

Abstract
The COP9 signalosome (CSN) is a highly conserved protein complex involved in the ubiquitin-proteasome system. Its metalloisopeptidase activity resides in subunit 5 (CSN5). Functions of csn5 in phytopathogenic fungi are poorly understood. Here, we knocked out the csn5 ortholog (Aacsn5) in the tangerine pathotype of Alternaria alternata. The ΔAacsn5 mutant showed a moderately reduced growth rate compared to the wildtype strain and was unable to produce conidia. The growth of ΔAacsn5 mutant was not affected in response to oxidative and osmotic stresses. Virulence assays revealed that ΔAacsn5 induced no or significantly reduced necrotic lesions on detached citrus leaves. The defects in hyphal growth, conidial sporulation, and pathogenicity of ΔAacsn5 were restored by genetic complementation of the mutant with wildtype Aacsn5. To explore the molecular mechanisms of conidiation and pathogenesis underlying Aacsn5 regulation, we systematically examined the transcriptomes of both ΔAacsn5 and the wildtype. Generally, 881 genes were overexpressed and 777 were underexpressed in the ΔAacsn5 mutant during conidiation while 694 overexpressed and 993 underexpressed during infection. During asexual development, genes related to the transport processes and nitrogen metabolism were significantly downregulated; the expression of csn1–4 and csn7 in ΔAacsn5 was significantly elevated; secondary metabolism gene clusters were broadly affected; especially, the transcript level of the whole of cluster 28 and 30 was strongly induced. During infection, the expression of the host-specific ACT toxin gene cluster which controls the biosynthesis of the citrus specific toxin was significantly repressed; many other SM clusters with unknown products were also regulated; 86 out of 373 carbohydrate-active enzymes responsible for breaking down the plant dead tissues showed uniquely decreased expression. Taken together, our results expand our understanding of the roles of csn5 on conidiation and pathogenicity in plant pathogenic fungi and provide a foundation for future investigations.
Keywords: csn5, conidiation, pathogenesis, Alternaria alternata, tangerine pathotype, RNA-Seq
Introduction
The COP9 signalosome (CSN) is an evolutionarily conserved protein complex initially identified as a photomorphogenic regulator of development in Arabidopsis thaliana (Wei et al., 1994) and subsequently characterized in mammals, invertebrates, and fungi (Oron et al., 2002; Kato and Yoneda-Kato, 2009; Braus et al., 2010). The CSN complex regulates ubiquitin-proteasome-mediated proteolysis. The best-studied function of the CSN is the control of the activity of cullin–RING ubiquitin E3 ligases (CRLs; Petroski and Deshaies, 2005). A typical CRL complex consists of a backbone cullin subunit, a RING protein, an adaptor protein, and a substrate-recognition subunit (Petroski and Deshaies, 2005). Nedd8 (neural precursor cell expressed, developmentally downregulated 8) is a ubiquitin-like protein which promotes the assembly of the CRL complex and the ubiquitylation of substrates by covalently attaching to a conserved lysine site on cullin proteins. The CSN functions as an isopeptidase enzyme to negatively regulate the activity of CRLs by removing the Ndedd8 from cullins (deneddylation) (Lyapina et al., 2001).
The CSN complex is composed of eight subunits in higher eukaryotic organisms, designated as CSN1–CSN8 (Wei and Deng, 2003). In humans, the CSN has two organizational centers: a horseshoe-shaped ring formed by CSN1–CSN4 and CSN7–CSN8 which contain a PCI (proteasome lid-CSN-initiation factor 3) domain and a large carboxy-terminal helical bundle created by all the subunits. CSN5 and CSN6, which have an MPN (Mpr1 and Pad1 N-terminal) domain, form a dimer and are embedded at the core of the helical bundle (Lingaraju et al., 2014). Specifically, the CSN5 subunit harbors a metalloprotease motif (JAMM, Jab1/MPN/Mov34) in the MPN domain responsible for the isopeptidase activity, making it the sole subunit that can provide the catalytic center for the CSN complex and cleave the Ndedd8 from cullins (Cope et al., 2002). Besides the basic role in regulating the activity of CRLs, the CSN complex also participates in many other biological processes. The botanic CSN is involved in photomorphogenesis, hormone signaling, flower development, iron deficiency, and plant pathogen response (Craig et al., 2009; Santner and Estelle, 2010; Lau and Deng, 2012; Stratmann and Gusmaroli, 2012; Duplan and Rivas, 2014; Tan et al., 2016). In mammals, the CSN is involved in several important biological functions such as cell cycle regulation, checkpoint control, signal transduction, autophagy, apoptosis, and tumorigenesis (Kato and Yoneda-Kato, 2009; Lee et al., 2011; Su et al., 2013).
The subunit composition of CSN in some fungi can be significantly different from that of the higher organisms. Neurospora crassa lacks the CSN8 subunit while the fission yeast Schizosaccharomyces pombe lacks CSN6 and CSN8 (Mundt et al., 2002; Liu et al., 2003; Wang et al., 2010). The budding yeast Saccharomyces cerevisiae possesses an alternative CSN complex composed of six subunits (Maytal-Kivity et al., 2002). In higher eukaryotes, the absence of any subunit is lethal in early stages of development (Wei et al., 2008; Zhao et al., 2011). However, fungi can survive without a CSN subunit (Gerke and Braus, 2014). Therefore, fungi are ideal model systems for understanding the evolution and biological roles of CSN. Functions of the fungal CSN have been characterized in several model species. In S. pombe, the CSN is required for the coordination of S phase and resistance to DNA damage (Mundt et al., 1999, 2002). In S. cerevisiae, the CSN plays a role in mating efficiency, pheromone sensitivity, lipid metabolism, and zinc absorption (Maytal-Kivity et al., 2002; Licursi et al., 2014). The N. crassa CSN is essential for conidiation and circadian rhythms (Wang et al., 2010), while the CSN in Aspergillus nidulans is involved in the light response, sexual development, and secondary metabolism (SM; Nahlik et al., 2010). Despite these advances, the molecular mechanism underlying the CSN’s control of development, pathogenicity, and SM in filamentous phytopathogenic fungi is poorly understood.
Alternaria alternata is a necrotrophic fungus, which can be ubiquitously isolated from soil and various plants and decaying plant materials (Thomma, 2003). Some of the strains in this species can cause diseases on plants and result in severe crop losses worldwide. At least seven pathogenic A. alternata pathotypes, each producing a unique host-selective toxin (HST), have been recognized to cause diseases in Japanese pear, strawberry, tangerine, apple, tomato, rough lemon, and tobacco, respectively (Tsuge et al., 2013). The HSTs in A. alternata pathotypes are crucial for their respective pathogenicity (Tsuge et al., 2013). The tangerine pathotype of Alternaria alternata is the causal agent of the Alternaria brown spot disease, which causes significant losses of both yield and marketability for tangerines and tangerine hybrids worldwide. In this study, we have identified the csn5 gene homolog in the tangerine pathotype of A. alternata (Aacsn5) and characterized its function using a gene deletion and complementation strategy. We also performed a transcriptome-wide gene expression analysis to explore the regulatory role of Aacsn5 in the development of conidia and pathogenicity in this important citrus pathogen.
Materials and Methods
Fungal Strains and Culture Conditions
The reference A. alternata strain, Z7, was isolated from an infected citrus fruit from Zhejiang, China (Huang et al., 2015; Wang et al., 2016). Z7 and its derived mutants were stored in 20% glycerol solutions at -80°C until use. Fungi were grown on regular solid PDA (potato dextrose agar) at 25°C and conidia were collected after incubating for 8 days. Mycelia were obtained by growing spores in liquid potato dextrose broth (PDB) incubated on a rotary shaker at 160 rpm at 25°C for 2 days.
Genetic Construction of Aacsn5 Deletion and Complementation Mutants
The Aacsn5 (accession number AALT_g2342) was identified using the A. nidulans (accession number AAM95164) csn5 homolog as a query to search the proteome of A. alternata by BLASTp. The Aacsn5 gene was knocked out using a fungal protoplast transformation as described previously (Chen et al., 2017; Ruan et al., 2017). Briefly, the deletion vectors were constructed by inserting two ∼1 kb flanking sequences of the Aacsn5 gene into the left and right sides of the neomycin-resistance gene in the pA1300-NEO plasmid (Wang et al., 2015). The resulting fragment was then amplified and directly introduced into the wildtype protoplasts using polyethylene glycol and CaCl2 (Supplementary Figure S2A). Transformants growing on a regeneration medium amended with 100 μg/mL neomycin were selected, examined by polymerase chain reaction (PCR) with specific primer pairs, and verified by southern blot (Supplementary Figures S2B,C). For genetic complementation, a full-length Aacsn5 fragment with its endogenous promoter was amplified by PCR and was inserted into the pTFCM plasmid carrying a hygromycin-resistance gene (Wang et al., 2015). The resultant plasmid was then transformed into protoplasts of a ΔAacsn5 mutant. Transformants were recovered from medium supplemented with hygromycin (150 mg/L) and examined by PCR and southern blot (Supplementary Figures S2B,C). All the primers used in this study are listed in Supplementary Table S1.
Fungal Growth Assays and Ergosterol Quantification
To examine oxidative and osmotic stress tolerance, ΔAacsn5, CPAacsn5, and wildtype A. alternata were grown on PDA plates supplemented with either 1.5 M NaCl, 0.5 M CaCl2, 1 mM H2O2, or 3 mM menadione (VK3). Each plate was inoculated with a 5 mm mycelial plug taken from the edge of a 5-day-old colony. The diameters of the colonies were measured after the plates were incubated at 25°C for 5 days. To examine differences in carbon and nitrogen utilization, fungal strains were grown on modified Czapek’s medium (30 g glucose, 3.0 g NaNO3, 1.0 g KH2PO4, 0.5 g MgSO4, 0.5 g KCl, 0.01 g FeSO4, and 15 g agar per liter). More specifically, the sole source of carbon (i.e., glucose) was replaced with equal mass sucrose, cellulose, hemicellulose, pectin, or citrus leaf to examine differences in carbon utilization and the sole source of nitrogen (i.e., NaNO3) was replaced with equal mass yeast extract, peptone, urea, acetamide, or phenylalanine to examine differences in nitrogen utilization. The diameters of the colonies were measured after the plates were incubated at 25°C for 6 days. For ergosterol extraction, fungal strains were added to a 150 mL yeast glucose (YG) medium (5 g/L yeast extract and 15 g/L glucose) and incubated at 25°C for 2 days on a rotary shaker. Mycelia were harvested by passing through a filter paper and washing three times with sterile water. Total ergosterol was extracted with hexane using a previously described method (Wang et al., 2014). Ergosterol samples were analyzed using an Agilent1100 high-performance liquid chromatography system. All experiments were repeated two times.
Pathogenicity and Toxin Assays
Fungal virulence was assessed by placing a 5 mm plug taken from the PDA media on ponkan (Citrus reticulata Blanco) leaves for 48 h or by placing the mycelia harvested from the liquid PDB media on the leaves for 24 h. Each strain was tested on at least 15 leaves and experiments were repeated two times. Host-selective ACT toxin was crudely extracted using Amberlite XAD-2 resin and ethyl acetate from culture filtrates (Kohmoto et al., 1993). The toxicity was assessed by spreading 40 μl ethyl acetate extracts onto detached ponkan leaves.
Transcriptome Analysis
To investigate the regulatory role of Aacsn5 in conidia formation, the colonies of wildtype and ΔAacsn5 mutants were scraped from the solid PDA medium after the incubation for 7 days at which time the conidia of the wildtype began to form. To investigate the regulatory role of Aacsn5 in pathogenesis, an equal number of mycelia from both wildtype and ΔAacsn5 mutants were inoculated on the citrus leaves, respectively, and were harvested at 24 h post-inoculation. Collected mycelia of each sample were then ground in liquid nitrogen and total RNA was extracted using an AxyPrepTM multisource total RNA miniprep kit. RNA-Seq was conducted for two biological replicates of each sample. The libraries were performed using an IlluminaTruSeq RNA Sample Preparation Kit and were sequenced on an Illumina Hiseq 2500 platform, generating 150 bp paired-end reads. Raw RNA-Seq reads were trimmed of adapter sequences and low-quality reads using trimmomatic (Bolger et al., 2014). Index of the A. alternata Z7 genome was built using Bowtie2 (Langmead and Salzberg, 2012) and cleaned reads were mapped to the reference genome using TopHat2 (Kim et al., 2013). The number of reads mapped to each gene was counted by HTSeq (Anders et al., 2014) and the resulting transcript count tables were subjected to DESeq R package for differential expression analysis (Anders and Huber, 2012). Transcripts with an adjusted P value less than 0.01 and the absolute value of log2FC (log2 fold change) greater than 2 were determined as differentially expressed. The differentially expressed genes (DEGs) were annotated by BLAST search against the NCBI non-redundant protein sequences database. Gene ontology (GO) enrichment analysis of DEGs was conducted using topGO (Alexa and Rahnenfuhrer, 2016). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed using KOBAS 3.0 (Xie et al., 2011). The transcriptome data reported in this study have been deposited in NCBI’s Sequence Read Archive with accession number SRP120511.
qRT-PCR Analysis
To validate the transcriptome data obtained by RNA sequencing, quantitative real-time PCR (qRT-PCR) was carried out on 12 A. alternata genes; 5 μg of each RNA sample was used for reverse transcription with the Prime Script RT reagent kit (TakaRa Biotechnology, Co., Dalian, China). The relative transcript level of the selected genes was quantified in triplicate on a 7300 Real-Time PCR system (ABI, United States). Primers used in this study were listed in Supplementary Table S1. The actin-encoding gene (KP341672) was used as an internal control and the resulting data were normalized using the comparative 2-ΔΔCT as described previously (Wang et al., 2014).
Results
Identification and Deletion of Aacsn5
By searching the whole genome of A. alternata Z7, we discovered a gene showing 58% amino acid identity and 97% coverage to the A. nidulans CSN5 homolog, and thus we designated this gene Aacsn5. Sequence alignment of CSN5 homologs from fungal species with different taxonomies confirmed that they share a highly conserved MPN domain (Supplementary Figure S1). The cloned Aacsn5 gene, which encodes a protein of 353 amino acids, has a 1149 bp ORF interrupted by one intron of 81 bp.
To investigate the functions of Aacsn5, an A. alternata mutant defective for Aacsn5 was generated by targeted gene disruption (Supplementary Figure S2A). In the mutant Aacsn5 allele, a 637 bp fragment spanning from Ala-55 to Arg-267 including the conserved MPN domain was replaced by the neomycin-resistance cassette. The disrupted and complemented mutants were first verified by PCR analysis with outside and inside primer pairs (Supplementary Figure S2B) and then further confirmed by southern blot analysis (Supplementary Figure S2C). As both mutants we got showed identical phenotype, only one mutant stain was used to perform the following experiments.
Involvement of Aacsn5 in Vegetative Growth and Conidiation
During growth on basic PDA medium, the ΔAacsn5 exhibited a moderate growth defect (29–34% reduction in growth) compared to the wildtype strain (Figure 1A). A similar phenotype (18–23% reduction in growth) was also observed when the ΔAacsn5 mutant was grown on the YG (yeast 5 g, glucose 15 g, 1 L) medium (Figure 1A). The defects in hyphal growth of ΔAacsn5 on solid media were restored by genetic complementation of the mutant with wildtype Aacsn5 (Figure 1A). Conidiation is very important for ascomycetous fungi to survive adversity and propagate. The wildtype strain Z7 can produce a large number of conidia after incubation in solid PDA for 7 days; however, no conidia were observed in the ΔAacsn5 mutant (Figure 1B). The ΔAacsn5 mutant was allowed to grow for two additional weeks but did not produce any conidia during this time period, demonstrating that conidiation is entirely abrogated and not merely delayed in this mutant. Thus, Aacsn5 is absolutely required for the formation of conidia in A. alaternata. The defects in conidial sporulation of ΔAacsn5 on solid media were partly (∼50%) restored by genetic complementation of the mutant with wildtype Aacsn5 (Figure 1B).
FIGURE 1.
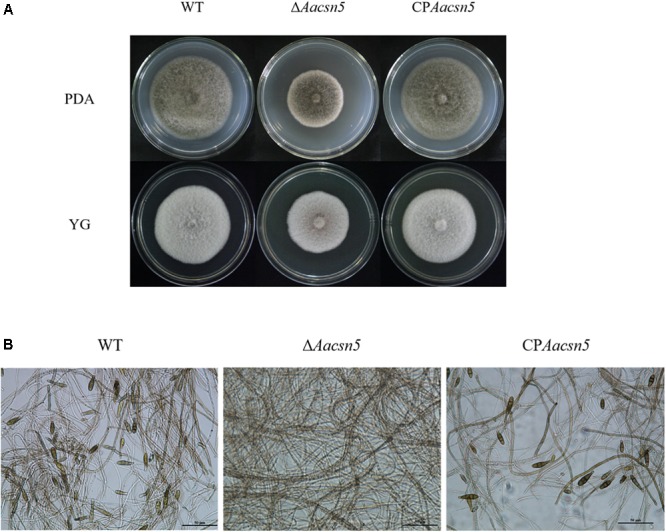
Aacsn5 is required for vegetative growth and conidiation. (A) Vegetative growth of the wildtype, ΔAacsn5, and CPAacsn5 mutant strains on PDA or YG at 25°C for 5 days. (B) Light microscopy images of the formation of conidia by A. alternata strains on PDA.
Aacsn5 Is Dispensable for Stress Response and Sterol Biosynthesis
To investigate whether Aacsn5 is involved in external stress tolerance, wildtype and mutant strains were grown on PDA media supplemented with salt stress inducers (1.5 M NaCl and 0.5 M CaCl2) and with oxidative stress inducers (15 mM H2O2 and 3 mM VK3) and analyzed for growth defects. No obvious change in stress tolerance was detected between the ΔAacsn5 strain and wildtype (Figure 2), indicating that Aacsn5 is not involved in the A. alternata response to osmotic and oxidative stresses.
FIGURE 2.

Effect of osmotic and oxidative stresses on the growth of the wildtype, ΔAacsn5, and CPAacsn5 mutant strains. Mycelial plugs were taken from the wildtype, ΔAacsn5, and CPAacsn5 mutants and grown individually on PDA medium supplemented with NaCl, CaCl2, H2O2, and menadione at the concentrations indicated in the figure. All plates were incubated for 5 days at 25°C.
Previously, it was reported that the csn5 homolog in S. cerevisiae is involved in the regulation of ergosterol metabolism (Licursi et al., 2014). To determine whether Aacsn5 regulates ergosterol, we quantified the total ergosterol content of the A. alternata wildtype and the ΔAacsn5 deletion mutant strains. However, no significant differences were found between them (2.25 μg/mg in the mutant versus 2.34 μg/mg in the wildtype).
Aacsn5 in Required for Full Pathogenicity of A. alternata
To study the possible effect of Aacsn5 on A. alternata virulence, infection assays were carried out on ponkan leaves. Most (26/30) ponkan leaves did not form necrotic lesions after inoculation with a ΔAacsn5 agar plug compared to all leaves inoculated with the wildtype strain, while four leaves formed significantly smaller lesions compared to the wildtype (Figure 3). To further test the virulence of the ΔAacsn5 mutant, we inoculated ponkan leaves with ΔAacsn5 and wildtype mycelia harvested from liquid PDB. We found that necrotic lesions caused by the wildtype mycelia were formed much faster than those caused by the agar plug and all leaves inoculated with ΔAacsn5 mycelia formed necrotic lesions (Figure 3). However, the ΔAacsn5 lesions were much less severe than those induced by the wildtype strain (Figure 3). Wounding the leaves prior to inoculation did not facilitate infection and lesion formation by the mutants (Supplementary Figure S3), suggesting that Aacsn5 is not involved in the capability of the fungus to penetrate the host. Taken together, our results indicate that Aacsn5 is required for the full virulence of A. alternata tangerine pathotype.
FIGURE 3.

The Aacsn5 gene is essential for fungal pathogenicity in the citrus. Necrosis symptoms in ponkan leaves inoculated with the PDA agar plug (48 h) and mycelia harvested from the liquid PDB (24 h) of the indicated strains.
Aacsn5 Regulates Expression of Genes During Conidiation and Plant–Pathogen Interaction
To explore the molecular mechanisms underlying the Aacsn5 regulation of conidiation, we performed a transcriptomic analysis comparing gene expression of the wildtype and ΔAacsn5 strains during asexual development. Overall, 1658 genes were differentially expressed in ΔAacsn5 compared to the wildtype strain (Supplementary Table S2), comprising 881 overexpressed and 777 underexpressed genes (Figure 4A). Genes expressed at a lower degree were enriched for the functional categories TRANSPORT, RESPONSE TO OXIDATIVE STRESS, RESPONSE TO DRUG, and OXIDATION–REDUCTION PROCESS while those expressed at a higher degree were enriched in the category OXIDATION–REDUCTION PROCESS (Figure 4B and Supplementary Table S3). After assigning these DEGs to the KEGG pathways, under-represented genes were significantly enriched in the nitrogen metabolism pathway while overexpressed genes were significantly enriched in the riboflavin metabolism, tyrosine metabolism, sulfur metabolism, and non-homologous end-joining pathways (Supplementary Table S3).
FIGURE 4.
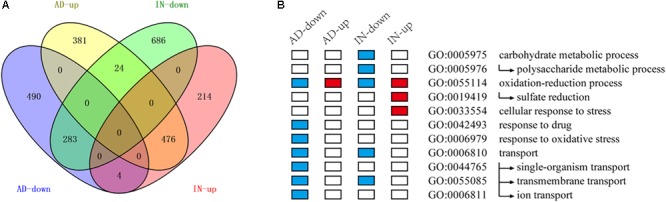
The RNA-seq analysis of ΔAacsn5 mutant during asexual development (AD) and infection (IN). (A) Venn diagram showing the upregulated and downregulated genes in ΔAacsn5 mutant compared with the wildtype strain. (B) Enriched functional categories for genes differentially expressed under different conditions. Red and blue boxes indicated GO terms enriched in overexpressed genes and underexpressed genes, respectively.
To further explore the effect of Aacsn5 on gene regulation during infection, we compared the transcriptomes of the ΔAacsn5 with the wildtype inoculated on ponkan leaves for 24 h. A total of 1687 DEGs were identified (Supplementary Table S2), comprising 694 overexpressed and 993 underexpressed genes (Figure 4A). Genes expressed at a lower degree were enriched for the functional categories CARBOHYDRATE METABOLIC PROCESS, TRANSPORT, and OXIDATION–REDUCTION PROCESS while those expressed at a higher degree were enriched in the category CELLULAR RESPONSE TO STRESS (Figure 4B and Supplementary Table S3). After assigning these DEGs to the KEGG pathways, underexpressed genes were significantly enriched in starch and sucrose metabolism, pentose and glucuronate interconversions, galactose metabolism, tyrosine metabolism, cyanoamino acid metabolism, phenylalanine metabolism, glycerolipid metabolism, fructose and mannose metabolism, arginine and proline metabolism, other glycan degradation, glycosphingolipid biosynthesis, and ascorbate and aldarate metabolism pathways while overexpressed genes were significantly enriched in the sulfur metabolism and non-homologous end-joining pathways (Supplementary Table S3).
After comparing two sets of DEGs related to conidiation and pathogenicity, 283 and 476 DGEs were upregulated and downregulated in both conditions, respectively, while 28 DEGs were oppositely regulated in both conditions (Figure 4A). More overexpressed genes overlapped between the two conditions (corresponding to 54% of all overexpressed genes during conidiation and 69% of all overexpressed genes during infection) than underexpressed genes (36% of all underexpressed genes during both conidiation and infection; Figure 4A).
To validate the RNA-seq results, 12 genes were selected and their expression levels were analyzed by qRT-PCR using gene-specific primers. These results showed that though the magnitude of fold changes between the two methods for some of the genes in the two conditions varied, overall both showed similar trends in transcript accumulation (Supplementary Figure S4).
Aacsn5 Affects the Expression of the CSN Complex During Asexual Development
The CSN complex is composed of at most eight subunits in fungi. To understand the expression pattern of these subunits in the absence of Aacsn5, we first searched the proteome of A. alternata Z7 by BLASTp using the A. nidulans CSN subunit proteins as queries. Like N. crassa, the A. alternata genome does not encode a csn8-like gene (Supplementary Table S4). As presented in Figure 5, with the exception of Aacsn6, the expression of other csn subunit genes in the ΔAacsn5 mutant was increased during infection and even more elevated during conidiation, indicating a negative feedback effect of Aacsn5 on the expression of csn1–4 and csn7, which may be involved in the conidiation process.
FIGURE 5.
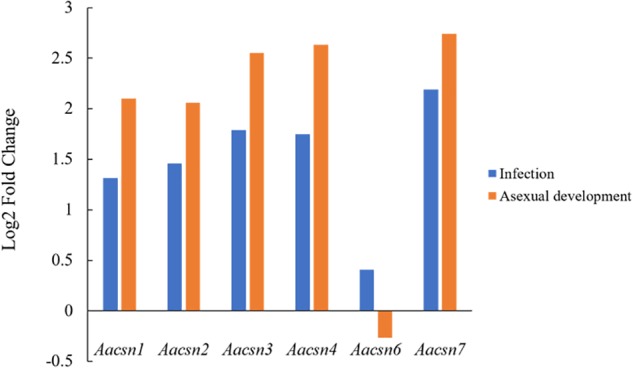
Expression of other csn subunit genes in the ΔAacsn5 mutant compared with wildtype strain during AD and IN.
Aacsn5 Does Not Mediate the Known Mechanism Underlying Sporulation in Other Fungi
In some model fungal species like A. nidulans and N. crassa, the BrlA > AbaA > WetA central regulatory pathway, the light-dependent signaling pathway, velvet family proteins, MAP kinase signaling pathway, G proteins, and FluG-mediated signaling pathway are important regulators of asexual development (Park and Yu, 2012). After looking into the expression pattern of these genes in the wildtype and ΔAacsn5 mutant, however, no significant difference was found between them except for the homolog of the flbC gene (AALT_g5513), which was expressed at an approximately eightfold lower level during asexual development in the ΔAacsn5 compared to wildtype (Supplementary Table S5).
Aacsn5 Widely Controls the Expression of CAZymes During Infection
Carbohydrate-active enzymes (CAZymes) play important roles in the breakdown of complex carbohydrates and are responsible for the acquisition of nutrients from the plant for phytopathogenic fungi. Previously, a total of 373 putative CAZyme genes were identified in A. alternata Z7 (Wang et al., 2016). Markedly, 98 of them showed significantly decreased expression levels during infection after the deletion of Aacsn5 (Figure 6A and Supplementary Table S6). Furthermore, 86 of these 98 CAZymes showed a unique expression pattern associated with pathogenicity, including 17 auxiliary activities, 12 carbohydrate esterases, 45 glycoside hydrolases, 4 glycosyl transferases (GTs), and 8 polysaccharide lyases (PLs) (Figure 6B). In particular, only 8% of the GT genes in the genome were downregulated in ΔAacsn5 during infection while 73% of the PL genes show a decreased expression level (Figure 6B). The transcript levels of 18 CAZymes were significantly elevated during infection while 33 and 26 CAZYmes overexpressed and underexpressed, respectively, during the asexual development (Figure 6A and Supplementary Table S6).
FIGURE 6.
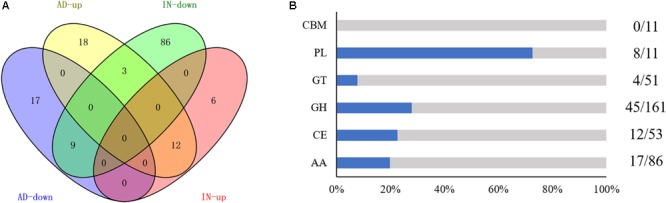
Differentially expressed CAZyme genes during AD and IN. (A) Venn diagram showing the upregulated and downregulated CAZyme genes in ΔAacsn5 mutant compared with the wildtype strain. (B) Percentage of exclusively down-regulated CAZyme genes (blue bars) in ΔAacsn5 mutant during infection. The corresponding number of exclusively downregulated CAZyme genes and the total number in the genome are shown on the right. GH, glycoside hydrolase; PL, polysaccharide lyase; CBM: carbohydrate-binding modules; GT: glycosyl transferase; CE: carbohydrate esterases; AA: auxiliary activities.
Aacsn5 Mediates the Expression of Some Transcription Factors
Aacsn5 may play its regulatory role by interacting with transcription factors. We then investigated which transcription factors were differentially expressed in the ΔAacsn5 strain. We found that 11 transcription factor genes are upregulated and 9 are downregulated during asexual development in Aacsn5 mutant while 12 and 6 transcription factors are upregulated and downregulated, respectively, during infection (Supplementary Table S7). Homologs of two functionally characterized C2H2-type transcription factors showed significantly downregulated gene expression in ΔAacsn5 during conidiation. One is the above-mentioned flbC (AALT_g5513, log2FC, -3.0), which is proposed to be involved in conidiophore development (Kwon et al., 2010), and the other is the amdA (AALT_g561, log2FC, -2.3), which controls the expression of the amdS and aciA structural genes required for acetamide catabolism (Lints et al., 1995). The nitrate-specific transcription factor NirA can activate the expression of the nitrate assimilation genes when nitrate or nitrite is present (Strauss et al., 1998). We discovered that the transcript level of its homolog (AALT_g8635, log2FC, 2.2) was increased in ΔAacsn5 mutant during asexual development. Interestingly, we found that the pH-responsive transcription factor PacC (AALT_g2998, log2FC, -2.1), whose homologs mediate not only the sensing and transduction of ambient pH but also the level of virulence in some plant pathogenic fungi (Caracuel et al., 2003; Zhang et al., 2013), was underexpressed in ΔAacsn5 during infection. The functions of the remaining transcription factors are largely uncharacterized and their relationship with Aacsn5 needs further investigation.
Aacsn5 Regulates Many SM Gene Clusters During Conidiation and Infection
To examine the relationship between SM and asexual development and pathogenicity mediated by Aacsn5, we examined the transcriptional responses of the 30 biosynthetic gene clusters in A. alternata predicted by anti-SMASH 4.0 (Blin et al., 2017; Supplementary Table S8). Overall, Aacsn5 showed a broad regulation of SM gene clusters in both conditions (Figure 7). The transcript level of cluster 28 and 30 was strongly induced in both conditions (Figure 7), suggested a negative regulation of Aacsn5 on both clusters. Clusters 15 and 25 are not regulated by Aacsn5 during conidiation; however, expression of some genes in these clusters is evidently decreased during infection (Figure 7). Conversely, clusters 3 and 7 are not regulated by Aacsn5 during infection while expression of a few genes in these clusters is underexpressed during conidiation (Figure 7). Only a few genes in clusters 2, 21, and ACT toxin were regulated by Aacsn5 during asexual development; however, a large number of genes in these clusters showed significantly underexpressed mRNA level during infection (Figure 7). ACT toxin is known to be the determinant for the pathogenicity of A. alternata to citrus (Kohmoto et al., 1993; Wang et al., 2016), so the reduced biosynthesis of this critical factor can help to explain the phenotype of less virulent ΔAacsn5. Only one or two genes in cluster 5 and 24 were regulated by Aacsn5 during infection while a large number of genes in these clusters are significantly downregulated and upregulated, respectively, during conidiation (Figure 7). Finally, a mixed regulation mode (an SM cluster has both upregulated and downregulated genes) was found only within cluster 6 in ΔAacsn5 during conidiation while this was very common (found in 11 clusters) during infection (Figure 7). Our results indicate that Aacsn5 regulates SM in a complex manner, which is associated with conidiation and infection.
FIGURE 7.

Differential expression of SM gene clusters in the ΔAacsn5 mutant during AD and IN. pks, polyketide synthase; nrps, non-ribosomal peptide synthetase; t1, Type 1; t3, Type 3.
Roles of Aacsn5 in Nutrients’ Utilization and Toxin Production During Development
Our RNA-Seq data suggested that the carbohydrate and nitrogen metabolic processes in ΔAacsn5 were significantly affected during infection (Supplementary Table S3). To determine if those were caused by the defect of the ΔAacsn5 mutant in nutrition utilization during development, we performed growth arrays of the wildtype and the ΔAacsn5 mutant on different carbon and nitrogen sources. The results showed that the ΔAacsn5 exhibited a growth defect of 2–15% depending on different nutrients compared to the wildtype strain (Figure 8A and Supplementary Figure S5). Especially, when the citrus leaves were used as the sole carbon source, the growth inhibition rate of the ΔAacsn5 mutant was ∼8% (Figure 8A and Supplementary Figure S5).
FIGURE 8.
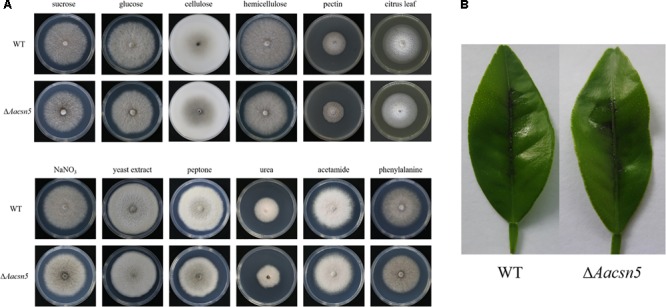
Roles of Aacsn5 in nutrient utilization and toxin production during development. (A) Growth of the wildtype and ΔAacsn5 on different carbon or nitrogen sources. Mycelial plugs were taken from the wildtype and ΔAacsn5 and grown individually on Czapek’s medium with the glucose, sucrose, cellulose, hemicellulose, pectin, or citrus leaf as the sole carbon source or, with NaNO3, yeast extract, peptone, urea, acetamide, or phenylalanine as the sole nitrogen source at 25°C for 6 days. (B) Necrotic lesions appearing on detached citrus leaves inoculated by spraying with the crude ACT toxin extracts from both the wildtype and ΔAacsn5 in the axenic culture.
The tangerine pathotype of A. alternata produces a host-selective ACT toxin which has been suggested as being the determining factor of pathogenicity (Miyamoto et al., 2008; Wang et al., 2016). Our transcriptome data also showed that the expression of the genes in the ACT gene cluster was significantly underexpressed during infection (Figure 7 and Supplementary Table S9). qRT-PCR analysis confirmed that the ACTT2 (AALT_g11743) and the ACTTS3 gene (AALT_g11750), which were reported to be critical for ACT toxin production in the tangerine pathotype of A. alternata (Miyamoto et al., 2008; Miyamoto et al., 2010), were significantly downregulated in ΔAacsn5 during infection (Supplementary Figure S4). To test whether Aacsn5 plays a role in regulating ACT production during development, we treated ponkan leaves with a dilution series of crude ACT toxin extracts from both wildtype and ΔAacsn5 strains in the axenic culture. The results showed that both strains induced similar necrotic lesions on the detached ponkan leaves (Figure 8B and Supplementary Figure S6).
Taken together, these results revealed that Aacsn5 plays a limited or no role in nutrients utilization and ACT toxin production during development. Combined with the transcriptome analysis, our data suggested that there might exist a unique host’s immune response during the process of pathogen–plant interaction, causing the attenuated virulence of the ΔAacsn5 mutant.
Discussion
As deficiency in any CSN subunits is lethal in higher eukaryotes, fungi are good model systems for an investigation into the molecular mechanisms of the CSN. Several studies have been performed to elucidate CSN composition, activity, and cellular functions in the model fungi including S. cerevisiae, N. crassa, and A. nidulans (Nahlik et al., 2010; Wang et al., 2010; Licursi et al., 2014). In this work, we investigated the biological roles that csn5 plays in a plant pathogenic fungus, the tangerine pathotype of A. alternata, which is an economically important agent of the citrus brown spot disease worldwide.
The production of asexual spores is of vital importance in the life cycle of phytopathogenic fungi. One of the major findings we present here is that the formation of conidia is totally blocked in the tangerine pathotype of A. alternata after functional inactivation of the csn5 homolog gene (Figure 1B), demonstrating that Aacsn5 is absolutely required for conidiation. A similar phenotype was also observed in the csn5 deletion mutant of another filamentous fungus, Pestalotiopsis fici (Zheng et al., 2017). However, conidiation was not affected in the A. nidulans csn5 deletion strain, though this mutant showed a severe defect in the sexual developmental cycle (Busch et al., 2003; Nahlik et al., 2010). In N. crassa, the csn5 mutant produces fewer conidia on race tubes than the wildtype (Wang et al., 2010). Taken together, these results indicate that the role of csn5 in conidiation has markedly diverged in different fungi.
A transcriptome analysis was performed to unveil the regulatory role Aacsn5 played during conidiation. Our results showed that genes downregulated in ΔAacsn5 during asexual development were significantly enriched in the transport-related processes and nitrogen metabolism (Figure 4B), and thus we speculate that the conidiation deficiency may be caused by failure to transport some nitrogen-containing compounds. Carbohydrate and nitrogen concentrations and their ratios can greatly influence the sporulation of many fungi (Elson et al., 1998; Gao et al., 2007). Further, SM is also an important factor for sporulation; the molecular mechanisms regulating SM are often involved in the control of asexual development (Keller, 2015; Macheleidt et al., 2016). For example, conidiogenone, an endogenous diterpenoid with conidiation-inducing activity, is continuously produced and released to the culture medium during the growth stage of Penicillium cyclopium, and it will trigger conidiation when it accumulates to a threshold concentration (Roncal et al., 2002). In our transcriptome data, we found that Aacsn5 has a broad regulation of SM gene clusters; especially, the transcript level of cluster 28 and 30 was strongly induced in ΔAacsn5 (Figure 7). Although the products of most SM clusters are not known, we speculate that there might be a relationship between SM and Aascn5-mediated sporulation.
The Velvet complex is a global regulator of morphogenesis and SM in filamentous fungi (Bayram et al., 2008). Deletions of either of two members of this complex, laeA and veA, greatly reduce conidia production in A. alternata (Estiarte et al., 2016). However, the expression of these genes or of any other Velvet family proteins was not affected in the ΔAacsn5 mutant (Supplementary Table S5). We also examined the expression of genes involved in the sporulation in other fungi, which include the BrlA > AbaA > WetA central regulatory pathway, the light-dependent signaling pathway, the velvet family proteins, MAP kinase signaling pathway, G proteins, and FluG-mediated signaling pathway (Park and Yu, 2012), but almost all of the genes in these pathways showed no difference at the transcriptional level (Supplementary Table S5). These data suggested that the Aacsn5 might mediate different mechanism underlying the sporulation from the important regulators in other fungi.
Our pathogenicity assays showed that the A. alternataΔAacsn5 mutants induced no or significantly reduced necrotic lesions on detached citrus leaves (Figure 3), demonstrating that Aacsn5 is required for wildtype levels of pathogenicity. To our knowledge, this is the first time that csn5 has been reported to be involved in virulence in phytopathogenic fungi. Previously, both the production of host-specific ACT toxin and the ability to detoxify reactive oxygen species (ROS) have been shown to be critical factors for the pathogenicity of the A. alternata tangerine pathotype (Miyamoto et al., 2008; Lin et al., 2009). The ability to synthesize the ACT toxin in the ΔAacsn5 mutant was not affected during vegetative growth in liquid medium (Figure 8B and Supplementary Figure S6). A similar situation was also observed in the virulence-attenuated A. alternata Yap1, Fus3, Hog1, Nps6, and Skn7 gene deletion mutants when grown in vitro (Lin et al., 2009, 2010; Lin and Chung, 2010; Chen et al., 2012, 2013). However, successfully synthesizing the ACT toxin in vitro does not guarantee an equivalent ACT toxin production in those mutants during infection. In those studies, no evidence was provided about whether the biosynthesis of ACT toxin in planta was impaired. Our transcriptome data showed that the expression of the genes in the ACT gene cluster was significantly underexpressed during infection (Figure 7), strongly suggesting the repression of the ACT toxin biosynthesis in planta. The discrepancy of toxin production between in vitro and in planta could be attributed to the host’s immune response during the process of pathogen–host interaction, causing the difficulty of ΔAacsn5 mutant in expressing those genes responsible for the ACT toxin biosynthesis, which coincides with the virulence assay results. Additionally, the expression of other clusters like cluster 2, 9, 15, 21, and 25 showed a very different expression pattern during infection compared to expression during asexual development (Figure 7), suggesting that these SM clusters might also be related to the pathogenicity of A. alternata. Our growth assays showed no change in the tolerance to H2O2 and menadione in the ΔAacsn5 strain compared to wildtype (Figure 2), suggesting that Aacsn5 plays no role in resistance to oxidative stress tolerance. Previously, two transcription factors SKN7 (AALT_g8622; Chen et al., 2012) and YAP1 (AALT_g912; Lin et al., 2009), HOG1 kinase (AALT_g10096; Lin and Chung, 2010), three NADPH oxidases NoxA (AALT_g5618; Yang and Chung, 2012), NoxB (AALT_g6028), and NoxR (AALT_g5214; Yang and Chung, 2013), glutathione peroxidase Gpx3 (AALT_g41; Yang et al., 2016), and major facilitator superfamily transporter Mfs19 (AALT_g2684; Chen et al., 2017) were shown to be essential for the detoxification of cellular stresses induced by ROS and for the pathogenesis in citrus. However, none of these genes showed significantly different expression in the Aacsn5 deletion mutants during infection (Supplementary Table S3). Thus, the reduced virulence of ΔAacsn5 is not associated with ROS scavenging.
In necrotrophic fungi, breaking down dead tissues for nutrient acquisition is carried out by various CAZymes and is one important process associated with pathogenesis. In our transcriptome data, we identified that a large number (86/373) of CAZymes encoding genes showed uniquely decreased expression in ΔAacsn5 mutant during infection (Figure 6), indicating that Aacsn5 positively regulates CAZymes during the infection of A. alternata to citrus leaves. Nitrogen availability and type can also be important factors for the virulence of fungal pathogens (Bolton and Thomma, 2008). For example, the A. alternata tobacco pathotype utilizes ammonia as a stimulator to form infection structures and to switch to a necrotrophic lifestyle (Duan et al., 2010). Studies using nitrogen-starved conditions to mimic the environment that a pathogen encounters during growth in planta have discovered a subset of genes related to fungal virulence (Stephenson et al., 2000; Donofrio et al., 2006; Bolton and Thomma, 2008; Brown et al., 2008). In this research, several amino acid metabolism pathways were identified to be underexpressed in ΔAacsn5 mutant during infection (Supplementary Table S3), implying the role of Aacsn5 in regulating nitrogen metabolism during infection.
Taken together, our results demonstrate that Aacsn5 plays a critical role in the conidiation and pathogenicity in the pathotype of A. alternata, the causal agent of Alternaria brown spot disease in citrus worldwide. To explore the potential mechanisms of Aacsn5 in regulating conidiation and pathogenicity, we performed RNA-Seq analyses and discovered candidate genes associated with conidiation and pathogenesis. Our analysis opens avenues into researches on the molecular function of the CSN in plant pathogenetic fungi. Ongoing research is needed to elucidate the precise molecular mechanism by which the CSN affects fungal development, nutrition metabolism, SM, and pathogenicity.
Author Contributions
MW and HL conceived and designed the experiments. XY, MW, RR, and HF performed the experiments. MW analyzed the data and wrote the paper. All authors reviewed the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank members of the Rokas Lab at Vanderbilt University for the helpful discussions. We also thank Dr. Abigail L. Lind and Matthew E. Mead for their critical comments on this paper.
Footnotes
Funding. This work was supported by the National Natural Science Foundation of China (Grant No. 31571948), the earmarked fund for China Agriculture Research System (Grant No. CARS-27), and the China Postdoctoral Science Foundation (Grant No. 2016M601945).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00508/full#supplementary-material
References
- Alexa A., Rahnenfuhrer J. (2016). topGO: Enrichment Analysis for Gene Ontology. (R Package version 2.28.0). [Google Scholar]
- Anders S., Huber W. (2012). Differential Expression of RNA-Seq Data at the Gene Level–the DESeq Package. Heidelberg: European Molecular Biology Laboratory (EMBL). [Google Scholar]
- Anders S., Pyl P. T., Huber W. (2014). HTSeq–A python framework to work with high-throughput sequencing data. Bioinformatics 31 166–169. 10.1093/bioinformatics/btu638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayram O., Krappmann S., Ni M., Bok J. W., Helmstaedt K., Valerius O., et al. (2008). VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 320 1504–1506. 10.1126/science.1155888 [DOI] [PubMed] [Google Scholar]
- Blin K., Wolf T., Chevrette M. G., Lu X., Schwalen C. J., Kautsar S. A., et al. (2017). antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 45 w36–w41. 10.1093/nar/gkx319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger A. M., Lohse M., Usadel B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30 2114–2120. 10.1093/bioinformatics/btu170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolton M. D., Thomma B. P. (2008). The complexity of nitrogen metabolism and nitrogen-regulated gene expression in plant pathogenic fungi. Physiol. Mol. Plant Pathol. 72 104–110. 10.1016/j.pmpp.2008.07.001 [DOI] [Google Scholar]
- Braus G. H., Irniger S., Bayram Ö. (2010). Fungal development and the COP9 signalosome. Curr. Opin. Microbiol. 13 672–676. 10.1016/j.mib.2010.09.011 [DOI] [PubMed] [Google Scholar]
- Brown S. H., Yarden O., Gollop N., Chen S., Zveibil A., Belausov E., et al. (2008). Differential protein expression in Colletotrichum acutatum: changes associated with reactive oxygen species and nitrogen starvation implicated in pathogenicity on strawberry. Mol. Plant Pathol. 9 171–190. 10.1111/j.1364-3703.2007.00454.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busch S., Eckert S. E., Krappmann S., Braus G. H. (2003). The COP9 signalosome is an essential regulator of development in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 49 717–730. 10.1046/j.1365-2958.2003.03612.x [DOI] [PubMed] [Google Scholar]
- Caracuel Z., Roncero M. I. G., Espeso E. A., Gonzalez-Verdejo C. I., Garcia-Maceira F. I., Di Pietro A. (2003). The pH signalling transcription factor PacC controls virulence in the plant pathogen Fusarium oxysporum. Mol. Microbiol. 48 765–779. 10.1046/j.1365-2958.2003.03465.x [DOI] [PubMed] [Google Scholar]
- Chen L. H., Lin C. H., Chung K. R. (2012). Roles for SKN7 response regulator in stress resistance, conidiation and virulence in the citrus pathogen Alternaria alternata. Fungal Genet. Biol. 49 802–813. 10.1016/j.fgb.2012.07.006 [DOI] [PubMed] [Google Scholar]
- Chen L. H., Lin C. H., Chung K. R. (2013). A nonribosomal peptide synthetase mediates siderophore production and virulence in the citrus fungal pathogen Alternaria alternata. Mol. Plant Pathol. 14 497–505. 10.1111/mpp.12021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L. H., Tsai H. C., Yu P. L., Chung K. R. (2017). A major facilitator superfamily transporter-mediated resistance to oxidative stress and fungicides requires Yap1, Skn7, and MAP kinases in the citrus fungal pathogen Alternaria alternata. PLoS One 12:e0169103. 10.1371/journal.pone.0169103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cope G. A., Suh G. S., Aravind L., Schwarz S. E., Zipursky S. L., Koonin E. V., et al. (2002). Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 298 608–611. 10.1126/science.1075901 [DOI] [PubMed] [Google Scholar]
- Craig A., Ewan R., Mesmar J., Gudipati V., Sadanandom A. (2009). E3 ubiquitin ligases and plant innate immunity. J. Exp. Bot. 60 1123–1132. 10.1093/jxb/erp059 [DOI] [PubMed] [Google Scholar]
- Donofrio N., Oh Y., Lundy R., Pan H., Brown D., Jeong J., et al. (2006). Global gene expression during nitrogen starvation in the rice blast fungus, Magnaporthe grisea. Fungal Genet. Biol. 43 605–617. 10.1016/j.fgb.2006.03.005 [DOI] [PubMed] [Google Scholar]
- Duan W., Zhang X., Yang T., Dou X., Chen T., Li S., et al. (2010). A novel role of ammonia in appressorium formation of Alternaria alternata (Fries) Keissler, a tobacco pathogenic fungus/Eine neue Rolle von Ammoniak bei der Appressoriumsbildung von Alternaria alternata (Fries) Keissler an Tabak. J. Plant Dis. Prot. 117 112–116. 10.1007/BF03356345 [DOI] [Google Scholar]
- Duplan V., Rivas S. (2014). E3 ubiquitin-ligases and their target proteins during the regulation of plant innate immunity. Front. Plant Sci. 5:42. 10.3389/fpls.2014.00042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elson M. K., Schisler D. A., Jackson M. A. (1998). Carbon-to-nitrogen ratio, carbon concentration, and amino acid composition of growth media influence conidiation of Helminthosporium solani. Mycologia 90 406–413. 10.2307/3761399 [DOI] [Google Scholar]
- Estiarte N., Lawrence C. B., Sanchis V., Ramos A. J., Crespo-Sempere A. (2016). LaeA and VeA are involved in growth morphology, asexual development, and mycotoxin production in Alternaria alternata. Int. J. Food Microbiol. 238(Suppl. C) 153–164. 10.1016/j.ijfoodmicro.2016.09.003 [DOI] [PubMed] [Google Scholar]
- Gao L., Sun M. H., Liu X. Z., Che Y. S. (2007). Effects of carbon concentration and carbon to nitrogen ratio on the growth and sporulation of several biocontrol fungi. Mycol. Res. 111 87–92. 10.1016/j.mycres.2006.07.019 [DOI] [PubMed] [Google Scholar]
- Gerke J., Braus G. H. (2014). Manipulation of fungal development as source of novel secondary metabolites for biotechnology. Appl. Microbiol. Biotechnol. 98 8443–8455. 10.1007/s00253-014-5997-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang F., Fu Y., Nie D., Stewart J. E., Peever T. L., Li H. (2015). Identification of a novel phylogenetic lineage of Alternaria alternata causing citrus brown spot in China. Fungal Biol. 119 320–330. 10.1016/j.funbio.2014.09.006 [DOI] [PubMed] [Google Scholar]
- Kato J. Y., Yoneda-Kato N. (2009). Mammalian COP9 signalosome. Genes Cells 14 1209–1225. 10.1111/j.1365-2443.2009.01349.x [DOI] [PubMed] [Google Scholar]
- Keller N. P. (2015). Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 11 671–677. 10.1038/nchembio.1897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim D., Pertea G., Trapnell C., Pimentel H., Kelley R., Salzberg S. L. (2013). TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14:R36. 10.1186/gb-2013-14-4-r36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohmoto K., Itoh Y., Shimomura N., Kondoh Y., Otani H., Kodama M., et al. (1993). Isolation and biological activities of two host-specific toxins from the tangerine pathotype of Alternaria alternata. Phytopathology 83 495–502. 10.1094/Phyto-83-495 [DOI] [Google Scholar]
- Kwon N.-J., Garzia A., Espeso E. A., Ugalde U., Yu J.-H. (2010). FlbC is a putative nuclear C2H2 transcription factor regulating development in Aspergillus nidulans. Mol. Microbiol. 77 1203–1219. 10.1111/j.1365-2958.2010.07282.x [DOI] [PubMed] [Google Scholar]
- Langmead B., Salzberg S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9 357–359. 10.1038/nmeth.1923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau O. S., Deng X. W. (2012). The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci. 17 584–593. 10.1016/j.tplants.2012.05.004 [DOI] [PubMed] [Google Scholar]
- Lee M. H., Zhao R., Phan L., Yeung S. C. (2011). Roles of COP9 signalosome in cancer. Cell Cycle 10 3057–3066. 10.4161/cc.10.18.17320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Licursi V., Salvi C., De Cesare V., Rinaldi T., Mattei B., Fabbri C., et al. (2014). The COP9 signalosome is involved in the regulation of lipid metabolism and of transition metals uptake in Saccharomyces cerevisiae. FEBS J. 281 175–190. 10.1111/febs.12584 [DOI] [PubMed] [Google Scholar]
- Lin C. H., Chung K. R. (2010). Specialized and shared functions of the histidine kinase- and HOG1 MAP kinase-mediated signaling pathways in Alternaria alternata, a filamentous fungal pathogen of citrus. Fungal Genet. Biol. 47 818–827. 10.1016/j.fgb.2010.06.009 [DOI] [PubMed] [Google Scholar]
- Lin C. H., Yang S. L., Chung K. R. (2009). The YAP1 homolog-mediated oxidative stress tolerance is crucial for pathogenicity of the necrotrophic fungus Alternaria alternata in citrus. Mol. Plant Microbe Interact. 22 942–952. 10.1094/MPMI-22-8-0942 [DOI] [PubMed] [Google Scholar]
- Lin C. H., Yang S. L., Wang N. Y., Chung K. R. (2010). The FUS3 MAPK signaling pathway of the citrus pathogen Alternaria alternata functions independently or cooperatively with the fungal redox-responsive AP1 regulator for diverse developmental, physiological and pathogenic processes. Fungal Genet. Biol. 47 381–391. 10.1016/j.fgb.2009.12.009 [DOI] [PubMed] [Google Scholar]
- Lingaraju G. M., Bunker R. D., Cavadini S., Hess D., Hassiepen U., Renatus M., et al. (2014). Crystal structure of the human COP9 signalosome. Nature 512 161–165. 10.1038/nature13566 [DOI] [PubMed] [Google Scholar]
- Lints R., Davis M. A., Hynes M. J. (1995). The positively acting amdA gene of Aspergillus nidulans encodes a protein with two C2H2 zinc-finger motifs. Mol. Microbiol. 15 965–975. 10.1111/j.1365-2958.1995.tb02365.x [DOI] [PubMed] [Google Scholar]
- Liu C., Powell K. A., Mundt K., Wu L., Carr A. M., Caspari T. (2003). Cop9/signalosome subunits and Pcu4 regulate ribonucleotide reductase by both checkpoint-dependent and-independent mechanisms. Genes Dev. 17 1130–1140. 10.1101/gad.1090803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyapina S., Cope G., Shevchenko A., Serino G., Tsuge T., Zhou C., et al. (2001). Promotion of NEDD-CUL1 conjugate cleavage by COP9 signalosome. Science 292 1382–1385. 10.1126/science.1059780 [DOI] [PubMed] [Google Scholar]
- Macheleidt J., Mattern D. J., Fischer J., Netzker T., Weber J., Schroeckh V., et al. (2016). Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 50 371–392. 10.1146/annurev-genet-120215-035203 [DOI] [PubMed] [Google Scholar]
- Maytal-Kivity V., Piran R., Pick E., Hofmann K., Glickman M. H. (2002). COP9 signalosome components play a role in the mating pheromone response of S. cerevisiae. EMBO Rep. 3 1215–1221. 10.1093/embo-reports/kvf235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyamoto Y., Masunaka A., Tsuge T., Yamamoto M., Ohtani K., Fukumoto T., et al. (2008). Functional analysis of a multicopy host-selective ACT-toxin biosynthesis gene in the tangerine pathotype of Alternaria alternata using RNA silencing. Mol. Plant Microbe Interact. 21 1591–1599. 10.1094/MPMI-21-12-1591 [DOI] [PubMed] [Google Scholar]
- Miyamoto Y., Masunaka A., Tsuge T., Yamamoto M., Ohtani K., Fukumoto T., et al. (2010). ACTTS3 encoding a polyketide synthase is essential for the biosynthesis of ACT-toxin and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant Microbe Interact. 23 406–414. 10.1094/MPMI-23-4-0406 [DOI] [PubMed] [Google Scholar]
- Mundt K. E., Liu C., Carr A. M. (2002). Deletion mutants in COP9/signalosome subunits in fission yeastSchizosaccharomyces pombe display distinct phenotypes. Mol. Biol. Cell 13 493–502. 10.1091/mbc.01-10-0521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mundt K. E., Porte J., Murray J. M., Brikos C., Christensen P. U., Caspari T., et al. (1999). The COP9/signalosome complex is conserved in fission yeast and has a role in S phase. Curr. Biol. 9 1427–1430. 10.1016/S0960-9822(00)80091-3 [DOI] [PubMed] [Google Scholar]
- Nahlik K., Dumkow M., Bayram O., Helmstaedt K., Busch S., Valerius O., et al. (2010). The COP9 signalosome mediates transcriptional and metabolic response to hormones, oxidative stress protection and cell wall rearrangement during fungal development. Mol. Microbiol. 78 964–979. 10.1111/j.1365-2958.2010.07384.x [DOI] [PubMed] [Google Scholar]
- Oron E., Mannervik M., Rencus S., Harari-Steinberg O., Neuman-Silberberg S., Segal D., et al. (2002). COP9 signalosome subunits 4 and 5 regulate multiple pleiotropic pathways in Drosophila melanogaster. Development 129 4399–4409. [DOI] [PubMed] [Google Scholar]
- Park H.-S., Yu J.-H. (2012). Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 15 669–677. 10.1016/j.mib.2012.09.006 [DOI] [PubMed] [Google Scholar]
- Petroski M. D., Deshaies R. J. (2005). Function and regulation of cullin-RING ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 6 9–20. 10.1038/nrm1547 [DOI] [PubMed] [Google Scholar]
- Roncal T., Cordobes S., Sterner O., Ugalde U. (2002). Conidiation in Penicillium cyclopium is induced by conidiogenone, an endogenous diterpene. Eukaryot. Cell 1 823–829. 10.1128/EC.1.5.823-829.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruan R., Wang M., Liu X., Sun X., Chung K.-R., Li H. (2017). Functional analysis of two sterol regulatory element binding proteins in Penicillium digitatum. PLoS One 12:e0176485. 10.1371/journal.pone.0176485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santner A., Estelle M. (2010). The ubiquitin-proteasome system regulates plant hormone signaling. Plant J. 61 1029–1040. 10.1111/j.1365-313X.2010.04112.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephenson S.-A., Hatfield J., Rusu A. G., Maclean D. J., Manners J. M. (2000). CgDN3: an essential pathogenicity gene of Colletotrichum gloeosporioides necessary to avert a hypersensitive-like response in the host Stylosanthes guianensis. Mol. Plant Microbe Interact. 13 929–941. 10.1094/MPMI.2000.13.9.929 [DOI] [PubMed] [Google Scholar]
- Stratmann J. W., Gusmaroli G. (2012). Many jobs for one good cop - the COP9 signalosome guards development and defense. Plant Sci. 186 50–64. 10.1016/j.plantsci.2011.10.004 [DOI] [PubMed] [Google Scholar]
- Strauss J., Muro-Pastor M. I., Scazzocchio C. (1998). The regulator of nitrate assimilation in ascomycetes is a dimer which binds a nonrepeated, asymmetrical sequence. Mol. Cell. Biol. 18 1339–1348. 10.1128/MCB.18.3.1339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su H., Li J., Osinska H., Li F., Robbins J., Liu J., et al. (2013). The COP9 signalosome is required for autophagy, proteasome-mediated proteolysis, and cardiomyocyte survival in adult mice. Circ. Heart Fail. 6 1049–1057. 10.1161/CIRCHEARTFAILURE.113.000338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan S., Liu F., Pan X. X., Zang Y. P., Jin F., Zu W. X., et al. (2016). CSN6, a subunit of the COP9 signalosome, is involved in early response to iron deficiency in Oryza sativa. Sci. Rep. 6:25485. 10.1038/srep25485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomma B. P. (2003). Alternaria spp.: from general saprophyte to specific parasite. Mol. Plant Pathol. 4 225–236. 10.1046/j.1364-3703.2003.00173.x [DOI] [PubMed] [Google Scholar]
- Tsuge T., Harimoto Y., Akimitsu K., Ohtani K., Kodama M., Akagi Y., et al. (2013). Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 37 44–66. 10.1111/j.1574-6976.2012.00350.x [DOI] [PubMed] [Google Scholar]
- Wang J., Hu Q., Chen H., Zhou Z., Li W., Wang Y., et al. (2010). Role of individual subunits of the Neurospora crassa CSN complex in regulation of deneddylation and stability of cullin proteins. PLoS Genet. 6:e1001232. 10.1371/journal.pgen.1001232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M., Chen C., Zhu C., Sun X., Ruan R., Li H. (2014). Os2 MAP kinase-mediated osmostress tolerance in Penicillium digitatum is associated with its positive regulation on glycerol synthesis and negative regulation on ergosterol synthesis. Microbiol. Res. 169 511–521. 10.1016/j.micres.2013.12.004 [DOI] [PubMed] [Google Scholar]
- Wang M., Sun X., Yu D., Xu J., Chung K., Li H. (2016). Genomic and transcriptomic analyses of the tangerine pathotype of Alternaria alternata in response to oxidative stress. Sci. Rep. 6:32437. 10.1038/srep32437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M., Sun X., Zhu C., Xu Q., Ruan R., Yu D., et al. (2015). PdbrlA, PdabaA and PdwetA control distinct stages of conidiogenesis in Penicillium digitatum. Res. Microbiol. 166 56–65. 10.1016/j.resmic.2014.12.003 [DOI] [PubMed] [Google Scholar]
- Wei N., Chamovitz D. A., Deng X.-W. (1994). Arabidopsis COP9 is a component of a novel signaling complex mediating light control of development. Cell 78 117–124. 10.1016/0092-8674(94)90578-9 [DOI] [PubMed] [Google Scholar]
- Wei N., Deng X. W. (2003). The COP9 signalosome. Annu. Rev. Cell Dev. Biol. 19 261–286. 10.1146/annurev.cellbio.19.111301.112449 [DOI] [PubMed] [Google Scholar]
- Wei N., Serino G., Deng X.-W. (2008). The COP9 signalosome: more than a protease. Trends Biochem. Sci. 33 592–600. 10.1016/j.tibs.2008.09.004 [DOI] [PubMed] [Google Scholar]
- Xie C., Mao X., Huang J., Ding Y., Wu J., Dong S., et al. (2011). KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 39 w316–w322. 10.1093/nar/gkr483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S. L., Chung K. R. (2012). The NADPH oxidase-mediated production of hydrogen peroxide (H2O2) and resistance to oxidative stress in the necrotrophic pathogen Alternaria alternata of citrus. Mol. Plant Pathol. 13 900–914. 10.1111/j.1364-3703.2012.00799.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S. L., Chung K. R. (2013). Similar and distinct roles of NADPH oxidase components in the tangerine pathotype of Alternaria alternata. Mol. Plant Pathol. 14 543–556. 10.1111/mpp.12026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang S. L., Yu P. L., Chung K. R. (2016). The glutathione peroxidase-mediated reactive oxygen species resistance, fungicide sensitivity and cell wall construction in the citrus fungal pathogen Alternaria alternata. Environ. Microbiol. 18 923–935. 10.1111/1462-2920.13125 [DOI] [PubMed] [Google Scholar]
- Zhang T., Sun X., Xu Q., Candelas L. G., Li H. (2013). The pH signaling transcription factor PacC is required for full virulence in Penicillium digitatum. Appl. Microbiol. Biotechnol. 97 9087–9098. 10.1007/s00253-013-5129-x [DOI] [PubMed] [Google Scholar]
- Zhao R., Yeung S.-C. J., Chen J., Iwakuma T., Su C.-H., Chen B., et al. (2011). Subunit 6 of the COP9 signalosome promotes tumorigenesis in mice through stabilization of MDM2 and is upregulated in human cancers. J. Clin. Invest. 121 851–865. 10.1172/JCI44111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng Y., Wang X., Zhang X., Li W., Liu G., Wang S., et al. (2017). COP9 signalosome subunit PfCsnE regulates secondary metabolism and conidial formation in Pestalotiopsis fici. Sci. China Life Sci. 60 656–664. 10.1007/s11427-017-9068-4 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.