensitivity, glucose metabolism, and the reduced risk of type 2 diabetes: A review. Nutrition (Burbank, Los Angeles County, Calif) 2016;32:732–9. doi: 10.1016/j.nut.2016.01.011. [DOI] [PubMed] [Google Scholar]
- 211.Gundberg CM, Lian JB, Booth SL. Vitamin K-dependent carboxylation of osteocalcin: friend or foe? Advances in nutrition (Bethesda, Md) 2012;3:149–57. doi: 10.3945/an.112.001834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Scheiber D, Veulemans V, Horn P, Chatrou ML, Potthoff SA, Kelm M, Schurgers LJ, Westenfeld R. High-Dose Menaquinone-7 Supplementation Reduces Cardiovascular Calcification in a Murine Model of Extraosseous Calcification. Nutrients. 2015;7:6991–7011. doi: 10.3390/nu7085318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Schurgers LJ, Spronk HM, Soute BA, Schiffers PM, DeMey JG, Vermeer C. Regression of warfarin-induced medial elastocalcinosis by high intake of vitamin K in rats. Blood. 2007;109:2823–31. doi: 10.1182/blood-2006-07-035345. [DOI] [PubMed] [Google Scholar]
- 214.Spronk HM, Soute BA, Schurgers LJ, Thijssen HH, De Mey JG, Vermeer C. Tissue-specific utilization of menaquinone-4 results in the prevention of arterial calcification in warfarin-treated rats. Journal of vascular research. 2003;40:531–7. doi: 10.1159/000075344. [DOI] [PubMed] [Google Scholar]
- 215.Schurgers LJ, Vermeer C. Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochimica et biophysica acta. 2002;1570:27–32. doi: 10.1016/s0304-4165(02)00147-2. [DOI] [PubMed] [Google Scholar]
- 216.Schurgers LJ, Teunissen KJ, Hamulyak K, Knapen MH, Vik H, Vermeer C. Vitamin K-containing dietary supplements: comparison of synthetic vitamin K1 and natto-derived menaquinone-7. Blood. 2007;109:3279–83. doi: 10.1182/blood-2006-08-040709. [DOI] [PubMed] [Google Scholar]
- 217.Beulens JW, Bots ML, Atsma F, Bartelink ML, Prokop M, Geleijnse JM, Witteman JC, Grobbee DE, van der Schouw YT. High dietary menaquinone intake is associated with reduced coronary calcification. Atherosclerosis. 2009;203:489–93. doi: 10.1016/j.atherosclerosis.2008.07.010. [DOI] [PubMed] [Google Scholar]
- 218.Geleijnse JM, Vermeer C, Grobbee DE, Schurgers LJ, Knapen MH, van der Meer IM, Hofman A, Witteman JC. Dietary intake of menaquinone is associated with a reduced risk of coronary heart disease: the Rotterdam Study. The Journal of nutrition. 2004;134:3100–5. doi: 10.1093/jn/134.11.3100. [DOI] [PubMed] [Google Scholar]
- 219.Gast GC, de Roos NM, Sluijs I, Bots ML, Beulens JW, Geleijnse JM, Witteman JC, Grobbee DE, Peeters PH, van der Schouw YT. A high menaquinone intake reduces the incidence of coronary heart disease. Nutrition, metabolism, and cardiovascular diseases: NMCD. 2009;19:504–10. doi: 10.1016/j.numecd.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 220.Chen M, Pan A, Malik VS, Hu FB. Effects of dairy intake on body weight and fat: a meta-analysis of randomized controlled trials. Am J Clin Nutr. 2012;96:735–47. doi: 10.3945/ajcn.112.037119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Abargouei AS, Janghorbani M, Salehi-Marzijarani M, Esmaillzadeh A. Effect of dairy consumption on weight and body composition in adults: a systematic review and meta-analysis of randomized controlled clinical trials. Int J Obes (Lond) 2012;36:1485–93. doi: 10.1038/ijo.2011.269. [DOI] [PubMed] [Google Scholar]
- 222.Scharf RJ, Demmer RT, DeBoer MD. Longitudinal evaluation of milk type consumed and weight status in preschoolers. Arch Dis Child. 2013;98:335–40. doi: 10.1136/archdischild-2012-302941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Noel SE, Ness AR, Northstone K, Emmett P, Newby PK. Associations between flavored milk consumption and changes in weight and body composition over time: differences among normal and overweight children. Eur J Clin Nutr. 2013;67:295–300. doi: 10.1038/ejcn.2012.123. [DOI] [PubMed] [Google Scholar]
- 224.Huh SY, Rifas-Shiman SL, Rich-Edwards JW, Taveras EM, Gillman MW. Prospective association between milk intake and adiposity in preschool-aged children. J Am Diet Assoc. 2010;110:563–70. doi: 10.1016/j.jada.2009.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Berkey CS, Rockett HR, Willett WC, Colditz GA. Milk, dairy fat, dietary calcium, and weight gain: a longitudinal study of adolescents. Arch Pediatr Adolesc Med. 2005;159:543–50. doi: 10.1001/archpedi.159.6.543. [DOI] [PubMed] [Google Scholar]
- 226.Noel SE, Ness AR, Northstone K, Emmett P, Newby PK. Milk intakes are not associated with percent body fat in children from ages 10 to 13 years. The Journal of nutrition. 2011;141:2035–41. doi: 10.3945/jn.111.143420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Niinikoski H, Viikari J, Ronnemaa T, Helenius H, Jokinen E, Lapinleimu H, Routi T, Lagstrom H, Seppanen R, Valimaki I, Simell O. Regulation of growth of 7- to 36-month-old children by energy and fat intake in the prospective, randomized STRIP baby trial. Pediatrics. 1997;100:810–6. doi: 10.1542/peds.100.5.810. [DOI] [PubMed] [Google Scholar]
- 228.Nupponen M, Pahkala K, Juonala M, Magnussen CG, Niinikoski H, Ronnemaa T, Viikari JS, Saarinen M, Lagstrom H, Jula A, Simell O, Raitakari OT. Metabolic syndrome from adolescence to early adulthood: effect of infancy-onset dietary counseling of low saturated fat: the Special Turku Coronary Risk Factor Intervention Project (STRIP) Circulation. 2015;131:605–13. doi: 10.1161/CIRCULATIONAHA.114.010532. [DOI] [PubMed] [Google Scholar]
- 229.Soedamah-Muthu SS, Verberne LD, Ding EL, Engberink MF, Geleijnse JM. Dairy consumption and incidence of hypertension: a dose-response meta-analysis of prospective cohort studies. Hypertension. 2012;60:1131–7. doi: 10.1161/HYPERTENSIONAHA.112.195206. [DOI] [PubMed] [Google Scholar]
- 230.Hu D, Huang J, Wang Y, Zhang D, Qu Y. Dairy foods and risk of stroke: a meta-analysis of prospective cohort studies. Nutr Metab Cardiovasc Dis. 2014;24:460–9. doi: 10.1016/j.numecd.2013.12.006. [DOI] [PubMed] [Google Scholar]
- 231.Qin LQ, Xu JY, Han SF, Zhang ZL, Zhao YY, Szeto IM. Dairy consumption and risk of cardiovascular disease: an updated meta-analysis of prospective cohort studies. Asia Pac J Clin Nutr. 2015;24:90–100. doi: 10.6133/apjcn.2015.24.1.09. [DOI] [PubMed] [Google Scholar]
- 232.Gao D, Ning N, Wang C, Wang Y, Li Q, Meng Z, Liu Y. Dairy products consumption and risk of type 2 diabetes: systematic review and dose-response meta-analysis. PLoS One. 2013;8:e73965. doi: 10.1371/journal.pone.0073965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Aune D, Norat T, Romundstad P, Vatten LJ. Dairy products and the risk of type 2 diabetes: a systematic review and dose-response meta-analysis of cohort studies. Am J Clin Nutr. 2013;98:1066–83. doi: 10.3945/ajcn.113.059030. [DOI] [PubMed] [Google Scholar]
- 234.Sluijs I, Forouhi NG, Beulens JW, van der Schouw YT, Agnoli C, Arriola L, Balkau B, Barricarte A, Boeing H, Bueno-de-Mesquita HB, Clavel-Chapelon F, Crowe FL, de Lauzon-Guillain B, Drogan D, Franks PW, Gavrila D, Gonzalez C, Halkjaer J, Kaaks R, Moskal A, Nilsson P, Overvad K, Palli D, Panico S, Quiros JR, Ricceri F, Rinaldi S, Rolandsson O, Sacerdote C, Sanchez MJ, Slimani N, Spijkerman AM, Teucher B, Tjonneland A, Tormo MJ, Tumino R, van der AD, Sharp SJ, Langenberg C, Feskens EJ, Riboli E, Wareham NJ, InterAct C. The amount and type of dairy product intake and incident type 2 diabetes: results from the EPIC-InterAct Study. Am J Clin Nutr. 2012;96:382–90. doi: 10.3945/ajcn.111.021907. [DOI] [PubMed] [Google Scholar]
- 235.Chen M, Sun Q, Giovannucci E, Mozaffarian D, Manson JE, Willett WC, Hu FB. Dairy consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. BMC Med. 2014;12:215. doi: 10.1186/s12916-014-0215-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Wang H, Troy LM, Rogers GT, Fox CS, McKeown NM, Meigs JB, Jacques PF. Longitudinal association between dairy consumption and changes of body weight and waist circumference: the Framingham Heart Study. Int J Obes (Lond) 2014;38:299–305. doi: 10.1038/ijo.2013.78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Smith JD, Hou T, Ludwig DS, Rimm EB, Willett WC, Hu FB, Mozaffarian D. Changes in intake of protein foods, carbohydrate amount and quality, and long-term weight change: results from 3 prospective cohorts. Am J Clin Nutr. 2015;101:1216–24. doi: 10.3945/ajcn.114.100867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Mensink RP, Zock PL, Kester AD, Katan MB. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: a meta-analysis of 60 controlled trials. The American journal of clinical nutrition. 2003;77:1146–55. doi: 10.1093/ajcn/77.5.1146. [DOI] [PubMed] [Google Scholar]
- 239.Thorning TK, Bertram HC, Bonjour JP, de Groot L, Dupont D, Feeney E, Ipsen R, Lecerf JM, Mackie A, McKinley MC, Michalski MC, Remond D, Riserus U, Soedamah-Muthu SS, Tholstrup T, Weaver C, Astrup A, Givens I. Whole dairy matrix or single nutrients in assessment of health effects: current evidence and knowledge gaps. The American journal of clinical nutrition. 2017;105:1033–1045. doi: 10.3945/ajcn.116.151548. [DOI] [PubMed] [Google Scholar]
- 240.Mozaffarian D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: a comprehensive review. Circulation. 2016;133:187–225. doi: 10.1161/CIRCULATIONAHA.115.018585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Ericson U, Hellstrand S, Brunkwall L, Schulz CA, Sonestedt E, Wallstrom P, Gullberg B, Wirfalt E, Orho-Melander M. Food sources of fat may clarify the inconsistent role of dietary fat intake for incidence of type 2 diabetes. Am J Clin Nutr. 2015;101:1065–80. doi: 10.3945/ajcn.114.103010. [DOI] [PubMed] [Google Scholar]
- 242.Diaz-Lopez A, Bullo M, Martinez-Gonzalez MA, Corella D, Estruch R, Fito M, Gomez-Gracia E, Fiol M, Garcia de la Corte FJ, Ros E, Babio N, Serra-Majem L, Pinto X, Munoz MA, Frances F, Buil-Cosiales P, Salas-Salvado J. Dairy product consumption and risk of type 2 diabetes in an elderly Spanish Mediterranean population at high cardiovascular risk. Eur J Nutr. 2015 doi: 10.1007/s00394-015-0855-8. Epub Feb 7. [DOI] [PubMed] [Google Scholar]
- 243.Nestel PJ, Mellett N, Pally S, Wong G, Barlow CK, Croft K, Mori TA, Meikle PJ. Effects of low-fat or full-fat fermented and non-fermented dairy foods on selected cardiovascular biomarkers in overweight adults. Br J Nutr. 2013;110:2242–9. doi: 10.1017/S0007114513001621. [DOI] [PubMed] [Google Scholar]
- 244.Mozaffarian D, Cao H, King IB, Lemaitre RN, Song X, Siscovick DS, Hotamisligil GS. Trans-palmitoleic Acid, metabolic risk factors, and new-onset diabetes in U.S. adults: a cohort study. Ann Intern Med. 2010;153:790–9. doi: 10.1059/0003-4819-153-12-201012210-00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Mozaffarian D, de Oliveira Otto MC, Lemaitre RN, Fretts AM, Hotamisligil G, Tsai MY, Siscovick DS, Nettleton JA. trans-Palmitoleic acid, other dairy fat biomarkers, and incident diabetes: the Multi-Ethnic Study of Atherosclerosis (MESA) Am J Clin Nutr. 2013;97:854–61. doi: 10.3945/ajcn.112.045468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Kratz M, Baars T, Guyenet S. The relationship between high-fat dairy consumption and obesity, cardiovascular, and metabolic disease. Eur J Nutr. 2013;52:1–24. doi: 10.1007/s00394-012-0418-1. [DOI] [PubMed] [Google Scholar]
- 247.Kratz M, Marcovina S, Nelson JE, Yeh MM, Kowdley KV, Callahan HS, Song X, Di C, Utzschneider KM. Dairy fat intake is associated with glucose tolerance, hepatic and systemic insulin sensitivity, and liver fat but not beta-cell function in humans. Am J Clin Nutr. 2014;99:1385–1396. doi: 10.3945/ajcn.113.075457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Forouhi NG, Koulman A, Sharp SJ, Imamura F, Kroger J, Schulze MB, Crowe FL, Huerta JM, Guevara M, Beulens JW, van Woudenbergh GJ, Wang L, Summerhill K, Griffin JL, Feskens EJ, Amiano P, Boeing H, Clavel-Chapelon F, Dartois L, Fagherazzi G, Franks PW, Gonzalez C, Jakobsen MU, Kaaks R, Key TJ, Khaw KT, Kuhn T, Mattiello A, Nilsson PM, Overvad K, Pala V, Palli D, Quiros JR, Rolandsson O, Roswall N, Sacerdote C, Sanchez MJ, Slimani N, Spijkerman AM, Tjonneland A, Tormo MJ, Tumino R, van der AD, van der Schouw YT, Langenberg C, Riboli E, Wareham NJ. Differences in the prospective association between individual plasma phospholipid saturated fatty acids and incident type 2 diabetes: the EPIC-InterAct case-cohort study. Lancet Diabetes Endocrinol. 2014;2:810–8. doi: 10.1016/S2213-8587(14)70146-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Khaw KT, Friesen MD, Riboli E, Luben R, Wareham N. Plasma phospholipid fatty acid concentration and incident coronary heart disease in men and women: the EPIC-Norfolk prospective study. PLoS Med. 2012;9:e1001255. doi: 10.1371/journal.pmed.1001255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.de Oliveira Otto MC, Nettleton JA, Lemaitre RN, Steffen LM, Kromhout D, Rich SS, Tsai MY, Jacobs DR, Mozaffarian D. Biomarkers of dairy fatty acids and risk of cardiovascular disease in the Multi-ethnic Study of Atherosclerosis. J Am Heart Assoc. 2013;2:e000092. doi: 10.1161/JAHA.113.000092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Chowdhury R, Warnakula S, Kunutsor S, Crowe F, Ward HA, Johnson L, Franco OH, Butterworth AS, Forouhi NG, Thompson SG, Khaw KT, Mozaffarian D, Danesh J, Di Angelantonio E. Association of dietary, circulating, and supplement fatty acids with coronary risk: a systematic review and meta-analysis. Ann Intern Med. 2014;160:398–406. doi: 10.7326/M13-1788. [DOI] [PubMed] [Google Scholar]
- 252.Yakoob MY, Shi P, Hu FB, Campos H, Rexrode KM, Orav EJ, Willett WC, Mozaffarian D. Circulating biomarkers of dairy fat and risk of incident stroke in U.S. men and women in 2 large prospective cohorts. Am J Clin Nutr. 2014;100:1437–47. doi: 10.3945/ajcn.114.083097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Mozaffarian D. Natural trans fat, dairy fat, partially hydrogenated oils, and cardiometabolic health – the Ludwigshafen Risk and Cardiovascular Health Study. Eur Heart J. 2015 doi: 10.1093/eurheartj/ehv595. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Kawser Hossain M, Abdal Dayem A, Han J, Yin Y, Kim K, Kumar Saha S, Yang GM, Choi HY, Cho SG. Molecular Mechanisms of the Anti-Obesity and Anti-Diabetic Properties of Flavonoids. International journal of molecular sciences. 2016;17:569. doi: 10.3390/ijms17040569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Manach C, Williamson G, Morand C, Scalbert A, Remesy C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. The American journal of clinical nutrition. 2005;81:230S–242S. doi: 10.1093/ajcn/81.1.230S. [DOI] [PubMed] [Google Scholar]