

Abstract
Research on the neurobiology of the kynurenine pathway has suffered years of relative obscurity because tryptophan degradation, and its involvement in both physiology and major brain diseases, was viewed almost exclusively through the lens of the well-established metabolite serotonin. With increasing recognition that kynurenine and its metabolites can affect and even control a variety of classic neurotransmitter systems directly and indirectly, interest is expanding rapidly. Moreover, kynurenine pathway metabolism itself is modulated in conditions such as infection and stress, which are known to induce major changes in well-being and behaviour, so that kynurenines may be instrumental in the etiology of psychiatric and neurological disorders. It is therefore likely that the near future will not only witness the discovery of additional physiological and pathological roles for brain kynurenines, but also ever-increasing interest in drug development based on these roles. In particular, targeting the kynurenine pathway with new specific agents may make it possible to prevent disease by appropriate pharmacological or genetic manipulations.
The following overview focuses on areas of kynurenine research which are either controversial, of major potential therapeutic interest, or just beginning to receive the degree of attention which will clarify their relevance to neurobiology and medicine. It also highlights technical issues so that investigators entering the field, and new research initiatives, are not misdirected by inappropriate experimental approaches or incorrect interpretations at this time of skyrocketing interest in the subject matter.
Keywords: 3-Hydroxykynurenine, Kynurenic acid, Neuroimmunology, Neurology, Psychiatry, Quinolinic acid
The early days
The kynurenine pathway, which is initiated by the oxidative metabolism of tryptophan, has been studied in some detail since the early part of the twentieth century. It was originally viewed primarily as the synthetic route to nicotinamide and hence nicotinamide adenine dinucleotide (NAD+), a crucial co-factor for a wide range of enzymes. The kynurenine pathway formed an intriguing link between the dietary availability of vitamin B6 and tryptophan metabolism (see Badawy, 2016) since many of the major enzymes in the cascade were B6-dependent, and B6 deficiency was usefully characterised by the increased excretion of xanthurenic acid in the urine (Bender et al., 1990). No specific neurobiological activity was demonstrated for kynurenine and its catabolic products until (Lapin, 1978) showed that several tryptophan metabolites could produce convulsions when injected directly into the brain. A number of publications had earlier reported abnormal concentrations of kynurenine in various brain disorders, although those results were interpreted in terms of their effects on tryptophan and serotonin levels (Curzon, 1969; Dewhurst, 1970; Wood et al., 1978).
By the mid-1970s, reports began to appear in support of the notion that glutamate, a simple amino acid, was likely to be an excitatory neurotransmitter in the central nervous system (CNS) (Stone, 1973, 1976). Furthermore, the use of selective antagonists to block synaptic excitation suggested the existence of more than one subtype of glutamate receptors (Davies et al., 1979; McLennan, 1983). It was during a screen of several dozen newly synthesised compounds that agonist activity was detected in one of them. This compound, pyridine-2,3-dicarboxylic acid, also named quinolinic acid and long known as a major component of the kynurenine pathway, produced an increase in neuronal firing rate in the rat cerebral cortex (Perkins and Stone, 1983b; Stone and Perkins, 1981). Neuronal excitation was blocked by 2-amino-5-phosphonopentanoic acid (AP5), a compound reported earlier the same year to block those glutamate receptors that were selectively sensitive to N-methyl-D-aspartate (NMDA) (Davies et al., 1981; Perkins et al., 1982; Perkins et al., 1981). Thus, quinolinic acid was identified as an endogenous, selective agonist at NMDA receptors (Stone and Perkins, 1981).
NMDA was already known to produce axon-sparing, “excitotoxic” loss of neurons in the CNS (Schwarcz et al., 1978), leading to the suggestion that an overexcitation of NMDA receptors might play a key role in the aetiology of neurodegenerative disorders such as Alzheimer’s disease or Huntington’s disease (HD), as well as epilepsy and stroke (Schwarcz and Meldrum, 1985). In line with its ability to act as a NMDA receptor agonist, quinolinic acid was also found to cause excitotoxic neuronal lesions, which could be prevented by the selective NMDA receptor antagonist 2-amino-7-phosphonoheptanoic acid (AP7) (Foster et al., 1983). As a result, the focus of attention for understanding the biological roles of quinolinic acid in the CNS became centered on disorders associated with neuronal damage and degeneration (Lee et al., 2016; Lovelace et al., 2016).
By testing other compounds in the kynurenine pathway, a second metabolite, kynurenic acid, was later characterised as a glutamate receptor antagonist, blocking the actions of glutamate, NMDA and quinolinic acid, as well as related agonists thought to define other glutamate receptor subtypes, namely quisqualic acid [acting at what are now known as α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors] and kainic acid (Perkins and Stone, 1982; Stone and Darlington, 2002). In line with the demonstration that NMDA receptor antagonists prevent excitotoxicity and seizures (Croucher et al., 1982; Schwarcz et al., 1982), kynurenic acid was then soon shown to possess pronounced neuroprotective and anticonvulsant effects (Foster et al., 1984).
Around the same time, interest developed in the biological function of one of the initial enzymes in the kynurenine pathway, indoleamine-2,3-dioxygenase (IDO), which converts tryptophan to kynurenine. Pfefferkorn (Pfefferkorn, 1984) observed that IDO was activated by interferon-ɣ during the immune response to infection, inhibiting the detrimental effects of the infectious agent. This insight triggered a long-running debate on whether the mechanism of the anti-infective effect was caused by the depletion of tryptophan or by the accumulation of kynurenine and its downstream metabolites (Badawy et al., 2016).
Mostly as a result of these early reports, interest in the kynurenine pathway has grown progressively and at an increasing rate, with over 15,000 articles published to date. The kynurenine field now encompasses a wide range of studies in physiology, pharmacology and immunology, with potential clinical relevance in areas as seemingly disparate as neurological and psychiatric diseases, diabetes and cancer (Strasser et al., 2016). In this review, we shall limit ourselves to highlighting a few selected areas of investigation in which there exist important yet largely unresolved questions with regard to basic biology and implications for brain pathology. Subsequent papers in this issue of “Neuropharmacology” address individual topics in more detail to provide a flavour of the growing depth and breadth of interest in the kynurenine pathway.
Kynurenine pathway metabolism: some unsettled basic issues
One important problem facing neurobiologists is that the traditionally accepted scheme of tryptophan metabolism via kynurenine (Figure 1) does not always agree with experimental findings, raising fundamental questions with regard to the origin and disposition of components of the pathway. For example, the concentration of quinolinic acid in the brain does not decline as predicted after pharmacological inhibition or genomic elimination of kynurenine 3-monooxygenase (KMO). Thus, despite large increases in the brain levels of kynurenic acid (10-100-fold), indicating a shift of pathway metabolism towards the formation of the neuroprotective agent, administration of a selective KMO inhibitor does not affect brain quinolinic acid levels at all (Clark et al., 2005), and even the complete genomic deletion of KMO reduces brain quinolinic acid levels only by around 20% (Giorgini et al., 2013). At the same time, brain levels of anthranilic acid and picolinic acid, two pathway metabolites which have received relatively little attention so far, increase several-fold in these experimental situations.
Figure 1.
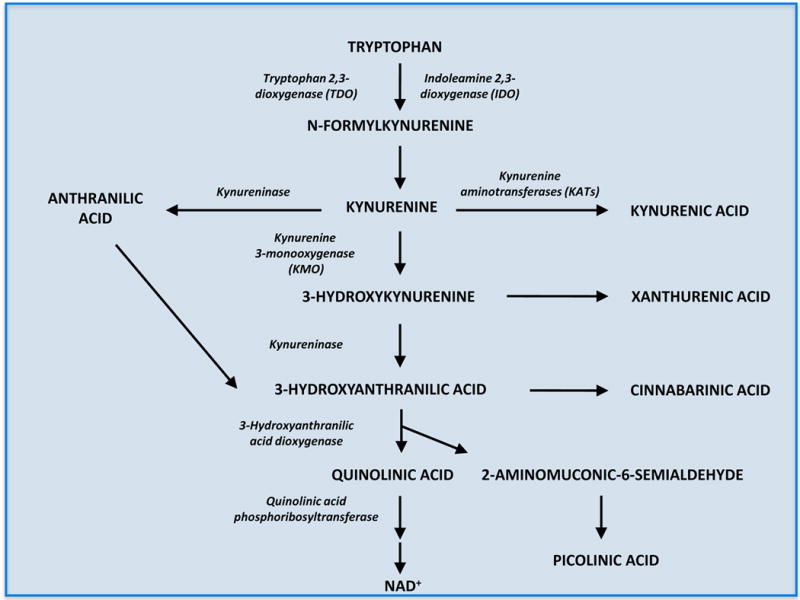
The kynurenine pathway of tryptophan degradation
Two conditions in which these phenomena may be pathophysiologically significant are cerebral malaria and trypanosomiasis. Mice treated with strains of the malaria parasite (Plasmodium), which induce a cellular and cerebral pathology closely resembling that seen in humans, develop progressive cerebral inflammation and behavioural impairments, and die within 7 days (Clark et al., 2005). When treated with the KMO inhibitor Ro 61-8048, however, the animals, which show 100-fold increases in the levels of kynurenic acid and anthranilic acid in the brain, survive for at least 21 days. This raises the possibility that not only kynurenic acid but also anthranilic acid may have beneficial effects in pathological situations that are associated with an inflammatory state. Perhaps it is no coincidence that the molecular structures of many non-steroidal anti-inflammatory and analgesic drugs currently available are built around the anthranilate nucleus (Mohamed Eissa et al., 2012; Sharma et al., 2002).
Many questions remain. If kynurenines are key players in the inflammatory process, are they drivers of functional changes or merely biologically inert bystanders? Are changes in the levels of the metabolites a direct consequence of immune regulations of individual kynurenine pathway enzymes or secondary reactions to entirely different stimuli? What are the differences between acute and chronic inflammation, given that chronic conditions are often associated with compensatory metabolic adjustments?
Another issue of fundamental relevance to neuroscientists pertains to the cellular localization of kynurenine pathway enzymes and their products in the mammalian brain. With exceptions (Guillemin et al., 2007), most studies indicate that brain kynurenines are synthesized or degraded in glial cells rather than in neurons under both normal and pathological conditions. Thus, neuronal lesions, which are typically accompanied by astrogliosis and microglial proliferation, are mostly associated with elevated enzyme activities and higher, rather than lower, levels of pathway metabolites (Ceresoli et al., 1997; Guidetti et al., 1995). The generally accepted view holds that the main branch of the cascade, leading to 3-hydroxykynurenine, 3-hydroxyanthranilic acid, quinolinic acid and beyond, is normally contained in microglial cells whereas kynurenic acid is formed in astrocytes in a second, competing branch of the pathway (Figure 1). This conclusion is largely based on in vitro studies using individual cell types in culture (Chen et al., 2011; Chiarugi et al., 2001; Heyes et al., 1997; Kiss et al., 2003; Kocki et al., 2002) and, unfortunately to a far lesser extent, immunocytochemical analyses with antibodies directed against purified enzymes or against metabolites themselves (Guidetti et al., 2007; Lehrmann et al., 2001). Specifically, in the absence of anti-KMO antibodies that can be used for unambiguous microscopic localization of the enzyme, evidence for a microglial localization of this pivotal pathway enzyme rests mostly on ex vivo studies (Guillemin et al., 2005; Parrott and O’Connor, 2015). Notably, however, though the concept of a physical segregation of the two arms of the pathway in the brain is intuitively attractive, the dynamics of cerebral kynurenine pathway metabolism change substantially under abnormal conditions (Hoshi et al., 2009; Parrott and O’Connor, 2015; Saito et al., 1993; Wu et al., 2013). It follows that the cellular localisation of individual pathway enzymes, as well as the functional relationship between the two pathway branches, in the brain ought to be carefully re-assessed and interpreted in future studies.
A third major challenge for neuroscientists is related to the fact that the influence of peripheral kynurenines on brain function is still not fully understood. Although circulating tryptophan as well as some pathway metabolites (including the pivotal kynurenine and 3-hydroxykynurenine) can enter the brain quite easily under physiological conditions (Fukui et al., 1991; Pardridge, 1998), important open questions abound. For example, since most challenge studies so far have only examined the central effects of acute fluctuations in plasma kynurenines, far too little is known about possible qualitative and quantitative adaptations of the blood-brain barrier to persistent up- or downregulation of kynurenine pathway metabolism in the periphery. What about the effects of pathological changes in blood-brain barrier permeability, causing abnormalities in endothelial cells and capillary function in specific parts of the brain (Owe-Young et al., 2008)? With regard to possible neuropharmacological applications, can targeted manipulations of the large neutral amino acid transporter, which normally mediates the brain uptake of blood-derived kynurenines (Pardridge, 1983, 1998), be used to influence cerebral pathway metabolism and function?
Molecular sites of action
Over the last decade, a plethora of newly identified molecular sites and receptors have been suggested to mediate the biological effects of the various metabolites of the kynurenine pathway. These novel molecular targets may exist on neurons or various non-neuronal brain cells, or on immune cells in the periphery, presenting significant impediments for the unambiguous interpretation of data involving pharmacological or genetic manipulations of the pathway.
While quinolinic acid is certainly an agonist at NMDA receptors, various subtypes of NMDA receptors and subunit combinations are differentially sensitive to the metabolite. In particular, quinolinic acid is relatively selective for NMDA receptors in the neocortex and hippocampus, with much weaker activity in the cerebellum and spinal cord (Perkins and Stone, 1983a, b), correlating with the weakness of quinolinate’s activity at NR2C subunits (de Carvalho et al., 1996; Monaghan and Beaton, 1991) and the low expression of NR2C and NR2B subunits in the cerebellum (Scherzer et al., 1997; Wang et al., 1995). Quinolinic acid can also generate reactive oxygen species, which may contribute to neuronal damage under pathological conditions (Santamaria et al., 2001).
In addition, it is becoming clear that quinolinic acid contributes to many areas of neurobiology and related systems. Many of these effects, which have been admirably reviewed elsewhere (Guillemin, 2012), involve important interactions with the immune system. Pro-inflammatory mediators such as TNFα can promote quinolinic acid production (Guillemin et al., 2001) and interleukin-1β potentiates quinolinate excitotoxicity (Stone and Behan, 2007) while suppressors of inflammation such as interleukin-4 reduces quinolinate neosynthesis by inhibiting indoleamine-2,3-dioxygenase (IDO) activity upstream (Musso et al., 1994). Interestingly, a number of indole-derived compounds and structurally related agents can suppress quinolinic acid production by inhibiting tryptophan-2,3-dioxygenase (TDO; Botting, 1995).
Although the direct effects of quinolinic acid are mediated primarily by NMDA receptors, quinolinic acid can also promote the release and inhibit the re-uptake of glutamate (Tavares et al., 2002), potentially facilitating the activation of kainate and AMPA receptors as well as metabotropic glutamate receptors. Furthermore, quinolinic acid has some intracellular actions, which are probably independent of membrane receptors. For example, quinolinic acid increases the phosphorylation of several proteins, including tau filament proteins which implicated in Alzheimer’s disease (Rahman et al., 2009; Pierozan et al., 2010).
Kynurenic acid is well established as an antagonist at all ionotropic glutamate receptor subtypes, with greatest potency blocking the obligatory glycine-B co-agonist site on NMDA receptors (Birch et al., 1988a, b). An argument against the physiological relevance of this action is that inhibition is only seen at concentrations that are substantially higher than those normally found endogenously in the CNS. However, upon efflux (whether active or passive) from neurons or glia, concentrations in the immediately surrounding tissue could easily reach levels similar to, or greater than, those known to be pharmacologically active and sufficient to directly affect NMDA receptor-mediated synaptic transmission. By the time kynurenic acid has diffused away from its sites of production, diluted by several orders of magnitude in the extracellular fluid and eventually the cerebrospinal fluid (CSF), its levels will have fallen substantially. Extracellular concentrations of kynurenic acid will also be reduced by active removal from the brain (Moroni et al., 1988) and cellular re-uptake (Uwai et al., 2012). Together, these factors make it difficult to rely on measurements of absolute concentrations of kynurenic acid and other pathway metabolites in tissue, microdialysate, CSF or plasma when attempting to assess target engagement in the context of physiological effects, pathological symptoms, and treatment.
Understanding the mechanism of action of kynurenic acid is especially difficult since the compound appears to have several biological targets in addition to glutamate receptors. One argument against a preferential role of NMDA receptor inhibition is that 7-chlorokynurenic acid, which blocks the glycine-B co-agonist site with approximately 20-fold higher affinity than kynurenic acid itself (Leeson et al., 1991), does not duplicate the actions of kynurenic acid in several experimental paradigms in vitro and in vivo (Beggiato et al., 2013; Beggiato et al., 2014; Hilmas et al., 2001). On the other hand, 7-chlorokynurenic acid itself may not act as a selective glycine-B antagonist in vivo (Lehmann et al., 1993) and even potentiates NMDA receptor activation in human embryonic kidney cells (Smothers and Woodward, 2007). Differential affinity of 7-chlorokynurenic acid at glycine binding sites of various NMDA receptor subtypes (Nilsson et al., 2007; O’Shea et al., 1991), possible brain region-specific effects (Linderholm et al., 2007), and the ability of kynurenic acid to modulate extracellular glutamate levels in the brain (see Schwarcz, 2016, for review), further complicate interpretation.
Another potential target is the α7 homomeric nicotinic receptor (Hilmas et al., 2001) though this site, too, has not yet been unequivocally validated. Thus, several independent studies have failed to reproduce the original observations (see Albuquerque and Schwarcz, 2013, for discussion; Arnaiz-Cot et al., 2008; Dobelis et al., 2012; Mok et al., 2009). Other investigators used pharmacological tools such as galantamine, an allosteric enhancer of α7 nicotinic receptor function that acts as an agonist at a site that overlaps with the site targeted by kynurenic acid (Lopes et al., 2007). But the drug also inhibits the activity of cholinesterase and other esterases at higher concentrations (Darvesh et al., 2003; Nordberg and Svensson, 1998), possibly limiting its ability to provide definitive proof for the involvement of α7 receptors in the actions of kynurenic acid. Moreover, galantamine blocks some potassium channels (Vicente et al., 2010; Vigneault et al., 2012), which may be sufficient to explain its facilitatory effect on neuronal transmission and its apparent reversal of the effects of kynurenate.
To add to these complexities, kynurenic acid can also activate the G-protein coupled receptor GPR35 (Stone et al., 2013; Wang et al., 2006). Although found primarily on immune cells and in the gastrointestinal tract, this protein does occur in the CNS where it is located on astrocytes (Moroni et al., 2012) and has been linked to nociceptive phenomena and neurotransmission in the hippocampus (Alkondon et al., 2015). Here, too, the extent to which this activity is relevant physiologically or under pathological conditions remains unclear, requiring further in-depth evaluation. Finally, kynurenic acid is an agonist at the aryl hydrocarbon receptor (AHR) (DiNatale et al., 2010), a xenobiotic-sensing receptor that promotes the metabolism of environmental toxins and functions in immune-regulation (Stephens et al., 2013). This site is present on tumour cells and cells of the immune system, sparking considerable interest in the role of kynurenines in the initiation and development of cancers (Bessede et al., 2014; Opitz et al., 2011). Implications for neurobiology, including neuropharmacology, are not unlikely but have not been explored so far.
One additional aspect that makes the kynurenine pathway increasingly interesting is that kynurenine itself, as well as its metabolites 3-hydroxykynurenine and 3-hydroxyanthranilic acid, are biologically active. Like kynurenic acid, kynurenine is an AHR agonist with unknown downstream effects (Opitz et al., 2011). 3-hydroxykynurenine and 3-hydroxyanthranilic acid are both redox active and may function to regulate and maintain the oxidative status of tissues in order to prevent cell damage by reactive oxygen species (Chobot et al., 2015; Gonzalez-Esquivel et al., 2016; Shibata et al., 2014). In addition, 3-hydroxyanthranilic acid also has distinct actions within the immune system. Target molecules which may function as 3-hydroxyanthranilic acid receptors have been demonstrated on dendritic cells and a range of T cells including Th1 cells and FoxP3+ Treg cells (Xie et al., 2015). While the precise nature of the effects mediated through these sites is still unclear, the overall effect of 3-hydroxyanthranilic acid seems to be anti-inflammatory (Krause et al., 2011). This concept is consistent with studies showing profound changes in 3-hydroxyanthranilic acid concentrations in inflammatory disorders and their normalisation on treatment (Forrest et al., 2006), especially when considered in terms of the ratio with anthranilic acid (Darlington et al., 2010).
Clarification of the differential roles and relevance of these various target sites is clearly a pressing problem and should be pursued with high urgency. Ideally, newly produced selective agonists and antagonists can soon be used as tools to achieve this objective in the laboratory. These experimental agents might then also constitute lead compounds for the development of new drugs which might be suitable in a range of clinical conditions involving kynurenine pathway metabolites.
Technical and methodological considerations
In studies of the role of kynurenines in the CNS, the majority of useful information has been gleaned from work on in vivo models or acutely prepared tissue slices, though studies with freshly dissociated cells (neurons, astrocytes and microglia) have also provided relevant information (Guillemin et al., 2007; Guillemin et al., 2005). In contrast, cell culture systems have proved less valuable and are often controversial. This is a reflection of the fact that cell lines – even those that are purportedly from the same original stock and are labelled as the same cell type – are quite heterogeneous and show progressively changing characteristics with repeated passages (Nugoli et al., 2003). This problem has received most attention in the cancer field, where some of the most widely used cell lines are now known to exhibit substantially different growth patterns and molecular signatures between suppliers and research laboratories (Nugoli et al., 2003; Xie et al., 2012), but often also muddies research of CNS kynurenines. Thus, though observations of intracellular biochemical events or effector targets (receptors, ion channels) of kynurenines in neuronal and glial cultures continue to be useful, great caution must be exercised when extrapolating such data with regard to functional interactions between various cell types. The continuous biochemical dialogue between neurons and glia, and between intra- and extracellular compartments of these cells, as well as the presence in the CNS of a considerable number of small molecule regulators of kynurenine pathway metabolism (Chen et al., 2011; Guillemin et al., 2007; Guillemin et al., 2005), present far greater challenges to experimental reproducibility and interpretation than studies using dissociated cells of peripheral origin, such as hepatocytes or myocytes.
Apart from an increased attention to the role of individual cell populations, it is not difficult to predict that the development of new experimental tools, too, will contribute greatly to solving the many open questions of kynurenine neurobiology. This includes approaches involving the genes of specific pathway enzymes, such as the generation of knockout animals, the search for informative spontaneous mutations and single nucleotide polymorphisms (Aoyama et al., 2006; Holtze et al., 2012; Wonodi et al., 2014), and gene silencing methodologies (Vidal et al., 2015). Moreover, conditional knockouts or knockdown models of each of the kynurenine pathway enzymes are expected to clarify the functional dynamics of pathway metabolism in the CNS in the near future.
The value of the new information obtained from genetic observations and manipulations will be further enhanced by the use of human or small animal imaging techniques, which can be used to visualize and quantify enzyme expression or activity non-invasively in vivo. This will require the design and radioactive labelling of appropriate specific ligands, and may include agonists/activators and antagonists/inhibitors of receptors, enzymes and other proteins that are known to define and determine the functions of kynurenines in the brain. Among many other conceivable applications, successful development of these tools will make it possible to discriminate between splice variants of proteins or between enzyme isoforms – for example the four subtypes of kynurenine aminotransferase (Han et al., 2010).
The gut-brain axis
As investigators pay increasing attention to the relationship between the gastrointestinal microbiome and the brain, with thousands of bacterial species under constant flux and variation with changes in the diet and nutritional status of the host (Bauer et al., 2016; Ghaisas et al., 2016; Ojeda et al., 2016), it is important to explore the potential role of microbes in the metabolism and function of the kynurenine pathway. Humans and most other mammals do not have the ability to synthesise tryptophan, which is therefore considered an “essential” amino acid. In contrast, tryptophan is readily neosynthesised by most plants and bacteria (Barends et al., 2008; Marinoni et al., 2008; Turski et al., 2011).
Interestingly, bacteria also utilize other mechanisms which account for the formation and degradation, respectively, of neuroactive kynurenines. For example, bacteria can neo-synthesize quinolinic acid (Marinoni et al., 2008; Reichmann et al., 2015; Sakuraba et al., 2005; Saunders et al., 2008), and unique prokaryotic enzymes can degrade kynurenic acid (Taniuchi and Hayaishi, 1963). Bacterial neosynthesis of quinolinic acid begins with the condensation of iminoaspartate with dihydroxyacetone, and the latter compound also inhibits α-amino-β-carboxymuconate-ε-semialdehyde decarboxylase (ACMSD) (Garavaglia et al., 2009). Normally, ACMS cyclises non-enzymatically to quinolinic acid, but is converted to picolinic acid in the presence of ACMSD. Inhibition of ACMSD by bacterial dihydroxyacetone will therefore increase the synthesis of quinolinic acid. Another source of quinolinic acid may be anthranilic acid (first converted to 3-hydroxyanthranilic acid by anthranilate oxidase; Liu et al., 2010), and bacteria can also produce quinolinic acid non-enzymatically (Cleaves and Miller, 2001). These bacterial processes may play a part in – still ill-defined – biological roles of endogenous quinolinic acid in the mammalian brain. We predict that more detailed information on these alternative processes involving the gut microbiome will be essential to understand fully the dynamics of cerebral tryptophan metabolism and, in particular, the balance between quinolinic acid and kynurenic acid production and function in the brain. In other words, it is tempting to speculate that microbially-regulated tryptophan and kynurenines significantly influence CNS function and dysfunction in mammals (Kennedy et al., 2016; O’Mahony et al., 2015).
CNS development
The nature and possible functional role of kynurenines in the developing brain has begun to receive increased attention in recent years. In rodents, the fetal brain contains remarkably high concentrations of neuroactive kynurenines compared to the adult brain (Ceresoli-Borroni and Schwarcz, 2000), and a similar observation has been made with regard to prenatal cerebral kynurenic acid levels in non-human primates (Beal et al., 1992). As the brain content of all these metabolites falls off precipitously immediately after birth, the high fetal brain levels likely originate either from the maternal circulation or from the placenta. Still, various scenarios may account for the high prenatal levels of brain kynurenines. Possibilities include, for example, relatively unimpeded transport of kynurenine and/or 3-hydroxykynurenine from the maternal blood or the placenta to the fetal brain and, subsequently, robust production of downstream metabolites due to high activities of kynurenine pathway enzymes in the fetal brain. Alternatively or in addition, and in contrast to their very low ability to penetrate the blood-brain barrier in adulthood (Fukui et al., 1991), the polar compounds kynurenic acid and quinolinic acid, derived from the mother or the placenta, may be able to enter the fetal brain relatively freely. These fundamental principles of kynurenine neurobiology are readily testable experimentally. Detailed knowledge is overdue and will significantly impact our thinking about the possible physiological role of the high brain levels of kynurenines during the prenatal period.
The term ‘neuro-developmental diseases’ has been coined to refer to disorders that appear in late childhood or adulthood but are caused by abnormal early development of the CNS. In principle, early developmental impairments may be genetic in origin or caused by adverse external influences including bacterial or viral infections, exposure to environmental or dietary toxins, or experience of stressful situations (O’Farrell and Harkin, 2016). It is now recognised that these factors directly affect the kynurenine pathway in at least three general ways. Firstly, infections induce an immune response which raises the production of several cytokines such as interferon-γ, interleukin-1β and tumour necrosis factor-α (TNF-α). These as well as other pro-inflammatory cytokines in turn induce TDO and KMO and therefore increase pathway flux (Pemberton et al. 1997; Smith et al., 2001)
Secondly, psychological stress causes increased synthesis and secretion of adrenal glucocorticoid hormones, which in turn affect kynurenines both in the periphery and in the brain (Laugeray et al., 2010). Thirdly, some naturally occurring compounds such as the brassinins, found in a number of vegetables, can inhibit IDO and thus reduce activity along the entire kynurenine pathway (Banerjee et al., 2008; Gaspari et al., 2006).
The fundamental importance of these links between kynurenine pathway metabolism and brain development is particularly apparent when considering the influence of environmental factors on the embryonic brain. Infection by viruses, bacteria or parasites is associated with the activation of Toll-Like Receptors (TLRs), especially TLR3 and TLR4, which can be activated experimentally by agents such as bacterial lipopolysaccharide (LPS) or the viral-mimetic double-stranded RNA poly(inosinic:cytidylic) acid (poly I:C). TLR activation results in stimulation of the kynurenine pathway (Asp et al., 2010; Clarke et al., 2012; Opitz et al., 2009; Suh et al., 2007; Wang et al., 2011), so that any infection contracted during pregnancy is likely to be accompanied by increased production of kynurenines. This effect may depend on the age at which TLR activation occurs (Khalil et al., 2013). In light of the fact that changes in kynurenine metabolism affect brain development (Forrest et al., 2013a; Forrest et al., 2013b), with consequences which persist into adulthood (Forrest et al., 2013b; Notarangelo and Pocivavsek, 2016; Pisar et al., 2014), any infection during the very early stages of brain development may therefore result in CNS dysfunction, or increased susceptibility to dysfunction, later in life. For example, in rats, perinatal administration of influenza virus stimulates cerebral kynurenine pathway metabolism acutely, resulting in impaired sensorimotor function in adulthood (Asp et al., 2010). As human brain development continues for several years after birth (Ballesteros et al., 1993; Gogtay et al., 2004; Jernigan et al., 2011), spanning times when children are exposed to a range of harmful environmental influences, these studies suggest that the vulnerable period may, in fact, extend into the early years of childhood in humans. Activation of the kynurenine pathway could certainly explain the emergence of psychiatric disorders in people born to mothers exposed to a range of infectious organisms including Toxoplasma gondii,rubella, influenza and cytomegalovirus (Hunt et al., 2016; Libbey et al., 2005; Torrey et al., 2007; Torrey et al., 2012; Yolken and Torrey, 2008) or to persistent stressful events in utero and beyond.
In view of the central role of glutamatergic mechanisms in brain development (Metzger, 2010), impaired glutamate function in the brain could constitute a common molecular mechanism linking perinatal imbalance of kynurenines to subsequent vulnerability and pathology. Indeed, prenatal interventions affecting kynurenine pathway metabolism affect the expression of glutamate receptor subunits (Forrest et al., 2013a; Forrest et al., 2013b), and abnormalities of glutamate-related cognitive performance are seen after repeated early stress (Tse et al., 2011; Yuen et al., 2012). It is therefore not difficult to predict that the nature and effects of imbalanced glutamatergic neurotransmission following abnormal kynurenine pathway metabolism during brain development will become a major focus of investigation for many years to come.
Finally, although not directly related to CNS function, it is important to note the crucial role played by IDO and its regulation at the earliest stages of implantation, embryogenesis, maternal immune tolerance and the maternal-fetal balance. These factors and interactions (reviewed by Badawy et al., 2016; see Stone, 2016) are important influences on the initiation and maintenance of a successful pregnancy and hence a complete and normal CNS development.
CNS diseases: Huntington’s disease as an example
As described and discussed in several articles in this issue of “Neuropharmacology”, dysfunctions in kynurenine pathway metabolism have been tentatively linked to the etiology of a considerable number of CNS diseases. Paradigmatically, we are limiting our comments here to the potential role of kynurenines in the pathogenesis of the neurodegenerative disorder HD, which is inherited in an autosomal dominant fashion. As noted earlier, this hypothesis originated more than three decades ago with the demonstration that an intrastriatal injection of quinolinic acid in rats duplicates an array of neuropathological, neurochemical and behavioural features that are characteristic of the human disease (Schwarcz et al., 2010). Subsequent post-mortem studies in humans revealed a significantly increased activity of 3-hydroxyanthranilic acid dioxygenase, the immediate biosynthetic enzyme of quinolinic acid, in brain tissue of HD victims compared to controls (Schwarcz et al., 1988), and found substantially elevated levels of quinolinic acid in cortex and striatum of patients who had died in the early stages of the disease (Guidetti et al., 2004). In apparent further support of a pathophysiologically significant connection to kynurenines, the same HD samples also showed greatly increased levels of 3-hydroxykynurenine, which generates highly reactive free radicals and thereby potentiates the neurotoxic effects of quinolinic acid (Guidetti and Schwarcz, 1999).
The fact that HD is well understood from a genetic perspective offers new opportunities for assessing the role of the kynurenine pathway in pathology. The protein huntingtin, which holds the key to the disorder, is located on the short arm of chromosome 4 and contains a terminal poly-glutamine repeat chain, which is normally up to about 35 residues in length. If the chain length exceeds 40 residues, individuals exhibit HD symptoms. The severity of symptoms and disease progression increase with increasing numbers of glutamine repeats on mutant huntingtin (Forrest et al., 2010). This linkage allows investigators to directly test the involvement of molecular entities such as kynurenines in the disease process, and supportive data have been obtained in several genetic HD mouse models (Schwarcz et al., 2010). In a study in humans, HD patients and their healthy relatives (≤ 35 glutamine repeats) were classified into four groups based on symptom severity – no, few, moderate or severe. Samples of saliva were then collected for the determination of the number of poly-glutamine repeats, and blood was taken for the analysis of tryptophan and kynurenine in the serum. The results revealed highly significant correlations between the kynurenine/tryptophan ratio, i.e. the activity of the initial oxidative step in the kynurenine pathway, and both symptom severity and – crucially – the length of the poly-glutamine expansion in mutant huntingtin (Forrest et al., 2010). These results naturally raise a host of interesting questions and are especially intriguing on principal grounds, supporting the hypothesis that a genetically determined increase in kynurenine pathway activity may indeed contribute to the symptoms and disease progression in HD. More generally, this comprehensive approach, combining genetic assays and systematic clinical evaluations with chemical analyses in blood or other readily accessible body fluids, represents a promising model for the study of other CNS disorders with suspected etiological connections to kynurenines.
Future directions
As demonstrated above using selected examples, studies of kynurenine neurobiology have begun to diversify into many areas of interest. Additional recent discoveries illustrate the unpredictability of the field and at the same time highlight entirely new, translationally relevant research trajectories. Thus, work by Ruas and collaborators recently revealed that the skeletal muscle enzyme PGC1α1 modulates kynurenine metabolism and at the same time causes resilience to depression caused by chronic mild stress (Agudelo et al., 2014). Specifically, the authors report that the enzyme, when induced by exercise, up-regulates kynurenine aminotransferase activity and reduces circulating kynurenine levels, and propose that this effect is critical for the well-established antidepressant consequences of exercise in animals and humans. Although the study does not clarify the mechanism(s) by which brain kynurenines might be involved, this intriguing study implies that the treatment of depression might be possible by pharmacological modulation of PGC activity – in effect postulating an “exercise pill” based on modulating kynurenine pathway metabolism.
Another recent study, describing the effects of nutritional restriction on tryptophan degradation along the kynurenine pathway, may also be relevant for the pathophysiology of depression since altered appetite is a common symptom of the disease. Thus, in the nematode C. elegans, reduced intake of tryptophan was found to lower the levels of kynurenic acid, allowing increased activation of NMDA receptors. This, in turn, resulted in higher activity of neuropeptide Y-containing cells, which secondarily promoted feeding behaviour by stimulating serotonergic neurons (Lemieux et al., 2015). This intriguing neurochemical cascade is consistent with studies in mammals, which show that feeding is promoted by the activation of serotonin 2C receptors (Voigt and Fink, 2015). Indeed over-activation of these receptors may contribute to abnormally high appetite and, ultimately, obesity. In line with these findings and observations, it will be interesting to explore whether modification of kynurenine pathway metabolism, and specifically a reduction of kynurenic acid synthesis, holds promise as an intervention to achieve appetite reduction in humans.
Another noteworthy study in C. elegans demonstrated that a loss of TDO2 activity reduces neuronal death associated with three potent neurotoxic peptides: α-synuclein, amyloid-β and extended poly-glutamine peptides (van der Goot and Nollen, 2013; van der Goot et al., 2012). These findings imply that TDO2 may normally facilitate the toxicity of the peptides, possibly involving a distinct site on the TDO2 molecule which normally regulates protein metabolism. Intriguingly, a reduction in TDO2 expression also results in a prolongation of lifespan in the worms (see Badawy, 2016; Oxenkrug et al., 2011). If these remarkable effects can be duplicated in mammals, efforts to decipher the underlying molecular mechanism(s) will certainly multiply. One of the many questions to be addressed is whether similarly impressive effects can be achieved by down-regulation of IDO and TDO1, which are highly conserved orthologs of TDO2.
Another topic that deserves careful study and is beginning to receive increasing attention is the link between kynurenines and stem cell biology. Human and mouse mesenchymal and neural stem cells express the entire kynurenine pathway (Croitoru-Lamoury et al., 2011), and pluripotent adult progenitor cells have IDO-dependent immunomodulatory properties, implicating kynurenine, and possibly its downstream metabolites, as effectors (Jacobs et al., 2013). Of the many open questions, clarification of the mechanisms by which kynurenines influence stem cell proliferation and differentiation may be of greatest interest to cell biologists and other pre-clinical researchers. However, a thorough understanding of these effects will also increase the likelihood of success of stem cell-based therapies in a variety of infectious and inflammatory diseases affecting the brain.
In the context of depressive illness, there is a pressing need to clarify whether kynurenines are unequivocally involved in pathophysiology. The existence of this connection, specifically a role of an immune-stimulated increase in cerebral quinolinic acid levels in pathogenesis, has been repeatedly proposed, but the hypothesis has not been fully confirmed so far (Clark et al., 2016). Notably, changes in kynurenine pathway metabolism may be preferentially seen in depressed patients with suicidal intent. Here, the levels of kynurenic acid are significantly reduced in patients and correlated with the severity of depressive symptoms (Bradley et al., 2015; Bryleva and Brundin, 2016; Sublette et al., 2011). Conceivably, therefore, the disease may be causally related to an imbalance between quinolinic acid and kynurenic acid in the brain, with a relative abundance of the former leading to enhanced glutamatergic activity and symptoms of depression. More generally, it is interesting to speculate that depressive states may be associated with reduced kynurenic acid synthesis, causing kynurenine to be increasingly converted to quinolinic acid. In people with schizophrenia, in contrast, quinolinic acid production is less efficient due to the dysfunction of KMO, diverting kynurenine pathway metabolism towards enhanced kynurenic acid formation (Holtze et al. 2012; Wonodi et al. 2011, 2014). Of note, this scenario would not only be in line with the efficacy of NMDA antagonists as rapidly acting antidepressants (Coyle and Laws, 2015; Drewniany et al., 2015; Machado-Vieira et al., 2009) but raises the possibility that interventions causing an elevation of brain kynurenic acid levels may provide clinical benefits in major depressive disorders.
New genetic information, too, will provide valuable insights into the roles of kynurenines in physiological processes and, in particular, in pathological phenomena affecting the CNS. The presence and remarkable conservation of most elements of the kynurenine pathway in organisms ranging from prokaryotes to humans, together with the recent development of highly sophisticated and precise molecular tools to study and manipulate DNA or RNA, will undoubtedly generate novel – and testable – biological hypotheses. One of the most interesting experimental targets in this regard is the pivotal pathway enzyme KMO. In fact, specific genetic features such as distinct single nucleotide polymorphisms in Kmo in patients with schizophrenia (see above), and reduced levels of the mRNA for KMO in prefrontal cortex tissue from patients with bipolar disorder with psychotic features (Lavebratt et al., 2014), have been identified in recent years. Interestingly, the latter study revealed, specifically, an association between a KMO Arg(452) allele and manic psychosis, and between this allele and raised levels of kynurenic acid in these patients. All these findings await verification in larger cohorts and, subsequently, careful evaluation of functional consequences.
Finally, we assert that thorough investigations of the relationship between kynurenines and other (neuro)biologically active tryptophan metabolites are overdue and of fundamental importance. Although functionally significant interactions between kynurenine pathway metabolites and other prominent tryptophan products such as serotonin and melatonin are not unlikely, far too little is known in this respect. In the brain as in the periphery, cross-talk of the various catabolic branches of tryptophan, perhaps also involving additional metabolites such as tryptamine, will surely turn out to be complex, and elucidation will be challenging. Uni- or bi-directional influences could well vary with brain region, life stage, circadian rhythm and physiological requirements, and may differ among species. Most likely, pathological events will need to be assessed individually in this respect, though commonalities may exist in maladies with shared features, for example excitotoxic neurodegeneration or abnormalities of the immune system.
Summary and final comments
Research on the neurobiology of the kynurenine pathway has suffered years of relative obscurity because tryptophan degradation, and its involvement in both physiology and major brain diseases such as depression, schizophrenia (Erhardt et al., 2016) and neurodegenerative disorders, was viewed almost exclusively through the lens of the well-established metabolite serotonin (and to some extent melatonin). With increasing recognition that kynurenine and its metabolites can affect and even control a variety of classic neurotransmitter systems directly and indirectly, interest is expanding rapidly. Moreover, as evidence accumulates that, in turn, kynurenine pathway metabolism itself is modulated in conditions such as infection and stress, which are known to induce major changes in well-being and behaviour, it is also becoming apparent that kynurenines may be instrumental in the etiology of psychiatric and neurological disorders (Fujigaki et al., 2016; Hunt et al., 2016). It is therefore likely that the near future will not only witness the discovery of additional physiological and pathological roles for brain kynurenines, but also ever-increasing interest in drug development based on these roles. In particular, targeting the kynurenine pathway with new specific agents may make it possible to prevent disease by appropriate pharmacological manipulations (Dounay et al., 2012; Dounay et al., 2015; Jayawickrama et al., 2015; Stone and Darlington, 2013; Wu et al., 2014; Zwilling et al., 2011). Knowledge of the kynurenine pathway will be central to one of the cardinal achievements of medical research – averting disease rather than treating its symptoms.
We focused here on areas of kynurenine research which are either controversial, of major potential therapeutic interest, or just beginning to receive the degree of attention which will clarify their relevance to neurobiology and medicine. We also highlighted technical issues which require careful consideration in study design, experimental approaches and data evaluation. Although several of these concerns are not unique to the kynurenine pathway, we feel that it is important that investigators entering the field, and new research initiatives, are not misdirected by misleading results or incorrect interpretations at this time of skyrocketing interest in the subject matter.
Acknowledgments
Work in our laboratories has been funded mainly by the Medical Research Council (UK), Biotechnology and Biology Research Council (UK), the Wellcome Trust and Epsom Medical Research, and the National Institutes of Health (USA).
References
- Agudelo LZ, Femenia T, Orhan F, Porsmyr-Palmertz M, Goiny M, Martinez-Redondo V, Correia JC, Izadi M, Bhat M, Schuppe-Koistinen I, Pettersson AT, Ferreira DM, Krook A, Barres R, Zierath JR, Erhardt S, Lindskog M, Ruas JL. Skeletal muscle PGC-1alpha1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell. 2014;159:33–45. doi: 10.1016/j.cell.2014.07.051. [DOI] [PubMed] [Google Scholar]
- Albuquerque EX, Schwarcz R. Kynurenic acid as an antagonist of alpha7 nicotinic acetylcholine receptors in the brain: facts and challenges. Biochem Pharmacol. 2013;85:1027–1032. doi: 10.1016/j.bcp.2012.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alkondon M, Pereira EF, Todd SW, Randall WR, Lane MV, Albuquerque EX. Functional G-protein-coupled receptor 35 is expressed by neurons in the CA1 field of the hippocampus. Biochem Pharmacol. 2015;93:506–518. doi: 10.1016/j.bcp.2014.12.009. [DOI] [PubMed] [Google Scholar]
- Aoyama N, Takahashi N, Saito S, Maeno N, Ishihara R, Ji X, Miura H, Ikeda M, Suzuki T, Kitajima T, Yamanouchi Y, Kinoshita Y, Yoshida K, Iwata N, Inada T, Ozaki N. Association study between kynurenine 3-monooxygenase gene and schizophrenia in the Japanese population. Genes Brain Behav. 2006;5:364–368. doi: 10.1111/j.1601-183X.2006.00231.x. [DOI] [PubMed] [Google Scholar]
- Arnaiz-Cot JJ, Gonzalez JC, Sobrado M, Baldelli P, Carbone E, Gandia L, Garcia AG, Hernandez-Guijo JM. Allosteric modulation of alpha 7 nicotinic receptors selectively depolarizes hippocampal interneurons, enhancing spontaneous GABAergic transmission. Eur J Neurosci. 2008;27:1097–1110. doi: 10.1111/j.1460-9568.2008.06077.x. [DOI] [PubMed] [Google Scholar]
- Asp L, Holtze M, Powell SB, Karlsson H, Erhardt S. Neonatal infection with neurotropic influenza A virus induces the kynurenine pathway in early life and disrupts sensorimotor gating in adult Tap1−/− mice. Int J Neuropsychopharmacol. 2010;13:475–485. doi: 10.1017/S1461145709990253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badawy A. Tryptophan availability for kynurenine pathway metabolism across the life span: Control mechanisms and focus on aging, exercise, diet and nutritional supplements. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2015.11.015. this issue. [DOI] [PubMed] [Google Scholar]
- Badawy A, Namboodiri MA, Moffett JR. The end of the road for the tryptophan depletion concept in pregnancy and infection. Clin Sci. 2016;130:1327–1333. doi: 10.1042/CS20160153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ballesteros MC, Hansen PE, Soila K. MR imaging of the developing human brain. Part 2 Postnatal development. Radiographics. 1993;13:611–622. doi: 10.1148/radiographics.13.3.8316668. [DOI] [PubMed] [Google Scholar]
- Banerjee T, Duhadaway JB, Gaspari P, Sutanto-Ward E, Munn DH, Mellor AL, Malachowski WP, Prendergast GC, Muller AJ. A key in vivo antitumor mechanism of action of natural product-based brassinins is inhibition of indoleamine 2,3-dioxygenase. Oncogene. 2008;27:2851–2857. doi: 10.1038/sj.onc.1210939. [DOI] [PubMed] [Google Scholar]
- Barends TR, Dunn MF, Schlichting I. Tryptophan synthase, an allosteric molecular factory. Curr Opin Chem Biol. 2008;12:593–600. doi: 10.1016/j.cbpa.2008.07.011. [DOI] [PubMed] [Google Scholar]
- Bauer PV, Hamr SC, Duca FA. Regulation of energy balance by a gut-brain axis and involvement of the gut microbiota. Cell Mol Life Sci. 2016;73:737–755. doi: 10.1007/s00018-015-2083-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beal MF, Swartz KJ, Isacson O. Development changes in brain kynurenic acid concentrations. Brain Res Dev Brain Res. 1992;68:136–139. doi: 10.1016/0165-3806(92)90256-v. [DOI] [PubMed] [Google Scholar]
- Beggiato S, Antonelli T, Tomasini MC, Tanganelli S, Fuxe K, Schwarcz R, Ferraro L. Kynurenic acid, by targeting alpha7 nicotinic acetylcholine receptors, modulates extracellular GABA levels in the rat striatum in vivo. Eur J Neurosci. 2013;37:1470–1477. doi: 10.1111/ejn.12160. [DOI] [PubMed] [Google Scholar]
- Beggiato S, Tanganelli S, Fuxe K, Antonelli T, Schwarcz R, Ferraro L. Endogenous kynurenic acid regulates extracellular GABA levels in the rat prefrontal cortex. Neuropharmacology. 2014;82:11–18. doi: 10.1016/j.neuropharm.2014.02.019. [DOI] [PubMed] [Google Scholar]
- Bender DA, Njagi EN, Danielian PS. Tryptophan metabolism in vitamin B6-deficient mice. Br J Nutr. 1990;63:27–36. doi: 10.1079/bjn19900089. [DOI] [PubMed] [Google Scholar]
- Bessede A, Gargaro M, Pallotta MT, Matino D, Servillo G, Brunacci C, Bicciato S, Mazza EM, Macchiarulo A, Vacca C, Iannitti R, Tissi L, Volpi C, Belladonna ML, Orabona C, Bianchi R, Lanz TV, Platten M, Della Fazia MA, Piobbico D, Zelante T, Funakoshi H, Nakamura T, Gilot D, Denison MS, Guillemin GJ, DuHadaway JB, Prendergast GC, Metz R, Geffard M, Boon L, Pirro M, Iorio A, Veyret B, Romani L, Grohmann U, Fallarino F, Puccetti P. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature. 2014;511:184–190. doi: 10.1038/nature13323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birch PJ, Grossman CJ, Hayes AG. Kynurenate and FG9041 have both competitive and non-competitive antagonist actions at excitatory amino acid receptors. Eur J Pharmacol. 1988a;151:313–315. doi: 10.1016/0014-2999(88)90814-x. [DOI] [PubMed] [Google Scholar]
- Birch PJ, Grossman CJ, Hayes AG. Kynurenic acid antagonises responses to NMDA via an action at the strychnine-insensitive glycine receptor. Eur J Pharmacol. 1988b;154:85–87. doi: 10.1016/0014-2999(88)90367-6. [DOI] [PubMed] [Google Scholar]
- Botting NP. Chemistry and neurochemistry of the kynurenine pathway of tryptophan metabolism. Chem Soc Rev. 1995;24:401–412. [Google Scholar]
- Bradley KA, Case JA, Khan O, Ricart T, Hanna A, Alonso CM, Gabbay V. The role of the kynurenine pathway in suicidality in adolescent major depressive disorder. Psychiatry Res. 2015;227:206–212. doi: 10.1016/j.psychres.2015.03.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryleva EY, Brundin L. Kynurenine pathway metabolites and suicidality. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.01.034. this issue. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceresoli-Borroni G, Schwarcz R. Perinatal kynurenine pathway metabolism in the normal and asphyctic rat brain. Amino Acids. 2000;19:311–323. doi: 10.1007/s007260070062. [DOI] [PubMed] [Google Scholar]
- Ceresoli G, Guidetti P, Schwarcz R. Metabolism of [5-(3)H]kynurenine in the developing rat brain in vivo: effect of intrastriatal ibotenate injections. Brain Res Dev Brain Res. 1997;100:73–81. doi: 10.1016/s0165-3806(97)00029-1. [DOI] [PubMed] [Google Scholar]
- Chen Y, Brew BJ, Guillemin GJ. Characterization of the kynurenine pathway in NSC-34 cell line: implications for amyotrophic lateral sclerosis. J Neurochem. 2011;118:816–825. doi: 10.1111/j.1471-4159.2010.07159.x. [DOI] [PubMed] [Google Scholar]
- Chiarugi A, Calvani M, Meli E, Traggiai E, Moroni F. Synthesis and release of neurotoxic kynurenine metabolites by human monocyte-derived macrophages. J Neuroimmunol. 2001;120:190–198. doi: 10.1016/s0165-5728(01)00418-0. [DOI] [PubMed] [Google Scholar]
- Chobot V, Hadacek F, Weckwerth W, Kubicova L. Iron chelation and redox chemistry of anthranilic acid and 3-hydroxyanthranilic acid: A comparison of two structurally related kynurenine pathway metabolites to obtain improved insights into their potential role in neurological disease development. J Organomet Chem. 2015;782:103–110. doi: 10.1016/j.jorganchem.2015.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark CJ, Mackay GM, Smythe GA, Bustamante S, Stone TW, Phillips RS. Prolonged survival of a murine model of cerebral malaria by kynurenine pathway inhibition. Infect Immun. 2005;73:5249–5251. doi: 10.1128/IAI.73.8.5249-5251.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark SM, Pocivavsek A, Nicholson JD, Notarangelo F, Langenberg P, McMahon RP, Kleinman JE, Hyde TM, Stiller J, Postolache TT, Schwarcz R, Tonelli LH. Reduced kynurenine pathway metabolism and cytokine expression in the prefrontal cortex of depressed individuals. J Psychiatry Neurosci. 2016 doi: 10.1503/jpn.150226. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke G, McKernan DP, Gaszner G, Quigley EM, Cryan JF, Dinan TG. A distinct profile of tryptophan metabolism along the kynurenine pathway downstream of toll-like receptor activation in irritable bowel syndrome. Front Pharmacol. 2012;3:90. doi: 10.3389/fphar.2012.00090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleaves HJ, Miller SL. The nicotinamide biosynthetic pathway is a by-product of the RNA world. J Mol Evol. 2001;52:73–77. doi: 10.1007/s002390010135. [DOI] [PubMed] [Google Scholar]
- Coyle CM, Laws KR. The use of ketamine as an antidepressant: a systematic review and meta-analysis. Hum Psychopharmacol. 2015;30:152–163. doi: 10.1002/hup.2475. [DOI] [PubMed] [Google Scholar]
- Croitoru-Lamoury J, Lamoury FM, Caristo M, Suzuki K, Walker D, Takikawa O, Taylor R, Brew BJ. Interferon-gamma regulates the proliferation and differentiation of mesenchymal stem cells via activation of indoleamine 2,3 dioxygenase (IDO) PLoS One. 2011;6:e14698. doi: 10.1371/journal.pone.0014698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croucher MJ, Collins JF, Meldrum BS. Anticonvulsant action of excitatory amino acid antagonists. Science. 1982;216:899–901. doi: 10.1126/science.7079744. [DOI] [PubMed] [Google Scholar]
- Curzon G. Tryptophan pyrrolase–a biochemical factor in depressive illness? Br J Psychiatry. 1969;115:1367–1374. doi: 10.1192/bjp.115.529.1367. [DOI] [PubMed] [Google Scholar]
- Darlington LG, Forrest CM, Mackay GM, Smith RA, Smith AJ, Stoy N, Stone TW. On the biological importance of the 3-hydroxyanthranilic acid: anthranilic acid ratio. Int J Tryptophan Res. 2010;3:51–59. doi: 10.4137/ijtr.s4282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darvesh S, Walsh R, Kumar R, Caines A, Roberts S, Magee D, Rockwood K, Martin E. Inhibition of human cholinesterases by drugs used to treat Alzheimer disease. Alzheimer Dis Assoc Disord. 2003;17:117–126. doi: 10.1097/00002093-200304000-00011. [DOI] [PubMed] [Google Scholar]
- Davies J, Evans RH, Francis AA, Watkins JC. Excitatory amino acid receptors and synaptic excitation in the mammalian central nervous system. J Physiol. 1979;75:641–654. [PubMed] [Google Scholar]
- Davies J, Francis AA, Jones AW, Watkins JC. 2-Amino-5-phosphonovalerate (2APV), a potent and selective antagonist of amino acid-induced and synaptic excitation. Neurosci Lett. 1981;21:77–81. doi: 10.1016/0304-3940(81)90061-6. [DOI] [PubMed] [Google Scholar]
- de Carvalho LP, Bochet P, Rossier J. The endogenous agonist quinolinic acid and the non endogenous homoquinolinic acid discriminate between NMDAR2 receptor subunits. Neurochem Int. 1996;28:445–452. doi: 10.1016/0197-0186(95)00091-7. [DOI] [PubMed] [Google Scholar]
- Dewhurst WG. Tryptophan pyrrolase–a biochemical factor in depressive illness? Br J Psychiatry. 1970;116:569–571. doi: 10.1192/bjp.116.534.569-a. [DOI] [PubMed] [Google Scholar]
- DiNatale BC, Murray IA, Schroeder JC, Flaveny CA, Lahoti TS, Laurenzana EM, Omiecinski CJ, Perdew GH. Kynurenic acid is a potent endogenous aryl hydrocarbon receptor ligand that synergistically induces interleukin-6 in the presence of inflammatory signaling. Toxicol Sci. 2010;115:89–97. doi: 10.1093/toxsci/kfq024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobelis P, Staley KJ, Cooper DC. Lack of modulation of nicotinic acetylcholine alpha-7 receptor currents by kynurenic acid in adult hippocampal interneurons. PLoS One. 2012;7:e41108. doi: 10.1371/journal.pone.0041108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dounay AB, Anderson M, Bechle BM, Campbell BM, Claffey MM, Evdokimov A, Evrard E, Fonseca KR, Gan X, Ghosh S, Hayward MM, Horner W, Kim JY, McAllister LA, Pandit J, Paradis V, Parikh VD, Reese MR, Rong S, Salafia MA, Schuyten K, Strick CA, Tuttle JB, Valentine J, Wang H, Zawadzke LE, Verhoest PR. Discovery of brain-penetrant, irreversible kynurenine aminotransferase II inhibitors for schizophrenia. ACS Med Chem Lett. 2012;3:187–192. doi: 10.1021/ml200204m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dounay AB, Tuttle JB, Verhoest PR. Challenges and opportunities in the discovery of new therapeutics targeting the kynurenine pathway. J Med Chem. 2015;58:8762–8782. doi: 10.1021/acs.jmedchem.5b00461. [DOI] [PubMed] [Google Scholar]
- Drewniany E, Han J, Hancock C, Jones RL, Lim J, Nemat Gorgani N, Sperry JK, 3rd, Yu HJ, Raffa RB. Rapid-onset antidepressant action of ketamine: potential revolution in understanding and future pharmacologic treatment of depression. J Clin Pharm Ther. 2015;40:125–130. doi: 10.1111/jcpt.12238. [DOI] [PubMed] [Google Scholar]
- Erhardt S, Schwieler L, Imbeault S, Engberg G. The kynurenine pathway and schizophrenia. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.05.020. this issue. [DOI] [PubMed] [Google Scholar]
- Forrest CM, Khalil OS, Pisar M, Darlington LG, Stone TW. Prenatal inhibition of the tryptophan-kynurenine pathway alters synaptic plasticity and protein expression in the rat hippocampus. Brain Res. 2013a;1504:1–15. doi: 10.1016/j.brainres.2013.01.031. [DOI] [PubMed] [Google Scholar]
- Forrest CM, Khalil OS, Pisar M, McNair K, Kornisiuk E, Snitcofsky M, Gonzalez N, Jerusalinsky D, Darlington LG, Stone TW. Changes in synaptic transmission and protein expression in the brains of adult offspring after prenatal inhibition of the kynurenine pathway. Neuroscience. 2013b;254:241–259. doi: 10.1016/j.neuroscience.2013.09.034. [DOI] [PubMed] [Google Scholar]
- Forrest CM, Mackay GM, Oxford L, Stoy N, Stone TW, Darlington LG. Kynurenine pathway metabolism in patients with osteoporosis after 2 years of drug treatment. Clin Exp Pharmacol Physiol. 2006;33:1078–1087. doi: 10.1111/j.1440-1681.2006.04490.x. [DOI] [PubMed] [Google Scholar]
- Forrest CM, Mackay GM, Stoy N, Spiden SL, Taylor R, Stone TW, Darlington LG. Blood levels of kynurenines, interleukin-23 and soluble human leucocyte antigen-G at different stages of Huntington’s disease. J Neurochem. 2010;112:112–122. doi: 10.1111/j.1471-4159.2009.06442.x. [DOI] [PubMed] [Google Scholar]
- Foster AC, Collins JF, Schwarcz R. On the excitotoxic properties of quinolinic acid, 2,3-piperidine dicarboxylic acids and structurally related compounds. Neuropharmacology. 1983;22:1331–1342. doi: 10.1016/0028-3908(83)90221-6. [DOI] [PubMed] [Google Scholar]
- Foster AC, Vezzani A, French ED, Schwarcz R. Kynurenic acid blocks neurotoxicity and seizures induced in rats by the related brain metabolite quinolinic acid. Neurosci Lett. 1984;48:273–278. doi: 10.1016/0304-3940(84)90050-8. [DOI] [PubMed] [Google Scholar]
- Fujigaki H, Yamamoto Y, Saito K. L-tryptophan-kynurenine pathway enzymes are therapeutic target for neuropsychiatric diseases: Focus on cell type differences. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.01.011. this issue. [DOI] [PubMed] [Google Scholar]
- Fukui S, Schwarcz R, Rapoport SI, Takada Y, Smith QR. Blood-brain barrier transport of kynurenines: implications for brain synthesis and metabolism. J Neurochem. 1991;56:2007–2017. doi: 10.1111/j.1471-4159.1991.tb03460.x. [DOI] [PubMed] [Google Scholar]
- Garavaglia S, Perozzi S, Galeazzi L, Raffaelli N, Rizzi M. The crystal structure of human alpha-amino-beta-carboxymuconate-epsilon-semialdehyde decarboxylase in complex with 1,3-dihydroxyacetonephosphate suggests a regulatory link between NAD synthesis and glycolysis. FEBS J. 2009;276:6615–6623. doi: 10.1111/j.1742-4658.2009.07372.x. [DOI] [PubMed] [Google Scholar]
- Gaspari P, Banerjee T, Malachowski WP, Muller AJ, Prendergast GC, DuHadaway J, Bennett S, Donovan AM. Structure-activity study of brassinin derivatives as indoleamine 2,3-dioxygenase inhibitors. J Med Chem. 2006;49:684–692. doi: 10.1021/jm0508888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghaisas S, Maher J, Kanthasamy A. Gut microbiome in health and disease: Linking the microbiome-gut-brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol Ther. 2016;158:52–62. doi: 10.1016/j.pharmthera.2015.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giorgini F, Huang SY, Sathyasaikumar KV, Notarangelo FM, Thomas MA, Tararina M, Wu HQ, Schwarcz R, Muchowski PJ. Targeted deletion of kynurenine 3-monooxygenase in mice: a new tool for studying kynurenine pathway metabolism in periphery and brain. J Biol Chem. 2013;288:36554–36566. doi: 10.1074/jbc.M113.503813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, Nugent TF, 3rd, Herman DH, Clasen LS, Toga AW, Rapoport JL, Thompson PM. Dynamic mapping of human cortical development during childhood through early adulthood. Proc Natl Acad Sci U S A. 2004;101:8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez-Esquivel D, Ramirez-Ortega D, Pineda B, Castro N, Rios C, Perez de la Cruz V. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.03.013. this issue. [DOI] [PubMed] [Google Scholar]
- Guidetti P, Eastman CL, Schwarcz R. Metabolism of [5-3H]kynurenine in the rat brain in vivo: evidence for the existence of a functional kynurenine pathway. J Neurochem. 1995;65:2621–2632. doi: 10.1046/j.1471-4159.1995.65062621.x. [DOI] [PubMed] [Google Scholar]
- Guidetti P, Hoffman GE, Melendez-Ferro M, Albuquerque EX, Schwarcz R. Astrocytic localization of kynurenine aminotransferase II in the rat brain visualized by immunocytochemistry. Glia. 2007;55:78–92. doi: 10.1002/glia.20432. [DOI] [PubMed] [Google Scholar]
- Guidetti P, Luthi-Carter RE, Augood SJ, Schwarcz R. Neostriatal and cortical quinolinate levels are increased in early grade Huntington’s disease. Neurobiol Dis. 2004;17:455–461. doi: 10.1016/j.nbd.2004.07.006. [DOI] [PubMed] [Google Scholar]
- Guidetti P, Schwarcz R. 3-Hydroxykynurenine potentiates quinolinate but not NMDA toxicity in the rat striatum. Eur J Neurosci. 1999;11:3857–3863. doi: 10.1046/j.1460-9568.1999.00806.x. [DOI] [PubMed] [Google Scholar]
- Guillemin GJ. Quinolinic acid: the inescapable neurotoxin. FEBS J. 2012;279:1356–1365. doi: 10.1111/j.1742-4658.2012.08485.x. [DOI] [PubMed] [Google Scholar]
- Guillemin GJ, Cullen KM, Lim CK, Smythe GA, Garner B, Kapoor V, Takikawa O, Brew BJ. Characterization of the kynurenine pathway in human neurons. J Neurosci. 2007;27:12884–12892. doi: 10.1523/JNEUROSCI.4101-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillemin GJ, Kerr SJ, Smythe GA, Smith DG, Kapoor V, Armati PJ, Croitoru J, Brew BJ. Kynurenine pathway metabolism in human astrocytes: a paradox for neuronal protection. J Neurochem. 2001;78:1–13. doi: 10.1046/j.1471-4159.2001.00498.x. [DOI] [PubMed] [Google Scholar]
- Guillemin GJ, Smythe G, Takikawa O, Brew BJ. Expression of indoleamine 2,3-dioxygenase and production of quinolinic acid by human microglia, astrocytes, and neurons. Glia. 2005;49:15–23. doi: 10.1002/glia.20090. [DOI] [PubMed] [Google Scholar]
- Han Q, Cai T, Tagle DA, Li J. Structure, expression, and function of kynurenine aminotransferases in human and rodent brains. Cell Mol Life Sci. 2010;67:353–368. doi: 10.1007/s00018-009-0166-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heyes MP, Chen CY, Major EO, Saito K. Different kynurenine pathway enzymes limit quinolinic acid formation by various human cell types. Biochem J. 1997;326:351–356. doi: 10.1042/bj3260351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilmas C, Pereira EF, Alkondon M, Rassoulpour A, Schwarcz R, Albuquerque EX. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: physiopathological implications. J Neurosci. 2001;21:7463–7473. doi: 10.1523/JNEUROSCI.21-19-07463.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holtze M, Saetre P, Engberg G, Schwieler L, Werge T, Andreassen OA, Hall H, Terenius L, Agartz I, Jonsson EG, Schalling M, Erhardt S. Kynurenine 3-monooxygenase polymorphisms: relevance for kynurenic acid synthesis in patients with schizophrenia and healthy controls. J Psychiatry Neurosci. 2012;37:53–57. doi: 10.1503/jpn.100175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoshi M, Saito K, Murakami Y, Taguchi A, Fujigaki H, Tanaka R, Takemura M, Ito H, Hara A, Seishima M. Marked increases in hippocampal neuron indoleamine 2, 3-dioxygenase via IFN-gamma-independent pathway following transient global ischemia in mouse. Neurosci Res. 2009;63:194–198. doi: 10.1016/j.neures.2008.12.003. [DOI] [PubMed] [Google Scholar]
- Hunt NH, Too LK, Khaw IT, Gun J, Hee L, Mitchell AJ, Grau GE, Ball HJ. The kynurenine pathway and parasitic infections that affect CNS function. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.02.029. this issue. [DOI] [PubMed] [Google Scholar]
- Jacobs SA, Pinxteren J, Roobrouck VD, Luyckx A, van’t Hof W, Deans R, Verfaillie CM, Waer M, Billiau AD, Van Gool SW. Human multipotent adult progenitor cells are nonimmunogenic and exert potent immunomodulatory effects on alloreactive T-cell responses. Cell Transplant. 2013;22:1915–1928. doi: 10.3727/096368912X657369. [DOI] [PubMed] [Google Scholar]
- Jayawickrama GS, Sadig RR, Sun G, Nematollahi A, Nadvi NA, Hanrahan JR, Gorrell MD, Church WB. Kynurenine aminotransferases and the prospects of inhibitors for the treatment of schizophrenia. Curr Med Chem. 2015;22:2902–2918. doi: 10.2174/0929867322666150608094054. [DOI] [PubMed] [Google Scholar]
- Jernigan TL, Baare WF, Stiles J, Madsen KS. Postnatal brain development: structural imaging of dynamic neurodevelopmental processes. Prog Brain Res. 2011;189:77–92. doi: 10.1016/B978-0-444-53884-0.00019-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy PJ, Cryan JF, Dinan TG, Clarke G. Kynurenine pathway metabolism and the microbiome-gut-brain axis. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.07.002. this issue. [DOI] [PubMed] [Google Scholar]
- Khalil OS, Forrest CM, Pisar M, Smith RA, Darlington LG, Stone TW. Prenatal activation of maternal TLR3 receptors by viral-mimetic poly(I:C) modifies GluN2B expression in embryos and sonic hedgehog in offspring in the absence of kynurenine pathway activation. Immunopharmacol Immunotoxicol. 2013;35:581–593. doi: 10.3109/08923973.2013.828745. [DOI] [PubMed] [Google Scholar]
- Kiss C, Ceresoli-Borroni G, Guidetti P, Zielke CL, Zielke HR, Schwarcz R. Kynurenate production by cultured human astrocytes. J Neural Transm. 2003;110:1–14. doi: 10.1007/s00702-002-0770-z. [DOI] [PubMed] [Google Scholar]
- Kocki T, Dolinska M, Dybel A, Urbanka EM, Turski WA, Albrecht J. Regulation of kynurenic acid synthesis in C6 glioma cells. J Neurosci Res. 2002;68:622–626. doi: 10.1002/jnr.10243. [DOI] [PubMed] [Google Scholar]
- Krause D, Suh HS, Tarassishin L, Cui QL, Durafourt BA, Choi N, Bauman A, Cosenza-Nashat M, Antel JP, Zhao ML, Lee SC. The tryptophan metabolite 3-hydroxyanthranilic acid plays anti-inflammatory and neuroprotective roles during inflammation: role of hemeoxygenase-1. Am J Pathol. 2011;179:1360–1372. doi: 10.1016/j.ajpath.2011.05.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapin IP. Stimulant and convulsive effects of kynurenines injected into brain ventricles in mice. J Neural Transm. 1978;42:37–43. doi: 10.1007/BF01262727. [DOI] [PubMed] [Google Scholar]
- Laugeray A, Launay JM, Callebert J, Surget A, Belzung C, Barone PR. Peripheral and cerebral metabolic abnormalities of the tryptophan-kynurenine pathway in a murine model of major depression. Behav Brain Res. 2010;210:84–91. doi: 10.1016/j.bbr.2010.02.014. [DOI] [PubMed] [Google Scholar]
- Lavebratt C, Olsson S, Backlund L, Frisen L, Sellgren C, Priebe L, Nikamo P, Traskman-Bendz L, Cichon S, Vawter MP, Osby U, Engberg G, Landen M, Erhardt S, Schalling M. The KMO allele encoding Arg452 is associated with psychotic features in bipolar disorder type 1, and with increased CSF KYNA level and reduced KMO expression. Mol Psychiatry. 2014;19:334–341. doi: 10.1038/mp.2013.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J, Tan V, Braidy N, Brew BJ, Guillemin GJ. Implications of the neurotoxin quinolinic acid in the neuropathogenesis of amyotrophic lateral sclerosis. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.05.011. this issue. [DOI] [PubMed] [Google Scholar]
- Leeson PD, Baker R, Carling RW, Curtis NR, Moore KW, Williams BJ, Foster AC, Donald AE, Kemp JA, Marshall GR. Kynurenic acid derivatives. Structure-activity relationships for excitatory amino acid antagonism and identification of potent and selective antagonists at the glycine site on the N-methyl-D-aspartate receptor. J Med Chem. 1991;34:1243–1252. doi: 10.1021/jm00108a002. [DOI] [PubMed] [Google Scholar]
- Lehmann JC, Procureur D, Wood PL. 7-Chlorokynurenate prevents NMDA-induced and kainate-induced striatal lesions. Brain Res. 1993;620:1–6. doi: 10.1016/0006-8993(93)90263-m. [DOI] [PubMed] [Google Scholar]
- Lehrmann E, Molinari A, Speciale C, Schwarcz R. Immunohistochemical visualization of newly formed quinolinate in the normal and excitotoxically lesioned rat striatum. Exp Brain Res. 2001;141:389–397. doi: 10.1007/s002210100887. [DOI] [PubMed] [Google Scholar]
- Lemieux GA, Cunningham KA, Lin L, Mayer F, Werb Z, Ashrafi K. Kynurenic acid is a nutritional cue that enables behavioral plasticity. Cell. 2015;160:119–131. doi: 10.1016/j.cell.2014.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Libbey JE, Sweeten TL, McMahon WM, Fujinami RS. Autistic disorder and viral infections. J Neurovirol. 2005;11:1–10. doi: 10.1080/13550280590900553. [DOI] [PubMed] [Google Scholar]
- Linderholm KR, Andersson A, Olsson S, Olsson E, Snodgrass R, Engberg G, Erhardt S. Activation of rat ventral tegmental area dopamine neurons by endogenous kynurenic acid: a pharmacological analysis. Neuropharmacology. 2007;53:918–924. doi: 10.1016/j.neuropharm.2007.09.003. [DOI] [PubMed] [Google Scholar]
- Liu X, Dong Y, Li X, Ren Y, Li Y, Wang W, Wang L, Feng L. Characterization of the anthranilate degradation pathway in Geobacillus thermodenitrificans NG80-2. Microbiology. 2010;156:589–595. doi: 10.1099/mic.0.031880-0. [DOI] [PubMed] [Google Scholar]
- Lopes C, Pereira EF, Wu HQ, Purushottamachar P, Njar V, Schwarcz R, Albuquerque EX. Competitive antagonism between the nicotinic allosteric potentiating ligand galantamine and kynurenic acid at alpha7* nicotinic receptors. J Pharmacol Exp Ther. 2007;322:48–58. doi: 10.1124/jpet.107.123109. [DOI] [PubMed] [Google Scholar]
- Lovelace MD, Varney B, Sundaram G, Lennon MJ, Lim CK, Jacobs K, Guillemin GJ, Brew BJ. Recent evidence for an expaned role of the kynruenine pathway of tryptophan metabolism in neurological diseases. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.03.024. this issue. [DOI] [PubMed] [Google Scholar]
- Machado-Vieira R, Salvadore G, Diazgranados N, Zarate CA., Jr Ketamine and the next generation of antidepressants with a rapid onset of action. Pharmacol Ther. 2009;123:143–150. doi: 10.1016/j.pharmthera.2009.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinoni I, Nonnis S, Monteferrante C, Heathcote P, Hartig E, Bottger LH, Trautwein AX, Negri A, Albertini AM, Tedeschi G. Characterization of L-aspartate oxidase and quinolinate synthase from Bacillus subtilis. FEBS J. 2008;275:5090–5107. doi: 10.1111/j.1742-4658.2008.06641.x. [DOI] [PubMed] [Google Scholar]
- McLennan H. Receptors for the excitatroy amino acids in the mammalian central nervous system. Prog Neurobiol. 1983;20:251–271. doi: 10.1016/0301-0082(83)90004-7. [DOI] [PubMed] [Google Scholar]
- Metzger F. Molecular and cellular control of dendrite maturation during brain development. Curr Mol Pharmacol. 2010;3:1–11. doi: 10.2174/1874467211003010001. [DOI] [PubMed] [Google Scholar]
- Mohamed Eissa AA, Soliman GA, Khataibeh MH. Design, synthesis and anti-inflammatory activity of structurally simple anthranilic acid congeners devoid of ulcerogenic side effects. Chem Pharm Bull (Tokyo) 2012;60:1290–1300. doi: 10.1248/cpb.c12-00516. [DOI] [PubMed] [Google Scholar]
- Mok MH, Fricker AC, Weil A, Kew JN. Electrophysiological characterisation of the actions of kynurenic acid at ligand-gated ion channels. Neuropharmacology. 2009;57:242–249. doi: 10.1016/j.neuropharm.2009.06.003. [DOI] [PubMed] [Google Scholar]
- Monaghan DT, Beaton JA. Quinolinate differentiates between forebrain and cerebellar NMDA receptors. Eur J Pharmacol. 1991;194:123–125. doi: 10.1016/0014-2999(91)90134-c. [DOI] [PubMed] [Google Scholar]
- Moroni F, Cozzi A, Sili M, Mannaioni G. Kynurenic acid: a metabolite with multiple actions and multiple targets in brain and periphery. J Neural Transm. 2012;119:133–139. doi: 10.1007/s00702-011-0763-x. [DOI] [PubMed] [Google Scholar]
- Moroni F, Russi P, Lombardi G, Beni M, Carla V. Presence of kynurenic acid in the mammalian brain. J Neurochem. 1988;51:177–180. doi: 10.1111/j.1471-4159.1988.tb04852.x. [DOI] [PubMed] [Google Scholar]
- Musso T, Gusella GL, Brooks A, Longo DL, Varesio L. Interleukin-4 inhibits indoleamine 2,3-dioxygenase expression in human monocytes. Blood. 1994;83:1408–1411. [PubMed] [Google Scholar]
- Nilsson A, Duan J, Mo-Boquist LL, Benedikz E, Sundstrom E. Characterisation of the human NMDA receptor subunit NR3A glycine binding site. Neuropharmacology. 2007;52:1151–1159. doi: 10.1016/j.neuropharm.2006.12.002. [DOI] [PubMed] [Google Scholar]
- Nordberg A, Svensson AL. Cholinesterase inhibitors in the treatment of Alzheimer’s disease: a comparison of tolerability and pharmacology. Drug Saf. 1998;19:465–480. doi: 10.2165/00002018-199819060-00004. [DOI] [PubMed] [Google Scholar]
- Notarangelo FM, Pocivavsek A. Elevated kynurenine pathway metabolism during neurodevelopment: Implications for brain and behavior. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.03.001. this issue. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nugoli M, Chuchana P, Vendrell J, Orsetti B, Ursule L, Nguyen C, Birnbaum D, Douzery EJ, Cohen P, Theillet C. Genetic variability in MCF-7 sublines: evidence of rapid genomic and RNA expression profile modifications. BMC Cancer. 2003;3:13. doi: 10.1186/1471-2407-3-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Farrell K, Harkin A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disroders. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2015.12.004. this issue. [DOI] [PubMed] [Google Scholar]
- O’Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res. 2015;277:32–48. doi: 10.1016/j.bbr.2014.07.027. [DOI] [PubMed] [Google Scholar]
- O’Shea RD, Manallack DT, Conway EL, Mercer LD, Beart PM. Evidence for heterogenous glycine domains but conserved multiple states of the excitatory amino acid recognition site of the NMDA receptor: regional binding studies with [3H]glycine and [3H]L-glutamate. Exp Brain Res. 1991;86:652–662. doi: 10.1007/BF00230539. [DOI] [PubMed] [Google Scholar]
- Ojeda P, Bobe A, Dolan K, Leone V, Martinez K. Nutritional modulation of gut microbiota - the impact on metabolic disease pathophysiology. J Nutr Biochem. 2016;28:191–200. doi: 10.1016/j.jnutbio.2015.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opitz CA, Litzenburger UM, Lutz C, Lanz TV, Tritschler I, Koppel A, Tolosa E, Hoberg M, Anderl J, Aicher WK, Weller M, Wick W, Platten M. Toll-like receptor engagement enhances the immunosuppressive properties of human bone marrow-derived mesenchymal stem cells by inducing indoleamine-2,3-dioxygenase-1 via interferon-beta and protein kinase R. Stem Cells. 2009;27:909–919. doi: 10.1002/stem.7. [DOI] [PubMed] [Google Scholar]
- Opitz CA, Litzenburger UM, Sahm F, Ott M, Tritschler I, Trump S, Schumacher T, Jestaedt L, Schrenk D, Weller M, Jugold M, Guillemin GJ, Miller CL, Lutz C, Radlwimmer B, Lehmann I, von Deimling A, Wick W, Platten M. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature. 2011;478:197–203. doi: 10.1038/nature10491. [DOI] [PubMed] [Google Scholar]
- Owe-Young R, Webster NL, Mukhtar M, Pomerantz RJ, Smythe G, Walker D, Armati PJ, Crowe SM, Brew BJ. Kynurenine pathway metabolism in human blood-brain-barrier cells: implications for immune tolerance and neurotoxicity. J Neurochem. 2008;105:1346–1357. doi: 10.1111/j.1471-4159.2008.05241.x. [DOI] [PubMed] [Google Scholar]
- Oxenkrug GF, Navrotskaya V, Voroboyva L, Summergrad P. Extension of life span of Drosophila melanogaster by the inhibitors of tryptophan-kynurenine metabolism. Fly (Austin) 2011;5:307–309. doi: 10.4161/fly.5.4.18414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pardridge WM. Brain metabolism: a perspective from the blood-brain barrier. Physiol Rev. 1983;63:1481–1535. doi: 10.1152/physrev.1983.63.4.1481. [DOI] [PubMed] [Google Scholar]
- Pardridge WM. Blood-brain barrier carrier-mediated transport and brain metabolism of amino acids. Neurochem Res. 1998;23:635–644. doi: 10.1023/a:1022482604276. [DOI] [PubMed] [Google Scholar]
- Parrott JM, O’Connor JC. Kynurenine 3-monooxygenase: An influential mediator of neuropathology. Front Psychiatry. 2015;6:116. doi: 10.3389/fpsyt.2015.00116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pemberton LA, Kerr SJ, Smythe G, Brew BJ. Quinolinic acid production by macrophages stimulated with IFN-gamma, TNF-alpha, and IFN-alpha. J Interferon Cytokine Res. 1997;17:589–595. doi: 10.1089/jir.1997.17.589. [DOI] [PubMed] [Google Scholar]
- Perkins MN, Collins JF, Stone TW. Isomers of 2-amino-7-phosphonoheptanoic acid as antagonists of neuronal excitants. Neurosci Lett. 1982;32:65–68. doi: 10.1016/0304-3940(82)90230-0. [DOI] [PubMed] [Google Scholar]
- Perkins MN, Stone TW. An iontophoretic investigation of the actions of convulsant kynurenines and their interaction with the endogenous excitant quinolinic acid. Brain Res. 1982;247:184–187. doi: 10.1016/0006-8993(82)91048-4. [DOI] [PubMed] [Google Scholar]
- Perkins MN, Stone TW. Pharmacology and regional variations of quinolinic acid-evoked excitations in the rat central nervous system. J Pharmacol Exp Ther. 1983a;226:551–557. [PubMed] [Google Scholar]
- Perkins MN, Stone TW. Quinolinic acid: regional variations in neuronal sensitivity. Brain Res. 1983b;259:172–176. doi: 10.1016/0006-8993(83)91084-3. [DOI] [PubMed] [Google Scholar]
- Perkins MN, Stone TW, Collins JF, Curry K. Phosphonate analogues of carboxylic acids as amino acid antagonists on rat cortical neurones. Neurosci Lett. 1981;23:333–336. doi: 10.1016/0304-3940(81)90021-5. [DOI] [PubMed] [Google Scholar]
- Pfefferkorn ER. Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proc Natl Acad Sci U S A. 1984;81:908–912. doi: 10.1073/pnas.81.3.908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierozan P, Zamoner A, Soska AK, Silvestrin RB, Loureiro SO, Heimfarth L, Mello e Souza T, Wajner M, Pessoa-Pureur R. Acute intrastriatal administration of quinolinic acid provokes hyperphosphorylation of cytoskeletal intermediate filament proteins in astrocytes and neurons of rats. Exp, Neurol. 2010;224:188–196. doi: 10.1016/j.expneurol.2010.03.009. [DOI] [PubMed] [Google Scholar]
- Pisar M, Forrest CM, Khalil OS, McNair K, Vincenten MC, Qasem S, Darlington LG, Stone TW. Modified neocortical and cerebellar protein expression and morphology in adult rats following prenatal inhibition of the kynurenine pathway. Brain Res. 2014;1576:1–17. doi: 10.1016/j.brainres.2014.06.016. [DOI] [PubMed] [Google Scholar]
- Rahman A, Ting K, Cullen KM, Braidy N, Brew BJ, Guillemin GJ. The excitotoxin quinolinic acid induces tau phosphorylation in human neurons. PLoS ONE. 2009;4:e6344. doi: 10.1371/journal.pone.0006344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reichmann D, Coute Y, Ollagnier de Choudens S. Dual activity of quinolinate synthase: triose phosphate isomerase and dehydration activities play together to form quinolinate. Biochemistry (Mosc) 2015;54:6443–6446. doi: 10.1021/acs.biochem.5b00991. [DOI] [PubMed] [Google Scholar]
- Saito K, Nowak TSJ, Markey SP, Heyes MP. Mechanism of delayed increases in kynurenine pathway metabolism in damaged brain regions following transient cerebral ischemia. J Neurochem. 1993;60:180–192. doi: 10.1111/j.1471-4159.1993.tb05836.x. [DOI] [PubMed] [Google Scholar]
- Sakuraba H, Tsuge H, Yoneda K, Katunuma N, Ohshima T. Crystal structure of the NAD biosynthetic enzyme quinolinate synthase. J Biol Chem. 2005;280:26645–26648. doi: 10.1074/jbc.C500192200. [DOI] [PubMed] [Google Scholar]
- Santamaria A, Galvan-Arzate S, Lisy V, Ali SF, Duhart HM, Osorio-Rico L, Rios C, St’astny F. Quinolinic acid induces oxidative stress in rat brain synaptosomes. Neuroreport. 2001;12:871–874. doi: 10.1097/00001756-200103260-00049. [DOI] [PubMed] [Google Scholar]
- Saunders AH, Griffiths AE, Lee KH, Cicchillo RM, Tu L, Stromberg JA, Krebs C, Booker SJ. Characterization of quinolinate synthases from Escherichia coli, Mycobacterium tuberculosis, and Pyrococcus horikoshii indicates that [4Fe-4S] clusters are common cofactors throughout this class of enzymes. Biochemistry (Mosc) 2008;47:10999–11012. doi: 10.1021/bi801268f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherzer CR, Landwehrmeyer GB, Kerner JA, Standaert DG, Hollingsworth ZR, Daggett LP, Velicelebi G, Penney JB, Jr, Young AB. Cellular distribution of NMDA glutamate receptor subunit mRNAs in the human cerebellum. Neurobiol Dis. 1997;4:35–46. doi: 10.1006/nbdi.1997.0136. [DOI] [PubMed] [Google Scholar]
- Schwarcz R. Kynurenines and glutamate: multiple links and therapeutic implications. In: Schwarcz R, Enna SJ, editors. Advances in Pharmacology; Neuropsychopharmacology: a tribute of Joseph T Coyle. Elsevier; 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarcz R, Collins JF, Parks DA. alpha-Amino-omega-phosphono carboxylates block ibotenate but not kainate neurotoxicity in rat hippocampus. Neurosci Lett. 1982;33:85–90. doi: 10.1016/0304-3940(82)90134-3. [DOI] [PubMed] [Google Scholar]
- Schwarcz R, Guidetti P, Sathyasaikumar KV, Muchowski PJ. Of mice, rats and men: Revisiting the quinolinic acid hypothesis of Huntington’s disease. Prog Neurobiol. 2010;90:230–245. doi: 10.1016/j.pneurobio.2009.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarcz R, Meldrum B. Excitatory amino acid antagonists provide a therapeutic approach to neurological disorders. Lancet. 1985;2:140–143. doi: 10.1016/s0140-6736(85)90238-7. [DOI] [PubMed] [Google Scholar]
- Schwarcz R, Okuno E, White RJ, Bird ED, Whetsell WO., Jr 3-Hydroxyanthranilic oxygenase activity is increased in the brains of Huntington disease victims. Proc Natl Acad Sci U S A. 1988;85:4079–4081. doi: 10.1073/pnas.85.11.4079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarcz R, Scholz D, Coyle JT. Structure-activity relations for the neurotoxicity of kainic acid derivatives and glutamate analogues. Neuropharmacology. 1978;17:145–151. doi: 10.1016/0028-3908(78)90127-2. [DOI] [PubMed] [Google Scholar]
- Sharma S, Srivastava VK, Kumar A. Newer N-substituted anthranilic acid derivatives as potent anti-inflammatory agents. Eur J Med Chem. 2002;37:689–697. doi: 10.1016/s0223-5234(02)01340-5. [DOI] [PubMed] [Google Scholar]
- Shibata K, Ohno T, Sano M, Fukuwatari T. The urinary ratio of 3-hydroxykynurenine/3-hydroxyanthranilic acid is an index to predicting the adverse effects of D-tryptophan in rats. J Nutr Sci Vitaminol (Tokyo) 2014;60:261–268. doi: 10.3177/jnsv.60.261. [DOI] [PubMed] [Google Scholar]
- Smith DG, Guillemin GJ, Pemberton L, Kerr S, Nath A, Smythe GA, Brew BJ. Quinolinic acid is produced by macrophages stimulated by platelet activating factor, Nef and Tat. J Neurovirol. 2001;7:56–60. doi: 10.1080/135502801300069692. [DOI] [PubMed] [Google Scholar]
- Smothers CT, Woodward JJ. Pharmacological characterization of glycine-activated currents in HEK 293 cells expressing N-methyl-D-aspartate NR1 and NR3 subunits. J Pharmacol Exp Ther. 2007;322:739–748. doi: 10.1124/jpet.107.123836. [DOI] [PubMed] [Google Scholar]
- Stephens GL, Wang Q, Swerdlow B, Bhat G, Kolbeck R, Fung M. Kynurenine 3-monooxygenase mediates inhibition of Th17 differentiation via catabolism of endogenous aryl hydrocarbon receptor ligands. Eur J Immunol. 2013;43:1727–1734. doi: 10.1002/eji.201242779. [DOI] [PubMed] [Google Scholar]
- Stone TW. Cortical pyramidal tract interneurones and their sensitivity to L-glutamic acid. J Physiol. 1973;233:211–225. doi: 10.1113/jphysiol.1973.sp010306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone TW. Blockade by amino acid antagonists of neuronal excitation mediated by the pyramidal tract. J Physiol. 1976;257:187–198. doi: 10.1113/jphysiol.1976.sp011363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone TW. Tryptophan and kynurenines: continuing to court controversy. Clin Sci. 2016;130:1335–1337. doi: 10.1042/CS20160294. [DOI] [PubMed] [Google Scholar]
- Stone TW, Behan WMH. Interkeukin-1β but not TNF-α potentiates neuronal damage by quinolinic acid: protection by an adenosine A(2A) receptor antagonist. J Neurosci Res. 2007;85:1077–1085. doi: 10.1002/jnr.21212. [DOI] [PubMed] [Google Scholar]
- Stone TW, Darlington LG. Endogenous kynurenines as targets for drug discovery and development. Nat Rev Drug Discov. 2002;1:609–620. doi: 10.1038/nrd870. [DOI] [PubMed] [Google Scholar]
- Stone TW, Darlington LG. The kynurenine pathway as a therapeutic target in cognitive and neurodegenerative disorders. Br J Pharmacol. 2013;169:1211–1227. doi: 10.1111/bph.12230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone TW, Perkins MN. Quinolinic acid: a potent endogenous excitant at amino acid receptors in CNS. Eur J Pharmacol. 1981;72:411–412. doi: 10.1016/0014-2999(81)90587-2. [DOI] [PubMed] [Google Scholar]
- Stone TW, Stoy N, Darlington LG. An expanding range of targets for kynurenine metabolites of tryptophan. Trends Pharmacol Sci. 2013;34:136–143. doi: 10.1016/j.tips.2012.09.006. [DOI] [PubMed] [Google Scholar]
- Strasser B, K B, Fuchs D, Gostner JM. Kynurenine pathway metabolism and immune activation: Peripheral measurements in psychiatric and co-morbid conditions. Neuropharmacology. 2016 doi: 10.1016/j.neuropharm.2016.02.030. this issue. [DOI] [PubMed] [Google Scholar]
- Sublette ME, Galfalvy HC, Fuchs D, Lapidus M, Grunebaum MF, Oquendo MA, Mann JJ, Postolache TT. Plasma kynurenine levels are elevated in suicide attempters with major depressive disorder. Brain Behav Immun. 2011;25:1272–1278. doi: 10.1016/j.bbi.2011.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh HS, Zhao ML, Rivieccio M, Choi S, Connolly E, Zhao Y, Takikawa O, Brosnan CF, Lee SC. Astrocyte indoleamine 2,3-dioxygenase is induced by the TLR3 ligand poly(I:C): mechanism of induction and role in antiviral response. J Virol. 2007;81:9838–9850. doi: 10.1128/JVI.00792-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taniuchi H, Hayaishi O. Studies on the metabolism of kynurenic acid. J Biol Chem. 1963;238:283–293. [PubMed] [Google Scholar]
- Tavares RG, Tasca CI, Santos CE, Alves LB, Porciuncula LO, Emanuelli T, Souza DO. Quinolinic acid stimulates synaptosomal glutamate release and inhibits glutamate uptake into astrocytes. Neurochem Int. 2002;40:621–627. doi: 10.1016/s0197-0186(01)00133-4. [DOI] [PubMed] [Google Scholar]
- Torrey EF, Bartko JJ, Lun ZR, Yolken RH. Antibodies to Toxoplasma gondii in patients with schizophrenia: a meta-analysis. Schizophr Bull. 2007;33:729–736. doi: 10.1093/schbul/sbl050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torrey EF, Bartko JJ, Yolken RH. Toxoplasma gondii and other risk factors for schizophrenia: an update. Schizophr Bull. 2012;38:642–647. doi: 10.1093/schbul/sbs043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tse YC, Bagot RC, Hutter JA, Wong AS, Wong TP. Modulation of synaptic plasticity by stress hormone associates with plastic alteration of synaptic NMDA receptor in the adult hippocampus. PLoS One. 2011;6:e27215. doi: 10.1371/journal.pone.0027215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turski MP, Turska M, Zgrajka W, Bartnik M, Kocki T, Turski WA. Distribution, synthesis, and absorption of kynurenic acid in plants. Planta Med. 2011;77:858–864. doi: 10.1055/s-0030-1250604. [DOI] [PubMed] [Google Scholar]
- Uwai Y, Honjo H, Iwamoto K. Interaction and transport of kynurenic acid via human organic anion transporters hOAT1 and hOAT3. Pharmacol Res. 2012;65:254–260. doi: 10.1016/j.phrs.2011.11.003. [DOI] [PubMed] [Google Scholar]
- van der Goot AT, Nollen EA. Tryptophan metabolism: entering the field of aging and age-related pathologies. Trends Mol Med. 2013;19:336–344. doi: 10.1016/j.molmed.2013.02.007. [DOI] [PubMed] [Google Scholar]
- van der Goot AT, Zhu W, Vazquez-Manrique RP, Seinstra RI, Dettmer K, Michels H, Farina F, Krijnen J, Melki R, Buijsman RC, Ruiz Silva M, Thijssen KL, Kema IP, Neri C, Oefner PJ, Nollen EA. Delaying aging and the aging-associated decline in protein homeostasis by inhibition of tryptophan degradation. Proc Natl Acad Sci U S A. 2012;109:14912–14917. doi: 10.1073/pnas.1203083109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicente MI, Costa PF, Lima PA. Galantamine inhibits slowly inactivating K+ currents with a dual dose-response relationship in differentiated N1E-115 cells and in CA1 neurones. Eur J Pharmacol. 2010;634:16–25. doi: 10.1016/j.ejphar.2010.02.021. [DOI] [PubMed] [Google Scholar]
- Vidal C, Li W, Santner-Nanan B, Lim CK, Guillemin GJ, Ball HJ, Hunt NH, Nanan R, Duque G. The kynurenine pathway of tryptophan degradation is activated during osteoblastogenesis. Stem Cells. 2015;33:111–121. doi: 10.1002/stem.1836. [DOI] [PubMed] [Google Scholar]
- Vigneault P, Bourgault S, Kaddar N, Caillier B, Pilote S, Patoine D, Simard C, Drolet B. Galantamine (Reminyl) delays cardiac ventricular repolarization and prolongs the QT interval by blocking the HERG current. Eur J Pharmacol. 2012;681:68–74. doi: 10.1016/j.ejphar.2012.02.002. [DOI] [PubMed] [Google Scholar]
- Voigt JP, Fink H. Serotonin controlling feeding and satiety. Behav Brain Res. 2015;277:14–31. doi: 10.1016/j.bbr.2014.08.065. [DOI] [PubMed] [Google Scholar]
- Wang B, Koga K, Osuga Y, Cardenas I, Izumi G, Takamura M, Hirata T, Yoshino O, Hirota Y, Harada M, Mor G, Taketani Y. Toll-like receptor-3 ligation-induced indoleamine 2, 3-dioxygenase expression in human trophoblasts. Endocrinology. 2011;152:4984–4992. doi: 10.1210/en.2011-0278. [DOI] [PubMed] [Google Scholar]
- Wang J, Simonavicius N, Wu X, Swaminath G, Reagan J, Tian H, Ling L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J Biol Chem. 2006;281:22021–22028. doi: 10.1074/jbc.M603503200. [DOI] [PubMed] [Google Scholar]
- Wang YH, Bosy TZ, Yasuda RP, Grayson DR, Vicini S, Pizzorusso T, Wolfe BB. Characterization of NMDA receptor subunit-specific antibodies: distribution of NR2A and NR2B receptor subunits in rat brain and ontogenic profile in the cerebellum. J Neurochem. 1995;65:176–183. doi: 10.1046/j.1471-4159.1995.65010176.x. [DOI] [PubMed] [Google Scholar]
- Wonodi I, Stine OC, Sathyasaikumar KV, Roberts RC, Mitchell BD, Hong LE, Kajii Y, Thaker GK, Gunvant K, Schwarcz R. Downregulated kynurenine 3-monooxygenase gene expression and enzyme activity in schizophrenia and genetic association with schizophrenia endophenotypes. Arch Gen Psych. 2011;68:665–674. doi: 10.1001/archgenpsychiatry.2011.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wonodi I, McMahon RP, Krishna N, Mitchell BD, Liu J, Glassman M, Hong LE, Gold JM. Influence of kynurenine 3-monooxygenase (KMO) gene polymorphism on cognitive function in schizophrenia. Schizophr Res. 2014;160:80–87. doi: 10.1016/j.schres.2014.10.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood K, Harwood J, Coppen A. The effect of antidepressant drugs on plasma kynurenine in depressed patients. Psychopharmacology (Berl) 1978;59:263–266. doi: 10.1007/BF00426632. [DOI] [PubMed] [Google Scholar]
- Wu HQ, Okuyama M, Kajii Y, Pocivavsek A, Bruno JP, Schwarcz R. Targeting kynurenine aminotransferase II in psychiatric diseases: promising effects of an orally active enzyme inhibitor. Schizophr Bull. 2014;40(Suppl 2):S152–158. doi: 10.1093/schbul/sbt157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu W, Nicolazzo JA, Wen L, Chung R, Stankovic R, Bao SS, Lim CK, Brew BJ, Cullen KM, Guillemin GJ. Expression of tryptophan 2,3-dioxygenase and production of kynurenine pathway metabolites in triple transgenic mice and human Alzheimer’s disease brain. PLoS One. 2013;8:e59749. doi: 10.1371/journal.pone.0059749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie FT, Cao JS, Zhao J, Yu Y, Qi F, Dai XC. IDO expressing dendritic cells suppress allograft rejection of small bowel transplantation in mice by expansion of Foxp3+ regulatory T cells. Transpl Immunol. 2015;33:69–77. doi: 10.1016/j.trim.2015.05.003. [DOI] [PubMed] [Google Scholar]
- Xie G, Zhan J, Tian Y, Liu Y, Chen Z, Ren C, Sun Q, Lian J, Chen L, Ruan J, Ye C, Sun A, Yuan Y. Mammosphere cells from high-passage MCF7 cell line show variable loss of tumorigenicity and radioresistance. Cancer Lett. 2012;316:53–61. doi: 10.1016/j.canlet.2011.10.018. [DOI] [PubMed] [Google Scholar]
- Yolken RH, Torrey EF. Are some cases of psychosis caused by microbial agents? A review of the evidence Mol Psychiatry. 2008;13:470–479. doi: 10.1038/mp.2008.5. [DOI] [PubMed] [Google Scholar]
- Yuen EY, Wei J, Liu W, Zhong P, Li X, Yan Z. Repeated stress causes cognitive impairment by suppressing glutamate receptor expression and function in prefrontal cortex. Neuron. 2012;73:962–977. doi: 10.1016/j.neuron.2011.12.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwilling D, Huang SY, Sathyasaikumar KV, Notarangelo FM, Guidetti P, Wu HQ, Lee J, Truong J, Andrews-Zwilling Y, Hsieh EW, Louie JY, Wu T, Scearce-Levie K, Patrick C, Adame A, Giorgini F, Moussaoui S, Laue G, Rassoulpour A, Flik G, Huang Y, Muchowski JM, Masliah E, Schwarcz R, Muchowski PJ. Kynurenine 3-monooxygenase inhibition in blood ameliorates neurodegeneration. Cell. 2011;145:863–874. doi: 10.1016/j.cell.2011.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]