

SUMMARY
Experiences during early development can influence neuronal functions and modulate adult behaviors [1, 2]. However, the molecular mechanisms underlying the long-term behavioral effects of these early experiences are not fully understood. The C. elegans ascr#3 (asc-0394C9, C9) pheromone triggers avoidance behavior in adult hermaphrodites [3–7]. Here, we show that hermaphrodites that are briefly exposed to ascr#3 immediately after birth exhibit increased ascr#3-specific avoidance as adults indicating that ascr#3-experienced animals form a long lasting memory or imprint of this early ascr#3 exposure [8]. Ascr#3 imprinting is mediated by increased synaptic activity between the ascr#3-sensing ADL neurons and their post-synaptic SMB motor neuron partners via increased expression of the odr-2 GPI-linked signaling gene in the SMB neurons. Our study suggests that the memory for early ascr#3 experience is imprinted via alteration of activity of a single synaptic connection, that in turn shapes experience-dependent plasticity in adult ascr#3 responses.
Keywords: sensory imprinting, pheromone, synapse, neuronal activity, GPI-anchored protein
RESULTS
Adult C. elegans transiently exposed to ascr#3 during larval stages exhibits increased acsr#3 pheromone avoidance
To assess whether early experience of pheromones affects ascr#3 avoidance behavior in adult worms, we transiently exposed first larval stage (L1) wild-type worms to ascr#3 and examined their responses to ascr#3 as adults (Figure 1A and see STAR methods). To test avoidance, a pheromone diluted in buffer was applied to a freely moving animal and the fraction of animals that reverse was calculated (drop test assay; See STAR Methods) [9]. Pre-exposure to 600 nM ascr#3 during the L1 stage significantly improved adult avoidance of ascr#3 at concentrations that elicit only weak avoidance by adults that were not similarly pre-exposed, but did not further increase avoidance at higher ascr#3 concentrations (Figure 1B). Increased ascr#3 avoidance appeared to be mediated by increased long reversals and omega turns but not short reversals in pre-exposed animals (Figure S1A). Henceforth, we calculate increased avoidance as a learning index (LI: pre-exposed reversing rate minus naive reversing rate/100) (Figure 1C).
Figure 1. Transient exposure to the ascr#3 pheromone after birth specifically enhances ascr#3 avoidance behavior of adults.
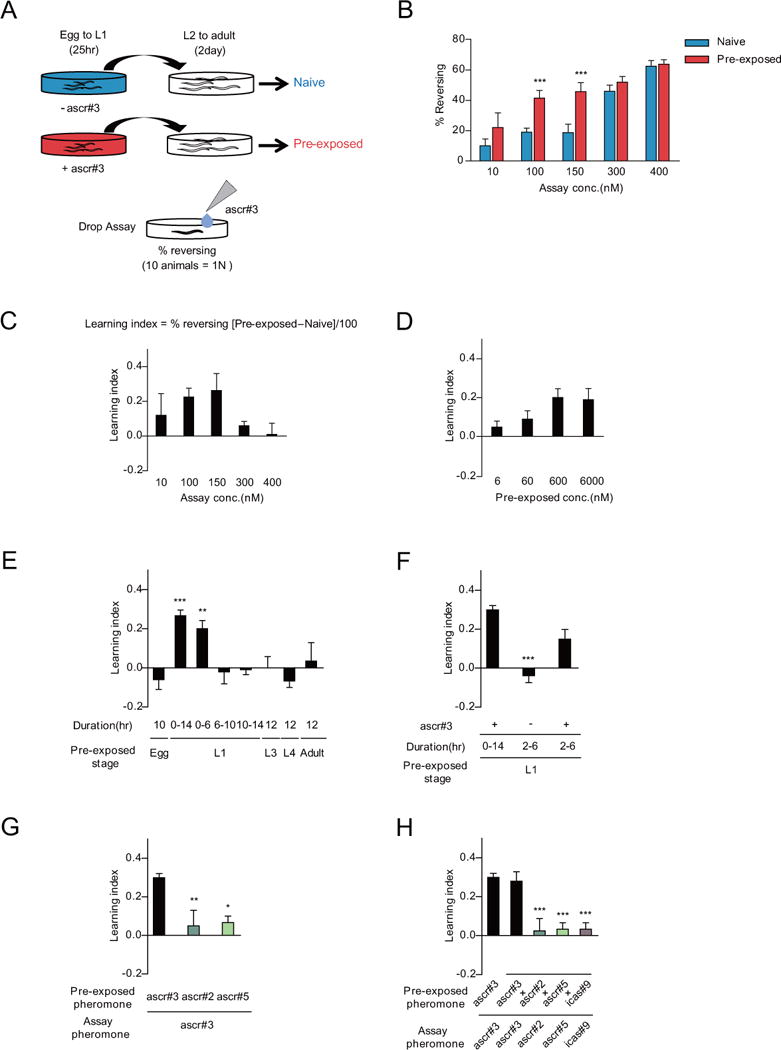
A. Experimental scheme of pheromone imprinting assay. The animals are exposed to dH2O (naive: shown in blue) and ascr#3 diluted with dH2O (pre-exposed: shown in red) from egg to the L1 stage. The percentage of reversal is calculated by measuring avoidance frequencies to ascr#3 exposure at the adult stage.
B and C. Percentage of reversal (B) and learning index (C) of naive and pre-exposed adult animals to 10 nM, 100 nM, 150 nM, 300 nM, and 400 nM ascr#3. Learning index is calculated by the percentage of the reversal rate of pre-exposed minus the reversal rate of naive divided by 100. *** indicates different ascr#3 avoidance from naive at p<0.001 by one-way ANOVA with Bonferroni’s post hoc test. n=50–200 each.
D. Learning index of adult animals pre-exposed to 6 nM, 60 nM, 600 nM and 6000 nM ascr#3 at L1. n=80–200 each.
E. Learning index of adult animals pre-exposed to 600 nM ascr#3 at different developmental stages. Exposure times during the specific developmental stage are indicated as egg (10 hours), L3 (12 hours), L4 (12 hours) and adult (12 hours). For the L1 stage, exposure times are indicated as between starting and ending time of pheromone exposure. ** and *** indicate different from egg (10 hours) at p<0.01 and p<0.001 by one-way ANOVA with Dunnett’s post hoc test, respectively. n=30–190 each.
F. Learning index of adult animals exposed to ascr#3 for 4 hours in the L1 stage. Pre-exposure for 2~6 hour in the L1 stage is sufficient for ascr#3 imprinting. *** indicates different from ascr#3 pre-exposure for 14 hours at p<0.001 by one-way ANOVA with Dunnett’s post hoc test. n=80–120 each.
G and H. Learning index of adult animals pre-exposed and/or assayed with other pheromone components. Animals are pre-exposed to either 600 nM ascr#3, ascr#2 or ascr#5 and assayed with 100 nM ascr#3 (G) and pre-exposed to pheromone mixture containing 600 nM ascr#3, ascr#2, ascr#5 and icas#9, and assayed with 100 nM ascr#3, ascr#2, ascr#5 or icas#9 (H). *, ** and *** indicate different from the control (ascr#3 pre-exposed and ascr#3 assayed) at p<0.05, p<0.01 and p<0.001, respectively, by one-way ANOVA with Dunnett’s post hoc test. (G) n=30–100 each. (H) n=30–50 each. (B–H) Error bars represent SEM.
See also Figure S1.
We next defined the optimal ascr#3 concentration that elicits the pre-exposure effect. Animals exhibited maximal adult ascr#3 avoidance when they were pre-exposed to ascr#3 concentrations higher than 600 nM (Figure 1D) [4]. Although exposure of L1 larvae to ascr#3 pheromone also promotes entry into the alternate dauer developmental stage, we note that the conditions used for imprinting are distinct from those used for dauer induction, and few if any ascr#3-imprinted animals enter into the dauer stage. These results indicate that early experience of ascr#3 pheromone appears to be translated into behavioral changes in the adult stage of hermaphrodites, suggesting that ascr#3-experienced animals form a long-lasting memory or imprint for ascr#3. Wild-type C. elegans males exhibit neutral responses to 100 nM ascr#3 [7, 10]. We found that ascr#3-experienced adult males continue to exhibit neutral responses to 100 nM ascr#3 (Figure S1B), indicating that early ascr#3 experience appears to increase ascr#3 avoidance only by adult hermaphrodites but not by adult males.
The memory for ascr#3 is acquired during the L1 larval stage
To determine whether the long-lasting memory for ascr#3 is formed during a specific developmental time window, worms were pre-exposed to ascr#3 at multiple developmental stages and their ascr#3 responses were assessed as adults. All assays were conducted by pre-exposing animals to 600 nM pheromone and assessing responses to 100 nM pheromone, unless noted otherwise.
Compared to the L1 stage, ascr#3 pre-exposure at the L3, L4, or adult stages did not enhance ascr#3 avoidance in adults (Figure 1E), suggesting that the L1 stage is critical for acquiring the ascr#3 memory. We could not test the L2 stage because of complexity with dauer formation [11]. We further defined the time window within the L1 stage necessary for this behavior, and found that pre-exposure in a defined time period during the early L1 stage (up to 6 hours following hatching at 20 °C) appears to represent the critical period for formation of the ascr#3 memory (Figure 1E). More specifically, exposure of ascr#3 to worms at 2–6 hour after birth appears to be necessary and sufficient for the increased ascr#3 avoidance (Figure 1F).
C. elegans secretes additional pheromones, including ascr#2, ascr#5, and icas#9. Similar to ascr#3, these pheromones are also potent inducers of entry into the alternate dauer developmental stage under limiting food conditions [4]. Pre-exposure to 600 nM ascr#2 or ascr#5 alone did not affect ascr#3 avoidance by adults, suggesting that pheromone imprinting does not reflect a memory of general unfavorable dauer inducing conditions but specifically encodes experience of the ascr#3 pheromone (Figure 1G). Pre-exposure to mixtures of ascr#2, ascr#5, and icas#9 with ascr#3 also did not affect ascr#3 imprinting, further supporting that imprinting is specific to ascr#3 (Figure 1H). Moreover, pre-exposure to ascr#2 did not result in altered adult avoidance of ascr#2 (Figure S1C), further confirming specificity of this process to ascr#3. In addition, pre-exposure to ascr#3 did not affect high-osmolality glycerol avoidance (Figure S1D), suggesting that early ascr#3 experience does not affect broad avoidance behaviors to other repulsive chemicals by adults. However, we are unable to exclude the possibility that imprinting by other ascarosides may occur at developmental stages other than the L1 stage.
Since the memory for a specific pheromone component is acquired only at the critical developmental period, we further define this altered ascr#3 avoidance as sensory imprinting [8]. Interestingly, when animals recovered from the developmentally arrested dauer stage that was induced by limited food supply in the presence of pheromones including ascr#3 at the L1 and L2 stages, their ascr#3 avoidance was comparable to or even weaker than that of naive animals (Figure S1E) [11]. These data indicate that the ascr#3 imprint appears to be removed following dauer experience and/or ascr#3 imprinting requires non dauer-inducing conditions at the L1 stage. Moreover, it is likely that passage through the dauer stage results in genome-wide changes in gene expression patterns [12, 13], which may mask or erase ascr#3 imprinting phenotypes.
We next investigated perdurance of the ascr#3 memory. We found that pre-exposed 3 or 6 day-old adults still exhibited increased ascr#3 avoidance (Figure S1F). We also asked whether memories are transmitted to the next generation but found that progeny from imprinted mothers did not exhibit improved ascr#3 avoidance (Figure S1G). These results indicate that the ascr#3 imprint lasts with aging but is not inherited under these assay conditions.
odr-2 acts in the SMB neurons to increase ascr#3 avoidance in ascr#3 imprinted animals
We next performed a candidate gene search to identify genes and molecules required for ascr#3 imprinting. We first tested genes that have been shown to be required for distinct forms of learning and memory in C. elegans. These include the egl-4 (cyclic GMP-dependent protein kinase) gene implicated in sensory adaptation, the casy-1 (calsyntenin) gene involved in associative learning, and the sra-11 (7-TM G protein-coupled receptor), ttx-3 (LIM homeodomain protein), or tdc-1 (tyrosine decarboxylase) genes involved in olfactory imprinting [14–17]. However, loss of function of these genes did not affect ascr#3 imprinting (Figure S2A).
In the course of analyzing mutants defective in odorant responses, we found that odr-2 (n2145) [18, 19] mutants exhibited defects in ascr#3 imprinting although the ability of these mutants to avoid ascr#3 was unaffected (Figure 2A, 2B). The ascr#3 imprinting defects in odr-2 mutants were fully rescued upon expression of wild-type odr-2 cDNA driven under its upstream regulatory sequences (Figure 2C). The odr-2 gene encodes a membrane-associated protein related to the Ly-6 (leukocyte antigen-6) superfamily of GPI (glycosylated phosphatidylinositol)-linked proteins, and odr-2 mutants have previously been shown to exhibit decreased chemotaxis towards a set of volatile attractive chemicals [19]. odr-2 is expressed in a set of head neurons including RIG, RME and SMB [19, 20]. To determine where ODR-2 acts to regulate ascr#3 imprinting, we tested transgenic animals expressing odr-2 wild-type sequences for rescue of the ascr#3 imprinting defects. We found that while expression of odr-2 in RIG and RME under the control of odr-2 (−377) promoter [20] did not rescue these behavioral defects, ascr#3 imprinting defects were fully restored upon expression of odr-2 exclusively in SMB under the control of flp-12 (−339) promoter (Figure 2D) [20], indicating that ODR-2 acts in the SMB neurons to mediate ascr#3 imprinting.
Figure 2. The glycosylated phosphatidylinositol-linked protein, ODR-2, is required for increased ascr#3 avoidance in ascr#3 imprinted animals.
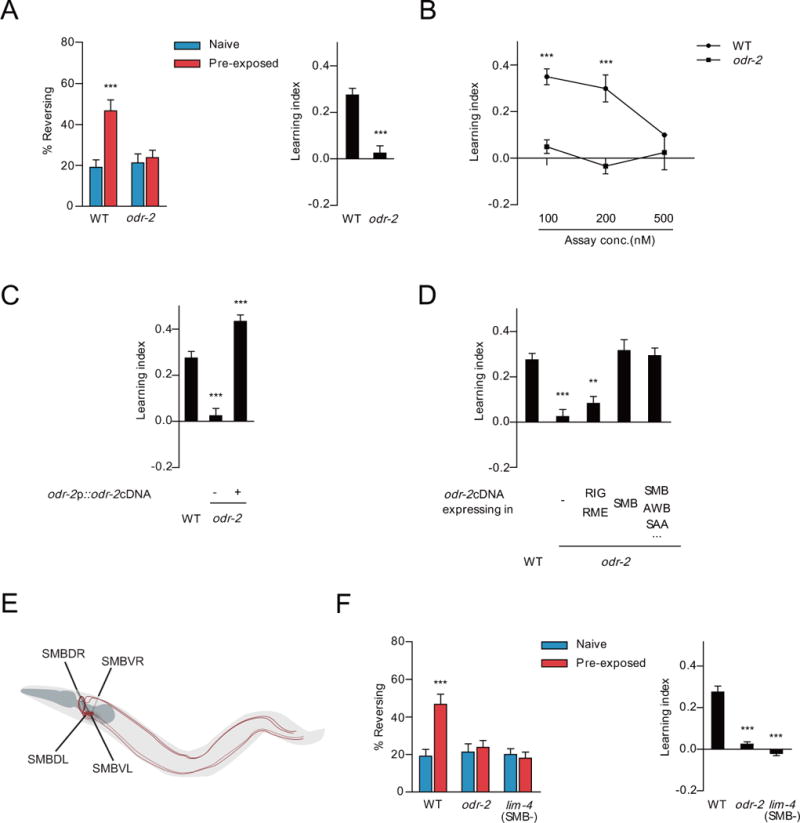
A. Percentage of reversal (left) and learning index (right) in wild-type and odr-2 mutants. *** indicates significantly different from naive (left) and WT (right) at p<0.001 by one-way ANOVA with Bonferroni’s post hoc test (left) and by Student’s t-test (right), respectively. n=80–120 each.
B. Learning index of wild-type and odr-2 mutants at 100 nM, 200 nM or 500 nM ascr#3. *** indicates different from WT at p<0.001 by one-way ANOVA with Bonferroni’s post hoc test. n=30–60 each.
C and D. Learning index of wild-type and odr-2 mutants expressing odr-2 cDNA under the control of odr-2 promoter (RIG, RME and SMB) (C) or cell specific promoters including odr-2 (−377) (RIG and RME), flp-12 (−339) (SMB), and lim-4 (AWB, SAA, RID, RIV, RMD and SMB) (D). ** and *** indicate different from WT at p<0.01 and p<0.001, respectively, by one-way ANOVA with Dunnett’s post hoc test. (C) n=80–120 each. (D) n=60–140 each.
E. Schematic diagram of the SMB neurons. A pair of dorsal and ventral cell bodies (SMBDs and SMBVs) are located in the head and their processes innervate head muscle and run posteriorly.
F. Percentage of reversal of naive and pre-exposed animals (left) and learning index (right) in wild-type, odr-2, and lim-4 mutants. *** indicates different from naive (left) and WT (right) at p<0.001 by one-way ANOVA with Bonferroni’s (left) and Dunnett’s post hoc tests (right), respectively. n=80–120 each. All error bars represent SEM.
See also Figure S2.
The SMB neurons consist of two left and right pairs (dorsal or ventral) of sensory/inter/motor neuron types that are located in the head, and that innervate the head and neck muscles (Figure 2E) [21]. While their synapse-free processes extend along the ventral or dorsal sublateral cords to the tail, the SMB neurons have extensive electric and chemical synaptic contacts to other neurons in the head [21]. These neurons regulate head locomotion [20–22]. However, their roles in chemosensory behaviors have not been explored. We next investigated that the SMB neurons are required for ascr#3 imprinting. To address this issue, we examined lim-4 mutants in which the SMB neurons are not fully differentiated and in which the functions of SMB are completely abolished [20]. lim-4 mutants had been shown to exhibit additional defects including neuronal specification [23]. While lim-4 mutants were still able to avoid ascr#3, pre-exposure to ascr#3 did not enhance ascr#3 avoidance in lim-4 mutants (Figure 2F). These results together with odr-2 rescue results support that the SMB neurons may play an important role in ascr#3 imprinting.
Ascr#3-induced responses in the ADL chemosensory neurons are unaltered in ascr#3 imprinted worms
In adults, ascr#3 elicits avoidance behavior in hermaphrodites via the nociceptive ADL chemosensory neurons (Figure 3A) [7]. To describe the neuronal basis of pheromone imprinting, we first monitored intracellular Ca2+ dynamics in response to ascr#3 in transgenic animals expressing the genetically encoded calcium sensor GCaMP3 in the ascr#3-sensing ADL neurons. The ADL neurons exhibit a rapid and transient Ca2+ increase upon exposure to nano-molar concentrations of ascr#3 (Figure 3B) [7]. Both naive animals and ascr#3-imprinted animals displayed similar Ca2+ transients in adult ADL neurons upon addition of ascr#3 (Figure 3B). These results suggest that increased ascr#3 avoidance is not due to enhanced sensory responsiveness of ADL. Furthermore, consistent with the normal ascr#3 avoidance behavior of odr-2 mutants, the ADL Ca2+ response to ascr#3 was not altered in odr-2 mutants (Figure 3C).
Figure 3. Ca2+ response to acute ascr#3 exposure is not altered in the ascr#3-sensing ADL neurons and their downstream command interneurons of ascr#3 imprinted animals.
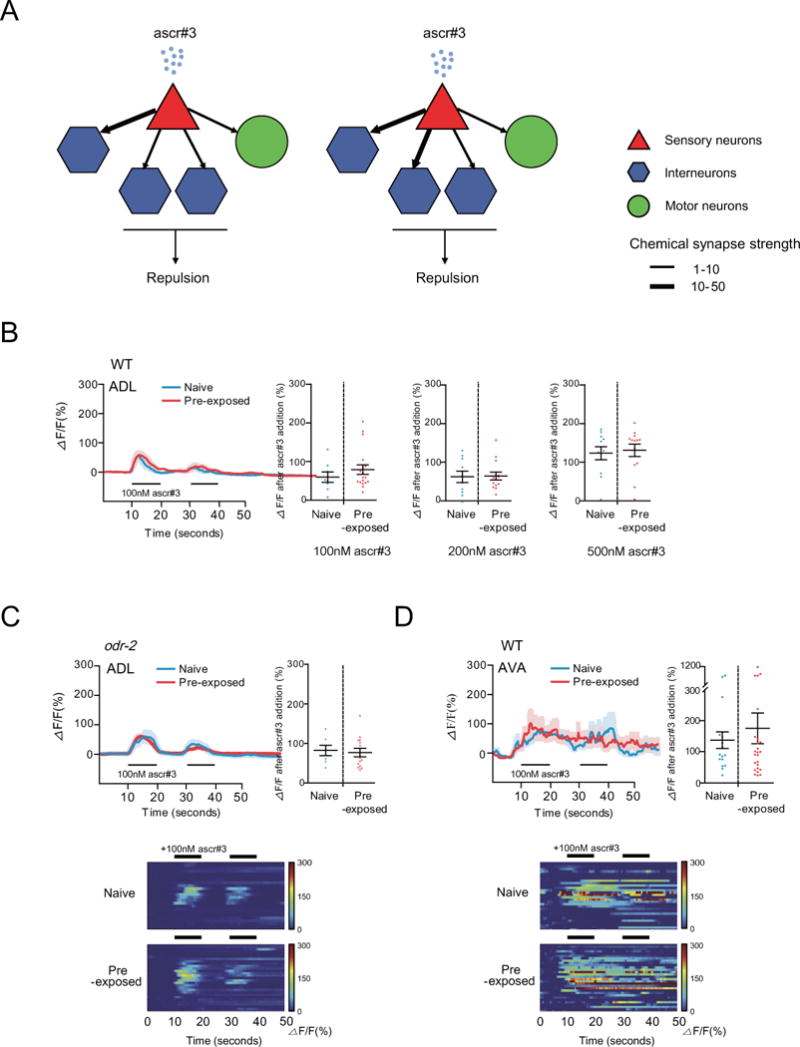
A. Post-synaptic connections of the ADLL and ADLR neurons. The AIB, AVD, AVA and SMBV neurons are post-synaptic to ADL. Chemical synapses between sensory (triangles), inter-(hexagons) and motor (circles) are indicated as arrows. Synaptic strength is indicated in thickness of lines (www.wormwiring.org).
B. Ca2+ transients of ADL in response to 100 nM ascr#3 exposure. The average traces of Ca2+ responses during two pulses of 100 nM ascr#3 (left) and the maximum value of Ca2+ responses to 100 nM, 200 nM or 500 nM ascr#3 (right) of naive (shown in blue) or pre-exposed animals (shown in red) in 50 seconds are shown. n=8 (naive) and 18 (pre-exposed) each.
C. Ca2+ transients of ADL in response to 100 nM ascr#3 exposure in odr-2 mutants. The average traces of Ca2+ responses during two pulses of 100 nM ascr#3 (left), heat map of individual Ca2+ responses to ascr#3 (bottom), and the maximum value of Ca2+ responses to 100 nM ascr#3 (right) in naive (shown in blue) or pre-exposed (shown red) odr-2 mutants are shown. n=7 (naive) and 14 (pre-exposed) each.
D. Ca2+ transients of AVA in response to 100 nM ascr#3 exposure. The average traces of Ca2+ responses during two pulses of 100 nM ascr#3 (left), heat map of individual Ca2+ responses to ascr#3 (bottom), and the maximum value of Ca2+ responses to 100 nM ascr#3 (right) in naive (shown in blue) or pre-exposed animals (shown in red) are shown. n=7 (naive) and 8 (pre-exposed) each. All error bars represent SEM.
See also Figure S3.
The ADL sensory neurons drive ascr#3 avoidance through their chemical synapses [7]. Inspection of the anatomical wiring data from the C. elegans connectome suggests that ascr#3 signals from the ADL neurons are transmitted to a few major postsynaptic target neurons including AIB (1st layer interneurons), and the AVA and AVD backward command interneurons (Figure 3A). Previous studies have shown that activity in AIB, AVA and AVD is increased during backward movement or reversals [24]. We found that while the AIB or AVD neurons exhibited noisy but low levels of Ca2+ responses upon exposure to ascr#3 (Figure S3A, S3B), the AVA cell bodies responded strongly and consistently to repeated ascr#3 addition with increased Ca2+ levels (Figure 3D). However, these responses were unaltered upon ascr#3 imprinting (Figure 3D, Figure S3A, S3B). These results imply that ascr#3 signals in the ADL neurons may be transmitted mainly via AVA, but that imprinting does not enhance ascr#3 avoidance via this ADL-AVA circuit.
The SMB neurons mediate increased ascr#3 avoidance in ascr#3 imprinted animals
The C. elegans connectome data indicate that the SMBV neurons are additional post-synaptic partners of the ADL sensory neurons (Figure 3A) although the synaptic strength appears to be weak. Since the SMB neurons appear to play a role in ascr#3 imprinting, we then examined how SMB is involved in ascr#3 avoidance behavior.
We first expressed GCaMP3 in SMB under the control of flp-12 (−339) promoter that drives transgene expression in all four SMB neurons [20]. We note that a SMBV-specific promoter is not currently available. Moreover, since the cell bodies of SMBD and SMBV are located in close proximity, differentiating among these neurons is challenging [20]. We thus examined ascr#3 responses in all SMB neurons together. Acute ascr#3 exposure did not elicit Ca2+ transients in the SMB neurons of naive control animals (Figure 4A, Movie S1). However, we detected consistent and robust Ca2+ transients in response to acute ascr#3 exposure in ascr#3 imprinted animals (Figure 4A, Movie S2). Since SMB may detect body muscle contractions that could be responsible for the observed Ca2+ responses in SMB, we paralyzed worms with the nicotinic acetylcholine receptor agonist levamisole. Levamisole-treated ascr#3 imprinted animals still exhibited similar or even enhanced Ca2+ dynamics upon ascr#3 exposure (Figure S4A).
Figure 4. The SMB motor neurons mediate enhanced ascr#3 avoidance in ascr#3 imprinted animals.
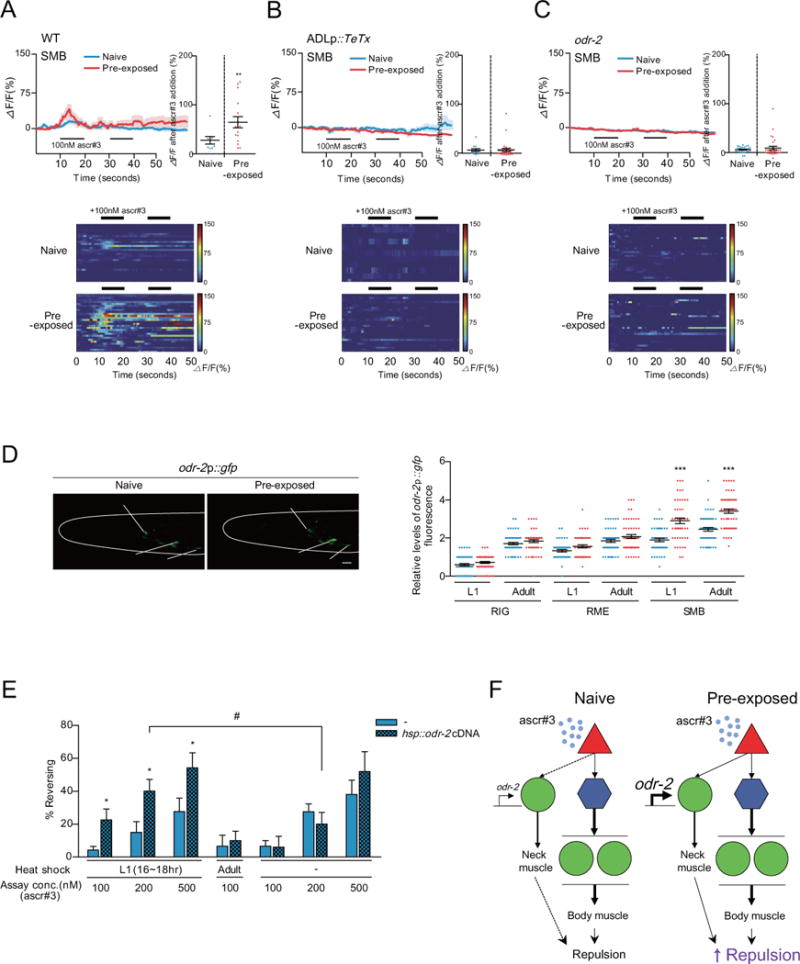
A–C. Ca2+ transients of the SMB neurons in response to 100 nM ascr#3 in wild-type animals (A), wild-type animals expressing ADLp∷TeTx (B) or odr-2 mutants (C). The average traces of Ca2+ responses during two pulses of 100 nM ascr#3 (left), heat map of individual Ca2+ responses to ascr#3 (bottom), and the maximum value of Ca2+ responses to 100 nM ascr#3 (right) in naive (blue) or pre-exposed animals [10] are shown. ** indicates different from naive at p<0.01 by Student’s t-test. n=8–30 each.
D. Representative images (left) and GFP quantification (right) of odr-2p∷gfp expression in RIG, RME, and SMB of naive (shown in blue) and pre-exposed (shown in red) animals at the L1 or adult stages are shown. The images were taken of adults. *** indicates different from WT at p<0.001 by one-way ANOVA with Bonferroni’s post hoc test. n=50–60 each. Scale bar: 10 m.
E. Percentage of reversal of adult transgenic animals expressing hsp∷odr-2 cDNA under heat shock at the L1 and adult stages. * indicates different from hsp∷odr-2 cDNA (−) and # indicates different from no heat shock at p<0.05 by Student’s t-test. n=50–70 each. (A–E) Error bars represent SEM.
F. Model for circuit mechanisms underlying ascr#3 avoidance in naive or ascr#3 imprinted animals. In naive animals, ADL detects ascr#3 and transmits the signals to the post-synaptic AVA command interneurons and downstream VA/DA motor neurons. In pre-exposed animals, the increased expression of odr-2 in SMB initiates synaptic transmission from ADL to SMB that mediate ascr#3 signal transmission from ADL not only to AVA but SMB. SMB activation may serve to sensitize background for backward movement via excitation of head muscles.
See also Figure S4.
To test whether the observed Ca2+ transients in SMB in ascr#3 imprinted animals are transmitted from ADL, we examined Ca2+ responses in transgenic animals expressing TeTx (tetanus toxin light chain) specifically in the ADL neurons to block synaptic transmission from ADL [7]. Ca2+ transients of SMB in ascr#3 imprinted animals were strongly suppressed by blocking ADL synaptic transmission (Figure 4B, Figure S4B), suggesting that ascr#3 imprinting results in enhanced ascr#3 signal transmission from ADL to SMB. These results indicate that ascr#3 imprinting specifically alters the ADL-SMB synaptic transmission.
Moreover, the ascr#3 imprinting-dependent Ca2+ response in SMB was completely abolished in odr-2 mutants (Figure 4C), indicating that odr-2 elicits increased activity of SMB to ascr#3 exposure in imprinted animals by enhancing the ADL-SMB synaptic transmission.
Upregulation of odr-2 expression of L1 larvae is sufficient for increased ascr#3 avoidance in adults
To further investigate the role of odr-2 in mediating increased SMB activity in of ascr#3 imprinted animals, we monitored odr-2 expression in SMB and found that odr-2 expression in SMB was increased after transient exposure to ascr#3 at the L1 stage and was maintained through adulthood (Figure 4D). In contrast, odr-2p∷gfp expression was unaffected in other odr-2-expressing cells including RIG and RME (Figure 4D). To examine whether upregulation of odr-2 at the L1 stage is sufficient for the increased ascr#3 avoidance in adults, we expressed odr-2 cDNA in wild-type animals with the inducible expressed heat-shock promoter. Compared to animals with no heat-shock treated or those heat-shocked only at the adult stage, transient induction of odr-2 gene activity at the L1 larval stage was sufficient to increase ascr#3 avoidance in adults (Figure 4E). Thus far, our results are consistent with a model in which the increased odr-2 expression in SMB of the ascr#3 imprinted animals causes functional changes in the ADL-SMB synaptic activities.
DISCUSSION
The well-defined nervous system of C. elegans with only 302 neurons mediates a broad spectrum of behaviors. However, these behaviors are plastic and can be modulated by the animal’s experience [25, 26]. For example, C. elegans is able to constantly monitor and adapt to their environment by forming short-term and/or long-term memories in their hard-wired neural circuits (See a review by [27]). C. elegans has also been shown to imprint long-term memories of environments, including odor or pathogen exposure experienced during development, via largely uncharacterized mechanisms [15, 17]. Here, we show a novel imprinting paradigm in C. elegans where exposure of newly hatched animals to the ascr#3 pheromone increases adult neuronal activity in response to ascr#3, resulting in altered behavior. Since the long-lasting memory for ascr#3 pheromone is formed during a critical period of early development and shapes the ascr#3 avoidance behavior of adults, this learning and long-term memory process for the pheromone could be referred to as sensory imprinting [8]. Sensory imprinting is a form of long-term memory that occurs in many animals: for example, homing of salmon and neonatal attachment of rodents [28, 29]. This type of behavioral plasticity could be essential for animal survival in natural environments. Ascr#3 is the most abundant and potent aversive pheromone in C. elegans hermaphrodites and may represent conditions of overcrowding [7, 30, 31]. Thus, newly hatched worms may pair this specific ascr#3 pheromone memory to their neuronal activity to promote searching behavior for more favorable environments as adults or to shape currently unidentified behavioral or physiological states for optimal fitness.
We previously showed that the ADL sensory neurons detect C. elegans pheromone ascr#3 and drive ascr#3 avoidance through their chemical synapses [7]. The ADL neurons directly connect to more than twenty postsynaptic neurons, and inspection of the anatomical wiring data including the number of synapses led us to hypothesize that ascr#3 signals from ADL are transmitted to the three major post-synaptic target neurons including the AIB, AVA and AVD interneurons. In this work, we have found that under non-ascr#3 pre-exposed (naive) conditions the ascr#3 signals are transmitted via the ADL-AVA synapses (Figure 4F). However, while transient exposure to ascr#3 at the L1 larval stage does not appear to affect this ADL-AVA synaptic transmission, this experience instead increases the ADL-SMB synaptic activities which are normally inactive in naive animals, and may account for the observed enhanced ascr#3 avoidance behavior in adult worms (Figure 4F).
The conclusion that the SMB neurons are the critical site for mediating ascr#3 imprinting is supported by these findings: 1) The SMB neurons respond to ascr#3 exposure in ascr#3-imprinted but not in naive animals; 2) Expression of odr-2 that is necessary and partly sufficient for ascr#3 imprinting is increased specifically in SMB of ascr#3-imprinted animals; 3) lim-4 mutants in which SMB functions are abolished fail to imprint ascr#3 experience. Laser ablation of the SMB motor neurons causes increased reversal frequency and wave amplitude of forward locomotion [20, 22]. Since the SMB neurons innervate head and neck muscles, and ascr#3 exposure now activates the SMB neurons in ascr#3 imprinted animals, our results suggest that early ascr#3 experience causes the SMB neurons to sensitize to subsequent ascr#3 exposure and modulate backward movement via the alteration of head/neck muscle activity (Figure 4F). For example, in addition to body muscle contraction triggered by ADL-AVA synaptic transmission, head/neck muscle contraction due to enhanced SMB neuronal activities upon ascr#3 imprinting may increase reversal rates further [24, 32] or the SMB neurons could directly activate backward motor circuits via interneurons including the SAA neurons [21].
Our data show that odr-2 is essential for ascr#3 imprinting. odr-2 encodes a novel protein distantly related to murine Ly6 lymphocyte antigen protein that is tethered to membrane by a GPI (glycosylphosphatidylinositol) anchor [19]. In vertebrates, GPI anchored proteins have roles in development and neurogenesis by regulating several signaling pathways including Notch, Wnt or TGFβ [33]. Moreover, a set of Ly6 related genes including Lynx1 and Lynx2 have been shown to be expressed in the nervous system and appear to be accessory components of nicotinic acetylcholine receptors, thus modulating their functions [34, 35]. Our results indicate that the ODR-2 GPI anchored protein changes synaptic activity between ADL and SMB. ODR-2 proteins are localized on cell bodies and neuronal processes [36]. Thus, it is possible that ODR-2 may be co-localized with and regulate functions of neurotransmitter receptors in SMB.
Understanding how individual synapses are functionally or anatomically altered upon sensory imprinting is the essential first steps towards being able to dissect molecular and neuronal mechanisms underlying sensory imprinting and other forms of behavioral plasticity. Given that structures and functions of neural circuits are evolutionarily conserved, we expect that our work will lead to a general framework for understanding how circuits are modulated in higher animals including humans.
STAR METHODS
KEY RESOURCES TABLE
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Chemicals, Peptides, and Recombinant Proteins | ||
| Levamisole | Sigma | Cat# 16595-80-5 |
| ascr #3 | This paper | |
| Critical Commercial Assays | ||
| PDMS Sylgard 184 Silicone Elastomer Kit | Dow Corning | Product code: 1064291 |
| Experimental Models: Organisms/Strains : C. elegans strains | ||
| odr-2(n2145) V | Caenorhabditis Genetics Center | RRID:WB-STRAIN:CX2304 |
| odr-2(n2145) V; lskEx606[odr-2Δ3p∷odr-2cDNA] | This paper | N/A |
| odr-2(n2145) V; lskEx607[flp-12Δ3p∷odr-2cDNA] | This paper | N/A |
| odr-2(n2145) V ; lskEx608[lim-4p∷odr-2cDNA] | This paper | N/A |
| lim-4(ky403) X | Caenorhabditis Genetics Center | RRID:WB-STRAIN:CX3937 |
| lskEx218[sre-1p∷GCaMP3; unc-122p:dsRed] | This paper | N/A |
| odr-2(n2145) V; lskEx218[sre-1p∷GCaMP3; unc-122p:dsRed] | This paper | N/A |
| lskEx597[nmr-1p∷GCaMP3; unc-122p∷dsRed] | This paper | N/A |
| lskEx113[flp-12Δ3p∷GCaMP3; unc-122p∷dsRed] | This paper | N/A |
| lskEx611[sre-1p∷TeTx; unc-122p∷gfp]; lskEx113[flp12Δ3p∷GCaMP3] | This paper | N/A |
| odr-2(n2145) V; lskEx113[flp-12Δ3p∷GCaMP3] | This paper | N/A |
| lsk Is11[odr-2p∷gfp; unc-122p∷dsRed] | This paper | N/A |
| lskEx418[hsp∷odr-2cDNA; unc-122p∷gfp] | This paper | N/A |
| lskEx895[avr-14p∷gfp; unc-122p∷dsRed]; lskEx28[flp-12p∷mcherry; unc-122p∷gfp] | This paper | N/A |
| odr-2(n2145); lskEx895[avr-14p∷gfp; unc-122p∷dsRed]; lskEx28[flp-12p∷mcherry; unc-122p∷gfp] | This paper | N/A |
| egl-4(ks60) IV | Caenorhabditis Genetics Center | RRID:WB-STRAIN:FK223 |
| sra-11(ok630) II | Caenorhabditis Genetics Center | RRID:WB-STRAIN:RB816 |
| odr-3(n2150) V | Caenorhabditis Genetics Center | RRID:WB-STRAIN:CX2205 |
| odr-7(ky4) X | Caenorhabditis Genetics Center | RRID:WB-STRAIN:CX4 |
| odr-10(ky32) X | Caenorhabditis Genetics Center | RRID:WB-STRAIN:CX32 |
| casy-1(tm718) II | National BioResource Project | N/A |
| casy-1(pe401) II | Caenorhabditis Genetics Center | RRID:WB-STRAIN:JN414 |
| ttx-3(ks5) X | Caenorhabditis Genetics Center | RRID:WB-STRAIN:FK134 |
| tdc-1(n3419) II | Caenorhabditis Genetics Center | RRID:WB-STRAIN:MT13113 |
| lskEx610[npr-9p∷GCaMP3; unc-122p∷dsRed] | This paper | N/A |
| Software and Algorithms | ||
| AxioVision software | Zeiss | RRID:SCR_002677 |
| ZEN 2010 Lite Edition software | Zeiss | RRID:SCR_013672 |
| ImageJ | https://imagej.nih.gov/ij/ | RRID:SCR_003070 |
| MATLAB | MathWorks | RRID:SCR_001622 |
| Prism 5.0 | GraphPad Software | RRID:SCR_002798 |
| Adobe Illustrator CS6 | Adobe | RRID:SCR_014198 |
Contact for Reagent and Resource Sharing
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Dr. Kyuhyung Kim (khkim@dgist.ac.kr).
Experimental Model and Subject Details
C. elegans N2 strain was used as wild-type. Wild-type and transgenic animals were maintained at 20°C with abundant E. coli OP50 as food following standard conditions [37]. Day 1 (~24 hours after mid-L4 larval stage) young adult, hermaphrodite animals were used for experimentation, with the exception of the odr-2p∷gfp quantification study in which the L1 stage animals were also assayed (Figure 4D).
Method Details
Generation of C. elegans transgenic lines
odr-2 18a cDNA was amplified with a set of primers containing restriction sites, 5′-AgeI and 3′-NotI using PCR and inserted into pMC10 for performing the odr-2 rescue experiment [19]. Upstream regulatory sequences, including odr-2 (RIG, RME and SMB), odr-2 (−377) (RIG and RME), flp-12 (−339) (SMB), and lim-4 (AWB, SAA, RID, RIV, RMD and SMB), were fused with odr-2cDNA to drive its expression in specific neurons [20]. To generate the hsp∷odr-2cDNA transgene, the hsp16.2 promoter was amplified from the hsp16.2p∷unc-3cDNA transgene [38].
For Ca2+ imaging experiments of the AIB, AVD and AVA neurons, the promoter region of sre-1p∷GCaMP3 transgene was exchanged with npr-9 (AIB) or nmr-1 (AVD and AVA) promoters [7, 39]. nmr-1p was amplified from pCZGY1553 (a kind gift from Yishi Jin), and npr-9p was 2076 bp of upstream regulatory sequence amplified from genomic DNA by PCR. These promoters were ligated with GCaMP3 using the restriction sites, HindIII and BamHI.
Each transgenic C. elegans strain was generated by microinjection of the rescue construct (10 ng/μl) with unc-122p∷dsRed (50 ng/μl). The hsp∷odr-2cDNA construct was injected with the unc-122p∷gfp marker (50 ng/μl).
Pheromone imprinting
For pheromone imprinting, two types of 35 mm noble agar plates seeded with E. coli OP50 were made: the negative control plate contained distilled water (naive) and the experimental plate contained pheromones (pre-exposed). Figures 1G and 1H utilized experimental plates containing 600 nM ascr#3, ascr#2 and ascr#5, and 600 nM ascr#3, ascr#2, ascr#5 and icas#9, respectively. All other assays in this study used experimental plates containing 600 nM ascr#3 for imprinting. To pre-expose animals to ascr#3, five 1-day-old young adult hermaphrodites were placed onto each control and experimental plate for 2~3 hours until 30 to 50 eggs were laid. After 1 day (~26 hr), the naive or pre-exposed L1 stage animals were transferred onto 60 mm nematode growth media plates seeded with E. coli OP50 food and grown for 2.5 days until the young adult stage. For male imprinting experiments, we added ~20 males with L4 hermaphrodites for 1 day to mate and picked 5 adult hermaphrodites onto each naive and pre-exposed plates. For imprinting analysis of the next generation, 5 adults from naive or pre-exposed group (P0) were placed onto new control plates and allowed to lay 30~50 eggs (F1). The adults were then subsequently removed and the plates were incubated for 2.5 days at 20°C.
Post-dauer assay
Dauer formation assay was based on the protocol in [40]. 20 μl heat-killed OP50 was seeded onto 3.5 mm 600 nM ascr#3 noble agar plates. 5~10 young adults were placed and discarded when over 50 eggs were obtained. The eggs were grown at 25°C for 68~72hr. Animals in dauer formation were picked onto new OP50 seeded 60 mm plates and grown at 20°C for 2~3 days. We assayed ascr#3 avoidance with adult animals which were grown at 20°C as control.
Behavioral assay
The drop assay was performed as described with modifications [7]. All behavioral assays were performed in the absence of food and used 100 nM ascr#3 diluted in M13 buffer (30 mM Tris-HCl [pH 7.0], 100 mM NaCl, 10 mM KCl), except Figure S1D which 200 mM glycerol was used [41]. To measure the percent of animals reversing upon ascr#3 exposure, a young adult was transferred onto a plate without food and M13 buffer was dropped near the head of worm. If the worm failed to reverse in response to M13 after 10 seconds, ascr#3 was dropped onto the front of the head and avoidance behaviors (reversal) were monitored for 10 seconds. Short or long reversals were defined as reversals with fewer than two head bends or more than two head bends, respectively [42]. Long reversals were counted as repulsion in this study. The imprinting index was calculated as the percentage of reversals of pre-exposed animals minus percentage of reversals of naive animals. Each assay tested the avoidance response of 10 young adult hermaphrodite animals, and at least 3 independent trials were performed.
In vivo calcium imaging
Calcium imaging experiments were performed as described previously [7], using microfluidics chips that were produced in-house with a custom made chrome mask and master mold [43]. Briefly, PDMS Sylgard 184 Silicone Elastomer Kit (Dow Corning) was solidified on the master mold at 70°C for 2 hours, and then was attached on a 24 × 24 mm glass coverslip using CUTE plasma equipment (Air) (FEMTOSCIENCE). The C. elegans transgenic strains used for calcium imaging experiments included sre-1p∷GCaMP3 (ADL), flp-12 (−339)p∷GCaMP3 (SMB), npr-9p∷GCaMP3 (AIB) and nmr-1p∷GCaMP3 (AVA and AVD) were used [7]. Each animal was exposed to fluorescent light for 1~2 minutes and the images were captured under fixed exposure time (100ms) for 1 min with Zeiss Axioplan microscope using a 40X objective and a Zeiss AxioCam HR. The images were analyzed by AxioVision software, Image J, and custom-written scripts in MATLAB [7].
Levamizole treatment
25 mM levamisole (Sigma) was used to paralyze animals. 25 mM levamisole was diluted in M9 buffer from 0.25 M levamisole. The animals were placed into the solution for about 1 minute, during which pharyngeal pumping ceased, indicating paralysis.
Heat shock treatment
Heat shock treatment was performed as previously described [44]. After 16 hours from egg laying, animals were twice exposed to a 33°C heat shock for 30 minutes with a 1 hour recovery at 20°C in between. After the second heat shock treatment, the worms were incubated at 20°C until the adult stage and behavioral assays were performed (see above).
Quantification and Statistical analysis
Representative images
Images in Figures 4D and 4G were acquired using a Zeiss LSM700 Confocal microscope and Zeiss LSM780 Confocal microscope, respectively. The images were taken and analyzed through ZEN 2010 and Image J.
GFP quantification
For GFP quantification, the animals were anaesthetized by using 1 M NaN3 and placed on 2 % agarose pad with a 24 × 24 mm cover slip. odr-2p∷gfp strain was integrated using UV crosslinker (UVP). A minimum of 30 odr-2p∷gfp animals were exposed to 100 mM ascr#3 or control plates at the L1 larval stage following the imprinting protocol described above and quantified for GFP fluorescence. GFP expression was scored under 40X objective Zeiss Axioplan microscopy by observing the brightness of the neuron cell body and process. The animals were observed under 40X objective Zeiss Axioplan microscopy and LSM700 Confocal microscope. The images were taken and acquired by AxioVision or Zen 2010. All strains were assayed in parallel in two independent experiments.
Statistical tests
All of statistics was analyzed using Prism 5.0. When only two groups were compared, two tailed unpaired Student’s t-test was used. One-way ANOVA test was utilized to evaluate variation among more than two groups of datasets. The post hoc tests used include Dunnett’s test for multiple comparisons obtained at the same experimental condition, and Bonferroni’s test when the multiple comparisons were made from data produced at different conditions. All error bars indicate S.E.M. Detailed information of each statistical analysis is described in the figure legends.
Supplementary Material
Movie S1. Ca2+ responses to ascr#3 exposure in the SMB neurons of naive control. Related to Figure 4.
Movie S2. Ca2+ responses to ascr#3 exposure in the SMB neurons of ascr#3 imprinted animals. Related to Figure 4.
HIGHLIGHTS.
Early pheromone exposure modulates behavioral responses to the pheromone as adults.
Pheromone experience is imprinted as increased synaptic activity.
The odr-2 GPI-linked signaling protein mediates pheromone imprinting.
Acknowledgments
We are grateful to Yishi Jin for reagents, the Caenorhabditis Genetics Center (NIH Office of Research Infrastructure Programs, P40 OD010440) and the National BioResource Project (Japan) for strains, and Seung-Jae V. Lee, Sunkyung Lee, members of the S.V. Lee and K. Kim Lab for helpful discussion and/or critical comments on the manuscript. We also thank members of the Advanced Neural Imaging Center in KBRI for their technical support. This work was supported by the DGIST R&D Program of the Ministry of Science, ICT and Future Planning (17-BD-06), KBRI Basic Research Program of the Ministry of Science, ICT and Future Planning (17-BR-04), and the National Research Foundation of Korea (NRF-2015R1D1A1A09061430, NRF-2017R1A4A1015534) (K.K.), NIH R15GM111094 (S.E.H), NIH R01GM087533 (R.A.B), KBSI Grant T37416 (Y.H.H), and KBRI Basic Research Program of the Ministry of Science, ICT and Future Planning (17-BR-02), and the Korea Health Technology R&D Project of the Ministry for Health and Welfare (HI14C1135) (K.J.L).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Author contributions
M.H., L.S., M.O., and A.R.J. performed the experiments; J.K., S.C., R.A.B. and H.C. provides reagents; M.H., L.S., M.O., Y.H.H., K.J.L. P.S., S.E.H. and K.K. analyzed and interpreted data; M.H., L.S., P.S., S.E.H., and K.K wrote the manuscript.
References
- 1.Beach FA, Jaynes J. Effects of early experience upon the behavior of animals. Psychol Bull. 1954;51:239–263. doi: 10.1037/h0061176. [DOI] [PubMed] [Google Scholar]
- 2.Hensch TK. Critical period regulation. Annu Rev Neurosci. 2004;27:549–579. doi: 10.1146/annurev.neuro.27.070203.144327. [DOI] [PubMed] [Google Scholar]
- 3.Jeong PY, Jung M, Yim YH, Kim H, Park M, Hong E, Lee W, Kim YH, Kim K, Paik YK. Chemical structure and biological activity of the Caenorhabditis elegans dauer-inducing pheromone. Nature. 2005;433:541–545. doi: 10.1038/nature03201. [DOI] [PubMed] [Google Scholar]
- 4.Butcher RA, Fujita M, Schroeder FC, Clardy J. Small-molecule pheromones that control dauer development in Caenorhabditis elegans. Nat Chem Biol. 2007;3:420–422. doi: 10.1038/nchembio.2007.3. [DOI] [PubMed] [Google Scholar]
- 5.Macosko EZ, Pokala N, Feinberg EH, Chalasani SH, Butcher RA, Clardy J, Bargmann CI. A hub-and-spoke circuit drives pheromone attraction and social behaviour in C. elegans. Nature. 2009;458:1171–1175. doi: 10.1038/nature07886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Edison AS. Caenorhabditis elegans pheromones regulate multiple complex behaviors. Curr Opin Neurobiol. 2009;19:378–388. doi: 10.1016/j.conb.2009.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jang H, Kim K, Neal SJ, Macosko E, Kim D, Butcher RA, Zeiger DM, Bargmann CI, Sengupta P. Neuromodulatory state and sex specify alternative behaviors through antagonistic synaptic pathways in C. elegans. Neuron. 2012;75:585–592. doi: 10.1016/j.neuron.2012.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lorenz K. Der Kumpan in der Umwelt des Vogels. Journal für Ornithologie. 1935;83:137–213. [Google Scholar]
- 9.Hilliard MA, Bargmann CI, Bazzicalupo P. C. elegans responds to chemical repellents by integrating sensory inputs from the head and the tail. Curr Biol. 2002;12:730–734. doi: 10.1016/s0960-9822(02)00813-8. [DOI] [PubMed] [Google Scholar]
- 10.Srinivasan J, Kaplan F, Ajredini R, Zachariah C, Alborn HT, Teal PE, Malik RU, Edison AS, Sternberg PW, Schroeder FC. A blend of small molecules regulates both mating and development in Caenorhabditis elegans. Nature. 2008;454:1115–1118. doi: 10.1038/nature07168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sims JR, Ow MC, Nishiguchi MA, Kim K, Sengupta P, Hall SE. Developmental programming modulates olfactory behavior in C elegans via endogenous RNAi pathways. Elife. 2016;5 doi: 10.7554/eLife.11642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hall SE, Beverly M, Russ C, Nusbaum C, Sengupta P. A cellular memory of developmental history generates phenotypic diversity in C. elegans. Curr Biol. 2010;20:149–155. doi: 10.1016/j.cub.2009.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hall SE, Chirn GW, Lau NC, Sengupta P. RNAi pathways contribute to developmental history-dependent phenotypic plasticity in C. elegans. RNA. 2013;19:306–319. doi: 10.1261/rna.036418.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.L’Etoile ND, Coburn CM, Eastham J, Kistler A, Gallegos G, Bargmann CI. The cyclic GMP-dependent protein kinase EGL-4 regulates olfactory adaptation in C. elegans. Neuron. 2002;36:1079–1089. doi: 10.1016/s0896-6273(02)01066-8. [DOI] [PubMed] [Google Scholar]
- 15.Remy JJ, Hobert O. An interneuronal chemoreceptor required for olfactory imprinting in C. elegans. Science. 2005;309:787–790. doi: 10.1126/science.1114209. [DOI] [PubMed] [Google Scholar]
- 16.Ikeda DD, Duan Y, Matsuki M, Kunitomo H, Hutter H, Hedgecock EM, Iino Y. CASY-1, an ortholog of calsyntenins/alcadeins, is essential for learning in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2008;105:5260–5265. doi: 10.1073/pnas.0711894105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jin X, Pokala N, Bargmann CI. Distinct Circuits for the Formation and Retrieval of an Imprinted Olfactory Memory. Cell. 2016;164:632–643. doi: 10.1016/j.cell.2016.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bargmann CI, Hartwieg E, Horvitz HR. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 1993;74:515–527. doi: 10.1016/0092-8674(93)80053-h. [DOI] [PubMed] [Google Scholar]
- 19.Chou JH, Bargmann CI, Sengupta P. The Caenorhabditis elegans odr-2 gene encodes a novel Ly-6-related protein required for olfaction. Genetics. 2001;157:211–224. doi: 10.1093/genetics/157.1.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kim J, Yeon J, Choi SK, Huh YH, Fang Z, Park SJ, Kim MO, Ryoo ZY, Kang K, Kweon HS, et al. The Evolutionarily Conserved LIM Homeodomain Protein LIM-4/LHX6 Specifies the Terminal Identity of a Cholinergic and Peptidergic C. elegans Sensory/Inter/Motor Neuron-Type. PLoS Genet. 2015;11:e1005480. doi: 10.1371/journal.pgen.1005480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.White JG, Southgate E, Thomson JN, Brenner S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci. 1986;314:1–340. doi: 10.1098/rstb.1986.0056. [DOI] [PubMed] [Google Scholar]
- 22.Gray JM, Hill JJ, Bargmann CI. A circuit for navigation in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2005;102:3184–3191. doi: 10.1073/pnas.0409009101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sagasti A, Hobert O, Troemel ER, Ruvkun G, Bargmann CI. Alternative olfactory neuron fates are specified by the LIM homeobox gene lim-4. Genes Dev. 1999;13:1794–1806. doi: 10.1101/gad.13.14.1794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Piggott BJ, Liu J, Feng Z, Wescott SA, Xu XZ. The neural circuits and synaptic mechanisms underlying motor initiation in C. elegans. Cell. 2011;147:922–933. doi: 10.1016/j.cell.2011.08.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Bono M, Maricq AV. Neuronal substrates of complex behaviors in C. elegans. Annu Rev Neurosci. 2005;28:451–501. doi: 10.1146/annurev.neuro.27.070203.144259. [DOI] [PubMed] [Google Scholar]
- 26.Giles AC, Rose JK, Rankin CH. Investigations of learning and memory in Caenorhabditis elegans. Int Rev Neurobiol. 2006;69:37–71. doi: 10.1016/S0074-7742(05)69002-2. [DOI] [PubMed] [Google Scholar]
- 27.Ardiel EL, Rankin CH. An elegant mind: learning and memory in Caenorhabditis elegans. Learn Mem. 2010;17:191–201. doi: 10.1101/lm.960510. [DOI] [PubMed] [Google Scholar]
- 28.Nevitt GA, Dittman AH, Quinn TP, Moody WJ., Jr Evidence for a peripheral olfactory memory in imprinted salmon. Proc Natl Acad Sci U S A. 1994;91:4288–4292. doi: 10.1073/pnas.91.10.4288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wilson DA, Sullivan RM. Neurobiology of associative learning in the neonate: early olfactory learning. Behav Neural Biol. 1994;61:1–18. doi: 10.1016/s0163-1047(05)80039-1. [DOI] [PubMed] [Google Scholar]
- 30.Golden JW, Riddle DL. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science. 1982;218:578–580. doi: 10.1126/science.6896933. [DOI] [PubMed] [Google Scholar]
- 31.Izrayelit Y, Robinette SL, Bose N, von Reuss SH, Schroeder FC. 2D NMR-based metabolomics uncovers interactions between conserved biochemical pathways in the model organism Caenorhabditis elegans. ACS Chem Biol. 2013;8:314–319. doi: 10.1021/cb3004644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Guo ZV, Hart AC, Ramanathan S. Optical interrogation of neural circuits in Caenorhabditis elegans. Nat Methods. 2009;6:891–896. doi: 10.1038/nmeth.1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fujihara Y, Ikawa M. GPI-AP release in cellular, developmental, and reproductive biology. J Lipid Res. 2016;57:538–545. doi: 10.1194/jlr.R063032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Miwa JM, Ibanez-Tallon I, Crabtree GW, Sanchez R, Sali A, Role LW, Heintz N. lynx1, an endogenous toxin-like modulator of nicotinic acetylcholine receptors in the mammalian CNS. Neuron. 1999;23:105–114. doi: 10.1016/s0896-6273(00)80757-6. [DOI] [PubMed] [Google Scholar]
- 35.Tekinay AB, Nong Y, Miwa JM, Lieberam I, Ibanez-Tallon I, Greengard P, Heintz N. A role for LYNX2 in anxiety-related behavior. Proc Natl Acad Sci U S A. 2009;106:4477–4482. doi: 10.1073/pnas.0813109106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gottschalk A, Schafer WR. Visualization of integral and peripheral cell surface proteins in live Caenorhabditis elegans. J Neurosci Methods. 2006;154:68–79. doi: 10.1016/j.jneumeth.2005.11.016. [DOI] [PubMed] [Google Scholar]
- 37.Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77:71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Coates JC, de Bono M. Antagonistic pathways in neurons exposed to body fluid regulate social feeding in Caenorhabditis elegans. Nature. 2002;419:925–929. doi: 10.1038/nature01170. [DOI] [PubMed] [Google Scholar]
- 39.Tian L, Hires SA, Mao T, Huber D, Chiappe ME, Chalasani SH, Petreanu L, Akerboom J, McKinney SA, Schreiter ER, et al. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat Methods. 2009;6:875–881. doi: 10.1038/nmeth.1398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kim K, Sato K, Shibuya M, Zeiger DM, Butcher RA, Ragains JR, Clardy J, Touhara K, Sengupta P. Two chemoreceptors mediate developmental effects of dauer pheromone in C. elegans. Science. 2009;326:994–998. doi: 10.1126/science.1176331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Srinivasan J, Durak O, Sternberg PW. Evolution of a polymodal sensory response network. BMC Biol. 2008;6:52. doi: 10.1186/1741-7007-6-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ibsen S, Tong A, Schutt C, Esener S, Chalasani SH. Sonogenetics is a non-invasive approach to activating neurons in Caenorhabditis elegans. Nat Commun. 2015;6:8264. doi: 10.1038/ncomms9264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chronis N, Zimmer M, Bargmann CI. Microfluidics for in vivo imaging of neuronal and behavioral activity in Caenorhabditis elegans. Nat Methods. 2007;4:727–731. doi: 10.1038/nmeth1075. [DOI] [PubMed] [Google Scholar]
- 44.Kratsios P, Stolfi A, Levine M, Hobert O. Coordinated regulation of cholinergic motor neuron traits through a conserved terminal selector gene. Nat Neurosci. 2011;15:205–214. doi: 10.1038/nn.2989. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Movie S1. Ca2+ responses to ascr#3 exposure in the SMB neurons of naive control. Related to Figure 4.
Movie S2. Ca2+ responses to ascr#3 exposure in the SMB neurons of ascr#3 imprinted animals. Related to Figure 4.