

Abstract
The oral microbiome is a dynamic environment inhabited by both commensals and pathogens. Among these is Streptococcus mutans, the causative agent of dental caries, the most prevalent childhood disease. Carolacton has remarkably specific activity against S. mutans, causing acid-mediated cell death during biofilm formation; however, its complex structure limits its utility. Herein, we report the diverted total synthesis and biological evaluation of a rationally designed library of simplified analogs that unveiled three unique biofilm phenotypes further validating the role of natural product synthesis in the discovery of new biological phenomena.
Natural products have served as starting points for both the discovery of new drugs and the identification of biological targets for decades.1 An exemplary case of the former is penicillin, initially discovered in 1928, which has since been reprogrammed into four new classes of derivatives all in an effort to overcome resistance.2 Recently, natural products have been postulated to serve as excellent starting points for narrow-spectrum therapeutics and also as chemical probes to help deconvolute complex microbiomes and aid in their study.3 The oral microbiome is home to around 1000 species and has been identified as one of the most diverse biotic habitats in the human body.4 Therefore, the identification of small molecules that can specifically target a small subset of pathogenic species would further our understanding of their role in the microbiota. Carolacton is a natural product that displays remarkable bioactivity against the pathogen Streptococcus mutans whereby it does not affect the viability of planktonic bacteria but instead causes acid-mediated cell death during biofilm formation at low nanomolar concentration (Figure 1).5 S. mutans plays a significant role in human health as it is the causative agent in the formation of dental caries,6 the single most common childhood chronic disease.7 Recently, Wagner-Dobler showed that dental composites impregnated with carolacton could inhibit S. mutans biofilm growth8 demonstrating a future role for the natural product in the development of preventative materials.
Figure 1.
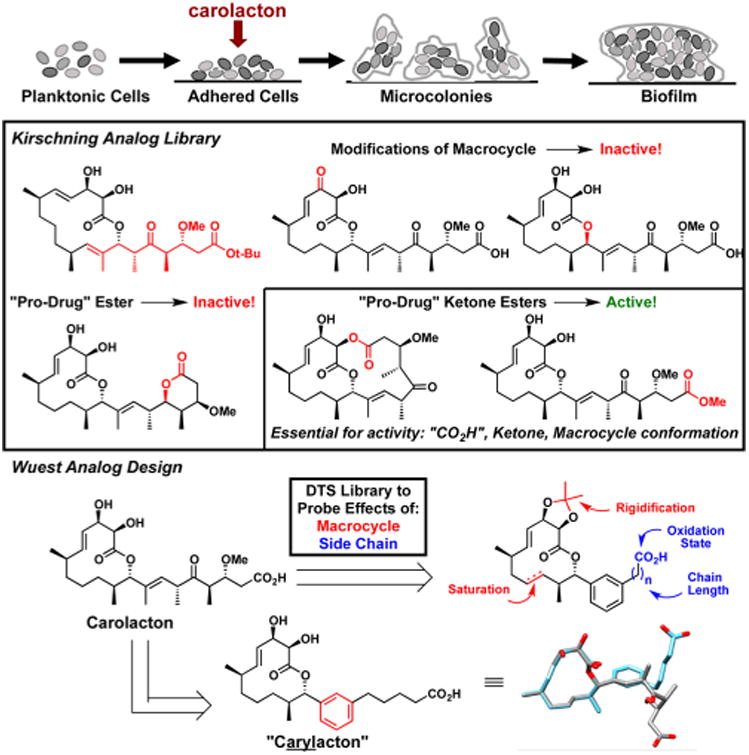
S. mutans biofilm lifecycle (top); Kirschning Analog Library (middle); overview of the diverted total synthesis (DTS) of aryl-analogs; and modeling of simplified analog and carolacton (bottom).
This exquisite bioactivity of carolacton has drawn the interest of the synthetic community as a number of groups have targeted the natural product.9 Kirschning in particular has been quite prolific as they have disclosed both the first total synthesis10 and uncovered useful information regarding the structure—activity relationship (SAR) by analog synthesis (Figure 1).11 These studies elucidated that both drastic (increased ring size) and even small conformational changes to the 12-membered macrocycle, for example, introduction of an α,β-unsaturated ketone or an epimeric allylic alcohol, abolish activity. Further, the side chain ketone was thought to be essential and “pro-drug” ester derivatives affected biofilm development at concentrations relative to their rate of hydrolysis indicating that carolacton, and not the analogs, were responsible for activity. Taken in sum, earlier SAR work demonstrated that significant changes to the macrocycle were not tolerated; however, it was not clear if side chain modifications would be tolerated.
Our initial synthesis of carolacton was accomplished in collaboration with the Phillips group in 2014.12 The strategic design of our synthesis allowed for the late-stage derivatization of the natural product scaffold to elaborate on earlier SAR work, an approach referred to as Diverted Total Synthesis (DTS) (Figure 1).13 Both Danishefsky and Myers have beautifully applied DTS to the development of novel anticancer14 and antibiotic15 compounds, respectively, with improved bioactivity. DTS endeavors from our laboratory have uncovered new phenotypes for the antibiotic promysalin16 and we sought to use our initial total synthesis of carolacton toward this end. We were inspired by the seminal work of Chandrasekhar et al. in their disclosure of simplified pladienolide analogs wherein they rigidified the side chain with an aryl isostere and we determined that a similar replacement was feasible based on computational modeling (Figure 1).17 We and others18 have been drawn to this substitution as it would both simplify the synthesis and effectively evaluate the importance of the elaborate side chain. Herein, we report the DTS of 16 simplified carolacton-inspired analogs that have resulted in the discovery of three independent biofilm phenotypes: (1) inhibition of their formation, (2) inhibition of their maturation, and (3) acid-mediated cell death during formation, akin to carolacton, by an analog coined “carylacton”.
Our first generation DTS library focused on not only conserving the connectivity and stereochemistry of the 12-membered macrolactone, but also significantly simplifying the carolacton side chain by introducing an aryl moiety. We rationally selected 1,3-disubstituted aryl motifs to mimic the trisubstituted olefin and chose to vary both the length and oxidation state of the side chain. This isosteric substitution would thereby eliminate five synthetic operations while maintaining the structural integrity of the compound. Toward this end, we optimized our previously published route to the key intermediate, acid 3, which now proceeds in 36% overall yield (a 50% improvement) and provides gram-quantities in under 2 weeks from commercial material (Scheme 1; for more details, see SI). With both a reliable and scalable route to 3 in hand, we turned our attention to the synthesis of the simplified side chains. We initially chose two aryl diols to use as side chain precursors: a pentyl moiety (closely mimicking carolacton) and a methyl-derivative to interrogate the importance of chain length. Toward this end, monoprotection followed by oxidation of the corresponding 1,3-disubstituted benzene diols provided the known compounds 4a/b. Roush crotylation stereospecifically converts the benzylic aldehydes to the desired alcohols, furnishing structures 5a/b. Leveraging our previous endgame strategy, an esterification united the side chain alcohols with acid 3. Finally, cyclization using ring-closing metathesis resulted in the selective formation of the (E)-olefin, providing the fully protected macrocyclic analog precursors 6a/b, which can then be hydrogenated whereby the more sterically accessible olefin is reduced providing 7a/b. Thus, these four intermediates bearing the fully protected analog skeleton represent the main branching points from which we would synthesize our analog library.
Scheme 1. Diverted Total Synthesis of Carolacton Analogsa.
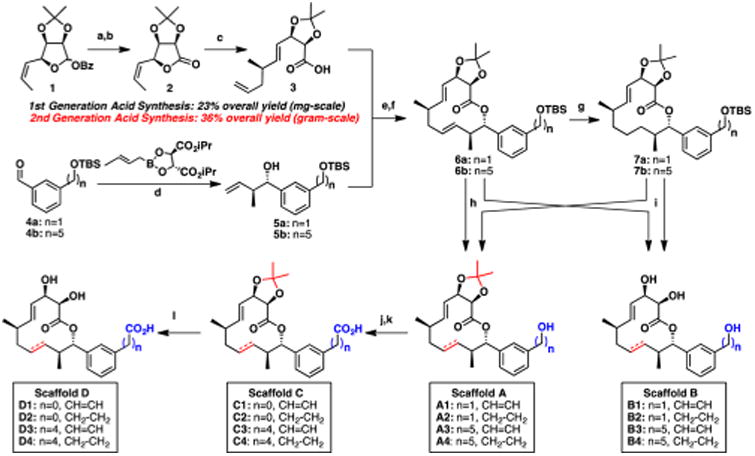
a(a) K2CO3, MeOH/THF, >99% (b) DMSO, (COCl)2, CH2Cl2, 94% (c) CuCN, Allyl-MgBr, THF, 90% (d) 4 Å MS, NaOH, PhMe, 83– 85% (e) EDCI, DMAP, CH2Cl2, 89–96% (f) Grubbs 2nd generation catalyst, CH2Cl2, 78–99% (g) Pd/C, H2, 55-69% (h) TBAF, 64–94% (i) HCl, THF/MeOH, 50–82% (j) DMSO, SO3·pyr, CH2Cl2, 63–89% (k) t-BuOH, 2-methyl-2-butene, NaH2PO4, NaClO2, MeCN, H2O, 65-96% (l) TFA/H2O, 59–79%.
It is well-known that slight structural modifications to macrolactones can result in significant biological outcomes;19 for example, the simple deletion of a methyl group is enough to render a molecule inactive, as was nicely demonstrated by the Andrade lab in their syntheses of desmethyl telithromycin analogs.20 On the basis of these prior reports, we sought to evaluate fully all desilyated compounds en route to “carylacton” to understand better the SAR of such perturbations. We envisioned three logical branching points that would facilitate the production of four distinct scaffolds whereby we could evaluate the role that the acetonide, olefin, and oxidation state play in bioactivity. Facile deprotection of silyl-ethers 6 and 7 generate Scaffolds A and B, respectively, which differ by an acetonide group. Scaffold A, bearing the primary alcohol and acetonide, can then undergo a two-step oxidation, providing the corresponding acid found in Scaffold C. Finally, the acetonide is treated with trifluoroacetic acid in water to furnish Scaffold D (Scheme 1). In total, milligram quantities of 16 analogs were synthesized and further evaluated for their biological activity in S. mutans UA159 assays described below.
We sought to evaluate initially the biological activity of the analog library via minimum inhibitory concentration (MIC) and minimum biofilm inhibitory concentration (MBIC) assays. We first wanted to evaluate the ability of the compounds to affect biofilm formation. As we expected D4 and carolacton were both inactive, however, we serendipitously discovered that, compounds C3 and C4 inhibited biofilm adherence (Figure 2A). We quantified their activity by measuring the concentration at which 50% inhibition of biofilm formation (MBIC50) occurs and found it to be 63 and 500 μM, respectively (see Figures S1 and S2 for details). To confirm that the compounds were specifically targeting biofilms and not simply killing planktonic cells we performed MIC assays against planktonic bacteria wherein the media lacked sucrose. As expected, all 16 analogs and carolacton were ineffective at inhibiting planktonic growth (MIC ≥ 500 μM). The discovery of a simplified carolacton-inspired analog (C3) available in ten steps that inhibits biofilm formation is notable.
Figure 2.
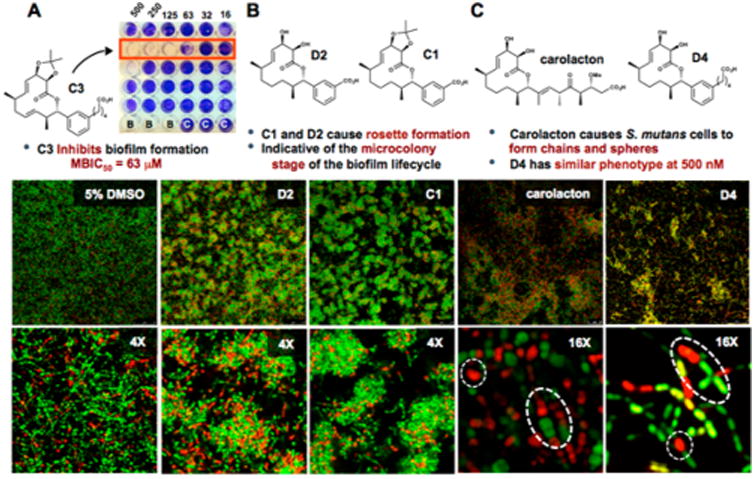
Carolacton analogs elicit a range of phenotypic responses. (A) C3 inhibits biofilm formation with an MBIC50 of 63 μM, “B” = Blank wells; “C” = S. mutans control. (B) Confocal microscopy highlights the effect of C1 and D2 on the growth of S. mutans biofilm at 500 μM (top, field view; bottom, 4× zoom). (C) Carolacton and D4 (top, field view; bottom, 16× zoom) display similar phenotypes at 500 nM. White circles indicate spherical cell morphology and ovals highlight chains of cells. See Figures S3 and S4 for additional representative images.
The eradication of biofilms is a longstanding challenge that has inspired many research programs to understand better their chemical underpinnings. We sought to screen our analogs in whole cell phenotypic biofilm assays to identify unique biological responses that could potentially unveil new targets or stall biofilm maturation processes for future microbiological analysis. We rationalized that the structural diversity of our library may provide leads for probe development and/or proteomic analysis and wanted to evaluate them fully via microscopy. Figure 2 depicts biofilms treated with 500 μM of carolacton and stained with propidium iodide/SYTO9 to demonstrate representative phenotypic results. All compounds except C3 and C4 (which inhibit biofilm formation) were able to be imaged at this concentration. Analogs C1 and D2 shared a common and unique phenotypic response that we had not previously been seen through our studies with carolacton. The two analogs appear to arrest growth at the stage of microcolony formation, which can be seen in the field view by their rosettelike pattern in Figure 2B, thereby halting biofilm maturation. At 4× zoom, a haze is apparent surrounding the microcolonies (as compared to the DMSO control) indicating possible increased SYTO9-staining extracellular DNA (eDNA) in the extracellular polymeric substance (EPS). This biofilm phenotype is distinctly different to that of carolacton-treated cells. On the basis of these findings, we postulate that this subset of analogs is affecting S. mutans biofilm growth via separate modes of action compared to carolacton and is the subject of ongoing investigation. Of note, these analogs contain a carboxylic acid that appears to be important for bioactivity as the corresponding alcohols are inactive (Figure S1), consistent with the findings of Kirschning.11
At the onset, we postulated that the aryl isostere of carolacton could serve as a simplified analog; therefore, we sought to investigate the range of activity for compound D4 (“carylacton”). It has been shown previously that at nanomolar concentrations of carolacton S. mutans cells begin to form long chains and become significantly more spherical than untreated cells within a biofilm. To our delight, D4 displayed a phenotype consistent to that of carolacton at concentrations as low as 500 nM (highlighted by white dotted circles/ovals in Figure 2C). This observation lends credence to our earlier hypothesis that our rationally designed isosteric structure would maintain biological activity similar to carolacton. Further testing is needed to validate that D4 is acting by the same mechanism of action as the natural product and is currently ongoing. Nevertheless, this work reveals that significant side chain modifications are tolerated without the loss of biological activity. It also serves as another example that meta-substituted aryl rings can serve as isosteres of trisubstituted olefins, which are present in a number of structurally diverse natural products.
The results disclosed herein highlight our DTS endeavor that revealed a multitude of compounds that each elicited distinct biological responses in S. mutans. Figure 3 summarizes the findings of this current bioorganic exploration. Our work demonstrates that subtle changes to the macrocyclic ring of carolacton results in substantial phenotypic outcomes (see Figure S5 for computational modeling of analogs). We initially discovered that compound C3 inhibits the formation of biofilms at concentrations as low as 63 μM. Several molecules have been shown to inhibit adherence of S. mutans biofilm such as betulin, vizantin, and quercitrin.21 Surprisingly, we identified two truncated carboxylate-bearing compounds (C1, D2) that arrested biofilm growth at the microcolony stage. It is well-established that hydrogen fluoride inhibits the growth of streptococci by dissipating the bacterial trans-membrane proton-motive force, thereby disrupting the membrane-bound, proton-pumping F-ATPase, and effectively acidifying the organism.22 We postulate that the benzoic acid derivatives containing the carolacton macrocycle saturation state, which are inherently more acidic than their alkyl counterparts, may act to inhibit growing cells of S. mutans by an analogous mechanism. Finally, our rationally designed target molecule, “carylacton” (D4), mimics carolacton both in structure and bioactivity, exhibiting a similar effect on S. mutans biofilms at nanomolar concentration. These findings demonstrate that our analogs can potentially control S. mutans growth at discrete points in biofilm maturation.
Figure 3.
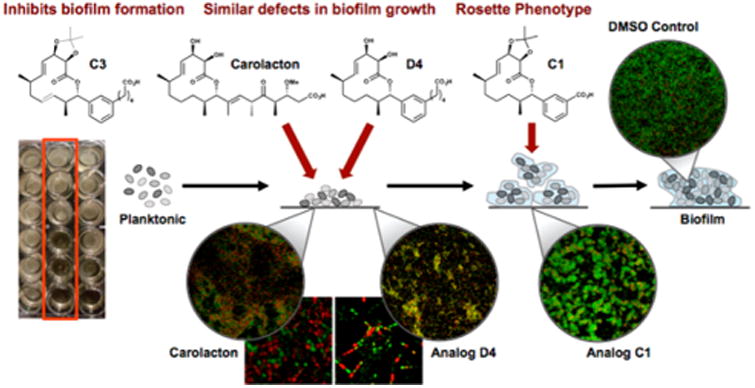
Summary of the unique phenotypes, and proposed stages of activity, of the carolacton-inspired analogs during the maturation of S. mutans biofilms. C3 inhibits biofilm growth at 63 μM (left). D4 kills cells during the growth of the biofilm similar to that of the natural product, carolacton (center). C1 inhibits biofilm maturation (bottom right).
The composition of the genome and proteome of a growing biofilm is constantly fluctuating.23 Having the ability to arrest the growth at a specific point of the biofilm formation will allow for more in-depth studies on how biofilms grow, thus providing “snapshots” of potential targets present only briefly in the biofilm lifecycle. Forthcoming work will investigate both the mechanism of action and biological targets of these analogs. In summary, the DTS of simplified carolacton analogs yielded a surprising diversity of bioactivity. Four lead compounds that are unavailable via isolation or derivatization were identified, further highlighting the value of natural product total synthesis and validating its role in biological discoveries. Two of these compounds, C3 and D4, are greatly simplified analogs and make for viable options for the future development of dental materials.
Supplementary Material
Acknowledgments
We thank K.P.C. Minbiole and S.E. Wengryniuk for valuable feedback, A. Hess, A. Plaster, and C. Burns-Lynch for synthetic assistance, and Materia, Inc. for catalysts. National Science Foundation CHE-1454116 (W.M.W.) and MCB-1412508 (V.AV.), the National Institute of Dental and Craniofacial Research DE-025837 (W.M.W.), DE017425 and DE013683 (R.G.Q.), and Temple University.
Footnotes
Associated Content: Supporting Information: The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/jacs.7b03879. Methods and synthesis (PDF)
Notes: The authors declare no competing financial interest.
References
- 1.(a) Newman DJ, Cragg GM. J Nat Prod. 2007;70:461. doi: 10.1021/np068054v. [DOI] [PubMed] [Google Scholar]; (b) Butler MS, Robertson AAB, Cooper MA. Nat Prod Rep. 2008;25:475. [Google Scholar]
- 2.Lewis K. Nat Rev Drug Discovery. 2013;12:371. doi: 10.1038/nrd3975. [DOI] [PubMed] [Google Scholar]
- 3.Brown EJ, Albers MW, Shin TB, Ichikawa K, Keith CT, Lane WS, Schreiber SL. Nature. 1994;369:756. doi: 10.1038/369756a0. [DOI] [PubMed] [Google Scholar]
- 4.Keohane CE, Steele AD, Wuest WM. Synlett. 2015;26:2739. [Google Scholar]
- 5.(a) Kunze B, Reck M, Dotsch A, Schummer D, Irschik H, Steinmetz H, Wagner-Dobler I. BMC Microbiol. 2010;10:1. doi: 10.1186/1471-2180-10-199. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Reck M, Rutz K, Kunze B, Tomasch J, Surapaneni SK, Schulz S, Wagner-Dobler I. J Bacteriol. 2011;193:5692. doi: 10.1128/JB.05424-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lemos JA, Quivey RG, Koo H, Abranches J. Microbiology. 2013;159:436. doi: 10.1099/mic.0.066134-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.(a) Bagramian RA, Garcia-Godoy F, Volpe AR. Am J Dent. 2009;22:3. [PubMed] [Google Scholar]; (b) Listl S, Galloway J, Mossey PA, Marcenes W. J Dent Res. 2015;94:1355. doi: 10.1177/0022034515602879. [DOI] [PubMed] [Google Scholar]
- 8.Apel C, Barg A, Rheinberg A, Conrads G, Wagner-Döbler I. Dent Mater. 2013;29:1188. doi: 10.1016/j.dental.2013.09.005. [DOI] [PubMed] [Google Scholar]
- 9.(a) Sabitha G, Shankaraiah K, Prasad MN, Yadav JS. Synthesis. 2013;45:251. [Google Scholar]; (b) Rao KS, Ghosh S. Synthesis. 2013;45:2745. [Google Scholar]; (c) Sharma GVM, Reddy SV. RSC Adv. 2013;3:21759. [Google Scholar]; (d) Kuilya TK, Goswami RK. Org Lett. 2017;19:2366. doi: 10.1021/acs.orglett.7b00903. [DOI] [PubMed] [Google Scholar]
- 10.Schmidt T, Kirschning A. Angew Chem, Int Ed. 2012;51:1063. doi: 10.1002/anie.201106762. [DOI] [PubMed] [Google Scholar]
- 11.Stumpp N, Premnath P, Schmidt T, Ammermann J, Dräger G, Reck M, Jansen R, Stiesch M, Wagner-Döbler I, Kirschning A. Org Biomol Chem. 2015;13:5765. doi: 10.1039/c5ob00460h. [DOI] [PubMed] [Google Scholar]
- 12.Hallside MS, Brzozowski RS, Wuest WM, Phillips A. J Org Lett. 2014;16:1148. doi: 10.1021/ol500004k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Szpilman AM, Carreira EM. Angew Chem, Int Ed. 2010;49:9592. doi: 10.1002/anie.200904761. [DOI] [PubMed] [Google Scholar]
- 14.Cho YS, Wu KD, Moore MAS, Chou TC, Danishefsky SJ. Drugs Future. 2005;30:737. [Google Scholar]
- 15.Sun C, Wang Q, Brubaker JD, Wright PM, Lerner CD, Noson K, Charest M, Siegel DR, Wang YM, Myers AG. J Am Chem Soc. 2008;130:17913. doi: 10.1021/ja806629e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Steele AD, Keohane CE, Knouse KW, Rossiter SE, Williams SJ, Wuest WM. J Am Chem Soc. 2016;138:5833. doi: 10.1021/jacs.6b03373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kumar PV, Chandrasekhar S. Org Lett. 2013;15:3610. doi: 10.1021/ol401458d. [DOI] [PubMed] [Google Scholar]
- 18.(a) Erdelyi M, Navarro-Vazquez A, Pfeiffer B, Kuzniewski CN, Felser A, Widmer T, Gertsch J, Pera B, Diaz JF, Altmann KH, Carlomagno T. ChemMedChem. 2010;5:911. doi: 10.1002/cmdc.201000050. [DOI] [PubMed] [Google Scholar]; (b) Maier ME. Org Biomol Chem. 2015;13:5302. doi: 10.1039/c5ob00169b. [DOI] [PubMed] [Google Scholar]
- 19.Brzozowski RS, Wuest WM. Chem Biol Drug Des. 2017;89:169. doi: 10.1111/cbdd.12783. [DOI] [PubMed] [Google Scholar]
- 20.Andrade RB. Synlett. 2015;26:2199. [Google Scholar]
- 21.(a) Hasan NA, Young BA, Minard-Smith AT, Saeed K, Li H, Heizer EM, McMillan NJ, Isom R, Abdullah AS, Bornman DM, Faith SA, Choi SY, Dickens ML, Cebula TA, Colwell RR. PLoS One. 2014;9:e106124. doi: 10.1371/journal.pone.0097699. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Viszwapriya D, Subramenium GA, Radhika S, Pandian SK. Antonie van Leeuwenhoek. 2017;110:153. doi: 10.1007/s10482-016-0785-3. [DOI] [PubMed] [Google Scholar]; (c) Takenaka S, Oda M, Domon H, Ohsumi T, Suzuki Y, Ohshima H, Yamamoto H, Terao Y, Noiri Y. Biochem Biophys Res Commun. 2016;480:173. doi: 10.1016/j.bbrc.2016.10.021. [DOI] [PubMed] [Google Scholar]
- 22.Marquis RE. J Dent Res. 1990;69:627. doi: 10.1177/00220345900690S130. [DOI] [PubMed] [Google Scholar]
- 23.Sauer K. Genome Biol. 2003;4:1. doi: 10.1186/gb-2003-4-6-219. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.